Epigenetics and cell fate in JIA and pulmonary fibrosis
Jim HagoodUCSD/RCHSD Division of Respiratory Medicine
Caring, Curing, Discovering

Outline
• Lung remodeling in fibrosis
• Possible role of epigenetic mechanisms in IPF and autoimmunity, JIA
• What can we learn from epigenomics?
• miRNA and other non-coding RNA will not be covered
• Promise and pitfalls of epigenetics targeted therapy
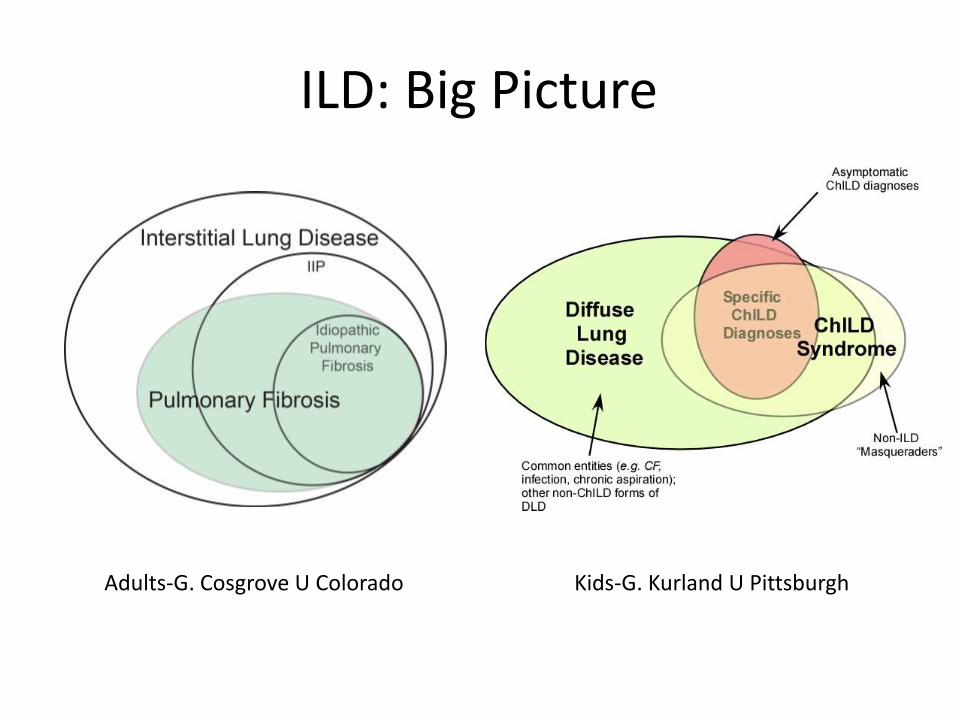
Adults-G. Cosgrove U Colorado Kids-G. Kurland U Pittsburgh
ILD: Big Picture

IPF: Impact
• Affects more than 120,000 people in the U.S., with about 48,000 new cases diagnosed annually. 40,000 people die each year to IPF, the same as to breast cancer
• IPF is five times more common than cystic fibrosis and Lou Gehrig’s Disease (ALS), yet the disease remains virtually unknown to general public.
• IPF receives a fraction of the research funding (IPF: approx. $18 million per year; Cystic Fibrosis and ALS: $85 million and $48 million per year respectively.
• There is no known cause, no cure. New FDA-approved treatments slow progression but no impact on mortality.
www.coalitionforpf.org
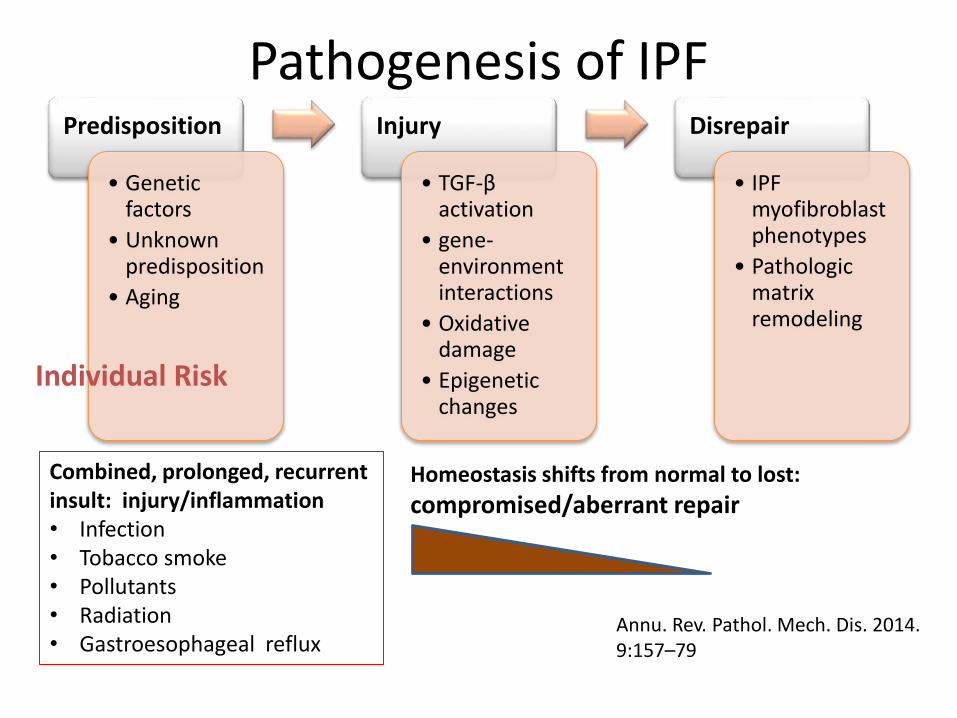
Pathogenesis of IPFPredisposition
• Genetic factors
• Unknown predisposition
• Aging
Injury
• TGF-β activation
• gene-environment interactions
• Oxidative damage
• Epigenetic changes
Disrepair
• IPF myofibroblastphenotypes
• Pathologic matrix remodeling
Combined, prolonged, recurrent insult: injury/inflammation• Infection• Tobacco smoke• Pollutants• Radiation• Gastroesophageal reflux
Individual Risk
Annu. Rev. Pathol. Mech. Dis. 2014. 9:157–79
Homeostasis shifts from normal to lost:
compromised/aberrant repair

Lung Cell Phenotype Regulation
• Lung development begins as a simple epithelial tube invading a mesenchymal matrix
• Subsequently there is a marked increase in structural complexity, accompanied by cellular differentiation, that persist into adolescence
• In addition to genetic influences, interaction with the environment (e.g., infection, toxicants, oxyradicals, mechanical environment) can have major effects on cell phenotype, lung development, and remodeling
• Most diffuse/interstitial lung disease is characterized by marked alteration in cellular phenotypes

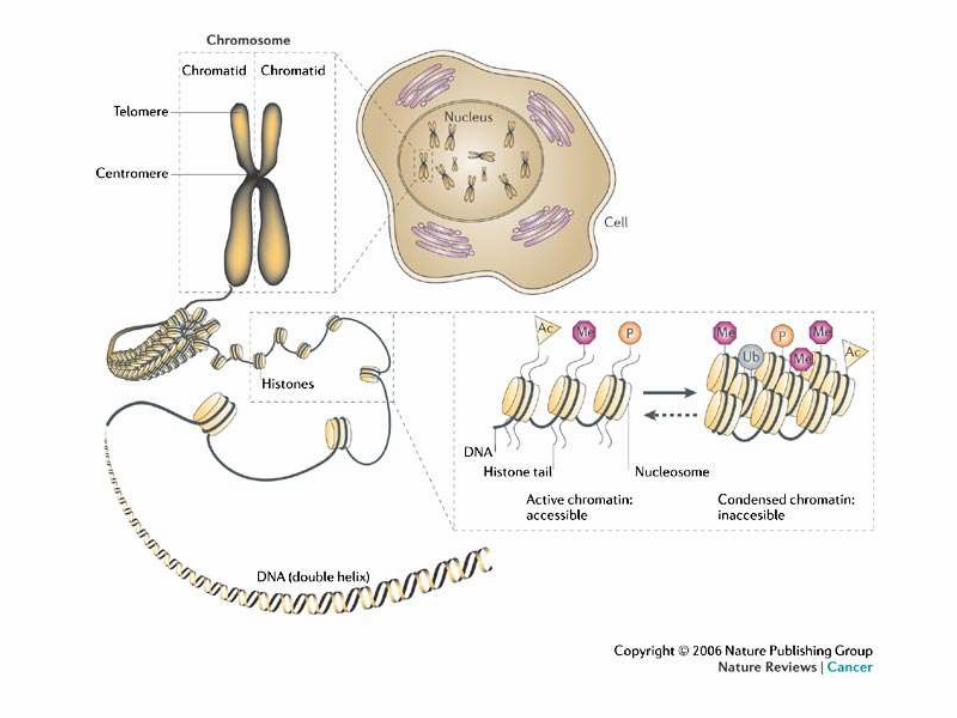
Epigenetics
• study of heritable changes in gene function that occur without a change in the DNA sequence
• “The structural adaptation of chromosomal regions so as to register, signal or perpetuate altered activity states”
• DNA methylation, histone acetylation, and RNA interference, and their effects in gene activation and inactivation
• DNA is not just a string of bases
Bird A, Nature 2007, 447:396–398

Why Epigenetics?
• From single cell to 50-75 x 1012 cells, >200 cell types; genome remains the same, for the most part
• Disease phenotype variability within single genomic abnormalities
• Genetic variants collectively account for a small fraction of the heritability of complex phenotypes
• Epigenetic modifications (DNA methylation, histone tail modifications, chromatin remodeling and noncoding RNA expression) have major influence on gene expression, which drives cell phenotype alteration
• All disease paradigms (inflammation, wound repair, etc.) relevant to CTD and ILD involve changes in cell phenotype
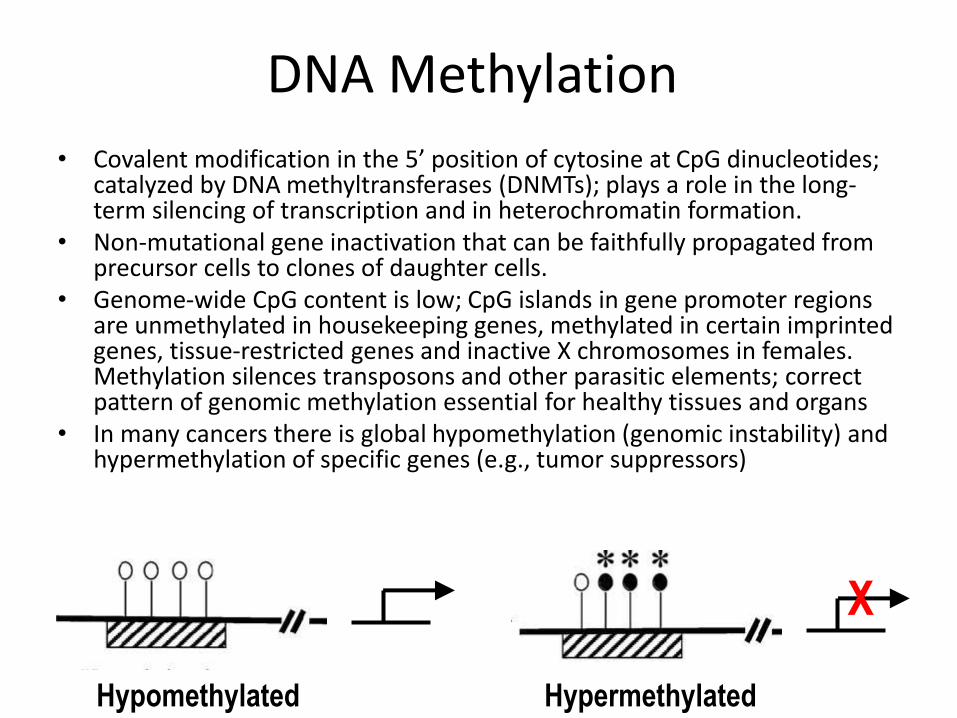
DNA Methylation
• Covalent modification in the 5’ position of cytosine at CpG dinucleotides; catalyzed by DNA methyltransferases (DNMTs); plays a role in the long-term silencing of transcription and in heterochromatin formation.
• Non-mutational gene inactivation that can be faithfully propagated from precursor cells to clones of daughter cells.
• Genome-wide CpG content is low; CpG islands in gene promoter regions are unmethylated in housekeeping genes, methylated in certain imprinted genes, tissue-restricted genes and inactive X chromosomes in females. Methylation silences transposons and other parasitic elements; correct pattern of genomic methylation essential for healthy tissues and organs
• In many cancers there is global hypomethylation (genomic instability) and hypermethylation of specific genes (e.g., tumor suppressors)
Hypomethylated Hypermethylated
X
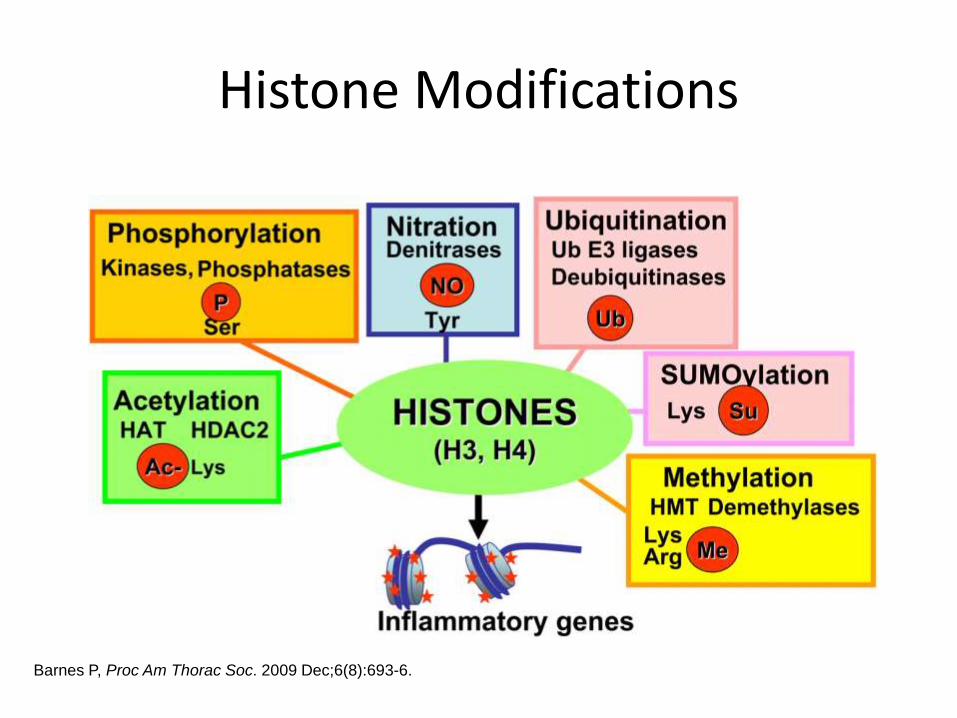
Histone Modifications
Barnes P, Proc Am Thorac Soc. 2009 Dec;6(8):693-6.
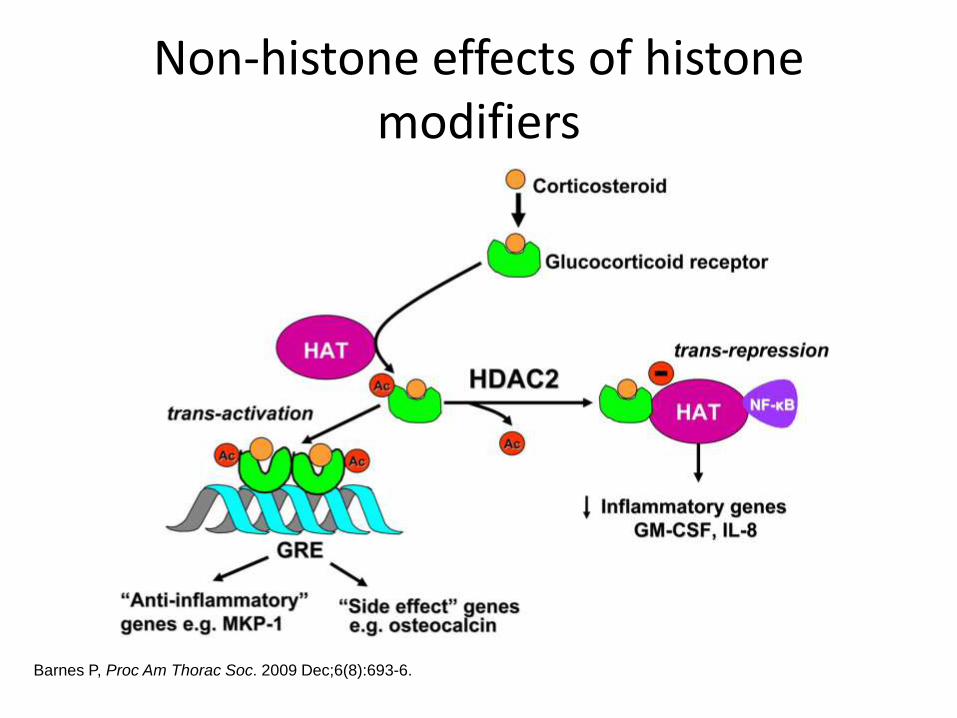
Non-histone effects of histone modifiers
Barnes P, Proc Am Thorac Soc. 2009 Dec;6(8):693-6.
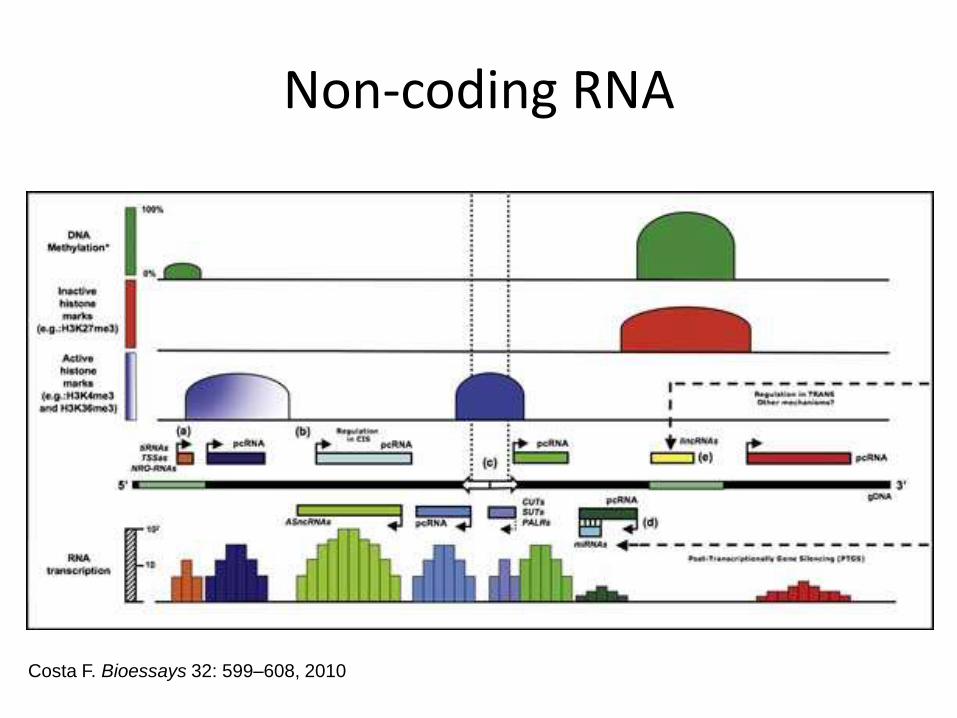
Non-coding RNA
Costa F. Bioessays 32: 599–608, 2010

Chromatin and Nuclear Architecture
• Chromatin: highly ordered structure that contains DNA, RNA, histones and other chromosomal proteins.
• Originally classified into two domains, euchromatin and heterochromatin, based on the density of staining in micrographs
• Euchromatin is gene-rich, transcriptionally active, hyperacetylated, hypomethylated chromatin.
• Heterochromatin is transcriptionally inactive, gene-poor, hypoacetylated and hypermethylated
• Lamins (A, B1, B2 and C3) interact with chromatin and each other to create a specific three-dimensional nuclear architecture, disruption of which leads to deformed nuclei, genome instability, age-related diseases and cancer
Black JC Epigenetics 6:1, 9-15; January 2011

IPF and epigenetics
• IP-10 expression is decreased in F-IPF due to histone modifications and altered recruitment of HATs and HDAC-containing repressor complexes to the IP-10 promoter; expression is restored by HDAC and G9a inhibitors
• Suberoylanilide hydroxamic acid (SAHA, an HDACi) abrogates TGF-β1 effects on IPF and normal lung fibroblasts by preventing transdifferentiation into α-SMA positive myofibroblasts and increased collagen deposition
• THY1 is silenced in IPF fibroblasts; DNMT and HDAC inhibitors restore expression and suppress myofibroblastphenotype
• Interaction between DNMT-1 and miR-17~92 regulates multiple profibrotic pathways in IPF lung tissue
Coward WR, Mol Cell Biol. 30(12):2874, 2010; Wang Z, Eur Respir J. 34(1):145, 2009;
Sanders Y, Am J Respir Cell Mol Biol 39:610, 2008; Marsh CB

Other diseases
• Rheumatoid arthritis synovial fibroblasts (RASF): hypermethylation of DR3, hypomethylation of IL6, reversible histone acetylation and apoptosis; altered methylation in mononuclear cells, T cells
• Myofibroblastic activation of hepatic stellate cells by epigenetic mechanisms; methylation silencing of SOCS-1 in hepatic fibrosis, hepatocellular carcinoma
• HDAC4 required for TGF-b-induced myofibroblasticdifferentiation of skin fibroblasts
• Methylation of FLI1 associated with increased collagen expression in scleroderma fibroblasts
Sánchez-Pernaute O, J Autoimmunity 30: 12, 2008; Ellis et al. Clinical
Epigenetics 2012, 4:20; Mann DA J Gastroenterol Hepatol 23: S108, 2008; Ogata
H, Oncogene 25: 2520, 2006; Glenisson W, BBA-MCR 1773: 1572, 2007; Wang
Y, Arthritis Rheum 54: 2271, 2006
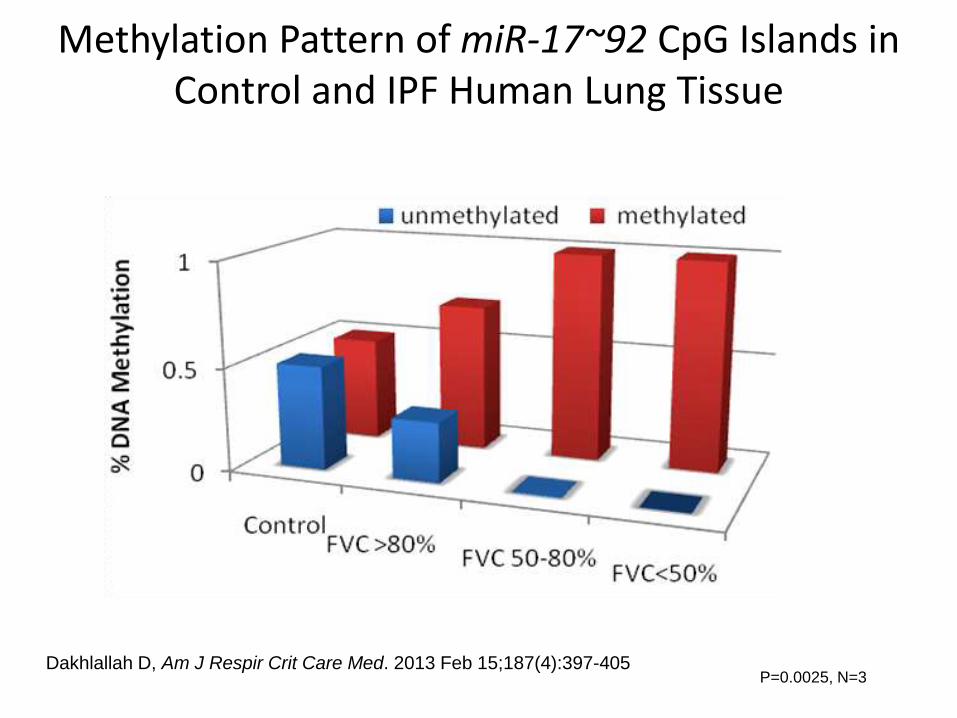
Methylation Pattern of miR-17~92 CpG Islands in Control and IPF Human Lung Tissue
P=0.0025, N=3Dakhlallah D, Am J Respir Crit Care Med. 2013 Feb 15;187(4):397-405

Epigenomics: the “methylome”: searching for new targets
• Sanders YY, Am J Respir Crit Care Med 2012;186:525–535
– Lung tissue IPF (12, severe, explant, 60.3y) v. normal (7, failed donor, 39y)
– Illumina human Methylation27 BeadChip (bisulfite modification, identifies known CpG sites) and human HT-12 BeadChip (RNA)
– Validation of selected genes with RT-PCR, methylation-specific PCR, WB, IHC

IPF Normal
Up-RegulatedDown-Regulated
16
DNA
methylation
array
RNA
expression
array
RNA Expression Array-IPF

-0.150
-0.100
-0.050
0.000
0.050
0.100
0.150
-8.000
-6.000
-4.000
-2.000
0.000
2.000
4.000
6.000
8.000
10.000
12.000
Δ B
eta
Fold
Ch
ange
Fold Change (IPF vs Normal) IPF Delta Beta
Overlap: Methylation/Expression
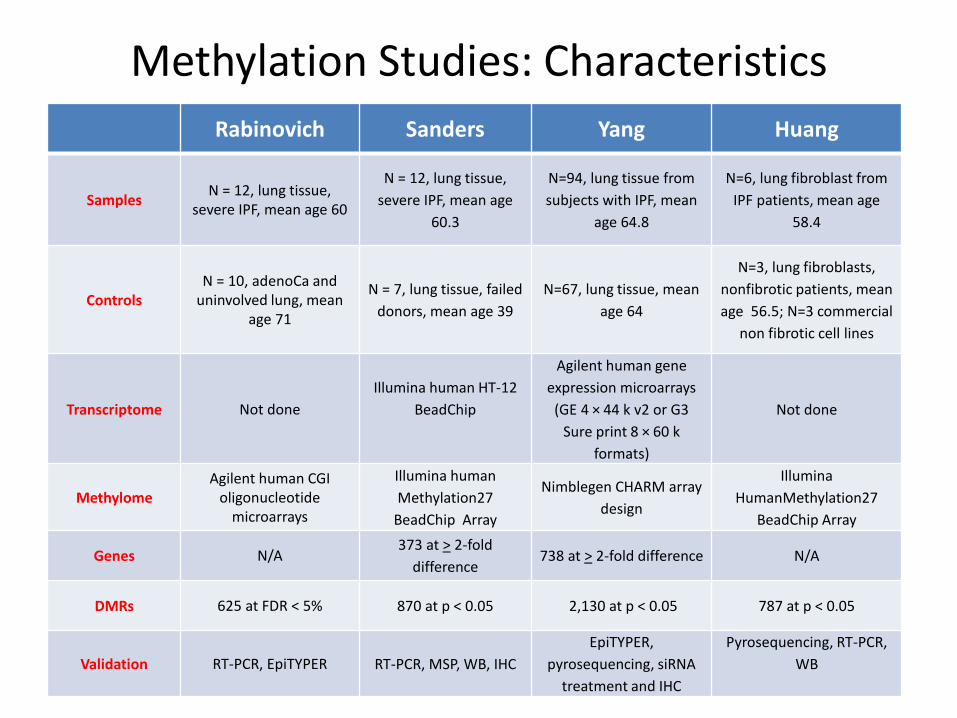
Rabinovich Sanders Yang Huang
SamplesN = 12, lung tissue,
severe IPF, mean age 60
N = 12, lung tissue,
severe IPF, mean age
60.3
N=94, lung tissue from
subjects with IPF, mean
age 64.8
N=6, lung fibroblast from
IPF patients, mean age
58.4
ControlsN = 10, adenoCa and
uninvolved lung, mean age 71
N = 7, lung tissue, failed
donors, mean age 39
N=67, lung tissue, mean
age 64
N=3, lung fibroblasts,
nonfibrotic patients, mean
age 56.5; N=3 commercial
non fibrotic cell lines
Transcriptome Not done
Illumina human HT-12
BeadChip
Agilent human gene
expression microarrays
(GE 4 × 44 k v2 or G3
Sure print 8 × 60 k
formats)
Not done
MethylomeAgilent human CGI
oligonucleotide microarrays
Illumina human
Methylation27
BeadChip Array
Nimblegen CHARM array
design
Illumina
HumanMethylation27
BeadChip Array
Genes N/A373 at > 2-fold
difference738 at > 2-fold difference N/A
DMRs 625 at FDR < 5% 870 at p < 0.05 2,130 at p < 0.05 787 at p < 0.05
Validation RT-PCR, EpiTYPER RT-PCR, MSP, WB, IHC
EpiTYPER,
pyrosequencing, siRNA
treatment and IHC
Pyrosequencing, RT-PCR,
WB
Methylation Studies: Characteristics

Rabinovich Sanders Yang
Cellular Assembly and
OrganizationHumoral Immune Response Gene Expression
Cellular Growth and
ProliferationEnergy Production Cellular Development
Cell MorphologyCellular Assembly and
OrganizationCellular Growth and
Proliferation
Cancer Molecular TransportHematological System
Development and Function
Cell Signaling DNA ReplicationCardiovascular System
Development and Function
Gene ExpressionCellular Growth and
ProliferationOrganismal Development
Cell Death Protein Trafficking Hematopoiesis
Methylation Studies: Functional Analysis

“Methylome” Studies: Key Points
Limitations Key Insights
Based on whole tissue (signals from mixtures of cells)
Differential methylation at CpG sites across the genome; confirmed by alternate techniques
Different platforms may yield different results
Many of the DMRs are outside promoters
Omit hydroxymethylcytosine and N6-methyladenine
Can be used to identify novel mediators and pathways
Confirmation and biological plausibility of differentially methylated genes
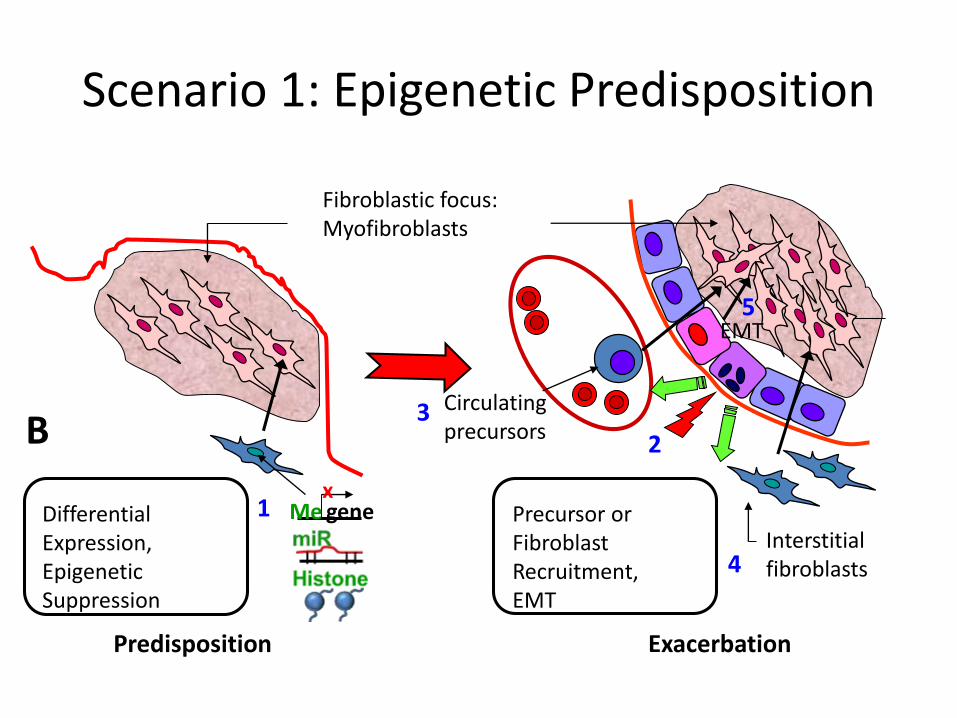
Differential Expression,Epigenetic Suppression
Fibroblastic focus:Myofibroblasts
Predisposition Exacerbation
Circulatingprecursors
2
Interstitialfibroblasts
Precursor orFibroblastRecruitment,EMT
1
3
genex
Me
4
B
Scenario 1: Epigenetic Predisposition
5EMT
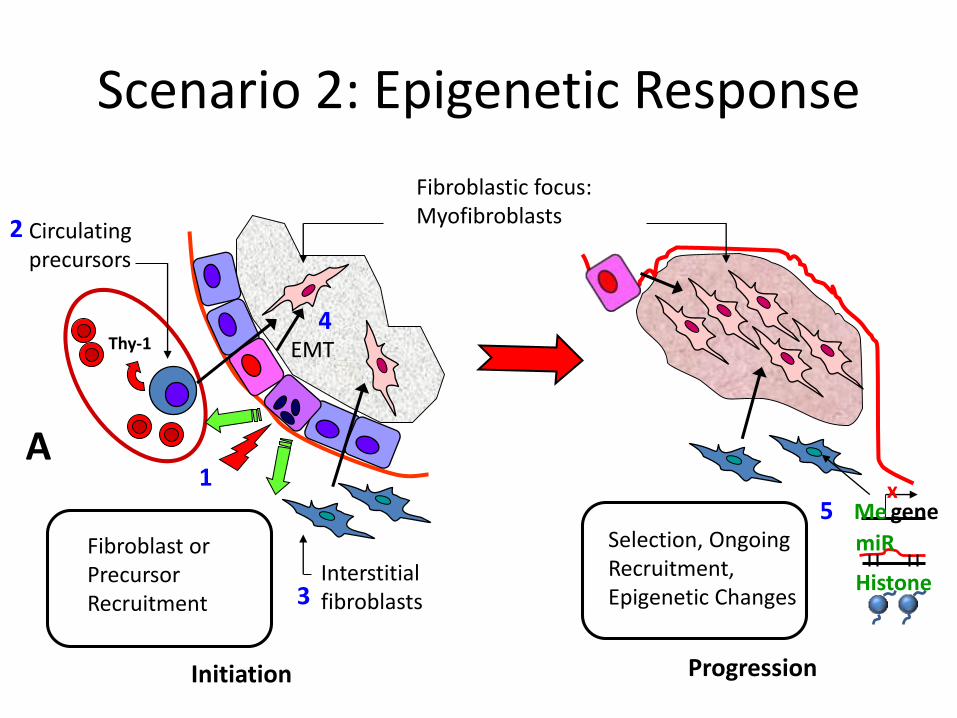
Interstitialfibroblasts
Thy-1
Selection, OngoingRecruitment,Epigenetic Changes
Fibroblast or PrecursorRecruitment
Circulatingprecursors
Fibroblastic focus:Myofibroblasts
Initiation Progression
genex
Me
1
2
3
4
A
Scenario 2: Epigenetic Response
EMT
5miR
Histone
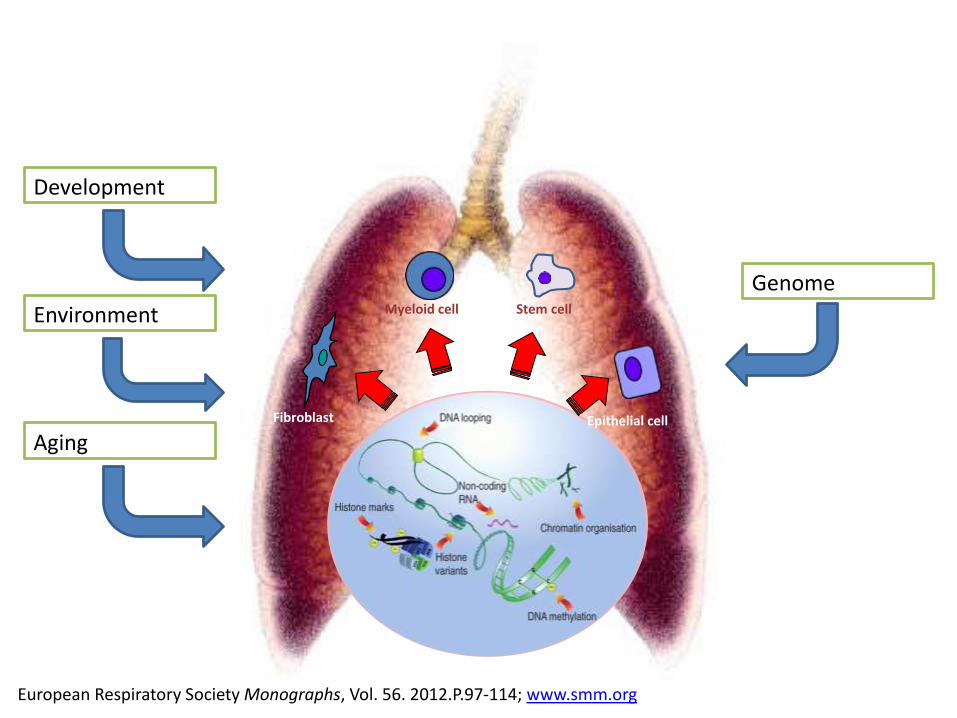
European Respiratory Society Monographs, Vol. 56. 2012.P.97-114; www.smm.org
Genome
Development
Environment
AgingFibroblast
Myeloid cell Stem cell
Epithelial cell

Epigenetics and JIA
• T cell differentiation is in part epigenetically controlled
• T cell methylation different at 145 loci vs. controls (11 after adjusting for methotrexate)
• Top networks with differentially methylated loci included ‘immunological disease’ (21), ‘cellular growth and proliferation’ (16), ‘antigen presentation’ (15) and ‘cell-to-cell signalling and interaction’ (15)
• Differential IL32 methylation and expression confirmed
Ellis et al. Clinical Epigenetics 2012, 4:20

Epigenetics and Autoimmunity
• Gender bias in some autoimmune diseases, modest concordance in MZ twins suggest epigenetic contribution
• Demethylation of inflammatory loci in SLE T cells and B Cells
• Neutrophil “methylome” in SLE has significant demethylation in “interferon signature” loci; similar to prior findings in CD4+ T cells
• Multiple alterations in histone acetylation and histone lysine methylation in SLE monocytes
• Significant alterations in DNA methylation in RA monocytes and RASF
• HDACi block inflammatory cytokine production by RA macrophages
Mau T, Front Genet. 2014 Dec 19; Coit P, J Autoimmun. 2015 Jan 28; Grabiec et al, J. Immunol. 184, 2718–2728 ; Jeffries MA, Expert Rev Clin Immunol. 2015 Jan

Epigenetic therapies
• DNA methyltransferase (DNMT) inhibitors
• Histone deacetylase (HDAC) inhibitors
• Many are already in clinical trials for a number of malignancies; many have been tested in animal models of systemic inflammatory disorders or in vitro
• Many other chromatin modifications can be targeted by small molecule inhibitors
• miRNA-based therapeutics in development
• Specificity and targeting are critical
• Ongoing study of critical “nodes” controlling epigenetic modifications

The epigenetic therapy pipeline
Dawson, MA. Cell 150: 12, 2012
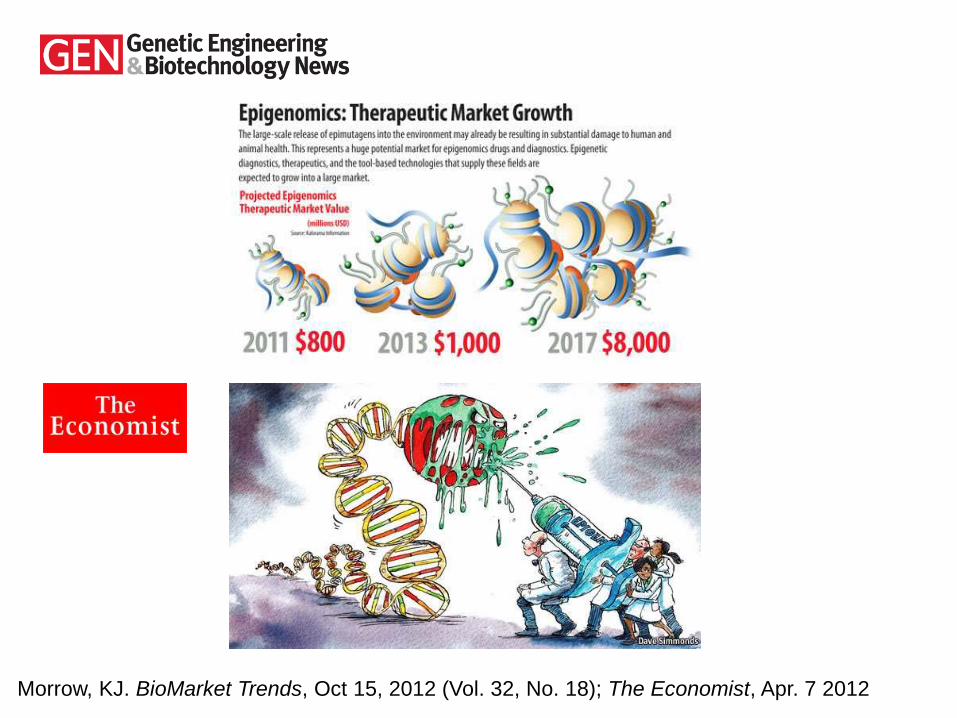
Morrow, KJ. BioMarket Trends, Oct 15, 2012 (Vol. 32, No. 18); The Economist, Apr. 7 2012
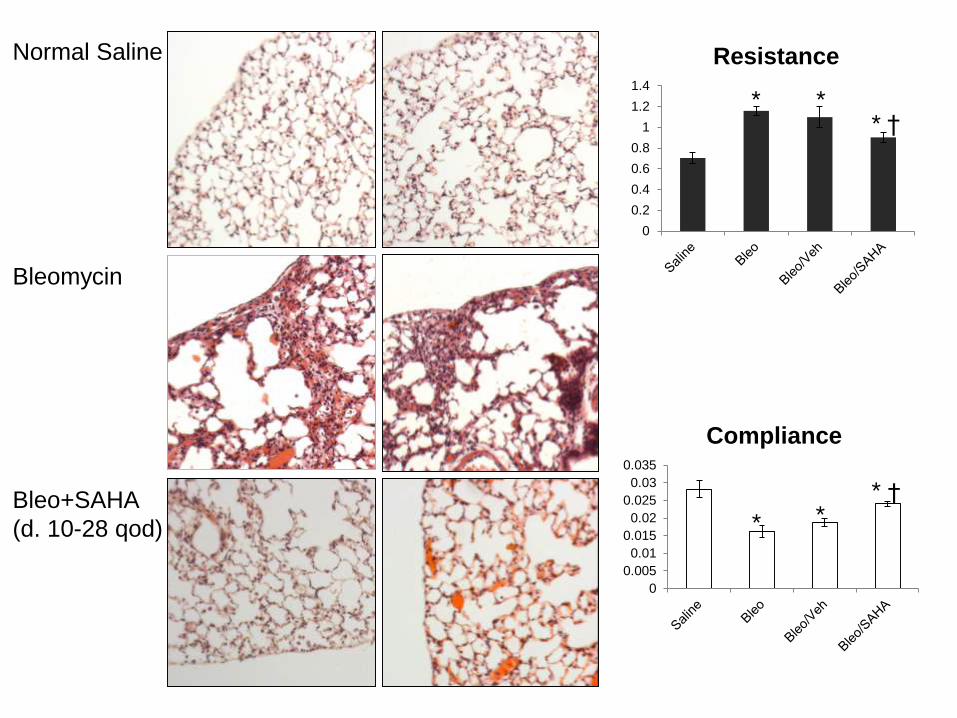
Normal Saline
Bleomycin
Bleo+SAHA
(d. 10-28 qod)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Resistance
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
Compliance
* **
* **
†
†

Next Steps: Sequence-Based Approaches-Potential and Challenges
• Non-CpG methylation, hydroxymethylcytosine(5hMC), 5-methyladenine
• Chromatin modifications (ChIP-Seq) yield much larger datasets
• Limitations of tissue-based studies; dynamic nature of epigenetic alterations
• Understanding hierarchy of epigenetic alterations and “epigenome code”

What is needed: JIA
• Analysis of DNA methylation, histone modifications, miRs, chromatin organization in well-defined samples
• Temporal variation; response to “biologics”
• Interaction of epigenetic paradigms, interaction with genome variants, response to environment
• Mechanisms of epigenomic alteration and targetable “nodes”
• Epigenome as biomarker; especially circulating RNA
• Preclinical models and clinical trials of epigenetic-targeted therapies
• Funding for additional research!







