donald n. duvick - university of wisconsin– · pdf filethe contribution of breeding to...
TRANSCRIPT

THE CONTRIBUTION OF BREEDING
TO YIELD ADVANCES IN MAIZE
(ZEA MAYS L.)
Donald N. Duvick
Iowa State UniversityAmes, Iowa 50011
I. I
ntroduction83
Advances in Agronomy, Volume 86Copyright 2005, Elsevier Inc. All rights reserved.
0065-2113/05 $35.00
A
. M aize Yield Trends During the Past Century B . F actors Responsible for Upward Yield TrendsII. G
enetic Gains in Grain Yield of Hybrids A . P reviously Reported Genetic Yield Gains B . R ecent Estimates of Genetic Yield Gains C . E stimates of the Contribution of Breeding to Total Yield Gains D . C hanges that Have Accompanied Genetic Yield Gains in HybridsI
II. G enetic Gains from Population Improvement A . C omparisons with Genetic Gains in Hybrids B . R elative Contributions of Population Improvement andPedigree Breeding
I V. A nalysis and ConclusionsA
. P ossible Reasons for Genetic Yield Gains B . P otential Helps or Hindrances to Future Gains in Yield C . P redictions R eferencesMaize (Zea mays L.) yields have risen continually wherever hybrid maize
has been adopted, starting in the U.S. corn belt in the early 1930s. Plant
breeding and improved management practices have produced this gain joint-
ly. On average, about 50% of the increase is due to management and 50% to
breeding. The two tools interact so closely that neither of them could have
produced such progress alone. However, genetic gains may have to bear a
larger share of the load in future years. Hybrid traits have changed over the
years. Trait changes that increase resistance to a wide variety of biotic and
abiotic stresses (e.g., drought tolerance) are the most numerous, but mor-
phological and physiological changes that promote efficiency in growth,
development, and partitioning (e.g., smaller tassels) are also recorded.
Some traits have not changed over the years because breeders have intended
to hold them constant (e.g., grain maturity date in U.S. corn belt). In other
instances, they have not changed, despite breeders’ intention to change them

84 DONALD N. DUVICK
(e.g., harvest index). Although breeders have always selected for high yield,
the need to select simultaneously for overall dependability has been a driving
force in the selection of hybrids with increasingly greater stress tolerance
over the years. Newer hybrids yield more than their predecessors in unfavor-
able as well as favorable growing conditions. Improvement in the ability of
the maize plant to overcome both large and small stress bottlenecks, rather
than improvement in primary productivity, has been the primary driving
force of higher yielding ability of newer hybrid. # 2005, Elsevier Inc.
I. INTRODUCTION
A. MAIZE YIELD TRENDS DURING THE PAST CENTURY
Maize yields began to rise markedly in many countries during the past
century, first in the United States in the 1930s and then in other parts of the
world in the 1950s and 1960s. For example:
• U.S. yields, level at approximately 1.5 mg ha�1 in the first three decades
of the 20th century, started to rise significantly in the 1930s, reaching
8.5 mg ha�1 by the end of the century (USDA-NASS, 2003b). The U.S.
yield gains averaged 63 kg ha�1 year�1 from 1930–1960 and 110 kg ha�1
year�1 during the next 40 years (Troyer, 2000).
• Maize yields in Canada tripled during the period 1940–2000, increasing
from 2.5 to 7.5 mg ha�1, a linear increase of 80 kg ha�1 year�1 (Bruulsema
et al., 2000).
• Maize yields in Germany doubled in the period 1965–2000, going from
4 to 8 mg ha�1 (Frei, 2000).
• Maize yields in France quadrupled in the period 1950–1984, increasing
from 1.5 to 6.0 mg ha�1 (Derieux et al., 1987).
• In Argentina, the national mean maize yield increased “at a rate of 2.3%
per year from 1970–1992” (Eyherabide et al., 1994).
Table I summarizes yield gain data for several regions of the world during
the period 1961–2002. Globally, maize yields doubled during this time, from
1.9 to 4.3 mg ha�1, a linear increase of 61 kg ha�1 year�1. Different regions
varied in the size of annual gain, as well as in average yields at the beginning
and the end of the interval, but all showed positive and significant gains with
the exception of eastern Europe (highly variable in the past decade) and
southern Africa (minimal gain and highly variable during entire period).
Yields in south Asia did not start to rise significantly until the 1980s; annual
gains since 1985 have averaged 38 kg ha�1 year�1.

Table I
Maize Yield Trends in Selected Regions (1961–2002)a
Region1961 mean
(mg ha�1)
2002 mean
(mg ha�1)
Annual gainb
(kg ha�1 year�1) R2c
European Union (15) 2.5 9.1 169 0.98
USA 3.9 8.2 109 0.83
China 1.2 5.0 103 0.96
Canada 4.6 7.6 69 0.77
World 1.9 4.3 61 0.95
South America 1.4 3.4 48 0.87
Eastern Europe 1.8 4.2 42 0.38
South Asia 1.0 1.7 20 0.78
Southern Africa 0.7 1.3 8 0.26
aFrom FAO Statistical Databases (2004) http://apps.fao.org/default.htm.bLinear regression coefficients, calculated from annual means, 1961–2002.cCoefficient of determination.
ADVANCES IN MAIZE 85
These exampl es and other data show that maiz e y ields have increa sed
signi ficantly in man y regions of the worl d dur ing the latter half of the 20th
century , especi ally in those places wher e maiz e is gro wn as a co mmercial crop.
B. FACTORS RESPONS IBLE FOR UPWARD Y IELD TRENDS
1. Cultural Practi ces
Change s in cultura l practi ces have been respo nsible for a signifi cant
porti on of maiz e yield gains. Crop managem ent practices, such as weed
and pest con trol, timeli ness of plan ting, and increa sed effici ency of harvest
equ ipment, have impro ved over the years, especially (but not exclus ively) in
the indust rialized coun tries (e.g., Car dwell, 198 2; Edm eades and Tollenaa r,
1990 ).
Perha ps most impor tantl y, the use of synthetic nitr ogen fert ilizers
increa sed marke dly starting in the years afte r World War II when plentiful
and affor dable supp lies be came avail able, first in the ind ustrialized coun tries
and then in many (but not all) of the developi ng countri es (e.g ., Car dwell,
1982; Edmeades and Tollenaa r, 1990; Miquel, 20 01 ). Total fert ilizer app lica-
tions on all cro ps worldw ide increa sed fiv efold during the pe riod 1961–199 2.
The linea r increa se started from an average applic ation of about 20 kg ha� 1
in 1962 and reached 105 kg ha� 1 in 1992 ( USDA -ERS, 2003 ). How ever, in
some co untries, app lication amoun ts of synthetic nitr ogen fertilizer did not
fit this gen eral trend; they be gan to level off in the 1980s. Applicati on of

86 DONALD N. DUVICK
commercial nitrogen fertilizer to maize plantings in the United States rose
from an average of 58 kg ha�1 in 1964 to 157 kg ha�1 in 1985, but since then
has stabilized at approximately 145–150 kg ha�1 (Daberkow et al., 2000;
USDA-ERS, 2003). It would seem, therefore, that yield gains of U.S. maize
since the mid-1980s cannot be attributed to application of increasing
amounts of nitrogen fertilizer on maize plantings.
Plant density—the number of maize plants per hectare—also increased
steadily through the years following World War II in the United States as
well as in other countries. The increase was more or less in step with increases
in application amounts of fertilizer nitrogen. In the central U.S. corn belt,
plant density averaged about 30,000 plants hectare�1 (or less) in the 1930s;
it began to increase in the late 1940s and 1950s, reaching about 40,000 plants
hectare�1 in the 1960s, 60,000 plants hectare�1 in the 1980s, and is often
at 80,000 plants hectare�1 or higher at present (Duvick, 1977, 1984a, 1992;
Duvick et al., 2004b; Paszkiewicz and Butzen, 2001; USDA, 1949–1992).
During the past 50 years, plant density in the central U.S. corn belt has
increased at an average rate of about 1000 plants hectare�1 year�1.
2. Plant Breeding
a. Farmer Breeding. Genetic improvements, as well as cultural
improvements, can contribute to an increased yield of maize. Farmer bree-
ders, beginning with the people who first domesticated maize, have selected
plants and cultivars to fit their wants and needs and, in so doing, have
developed thousands of landraces adapted to a multitude of environments,
as well as with a wide range of morphological and quality traits (e.g.,
Goodman and Brown, 1988; Grobman et al., 1961; Paterniani and Good-
man, 1977). We can assume that a higher yield, or at least an acceptable and
dependable level of yield, was always a desired trait for maize cultivars, as
well as for those of other staple grain crops.
Although long-term yield trends are not recorded for specific farmer
breeding programs, a general observation indicates that when crop varieties
are grown in a new environment (e.g., when migrants carry their favorite
cultivars to a new land), the cultivars often do not perform as well as
intended. Careful selection in the unadapted cultivars, often coupled with
hybridization to cultivars from elsewhere, then is used to develop genetically
different cultivars that are better adapted to the new environment and
therefore yield more (and more dependably) than the first introductions.
Examples in U.S. history are the 19th century development of hard red
winter wheat (Triticum aestivum L.) cultivars for Kansas (Malin, 1944) and
“Corn Belt Dent” maize open pollinated cultivars (OPCs) for U.S. corn belt
states such as Illinois and Iowa (Wallace and Brown, 1988).

Figure 1 United States maize yields, annual average, 1900–2003. FromUSDA-NASS (2003a).
ADVANCES IN MAIZE 87
Farmers developed adapted maize OPCs for the U.S. corn belt states in a
relatively short time (Hallauer and Miranda, 1988; Wallace and Brown,
1988). Within a few decades after settlement of the region in the early
years of the 19th century, maize yields and general performance of the new
“Corn Belt Dent” cultivars were at acceptable levels in most parts of the
region. However, from then on, gains in yield were small or nonexistent.
This is evidenced by the lack of gain in U.S. maize yields during the first
three decades of the 20th century (Fig. 1).
One could suppose that the lack of yield gain during those decades was
because maize-growing areas in the country changed in location and extent
over time and therefore were not always equivalent in productivity. How-
ever, in the states of Iowa and Illinois, where maize-growing areas and
cultural practices were relatively constant during this period, yields were
essentially unchanged also. Yields in those states were level at approximately
2.3 mg ha�1 during the years 1900–1930 (USDA-NASS, 2003a). It would
seem that farmer breeders in the corn belt, using selection techniques of that
time [primarily mass selection based on individual plant performance
(Sprague, 1952)], could not raise maize yields further than the levels attained
in the initial development of adapted cultivars.
New breeding methods were tried in the late 19th and early 20th centuries.
The production of varietal hybrids (first generation crosses of two maize
OPCs) was tried and abandoned because of unreliable results (e.g., Crabb,
1993; Richey, 1922). A few professional breeders in the public sector
(USDA) worked on variety improvement in the 1920s using relatively

88 DONALD N. DUVICK
unsophisticated methods of mass selection or ear-to-row breeding (Crow,
1998; Russell, 1991; Sprague, 1946, 1994). Their efforts did not increase
yields either, except when a program provided adaptation to a new environ-
ment. These breeders, working in the first decades of the 20th century, lacked
access to the present-day knowledge of experimental design, statistical anal-
ysis, and quantitative genetics. Lack of these tools must have hindered their
progress.
b. Hybrids. U.S. maize yields started to increase when maize hybrids
made from crosses of inbred lines were introduced in the early 1930s. Du-
ring the next few years the increase in maize yield was correlated with the
increase in the proportion of maize area planted to hybrids (USDA,
1944 – 1962; USDA-NASS, 2003a). Yields in Iowa increased from 2 mg
ha�1 to 3.5 mg ha�1 in the period 1933–1943, as the percentage of maize
area planted to hybrids went from 0.7 to 99%. U.S. maize yields rose from
1.5 mg ha�1 in 1933 to 2.4 mg ha�1 in 1950, as the percentage of area planted
to hybrids went from 0.1 to 78%. In either case, yield gains took place before
a significant increase in use of synthetic nitrogen fertilizers or chemical
control of weeds and insects (Cardwell, 1982; USDA, 1956), so it seems
likely that the yield gains primarily were caused by genetic improvements;
the new hybrids yielded more than the OPCs that they replaced, and succes-
sive hybrids yielded even more.
Maize yields began to rise in conjunction with the introduction of hybrids in
other countries as well (Cunha Fernandes and Franzon, 1997; Derieux et al.,
1987; Eyherabide et al., 1994; Frei, 2000; Tollenaar, 1989), although, as in the
United States, improved crop management techniques usually accompanied
the introduction of hybrid maize; plant breeding and crop management jointly
contributed to the sharp increases in maize yields. The proportion of gain
attributed to genetic improvements is treated in more detail in later sections,
with emphasis on hybrids and how sequential changes in their breeding and
genetics have contributed to increased on-farm yield.
c. Improved Populations. In the United States, the first hybrids were
made from inbreds that had been developed by selfing some of the better
OPCs of the 1920s. Breeders then worked to develop a second generation of
improved hybrids using new inbreds made by selfing the same OPCs. They
found that the second round of hybrids yielded little or no more than the
first; it seemed that breeders must have selected most of the superior geno-
types in the initial round of selfing in the OPCs. Some of the breeders
conjectured that it might be possible to make new “synthetic” OPCs, with
a potential for production of a superior second generation of inbred lines, by
intercrossing some of the best inbreds from the first round of OPC selfing
(Baker, 1990).

ADVANCES IN MAIZE 89
To this end, the breeders made several “synthetics” by intercrossing
the better inbreds of the day. Research in maize quantitative genetics
had begun by this time, and some of the populations were subjected to
various kinds of selection to make genetic improvements in the popula-
tions as such. The selection procedures were based on various assumptions
about gene action and genetic variability (Hallauer and Miranda, 1988;
Sprague, 1946, 1966). The Iowa State University Stiff Stalk Synthetic
(BSSS) (Eberhart et al., 1973; Sprague, 1946) is one of the best known of
these populations. Sprague (1946) lists the 16 progenitor inbred lines of this
synthetic.
Breeders practiced population improvement on other kinds of popu-
lations as well, such as locally adapted OPCs, exotic landraces, or compo-
sites of exotic landraces and/or inbred lines (e.g., Hallauer and Miranda,
1988; Sriwatanapongse et al., 1985). The name “recurrent selection” was
coined (Sprague, 1952) to distinguish these kinds of population improve-
ments from pedigree breeding (i.e., developing improved inbred lines from
crosses of proven inbreds). Depending on the prospective end user, breeders
intended to develop improved populations that would serve as sources of
superior inbred lines or that could be used directly as productive cultivars
per se. Results of their work are discussed in a later section.
II. GENETIC GAINS IN GRAIN YIELD OF HYBRIDS
A. PREVIOUSLY REPORTED GENETIC YIELD GAINS
Russell (1991) has summarized 16 independent estimates of genetic
yield gains of sequentially released maize hybrids. Most of the estimates
are based on comparisons of U.S. hybrids and were reported at intervals
during the 20-year period of 1971–1991. Estimates ranged from 25–92 kg
ha�1 year�1 with a mean of 57 kg ha�1 year�1. It seems likely that the wide
range in values was caused, in part, by differing growing conditions among
the several investigations and consequent differential interactions with old or
new genotypes. Other factors, as well, might explain some of the variation,
as follows:
• Breeding might have been less effective in some regions (such as those with
erratic and often severe abiotic stress) than in others.
• Choice of the time series of hybrids for comparison could have had major
effect on size of measured gain. For example, a short time series might
show less improvement than a long one if the short time series happened to
sample a period with small genetic gain.

90 DONALD N. DUVICK
• Plant density of trials could affect the results differentially; older hybrids
could have been disadvantaged if the plant density at which their yield
was measured was higher than that for which they were bred or new
hybrids could be disadvantaged if the density was below that which they
required for maximum yield.
• Harvest technology could be another source of difference; e.g., combine
harvested trials (as compared with hand-harvested trials) could under-
estimate yields of older hybrids if combines failed to pick up all downed
stalks (and ears) of older hybrids with poor standability. Conversely,
hand-harvested trials could overestimate yields of the older hybrids if a
standard shelling percentage was used to convert ear corn weight to grain
weight instead of shelling the ears and weighing the shelled grain. Use of a
standard shelling percentage could inflate estimates of grain weight on
poorly pollinated nubbins of the older hybrids.
However, these possibilities must remain conjecture. The salient fact is
that all of the experiments listed by Russell showed positive and linear
genetic yield gains, fluctuating around a mean of about 60 kg ha�1 year�1.
Additional estimates of genetic gain in hybrids have been made since
Russell’s review and are summarized in the following section.
B. RECENT ESTIMATES OF GENETIC YIELD GAINS
1. Argentina
Elite experimental maize hybrids tested in 154 regional trials in the
Argentine corn belt during the 1979–1991 period had an estimated linear
genetic gain of 105 kg ha�1 year�1 (Eyherabide et al., 1994). Estimates were
based on comparisons with a common check.
A second series of estimates extended the period (1979–1998) and showed
an estimated genetic gain of 107 kg ha�1 year�1, or 2.9% year�1. Further
analysis of these data indicated that gains were not linear during the entire
period; gains were greater in the second decade than in the first, perhaps in
part because of the introduction of single cross hybrids in the 1990s (Eyher-
abide and Damilano, 2001).
2. Brazil
Analysis of 30 years of national maize trials in Brazil (1963–1993) indi-
cates linear genetic progress of 123 kg ha�1 year�1 (Cunha Fernandes and
Franzon, 1997). Trials were grown in three locations, and estimates were
based on comparisons with a moving base of common entries.

ADVANCES IN MAIZE 91
3. United States
Duvick (1997), updating previous reports for hybrids adapted to central
Iowa in the U.S. corn belt, stated that a time series of hybrids and one OPC
representing the period from 1930–1991 showed a linear gain for grain yield
of 74 kg ha�1 year�1. The estimate was based on data from trials comparing
36 hybrids and one OPC, conducted over a period of 4 years, three locations
per year, at three plant densities per location. Yield for each hybrid was for
its “optimum density” in the trials: the plant density at which it gave its
highest mean yield.
A further update extended this time series through the year 2001; it
showed an estimated linear gain of 77 kg ha�1 year�1 (Duvick et al.,
2004b). This estimate applied “best linear unbiased predictors” (BLUPs) of
hybrid grain yield; it was based on trials of 51 hybrids and four OPCs grown
at three plant densities in the years 1991–2001, using yield of each hybrid at
its “optimum density” as described earlier.
C. ESTIMATES OF THE CONTRIBUTION OF BREEDING
TO TOTAL YIELD GAINS
Russell (1991) listed 14 estimates of genetic yield gain of hybrids as
percent of total yield gain. (Total yield gain was defined as on-farm gain
for appropriate regions during the time span of hybrids that were com-
pared.) Most of the comparisons were for the U.S. corn belt but the list
also included estimates for Ontario (Canada), France, and Yugoslavia.
Estimates of genetic gain varied from 29 to 94%, with a mean of 66%.
As noted by Russell (1991), several reasons can be advanced to show that
this broad variability could be caused by inconsistencies in planning and
executing the experiments, such as machine harvest vs hand harvest, or
whether experimental estimates of genetic gain were adjusted to on-farm
state averages. Nevertheless, all estimates agree in showing that hybrid maize
breeding (i.e., genetic improvement) has played a major part in raising
maize yields.
Among the reports of genetic gain since Russell’s summary (reviewed in
Section II.B), Cunha Fernandes and Franzon (1997) estimated that 57% of
total gain in yield in Brazil was due to genetics. The other reports did not
contain such an estimate, but further examination of data in Duvick et al.
(2004b) provides an estimate of 51% for the contribution of genetics, when
trial yields are adjusted to the equivalent of average on-farm yields for Iowa
during the period 1930–2001.
Based on these and earlier estimates, one can state that hybrid maize
breeding during the past six or seven decades has been responsible for 50 to

92 DONALD N. DUVICK
60% of the total on-farm yield gain. However, one also must acknowledge
that the interaction between breeding and management (cultural practices) is
such that neither tool could have caused the gains without aid of the other;
changes in breeding and management continually have interacted in positive
fashion.
D. CHANGES THAT HAVE ACCOMPANIED GENETIC YIELD GAINS
IN HYBRIDS
Breeders have noted that genetic gains in grain yield of hybrids may be
accompanied by changes in other traits, sometimes as a result of direct
selection, sometimes without direct intention by the breeders. And some
traits have stayed essentially unchanged over the generations. Three reviews
in the previous decade (Edmeades and Tollenaar, 1990; Russell, 1991;
Tollenaar et al., 1994) have given detailed accounts of such changes and
are recommended as sources of information and informed commentary on
the topic prior to the early 1990s. The following sections update those
accounts, as well as provide summaries and commentary for some of the
earlier research reports.
1. Plant and Ear Traits
a. Plant and Ear Height. Plant and ear height were reduced in the
second era but not thereafter in a study of single cross hybrids representing
U.S. corn belt hybrids of three eras: 1930s, 1950s, and 1970s (Meghji et al.,
1984). A study of 28 hybrids and four OPCs adapted to Iowa, representing
seven decades culminating in the 1980s, found no trend to reduction in plant
height but a continuing trend to reduction in ear height (Russell, 1984).
Plant height for a 1930–2001 time series of 51 hybrids and four OPCs
adapted to central Iowa likewise was essentially unchanged over the years,
but ear height showed a weak trend toward reduced height, approximately
�3 cm decade�1 (Duvick et al., 2004b).
b. Leaf Angle. Leaves became more upright in the 1970s era in a com-
parison of single crosses representing U.S. corn belt hybrids of three eras:
1930s, 1950s, and 1970s (Meghji et al., 1984). Leaf orientation below and
above the ear became more upright with time in a study of U.S. Midwestern
hybrids representing the decades from 1930–1970 (Crosbie, 1982; Russell,
1991). The trend to upright leaf orientationwas greatest above the ear. Russell
(1991) stated that the distinct increase in upright leaf orientation in the1970s
decade was probably because inbred B73, with upright leaves for its time, was

ADVANCES IN MAIZE 93
a parent in the set of 1970s single cross hybrids. The previously mentioned
1930–2001 time series of 51 hybrids and four OPCs for central Iowa (Duvick
et al., 2004b) showed a similar trend toward more upright leaf habit. Ratings
(as scores) in this study were for the entire plant.
c. Tassel Size. Tassel weight was least in the most recent era in com-
parisons of single cross hybrids representing U.S. corn belt hybrids of the
1930s, 1950s, and 1970s; tassel branch number decreased consistently over
the eras (Meghji et al., 1984). Tassel branch number and tassel weight
decreased over time in a 1930 to 1991 time series of hybrids for Iowa
(Duvick, 1997). Tassel branch numbers in the series averaged 2.5 fewer
branches per decade, and tassel dry weight declined, on average, 0.5 g per
decade. Reduction of tassel size continued in hybrids released during the
next 10 years, as evidenced by scores for tassel size of hybrids in the
1930–1991 time series extended to 2001 (Duvick et al., 2004b).
d. Leaf Number. Number of leaves per plant neither increased nor
decreased in a 1930–1991 time series of Iowa hybrids and OPCs (Duvick
et al., 2004b). Leaf number increased from 12.2 in the 1930s to 13.8 in the
1970s in comparisons of single cross hybrids representing U.S. corn belt
hybrids of the 1930s, 1950s, and 1970s (Meghji et al., 1984).
e. Leaf Area Index (LAI). Russell (1991) suggests that changes in LAI
“may be specific to the particular cultivars used, rather than a general
occurrence of all germplasm representative of similar eras.” This statement
is borne out by the lack of consistent trends across experiments conducted by
different researchers. LAI tended to be higher for recent hybrids than for
older ones in a time series of four hybrids grown in Ontario (Canada) from
1959 to 1989 (Dwyer et al., 1991; Tollenaar, 1991). In another investigation,
LAI increased over time in a set of eight maize hybrids that were commer-
cially important in central Ontario between 1959 and 1988 (Tollenaar, 1989).
However, a set of Iowa hybrids (20 single cross hybrids) representing the
decades of 1930–1970 showed no obvious trend (Crosbie, 1982; Russell,
1991), and a 1930–1991 time series of 36 commercial hybrids and one OPC
for Iowa also showed no change in LAI over time (Duvick, 1997).
f. Leaf Rolling. Leaf rolling of plants, especially in the vegetative stage,
is often seen when plants are subjected to drought. Leaf rolling consistently
increased in newer hybrids in a set of 18 commercial hybrids adapted to
central Iowa and representing the period 1953–2001 when the hybrids
(grown in a rain-free environment in Chile) were compared with and without
managed drought stress at various stages of development (Barker et al., 2005;
Edmeades et al., 2003). Commenting on this observation, Barker et al.

94 DONALD N. DUVICK
(2005) said “Apparently elite hybrids can reduce radiation interception and
water use by leaf rolling, while generating sufficient assimilate flux to the ear
to set adequate kernel numbers and conserving water for later in the season.”
g. Staygreen. Staygreen, also called delayed leaf senescence, or resis-
tance to premature death from unidentified causes, is consistently improved
in newer hybrids (Crosbie, 1982; Duvick et al., 2004b; Meghji et al., 1984;
Russell, 1991; Tollenaar, 1991). The improvement in each of these trials was
greatest under environmental stress such as that induced (or accentuated) by
high plant density. Staygreen improved over time in a set of 18 commercial
hybrids adapted to central Iowa and representing the period 1953–2001
when the hybrids, grown in a rain-free environment in Chile, were (a) well
watered, (b) subjected to induced drought at flowering time, or (c) subjected
to drought during the final grain-fill period (Barker et al., 2005). Genetic
gain for staygreen was greatest in the well-watered treatment and was least
under drought imposed during the grain-fill period.
h. Tillers. Number of tillers per 100 plants varied from hybrid to
hybrid, but decreased slightly on average in a 1930–1991 time series of
hybrids and OPCs for Iowa (Duvick et al., 2004b).
i. Anthesis. Date of anthesis varied among decades, but showed no
trend toward earlier or later dates in an Iowa series of four OPCs and
24 single cross hybrids representing the 1930s through 1980s (Russell,
1985). Similarly, heat units from planting to anthesis varied among hybrids
but showed no general trend toward earlier or later in a 1930–2001 time
series of 51 hybrids and four OPCs adapted to central Iowa (Duvick et al.,
2004b).
j. Silk Emergence. Silk emergence date trended toward earliness in an
Iowa series of four OPCs and 24 single cross hybrids representing the 1930s
through 1980s (Russell, 1985) and also in a 1930–1991 time series of 36
hybrids and one OPC for Iowa (Duvick, 1997). In the latter case, there was
little or no trend to an earlier silk date in absence of stress, such as at low
plant density, whereas higher plant density accentuated the trend, not be-
cause the new hybrids had earlier silking dates, but rather because the stress
of high plant density delayed silk emergence of the older hybrids.
k. Anthesis-Silking Interval (ASI). Anthesis usually precedes silk
emergence, and the interval between the two events is called the anthesis-
silking interval. (The term “silk delay” is also used.) ASI was unchanged in
hybrids representing earlier decades but was significantly shorter in hybrids

ADVANCES IN MAIZE 95
of the 1970s in a set of Iowa hybrids (20 single cross hybrids) representing
the decades of 1930–1970 (Crosbie, 1982). ASI became shorter in each
decade except the 1980s in an Iowa-adapted series of four OPCs and 24
single cross hybrids representing 1930–1980 (Russell, 1985). ASI was signifi-
cantly shorter in each interval in comparisons of single cross hybrids repre-
senting U.S. corn belt hybrids of the 1930s, 1950s, and 1970s (Meghji et al.,
1984). ASI showed a highly significant linear trend to shorter intervals in a
1930–2001 time series of 51 hybrids and four OPCs adapted to central Iowa
(Duvick et al., 2004b). The trend was greater in trials grown at higher plant
densities. ASI became shorter over time in a set of 18 commercial hybrids
adapted to central Iowa and representing the period 1953–2001 (Barker
et al., 2005; Edmeades et al., 2003). This trend was exhibited when the
hybrids were well watered and was accentuated when they were subjected
to induced drought at flowering time. The experiment was conducted in a
rain-free environment in Chile.
l. Ears per Plant. Both total and harvestable ears per plant increased
over the decades in a set of Iowa hybrids (20 single cross hybrids) representing
the decades of 1930–1970 (Crosbie, 1982). A 1930–2001 time series of 51
hybrids and four OPCs adapted to central Iowa showed a highly significant
trend toward more ears per 100 plants (+3.6 ears decade�1) (Duvick et al.,
2004b). However, ears per plant showed no change over the decades in an
Iowa-adapted series of four OPCs and 24 single cross hybrids representing
1930–1980 (Russell, 1985). The Russell experiment was planted at a single
density (51670 plants ha�1), whereas data for the other two experiments were
expressed as means of three densities in which the medium and high densities
were higher than optimum for the older hybrids and, therefore, were more
likely to cause barrenness in those hybrids. The end result would be a trend
towardmore ears per 100 plants (i.e., fewer barren plants) in the newer hybrids.
m. Grain-Filling Period. Newer hybrids had a longer period of grain
fill, calculated as time from silk emergence to black layer (physiological
maturity), in observations of four discrete time series of hybrids adapted to
the Midwestern United States (Cavalieri and Smith, 1985; Crosbie, 1982;
Meghji et al., 1984; Russell, 1985). The newer hybrids flowered at approxi-
mately the same time as the older hybrids, and although their grain-fill
periods were longer, they also exhibited a faster final dry-down rate, and
so had little or no delay in relative maturity (based on grain moisture
percentage at harvest time). The newer hybrids thus have more time to
devote to grain fill; they exhibit increased efficiency in use of the growing
season in the U.S. Midwest, which is limited on either end by average date of
last frost in spring and first frost in autumn.

96 DONALD N. DUVICK
n. Kernel Weight. Weight per 300 kernels increased in each decade
except the final one in an Iowa-adapted series of four OPCs and 24 single
cross hybrids representing seven eras from pre-1930s through the 1980s
(Russell, 1985). Kernel weight did not change significantly over the decades
except for a significant increase in the final (1970s) decade in comparisons of
20 single cross hybrids representing each decade from 1930–1970 (Crosbie,
1982). Weight per 100 kernels increased linearly in an Iowa-adapted series of
36 hybrids and one OPC representing the years 1930–1991, while number of
kernels per ear decreased slightly but not significantly (Duvick, 1997).
Weight per kernel exhibited a marked linear increase under well-watered
conditions and also under drought stress at flowering, early, and midfill
stages, but showed little or no increase under drought in late-fill and terminal
stages in a set of 18 Iowa-adapted hybrids (evaluated in Chile) representing
the period 1953–2002 (Barker et al., 2005; Edmeades et al., 2003). In ap-
praisal of the results summarized in this section: the general trend to
increased weight per kernel (and no increase in number of kernels per ear,
in the one series where this was measured) may indicate that assistance in
achieving genetic yield gain over time (and also gain in yield stability) is
given more efficiently by plants with increased kernel weight than by plants
with increased kernel number.
o. Grain Protein Percentage. Grain protein percentage declined con-
sistently in a series of 36 hybrids and one OPC for central Iowa spanning
the period 1930–1991 (Duvick, 1997). The loss averaged 0.3% protein dec-
ade�1, with a series mean of 9.8% protein.
p. Grain Starch Percentage. Grain starch percentage increased consis-
tently ina seriesof 36hybrids andoneOPCfor central Iowa spanning theperiod
1930–1991 (Duvick, 1997).The increaseaveraged0.3%starchperdecade,witha
series mean of 70.4% starch. Because production of starch requires less energy
than production of protein, selection for yield without attention to protein or
starch percentagemay have indirectly selected genotypeswith less grain protein
and more grain starch, giving a net increase in efficiency of grain production.
q. Harvest Index (HI). HI did not change consistently over time in
comparisons of single cross hybrids representing U.S. corn belt hybrids of
the 1930s, 1950s, and 1970s (Meghji et al., 1984). HI did not change consis-
tently in a set of 20 single cross hybrids representing Iowa hybrids of the
decades of the 1930s through 1970s (Crosbie, 1982) or in a series of nine
hybrids representing three decades (1959–1988) of maize production in
Ontario when hybrids were compared at optimum plant density for yield
for each hybrid (Tollenaar, 1989). HI did not change consistently over the
decades in an Iowa-adapted series of four OPCs and 24 single cross hybrids

ADVANCES IN MAIZE 97
representing 1930–1980 (Russell, 1985). HI improved on average to a small
degree in successive hybrids in a 1934–1985 time series of Iowa-adapted
hybrids (Duvick et al., 2004b) and in the same series extended to 1991
(Duvick, 1997). Higher plant density accentuated the trend. However, the
superior HI of the newer hybrids at higher plant densities was not because of
increased HI per se in the newer hybrids, but because of reduced HI in the
older hybrids—the result of barrenness induced by stress. At higher densities
the older hybrids maintained plant size but lost yield because of increased
percentages of barren or partially barren plants.
2. Resistance to Root Lodging
Hybrids improved over the years in resistance to root lodging in a series of
Iowa hybrids representing the decades 1930–1960 (Russell, 1974). However, a
later examination of a longer series of hybrids (1930–1980) showed no signifi-
cant improvement in root lodging resistance, although all hybrids were decid-
edly more resistant than the OPCs of the 1920s (Russell, 1984). Four
examinations of a successively lengthened time series of commercial hybrids
for central Iowa showed linear increases in resistance to root lodging, in each
examination (Duvick, 1977, 1984a, 1997; Duvick et al., 2004b). The four
experiments contained hybrids released in the years 1939–1971, 1934–1978,
1934–1991, and 1934–2001, respectively. However, in two other trials of this
series (for hybrids released in 1934–1989 and 1934–2000), improvement of
root strength ceased in the final decade at about the 95% nonroot-lodged level
(Duvick, 1992; Duvick et al., 2004a).
The intensity of root lodging in a given trial can influence the differentia-
tion between the older and the newer hybrids. Low levels of lodging (e.g.,
because of lack of the right combination of rain and wind or because of
insufficient plant density) will make it impossible to differentiate among the
more resistant hybrids because all will be in the range of 95 to 100% upright.
Also, from time to time new hybrids might be released with less resistance to
root lodging than intended, following which genetic improvements are
implemented in newer releases. Either of these conditions (abiotic or genetic)
could explain why apparent cessation of linear improvement in one time
series is followed by a later time series in which gains are linear.
3. Resistance to Stalk Lodging
Lodging resistance (not differentiated between root or stalk lodging)
improved significantly in a series of hybrids grown in France from
1950–1985 (Derieux et al., 1987). Higher plant densities accentuated the
difference between older and newer hybrids.

98 DONALD N. DUVICK
Stalk lodging resistance improved consistently in a time series of Iowa-
adapted hybrids representing 1930–1960 (Russell, 1974). However, in a
longer series (1930–1980), resistance to stalk lodging improved until the
1970s but did not improve in the 1980s (Russell, 1984).
Five different examinations of a successively lengthened time series of
commercial hybrids for central Iowa showed linear improvements in resis-
tance to stalk lodging (Duvick, 1977, 1984a, 1992, 1997; Duvick et al., 2004b).
The five series contained hybrids released in 1939–1971, 1934–1978,
1934–1989, 1934–1991, and 1934–2001. However, in one other trial for this
series (hybrids released from 1934–2000), improvement ceased in the final
decade at about the 95% nonstalk-lodged level (Duvick et al., 2004a).
So for both root lodging and stalk lodging, one can conclude that al-
though the overall trend is toward improved resistance to lodging, improve-
ment may seem to stop from time to time. Additional experiments involving
further extensions of the time series will be needed to test the validity of such
conclusions.
4. Tolerance to Abiotic Stress
a. High Temperatures. A 1930–1991 time series of 36 hybrids and one
OPC for Iowa exhibited a linear increase in grain yield in a low yield season
with a “hot and dry” summer, as well as in two highly favorable (exception-
ally high yield) seasons (Duvick, 1997). Weather records for the “hot and
dry” year (1991) indicate that temperatures during the flowering period were
higher than normal and precipitation was exceptionally low (Iowa State
University, 2003).
b. Low Temperatures. The aforementioned 1930–1991 time series also
exhibited a linear increase in grain yield in a “very cool and wet season”
(Fig. 2) (Duvick, 1997; Duvick et al., 2004b). Weather records for that trial
year (1993) indicate that precipitation amounts during the summer months
were at record-breaking high levels, and daytime high temperatures were
well below normal (Iowa State University, 2003).
Dwyer and Tollenaar (1989) stated that “genetic improvement in the
response to cold stress . . . has significant consequences for yield of field-
grown maize, since many Canadian seasons are subject to short seasons or
cool growing periods.” They showed (for a series of eight hybrids, released
during the years 1959–1988) that reduction in photosynthetic response to
irradiance (PRI) following a cold period during kernel fill was greater in
older than in newer hybrids. A subsequent study (Tollenaar et al., 2000)
showed similar results for a series of eight U.S. maize hybrids representing
the 1930s, 1950s, 1970s, and 1990s.
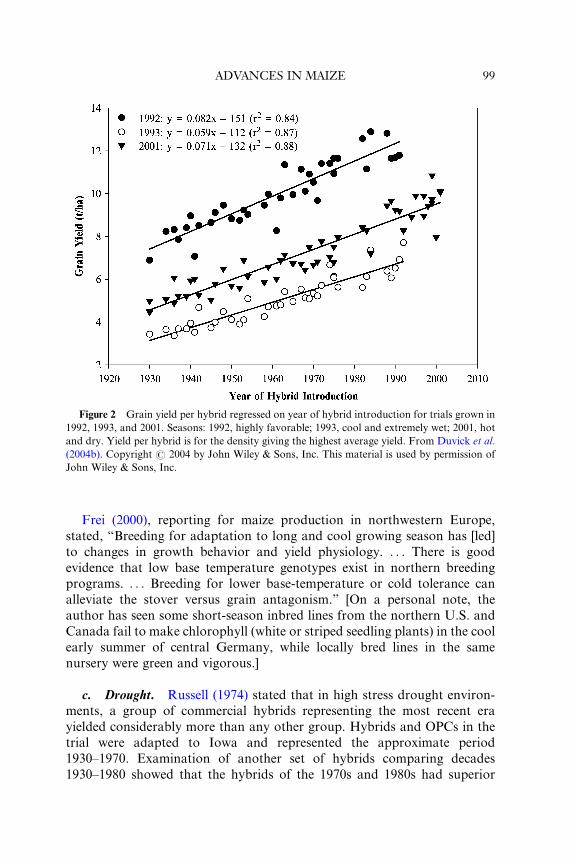
Figure 2 Grain yield per hybrid regressed on year of hybrid introduction for trials grown in
1992, 1993, and 2001. Seasons: 1992, highly favorable; 1993, cool and extremely wet; 2001, hot
and dry. Yield per hybrid is for the density giving the highest average yield. From Duvick et al.
(2004b). Copyright # 2004 by John Wiley & Sons, Inc. This material is used by permission of
John Wiley & Sons, Inc.
ADVANCES IN MAIZE 99
Frei (2000), reporting for maize production in northwestern Europe,
stated, “Breeding for adaptation to long and cool growing season has [led]
to changes in growth behavior and yield physiology. . . . There is good
evidence that low base temperature genotypes exist in northern breeding
programs. . . . Breeding for lower base-temperature or cold tolerance can
alleviate the stover versus grain antagonism.” [On a personal note, the
author has seen some short-season inbred lines from the northern U.S. and
Canada fail to make chlorophyll (white or striped seedling plants) in the cool
early summer of central Germany, while locally bred lines in the same
nursery were green and vigorous.]
c. Drought. Russell (1974) stated that in high stress drought environ-
ments, a group of commercial hybrids representing the most recent era
yielded considerably more than any other group. Hybrids and OPCs in the
trial were adapted to Iowa and represented the approximate period
1930–1970. Examination of another set of hybrids comparing decades
1930–1980 showed that the hybrids of the 1970s and 1980s had superior

100 DONALD N. DUVICK
yiel ds in all environm ents, which included two dro ught-stre ss locations an d
two high-y ieldin g en vironm ents ( Russell, 1991 ).
Com pariso n of a series of U.S. corn belt hybrids and OPC s repre-
senti ng the de cades 1930–198 0 showe d linear gains in grain yield unde r
eithe r dro ught stre ss or irrigat ed treatment s (Cas tleberry et al ., 1984 ). The
mean yield of the 1930s group was equal to 60% of the mean yield of
the 1980s grou p when both groups were subject ed to drough t stress, an d
63% of the mean y ield of the 1980s grou p when both gro ups were g iven full
irrigat ion.
In Onta rio, Canada , a ne wer hyb rid was more toleran t of sh ort dro ught
periods than an older hy brid (Dw yer et al ., 1992 ). Duri ng a dro ught period,
the ne wer hyb rid continued phot osynthes is for about 2 h longer than the
older one before star ting to decline . A furt her study indica ted that the tw o
hyb rids might adopt different mechani sms to tolerate mois ture stre ss
( Nissank a et al., 1997 ). The newer hy brid maint ained relative ly higher
rates of photo synthesis an d trans piration at a lower stem wate r potential.
Altho ugh one canno t con sider a compari son of only two hy brids as a “ time
seri es” that demonst rates trends over the years, exami nations like these
can give hints of pos sible trends and suggest profitabl e fie lds for futur e
invest igation.
Derie ux et al . (1987) , compari ng 33 maize hybrids (of three maturit y
group s) grow n in Fran ce from 1950–198 2, stat ed that modern hyb rids are
more adapted to stress, such as low temperature and drought. Regressions of
mean yield per decade of release on mean yield per location of trial consis-
tently showed that the newer the decade, the higher the yield at all locations.
Water stress limited yield in some locations, particularly for hybrids in the
semiearly category.
As not ed earlier in the sectio n “High Tem peratur es,” a 1930 to 1991 time
series of 36 hybrids and one OPC for Iowa exhibited a linear increase in
grain yield in a season (year 1991) with exceptionally low precipitation
during flowering, as well as in highly favorable seasons (Duvick, 1997).
The same series, further extended (1930–2001), showed a linear increase
in grain yield in another season (year 2000) when “heat and drought at
silking time caused reduced yields” (Duvick et al., 2004a), and also in a
third season (year 2001) when yields were low “because of a season-long
drought, especially severe at the sensitive anthesis-silking period” (Fig. 2)
(Duvick et al., 2004b). Weather records show that rainfall was well below
average during the anthesis-silking period in 2000, and also in 2001 (Iowa
State University, 2003).
A time series of 18 commercial hybrids adapted to central Iowa and
representing the period 1953–2001 showed linear gains in grain yield in
each of three different watering regimes. The hybrids (grown in a rain-free
environment in Chile) were (a) well watered, (b) subjected to drought at

ADVANCES IN MAIZE 101
flowering time, or (c) subjected to drought during the grain-fill period
(Barker et al., 2005; Edmeades et al., 2003). The hybrids in this experiment
were a subset of the series studied by Duvick et al. (2004b), discussed
previously. Gains in grain yield under optimal conditions were about twice
as large as gains when stress coincided with flowering or grain filling.
A time series of 2 OPCs and 52 hybrids adapted to central Iowa and
representing the years (for the hybrids) 1934–2001 was subjected to a man-
aged drought trial in Woodland, California (Barker et al., 2005). The
hybrids in this experiment were the same as those studied by Duvick et al.
(2004b), discussed previously. Watering regimes were similar to those de-
scribed for the experiment in Chile (described earlier). Trials were grown at
two plant densities. Both of the densities showed linear gains in grain yield
for all three watering regimes. Figure 3 shows results of the trial at high
density. Yield gain was greatest in the well-watered regime, although differ-
ences among the three regimes were not large. Annual genetic gains for all
watering regimes were greater in the trial grown at the higher density, typical
also of multidensity trials in rain-fed environments.
All of the experiments described in this section show that genetic yield
gains over time are expressed in drought as well as in favorable growing
Figure 3 Grain yield of two OPCs and 52 hybrids regressed against year of release. Hybrids
were grown in Woodland, California, at 90,000 plants ha�1 in three managed stress environ-
ments: full irrigation, flowering drought stress, or grain-filling drought stress. Adapted from
Barker et al. (2005).

102 DONALD N. DUVICK
seasons. On average, the newer the hybrid, the greater is its drought to-
lerance. Discussion in later sections suggests possible causes of this
improvement.
d. Excessi ve Soil Moisture. As noted ea rlier in this section (“Low Tem-
peratur es”), a 1930 to 1991 time series of Iowa hybrids showe d a linea r gain in
grain yield in a “very cool and wet season” (Duvick, 1997). That growing
season, 1993, was one of the wettest on record for the state of Iowa (precipi-
tation was two to three times normal), and soils were excessively moist (and in
some cases flooded, although not in these trials) during much of the summer
(Corrigan, 2003; Iowa State University, 2003). The linear gain in yield when
the 1930 to 1991 time series was grown in this unusually wet growing season
(see Fig. 2) indicates that although breeders had not selected directly for such
abnormally wet growing conditions, they must have done so indirectly,
perhaps through improvement in the ability of plants to set and develop
kernels in the presence of reduced photosynthesis per plant or in the ability
to tolerate a reduced uptake of key soil nutrients. One should note, however,
that yield gain was least (59 kg ha�1) in the flood year of 1993 (the lowest
yielding year) and greatest (82 kg ha�1) in the most favorable season, 1992.
e. Deficiency of Soil Nitrogen. Maize cultivars (OPCs and commercial
hybrids) typical of those grown in the U.S. corn belt in the decades 1930s
through 1980s were compared at high and low soil fertility levels (in trials
receiving approximately 200 kg ha�1 N, 90 kg ha�1 P2O5, and 150 kg ha�1
K2O versus trials in an area that, for two decades, had received no fertilizer
and was planted to continuous maize) (Castleberry et al., 1984). Yield gains
by decade were linear and positive under both of the soil fertility treatments
(high and low), although the average annual gain was greater in the high
fertility trial.
Single cross hybrids representing four decades (1940–1970) of U.S. corn
belt hybrids were compared at three levels (70, 130, and 200 kg ha�1) of
nitrogen (N) fertilizer application (Duvick, 1984a). The l970s decade gave
the highest grain yield at all N levels, and the 1960s decade produced the
second highest yield at all N levels. A second experiment, described in the
same report, compared five commercial hybrids spanning the period
1940–1978 at two treatments: high N, high plant density (215 kg N ha�1
and 54,000 plants ha�1) and low N, low plant density (70 kg N ha�1 and
35,000 plants ha�1) The two newest hybrids yielded more than the older ones
at either treatment level. In both experiments, the interaction of hybrids �N
rates was not statistically significant.
Carlone and Russell (1987) compared OPCs and a series of single cross
hybrids at three plant densities (34,445, 51,661, and 68,889 plants ha�1) and
four N fertilizer levels (0, 80, 160, and 240 kg ha�1). The single crosses were

ADVANCES IN MAIZE 103
chosen to represent hybrid genotypes of the decades 1930–1980. The trials
suffered severe moisture stress in both years of trial, 1983 and 1984. Com-
parisons used the yield of each era at its optimum plant density. Hybrids of
the older eras had their highest yield at lower densities; the newer hybrids
had their highest yield at the higher densities. The 1980s era had the highest
yield at each N level, and the 1970s era had the second highest yield at each
N level.
Carlone and Russell (1987) reported that levels of N fertilizer (0–240 kg
ha�1) interacted with plant densities and with hybrid genotype. The opti-
mum N level (level with highest yield) for hybrids of the 1940s, 1950s, and
1960s was higher than the optimum N level for hybrids of the 1970s and
1980s. However, the highest yields of the older group were lower than those
of the newer group at all N levels, so one might conclude that the newer
hybrids used N more efficiently than the older hybrids.
Carlone and Russell (1987) also showed that hybrids within an era
differed in response to densities and N levels. Two hybrids of the 1970s
group increased in yield as densities and N levels increased, but one hybrid
increased in yield significantly more than the other, such that the greatest
difference in yield between them was at the highest plant density and the
highest N level.
McCullough et al. (1994a) stated that when two hybrids were compared
in controlled environment chambers, an old hybrid (release year 1959) was
more sensitive than a new hybrid (release year 1988) to stress caused by low
soil N (0.5 mM) during early development. The new hybrid also maintained
a higher rate of leaf photosynthesis per unit of N regardless of N supply.
A second experiment (McCullough et al., 1994b) indicated that the higher
N-use efficiency of the new hybrid under low N supply “is associated with
higher N uptake and a higher leaf N per unit leaf area.” Field trials con-
firmed that the new hybrid yielded more than the old hybrid under both high
N and low N treatments (Tollenaar et al., 1994, 1997). The yield difference
between hybrids was accentuated when weeds were present, as compared
with weed-free conditions; one might conclude that the new hybrid was
also more “weed tolerant” than the old hybrid. As stated earlier, comparison
of only two hybrids representing “early” and “late” eras is not equivalent
to the study of trends in a time series of several hybrids, but the com-
parison can indicate possible changes over time and may suggest profitable
areas of future research to discover trait changes that accompany sequential
improvements in hybrid performance.
f. Unspecified Abiotic Stress—“Stress versus Nonstress” Environments.
Yield trial results can be categorized according to the average yield at
each test site. One can assume that the lower the yield at a given site
(in absence of obvious disease or insect problems), the greater the amount

104 DONALD N. DUVICK
of “unspecified” abiotic stress. A stability analysis of the kind proposed by
Eberhart and Russell (1966) can be used to compare yield response of
individual hybrids or of groups of hybrids such as those released in a given
decade. Yields of groups of hybrids (as in a decade) can be regressed on
mean yield at each test site (e.g., Figs. 4 and 5).
Russell (1991) cited such comparisons in several experiments. In general,
all experiments showed that the newest groups of hybrids had the highest
yields in all sites, regardless of average yield level at the site. However, linear
regressions (b), showing degree of response of each group of hybrids to
increasing site productivity, demonstrated no consistent trend of response.
In some cases, b values were similar for all eras, whereas in other cases, b
values were greater for new than for old eras, and in still other experiments,
the b values differed randomly among eras (e.g., Fig. 4). Russell concluded
that “there seems to be no distinct relationship between response and era of
the hybrids. More likely, the responses were specific for the genotypes.” He
Figure 4 Yield response, b, for open-pollinated cultivars (OPCs) and six hybrid groups of
10-year eras, 1930–1980, to eight environment indexes (four locations � 2 years). Reprinted
from Russell (1991), # 1991, with permission from Elsevier, and also with permission from W.
A. Russell and the Iowa State Journal of Research.

Figure 5 Mean grain yield of hybrids released within two-decade spans, and of three OPCs,
regressed on mean yield of all hybrids per environment. Trials were grown in a total of 13
environments during the years 1996–2000. Means of three densities per environment: 30,000,
54,000, and 79,000 plants ha�1. From Duvick et al. (2004b). Copyright# 2004 by JohnWiley &
Sons, Inc. This material is used by permission of John Wiley & Sons, Inc.
ADVANCES IN MAIZE 105
also consider ed the possibili ty that hand harvest vs machi ne harvest co uld
have intro duced a bias in some of the resul ts that he reviewed. (C ombines
might fail to ga ther ears of lodged plan ts, and because old er hybrids
tend to lodge mo re than newer hy brids, yiel ds of older hybrids woul d be
unde restima ted.)
Si nc e t he Russell (1991) report, Duvick et al. (2004b) p r e s en t e d r es ul ts f o r
42 commercial hybrids and four OPCs tested in 13 environments in central
Iowa during the years 1996–2000. They were grouped for stability analysis as
follows: OPCs, 1930s and 1940s, 1950s and 1960s, 1970s and 1980s, and 1990s
(Fig. 5). The regression for the OPCs was well below that for the hybrids,
at 0.65. Regression values were similar for all hybrid eras (b ¼ ca. 1.0), although
with a slight increase for the newest era. Thus, in this experiment the OPCs
showed markedly less response than the hybrids to higher yield environments
and the newest hybrid group gave the greatest response. However, in all cases,
the newer the era, the higher the yield in any location, low yield or high. Stress
tolerance increased significantly over the years.
In conc lusion, experi ments described in this section (“Toleran ce to
Abiot ic Stres s”) have shown that hyb rid toler ance to ab iotic stress has

106 DONALD N. DUVICK
increased consistently over the years. Although results are contradictory
regarding whether newer hybrids are more or less responsive than older
hybrids to higher yield environments, newer hybrids tend to be the most
responsive, sometimes strikingly so (e.g., Barker et al., 2005; Duvick, 1984a,
1992). Reasons for such variability in response are not known but probably
depend, as do most results, on interactions of genotype and environment.
5. Tolerance to Biotic Stress
a. Insects. A 1930–1991 time series of 36 hybrids and one OPC for
Iowa exhibited a linear increase in resistance to second-generation European
corn borer (Ostrinia nubilalis Hubner) (ECB2), as measured by tunnel
length following artificial infestation and by scores for evidence of natural
infestation in yield trials (Duvick, 1997). This improvement took place
even though breeders had not selected directly for resistance to ECB2.
The same series showed no improvement in resistance to first-generation
borer. Breeders and entomologists in the United States have collaboratively
produced inbreds and breeding populations with improved natural tolerance
and/or resistance to both generations of borer (see review in Russell, 1991),
but there is no record of how or if these materials were used in commer-
cial hybrids. They were not the source of increased resistance in the
aforementioned 1930–1991 time series.
In recent years (starting in 1996), seed companies have commercialized
transgenic maize hybrids that are resistant to ECB. These hybrids, common-
ly called Bt hybrids, have been genetically engineered to incorporate a gene
of Bacillus thuringiensis (Bt). Most of the first Bt hybrids contained the
gene that produces the insecticidal protein Cry1Ab. The toxic Bt protein is
effective against larvae from both first and second ECB generations
(Peferoen, 1992; Traore et al., 2000).
When subjected to artificial infestation, Bt hybrids showed significantly
less tunneling from second-generation borer than non-Bt hybrids (Traore
et al., 2000). They also had 9.7% more total plant weight in 1997 and 9.4%
more grain yield in 1998 than their non-Bt counterparts. However, the
amount of difference depended on the cultivar.
Under natural on-farm ECB infestation, Bt hybrids usually yield signifi-
cantly more than their isogenic counterparts in seasons when infestation is
relatively heavy, but not when infestation is light. In 14 yield trials in Iowa in
1997, nine Bt hybrids yielded 7% more than their near-isogenic counterparts
(Rice, 1997). Similar advantage for Bt hybrids was measured in several other
corn belt states in 1997, but in 1998 with lighter infestation the average yield
advantage was about one-fifth that of the 1997 amount (Gianessi and
Carpenter, 1999).

ADVANCES IN MAIZE 107
These data indicate that for Bt hybrids, as with other types of biotic and
abiotic stress tolerance, the amount of yield gain contributed by the benefi-
cial trait depends on the severity of the pertinent stress (in this case ECB
infestation). Thus, James (2003a) stated that global yield gains due to Bt
maize are currently estimated at 5% in the temperate maize-growing areas
and 10% in the tropical areas. The tropical areas have more and overlapping
generations of pests, leading to higher infestations and subsequently greater
yield loss, in absence of resistance contributed by Bt.
An additional restriction on potential yield gain from Bt hybrids is the
need to plant “refuge areas” (perhaps 20–30% of total) of non-Bt maize to
help prevent the development of resistance in the corn borer population
(Ostlie et al., 2002). Biologists theorize that the pest population will even-
tually develop/increase new genotypes that are not susceptible to the Bt
resistance genes, as has happened repeatedly (often in only a few seasons)
in other instances of major gene resistance [also called vertical resistance
(Simmonds, 1985; van der Plank, 1963)]. The use of refuge areas is intended
to delay such genotypic change as long as possible. The refuge areas of
course cannot provide the genetic yield advantage in the presence of corn
borer that is provided by the Bt hybrids.
More recently (in 2003), approval has been granted for use of a Bt transgene
that prevents root injury by larvae of two different species of rootworm
(Diabrotica barberi Smith & Lawrence, and Diabrotica virgifera virgifera
LeCont) (James, 2003b; Rice et al., 2003). This Bt protein is called Cry3Bb1;
it controls the rootworm larvae but not the adult beetle. As with Bt transgenic
protection against corn borer, genetic yield advantage of rootworm resistance
depends on the severity of infestation and its interaction with the environment.
“Yield trials demonstrated that under heavy rootworm pressure and moisture
stress the lack of corn rootworm larval injury in the [genetically engineered]
corn resulted in substantially higher yields than [in] cornwithout the Bt protein.
As rootworm pressure andmoisture deficits declined, the yield advantage of . . .genetically engineered corn declined.” (Rice et al., 2003).
Another similarity between the two kinds of Bt resistance is that refuge
areas will be needed to delay the development of rootworm populations that
are resistant to Cry3Bb1 (Rice et al., 2003). As with the corn borer Bt,
Cry3Bb1 imparts vertical resistance and presumably will lose its effectiveness
at some future date, thus necessitating replacement with a new genetic form
(or forms) of resistance.
b. Diseases. Frei (2000) stated that the minor presence of leaf diseases in
northern Europe allows increased emphasis on selection for yield performance
of maize hybrids for that area. This statement indirectly acknowledges that
maize breeders in other regions must select hybrids with tolerance or resis-
tance to locally prevalent diseases. The list of important diseases changes

108 DONALD N. DUVICK
from time to time, as new cultural methods and/or new genotypes encourage
diseases that had been absent or relatively unimportant (Dodd, 2000;
National Research Council (U.S.) Committee on Genetic Vulnerability of
Major Crops, 1972; Tatum, 1971).
Dodd (2000), speaking for maize in the United States, stated that during
the past 40 years at least 14 diseases of maize have had significant increase
in importance, although not all have endured or have proved to be wide-
spread. Their emergence as a problem is often encouraged by changes in
cultural practice, such as an increase in continuous maize growing and/or
in minimum tillage. At other times, widespread planting of a single genotype
will encourage spread of a particular disease. Breeders and farmers react
promptly to new disease problems; susceptible hybrids are dropped in favor
of resistant ones (if they are on hand) and further breeding ensures that new
releases have the needed level of resistance to the problem disease(s).
This battle will never end. Breeding for disease resistance shows its
success (and yield-enhancing contribution) most clearly when the disease is
active on susceptible hybrids (comparable to breeding for insect resistance).
It would be difficult or impossible to plot gradual gains in yield due to
gradual increases in disease resistance alone; nevertheless, the cumulative
effects of successful breeding for disease resistance surely must contribute to
the general increase in the level of on-farm yields. As Russell (1993) said,
“Selection for disease resistance has been an integral component of maize
breeding for many years, yet there are few data reflecting directly how
the success of this selection affects grain yield.” However, he does note
that “. . . improvement for stalk quality has been well documented . . . andstalk quality is highly dependent on plant health.”
One must acknowledge that interactions of disease resistance traits with
other beneficial genetic changes are perhaps the rule rather than the excep-
tion. For example, Clements et al. (2003) stated, “These results suggest that
Bt transformation events like MON810 are a useful supplement to hybrid
resistance to fumonisin contamination and fusarium [Fusarium spp.] ear
rot.” They went on to say that such benefits (reduced borer damage and
therefore less chance for disease entry) may accrue to susceptible hybrids
but not to hybrids with a relatively high level of resistance to fusarium.
The interactions of the two traits (disease resistance and insect resistance)
determine the outcome. One cannot credit either trait by itself.
6. Response to Changes in Plant Density
a. High Density. Genetic yield gain as a result of adaptation to contin-
ual increases in plant density is perhaps the most clear-cut and quantifiable
change in maize hybrids over the years. Cardwell (1982) calculated that

ADVANCES IN MAIZE 109
increased plant densities contributed 21% of the gain in maize yield in
Minnesota from 1930–1970. One must assume that not only the increase in
plant density but also the introduction of maize genotypes that could with-
stand and profit from the higher densities was essential to achieving the gain.
High plant density increases the deleterious effects of various kinds of stress
—abiotic and biotic—and so increases the need for genetic improvements in
stress tolerance (Troyer, 1996).
Several examinations of U.S. hybrids, summarized by Russell (1991),
showed that OPCs and old hybrids made their highest yields at lower
densities typical of their era, whereas the newest hybrids yielded the most
at the densities (always higher) typical of recent years. In other words, a
hybrid usually gave the highest yield when grown at the density for which it
was bred.
Similar results have been shown for late-maturing hybrids in France
and Ontario, Canada (essentially the same genotypes as grown in the
United States), but not so for early maturing hybrids in France or Ontario
(Derieux et al., 1987; Tollenaar et al., 1994). Tollenaar et al. (1994) suggested
that because newer hybrids of the early maturity group have greater leaf
area per plant as compared with the older hybrids of the same maturity,
they do not respond to (or need) higher plant density. He stated that, in
contrast, because the newer hybrids of the later maturity groups do not have
increased leaf area per plant compared with older hybrids in their maturity
group, the newer hybrids of the later maturity groups require more plants
per hectare to increase leaf area (and thereby photosynthetic surface) per
hectare.
As mentioned in Section II.D.4, optimum plant density can be affected by
the level of fertilizer N as well as by the hybrid genotype; hybrids differ
within and between eras in their response to various combinations of N level
and plant density (Carlone and Russell, 1987).
A 1930–1991 time series of 36 hybrids and one OPC for Iowa (Duvick,
1997) showed the same general trends as in earlier trials of shorter versions
of this series of hybrids (Duvick, 1977, 1984a, 1992); i.e., the older hybrids
yielded more at lower densities typical of their era, whereas the newer
hybrids yielded more at higher densities typical of their era.
However, the newest hybrids in the 1930–1991 time series made only a
very small gain in yield when planted at the highest density (79,000
plants ha�1) as compared with their performance at the intermediate density
(54,000 plants ha�1). This suggests the possibility that future yield gains
from breeding for adaptation to higher plant densities will come at a slower
pace and/or will require more breeding effort, at least with the present
breeding strategy. Breeders intending to increase genetic yield potentials
may need to modify or replace current breeding strategies and/or selection
criteria.
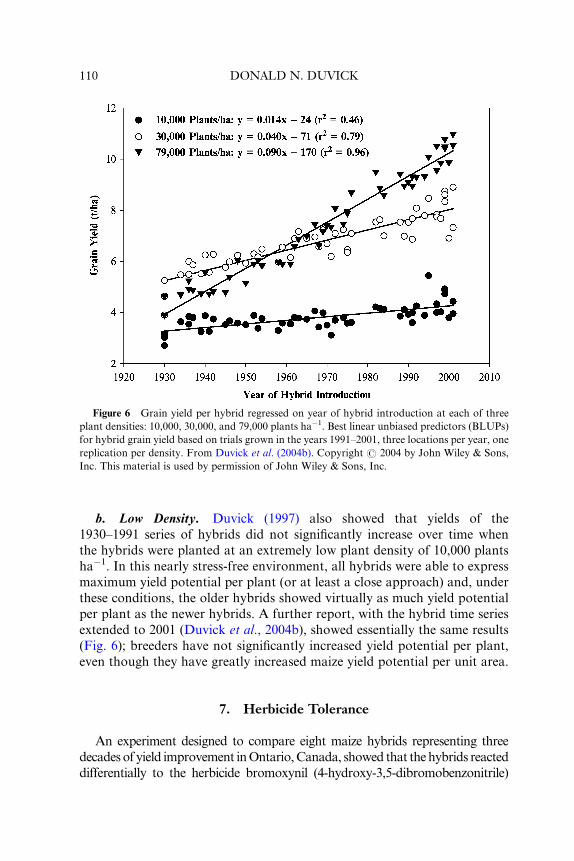
Figure 6 Grain yield per hybrid regressed on year of hybrid introduction at each of three
plant densities: 10,000, 30,000, and 79,000 plants ha�1. Best linear unbiased predictors (BLUPs)
for hybrid grain yield based on trials grown in the years 1991–2001, three locations per year, one
replication per density. From Duvick et al. (2004b). Copyright # 2004 by John Wiley & Sons,
Inc. This material is used by permission of John Wiley & Sons, Inc.
110 DONALD N. DUVICK
b. Low Density. Duvick (1997) also showed that yields of the
1930–1991 series of hybrids did not significantly increase over time when
the hybrids were planted at an extremely low plant density of 10,000 plants
ha�1. In this nearly stress-free environment, all hybrids were able to express
maximum yield potential per plant (or at least a close approach) and, under
these conditions, the older hybrids showed virtually as much yield potential
per plant as the newer hybrids. A further report, with the hybrid time series
extended to 2001 (Duvick et al., 2004b), showed essentially the same results
(Fig. 6); breeders have not significantly increased yield potential per plant,
even though they have greatly increased maize yield potential per unit area.
7. Herbicide Tolerance
An experiment designed to compare eight maize hybrids representing three
decadesof yield improvement inOntario,Canada, showed that thehybrids reacted
differentially to the herbicide bromoxynil (4-hydroxy-3,5-dibromobenzonitrile)

ADVANCES IN MAIZE 111
(Tollenaar and Mihailovic, 1991; Tollenaar et al., 1994). The hybrids in this
experiment, dating from 1959–1988, showed continuing improvement in
tolerance to bromoxynil, with a statistically significant trend of decreasing
phytotoxicity. They also showed continuing and significant improvement in
grain yield, especially at higher plant densities.
Commenting on these results, Tollenaar et al. (1994) suggested that
improved antioxidant defense mechanisms may be associated with increased
tolerance to bromoxynil and also with increased grain yield over the decades.
They also said, “The small and, in particular, gradual nature of the increased
bromoxynil tolerance suggests a highly complex, polygenic inheritance of
this particular kind of stress tolerance.”
Some maize hybrids now contain deliberately bred-in herbicide toler-
ance, primarily as transgenic resistance to broad-spectrum herbicides such
as glyphosate (e.g., Hetherington et al., 1999). Herbicide-tolerant maize
covered about 15% of the U.S. maize acreage in 2003 (ERS, 2003) and
in 2002 was planted on about 4% of all land planted to transgenic
crops globally (James, 2002). Strictly speaking, herbicide tolerance is
intended to improve the efficiency of weed management and not necessar-
ily to increase maize productivity, although better weed control could indi-
rectly result in higher yields, if weed levels were high with other kinds of
management.
In extreme cases, herbicide tolerance/resistance can increase maize yield
significantly and in strikingly large amounts. For example, initial experi-
ments in several African countries indicated that when maize is bred to be
resistant to a herbicide that normally is toxic to maize, seed coated with
that herbicide can provide effective season-long control of Striga spp.
(Kanampiu et al., 2003). Striga, a parasitic weed (sometimes called “witch-
weed”), can cause devastating crop loss in maize as well as other grain crops
in those countries. The herbicide resistance (imadazolinone resistance, “IR”)
is nontransgenic; it results from a mutation in an acetolactate synthase gene.
The specific herbicides used in these experiments were imazapyr and pyr-
ithiobac. When Striga density was high, the herbicide treatment resulted in a
three- to fourfold increase in yield. Kanampiu et al. (2003) stated, “When the
IR gene is incorporated into locally adapted varieties as in Kenya, this can
result in improvements in maize growth and hence high maize yield benefits
to small-scale farmers.”
Herbicide tolerance, indirectly, can produce undesired results and lower
yields. King and Hagood (2003) showed that postemergence control of
johnsongrass with glyphosate increased the severity of maize chlorotic
dwarf virus and maize dwarf mosaic virus in glyphosate-tolerant hybrids
that were susceptible to those diseases. They said, “The increased disease
severity resulted from greater transmission by insect vectors, which moved
from dying johnsongrass to the crop.” However, disease severity did not

112 DONALD N. DUVICK
increase in a virus-tolerant (and glyphosate-tolerant) hybrid subjected to
the same conditions. The authors concluded that for fields infested with
johnsongrass, the hybrid choice should be primarily for disease resistance
and secondarily for herbicide resistance.
8. Other Physiological Traits
a. Photosynthesis. As noted in earlier sections, leaf photosynthesis
seems to be more efficient in newer hybrids than in older hybrids when
they are compared in a range of stress conditions such as drought, low
temperature, or low N supply. Such an increase in efficiency could help
maize plants recover more rapidly from transient stresses such as those
induced by cold weather, overly wet soils, or drought. As summarized by
Tollenaar et al. (p. 215, 1994), “[These] findings, some of them preliminary in
nature, suggest that although hybrid differences in leaf photosynthesis under
unstressed conditions may not be indicative of actual or potential yield,
hybrid differences in response of leaf photosynthesis to stress conditions
may be a useful physiological indicator of high stable yields. To date,
selection for yield per se has apparently provided a selection pressure in
favor of stress-tolerant leaf photosynthesis.”
b. Canopy Gas Exchange, Temperature. Nissanka et al. (1997) com-
pared an old and a new hybrid (1959 vs 1988) in regard to whole plant gas
exchange and stem water potential throughout a water-deficit stress cycle
and during the subsequent recovery period upon rehydration. Under mois-
ture stress, the new hybrid maintained relatively higher rates of photosyn-
thesis and transpiration at a lower stem water potential than the old hybrid.
During the recovery day, canopy photosynthesis was 53% higher and
canopy transpiration was 31% higher in the new hybrid than in the old
hybrid. Respiration per unit CO2 fixed was lower in the new than in the
old hybrid in all conditions. The authors concluded that the new hybrid was
more tolerant to water stress and recovered faster upon rehydration than the
old hybrid. (As noted previously, comparison of only two hybrids is not
evidence of a long-term trend, but it may give useful suggestions for further
investigation.)
Canopy temperature under drought stress consistently decreased, going
from older to newer hybrids, in measurements of a set of 18 commercial
hybrids adapted to central Iowa and representing the period 1953 to 2001
(Barker et al., 2005). The hybrids were grown in a rain-free environ-
ment in Chile and were subjected to managed drought stress at various
stages of development. Barker et al. (2005) suggested that the trend to
lower canopy temperature under drought stress may result from the lower

ADVANCES IN MAIZE 113
radiation intensity on the more upright leaves of modern hybrids or on a
greater capacity by these hybrids to capture soil water.
9. Parentage and Genetic Diversity
As new hybrids replace those that preceded them, pedigrees and geno-
types change. It can be instructive to know more about the nature of these
changes. Which founder inbreds and/or OPCs remain in the pedigrees of
current hybrids and which ones disappear? Has genetic diversity decreased,
increased, or stayed about the same over the years? In a limited way some of
these questions were answered by Duvick et al. (2004a,b) with respect to the
time series of Iowa hybrids used for their studies of changes in hybrid
performance over time. For example:
• The 51 hybrids in the trials traced back to 53 founder sources: OPCs,
synthetic populations, and inbred lines. The founders came primarily from
the U.S. corn belt but a few were from the southeastern and northeastern
United States or from Latin America.
• Some families have persisted over the years and have contributed relatively
large amounts (by pedigree) to present-day hybrids, whereas others ap-
peared in pedigrees for only a few decades and then declined or disap-
peared. For example, Reid Yellow Dent, Iowa Stiff Stalk Synthetic, and
Reid Iodent have been important contributors since they first appeared in
pedigrees and they now contribute 33, 22, and 15%, respectively, to
hybrids of the 2000s. However, Maryland Yellow Dent, Boone Country
White, and the inbred Hy contributed briefly to pedigrees in the 1950s and
1960s but then disappeared completely from pedigrees of subsequent
hybrids in the time series. Krug reached a peak use of 23% in the 1940s
but declined rapidly to a steady level of about 3%, and some families
appeared late but have made significant, if not large, contributions;
for example, Argentinian Maiz Amargo appeared in the 1980s and has
contributed 4 to 5% in each of the past two decades.
• The pedigree information shows, therefore, that although certain lineages
have predominated over the years of hybrid improvement and replace-
ment, they have been supplemented significantly by additional, and di-
verse, lineages. On the whole, pedigree contributions have been broad and
volatile; the record provides strong evidence for “genetic diversity in time”
[as defined by Duvick (1984b)] in this 70-year time series of 51 hybrids for
central Iowa.Genetic diversity, including genetic diversity in time (also called
temporal genetic diversity), is widely acclaimed for its ability to provide
beneficial genetic response (timely and wide-ranging) to new and/or un-
usual kinds of biotic or abiotic stress (FAO, 1996; Gollin and Smale, 1998;

114 DONALD N. DUVICK
National Research Council (U.S.) Committee on Genetic Vulnerability
of Major Crops, 1972; Rosenow and Clark, 1987; Simmonds, 1962; Smale
et al., 2002).
10. Molecular Markers
Pedigree data, although informative, do not identify genetic materials in
the pedigree lineages, either qualitatively or quantitatively. Molecular mark-
er data, tracing a given DNA fragment from one generation to the next, can
enable quantification of the amount of founder germplasm that persists in
successive generations. Application of this technology to the Iowa time series
of hybrids (Duvick et al., 2004a,b) using simple sequence repeats (SSR)
showed that
• The number of alleles fluctuated from decade to decade, with about 40
to 50% of the total number of alleles present in any one decade. The
study identified 969 alleles at 100 SSR loci in the array of hybrids and
OPCs.
• The number of alleles per locus was similar for the female and male
parents of hybrids.
• Large-scale turnover of alleles took place in the first decades (1930s and
1940s) of hybrid breeding (which agrees with pedigree data), indicating
that many of the first inbred lines were not useful as parents for the next
generation of breeding. They were dropped from breeding pools, and new
and more successful breeding materials were brought in.
• The initial large-scale turnover of alleles was followed by a relatively
steady state of replacement until the 1970s (corresponding with the
changeover from double cross to single cross hybrids) when the number
of new alleles per decade again declined to what may be a new and lower
steady state of replacement.
• The alleles of the inbred parents of the modern hybrids (primarily single
crosses) could be separated (with multidimensional scaling analysis)
into two groups (called “stiff stalk” and “nonstiff stalk”), whereas alleles
of the older hybrids (primarily double crosses) sorted into an undifferenti-
ated third group (Fig. 7). This confirms the general observation that
breeders (using pedigree and experiential data) have established two breed-
ing pools to balance important traits (including those that enable use of
inbreds as either seed parent or pollen parent) and/or to maximize hetero-
sis when inbreds from the two groups are crossed with each other. The
primary goal of such divergent selection is to enable the development of
economically producible hybrids with improved on-farm yield and general
performance.
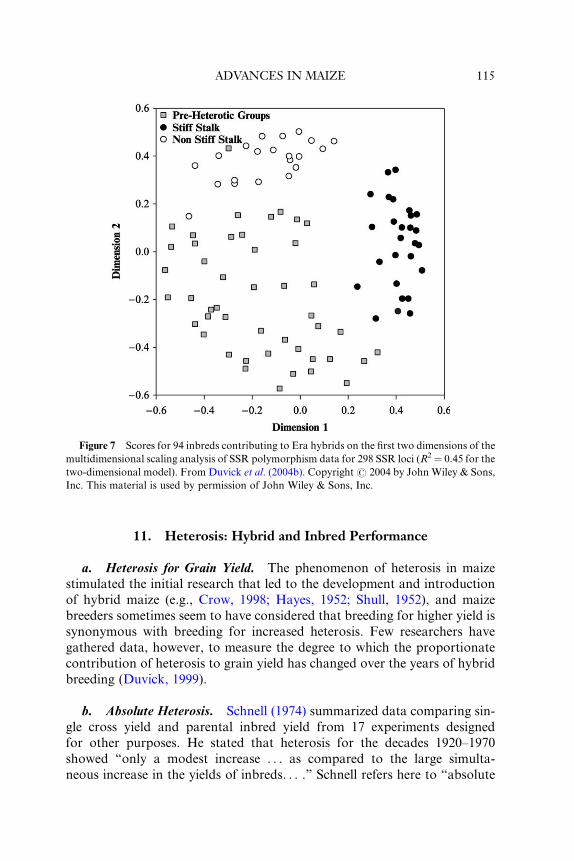
Figure 7 Scores for 94 inbreds contributing to Era hybrids on the first two dimensions of the
multidimensional scaling analysis of SSR polymorphism data for 298 SSR loci (R2 ¼ 0.45 for the
two-dimensional model). From Duvick et al. (2004b). Copyright# 2004 by John Wiley & Sons,
Inc. This material is used by permission of John Wiley & Sons, Inc.
ADVANCES IN MAIZE 115
11. Heterosis: Hybrid and Inbred Performance
a. Heterosis for Grain Yield. The phenomenon of heterosis in maize
stimulated the initial research that led to the development and introduction
of hybrid maize (e.g., Crow, 1998; Hayes, 1952; Shull, 1952), and maize
breeders sometimes seem to have considered that breeding for higher yield is
synonymous with breeding for increased heterosis. Few researchers have
gathered data, however, to measure the degree to which the proportionate
contribution of heterosis to grain yield has changed over the years of hybrid
breeding (Duvick, 1999).
b. Absolute Heterosis. Schnell (1974) summarized data comparing sin-
gle cross yield and parental inbred yield from 17 experiments designed
for other purposes. He stated that heterosis for the decades 1920–1970
showed “only a modest increase . . . as compared to the large simulta-
neous increase in the yields of inbreds. . . .” Schnell refers here to “absolute

116 DONALD N. DUVICK
heterosis,” the difference between the yield of a single cross and the mean
yield of its parental inbreds (midparent yield). The amount of annual increase
in midparent yield was nearly as great as that of the single cross hybrids.
Schnell also calculated heterosis as “relative heterosis” (absolute heterosis
divided by single cross yield) and noted that it decreased from 75% in the
1920s to about 50% in the 1970s. This is because the denominator (single
cross yield) increased at a faster rate than the numerator (absolute heterosis).
Meghji et al. (1984) studied changes in heterosis for inbreds and their
single crosses representing three decades (1930s, 1950s, and 1970s) of U.S.
corn belt hybrid germplasm. Six inbreds per decade represented the 1930s
and 1950s (two inbreds, WF9 and Os420, were in both decades) and seven
inbreds represented the 1970s (four of the six single crosses for the 1970s
contained the inbred Mo17). The trials were grown in Illinois; the year(s) of
the trial is not indicated. Yields of inbreds and single crosses (averaged
across densities) increased simultaneously over the decades. Absolute heter-
osis also increased over the decades; the increase averaged 51 kg ha�1 year�1
(Experiment 1, Table II). The increase was greater at a high density typical
of the 1970s than at a low density typical of the 1930s.
Duvick (1984a) compared inbreds and single crosses representing success-
ful Iowa hybrids for the decades 1930–1970. Five unrelated inbreds for each
decade were crossed in all possible combination to give 10 single crosses
per decade; trials were grown in 3 years (1977–1979) at three densities
Table II
Contributions of Absolute Heterosis and Relative Heterosis to Grain Yield in Three Experimentsa
Experimentb Categoryc
1930s 1940s 1950s 1960s 1970s 1980s
bdkg ha�1
1 SX yield 7097 7407 9538 61
Het Abs 4112 4438 6138 51
Het (%) (58) (60) (64)
2 SX yield 4600 5300 6900 7000 7900 83
Het Abs 2700 3200 4100 3600 4300 36
Het (%) (59) (60) (59) (51) (54)
3 SX yield 5941 6371 6865 7174 7929 9164 60
Het Abs 3787 3754 4143 4188 4102 4658 13
Het (%) (64) (59) (60) (58) (52) (51)
aAdapted from Duvick (1999), with permission from American Society of Agronomy, Crop
Science Society of America, and Soil Science Society of America.bExperiment 1: Means of two densities, data from Meghji et al. (1984). Experiment 2: Means of
three densities grown in 1977–1979, data from Duvick (1984). Experiment 3: Means of three
densities grown in 1992–1993, data from Duvick (1999).cSX yield, single cross yield; Het Abs, absolute heterosis; Het (%), relative heterosis.dLinear regression coefficient (kg ha�1 year�1).
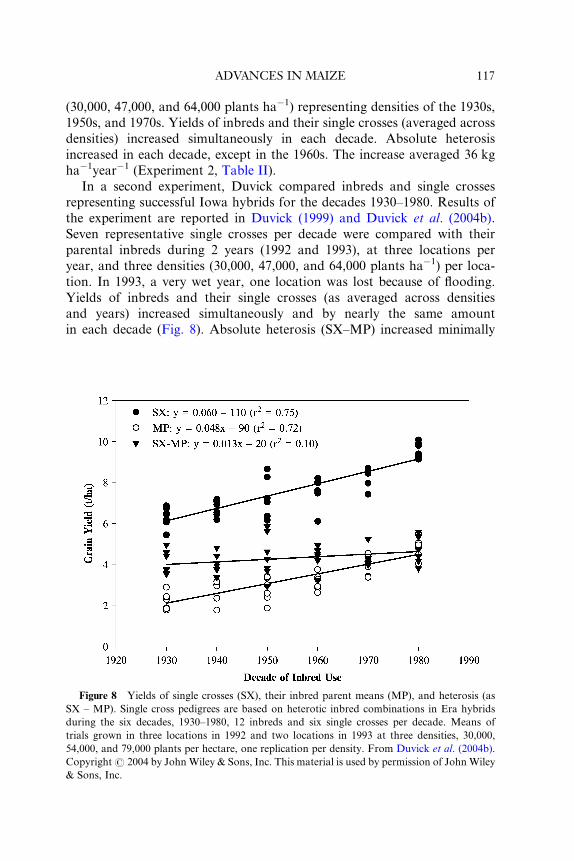
ADVANCES IN MAIZE 117
(30,000, 47,000, and 64,000 plants ha�1) representing densities of the 1930s,
1950s, and 1970s. Yields of inbreds and their single crosses (averaged across
densities) increased simultaneously in each decade. Absolute heterosis
increased in each decade, except in the 1960s. The increase averaged 36 kg
ha�1year�1 (Experiment 2, Table II).
In a second experiment, Duvick compared inbreds and single crosses
representing successful Iowa hybrids for the decades 1930–1980. Results of
the experiment are reported in Duvick (1999) and Duvick et al. (2004b).
Seven representative single crosses per decade were compared with their
parental inbreds during 2 years (1992 and 1993), at three locations per
year, and three densities (30,000, 47,000, and 64,000 plants ha�1) per loca-
tion. In 1993, a very wet year, one location was lost because of flooding.
Yields of inbreds and their single crosses (as averaged across densities
and years) increased simultaneously and by nearly the same amount
in each decade (Fig. 8). Absolute heterosis (SX–MP) increased minimally
Figure 8 Yields of single crosses (SX), their inbred parent means (MP), and heterosis (as
SX – MP). Single cross pedigrees are based on heterotic inbred combinations in Era hybrids
during the six decades, 1930–1980, 12 inbreds and six single crosses per decade. Means of
trials grown in three locations in 1992 and two locations in 1993 at three densities, 30,000,
54,000, and 79,000 plants per hectare, one replication per density. From Duvick et al. (2004b).
Copyright# 2004 by JohnWiley & Sons, Inc. This material is used by permission of JohnWiley
& Sons, Inc.
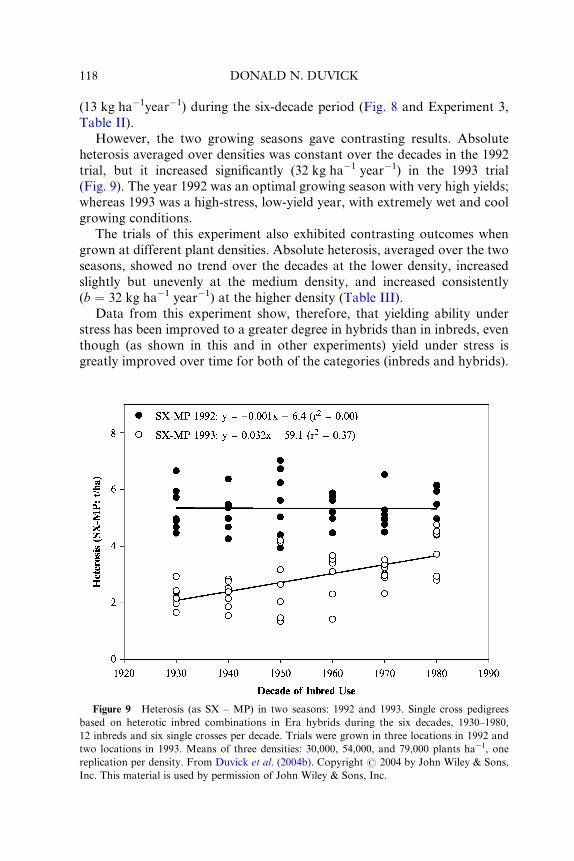
118 DONALD N. DUVICK
(13 kg ha�1year�1) during the six-decade period (Fig. 8 and Experiment 3,
Table II).
However, the two growing seasons gave contrasting results. Absolute
heterosis averaged over densities was constant over the decades in the 1992
trial, but it increased significantly (32 kg ha�1 year�1) in the 1993 trial
(Fig. 9). The year 1992 was an optimal growing season with very high yields;
whereas 1993 was a high-stress, low-yield year, with extremely wet and cool
growing conditions.
The trials of this experiment also exhibited contrasting outcomes when
grown at different plant densities. Absolute heterosis, averaged over the two
seasons, showed no trend over the decades at the lower density, increased
slightly but unevenly at the medium density, and increased consistently
(b ¼ 32 kg ha�1 year�1) at the higher density (Table III).
Data from this experiment show, therefore, that yielding ability under
stress has been improved to a greater degree in hybrids than in inbreds, even
though (as shown in this and in other experiments) yield under stress is
greatly improved over time for both of the categories (inbreds and hybrids).
Figure 9 Heterosis (as SX – MP) in two seasons: 1992 and 1993. Single cross pedigrees
based on heterotic inbred combinations in Era hybrids during the six decades, 1930–1980,
12 inbreds and six single crosses per decade. Trials were grown in three locations in 1992 and
two locations in 1993. Means of three densities: 30,000, 54,000, and 79,000 plants ha�1, one
replication per density. From Duvick et al. (2004b). Copyright # 2004 by John Wiley & Sons,
Inc. This material is used by permission of John Wiley & Sons, Inc.
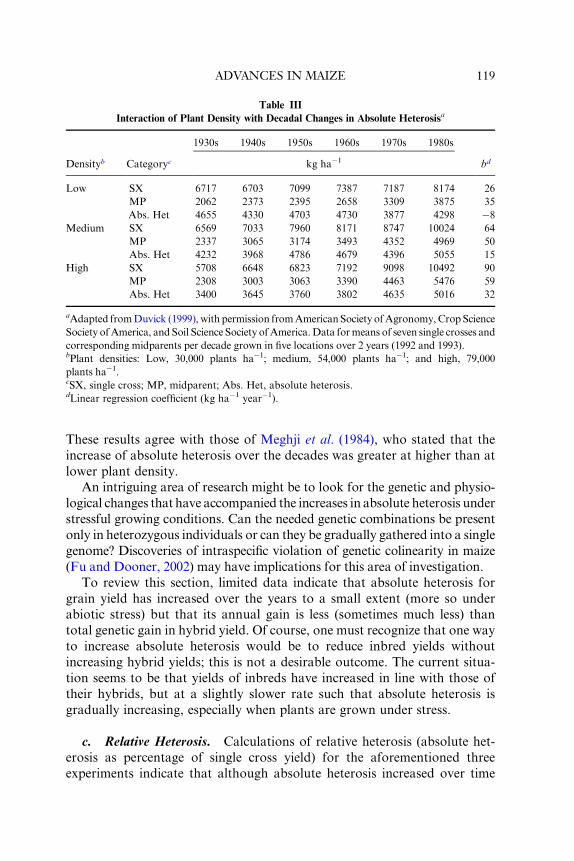
Table III
Interaction of Plant Density with Decadal Changes in Absolute Heterosisa
Densityb Categoryc
1930s 1940s 1950s 1960s 1970s 1980s
bdkg ha�1
Low SX 6717 6703 7099 7387 7187 8174 26
MP 2062 2373 2395 2658 3309 3875 35
Abs. Het 4655 4330 4703 4730 3877 4298 �8
Medium SX 6569 7033 7960 8171 8747 10024 64
MP 2337 3065 3174 3493 4352 4969 50
Abs. Het 4232 3968 4786 4679 4396 5055 15
High SX 5708 6648 6823 7192 9098 10492 90
MP 2308 3003 3063 3390 4463 5476 59
Abs. Het 3400 3645 3760 3802 4635 5016 32
aAdapted fromDuvick (1999), with permission fromAmericanSociety ofAgronomy,Crop Science
Society ofAmerica, and Soil Science Society ofAmerica.Data formeans of seven single crosses and
corresponding midparents per decade grown in five locations over 2 years (1992 and 1993).bPlant densities: Low, 30,000 plants ha�1; medium, 54,000 plants ha�1; and high, 79,000
plants ha�1.cSX, single cross; MP, midparent; Abs. Het, absolute heterosis.dLinear regression coefficient (kg ha�1 year�1).
ADVANCES IN MAIZE 119
These results agree with those of Meghji et al. (1984), who stated that the
increase of absolute heterosis over the decades was greater at higher than at
lower plant density.
An intriguing area of research might be to look for the genetic and physio-
logical changes that have accompanied the increases in absolute heterosis under
stressful growing conditions. Can the needed genetic combinations be present
only in heterozygous individuals or can they be gradually gathered into a single
genome? Discoveries of intraspecific violation of genetic colinearity in maize
(Fu and Dooner, 2002) may have implications for this area of investigation.
To review this section, limited data indicate that absolute heterosis for
grain yield has increased over the years to a small extent (more so under
abiotic stress) but that its annual gain is less (sometimes much less) than
total genetic gain in hybrid yield. Of course, one must recognize that one way
to increase absolute heterosis would be to reduce inbred yields without
increasing hybrid yields; this is not a desirable outcome. The current situa-
tion seems to be that yields of inbreds have increased in line with those of
their hybrids, but at a slightly slower rate such that absolute heterosis is
gradually increasing, especially when plants are grown under stress.
c. Relative Heterosis. Calculations of relative heterosis (absolute het-
erosis as percentage of single cross yield) for the aforementioned three
experiments indicate that although absolute heterosis increased over time

120 DONALD N. DUVICK
in most comparisons, relative heterosis did not increase in the two Iowa
experiments (Experiments 2 and 3, Table II), although it did increase in the
Illinois experiment (Experiment 1, Table II). Relative heterosis in the Iowa
experiment of 1992–1993 (Experiment 3, Table II) declined from 64% in the
1930s to 52% in the 1970s and 51% in the 1980s. Although absolute heterosis
increased minimally over the decades in this experiment, the gain in single
cross yield was greater than the gain for absolute heterosis, and so, as with
Schnell data, the proportionate contribution of heterosis to grain yield was
reduced over time. Interestingly, also, Schnell’s estimate of 50% for relative
heterosis in the 1970s corresponds closely with values of 54 and 52% for the
1970s in the two Iowa experiments.
Data summarized in this section suggest that relative heterosis for grain
yield has not increased markedly over the years; it more likely has stayed
constant or declined.
d. Heterosis for Other Traits. Plant height and flowering date exhibit
heterosis to a large degree in maize; crosses between two inbreds are always
taller and earlier than the mean of the parents. Heterosis for these traits may
(or may not) be related to some of the genetic interactions that produce
heterosis for grain yield and so it may be informative to examine changes
over time in heterosis for plant height, or other plant size measurements, and
flowering date.
Mean values for inbreeding depression in ear height and plant
height decreased over time, in comparisons made by Meghji et al. (1984).
This indicates that absolute heterosis for ear height and plant height de-
creased over time. In the same experiment, means for inbreeding depression
of tassel weight and tassel branch number increased in the 1950s but de-
creased in the 1970s to levels lower than in the 1930s. This would indicate
reduced heterosis for tassel weight and tassel branch number in the 1970s
genotypes.
Small but statistically significant trends toward reduced absolute heterosis
for plant height (�3 cm 10 year�1), ear height (�4 cm 10 year�1), and heat
units to anthesis (�11 heat units 10 year�1) were exhibited in the aforemen-
tioned comparison of inbreds and single crosses representing successful Iowa
hybrids for the decades 1930–1970 (Duvick, 1984a).
Absolute heterosis was reduced to a small degree for plant height but was
not reduced for heat units to anthesis in the aforementioned comparison of
inbreds and single crosses representing successful Iowa hybrids for the
decades 1930–1980 (Duvick et al., 2004b).
Data from these experiments indicate that, to a small degree, the size and
maturity differences between inbreds and their hybrid progeny were reduced
over time. However, the differences between the two classes remain (and
probably will remain) large. For example, reexamination of data for plant

ADVANCES IN MAIZE 121
height heterosis summarized in Duvick et al. (2004b) shows that the height
difference between the two classes was reduced from about 85 cm in the
1930s to about 70 cm in the 1980s, primarily because of a reduction in height
of the single crosses. If this rate of reduction in height difference could be
maintained (�0.3 cm year�1 or �15 cm 50 year�1), it would take about five
more 50-year cycles of selection to equalize inbred and hybrid height.
Data for changes over time in heterosis for plant size and maturity agree
in one respect with those for grain yield—neither category has exhibited
major increases (or decreases) in absolute heterosis. However, absolute
heterosis for grain yield has increased to a small degree (at least, under
stress), whereas absolute heterosis for plant size and maturity has decreased
to a small degree. Superficially, the two categories of heterosis do not seem
to answer to the same genetic stimuli.
III. GENETIC GAINS FROM POPULATIONIMPROVEMENT
A. COMPARISONS WITH GENETIC GAINS IN HYBRIDS
Although the emphasis of this review is on maize hybrids and how
successive changes in their breeding and genetics have contributed to
increased on-farm yield, recurrent selection to make improved populations
has interacted with and sometimes contributed to genetic improvements in
hybrids (e.g., via useful inbred lines bred from improved populations).
Additionally, for some farmers in some parts of the world, annual purchase
of hybrid seed is not an option. For these people, improved populations,
maintained by saving seed, are the only practical option for access to
improved cultivars. Therefore, this section briefly summarizes genetic gains
achieved by recurrent selection for population improvement in comparison
with genetic gains in hybrid performance and comments briefly on the
contribution of improved populations to hybrid maize yield.
A comparison of genetic gain for four recurrent selection experiments
with genetic gain for two time series of hybrids indicated that both methods
produced about the same annual genetic gain for grain yield: 71 kg ha�1 year�1
for recurrent selection and 68 kg ha�1 year�1 for the hybrids (Duvick, 1977).
Duvick described the two sets of experiments as follows: “Both took place
in central Iowa; both occurred in about the same time frame; both had as
a primary goal maximum improvement in yield.” For these reasons
he thought it appropriate to compare the outcomes of the two kinds of
breeding program. He also suggested, however, that selection pressure in the
recurrent selection programs might have placed less weight on nonyield

122 DONALD N. DUVICK
traits (e.g., root and stalk lodging) than was the case in the hybrid breed-
ing programs, thus allowing greater progress for yield per se in the recur-
rent selection programs. So the programs were not completely equivalent in
selection goals.
Duvick (1977) called for more comparisons of recurrent selection with
hybrid breeding (which typically is based on pedigree breeding), suggesting
that “Maize breeding probably could be helped by the results of quantitative
genetics studies specifically designed to compare ‘recurrent selection using
the pedigree method’ and ‘recurrent selection using the population pool
method.’ Good data, demonstrating the strong and weak points of these
two related and proven methods, would help the entire hybrid maize breed-
ing effort in its goal of producing good hybrids as quickly and efficiently as
possible.”
This request was easier to make than to grant, however, and to the
author’s knowledge, no such paired breeding programs have been designed
and executed. Even to compare estimates of genetic gain for hybrids with
estimates of genetic gain for recurrent selection is difficult because most
reports of progress in recurrent selection express yield gains in units cycle�1
rather than units year�1, and number of years per cycle usually is not stated
and must be inferred (if possible) from descriptions of the breeding cycle.
However, Edmeades and Tollenaar (1990) summarized 17 estimates of
genetic gain in temperate environments and 10 estimates of genetic gain in
tropical environments, with all estimates expressed as kg ha�1 year�1. With
one or two exceptions, the estimates for temperate environment are for time
series of commercial hybrids and the estimates for tropical environments are
for recurrent selection programs intended to produce improved populations.
Genetic gain in grain yield for the temperate programs averaged 66
kg ha�1 year�1, whereas genetic gain for the tropical programs averaged
145 kg ha�1 year�1. The average gain for the temperate experiments is iden-
tical to the average from the previously mentioned summary by Russell
(1991). This result is not too surprising because for the most part the two
lists cite the same reports, although the two lists are not identical. The list
of tropical experiments shows a broad range of estimates, from 51–310 kg
ha�1 year�1.
Edmeades and Tollenaar (1990) explained the high values for the esti-
mates of the tropical programs (primarily recurrent selection) as follows:
“The higher average rate of gain reported from the tropics is generally the
result of one, sometimes two and very occasionally three selection cycles per
year, the use of relatively unimproved germplasm with a broad genetic base,
and a selection scheme that was based on family performance.” Although it
is probably true that the first gains are the easiest in any professional
breeding program that starts with landrace materials, it also seems likely
that some of the high gains reported for the recurrent selection programs in

ADVANCES IN MAIZE 123
the tropics result from effective selection technology for important traits
such as drought tolerance (e.g., Banziger et al., 1999; Bolanos and
Edmeades, 1992), combined with rapid turnaround of breeding generations.
Edmeades and Tollenaar (1990) presented evidence for the possibility that
first gains are the easiest. They said that “. . . tropical maize . . . in its
unimproved state is tall, leafy, lodging-prone and has a harvest index of
about 0.35.” They stated that initial selection in unimproved populations in
the tropics results in shorter plants and reduced lodging, improved HI, and
reduction in barrenness.
The present review has shown that temperate hybrids have shown little or
no change over the years in plant height, leaf number, or HI when hybrids
are grown at the plant density for which they were bred. This may indicate
that farmer selectors had already changed these traits to a close approxima-
tion of “optimum” levels when they developed the corn belt OPCs that were
the basis of hybrid breeding. However, the first hybrids clearly surpassed
parent OPCs in root and stalk strength and in resistance to barrenness.
Although the stress of constantly increasing plant density continues to sort
out hybrid genotypes with increasingly greater resistance to lodging and
barrenness, these improvements are not as dramatic as those of the initial
hybrids compared with their parent OPCs. Taking into account these
changes (or lack thereof) in temperate breeding programs, one might predict
that annual yield gains of population improvement programs for tropical
materials eventually will move to lower (but nevertheless acceptable) levels,
similar to those for hybrid improvement in the temperate zones. Following
the relatively easy gains resulting from initial reductions in plant size and
lodging and from increases in HI, yield gains will need to come from more
gradual improvements in tolerance to locally important kinds of abiotic and
biotic stress.
Leaving aside comparisons between hybrid breeding and recurrent selec-
tion, numerous published reports testify clearly that recurrent selection for
grain yield and/or general performance can give positive results in temperate
as well as in tropical materials, and it can do so with acceptable investments
of time and effort (e.g., Hallauer and Miranda, 1988; Lamkey, 1992; Russell,
1991; Sriwatanapongse et al., 1985). Several methods have been tried; some
worked well and some did not (e.g., Edwards and Lamkey, 2002; Lamkey,
1992), much as has been true for various kinds of pedigree breeding applied
to hybrid development.
One difference between recurrent selection and hybrid breeding has been
that breeders sometimes have used recurrent selection primarily to intensify
expression of a single trait such as resistance to ECB or high grain oil
percentage (Klenke et al., 1986; Sprague, 1952). They often discovered that
other important traits such as grain yield or stalk digestibility could deterio-
rate, sometimes as an indirect response to strong selection for the intensified

124 DONALD N. DUVICK
trait or sometimes simply because of insufficient selection pressure for the
other important trait(s) (Hallauer and Miranda, 1988; Ostrander and Coors,
1997; Russell, 1991). Such populations could have only limited value as
sources of commercially useful inbred lines because inbreds (and the hybrids
they produce) must be as well balanced as possible for a full selection of
important traits. Transfer of the improved trait to useful inbred lines
may require a long-term effort or may not even be worth doing if its
intensification depends on the deterioration of another important trait.
B. RELATIVE CONTRIBUTIONS OF POPULATION IMPROVEMENT
AND PEDIGREE BREEDING
“Pedigree selection is the most widely used breeding method to develop
inbred lines for use as parents of hybrids.” . . . “Pedigree selection will always
bean important componentofmoderncorn-breedingprograms (Hallauer etal.,
1988, pp. 470–471),” “Pedigree selection was and is the most commonly used
selection method of line development (Hallauer andMiranda, 1988, p. 10).”
These statements, although in agreement with the general experience of
hybrid maize breeders, should not be construed to say that recurrent selection
for population improvement has notmade vital contributions to hybridmaize
breeding. Inbred lines such as B14, B37, and B73 have, in their time, been
parents of hybrids that dominated maize plantings in the U.S. corn belt, and
they also have been important sources of germplasm for further breeding by
pedigree selection. These inbreds are direct products of the previously de-
scribed population, BSSS (Russell, 1991). Other elite inbreds derived from
improved populations could be named as well (e.g., Hagdorn et al., 2003),
although their impact has been more limited than that of the “big three.”
Nevertheless, as stated earlier, pedigree breeding has been and remains
the backbone of hybrid maize breeding. [See Troyer (1996) for a detailed and
first-hand description of pedigree breeding in action.] Planned single crosses,
successful commercial hybrids, and crosses (or backcrosses) of an elite line to
an improved population are typical starting points for pedigree selection
to produce new inbred lines. The author knows of no detailed, data-based
exposition of the reasons for predominance and persistence of pedigree
breeding to develop inbred lines and so can only speculate about reasons
for this situation.
Perhaps one reason for the predominance of the pedigree method is that
its parental materials—inbred lines—have been very widely tested, not only
in breeders’ performance trials, but also by thousands of farmers. Breeders
typically have started selfing and pedigree selection from crosses of inbreds
(or their progeny) that were used in widely successful hybrids. Each hybrid
can be considered as a “test cross” for its parental inbreds. An unexpected

ADVANCES IN MAIZE 125
weakness in one or both parents is more likely to be discovered when a
hybrid is grown by thousands of farmers over a period of years, as compared
with growing an improved population in a relatively small number of yield
trials for one or two seasons. Conversely, one also can be more confident of
identifying genotypes that can give top performance over a wide variety
of environments if the replication number is in the thousands rather than in
the dozens (or fewer). (Commercial seed companies have a unique advantage
here in that they can gather trial data from hundreds or thousands of on-
farm “strip trial” comparisons. Although these data may be intended pri-
marily for potential use in sales promotion, they also provide the company’s
breeders with a deep mine of information about comparative performance of
a given genotype over a broad range of environments.)
A second consideration is that the odds of getting a superior inbred—with
good balance for all traits—from selfing a cross of two currently top-
performing inbreds are probably higher than the odds of getting a line of
equivalent merit from selfing an excellent but relatively heterogeneous im-
proved population, even though truly superior (and novel) genotypes may
well exist in the improved population.
This second possibility leads to further speculation that a new program
(such as some of those in the tropics) may have greater success in extracting
useful inbred lines from improved populations than has been exhibited in
mature temperate breeding programs. One reason could be that some of
today’s improved populations for the tropics are very high quality; they have
benefited from years of experience in designing and/or choosing proper
environments for performance trials and in the development of improved
designs for recurrent selection. The potential utility of such populations
(e.g., for production of stress tolerant hybrids) has been predicted by
Edmeades et al. (1997), who said “The probability of obtaining a hybrid
that yielded 40% greater than the trial mean under severe stress was 4-fold
greater when lines were extracted from a drought-tolerant source popula-
tion than from its conventional counterpart. . . . We conclude that drought-
or N-tolerant elite source populations provide a greater proportion of
drought- or N-tolerant inbred lines and hybrids.”
A second reason could be that there will be few or no truly superior
adapted inbred lines for a new breeding locality. If improved populations
(improved for the traits needed in the area where hybrids are to be grown)
are on hand, they may be the best available source of germplasm for the
development of inbred lines. They will be better than local OPCs and better
than crosses of unadapted inbreds that may be “elite” elsewhere but are not
well suited to the local environment. (This scenario contrasts with the course
of events in the U.S. corn belt, where two or three decades of pedigree
breeding had brought out many superior inbreds by the time improved
populations from recurrent selection programs were ready for use.)

126 DONALD N. DUVICK
In years to come, pedigree selection based on planned crosses of inbred
lines may become more competitive in new breeding localities (such as in
the tropics); however, further expertise in population improvement via
recurrent selection may enable continued development of populations that
are valuable sources of elite inbred lines for those environments.
A final speculation about the utility of improved populations is that as
expertise grows in molecular biology, breeders may learn how to “mine”
improved populations for valuable genes or gene combinations with more
precision and better odds of success than can be done with current methods
of breeding and selection. The improved populations could provide a wider
range of useful genetic diversity than can be achieved with pedigree breed-
ing and would have the diversity in much more useful forms than exist in
unadapted exotic cultivars and landraces. These latter materials are high
in genetic diversity but are also high in “useless” genetic diversity.
Remarks from Hallauer et al. (1988, pp. 531–532) make an appropriate
conclusion for this section. They said “Recurrent selection and pedigree selec-
tion . . . should not be considered in opposition to one another. Rather, the two
systems should complement each other. The goals of the two systems are
different, but the ultimate objective is the same—contribute to genetic gain.”
It seems safe to predict that as experience and technology progress,
breeders will find increasingly profitable ways to utilize recurrent selection
for development of a genetically diverse assortment of superior inbred lines
that can be parents of successful hybrids.
IV. ANALYSIS AND CONCLUSIONS
A. POSSIBLE REASONS FOR GENETIC YIELD GAINS
This review has shown that hybrid maize breeders have consistently
increased the yielding ability of hybrids during the past 70 years and that
genetic gains in grain yield are still linear. As the yielding ability of the
hybrids has increased, other traits have changed as well, in directions
that were sometimes intended and sometimes unintended or at least un-
planned. Conversely, some traits have not changed (or have changed very
little), sometimes at the intention of the breeders and sometimes despite the
breeders’ intent to make a change.
It will be instructive to categorize the various trait changes (or stabilities)
as described in Section II.D. They can be categorized as (1) trait changes that
promote the efficiency of grain production, (2) trait changes that increase
tolerance to biotic and abiotic stress, (3) intended trait stabilities, and (4)

ADVANCES IN MAIZE 127
unintended (or unplanned) trait stabilities. The various trait changes are
briefly summarized and arbitrarily sorted into these four groups as follows.
1. Trait Changes that Promote Efficiency of Grain Production
• Leaf angle has become significantly more upright, especially since about
the 1960s.
• Tassel size has been markedly reduced.
• Newer hybrids have longer period of grain fill but faster dry down;
therefore they are not later in harvest maturity and make better use of
the latter part of the growing season.
• Kernel weight is greater in newer hybrids except under drought stress at
late or terminal periods of grain fill.
• Newer hybrids have lower percentage grain protein.
• Newer hybrids have higher percentage grain starch.
• In some experiments, newer hybrids are markedly more responsive to
favorable environments (they make more efficient use of bountiful inputs),
although results are not consistent in this regard.
2. Trait Changes that Increase Tolerance (or Exhibit Evidenceof Increased Tolerance) to Biotic and Abiotic Stress
• Grain yield has increased in linear fashion; increases are greatest at
high plant density and are exhibited in high stress as well as low stress
environments, in poorly fertilized as well as in well-fertilized environments.
• Leaf rolling during drought stress is increased, perhaps because of changed
leaf orientation; potentially this can help maintain lower leaf temperature
and reduce water use.
• Staygreen (resistance to stress-induced premature death) is markedly
improved.
• Anthesis-silking interval is shortened, especially when hybrids are sub-
jected to conditions of abiotic stress such as drought or high plant density.
• Newer hybrids show increased resistance to barrenness when trials are
subjected to abiotic stress such as drought at flowering time or higher plant
density.
• Newer hybrids have higher HI than older ones if trials are subjected to
biotic stresses that induce barrenness.
• Hybrids show linear improvements in resistance to root lodging, although
some trials indicate a ceiling at about 95% nonroot-lodged plants.

128 DONALD N. DUVICK
• Hybrids show linear improvements in resistance to stalk lodging, although
some trials indicate a ceiling at about 95% nonstalk-lodged plants.
• Hybrids show linear improvement in yield in seasons with above-average
temperature during the growing season.
• Hybrids show linear improvement in yield in seasons with below-average
temperature during the growing season.
• Newer hybrids are more drought tolerant.
• Newer hybrids are more tolerant of excessive soil moisture (water-logged
soils).
• Newer hybrids are more tolerant of soil nitrogen deficiency.
• Newer hybrids are more tolerant of unspecified abiotic stresses (“low-
yield” sites).
• Newer hybrids are more tolerant of ECB2, and, recently, transgenic
hybrids have expressed a sharply increased level of resistance to both
generations of European corn borer and (separately) to two species of
rootworm.
• Circumstantial records show that new kinds of disease resistance are added
continually in response to new disease problems, but the contributions to
the yield of sequential changes in disease resistance are not documented.
• Newer hybrids, more tolerant of the stresses of higher plant density, enable
the use of higher plant density to maximize yield and therefore the grain
yield potential per unit area is increased.
• Newer hybrids show increased tolerance to a specific herbicide, apparently
correlated with increased antioxidant defense mechanisms; hybrids can be
bred (using conventional genetics) to be resistant to another specific her-
bicide; and in recent years transgenesis has been used to impart tolerance
to a third herbicide.
• A newer hybrid has more efficient photosynthesis than an older one,
especially under stress, and shows an improved capacity to recover the
photosynthetic rate after stress.
• A newer hybrid has more efficient canopy gas exchange, stem water
potential, transpiration, and respiration than an older hybrid when plants
are subjected to water stress.
• Canopy temperature under drought stress is decreased.
• Pedigree contributions have been diverse and are in a state of constant
change, although with a steady core of persistent lineages. The consequent
increase in genetic diversity (in time and in place) theoretically will provide
an increased stability of performance in the presence of diverse biotic and
abiotic stresses.
• Molecular marker studies validate the pedigree information and show,
further, that parent inbreds in recent years of the hybrid time series can
be divided into two genetically different groups called stiff stalk and
nonstiff stalk.

ADVANCES IN MAIZE 129
• Absolute heterosis for grain yield usually has increased to a small degree,
with the greatest increase when trials are grown under stress of high plant
density or drought.
3. Intended Trait Stabilities
• Plant and ear height have been relatively stable with a weak trend to lower
plant and ear height.
• Anthesis date is relatively unchanged.
• Date of silk emergence is unchanged over time in absence of stress, but
newer hybrids silk earlier than older hybrids when trials are subjected to
abiotic stress such as drought or high plant density; this is because silk
emergence is delayed (or fails entirely) in the older hybrids when subjected
to these kinds of abiotic stress.
• Tillering ability is rarely expressed at modern plant densities; tillering is
slightly reduced at low plant density.
4. Unintended (or Unplanned) Trait Stabilities
• Leaf number is unchanged.
• LAI is unchanged in the U.S. corn belt, but may be increased in early
maturity regions of Ontario (Canada).
• Number of ears per plant is not changed in absence of stress.
• HI of temperate hybrids is not changed if hybrids are grown at the density
for which they were bred.
• Yield potential per plant is not increased; i.e., newer hybrids do not yield
more than older hybrids at super-low plant density in absence of abiotic
stress.
• Relative heterosis for grain yield has not increased and, in some cases, is
slightly reduced.
• Absolute heterosis for plant height, ear height, and flowering date has not
increased and, in some cases, is slightly reduced.
Comparison of the categories in this summary shows that the list of
improvements in stress tolerance is by far the longest. This may indicate
that yield advances in hybrid maize depends primarily on increase in stress
tolerance—more specifically, on increased tolerance to the stresses that
typically occur in the environments where the hybrids are grown. It is true
that yields of both older and newer hybrids are reduced in the presence of
stress, either biotic or abiotic, but it also is true that the newer hybrids

130 DONALD N. DUVICK
always yield more than the older ones in the presence of stress and so, by this
definition, are more stress tolerant.
Changes that impart efficiency, such as smaller tassels, more upright
leaves, faster dry down of grain (to enable longer kernel-fill period), and
lower percentage grain protein, may have also enabled higher yields. These
changes were not selected directly, but they might have been selected indi-
rectly as a consequence of selection for increased grain yield per unit area
because they (presumably) improve the efficiency of transforming sunlight,
CO2, and soil nutrients into plant constituents, and so help increase yields.
The author is not surprised by the stability of maturity and plant size over
the years despite the fact that increased plant size and later maturity corre-
late positively with increased grain yield. The average date of first frost and
farmer prejudice against tall plants have automatically set the limits to which
breeders can select for harvest maturity and plant height, at least for U.S.
corn belt hybrids.
Some of the unintended (or unplanned) stabilities present surprises, or at
least do not agree with conventional wisdom. Probably the breeders’
intended constraint on plant and ear height has indirectly held leaf number
and leaf area index constant. In theory, the more leaf area per unit land area
(up to a maximum LAI of about four or five), the more photosynthesis and
consequently the higher the grain production per unit land area. However,
additional leaf area per unit land area has been achieved not by increasing
leaf area per plant, but by crowding more plants together. One could
hypothesize that shorter internodes could allow more leaves and more leaf
area per plant without increasing plant and ear height. This could substitute
for (or extend) the effect of increased plant density. However, plant archi-
tecture (e.g., leaf dimensions, shape, or angle) might need to be changed if
such an approach were used. Because leaf number usually is positively
correlated with time to flowering, it might not be possible to increase leaf
number and still hold the flowering date constant (a requirement for nearly
all temperate maize production).
Some may express surprise that HI of maize hybrids in the United States
has not increased (in absence of stress-induced barrenness) because of fre-
quent statements that an increase in HI was an important reason for the
higher yields of green revolution wheat and rice cultivars (e.g., Donald, 1968;
Peng et al., 1999; Reynolds et al., 1999; Swaminathan, 1998). However, the
first maize hybrids (as well as their parent OPCs) had about the same HI as
the initial high yield rice and wheat cultivars. It would appear that farmer
selection (at least in U.S. corn belt OPCs) had already brought maize plants
to an approximation of the 40–50% HI of initial green revolution rice and
wheat cultivars. Interestingly, rice and wheat breeders at the international
centers now suggest that an increase in biomass should receive major (per-
haps primary) emphasis as they strive to effect further increases in yield for

ADVANCES IN MAIZE 131
their crops (e.g., Peng et al., 1994, 1999; Reynolds et al., 1999). “Clearly,
there are limits to how far HI can be further increased in improved varieties
[of rice] (Peng et al., 1994).”
The lack of increase in yield potential per plant is surprising until one
reflects on the fact that up until now, the sole method of increasing yield per
unit area has been to increase plant density while maintaining constant ear
size (grain weight per plant). Although theoretically it may be possible to
raise yields per unit area by increasing yield per plant while holding popula-
tion constant (at lower densities than present norms), for one reason or other
this has not been done except in experimental studies (e.g., Fasoula and
Fasoula, 2000; Tokatlidis et al., 2001). Such a goal might be practical,
however, for hybrids suited for drought-prone environments, where planting
at lower density is prudent but the ability to utilize occasional higher rainfall
by increasing yield per plant would be desirable.
The relative lack of increase in heterosis, either absolute or relative, also
will surprise those of us who have supposed that the primary way to increase
hybrid yield is to increase heterosis for grain yield. It would appear that
comments by Hallauer (1999, p. 486) about recurrent selection can apply to
hybrids as well. He stated, “the additive effects of alleles with partial
to complete dominance were of greater importance but dominant and epi-
static effects could not be discounted.” It is also intriguing to note that
absolute heterosis for grain yield is greatest under conditions of stress, in
company with the knowledge that an increase in stress tolerance of all kinds
has strongly accompanied gains in hybrid yield over the years. Both inbreds
and hybrids are greatly improved in stress tolerance, but hybrid gains are
greater that those of their parental inbreds.
B. POTENTIAL HELPS OR HINDRANCES TO FUTURE
GAINS in YIELD
For the past 70 years, breeders have improved the yielding ability of
hybrid maize by selecting new genotypes with adaptation to the ever-increas-
ing stresses of constantly increasing plant density as well as to other kinds of
prevalent abiotic and biotic stress. They have done so by exposing these
hybrids to an increasing diversity of environments as the capacity for wide
area testing (especially in the commercial sector during the past few decades)
has been dramatically increased by investments in mechanization and infor-
mation management. Breeders consistently have selected for higher average
yield with acceptable grain moisture, improved standability, resistance to
local diseases and insect pests (that often change in kind or intensity over the
years), and tolerance to increased plant density. (Farmers continually plant
the newest hybrids at a density higher than recommended, thus forcing

132 DONALD N. DUVICK
breeders to continually raise the density at which they select the next round
of hybrids.) Newer hybrids have managed the stress of high plant density
(which usually accentuates other kinds of stress) in two ways: increased
stress tolerance and increased efficiency of grain production.
Efficiencies in grain production that presumably are provided by more
upright leaves, smaller tassels, and lower grain protein percentage may have
gone about as far as they can go; leaves cannot be much more vertical
without clasping the stem, tassels on some hybrids have no side branches
at all, and it is possible that livestock feeders will not be willing to accept
grain with a smaller percentage of protein than is now at hand.
Therefore, breeders will need to make even greater progress in improving
traits for stress tolerance if they are to continue the linear increase in grain
yield that has prevailed during most of the past 70 years of hybrid maize
breeding. Increased emphasis on such traits as tolerance to extremes of
temperature, to drought, to excess soil moisture, and to deficiency of soil
nitrogen (without neglecting simultaneous selection under nonstressed, high-
yield conditions) will probably be worth the effort. For each of these traits,
development and deployment of managed stress levels (such as managed
irrigation in a rain-free climate) may increase the precision and reliability of
selection for the trait.
However, widespread on-farm testing in a full range of “natural” envir-
onments, from high yield to high stress, must be the rock on which all other
selections are based. Past experience has shown that overemphasis on a
single trait may produce correlated changes in other traits, sometimes with
unintended (and unwanted) results. There is no substitute for extensive
trials, in both small plot and strip test format, of hybrids in final stages
before release (and also just after release). Although one may not be able to
identify the different kinds of stress (or nonstress) in those trials with
precision, one can be sure that such testing will subject the genotypes to a
multitude of stresses common to the area of adaptation. Some of the stresses
will be severe to catastrophic and some will be so minimal that only the
hybrids can sense them. In the end the genotypes with the broadest tolerance
to these stresses will give the highest yields in both low-yield and high-yield
environments.
C. PREDICTIONS
Mark Twain is supposed to have said, “Prophecy is a good line of
business but it is full of risks.” This caution surely applies to predictions
about prospects for future yield gains in hybrid maize, or in other kinds of
maize cultivars.

ADVANCES IN MAIZE 133
Increased grain yield per semay be less important in the future than in the
past for an increasingly large part of maize production. Specialty products
such as maize with altered or higher oil or protein content, or high extract-
able starch, or maize bred for use as a biofuel may rise in importance in
addition to or sometimes in place of commodity feed grain production (e.g.,
Lambert et al., 1998; Ng et al., 1997; Whitt et al., 2002). Conceivably, some
of these novel kinds of maize could command a premium price. Although
yield of some kind would be important for these specialty categories of
maize, grain yield per se would take second place to yield of a specified
product—yield of a specific kind of oil, protein, starch, etc.
However, despite these possible new markets for specialty types of maize,
the demand for maize as a feed grain will increase robustly if the developing
world continues to improve its economy and therefore its appetite for meat,
milk, and eggs (Rosegrant et al., 2001; Taha, 2003). Consequently, produc-
tion of maize as a commodity grain for animal feed will continue to domi-
nate commercial maize production. Increasing the on-farm grain yield of
maize hybrids will persist as a primary goal of maize breeders. Farmers will
require (and demand) hybrids that dependably produce maximum yield
with minimum inputs—a key requirement for the profitable production of
commodity maize grain.
Although the past does not necessarily predict the future, 70 years of
linear genetic yield gain in many parts of the maize-growing world would
seem to predict that similar gains will continue for at least the next few
decades. This prediction is more likely to hold for regions that have recently
adopted hybrids and complementary intensive management practices. Pro-
duction in those regions will not be as near to the theoretical maximum yield
potential (Cassman, 1999; Duvick and Cassman, 1999) as may be true for
regions with a longer period of constantly increasing yields. However, even
in regions with longer periods of yield increase such as the U.S. corn belt,
continuation of the long-standing practice of remolding adapted germplasm,
and slowly and carefully importing useful pieces of exotic germplasm, will
guarantee increased yield potential and increased stability of yield for years
to come.
On-farm yields may not always rise in line with genetic improvements,
however. In the coming decades, residents of the wealthier countries may
force their farmers to reduce (or, in some cases, eliminate) applications of
synthetic fertilizers and/or pesticides, with the intention to improve environ-
mental and human health. Such reductions could reduce maize yields. Even
though the newer hybrids would yield more than the older ones (including
the OPCs) following such a reduction of inputs, their yields could very well
be lower than before reductions were put into effect, although the amount of
loss (if any) would depend on the extent of the input reductions.

134 DONALD N. DUVICK
If this speculated mandatory reduction of management inputs should
come to pass, one can envision a future in which genetic yield gains will
continue but on-farm yields will stagnate or decline. In other words, the
genetic improvement of critical traits might be needed simply to maintain
yields at an even level or to minimize reductions in yield. Without genetic
improvements, yields would drop even more. This scenario highlights the
importance of breeding for resistance to disease and insect pests, tolerance to
deficiency of soil nutrients, and tolerance to other locally important kinds of
abiotic stress.
One should note, however, that breeders have already made good prog-
ress in breeding for such kinds of tolerance/resistance. As shown repeatedly
in this review, breeding for increased resistance to abiotic and biotic stress
has been the basis for 70 years of yield increase and dependability in hybrid
maize. With stimulus from farmers and the marketplace, it seems reasonable
to suppose (and predict) that breeders can and would increase the intensity
of breeding for stress tolerance, with special emphasis on specific stresses
that were amplified by the reduction of specific inputs.
For example, data reviewed in earlier sections of this report indicate that
successive hybrids have shown steady and significant genetic improvement in
the efficient use of soil nitrogen, although the selection for improvement was
indirect (and unintentional). If application of nitrogen fertilizers should be
curtailed, maize breeders could select directly for efficient use of soil nitro-
gen, and should be able to make even faster progress than in earlier years.
Such progress might enable a continued increase (or at least prevent a
decline) in on-farm yields, despite mandated reductions in fertilizer use.
Likewise, with regard to pesticides, as noted earlier, a small number
of effective aids from biotechnology for genetic improvements in disease
and insect resistance are already in place and more are contemplated
(Fernandez-Cornejo and McBride, 2002; James, 2003a; Rice et al., 2003;
Runge and Ryan, 2003). Continuation and enhancement of these transgenic
breeding efforts could incrementally and significantly increase the ability of
farmers to maintain yields without the use of pesticides. (An example of
enhancement of transgenic protection would be to progress from the con-
struction of vertical resistance genotypes to building systems of horizontal—
more durable—resistance.) Importantly, the transgenic improvements will
be most useful if they are integrated with continuing achievements in con-
ventional breeding for pest resistance (which itself will be enhanced by new
knowledge and tools, contributed by molecular biology). In time, there will
be no distinction between “biotechnology protection” and “conventional
protection.”
For some maize producers, an even more drastic reduction in inputs may
occur in future years, as water for irrigation becomes less available in regions
where irrigation is important (and sometimes essential) for maize production

ADVANCES IN MAIZE 135
(Rosegrant et al., 2002). As with breeding for tolerance to nutrient imbal-
ance or biotic stresses, breeders in regions of water shortage will need to
increase their emphasis on breeding for drought tolerance, either by moving
from indirect to direct selection or by increasing the emphasis on already
existing direct selection for drought tolerance. Past experience indicates that
this trait can be improved without sacrificing the ability to produce high
yields with favorable water balance, and evidence now accumulating indi-
cates that these breeding efforts may someday be made even more efficient
because of new insights provided by molecular biology investigations. So, as
with postulated reductions in soil nutrient and pesticide application, bree-
ders should be able to mitigate yield reductions, or even maintain on-farm
yields, in many of the areas where irrigation is reduced or eliminated.
A less optimistic prediction, based on past experience, is that the price of
successive increases in genetic yielding ability will continue to rise, just as it
has risen during the past 70 years. In the United States, for example, today’s
crew of maize breeders (broadly defined to include those who work in
biotechnology applied to plant breeding) is many times larger than the
crew that made advances during the first two or three decades of hybrid
maize breeding (Crosby et al., 1985; Duvick, 1984a; Fernandez-Cornejo,
2004; Frey, 1996), yet the genetic yield gain per year is no larger than in
past times—the gain is linear. (And therefore the expenditure per unit gain is
many times larger now than in early years.) Judging from this past experi-
ence, today’s plant breeding crew will need to be enlarged even further if
future gains are to be made at the same pace as is now achieved unless much
more efficient methods of breeding are developed and implemented.
Some have predicted that significant gains in breeding efficiency will
occur as various tools of biotechnology are employed, utilizing new genetic
discoveries and new knowledge of genes and gene action in the maize plant.
However, the current state of the art primarily is in the development of
techniques and data (e.g., Emrich et al., 2004; Jansen et al., 2003; Lawrence
et al., 2004). Biotechnology is not the primary tool for the development of
improved cultivars. Although useful new traits have been added via trans-
genesis, the number actually in use is still small (but growing) and is limited
to such defensive traits as pest resistance and/or herbicide tolerance. Bree-
ders must continue to use routine empirical breeding methods to effect
broad-scale yield improvements in the “base germplasm,” which they then
can enhance with useful transgenes. Today’s maize breeders cannot reduce
the effort devoted to “conventional plant breeding” without also reducing
the rate of progress in the development and release of higher-yielding culti-
vars, cultivars with a full and well-balanced spectrum of the important traits
that in sum govern maize yield.
Additionally, the present large investment in the testing of transgenic
products for safety adds significantly to the cost of their use for genetic

136 DONALD N. DUVICK
improvement. Comments by soybean breeders on the use of biotechnology
for soybean breeding are relevant to its use in maize breeding, as follows:
“Despite all the opportunities, biotech soybeans face numerous challenges.
Because of the cost of technology and regulatory clearance, it is difficult
for developers to earn sufficient returns on research investment for many
biotech traits. Gaining acceptance of crops and grain derived through bio-
technology, particularly in Europe, is yet another challenge. Although
biotechnology acceptance is increasing around the world, significant chal-
lenges will be faced by those wanting to bring new transgenic traits to market
(Soper et al., 2003).”
One hesitates to predict how much time must pass before the current
investment in biotechnology can bring about significant savings in time and
money per unit gain in maize yield, even though it is obvious that biotech-
nology research and development indeed will give significant and innovative
support to maize breeding. New techniques, based on new knowledge in
molecular biology, are increasing breeding efficiency incrementally, and with
time the number and use of such new techniques can only grow. As stated by
Runge and Ryan (2003), “Plant biotech R&D in the pipeline as of 2001
through mid-2003 indicates almost a hundred new traits in testing. Repre-
sented in these activities are about 40 universities (mainly land grants) and
about 35 private sector companies. Without question, more research
and development as measured by field tests has been devoted to biotech
traits in corn than to any other crop, attracting scores of public and private
institutions. Among the traits in testing for corn were 19 new agronomic
properties, four traits for fungal resistance, seven for herbicide tolerance,
four for insect resistance, ten trials focusing on some form of marker genes,
and over 30 for output and other end-use traits.”
This leads to the author’s final prediction, that despite the efforts of some
segments of society (e.g., Turning Point Project, 1999a,b) to stop or other-
wise hinder the use of biotechnology as an aid in food production (for
detailed description of this subject, see Charles, 2001), the tools of molecular
biology increasingly will aid maize breeders in their efforts to develop
superior hybrids. The most important and long-reaching aid will come
not so much from transgenes per se as from the use of a wide range of
biotechnology-based tools to give breeders a deeper knowledge of the genet-
ics and physiology of the maize plant; with this knowledge they will be able
to fine-tune maize genomes to achieve desired ends with much greater speed
and efficiency.
Valuable assistance will continue to come directly from some classes of
transgenes, e.g., new and more broadly effective versions of Bt transgenes
(James, 2003a; Rice et al., 2003). Breeders in the tropics and subtropics will
have particular use for transgenes that impart effective resistance to the wide
array of disease and insect problems in those regions, especially for instances

ADVANCES IN MAIZE 137
in which adequate genetic resistance cannot be found in maize itself. In
all cases, these transgenes will prevent yield loss in the presence of epi-
demics and infestations of the pest in question. In those places where
pest depredation is chronic, yield levels on average will advance. However,
as noted earlier, the use of transgenes for pest resistance must move
beyond changes that contribute vertical resistance to those that impart
more durable kinds of horizontal resistance, not an easy task but one
that will become possible as biotechnology-based knowledge and insights
accumulate.
As biologists move beyond genomics to proteomics, metabolomics,
and other related disciplines (some as yet unnamed or undiscovered), they
will help maize breeders identify key genes and gene systems/interactions in
the maize plant, and then, working together, the molecular biologists and
breeders will learn how to regulate or reconstruct them in ways that
will intensify the expression of key traits, whether for tolerance to heat
and/or drought, to chronic disease problems, or to tolerance of deficiency
of soil nutrients such as nitrogen. Breeders will learn how to mine geneti-
cally diverse exotic maize populations for improved versions of key genes or
(more likely) their regulator systems and, with aid of molecular markers, to
move them into elite germplasm with precision and efficiency (see Tuberosa
et al., 2002). (Marker-assisted selection is already used extensively in some
crops, including maize, to move useful genes or linkage groups from exotic
to adapted germplasm and/or elite cultivars.) In some cases, knowledge of
the identity and/or function of important genes in other species (such as for
tolerance to certain kinds of abiotic stress) will enable biologists to identify
their counterparts in maize, enabling breeders and molecular biologists,
collaboratively, to fine-tune the actions of the maize genes, sometimes by
using key regulatory sequences from the exotic organism (e.g., Appenzeller
et al., 2004; Shou et al., 2004).
Of course, individual genetic changes will be useful only when they
efficiently interact with the complete genetic complex in ways that improve
the overall performance of the plant according to goals set by the breeder.
Knowledge of the ways in which expression of a key gene affects the action
and products of other genes (i.e., the pleiotropic and epistatic effects), and
consequent interactions with the environment, will be critical to the success
of improving the action of any individual gene.
Finally, beyond all these conjectured advances, the maize breeders
can always hope for the Holy Grail of plant physiologists, major improve-
ment in the efficiency of the primary steps of converting intercepted solar
radiation into stored carbon, effected without disrupting the rest of the
infinitely complicated network of interacting genetic systems and ensuing
physiological processes that operate the functioning maize plant or any
other organism.

138 DONALD N. DUVICK
However, in the end, after all the modern tools have been employed to the
maximum degree, maize breeders will still need to walk the fields, observing
their latest creations under the widest possible range of conditions that
commonly occur in the intended region of adaptation. (One could describe
this activity as “personal perusal and evaluation of the genotype � environ-
ment interaction.”) The breeders will collate this subjective and highly
personal information with objective information obtained from widespread
performance trials, laboratory analyses, and other factual tests of perfor-
mance. In brief, maize breeders will need to practice the art as well as the
science of breeding if they are to continue the genetic progress that has been
achieved by their predecessors during the past three-quarters of a century.
“As the joy of artistic creation begins to assert itself we may expect many
interesting developments in the newer methods of corn breeding (H. A.
Wallace, 1930, unpublished manuscript).”
REFERENCES
Appenzeller, L., Doblin, M., Barreiro, R., Wang, H., Niu, X., Kollipara, K., Carrigan, L.,
Tomes, D., Chapman, M., and Dhugga, K. S. (2004). Cellulose synthesis in maize: Isolation
and expression analysis of the cellulose synthase (CesA) gene family. Cellulose 11, 287–289.
Baker, R. F. (1990). D. N. Duvick interview with R. F. Baker. Pioneer Hi-Bred International,
Inc., Johnston, Iowa.
Banziger, M., Edmeades, G. O., and Lafitte, H. R. (1999). Selection for drought tolerance
increases maize yields across a range of nitrogen levels. Crop Sci. 39, 1035–1040.
Barker, T., Campos, H., Cooper, M., Dolan, D., Edmeades, G., Habben, J., Schussler, J.,
Wright, D., and Zinselmeier, C. (2005). Improving drought tolerance in maize. Plant Breed.
Rev. 25, 173–253.
Bolanos, J., and Edmeades, G. O. (1992). Eight cycles of selection for drought tolerance in
lowland tropical maize. I. Responses in grain yield, biomass, and radiation utilization. Field
Crop Res. 31, 233–252.
Bruulsema, T. W., Tollenaar, M., and Heckman, J. R. (2000). Boosting crop yields in the next
century. Better Crops 84, 9–13.
Cardwell, V. B. (1982). Fifty years of Minnesota corn production: Sources of yield increase.
Agron. J. 74, 984–990.
Carlone, M. R., and Russell, W. A. (1987). Response to plant densities and nitrogen levels for
four maize cultivars from different eras of breeding. Crop Sci. 27, 465–470.
Cassman, K. G. (1999). Ecological intensification of cereal production systems: Yield potential,
soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 96, 5952–5959.
Castleberry, R. M., Crum, C. W., and Krull, C. F. (1984). Genetic yield improvement of
U.S. maize cultivars under varying fertility and climatic environments. Crop Sci. 24,
33–36.
Cavalieri, A. J., and Smith, O. S. (1985). Grain filling and field drying of a set of maize hybrids
released from 1930 to 1982. Crop Sci. 25, 856–860.
Charles, D. (2001). “Lords of the Harvest: Biotech, Big Money, and the Future of Food”.
Perseus Press, Cambridge, MA.

ADVANCES IN MAIZE 139
Clements, M. J., Campbell, K. W., Maragos, C. M., Pilcher, C., Headrick, J. M., Pataky, J. K.,
and White, D. G. (2003). Influence of Cry1Ab protein and hybrid genotype on fumonisin
contamination and fusarium ear rot of corn. Crop Sci. 43, 1283–1293.
Corrigan, P. (2003). The Flood of ‘93: Comprehensive review of the 1993 flood and how it
impacted Iowa. [Online]. Available by National Weather Service, Des Moines, IA http://
www.crh.noaa.gov/dmx/WX_events/93floodsummary.shtml (viewed November 3, 2003).
Crabb, R. (1993). “The Hybrid Corn-Makers”, 2nd Ed., West Chicago Publishing Company,
West Chicago, IL.
Crosbie, T. M. (1982). Changes in physiological traits associated with long-term breeding efforts
to improve grain yield of maize. In “Proceedings of 37th Annual Corn & Sorghum
Research Conference, 5–9 Dec., 1982” (H. D. Loden and D. Wilkinson, Eds.), Vol. 37,
pp. 206–223. American Seed Trade Association, Washington, DC, Chicago, IL.
Crosby, E. A., Eddleman, B. R., Kalton, R. R., Ruttan, V. W., and Wilcke, H. L. (1985).
“A Survey of U.S. Agricultural Research by Private Industry III”. Agricultural Research
Institute.
Crow, J. F. (1998). 90 years ago: The beginning of hybrid maize. Genetics 148, 923–928.
Cunha Fernandes, J. S., and Franzon, J. F. (1997). Thirty years of genetic progress in maize
(Zea mays L.) in a tropical environment. Maydica 42, 21–27.
Daberkow, S., Taylor, H., and Huang, W.-Y. (2000). “Agricultural Resources and Environ-
mental Indicators: Nutrient use and Management”. Rep. No. AH722. USDA/ERS,
Washingon, DC.
Derieux, M., Darrigrand, M., Gallais, A., Barriere, Y., Bloc, D., and Montalant, Y. (1987).
Estimation du progres genetique realise chez le maıs grain en France entre 1950 et 1985.
Agronomie 7, 1–11.
Dodd, J. (2000). How to foresee corn disease outbreaks. In “55th Annual Corn & Sorghum
Industry Research Conference, December 6–8, 2000”, Vol. 55, pp. 91–98. American Seed
Trade Association, Chicago, IL.
Donald, C. M. (1968). The breeding of crop ideotypes. Euphytica 17, 385–403.
Duvick, D. N. (1977). Genetic rates of gain in hybrid maize yields during the past 40 years.
Maydica 22, 187–196.
Duvick, D. N. (1984a). Genetic contributions to yield gains of U.S. hybrid maize, 1930 to 1980.
In “Genetic Contributions to Yield Gains of Five Major Crop Plants. CSSA Special
Publication No. 7” (W. R. Fehr, Ed.), pp. 15–47. Crop Science Society of America,
American Society of Agronomy, Madison, WI.
Duvick, D. N. (1984b). Genetic diversity in major farm crops on the farm and in reserve. Econ.
Bot. 38, 161–178.
Duvick, D. N. (1992). Genetic contributions to advances in yield of U.S. maize. Maydica 37,
69–79.
Duvick, D. N. (1997). What is yield? In “Developing Drought- and Low N-Tolerant Maize.
Proceedings of a Symposium, March 25–29, 1996, CIMMYT, El Batan, Mexico” (G. O.
Edmeades, M. Banziger, H. R. Mickelson, and C. B. Pena–Valdivia, Eds.), pp. 332–335.
CIMMYT, Mexico, D.F.
Duvick, D. N. (1999). Heterosis: Feeding people and protecting natural resources. In “The
Genetics and Exploitation of Heterosis in Crops” (J. G. Coors and S. Pandey, Eds.),
pp. 19–29. American Society of Agronomy, Inc., Crop Science Society of America, Inc.,
Soil Science Society of America, Inc., Madison, WI.
Duvick, D. N., and Cassman, K. G. (1999). Post-green revolution trends in yield potential of
temperate maize in the north-central United States. Crop Sci. 39, 1622–1630.
Duvick, D. N., Smith, J. S. C., and Cooper, M. (2004a). Changes in performance, parentage,
and genetic diversity of successful corn hybrids, from 1930 to 2000. In “Corn: Origin,

140 DONALD N. DUVICK
History, Technology and Production” (C. W. Smith, J. Betran, and E. Runge, Eds.),
pp. 65–97. Wiley, New York.
Duvick, D. N., Smith, J. S. C., and Cooper, M. (2004b). Long-term selection in a commercial
hybrid maize breeding program. In “Plant Breeding Reviews” (J. Janick, Ed.), Vol. 24,
pp. 109–151. Wiley, New York.
Dwyer, L. M., Stewart, D. W., and Tollenaar, M. (1992). Analysis of maize leaf photosynthesis
under drought stress. Can. J. Plant Sci. 72, 477–481.
Dwyer, L. M., and Tollenaar, M. (1989). Genetic improvement in photosynthetic response of
hybrid maize cultivars, 1959 to 1988. Can. J. Plant Sci. 69, 81–91.
Dwyer, L. M., Tollenaar, M., and Stewart, D. W. (1991). Changes in plant density dependence
of leaf photosynthesis of maize (Zea mays L.) hybrids, 1959 to 1988. Can. J. Plant Sci. 71,
1–11.
Eberhart, S. A., Debela, S., and Hallauer, A. R. (1973). Reciprocal recurrent selection in BSSS
and BSCB1 maize varieties and half sib selection in BSSS. Crop Sci. 13, 451–456.
Eberhart, S. A., and Russell, W. A. (1966). Stability parameters for comparing varieties. Crop
Sci. 6 , 36–40.
Edmeades, G. O., Ba nziger, M., and M. Cortes, C. (1997). From stress-tolerant populations to
hybrids: The role of source germplasm. In “Developing Drought– and Low N-Tolerant
Maize: Proceedings of a Symposium, March 25–29, 1996, CIMMYT, El Bata n, Mexico”
(G. O. Edmeades, M. Ba nziger, H. R. Mickelson, and C. B. Pen a-Valdiva, Eds.),
pp. 263–272. CIMMYT, Mexico, D.F.
Edmeades, G. O., Schussler, J., Campos, H., Zinselmeier, C., Habben, J., Collinson, S., Cooper, M.,
Hoffbeck, M., and Smith, O. (2003). Increasing the odds of success in selecting for abiotic stress
tolerance in maize. In “Versatile Maize—Golden Opportunities: Proceedings 5th Australian
Maize Conference, Toowoomba, 18–20 Feb. 2003” (C. J. Birch and S. R. Wilson, Eds.),
pp. 16–28. Maize Association of Australia.
Edmeades, G. O., and Tollenaar, M. (1990). Genetic and cultural improvements in maize
production. In “Proceedings of the International Congress of Plant Physiology ” (S. K.
Sinha, P. V. Sane, S. C. Bhargava, and P. K. Agrawal, Eds.), pp. 164 –180. Society for Plant
Physiology and Biochemistry, New Delhi, India.
Edwards, J. W., and Lamkey, K. R. (2002). Quantitative genetics of inbreeding in a synthetic
maize population. Crop Sci. 42, 1094–1104.
Emrich, S. J., Aluru, S., Fu, Y., Wen, T.-J., Narayanan, M., Guo, L., Ashlock, D. A., and
Schnable, P. S. (2004). A strategy for assembling the maize (Zea mays L.) genome.
Bioinformatics 20, 140–147.
ERS (2003). Data: Adoption of genetically engineered crops in the U.S. [Online]. Available by
Economic Research Service/USDA http://www.ers.usda.gov/data/BiotechCrops/ (viewed
November 20, 2003).
Eyherabide, G. H., and Damilano, A. L. (2001). Comparison of genetic gain for grain yield of
maize between the 1980s and 1990s in Argentina. Maydica 46, 277–281.
Eyherabide, G. H., Damilano, A. L., and Colazo, J. C. (1994). Genetic gain for grain yield of
maize in Argentina. Maydica 39, 207–211.
FAO (1996). “The State of the World’s Plant Genetic Resources for Food and Agriculture”.
Food and Agriculture Organization of the United Nations, Rome, Italy.
Fasoula, V. A., and Fasoula, D. A. (2000). Honeycomb breeding: Principles and applications.
In “Plant Breeding Reviews” (J. Janick, Ed.), Vol. 18, pp. 177–250. Wiley, New York.
Fernandez-Cornejo, J. (2004). “The Seed Industry in U.S. Agriculture: An Exploration of Data
and Information on Crop Seed Markets, Regulation, Industry Structure, and Research and
Development”. Rep. No. 786. USDA–ERS, Washington, DC.
Fernandez-Cornejo, J., and McBride, W. D. (2002). “Adoption of Bioengineered Crops”. Rep.
No. 810. Economic Research Service/USDA, Washington, DC.

ADVANCES IN MAIZE 141
Frei, O. M. (2000). Changes in yield physiology of corn as a result of breeding in northern
Europe. Maydica 45, 173–183.
Frey, K. J. (1996). “National plant breeding study. I. Human and financial resources devoted to
plant breeding research and development in the United States in 1994 ”. Rep. No. 98. Iowa
Agriculture and Home Economics Experiment Station, Ames, IA.
Fu, H., and Dooner, H. K. (2002). Intraspecific violation of genetic colinearity and its implica-
tions in maize. Proc. Natl. Acad. Sci. USA 99, 9573–9578.
Gianessi, L. P., and Carpenter, J. E. (1999). “Agricultural Biotechnology: Insect Control
Benefits”. National Center for Food and Agricultural Policy, Washington, DC.
Gollin, D., and Smale, M. (1998). Valuing genetic diversity: crop plants and agroecosystems.
In “Biodiversity in Agroecosystems” (W. Collins and C. O. Qualset, Eds.), pp. 237–265.
CRC Press LLC, Boca Raton, FL.
Goodman, M. M., and Brown, W. L. (1988). Races of corn. In “Corn and Corn Improvement” (G.
F. Sprague and J. W. Dudley, Eds.), pp. 33–79. American Society of Agronomy, Madison, WI.
Grobman, A., Salhuana, W., Sevilla, R., and Mangelsdorf, P. (1961). “Races of Maize in Peru:
Their Origins, Evolution, and Classification”. National Academy of Sciences – National
Research Council, Washington, DC.
Hagdorn, S., Lamkey, K. R., Frisch, M., Guimaraes, P. E. O., and Melchinger, A. E. (2003).
Molecular genetic diversity among progenitors and derived elite lines of BSSS and BSCB1
maize populations. Crop Sci. 43, 474–482.
Hallauer, A. R. (1999). Heterosis: What have we learned? What have we done? Where are we
headed? In “Genetics and Exploitation of Heterosis in Crops” (J. G. Coors and S. Pandey,
Eds.), pp. 483–492. American Society of Agronomy, Inc., Crop Science Society of America,
Inc., Soil Science Society of America, Inc., Madison, WI.
Hallauer, A. R., and Miranda, J. B. (1988). “Quantitative Genetics in Maize Breeding”, 2nd Ed.,
Iowa State University Press, Ames, IA.
Hallauer, A. R., Russell, W. A., and Lamkey, K. R. (1988). Corn breeding. In “Corn and Corn
Improvement” (G. F. Sprague and J. W. Dudley, Eds.), pp. 469–565. American Society of
Agronomy, Inc., Crop Science Society of America, Inc., Soil Science Society of America,
Inc., Madison, WI.
Hayes, H. K. (1952). Development of the heterosis concept. In “Heterosis” (J. W. Gowen, Ed.),
pp. 49–65. Iowa State College Press, Ames, IA.
Hetherington, P., Reynolds, T., Marshall, G., and Kirkwood, R. (1999). The absorption,
translocation and distribution of the herbicide glyphosate in maize expressing the CP-4
transgene. J. Exp. Bot. 50, 1567–1576.
Iowa State University (2003). University Extension Weather Information: Iowa Averages,
Climatology Data [Online]. Available by ISU Extension http://mesonet.agron.iastate.edu/
climodat/ (viewed November 6, 2003).
James, C. (2002). “Preview: Global Status of Commercialized Transgenic Crops 2002. ISAAA
Briefs”. Rep. No. 27. ISAAA, Ithaca, NY.
James, C. (2003a). “Global Review of Commercialized Transgenic Crops: 2002 Feature: Bt
maize”. The International Service for the Acquisition of Agri-biotech Applications
(ISAAA), Ithaca, NY.
James, C. (2003b). “Preview: Global Status of Commercialized Transgenic Crops: 2003. ISAAA
Briefs”. Rep. No. 30. ISAAA, Ithaca, NY.
Jansen, R. C., Jannink, J.-L., and Beavis, W. D. (2003). Mapping quantitative trait loci in plant
breeding populations: Use of parental haplotype sharing. Crop Sci. 43, 829–834.
Kanampiu, F.K.,Kabambe, V.,Massawe, C., Jasi, L., Friesen,D., Ransom, J.K., andGressel, J.
(2003).Multi-site, multi-season field tests demonstrate that herbicide seed-coating herbicide-
resistance maize controls Striga spp. and increases yields in several African countries. Crop
Prot. 22, 697–706.

142 DONALD N. DUVICK
King, S. R., and Hagood, E. S. (2003). The effect of johnsongrass ( Sorghum halepense) control
method on the incidence and severity of virus diseases in glyphosate-tolerant corn (Zea
mays). Weed Technol. 17, 503–508.
Klenke, J. R., Russell, W. A., and Guthrie, W. D. (1986). Recurrent selection for resistance to
European corn borer in a corn synthetic and correlated effects on agronomic traits. Crop
Sci. 26 , 864–868.
Lambert, R. J., Alexander, D. E., and Han, Z. J. (1998). A high oil pollinator enhancement of
kernel oil and effects on grain yields of maize hybrids. Agron. J. 90, 211–215.
Lamkey, K. R. (1992). Fifty years of recurrent selection in the Iowa Stiff Stalk Synthetic maize
population. Maydica 37, 19–28.
Lawrence, C. J., Dong, Q., Polacco, M. L., Seigfried, T. E., and Brendel, V. (2004). MaizeGDB,
the community database for maize genetics and genomics. Nucleic Acids Res. 32, 393–397.
Malin, J. C. (1944). “Winter Wheat in the Golden Belt of Kansas”. University of Kansas Press,
Lawrence, KS.
McCullough, D. E., Aguilera, A., and Tollenaar, M. (1994a). N uptake, N partitioning, and
photosynthetic N-use efficiency of an old and a new maize hybrid. Can. J. Plant Sci. 74,
479–484.
McCullough, D. E., Giradin, P., Mihajlovic, M., Aguilera, A., and Tollenaar, M. (1994b).
Influence of N supply on development and dry matter accumulation of an old and a new
maize hybrid. Can. J. Plant Sci. 74, 471–477.
Meghji, M. R., Dudley, J. W., Lambert, R. J., and Sprague, G. F. (1984). Inbreeding depression,
inbred and hybrid grain yields, and other traits of maize genotypes representing three eras.
Crop Sci. 24, 545–549.
Miquel, P. (2001). “La France et ses paysans: Une histoire du monde rural au XXe sie cle”.
L’Archipel, Paris.
National Research Council (U.S.) Committee on Genetic Vulnerability of Major Crops (1972).
“Genetic Vulnerability of Major Crops”. National Academy of Sciences, Washington, DC.
Ng, K.-Y., Pollak, L. M., Duvick, S. A., and White, P. J. (1997). Thermal properties of starch
from 62 exotic maize (Zea mays L.) lines grown in two locations. Cereal Chem. 74, 1997.
Nissanka, S. P., Dixon, M. A., and Tollenaar, M. (1997). Canopy gas exchange response to
moisture stress in old and new maize hybrid. Crop Sci. 37, 172–181.
Ostlie, K. R., Hutchison, W. D., and Hellmich, R. L. (2002). “Bt Corn and European Corn
Borer: Long-Term Success Through Resistance Management”. Rep. No. BU-07055-GO
1997. Regents of the University of Minnesota, St. Paul, MN.
Ostrander, B. O., and Coors, J. G. (1997). Relationship between plant composition and
European corn borer resistance in three maize populations. Crop Sci. 37, 1741–1745.
Paszkiewicz, S., and Butzen, S. (2001). Crop Insights Vol. 11, No. 6: Corn Hybrid Response to
Plant Population [Online]. Available by Pioneer Hi-Bred International, Inc. http://www.
pioneer.com/usa/agronomy/corn/1106.htm (viewed January 13, 2004).
Paterniani, E., and Goodman, M. M. (1977). “Races of Maize in Brazil and Adjacent Areas”.
CIMMYT, Mexico D.F.
Peferoen, M. (1992). Engineering of insect-resistant plants with Bacillus thuringiensis
crystal protein genes. In “Plant Genetic Manipulation for Crop Protection” (A. M. R.
Gatehouse, V. A. Hilderand, and D. Boulter, Eds.), Vol. 7, pp. 135–153. CAB Internation-
al, Wallingford, UK.
Peng, S., Cassman, K. G., Virmani, S. S., Sheehy, J., and Khush, G. S. (1999). Yield potential
trends of tropical rice since the release of IR8 and the challenge of increasing rice yield
potential. Crop Sci. 39, 1552–1559.
Peng, S., Khush, G. S., and Cassman, K. G. (1994). Evolution of the new plant type for
increased yield potential. In “Breaking the Yield Barrier: Proceedings of a Workshop on

ADVANCES IN MAIZE 143
Rice Yield Potential in Favorable Environments, IRRI, 29 November–4 December 1993”,
pp. 5–20. IRRI, Manila, Phillipines.
Reynolds, M. P., Rajaram, S., and Sayre, K. D. (1999). Physiological and genetic changes of
irrigated wheat in the post-green revolution period and approaches for meeting projected
global demand. Crop Sci. 39, 1611–1621.
Rice, M. E. (1997). Perception and performance of Bt corn. In “Proceedings of the 52nd Annual
Corn and Sorghum Research Conference”, Vol. 52, pp. 144–156. American Seed Trade
Association, Washington, DC.
Rice, M. E., Tollefson, J. J., and Oleson, J. (2003). Corn rootworm management with Yield-
Gard rootworm corn: This changes everything. In “Proceedings 15th Annual Integrated
Crop Management Conference, Iowa State University, Ames, Iowa ”, Vol. 15, pp. 75–84.
Iowa State University, University Extension, Ames, IA.
Richey, F. D. (1922). The experimental basis for the present status of corn breeding. J. Am. Soc.
Agron. 14 , 1–17.
Rosegrant, M. W., Cai, X., and Cline, S. A. (2002). “Global Water Outlook to 2025: Averting
an Impending Crisis”. International Food Policy Research Institute and International
Water Management Institute, Washington, DC and Colombo, Sri Lanka.
Rosegrant, M. W., Paisner, M. S., Meijer, S., and Witcover, J. (2001). “Global Food Projections
to 2020: Emerging Trends and Alternative Futures”. International Food Policy Research
Institute, Washington, DC.
Rosenow, D. T., and Clark, L. E. (1987). Utilization of exotic germplasm in breeding for
yield stability. In “Fifteenth Biennial Grain Sorghum Research and Utilization Conference,
15–17 Feb. 1987” (R. R. Duncan, Ed.), pp. 49–56. National Grain Sorghum Producers
Association, Abernathy, TX, Lubbock, TX.
Runge, C. F., and Ryan, B. (2003). The Economic Status and Performance of Plant Biotech-
nology in 2003: Adoption, Research and Development in the United States [Online].
Available by Council for Biotechnology Information (CBI) report at http://www.apec.
umn.edu/faculty/frunge/plantbiotech.pdf (viewed February 2, 2004).
Russell, W. A. (1974). Comparative performance for maize hybrids representing different eras
of maize breeding. In “29th Annual Corn and Sorghum Research Conference”, Vol. 29,
pp. 81–101. American Seed Trade Association, Chicago, IL.
Russell, W. A. (1984). Agronomic performance of maize cultivars representing different eras of
breeding. Maydica 29, 375–390.
Russell, W. A. (1985). Evaluations for plant, ear, and grain traits of maize cultivars representing
seven eras of breeding. Maydica 30, 85–96.
Russell, W. A. (1991). Genetic improvement of maize yields. Adv. Agron. 46, 245–298.
Russell, W. A. (1993). Achievements of maize breeders in North America. In “International
Crop Science I” (P. S. Baenziger, Ed.), pp. 225–233. Crop Science Society of America, Inc.,
Madison, WI.
Schnell, F. W. (1974). Trends and problems in breeding methods for hybrid corn. In “XVI
British Poultry Breeders Roundtable”, Vol. 16, pp. 86–98. Birmingham, England.
Shou, H., Bordallo, P., Fan, J.-B., Yeakley, J. M., Bibikova, M., Sheen, J., and Wang, K.
(2004). Expression of an active tobacco mitogen-activated protein kinase enhances freezing
tolerance in transgenic maize. Proc. Natl. Acad. Sci. USA 101, 3298–3303.
Shull, G. H. (1952). Beginnings of the heterosis concept. In “Heterosis” (J. W. Gowen, Ed.),
pp. 14–48. Iowa State College Press, Ames, IA.
Simmonds, N. W. (1962). Variability in crop plants, its use and conservation. Biol. Rev. 37,
422–465.
Simmonds, N. W. (1985). A plant breeder’s perspective of durable resistance. FAO Plant Prot.
Bull. 33, 13–17.

144 DONALD N. DUVICK
Smale, M., Reynolds, M. P., Warburton, M., Skovmand, B., Trethowan, R., Singh, R. P., Ortiz-
Monasterio, I., and Crossa, J. (2002). Dimensions of diversity in modern spring bread
wheat in developing countries from 1965. Crop Sci. 42, 1766–1779.
Soper, J., Judd, D., Schmidt, D., and Sullivan, S. (2003). The future of biotechnology in
soybeans. Ag. Bio. Forum 6, 8–10.
Sprague, G. F. (1946). Early testing of inbred lines of corn. J. Am. Soc. Agron. 38, 108–117.
Sprague, G. F. (1952). Early testing and recurrent selection. In “Heterosis” (J. W. Gowen, Ed.),
pp. 400–417. Iowa State College Press, Ames, IA.
Sprague, G. F. (1966). Quantitative genetics in plant improvement. In “Plant Breeding” (K. J.
Frey, Ed.), pp. 315–355. Iowa State University Press, Ames, IA.
Sprague, G. F. (1994). Recorded Interview with George F. Sprague (D. N. Duvick, Ed.).
Johnston, IA.
Sriwatanapongse, S., Chutkaew, C., Jampathong, S., Paisuwan, B., Aekatasanawan, C., and
Jihnayon, S. (1985). Long-term improvement of a maize composite, Suwan I.
In “Proceedings of the Fifth International Congress of Society for the Advancement of
Breeding Researches in Asia and Oceania (SABRAO), November 25 –29, 1985”,
pp. 257–261. Kasetsart University, Bangkok, Thailand.
Swaminathan, M. S. (1998). Crop production and sustainable food security. In “Crop Produc-
tivity and Sustainability: Shaping the Future: Proceedings of the 2nd International Crop
Science Congress” (V. L. Chopra, R. B. Singh, and A. Varma, Eds.), pp. 3–18. Oxford &
IBH Publishing Company. Pvt. Ltd., New Delhi.
Taha, F. A. (2003). The Poultry Sector in Middle-Income Countries and Its Feed Requirements:
The Case of Egypt [Online]. Available by Economic Research Service, USDA http://www.
ers.usda.gov/publications/WRS03/dec03/wrs0302/ (viewed January 7, 2004).
Tatum, L. A. (1971). The southern corn leaf blight epidemic. Science 171, 1113–1116.
Tokatlidis, I. S., Koutsika-Sotiriou, M., and Fasoulas, A. C. (2001). The development of
density-independent hybrids in maize. Maydica 46, 21–25.
Tollenaar, M. (1989). Genetic improvement in grain yield of commercial maize hybrids grown in
Ontario from 1959 to 1988. Crop Sci. 29, 1365–1371.
Tollenaar, M. (1991). Physiological basis of genetic improvement of maize hybrids in Ontario
from 1959 to 1988. Crop Sci. 31, 119–124.
Tollenaar, M., Aguilera, A., and Nissanka, S. P. (1997). Grain yield is reduced more by weed
interference in an old than in a new maize hybrid. Agron. J. 89, 239–246.
Tollenaar, M., McCullough, D. E., and Dwyer, L. M. (1994). Physiological basis of the genetic
improvement of corn. In “Genetic Improvement of Field Crops” (G. A. Slafer, Ed.),
pp. 183–236. Dekker, New York.
Tollenaar, M., and Mihailovic, M. (1991). Bromoxynil tolerance during the seedling phase is
associated with genetic grain yield improvement in maize. Can. J. Plant Sci. 71, 1021–1027.
Tollenaar, M., Ying, J., and Duvick, D. N. (2000). Genetic gain in corn hybrids from
the northern and central corn belt. In “55th Annual Corn and Sorghum Research Confer-
ence”, Vol. 55, pp. 53–62. American Seed Trade Association, Chicago.
Traore, S. B., Carlson, R. E., Pilcher, C. D., and Rice, M. E. (2000). Bt and non-Bt maize
growth and development as affected by temperature and drought stress. Agron. J. 92,
1027–1035.
Troyer, A. F. (1996). Breeding widely adapted, popular maize hybrids. Euphytica 92, 163–174.
Troyer, A. F. (2000). Temperate corn: Background, behavior, and breeding. In “Specialty
Corns” (A. R. Hallauer, Ed.), pp. 393–466. CRC Press, Washington, DC.
Tuberosa, R., Salvi, S., Sanguineti, M. C., Landi, P., Maccaferri, M., and Conti, S. (2002).
Mapping QTLs regulating morpho-physiological traits and yield: Case studies, shortcom-
ings and perspectives in drought-stressed maize. Ann. Bot. 89, 941–963.

ADVANCES IN MAIZE 145
Turning Point Project (1999a). Biotechnology ¼ Hunger (paid advertisement by Turning
Point Project on behalf of 22 non–governmental organizations). In “The New York
Times”, p. A5, New York.
Turning Point Project (1999b). Who plays God in the 21st century? (paid advertisement by
Turning Point Project on behalf of 19 non-governmental organizations). In “The New York
Times”, p. A11, New York.
USDA (1944–1962). “Agricultural Statistics”. United States Government Printing Office,
Washington, DC.
USDA (1949–1992). “Agricultural Statistics”. United States Government Printing Office,
Washington, DC.
USDA (1956). “Agricultural Statistics 1956”. United States Government Printing Office,
Washington, DC.
USDA-ERS (2003). Fertilizer Use and Price Statistics [Online]. Available by USDA Economics
and Statistics System http://usda.mannlib.cornell.edu/ (viewed February 3, 2004).
USDA-NASS (2003a). Agricultural Statistics Data Base [Online]. Available by USDA–NASS
http://www.nass.usda.gov:81/ipedb/ (viewed February 2, 2004).
USDA-NASS (2003b). Corn: Acreage, Utilization, Price, and Value of Production. United
States, 1866 to Date [Online]. Available by USDA-National Agricultural Statistical Service
http://www.usda.gov/nass/pubs/trackrec/track03a.htm (viewed February 2, 2004).
van der Plank, J. E. (1963). “Plant Diseases: Epidemics and Control”. Academic Press, New
York.
Wallace, H. A. (1930). Practical Aspects of the Newer Methods of Corn Breeding, p. 5
(unpublished manuscript). The Hi-Bred Corn Company, Des Moines, IA.
Wallace, H. A., and Brown, W. L. (1988). “Corn and Its Early Fathers”. Revised/Ed. Iowa State
University Press, Ames, IA.
Whitt, S. R., Wilson, L. M., TenaIllon, M. I., Gaut, B. S., and IV, E. S. B. (2002). Genetic
diversity and selection in the maize starch pathway. Proc. Natl. Acad. Sci. USA 99,
12959–12962.