dodecameric structure and atpase activity of the · pdf filedodecameric structure and atpase...
TRANSCRIPT

doi:10.1016/j.jmb.2006.11.030 J. Mol. Biol. (2007) 366, 179–192
Dodecameric Structure and ATPase Activity of theHuman TIP48/TIP49 Complex
Teena Puri1†, Petra Wendler2†, Barbara Sigala1, Helen Saibil2
and Irina R. Tsaneva1⁎
1Department of Biochemistryand Molecular Biology,University College London,London WC1E 6BT, UK2School of Crystallography,Birkbeck College,University of London,London WC1E 7HX, UK† T. P. and P. W. contributed equaPresent address: B. Sigala, UCL In
Royal Free Medical School, RowlandNW3 2PF, UK.Abbreviations used: TIP, TBP/tra
interacting protein; IPTG,isopropyl-1-thio-β-D-galactopyranosphenylmethylsulphonyl fluoride; BSalbumin; ss, single-stranded; ds, douATPases associated with different ceelectron microscopy; snoRNP, smallribonucleoprotein complex.E-mail address of the correspondi
0022-2836/$ - see front matter © 2006 E
TIP48 and TIP49 are two related and highly conserved eukaryotic AAA+
proteins with an essential biological function and a critical role in majorpathways that are closely linked to cancer. They are found together ascomponents of several highly conserved chromatin-modifying complexes.Both proteins show sequence homology to bacterial RuvB but the natureand mechanism of their biochemical role remain unknown. Recombinanthuman TIP48 and TIP49 were assembled into a stable high molecular massequimolar complex and tested for activity in vitro. TIP48/TIP49 complexformation resulted in synergistic increase in ATPase activity but ATPhydrolysis was not stimulated in the presence of single-stranded, double-stranded or four-way junction DNA and no DNA helicase or branchmigration activity could be detected. Complexes with catalytic defects ineither TIP48 or TIP49 had no ATPase activity showing that both proteinswithin the TIP48/TIP49 complex are required for ATP hydrolysis. Thestructure of the TIP48/TIP49 complex was examined by negative stainelectron microscopy. Three-dimensional reconstruction at 20 Å resolutionrevealed that the TIP48/TIP49 complex consisted of two stacked hexamericrings with C6 symmetry. The top and bottom rings showed substantialstructural differences. Interestingly, TIP48 formed oligomers in the presenceof adenine nucleotides, whilst TIP49 did not. The results point tobiochemical differences between TIP48 and TIP49, which may explain thestructural differences between the two hexameric rings and could besignificant for specialised functions that the proteins perform individually.
© 2006 Elsevier Ltd. All rights reserved.
Keywords: TIP48 (RUVBL2); TIP49 (RUVBL1); AAA+ protein; dodecamer;ATPase
*Corresponding authorlly to this work.stitute of Hepatology,Hill Street, London
nscription activating
ide; PMSF,A, bovine serumble-stranded; AAA+,llular activities; EM,nucleolar
ng author:
lsevier Ltd. All rights reserve
Introduction
TIP48 (RUVBL2) and TIP49 (RUVBL1) are twoclosely related and highly conserved eukaryoticproteins discovered by different groups.1–10 Theyare members of the AAA+ family (ATPases asso-ciated with different cellular activities+)11 and showmoderate homology to the bacterial RuvB, the ATP-dependent motor of the RuvAB complex that drivesbranch migration of Holliday junctions.12 TIP48and TIP49 are found in large nuclear complexes,such as chromatin modifying complexes,4,9,13–16transcription-activating complexes1,2,10,17–19 andsmall nucleolar ribonucleoprotein complexes(snoRNPs).20–22 In addition, TIP48 and TIP49 co-localizewith centrosomes and elements of themitoticapparatus in mitosis.23,24 Both proteins are essentialfor viability in yeast,5,8 Drosophila melanogaster1 and
d.

180 Dodecameric TIP48/TIP49 Complex
Caenorhabditis elegans‡ and are likely to have similarimportant roles in humans.Based on some sequence homology with RuvB it is
speculated that TIP48 and TIP49 may be potentialDNA translocation or branch migration motors ineukaryotes, similar to RuvB.3,6,7,25 However, thedata reporting DNA helicase activities of TIP485 andTIP4925 are contradictory.4,8 ATP hydrolysis isclearly essential for the biological activity of bothproteins10,20,26,27 but their biochemical functions inthe cell remain unknown.TIP48 and TIP49 interact and form a complex in
vitro and in vivo.1,3–5,10 With few exceptions, the twoproteins are found together in chromatin remodel-ling complexes (INO809,28,29 p40013 and SWR116,30),the mammalian histone acetylase complex TIP604,31
and with c-Myc.10 Within the INO80 complex, TIP48and TIP49 are present in a 6:1 stoichiometry relativeto other components of the complex.9 INO80 com-plexes purified from yeast cells depleted of scTIP48(Rvb1p) also lacked scTIP49(Rvb2p)27 indicatingthat the two proteins need to be incorporatedtogether into the INO80 complex. AAA+ proteinsoften function as molecular motor proteins in largemolecular machines that drive the assembly, opera-tion and disassembly of protein complexes.11 It hasbeen speculated that TIP48 and TIP49 (together andindividually) function in remodelling specific pro-tein–protein or protein–DNA interactions, andevidence for this has emerged through studies ofboth the INO80 complex27 and snoRNP assembly.32
As well as functioning together, the two proteinsmay have specialised individual roles. TIP48 andTIP49 exhibit antagonistic effects on β-catenin-mediated transcription,1 heart development,33 andgene expression.34 TIP48 interacts with and facil-itates the function of the transcription activatingprotein ATF2.17 TIP48, but not TIP49, accumulates atthe midzone and mid-body of dividing HeLa cells.24
In this work we examined the molecular archi-tecture of the TIP48/TIP49 complex and thecontribution of the individual proteins to theATPase activity of the complex. The TIP48/TIP49complex assembled from recombinant proteinsshowed a synergistic increase in ATPase activity,compared to that of the constituent proteins, asreported.4 In the TIP48/TIP49 complex, both pro-teins bind ATP and the catalytic activity of both isessential for ATP hydrolysis. DNA had no effect onthe ATPase activity of the complex. Electron micro-scopy and 3D reconstruction of the complex showedtwo distinct stacked hexameric rings with C6symmetry. Interestingly, TIP48 formed oligomersin the presence of adenine nucleotides, whilst TIP49did not. These results point to biochemical differ-ences between the two components, which mayexplain the structural differences between the twohexameric rings and could be significant forspecialised functions that the proteins performindividually.
‡http://www.wormbase.org/
Results
Purification and ATPase activity of theTIP48/TIP49 complex
The TIP48/TIP49 complex was assembled frompurified TIP48-His6 and partially purified TIP49 (seeMaterials and Methods). The proteins were mixed ata twofold excess of the non-tagged TIP49 overTIP48-His6 and applied to a TALON™metal affinitycolumn. Fractions eluting from the column con-tained almost equal amounts of both TIP49 andTIP48-His6. The presence of the proteins in differentfractions and their positions on the gel were verifiedusing specific antibodies.24 Most of the His-taggedTIP48 was found associated with TIP49, whichdemonstrates the formation of a stable TIP48/TIP49 complex.The TIP48/TIP49 complex was fractionated by
size exclusion chromatography on a Superdex 200HR column. In these experiments it was not possibleto record the protein elution profile as absorbance at280 nm due to the inclusion of 0.05% (v/v) TritonX-100 in the chromatography buffer, used to preventnon-specific adsorption and loss of protein. Theprotein composition of the fractions was examinedby SDS– PAGE using equal protein loading (∼ 2 μg)as shown in Figure 1(a). TIP48 and TIP49 co-elutedin the high molecular range of the column infractions 8 to 10. Fraction 9 contained equal amountsof TIP48 and TIP49 as judged by densitometry of theprotein bands on the gel. This fraction correspondsto a range of molecular masses from 850 kDa to650 kDa. From fraction 10 onwards, the amount ofTIP49 decreased and fractions 13 and 14 containedTIP48-His6 only (upper band), as verified byimmunoblotting. Therefore, the TIP48/TIP49 com-plex was present as a high molecular mass oligomer.It should be emphasised that the complex wasassembled without addition of nucleotide cofactors.While the presence of tightly bound nucleotides inthe recombinant proteins cannot be ruled outcompletely, both proteins had been dialysed againstEDTA-containing buffers during purification andthe assembled complex fractionated by size exclu-sion chromatography in EDTA-containing buffer.On their own, purified TIP48 and TIP49 elutedpredominantly as monomers under these conditions(see below).The ATPase activities of the fractions from size
exclusion chromatography are shown in Figure 1(b).Fraction 9, containing the equimolar TIP48/TIP49complex, showed significantly higher ATPase activ-ity, than later fractions containing reducing amountsof TIP49. Fraction 13, which consisted of TIP48 only,had approximately ten times lower ATPase activitycompared to fraction 9, while ATPase activity wasundetectable in fraction 14. For direct comparisonwith the TIP48/TIP49 complex we tested purifiedrecombinant TIP48-His6 and His6-TIP49 (see Mate-rials and Methods) fractioned by Superdex 200FPLC as monomers. The activity of purified TIP48

Figure 1. Composition and ATPase activity of theTIP48-His6/TIP49 complex. (a) Protein composition of the1 ml fractions obtained by Superdex 200 size exclusionchromatography. The TIP48-His6/TIP49 complex cap-tured on TALON™ column (ON) and 2 μg of eachfraction were analysed by SDS–18% PAGE and stainedwith Coomassie brilliant blue. The positions of molecularmass markers (in kDa) are indicated above the gel. (b)ATPase activities of the fractions and purified TIP48 (T48)and TIP49 (T49) proteins, obtained as described inMaterials and Methods. Protein was added to a finalconcentration of 1 μM in terms of TIP48/TIP49 monomers.Reactions were incubated at 37 °C for 30 min. ATPaseactivity is expressed as moles of ATP hydrolyzed per moleof TIP48/TIP49 monomer.
Figure 2. ATP binding and the effect of mutations inthe Walker B motif on the ATPase activity of the TIP48/TIP49 complex. (a) Approximately 1 μg of TIP48/TIP49complex, in 20 μl, were incubated with [γ-32P]2-azido ATPat a final concentration of 10 μM. Reactions wereirradiated with UV light and processed as described inMaterials and Methods. The proteins were fractionated bySDS–18% PAGE. The gel was stained with Coomassiebrilliant blue (CBB), dried and the 32P radioactivitydetected using FLA 2000 phosphorimager. Quantitationof the 32P signal was by phosphorimaging and the inten-sity of the stained protein bands was measured by densi-tometry. (b) Time courses of ATP hydrolysis by wild-typeTIP48/TIP49 complex (squares) and mutant complexesTIP49/TIP48D299N (circles) and TIP48/TIP49D302N(triangles). Reactions contained 1 μM protein (TIP48/TIP49 monomer) and were incubated at 37 °C for the timeindicated.
181Dodecameric TIP48/TIP49 Complex
(T48) was similar to that of fraction 13, whichcontained free TIP48. The activity of purified TIP49(T49) was about five times lower than that of theTIP48/TIP49 complex. The difference between theATPase activity of the complex and TIP49 is likely tobe much greater as TIP49 co-purified with ATPasecontaminants from Escherichia coli through a numberof chromatography steps.35 The results are in fullagreement with the data of Ikura et al. who reportedsimilar levels of ATPase activities both for thecomplex and the individual proteins.4
Within the TIP48/TIP49 complex both TIP48 andTIP49 bind and hydrolyse ATP
Hexameric ring assemblies have been shown tobe the active form of many AAA+ proteins.36–38
There are some examples in eukaryotes wherehexamers assemble from different subunits, includ-ing catalytically inactive subunits (e.g. the MCMcomplex).36,39 Therefore, it is not obvious that bothproteins within the TIP48/TIP49 complex need tobind and hydrolyse ATP. It has been shown in yeastthat the catalytic activity of Rvb1 (TIP49) and Rvb2
(TIP48) are both required for viability.10,20 However,TIP48 and TIP49 play functionally diverse roles andthere are indications that in some instances theircatalytic activity may not be required.27 Moreover,TIP48 and TIP49 may have separate functions ontheir own.24 To test whether both proteins bind ATP,the assembled TIP48/TIP49 complex was incubatedwith the ATP analogue [γ-32P]2 azido-ATP and UV-irradiated to induce cross-linking to the proteins.The labelled ATP analogue cross-linked to bothTIP48 and TIP49 within the complex (Figure 2(a)).Binding was specific to the ATP binding sites as thesignal could be competed out with unlabelled ATP(not shown). Furthermore, the intensities of the 32Psignals from the two bands appear equivalent(Figure 2(a)), suggesting that the two proteins bindequivalent amounts of ATP.

Figure 3. Effect of DNA on the ATPase activity of theTIP48/TIP49 complex. (a) Time courses of ATP hydrolysisby the TIP48/TIP49 complex, in the absence (squares) orpresence (triangles) of 5 ng/μl of φX174 ssDNA. (b) Bargraph representing the ATPase activity of the TIP48/TIP49complex in the presence of different DNA substrates: (1)no DNA; (2) φX174 ssDNA; (3) pUC19 super-coileddsDNA; (4) pUC19 linearised dsDNA; (5) syntheticHolliday junction. DNA was added to a final concentra-tion of 5 ng/μl and the reactions were incubated for20 min at 37 °C. All reactions contained TIP48/TIP49complex at a final concentration of 1 μM, in terms ofTIP48/TIP49 monomers. ATPase activity is expressed asmoles of ATP hydrolyzed per mole of protein monomer.
182 Dodecameric TIP48/TIP49 Complex
In order to test directly the contribution of eachcomponent to the ATPase activity, TIP48/TIP49complexes containing one wild-type and one cataly-tically inactive protein were assembled. A conservedaspartic acid residue in the Walker B motif in eachprotein was mutated to asparagine (TIP48D299N-His6 and TIP49D302N). The equivalent mutationwas shown previously to inactivate TIP49 proteinin vivo10,18,26 and to inhibit the ATPase activity ofthe bacterial RuvB.40 The mutant proteins wereexpressed and purified under the same conditions asthe wild-type proteins and the mutant complexes(TIP48-His6/TIP49D302N and TIP49/TIP48D299N-His6) were assembled and purified as describedabove for the wild-type complex. The results inFigure 2(b) show a steady increase of ATP hyd-rolysed over time for the wild-type complex butATP hydrolysed by the two mutant complexes,TIP48D299N/TIP49 and TIP48/TIP49D302N,remained at background levels. The individualproteins, wild-type or mutants, showed very littleor no ATPase activity when tested in similar experi-ments.35 Within the mutant complexes both proteinsbound [γ-32P]2 azido-ATP (not shown), signifyingthat the mutations block ATP hydrolysis but notbinding. These results indicate that the catalyticactivities of both TIP48 and TIP49 are needed for theATPase activity of the complex.
The ATPase activity of the TIP48/TIP49 complexis not stimulated by DNA
The effect of DNA on the ATPase activity of theassembled TIP48/TIP49 complex was tested usingdifferent forms of DNA including ϕX174 ssDNA,pUC19 supercoiled dsDNA, linearised pUC19dsDNA, and a synthetic four-way Holliday junction(Figure 3). The presence of ssDNA had no effect onthe time course of ATP hydrolysis by the TIP48/TIP49 complex (Figure 3(a)) and no significantstimulation of the ATPase activity could be observedin the presence of any of the different forms of DNAused, as summarized in Figure 3(b). We did not findany stimulation of the ATPase activity in thepresence of DNA when TIP48 or TIP49 were testedindividually either (data not shown). Branch migra-tion and helicase activity were also tested butneither the individual proteins nor the complexdisplayed any of these activities.35 However, thebacterially expressed proteins frequently co-purifiedwith ssDNA-dependent ATPases that could beremoved by ssDNA cellulose chromatography.35
Our results did not reproduce the data published byKanemaki et al. and Makino et al., who reportedrobust ssDNA-dependent ATPase and DNA heli-case activities of opposite polarities for humanTIP48 and rat TIP49 expressed in bacteria.5,25
However, other groups were also unable to detecthelicase or branch migration activities with theindividual proteins4,8 or their complex.4 Signifi-cantly, Makino et al. did not observe ATPase activitywith TIP49 purified from insect cells.25 All theseresults make it unlikely that on their own TIP48,
TIP49, or the TIP48/49 complex can act on DNA invitro.
Effect of nucleotide cofactors on the oligomericstates of TIP48 and TIP49
Fractionation of the TIP48/TIP49 complex by sizeexclusion chromatography clearly indicates that thecomplex contains oligomers of each component. Tounderstand the higher-order architecture of theTIP48/TIP49 complex and its significance, it wasimportant to determine the oligomeric state of theindividual proteins. It was of particular interest totest the effect of adenine nucleotides on thequaternary structure, as some AAA+ proteins, forexample RuvB, require adenine nucleotide cofactorsfor oligomerization.37 Histidine-tagged TIP48-His6and His6-TIP49 were expressed in E. coli andpurified as described in Materials and Methods.
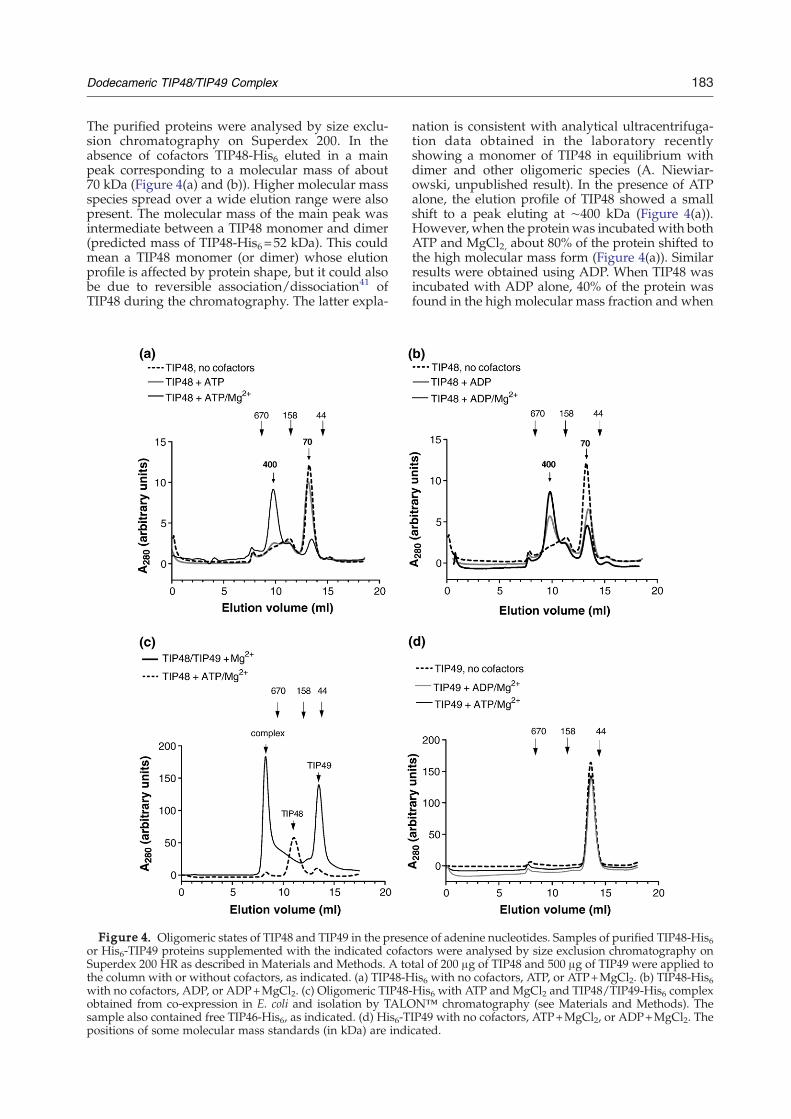
183Dodecameric TIP48/TIP49 Complex
The purified proteins were analysed by size exclu-sion chromatography on Superdex 200. In theabsence of cofactors TIP48-His6 eluted in a mainpeak corresponding to a molecular mass of about70 kDa (Figure 4(a) and (b)). Higher molecular massspecies spread over a wide elution range were alsopresent. The molecular mass of the main peak wasintermediate between a TIP48 monomer and dimer(predicted mass of TIP48-His6=52 kDa). This couldmean a TIP48 monomer (or dimer) whose elutionprofile is affected by protein shape, but it could alsobe due to reversible association/dissociation41 ofTIP48 during the chromatography. The latter expla-
Figure 4. Oligomeric states of TIP48 and TIP49 in the preseor His6-TIP49 proteins supplemented with the indicated cofacSuperdex 200 HR as described in Materials and Methods. A tothe column with or without cofactors, as indicated. (a) TIP48-Hwith no cofactors, ADP, or ADP+MgCl2. (c) Oligomeric TIP48obtained from co-expression in E. coli and isolation by TALOsample also contained free TIP46-His6, as indicated. (d) His6-Tpositions of some molecular mass standards (in kDa) are indi
nation is consistent with analytical ultracentrifuga-tion data obtained in the laboratory recentlyshowing a monomer of TIP48 in equilibrium withdimer and other oligomeric species (A. Niewiar-owski, unpublished result). In the presence of ATPalone, the elution profile of TIP48 showed a smallshift to a peak eluting at ∼400 kDa (Figure 4(a)).However, when the proteinwas incubatedwith bothATP and MgCl2, about 80% of the protein shifted tothe high molecular mass form (Figure 4(a)). Similarresults were obtained using ADP. When TIP48 wasincubated with ADP alone, 40% of the protein wasfound in the high molecular mass fraction and when
nce of adenine nucleotides. Samples of purified TIP48-His6tors were analysed by size exclusion chromatography ontal of 200 μg of TIP48 and 500 μg of TIP49 were applied tois6 with no cofactors, ATP, or ATP+MgCl2. (b) TIP48-His6
-His6 with ATP and MgCl2 and TIP48/TIP49-His6 complexN™ chromatography (see Materials and Methods). TheIP49 with no cofactors, ATP+MgCl2, or ADP+MgCl2. Thecated.

184 Dodecameric TIP48/TIP49 Complex
both ADP andMgCl2 were present, about 65% of theprotein was oligomeric (Figure 4(b)). No oligomer-ization was observed when TIP48 was incubatedwith MgCl2 alone (not shown). These resultsdemonstrate that both ATP and ADP promoteoligomerisation of TIP48, which in the case of ATPrequires Mg2+.A direct comparison of oligomeric TIP48 with
TIP48/TIP49 complex is shown in Figure 4(c). Thecomplex consisted of untagged TIP48 and TIP49-His6 co-expressed in E. coli and captured on TALONmetal affinity column, as described in Materials andMethods. (This procedure improved significantlythe yield of complex and its performance onSuperdex 200 size exclusion chromatography.)
Figure 5. Binding of adenine nucleotides to TIP48 and TIP(d) and (e) were supplemented with 2 mM MgCl2 and [γ-32P]along with increasing concentrations of ATP (a) and (d) or Adescribed in Materials and Methods. Proteins were separated(top panels), dried and the 32P radioactivity detected using FLeach lane was measured from the phosphorimages and the resuused in the binding buffer can be seen on the stained gel, as i
Clearly, the size of the complex differed significantlyfrom that of oligomeric TIP48, which was theexpected size of a TIP48 hexamer (320 kDa in thisexperiment). The higher molecular mass (∼400 kDa)observed in Figure 4(a) and (b) could be due toexperimental differences, such as the Superdex 200column used and calibration, but it could also bedue to variations between different preparationswhich showed different amounts of TIP48 in the70 kDa peak. The size of the TIP48/TIP49 complexwas difficult to determine as it eluted before the670 kDa molecular size standard. A complexcontaining six subunits of each TIP48 and TIP49-His6 would have a predicted molecular mass of608.9 kDa.
49. Reactions containing 1 μg of TIP48 (a) and (b) or TIP492-azido ATP was added to a final concentration of 10 μM,DP (b) and (e), as indicated. Reactions were processed asby SDS–12% PAGE, stained with Coomassie brilliant blueA 2000 phosphorimager (bottom panels). The 32P signal inlts expressed as a percentage of lane 1. (e) and (f) The BSAndicated.

185Dodecameric TIP48/TIP49 Complex
Surprisingly, TIP49 behaved differently. It elutedas a monomer of 55 kDa (expected molecular mass51.2 kDa) and was unable to form stable oligomersin the presence of adenine nucleotides under allconditions tested (Figure 4(d)). These results indi-cate that although TIP48 and TIP49 are highlyhomologous, they differ in their ability to formstable oligomers upon nucleotide binding. To ruleout the possibility that the results were due to failureof the recombinant TIP49 to bind nucleotides, it wasimportant to test for nucleotide binding directly. Ithas been shown before that both TIP49 and TIP48bind ATP5,25 but ADP binding had not been tested.Nucleotide binding experiments using 0.01 mM
[γ-32P]2-azido ATP in competition with unlabelledATP and ADP are shown in Figure 5. The amountsof 32P retained in the presence of non-labelled ATPor ADP competitors were measured from thephosphorimages and are presented as bar charts.In the absence of nucleotide competitor TIP48 binds[γ-32P]2-azido ATP (Figure 5(a), lane 2). When equalconcentrations of ATP and [γ-32P]2-azido ATP wereused (0.01 mM), the 32P signal was clearly reduced(Figure 5(a), lane 3) and decreased further as ATPconcentration increased up to 0.5 mM. Similarobservations were made when ADP was used as acompetitor (Figure 5(b)). However, as seen on thegels and from phosphorimaging (Figure 5(c)), ADPappeared to compete for azido-ATP binding moreefficiently than ATP (compare lanes 3, Figure 5(a)and (b)), indicating that TIP48 may have a higheraffinity for ADP than ATP. Similar results were ob-tained with TIP49 which cross-linked efficiently and
Figure 6. Image analysis of the TIP48/ TIP49 complex negTIP48/TIP49 after low pass filtering. The selection showsdisplayed in (b). (b) Examples of class averages obtained by mu(c) Re-projections of the final 3D structure in the Euler- anglerepresentations of the final 3D structure viewed in the corresp
specifically to [γ-32P]2-azido ATP (Figure 5(d)–(f)).The 32P signal was competed out with increasingconcentrations of both ATP and ADP. As seen withTIP48, TIP49 showed a higher affinity for ADP thanATP (compare lanes 3, Figure 5(d) and (e)). Takentogether, our experiments indicate that both TIP48and TIP49 can bind adenine nucleotides, but onlyTIP48 forms stable oligomers in the presence of thesenucleotides, under the conditions tested.
Structure of TIP48/TIP49 complex determined byelectron microscopy
Samples of the TIP48/TIP49 complex obtainedby size-exclusion chromatography (fraction 9 inFigure 1) were examined by negative stainelectron microscopy and image processing. Thesymmetry analysis of selected top view projectionsafter multiple rounds of alignment by classifi-cation42 suggests that the complex has a 6-foldsymmetry. Applying this symmetry, an initial 3Dstructure was obtained using classification andangular reconstitution in IMAGIC. The map wasrefined in multiple rounds of projection matching inSPIDER and the final reconstruction includes 1765 of2267 particles yielding a resolution of 20 Å. Classaverages of 7–16 images which aligned to the samereference in the last round of refinement as well asindividual images extracted from these classesmatch the equivalent re-projections and 2-D viewsof the reconstruction in detail (Figure 6).The TIP48/49 complex is made up of two
hexameric rings with clearly distinct upper and
atively stained in uranyl acetate. (a) Individual images ofrepresentative members of classes, whose averages areltivariate statistical analysis and classification in IMAGIC.directions assigned to the class averages in (b). (d) Surfaceonding orientations.

186 Dodecameric TIP48/TIP49 Complex
lower layers (Figure 7). The upper ring resemblesclassical AAA+ hexamers such as p97/VCP,whereas the structural organisation in the lowerring is less obvious. A central channel runs throughthe entire complex and is wide enough toaccommodate double-stranded DNA. The heightof the double layer is ∼120 Å along the 6-fold axisand the diameter of the central channel is con-stricted at both ends to ∼30 Å. The maximumdiameter of the complex is ∼160 Å when thedensity threshold is chosen to enclose the expectedmolecular volume. Equal fractions of the molecularvolume are taken up by the upper and lower ring,in accordance with the assumption that equimolarrings of TIP48 and TIP49 assemble to form thecomplex.The structure of TIP49/RuvBL1 has very recently
been determined by X-ray crystallography.43 SinceTIP48 shares 40% sequence identity with TIP49, this
Figure 7. Three-dimensional reconstruction of the TIP48different orientations show the density rendered at a thresholPossible fitted positions for the AAA+ domain of the Thermotogand the human RuvBL1/TIP49 crystal structure (blue) in the lohexameric ring of the TIP proteins. The asterisk marks the unβ-sheet rich TIP48/TIP49 insertion. The hand of the map is u16 Å thick central slice. The central channel narrows to 26 Å at bthe upper hexameric ring.
structure is likely to define all the domains in ourmap. To examine whether the TIP49 structure can beaccommodated in the 3Dmap, the hexameric ring ofTIP49 was manually docked into both rings of themap. It did not fit into the upper ring, and partiallyfitted the lower ring. With a 90° rotation of eachsubunit, a reasonable fit could be achieved to thelower ring (Figure 7). A hexamer of RuvB AAA+
domains,44 based on the oligomeric structure of p97ND1,45 fits into the upper half of the density andallows contact of the arginine finger with the ATPbinding pocket of the adjacent subunit upon ATPbinding.The structure of the complex shows openings
between the subunits of the upper ring, whichsuggests that the packing in this hexamer is loose.This could be due to the absence of boundnucleotide. Further investigations could involvedetermining the structure of the complex in the
/TIP49 complex at 20 Å resolution. (a) Surface views ind accounting for 100% of the expected molecular volume.a maritima RuvB crystal structure44 (red) in the upper ringwer ring demonstrate that each layer can accommodate afilled density in the top ring, which can accommodate thencertain. (b) Side view of the TIP48/TIP49 complex as aoth ends of the complex and extends up to 75 Å in width in

187Dodecameric TIP48/TIP49 Complex
presence of adenine nucleotide cofactors and look-ing for differences in the packing.
Discussion
In agreement with other published data, ourexperiments show that TIP48 and TIP49 form anequimolar complex in vitro, with a molecular massconsistent with a dodecamer of the two proteins.4,27
Electron microscopy studies of negatively stainedpreparations of the TIP48/TIP49 complex revealedparticles containing two stacked hexameric rings.This correlates well with data on other AAA+
proteins which have been shown to form hexamersor double hexamers in vitro, such as RuvB,46
mtMCM47 and other related AAA+ proteins.37,46–51
In our docking, the TIP48/49 AAA+ domains formtwo hexameric rings stacked on top of each other intandem. This arrangement resembles that of pro-teins with two covalently linked AAA domains, likep97514 or the Hsp100 family.52
The composition of the two hexameric rings couldnot be unequivocally determined in this investiga-tion. An equimolar ratio of TIP48 to TIP49 couldrepresent either two homohexamers or mixedhexamers containing equimolar amounts of eachprotein. The electron microscopy (EM) structureshows clear differences between the top and bottomrings and the lower ring in Figure 7 more closelyresembles the TIP49 hexamer of the crystal struc-ture. If the particles contain two heterohexamers ofthe same composition, the structural differencesobserved would reflect different conformationalstates. Two homohexamers, on the other hand,would point to some intrinsic differences betweenTIP48 and TIP49. The results presented here revealdifferences between the abilities of TIP48 and TIP49to form stable oligomers in the presence of nucleo-tides. In the TIP48/TIP49 complex the upperhexamer might be a homohexamer of TIP48, whichcould act as a scaffold promoting the hexameriza-tion of TIP49 in the lower ring. TIP49 in turn appearsto stabilise the hexameric form of TIP48 in theabsence of nucleotides.A 3D reconstruction of TIP48 would have greatly
facilitated the interpretation of the organization ofthe TIP48/TIP49 complex. We did, therefore,attempt to analyse the oligomeric fraction of TIP48by EM. Unfortunately the TIP48 sample did notappear to be homogenous enough for imageprocessing. This may arise from a lower stability ofisolated TIP48 compared to the TIP48/49 complex.Hexamer assembly has been found to stimulate
the ATPase activity of most AAA+ proteins. TheMCM proteins from Saccharomyces cerevisiae showedlittle or no ATPase activity on their own, but whenpart of the complex containing all six subunits, orsub-complexes containing two to three subunits, thisactivity was significantly stimulated.36 A similarobservation was made with the S. cerevisiae RFCclamp loader proteins.53 In both cases, complexformation was essential for activity, because one
subunit provided the ATP binding function throughthe Walker A motif (P-loop) and the other subunitprovided the arginine finger, which is important forcatalysis.38,39,53,54 These ATP hydrolysis sites arefound at the subunit interface of many AAA+
proteins including RuvB54 and explains why theactive form of many AAA+ proteins is hexameric.The synergistic effect on ATPase activity when thetwo hexameric rings assemble into the TIP48/TIP49a complex, however, is less obvious. Mutualstabilisation of the hexameric rings could explainin part the enhanced ATPase activity but clearly theneed for both proteins to be catalytically active andthe synergistic effect on enzymatic activity suggestintimate communications between the two proteinscoupled to conformational effects.The ATPase activity of the TIP48/TIP49 complex
is very low, about 0.5 mol ATP per TIP48 (TIP49)monomer per minute. The nucleotide bindingcompetition experiments shown in Figure 5 indicatethat both TIP48 and TIP49 bind ADP with a higheraffinity than ATP. It may be that the low turnovernumber for ATP hydrolysis is due to slow release ofADP after hydrolysis. These results are consistentwith the TIP49 crystal structure showing ADPtrapped in a closed nucleotide-binding pocket andthe protein exhibiting very low levels of ATPaseactivity in vitro.42 This suggests that interactionswith other proteins within larger complexes may berequired for ATP turnover. It might also be thatTIP48 and TIP49 play more of a structural role forthe assembly and activity of other proteins and,therefore, have slow turnover.In our model the sequences in TIP48 and TIP49
that are inserted between the Walker A and WalkerB motifs of the AAA+ domain would occupy densityregions that lie on the outside of the particle andwould be ideally situated to interact with otherproteins, consistent with the finding that part of theinserted domain is required for binding of c-Myc.10
The same region would be a prime candidate forspecific binding of other factors and the hexamericring architecture could facilitate interaction betweendifferent proteins.Our biochemical experiments did not show any
indications that TIP48 or TIP49 individually, or incomplex, can process DNA in vitro. It may be thatother proteins within a larger complex could recruitand load the proteins onto DNA. The channel thatruns through the TIP48/TIP49 complex is wideenough to accommodate ssDNA and possiblydsDNA. The activities of TIP48 and TIP49 couldalso be affected by post-translational modifications.Indeed, it was shown only recently that sumoylationof Reptin (TIP48) confers repressive function onTIP48 (reptin55). However, the sequence homologyto RuvB, which is clearly reflected in the hexamericring structure of the TIP48/TIP49 complex, does notnecessarily imply functional homology with RuvB.This structural organisation is typical formembers ofthe AAA+ family and TIP48 and TIP49 may equallywell act as molecular chaperones for the correctassembly of complex multi-protein complexes.

188 Dodecameric TIP48/TIP49 Complex
Recently, it was shown in yeast by chromatin im-munoprecipitation that Rvb1p (scTIP49) and Rvb2p(scTIP48) did not cross-link to the promoters theyregulate via the Ino80 complex, whereas the Ino80pdid.27 In the same study it was also suggested that inthe yeast Ino80 complex Rvb2p (scTIP48) and Rvb1p(scTIP49) do not have a catalytic role in chromatinremodelling but are required for the correct assem-bly of this complex.27 A different line of investigationhas shown that the association of TIP48 and TIP49with snoRNPs correlates with a restructuring event,which leads to the stabilization of the core box C/Dcomplex before it enters the nucleolus.32 It wasspeculated that TIP48 and TIP49 promote this restruc-turing event, acting as molecular motors. Clearly,further investigations are needed to identify thebiochemical substrates and reactions that are coupledto ATP hydrolysis by the TIP48/TIP49 complex.Our experiments reveal differences between the
closely related TIP48 and TIP49, which couldunderpin their functional specialisation. The antag-onistic effects that TIP48 and TIP49 exert on geneexpression regulated by β-catenin1,33,34 makes itunlikely that the two proteins work as a complex inthis pathway. If the active form of these proteins isthe hexameric ring, the ability of TIP48 to formstable oligomers, presumably hexamers, on its ownsuggests that it could function without TIP49. Onesuch example is presented in the finding that TIP48affects the transcriptional activities of ATF2, whereasTIP49 does not.17 We showed recently that in mitosisTIP48 relocalises to the midzone and, later incytokinesis, to the mid-body of dividing HeLacells, which suggested a possible novel role forTIP48 in mitosis.24 Sequence comparisons show thatthere is an evolutionary ancestor of TIP48 and TIP49in archaea.5,21 A duplication event could have led tothe evolution of the TIP48/TIP49 complex and thediversification of function, including functions thatinvolve complex formation alongside specialisedfunctions for TIP48 and TIP49 acting separately.
Materials and Methods
Expression constructs
The TIP48 and TIP49 coding sequences were PCRamplified using I.M.A.G.E library clones (IMAGE:2819778and IMAGE:2823568, respectively) as templates andcloned into T7 expression vectors. The TIP48 coningsequence was inserted into the NdeI and XhoI restrictionsites of the pET21b+ vector (Novagen) as a C-teminal sixhistidine (His6) fusion (pET21-TIP48-His6). The TIP49coding sequence was cloned into the pET21b+ vectorbetween the NdeI and BamHI restriction sites (pET21-TIP49) and into a pET15b+ (Novagen) vector between theNdeI and XhoI restriction sites creating an N-terminal His6fusion (pET15-His6-TIP49). A bi-cistronic construct for co-expression of TIP48 and TIP49-His6 was also engineeredbased on plasmid pET21a.Mutations were introduced into pET21-TIP48-His6,
pET21-TIP49 and pET15-His6-TIP49 constructs by sitedirected mutagenesis using the Quick-change® site-
directed mutagenesis kit (Stratagene). Aspartate residue299 of TIP48 was mutated to an asparagine (pET21-TIP48D299N-His6) and aspartate residue 302 of TIP49 wasmutated to an asparagine (pET21-TIP49D302N andpET15-His6-TIP49D302N).
Expression and purification of recombinantTIP48-His6, His6-TIP49 and their mutants
The pET21-TIP48-His6 and pET21-TIP48D299N-His6plasmids were transformed into E. coli BL21-Gold (DE3)pLysS cells (Novagen) in the presence of 100 μg/ml ofampicillin and 34 μg/ml of chloroamphenocol. ThepET15b-His6-TIP49 and pET15b-His6-TIP49D302N plas-mids were transformed into E. coli BL21-Gold (DE3) cells(Novagen) in the presence of 100 μg/ml of ampicillin and1% (w/v) glucose. The cells were grown at 37 °C in LBmedia containing the appropriate antibiotics to an A600 of0.6–0.8 and induced at 30 °C with 1 mM IPTG for 3–4 h.Induced cells were pelleted, frozen overnight at −20 °Cand thawed out in the presence of lysis buffer (20 mMTris–HCl (pH 8.0), 300 mM NaCl, 10% (v/v) glycerol,1 mM PMSF, 1 mM 2-β-mercaptoethanol), containingEDTA-free protease inhibitor tablets (Roche).For cells containing induced TIP48-His6 or TIP48D299N-
His6, Nonidet P40 was added to the cell suspension at afinal concentration of 0.2% (v/v) and cells were lysedfollowing incubation for 90min at 4 °C. For cells containinginducedHis6-TIP49 orHis6-TIP49D302N,Nonidet P40 andlysozyme (Sigma) were added to the cell suspension atfinal concentrations of 0.5% and 0.75 mg/ml, respectively,and cells were lysed following incubation for 60 min at4 °C. Following sonication, lysates were cleared bycentrifugation at 70,000 g for 30 min, in a Beckman 45 Tirotor. The cleared lysates were loaded onto a TALON™metal affinity column (Clontech), equilibrated in lysisbuffer. The column was washed with lysis buffer contain-ing 20 mM imidazole and the bound His6-tagged proteinswere eluted with a 20 mM–500 mM imidazole gradient.Peak fractions of TIP48-His6/TIP48D299N-His6 were
collected and dialysed overnight in buffer R (20 mM Tris–HCl (pH 8.0), 100 mM NaCl, 10% glycerol, 1 mM PMSF,1 mM DTT and 1 mM EDTA) and loaded onto a MonoQFPLC anion exchange column (Amersham Biosciences).The protein was eluted with a 0.1 M–1 MNaCl gradient inbuffer R. The purified protein was then dialysed intobuffer R and applied to ssDNA cellulose column (Sigma).The protein was collected in the flow-through fraction andwas concentrated using an Amicon® Ultra-15 Centrifugalfilter (Millipore). Lastly, the protein was fractionated on aSuperdex 200 HR 10/30 column (Amersham Biosciences).Peak fractions of His6-TIP49 or His6-TIP49D302N from
the TALON™ column were collected and dialysed over-night, into buffer H (10 mM potassium phosphate (pH6.8), 100 mM KCl, 1 mM DTT, 1 mM PMSF and 10%glycerol) and loaded onto a Hydroxyapatite column (Bio-Rad). Bound proteins were eluted with a 10 mM–600 mMpotassium phosphate gradient in buffer H. Peak fractionswere dialysed in buffer R and applied to ssDNA cellulosecolumn. The flow-through from the ssDNA cellulosecolumn was concentrated and fractionated further on aSuperdex 200 FPLC column.
Partial purification of non-tagged recombinant TIP49
The pET21-TIP49 and pET21-TIP49D302N plasmidswere transformed into the E. coli BL21-Gold (DE3) strain.Cell cultures were grown and induced with 1 mM IPTG as

189Dodecameric TIP48/TIP49 Complex
described above. Cells were lysed in buffer R containing0.3 M NaCl, protease inhibitor tablets (Roche), 0.2%Nonidet P40 and 0.75 mg/ml of lysozyme. After 60 min,the cells were soniciated and centrifuged at 66,000g for30 min in a Beckman 70 Ti rotor. To remove cellular DNA,the cleared lysate was loaded onto a DEAE Bio-Gelcolumn (Bio-Rad) pre-equilibrated in buffer R containing0.3 M NaCl. Proteins in the flow-through fraction werefractionated by 20% (w/v) ammonium sulphate precipita-tion followed by 70% ammonium sulphate precipitation.The 20%–70% ammonium sulphate pellet was resus-pended and dialyzed in buffer H and loaded onto a SPSepharose column (Amersham Biosciences), connected toa Hydroxyapatite column. Once the sample was loadedand the columns washed with buffer H, the SP Sepharosecolumnwas disconnected and the protein was eluted fromthe Hydroxyapatite column with a 10 mM–600 mMpotassium phosphate gradient in buffer H. Fractionscontaining TIP49 were pooled and precipitated with 70%ammonium sulphate.The ammonium sulphate pellet was resuspended and
dialyzed into lysis buffer. The amount of TIP49 in thesample was quantified by SDS–12% (w/v) PAGE, usingthe gel quantification program Gene tools (Syngene).
Assembly and isolation of TIP48/TIP49 complexes
For assembly of TIP48-His6/TIP49 complex partiallypurified TIP49 was incubated for 30 min on ice withTIP48-His6/TIP48D299N-His6 in a 2:1 ratio in lysis buffer.The mixture was loaded onto a TALON™ column andbound proteins were eluted with a 10 mM–500 mMimidazole gradient in lysis buffer. The presence of TIP49and TIP48-His6 in the eluted fractions was analyzed bySDS–18% PAGE and immunoblotting, using anti-TIP48and anti-TIP49 antibodies.24 The TIP48-His6/TIP49 com-plexes were fractionated further by size exclusion chro-matography on a Superdex 200 FPLC column in buffer Rcontaining 0.05% Triton X-100. The fractions wereconcentrated using Amicon® Ultra-15 Centrifugal filters.For the preparation of TIP48/TIP49-His6 complex TIP48
and TIP49-His6 were co-expressed from a bi-cistronicpET21a construct transformed into the E. coli BL21-Gold(DE3) strain. Cells were induced overnight with 0.5 mMIPTG at 20 °C. Cell lysis and metal affinity chromato-graphy was as described above.
Size exclusion chromatography
For each chromatography run 200–500 μl of purifiedHis6-TIP49 or TIP48-His6 (concentrated to 1 mg/ml orabove) were loaded on Superdex 200 FPLC columnequilibrated with buffer R alone, buffer R containing0.1 mM adenine nucleotide, or buffer R without EDTAcontaining 2 mM MgCl2, as appropriate. The proteinsample was supplemented with adenine nucleotides at1 mM final concentration and incubated on ice for 5 min,followed by incubation with MgCl2 at final concentrationof 2 mM, for a further 5 min. The amount of proteinpresent in different oligomeric forms was estimated fromthe area under the chromatograph peaks.
ATPase assays
Reactions contained ATPase reaction buffer (50 mMTris–HCl (pH 7.5), 50 mM NaCl, 2 mMMgCl2, 0.1 mg/mlBSA, 1 mM DTT, 1 mM ATP) and 0.0025 μCi/μl of [α-32P]
ATP (800 Ci/mmol; Perkin and Elmer). Protein was addedto each 10 μl or 20 μl single reaction tube to the finalconcentration, as specified. Where used, DNA substrateswere added to a final concentration of 5 ng/μl. Reactionswere incubated for the specific time at 37 °C and stoppedby adding 5 μl of 0.5 M EDTA (pH 8.0) per 20 μl. For timecourse experiments, master mixtures were pre-incubatedat 37 °C for 5 min. For the 0 min time point, the proteinwas added to the master mixture, mixed and a 10 μlaliquot was immediately removed and stopped withEDTA. Incubation continued at 37 °C with 10 μl aliquotswithdrawn at specific time points and stopped. DNAsubstrates used in assays were ϕX174 virion ssDNA(NEB), supercoiled pUC19 plasmid (NEB) and pUC19linearised by digestion with EcoRI. The synthetic 62-merfour-way Holliday junction X12 (unlabelled) was made asdescribed.56
ATP hydrolysis was determined by thin-layered chro-matography. Aliquots (1 μl) from each reaction werespotted onto PEI cellulose TLC plates (Sigma) anddeveloped with ATPase running buffer (1 M formic acidand 0.5M LiCl) for 2 h. Plates were dried and exposed on aphosphorimager screen (Fuji film) overnight. Spots werevisualized with a FUJI FLA-2000 phosphorimager andquantified using the FUJI FILM Image Gauge program. Amolecular mass of 50 kDa was used to calculate theamount of ATP hydrolyzed per mole of protein for TIP48,TIP49 and the TIP48/TIP49 complex.
ATP binding assays
One to 2 μg of protein was added to binding reactionscontaining ATP binding buffer (20 mM Tris–HCl (pH 7.5),100 mM NaCl, 1 mM DTT and 25 μg/ml BSA), 2 mMMgCl2 and 1 μCi of [γ-32P]2 azido-ATP (10–15 Ci/mmol;Affinity Labelling Technologies). The reactions wereincubated at room temperature for 3 min in the darkand then exposed for 30–60 s to UV light at the intensity of1800 μW/cm2. Protein was precipitated with 10% tri-chloroacetic acid (in 15 mM Tris–HCl (pH 7.5)), overnightfollowed by centrifugation for 30 min at 6000g. The pelletswere washed once with ice-cold acetone, dissolved in SDSprotein loading buffer (50 mM Tris–HCl (pH 6.8), 100 mMDTT, 4% SDS, 0.1% (w/v) bromophenol blue and 40%glycerol) and separated by SDS–PAGE. Gels were stainedwith Coomassie brilliant blue, dried and exposed over-night on a phosphorimager screen and visualised by FUJIFLA 2000 phosphorimager.For nucleotide competition binding experiments, a
master mixture was made up containing ATP bindingbuffer and protein. Amixture of 10 μM [γ-32P]2 azido-ATP,2 mM MgCl2 and the adenine nucleotide cofactor (finalconcentrations of 0.01, 0.1 or 0.5 mM), was added to eachsingle 20 μl reaction. Reactions were incubated at roomtemperature, exposed to UV light and processed as above.
Electron microscopy and 3D reconstruction
A 3 μl sample of the protein complex (63 μg/ml) inbuffer R without PMSF and EDTAwas applied to carboncoated copper grids (400 mesh) and stained twice with 2%(w/v) uranyl acetate. Micrographs were collected underlow dose conditions on a Tecnai T12 electron microscope(FEI, Eindhoven, The Netherlands) at 120 kV. Images weretaken at a magnification of 42,000x using defocus settingsfrom 0.4 μm −1 μm, and scanned on a SCAI micro-densitometer (Zeiss), resulting in a pixel size of 3.3 Å.

190 Dodecameric TIP48/TIP49 Complex
Image analysis was performed using the IMAGIC-557
and SPIDER58 software packages. A total of 2267 particleswere manually selected and extracted using the MRCprograms Ximdisp and Label.59 The images were band-pass filtered between 8 Å and 200 Å, translationallycentred and classified using multivariate statistical analy-sis. Individual images from classes representing top viewswere extracted and randomly rotated before beingsubjected to multivariate statistical analysis and symmetryanalysis in IMAGIC. A low-resolution 3D map wascreated by angular reconstitution with C6 point-groupsymmetry in IMAGIC. The map was refined by projectionmatching in SPIDER using a 3° angular step size. After fivecycles of refinement the orientations of the particlesstabilized, yielding a 3D structure at 20 Å resolutionestimated by Fourier shell correlation with a 0.5 correla-tion cut-off. The handedness of the EM structure waschosen to match that of the X-ray structure of p97/VCPND1 in the upper ring45 and the crystal structures wasfitted manually into each hexameric ring of the EMstructure using PyMol§.
Acknowledgements
We thank Elena Orlova for helping with thegeneration of the first 3D reconstruction in IMAGICand Andrew Niewiarowski for the TIP48/TIP49-His6 complex data. We are grateful for the supportobtained from the BBSRC (to T.P.) and the WellcomeTrust who funded parts of this work.
References
1. Bauer, A., Chauvet, S., Huber, O., Usseglio, F.,Rothbacher, U., Aragnol, D. et al. (2000). Pontin52 andReptin52 function as antagonistic regulators of beta-catenin signalling activity. EMBO J. 19, 6121–6130.
2. Bauer, A., Huber, O. & Kemler, R. (1998). Pontin52, aninteraction partner of beta-catenin, binds to the TATAbox binding protein. Proc. Natl Acad. Sci. USA, 95,14787–14792.
3. Gohshi, T., Shimada, M., Kawahire, S., Imai, N.,Ichimura, T., Omata, S. & Horigome, T. (1999).Molecular cloning of mouse p47, a second groupmammalian RuvB DNA helicase-like protein: homol-ogy with those from human and Saccharomycescerevisiae. J. Biochem. (Tokyo), 125, 939–946.
4. Ikura, T., Ogryzko, V. V., Grigoriev, M., Groisman, R.,Wang, J., Horikoshi, M. et al. (2000). Involvement ofthe TIP60 histone acetylase complex in DNA repairand apoptosis. Cell, 102, 463–473.
5. Kanemaki, M., Kurokawa, Y., Matsu-ura, T., Makino,Y., Masani, A., Okazaki, K. et al. (1999). TIP49b, a newRuvB-like DNA helicase, is included in a complextogether with another RuvB-like DNA helicase,TIP49a. J. Biol. Chem. 274, 22437–22444.
6. Kanemaki, M., Makino, Y., Yoshida, T., Kishimoto, T.,Koga, A., Yamamoto, K. et al. (1997). Molecularcloning of a rat 49-kDa TBP-interacting protein(TIP49) that is highly homologous to the bacterialRuvB. Biochem. Biophys. Res. Commun. 235, 64–68.
§www.pymol.org
7. Kikuchi, N., Gohshi, T., Kawahire, S., Tachibana, T.,Yoneda, Y., Isobe, T. et al. (1999). Molecular shape andATP binding activity of rat p50, a putative mammalianhomologue of RuvB DNA helicase. J .Biochem. (Tokyo),125, 487–494.
8. Qiu, X. B., Lin, Y. L., Thome, K. C., Pian, P., Schlegel,B. P.,Weremowicz, S. et al. (1998). An eukaryotic RuvB-like protein (RUVBL1) essential for growth. J. Biol.Chem. 273, 27786–27793.
9. Shen, X., Mizuguchi, G., Hamiche, A. & Wu, C.(2000). A chromatin remodelling complex involvedin transcription and DNA processing. Nature, 406,541–544.
10. Wood, M. A., McMahon, S. B. & Cole, M. D. (2000). AnATPase/helicase complex is an essential cofactor foroncogenic transformation by c-Myc. Mol. Cell, 5,321–330.
11. Neuwald, A. F., Aravind, L., Spouge, J. L. & Koonin,E. V. (1999). AAA+: a class of chaperone-like ATPasesassociated with the assembly, operation, and disas-sembly of protein complexes. Genome Res. 9, 27–43.
12. Tsaneva, I. R., Muller, B. & West, S. C. (1992). ATP-dependent branch migration of Holliday junctionspromoted by the RuvA and RuvB proteins of E. coli.Cell, 69, 1171–1180.
13. Fuchs, M., Gerber, J., Drapkin, R., Sif, S., Ikura,T., Ogryzko, V. et al. (2001). The p400 complex isan essential E1A transformation target. Cell, 106,297–307.
14. Jonsson, Z. O., Dhar, S. K., Narlikar, G. J., Auty, R.,Wagle, N., Pellman, D. et al. (2001). Rvb1p and Rvb2pare essential components of a chromatin remodelingcomplex that regulates transcription of over 5% ofyeast genes. J. Biol. Chem. 276, 16279–16288.
15. Kobor, M. S., Venkatasubrahmanyam, S., Meneghini,M. D., Gin, J. W., Jennings, J. L., Link, A. J. et al. (2004).A protein complex containing the conserved Swi2/Snf2-related ATPase Swr1p deposits histone variantH2A.Z into euchromatin. PloS. Biol. 2, E131.
16. Krogan, N. J., Keogh, M. C., Datta, N., Sawa, C., Ryan,O. W., Ding, H. et al. (2003). A Snf2 family ATPasecomplex required for recruitment of the histone H2Avariant Htz1. Mol. Cell, 12, 1565–1576.
17. Cho, S. G., Bhoumik, A., Broday, L., Ivanov, V.,Rosenstein, B. & Ronai, Z. (2001). TIP49b, a regulatorof activating transcription factor 2 response to stressand DNA damage. Mol. Cell Biol. 21, 8398–8413.
18. Dugan, K. A., Wood, M. A. & Cole, M. D. (2002).TIP49, but not TRRAP, modulates c-Myc and E2F1dependent apoptosis. Oncogene, 21, 5835–5843.
19. Lim, C. R., Kimata, Y., Ohdate, H., Kokubo, T.,Kikuchi, N., Horigome, T. & Kohno, K. (2000). TheSaccharomyces cerevisiae RuvB-like protein, Tih2p, isrequired for cell cycle progression and RNA poly-merase II-directed transcription. J. Biol. Chem. 275,22409–22417.
20. King, T. H., Decatur,W. A., Bertrand, E., Maxwell, E. S.& Fournier, M. J. (2001). A well-connected andconserved nucleoplasmic helicase is required forproduction of box C/D and H/ACA snoRNAs andlocalization of snoRNP proteins. Mol. Cell Biol. 21,7731–7746.
21. Newman, D. R., Kuhn, J. F., Shanab, G. M. &Maxwell,E. S. (2000). Box C/D snoRNA-associated proteins:two pairs of evolutionarily ancient proteins andpossible links to replication and transcription. RNA,6, 861–879.
22. Watkins, N. J., Dickmanns, A. & Luhrmann, R. (2002).Conserved stem II of the box C/Dmotif is essential for

191Dodecameric TIP48/TIP49 Complex
nucleolar localization and is required, along with the15.5 K protein, for the hierarchical assembly of the boxC/D snoRNP. Mol. Cell Biol. 22, 8342–8352.
23. Gartner, W., Rossbacher, J., Zierhut, B., Daneva, T.,Base, W., Weissel, M. et al. (2003). The ATP-dependenthelicase RUVBL1/TIP49a associates with tubulinduring mitosis. Cell Motil. Cytoskeleton. 56, 79–93.
24. Sigala, B., Edwards, M., Puri, T. & Tsaneva, I. R. (2005).Relocalization of human chromatin remodeling cofac-tor TIP48 in mitosis. Exp. Cell Res. 310, 357–369.
25. Makino, Y., Kanemaki, M., Kurokawa, Y., Koji, T. &Tamura, T. (1999). A rat RuvB-like protein, TIP49a, is agerm cell-enriched novel DNA helicase. J .Biol. Chem.274, 15329–15335.
26. Feng, Y., Lee, N. & Fearon, E. R. (2003). TIP49regulates beta-catenin-mediated neoplastic transfor-mation and T-cell factor target gene induction viaeffects on chromatin remodeling. Cancer Res. 63,8726–8734.
27. Jonsson, Z. O., Jha, S., Wohlschlegel, J. A. & Dutta, A.(2004). Rvb1p/Rvb2p recruit Arp5p and assemble afunctional Ino80 chromatin remodeling complex. Mol.Cell, 16, 465–477.
28. Bakshi, R., Mehta, A. K., Sharma, R., Maiti, S., Pasha,S. & Brahmachari, V. (2006). Characterization of ahuman SWI2/SNF2 like protein hINO80: Demonstra-tion of catalytic and DNA binding activity. Biochem.Biophys. Res. Commun. 339, 313–320.
29. Jin, J., Cai, Y., Yao, T., Gottschalk, A. J., Florens, L.,Swanson, S. K. et al. (2005). A mammalian chromatinremodeling complex with similarities to the yeastINO80 complex. J. Biol. Chem. 280, 41207–41212.
30. Mizuguchi, G., Shen, X., Landry, J., Wu, W. H., Sen, S.& Wu, C. (2004). ATP-driven exchange of histoneH2AZ variant catalyzed by SWR1 chromatin remo-deling complex. Science, 303, 343–348.
31. Doyon, Y., Selleck, W., Lane, W. S., Tan, S. & Cote, J.(2004). Structural and functional conservation of theNuA4 histone acetyltransferase complex from yeast tohumans. Mol. Cell Biol. 24, 1884–1896.
32. Watkins, N. J., Lemm, I., Ingelfinger, D., Schneider, C.,Hossbach, M., Urlaub, H. & Luhrmann, R. (2004).Assembly and maturation of the U3 snoRNP in thenucleoplasm in a large dynamic multiprotein com-plex. Mol. Cell, 16, 789–798.
33. Rottbauer, W., Saurin, A. J., Lickert, H., Shen, X.,Burns, C. G., Wo, Z. G. et al. (2002). Reptin and pontinantagonistically regulate heart growth in zebrafishembryos. Cell, 111, 661–672.
34. Kim, J. H., Kim, B., Cai, L., Choi, H. J., Ohgi, K. A.,Tran, C. et al. (2005). Transcriptional regulation of ametastasis suppressor gene by Tip60 and beta-catenincomplexes. Nature, 434, 921–926.
35. Puri, T. (2005). Characterisation of the human TIP48and TIP49 AAA+ proteins and their complex, PhDthesis, University of London.
36. Davey, M. J., Indiani, C. & O'Donnell, M. (2003).Reconstitution of the Mcm2-7p heterohexamer, sub-unit arrangement, and ATP site architecture. J. Biol.Chem. 278, 4491–4499.
37. Mitchell, A. H. & West, S. C. (1994). Hexameric ringsof Escherichia coli RuvB protein. Cooperative assembly,processivity and ATPase activity. J. Mol. Biol. 243,208–215.
38. Ogura, T. & Wilkinson, A. J. (2001). AAA+ super-family ATPases: common structure–diverse function.Genes Cells, 6, 575–597.
39. Schwacha, A. & Bell, S. P. (2001). Interactions betweentwo catalytically distinct MCM subgroups are essen-
tial for coordinated ATP hydrolysis and DNA repli-cation. Mol. Cell, 8, 1093–1104.
40. Mezard, C., Davies, A. A., Stasiak, A. & West, S. C.(1997). Biochemical properties of RuvBD113N: amutation in helicase motif II of the RuvB hexameraffects DNA binding and ATPase activities. J. Mol.Biol. 271, 704–717.
41. Yu, C. M., Mun, S. & Wang, N. H. (2006). Theoreticalanalysis of the effects of reversible dimerization in sizeexclusion chromatography. J. Chromatog. sect. A, 1132,99–108.
42. Dube, P., Tavares, P., Lurz, R. & van Heel, M. (1993).The portal protein of bacteriophage SPP1: a DNApump with 13-fold symmetry. EMBO J. 12, 1303–1309.
43. Matias, P. M., Gorynia, S., Donner, P. & Carrondo,M. A. (2006). Crystal structure of the human AAA+
protein RUVBL1. J. Biol. Chem. In the press.44. Putnam, C. D., Clancy, S. B., Tsuruta, H., Gonzalez, S.,
Wetmur, J. G. & Tainer, J. A. (2001). Structure andmechanism of the RuvB Holliday junction branchmigration motor. J. Mol. Biol. 311, 297–310.
45. Dreveny, I., Kondo, H., Uchiyama, K., Shaw, A.,Zhang, X. & Freemont, P. S. (2004). Structural basis ofthe interaction between the AAA ATPase p97/VCPand its adaptor protein p47. EMBO J. 23, 1030–1039.
46. Stasiak, A., Tsaneva, I. R., West, S. C., Benson, C. J., Yu,X. & Egelman, E. H. (1994). The Escherichia coli RuvBbranch migration protein forms double hexamericrings around DNA. Proc. Natl Acad. Sci. USA, 91,7618–7622.
47. Pape, T., Meka, H., Chen, S., Vicentini, G., van Heel,M. & Onesti, S. (2003). Hexameric ring structure of thefull-length archaeal MCM protein complex. EMBORep. 4, 1079–1083.
48. King, S. M. (2000). AAA domains and organization ofthe dynein motor unit. J. Cell Sci. 113, 2521–2526.
49. Chong, J. P., Hayashi, M. K., Simon, M. N., Xu, R. M. &Stillman, B. (2000). A double-hexamer archaeal mini-chromosome maintenance protein is an ATP-depen-dent DNA helicase. Proc. Natl Acad. Sci. USA, 97,1530–1535.
50. Valle, M., Gruss, C., Halmer, L., Carazo, J. M. &Donate, L. E. (2000). Large T-antigen double hexamersimaged at the simian virus 40 origin of replication.Mol. Cell Biol. 20, 34–41.
51. Beyer, A. (1997). Sequence analysis of the AAAprotein family. Protein Sci. 6, 2043–2058.
52. Schirmer, E. C., Glover, J. R., Singer,M. A. & Lindquist,S. (1996). HSP100/Clp proteins: a common mechan-ism explains diverse functions. Trends Biochem. Sci. 21,289–296.
53. Yao, N., Coryell, L., Zhang, D., Georgescu, R. E.,Finkelstein, J., Coman, M. M. et al. (2003). Replicationfactor C clamp loader subunit arrangement within thecircular pentamer and its attachment points toproliferating cell nuclear antigen. J. Biol. Chem. 278,50744–50753.
54. Hishida, T., Han, Y. W., Fujimoto, S., Iwasaki, H. &Shinagawa, H. (2004). Direct evidence that a con-served arginine in RuvB AAA+ ATPase acts as anallosteric effector for the ATPase activity of theadjacent subunit in a hexamer. Proc. Natl Acad. Sci.USA, 101, 9573–9577.
55. Kim, J. H., Choi, H. J., Kim, B., Kim, M. H., Lee, J. M.,Kim, I. S. et al. (2006). Roles of sumoylation of a reptinchromatin-remodelling complex in cancer metastasis.Nature Cell Biol. 8, 631–639.
56. Oram, M., Keeley, A. & Tsaneva, I. (1998). Hollidayjunction resolvase in Schizosaccharomyces pombe has

192 Dodecameric TIP48/TIP49 Complex
identical endonuclease activity to the CCE1 homo-logue YDC2. Nucl. Acids Res. 26, 594–601.
57. van Heel, M., Harauz, G., Orlova, E. V., Schmidt, R.& Schatz, M. (1996). A new generation of theIMAGIC image processing system. J. Struct. Biol. 116,17–24.
58. Frank, J., Radermacher, M., Penczek, P., Zhu, J., Li,
Y., Ladjadj, M. & Leith, A. (1996). SPIDER and WEB:processing and visualization of images in 3D electronmicroscopy and related fields. J. Struct. Biol. 116,190–199.
59. Crowther, R. A., Henderson, R. & Smith, J. M. (1996).MRC image processing programs. J. Struct. Biol. 116,9–16.
Edited by K. Morikawa
(Received 31 July 2006; received in revised form 4 November 2006; accepted 8 November 2006)Available online 11 November 2006