
^ Í , Z Í ¿ CJ
d Offprint from “M arine Biology”International Journal on Life in Oceans and Coastal Waters, Vol. 12, No. 3, February 1972, Pages 189— 193
© by Spring er-Verlag 1972 • Printed in Germany
Are some bacteria toxic for marine algae ?
B. R. B e r l a n d , D. J . B o n i n and S . Y. M a e s t r i n i
Station m arine d ’Endoum e; Marseille, France
\)<-c
Abstract
I t was observed in vitro th a t some bacteria from different origins, including th e sea, are capable of inhibiting th e growth o f several m arine algae. However, there is too much difference between in vitro experiments and in situ environmental conditions to assume th a t bacterial poisons may rule a great deal o f algae-bacteria relationships in th e sea as a whole. In the au thor’s opinion th e im portance of bacterial poison is lim ited to special and rare instances.
Introduction
There is now a g rea t deal of evidence th a t th e algal and bacterial environm ent in th e sea is no t only ruled by a simple trophic relationship; antibiosis m ay also p lay an im p o rtan t role in m arine ecology.
R ed-tide algal toxins have a ttrac ted atten tion over a long period because of the ir toxic effects on fishes and even m ammals, including m an (review by S h i l o , 1967). Some m arine algae have also been proved capable of antib io tic synthesis against bacteria (S t e e m a n -N i e l s e n , 1955; S i e b u r t h , 1959; B u r k h o l d e r e t ah, 1960; A u b e r t , 1965; D u f f e t al., 1966).
I t has long been recognised th a t sea w ater possesses bactericidal properties. R o s e n f e l d and Z o b e l l (1947) suggest th a t th is m ay be due to the existence of an autochthonous bacterial microflora which produces an tib io tic substances. K r a s i l ’n i k o v a (1964), B u r k h o l d e r e t al. (1966) and G a u t h i e r (1969, 1970) repo rt th e production of antibacterial substances by m arine bacteria.
In regard to organisms isolated from bo th fish and w aste ponds, S h i l o (1967) and S t e w a r t and B r o w n (1969) pointed ou t th a t, in m ixed-plate cultures, the bacteria m ay lyse C yanophyta or Chlorophyta cells.
In our in vitro experim ents we observed th a t the substances released b y some bacteria inh ib it m arine algae grow th un til death ensues.
MaterialThe following algae species were used. Chloro
phyceae: Chlamydomonas m agnusii1', Prasinophyceae:
Prasinocladus m arinus2, Tetraselmis striata-, Bacillariophyceae: Amphora sp ., Asterionella japonica, Chaetoceros lauderi, Hemiaulus sinensis, Nitzschia acicularis, Phaeodactylum tricornutum, Thalassiosira rotula-, Chrysophyceae: Stichochrysis immobilis1; X an thophyceae: Heterothrix sp.3, M onallantus salina1.
All strains were previously obtained in bacteria- free cultures and routinely grown in P r o v a s o l i ’s (1968) ES solution. Tetraselmis striata and Nitzschia acicularis are P r o v a s o l i ’s collection strains (Haskins Laboratories, New-Haven, USA) obtained through the courtesy of Dr. A u b e r t (C.E.R.B.O.M., Nice, France); th e o ther species were isolated from the G ulf of Marseilles.
Several of th e bacterial stra ins which have been tested were obtained from the In s ti tu t P asteur (Paris, France) by courtesy of Dr. B i a n c h i (Station marine d ’Endoum e): Pseudomonas aeruginosa (C.I.P. strain num ber 6352), P . fluorescens (5690), P . maltophilia (5490), P . putida (6323), Vibrio cholerae (6215). Dr. B i a n c h i also provided stra in 558, which is a Pseudomonas very closely related to Pseudomonas pseudo- alcaligenes, and stra in 601, a brevibacterium ; bo th were collected from deep m ud ( B i a n c h i , 1971). Tw enty stra ins were isolated from algae cultures as previously described ( B e r l a n d e t ah, 1969); 8 Pseudomonas, 2 Vibrio, 4 Achromobacter and 6 Flavobacteria were used. Three o ther strains of th e sam e origin, as ye t undeterm ined, were also used (reference num bers 00.27, 00.29, 00.30 of our collection). Two strains (references 26.32 and 26.51) were isolated off th e Gulf o f Fos, near Marseille. These 5 strains are presently being investigated as to their taxonom ic position.
Toxicity of P seu d o m o n a s a eru g in o sa to T e tra se lm is s tr ia ta
Pseudom onads are frequently reported to cause diseases in anim als and p lan ts (S h a c k l e t o n , 1968; S i n d e n and D u r b i n , 1969; H e c k l e y , 1970); we a t tem p ted to discover if they are also toxic to m arine algae. Pseudomonas aeruginosa was chosen, both
1 Determ ined b y Professor P. G a y r a l , Caen.
25 Marine Biology, Vol. 12
2 Determ ined b y Dr. M. J . Ch r e t i e n n o t , Marseille.3 Determ ined b y D r. P. B o u r r e l l y , Paris.
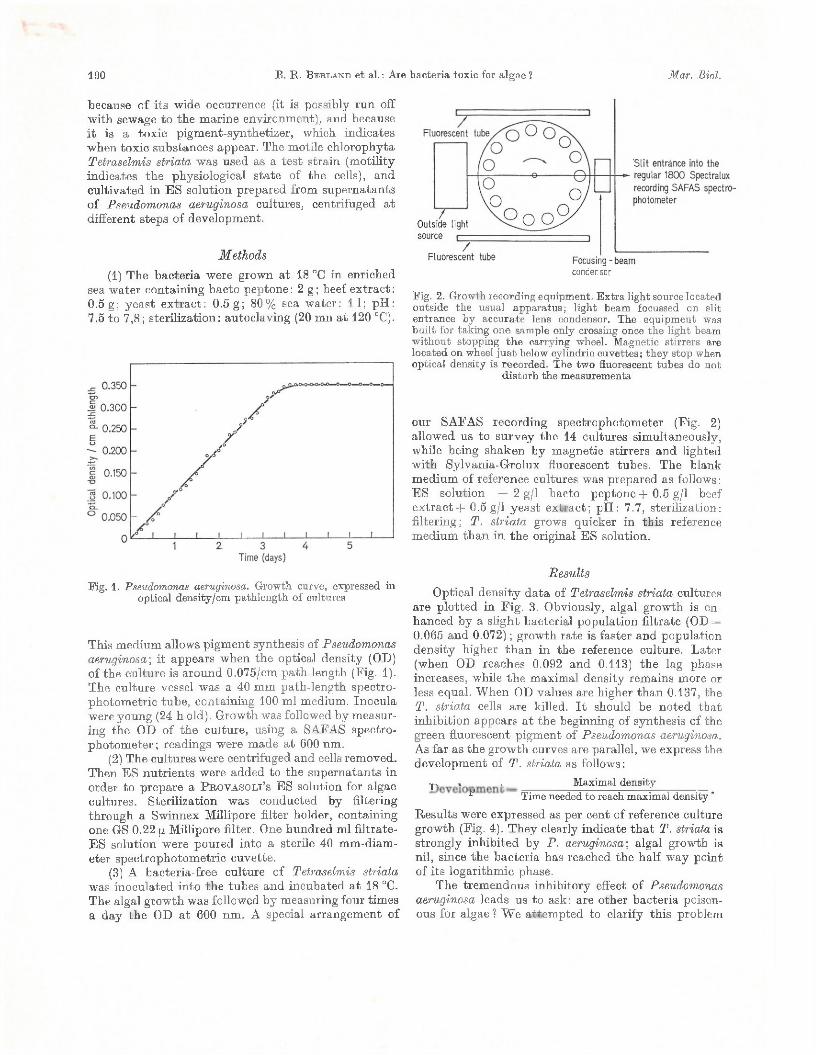
190 B . R . B e r l a n d e t al.: Are bacteria toxic for algae ? M ar. Biol.
because o f its wide occurrence (it is possibly ru n off w ith sewage to the m arine environm ent), and because i t is a tox ic pigm ent-synthetizer, which indicates when toxic substances appear. The motile chlorophyta Tetraselmis striata was used as a te s t stra in (m otility indicates the physiological s ta te of the cells), and cultivated in ES solution prepared from supernatants of Pseudomonas aeruginosa cultures, centrifuged a t different steps of development.
M ethods(1) The bacteria were grown a t 18 °C in enriched
sea w ater containing bacto peptone : 2 g ; beef ex trac t : 0.5 g ; yeast ex trac t: 0.5 g ; 80% sea w ater: 11; pH : 7.5 to 7,8; sterilization: autoclaving (20 mii a t 120 °C).
0.350
.S' 0 .300
0.250
- - 0.200
g 0.150“O
S 0.100
° 0.050
Time (days)
Fluorescent tube
'Slit entrance into the regular 1800 Spectralux recording SAFAS spectrophotometer
Outside light source i—
Fluorescent tube Focusing - beam
P o o
Fig. 1. Pseudomonas aeruginosa. Growth curve, expressed in optical density/cm pathlength of cultures
This m edium allows pigm ent synthesis of Pseudomonas aeruginosa-, i t appears when th e optical density (OD) of the culture is around 0.075/cm path-length (Fig. 1). The culture vessel was a 40 mm path-length spectro- photom etric tube , containing 100 ml medium. Inocula were young (24 h old). Growth was followed by m easuring the OD of the culture, using a SAFAS spectrophotom eter; readings were m ade a t 600 nm.
(2) The cultures were centrifuged and cells removed. Then ES nu trien ts were added to the supernatants in order to prepare a P r o v a s o l i ’s ES solution for algae cultures. Sterilization was conducted by filtering through a Swinnex Millipore filter holder, containing one GS 0.22 ¡a Millipore filter. One hundred ml filtrate- ES solution were poured into a sterile 40 mm-diam- eter spectrophotom etric cuvette.
(3) A bacteria-free culture of Tetraselmis striata was inoculated in to th e tubes and incubated a t 18 °C. The algal grow th was followed by measuring four tim es a day th e OD a t 600 nm . A special arrangem ent of
condensor
Fig. 2. Growth recording equipm ent. E x tra light source located outside th e usual apparatus; light beam focussed on slit entrance b y accurate lens condensor. The equipm ent was bu ilt for taking one sample only crossing once the light beam w ithout stopping th e carrying wheel. Magnetic stirrers are located on wheel ju s t below cylindric cuvettes; they stop when optical density is recorded. The tw o fluorescent tubes do not
d is tu rb th e measurements
our SAFAS recording spectrophotom eter (Fig. 2) allowed us to survey the 14 cultures sim ultaneously, while being shaken by m agnetic stirrers and lighted w ith Sylvania-G rolux fluorescent tubes. The blank m edium of reference cultures was prepared as follows : ES solution + 2 g/1 bacto pep tone-)-0.5 g/1 beef ex trac t + 0.5 g/1 yeast ex trac t; pH : 7.7, sterilization: filtering; T . striata grows quicker in th is reference m edium th an in th e original ES solution.
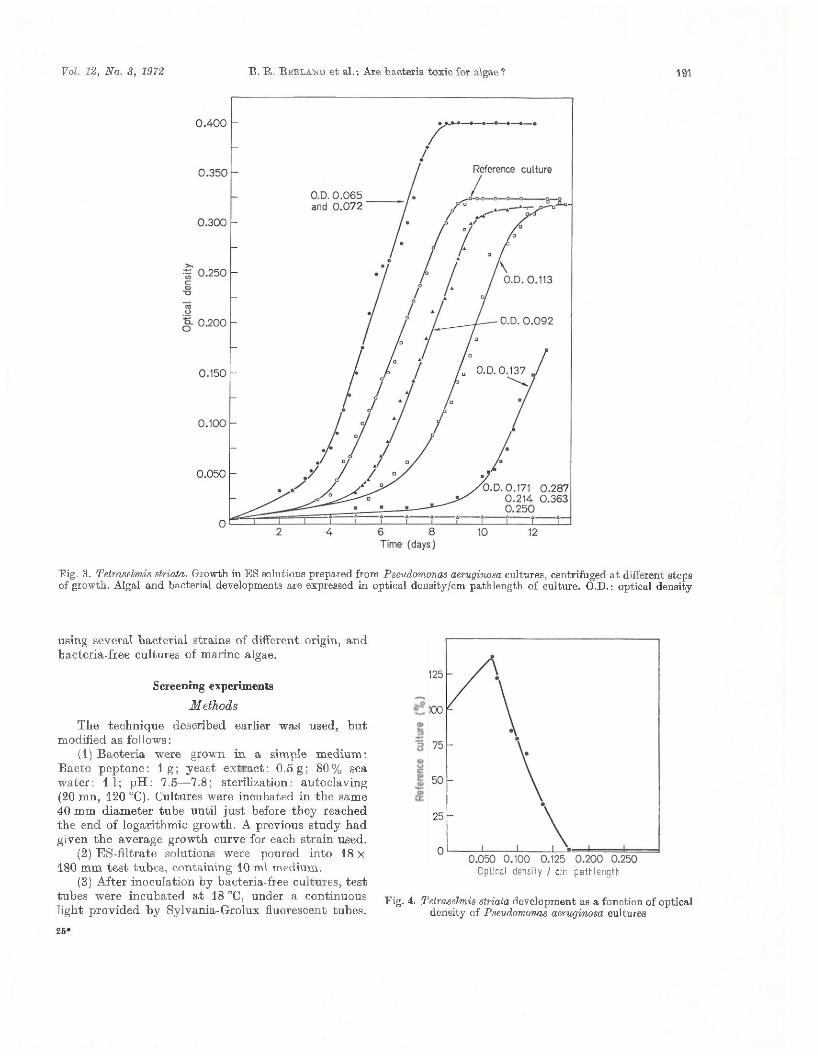
ResultsO ptical density d a ta of Tetraselmis striata cultures
are p lo tted in F ig . 3. Obviously, algal grow th is enhanced by a sligh t bacterial population filtrate (OD =0.065 and 0.072) ; growth rate is faster and population density higher th a n in the reference culture. Later (when OD reaches 0.092 and 0.113) the lag phase increases, while th e maximal density rem ains more or less equal. W hen OD values are higher th an 0.137, the T . striata cells are killed. I t should be no ted th a t inhibition appears a t the beginning of synthesis of the green fluorescent pigm ent of Pseudomonas aeruginosa. As far as the grow th curves are parallel, we express the developm ent of T . striata as follows :
-p. , , __ Maximal density^ Time needed to reach maximal density *
R esults were expressed as per cent of reference culture growth (Fig. 4). They clearly indicate th a t T . striata is strongly inhibited by P. aeruginosa; algal growth is nil, since the bacteria has reached th e half w ay point of its logarithm ic phase.
The trem endous inhibitory effect of Pseudomonas aeruginosa leads us to ask: are other bacteria poisonous for algae ? W e a ttem pted to clarify th is problem
Vol. 12, No. 3, 1972 B. R . B e r l a n d e t a í.: Are bacteria toxic for algae ? 191
0 .4 0 0
Reference culture0 .3 5 0
O.D. 0 .0 6 5 and 0 .0 7 2
0 .3 0 0
~ 0 .2 5 0O.D. 0.113
0.200 O.D. 0 .0 9 2
O.D. 0 .1370.1 50
0.100
0 .0 5 0O.D. 0.171 0 .2 8 7
0 .2 1 4 0 .3 6 3 0 .2 5 0
Time (days)
Fig. 3. Tetraselmis striata. Growth in ES solutions prepared from Pseudomonas aeruginosa cultures, centrifuged a t different steps of growth. Algal and bacterial developments are expressed in optical density/cm pathlength of culture. O.D.: optical density
using several bacterial strains of different origin, and bacteria-free cultures of m arine algae.
Screening experiments
M ethodsThe technique described earlier was used, bu t
modified as follows :(1) B acteria were grown in a simple m edium :
B acto pep tone: l g ; yeast ex trac t: 0.5 g ; 80% sea w ater: 11; p H : 7.5— 7.8; sterilization: autoclaving (20 mn, 120 °C). Cultures were incubated in th e same 40 mm diam eter tube until ju s t before th ey reached the end of logarithm ic growth. A previous study had given th e average grow th curve for each stra in used.
(2) E S-filtra te solutions were poured in to 18 x 180 mm te s t tubes, containing 10 ml medium.
(3) A fter inoculation by bacteria-free cultures, te s t tubes were incubated a t 18 °C, under a continuous fight p rovided by Sy 1 vania-Grolux fluorescent tubes.
125
100
75
50
25
00 .0 5 0 0.100 0.125 0 .2 0 0 0.250
Optical density / cm pathlength
Fig. 4. Tetraselmis striata developm ent as a function of optical density of Pseudomonas aeruginosa cultures
25*

192 B . R . B e r l a n d e t al.: Are bacteria toxic for algae ? M ar. Biol.
A fter a period o f a t least a week, results were read by direct observation, using a 4° scale as follows: complete inhibition (I) — the experim ental tubes are completely clear; inhibition (si) — there is evidence th a t te s t cultures are less deeply coloured th a n reference culture ; no inhibition (0) — tes t and reference cultures are sim ilar; growth enhancem ent (GE) of algae. The blank m edium was prepared as follows: ES solution + 1 g/1 peptone + 0.5 g/1 yeast ex trac t; p H : 7.7; sterilization: filtering.
R esultsAll d a ta of the large-scale survey will be published
la ter, b u t th e prim ary results, sum m arized in Table 1, clearly indicate th a t several bacteria are capable of strong inhibition against algae. Asterionella japonica and Chaetoceros lauderi appear to be dam aged by all th e strains tested . Regarding the bacteria, some strains badly dam age alm ost all the algae, especially Pseudomonas aeruginosa, P . marinoglutinosa and stra in reference 00.30.
Discussion and conclusion
The in vitro experim ents reveal an obvious inhibition of several m arine algae by some bacteria. Pseudomonas aeruginosa and related strains are n o t m arine organisms, b u t they can be transported to th e sea, especially in polluted areas.
In so far as there is no quan tita tive sim ilarity betw een in situ and culture densities of bacterial populations, bacterial poisons m ay only be of some im portance when concentrated, as, for exam ple, in narrow areas of w aters rich in suspended or dissolved organic m atter.
Table 1. List of marine algae tested and number of inhibiting bacteria strains. I : complete inhibition; e l : inhibition
Algal species Number ofbacteriatested
Number of (I)
N um ber o f (el)
Chlamydomonas magnusii 23 6 1Prasinocladus marinus 13 2 4Tetraselmis striata 32 6 3Amphora sp. 20 11Asterionella japonica 6 6Chaetoceros lauderi 5 5Hemiaulus sinensis 19 7 1Nitzschia acicularis 8 2 6Phaeodactylum tricornutum 16 2 1Thalassiosira rotula 1 1Stichochrysis immobilis 20 7 1Heterothrix sp. 20 2 3Monallantus salina 20 4
There is, how ever, no indication a t presen t th a t bacterial toxins a re capable o f playing a role in the algae-bacteria relationships in th e sea as a whole.
Summary
1. I n vitro experim ents indicate th a t th e pigm ented poison of the bacterium Pseudomonas aeruginosa is a strong growth in h ib ito r of th e alga Tetraselmis striata.
2. Several bac te ria stra in s from different origins are recognized to have th e sam e tox icity against various m arine algae.
3. Taking in to account th e very g rea t differences betw een in vitro experim ents and in situ environm ental conditions, i t is no t a t present possible to sta te th a t bacterial poisons really play a role in th e algae- bacteria relationships in the sea.
Acknowledgements. Our thanks are due to D r. P. B o u r - r e l l y (Museum, Paris), Dr. M. J . Ch r e t i e n n o t (Station m arine d’Endoume) and Professor P. G a y r a l (University of Caen) for determ ining several micro-algae; to D r. M. A u b e r t (C.E.R.B.O.M., N ice/ and Dr. A. B i a n c h i (Station marine d ’Endoume) for providing several strains tested.
Literature citedA u b e r t , M. : Le com portem ent des bactéries terrigènes en mer.
Relations avec le phytoplancton. Cah. C.E.R.B.O.M., 19/20, 8—285 (1965).
B e r l a n d , B. R ., M. G. B i a n c h i and S. Y. M a e s t r i n i : Etude des bactéries associées aux algues marines en culture. I. D éterm ination préliminaire des espèces. Mar. Biol. 2, 350—355 (1969).
B i a n c h i , A. : Ecologie e t taxonomie des bactéries hétérotrophes aérobies des sédiments marins. Leur participation à la dégradation des matières organiques. Thèse Dr.-es-Se. Univ. Marseille Luminy, n° A.O. 5223 (1971).
B u r k h o l d e r , P. R ., L. M. B u r k h o l d e r and L. R . A l m a - d o v a r : A ntibiotic activity of some marine algae of Puerto Rico. Botanica m ar. 2 (1/2), 149—156 (1960).
—, R. M. P f i s t e r and F. H . L e i t z : Production o f a pyrrole antibiotic by a m arine bacterium . Appl. Microbiol. 14 (4), 649—653 (1966).
D u f f , D. C. B., D . C. B r u c e and N. J . A n t i a : The antibacterial ac tiv ity of some planktonic algae. Can. J . Microbiol. 12, 877—884 (1966).
G a u t h i e r , M.: Substances antibactériennes produites par les bactéries marines. Première partie: E tude systématique de l’activité antagoniste de souches bactériennes marines vis-à-vis de germes telluriques aérobies. Revue int. Océanogr. méd. 15/16, 41— 59 (1969).
— Deuxième partie : Lipo-polysaccharides antibiotiques produits par certains germes marins appartenan t aux genres Pseudomonas e t Chromobacterium. Revue in ter. Océanogr. méd. 17, 23—45 (1970).
H e c k l e y , R . J . : Toxins of Pseudomonas. Microbiol, toxins 3, 473—491 (1970).
K r a s i l ’n i k o v a , Y e . N. : Antagonism in m arine microorganisms. In : Microbial population of oceans and seas, pp. 1—287, E d . by A. E. K r i s s , I. E . M i s h u s t i n a , N. M i t s k e v ic h and E. V. Z e m t s o v a . Moscow: N auka 1964. (Translated by G. E. F o g g , E. A r n o l d 1967).
P r o v a s o l i , L.: Media and prospects for the cultivation of marine algae. In : Cultures and collections o f algae, pp. 63—75, Ed. by A. W a t a n a b e and A. H a t t o r i . Proc. U .S.A.-Japan Conf. Hakone, Septem ber 1966. Jap . Soc. P lant. Physiol. (1968).

Vol. 12, No. 3, 1972 B . B . B e r l a n d e t a l . : A re b a c t e r i a to x ic f o r a lg a e ? 193
R o s e n f e l d , D. W. and C. E. Z o b e l l : A ntibiotic production by marine microorganisms. J . Bacteriol. 54,393—398 (1947).
S h a c k l e t o n , D. A.: A comparative study o f some plan t pathogenic pseudomonads. N . Z. J . Sei. 11 (2), 236—248 (1968).
S h l l o , M .: Eorm ation and mode of action of algal toxins. B act. Rev. 31 (3), 180—193 (1967).
S i e b t j r t h , J . M a c N. : Antibacterial activ ity o f A ntarctic m arine phytoplankton. Limnol. Oceanogr. 4 (4), 419—424 (1959).
S i n d e n , S. L. and R . D. D u b b i n : Some comparison of chlorosis inducing pseudomonas toxins. Phytopathology 59 (2), 249—250 (1969).
S t e e m a n -N i e l s e n , E .: T he production o f antibiotics by plankton algae and its effect upon bacterial activities in th e sea. Papers in m ar. Biol, and Ocean. Deep Sea. Res. Suppl. 3, 281—286 (1955).
S t e w a b t , J . R . and R . M. B r o w n , J r . : Cytophaga th a t kills or lyses algae. Science N .Y . 164, 1523—1524 (1969).
F i r s t a u t h o r ’s a d d r e s s : D r . B . R . B e r l a n dStation marine d ’Endoume Chemin de la B atterie des Lions Marseille (7e)France
D ate of final m anuscript acceptance: Septem ber 9,1971. Communicated b y J . M. P e r e s , Marseille








