diversitÉ gÉnÉtique de talpa europaea et de …
TRANSCRIPT

Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/ 277
COMMUNICATION
DIVERSITÉ GÉNÉTIQUE DE TALPA EUROPAEA
ET DE L’HANTAVIRUS NOVA (NVAV) EN FRANCE
GENETIC DIVERSITY OF TALPA EUROPAEA
AND NOVA HANTAVIRUS (NVAV) IN FRANCE
Par Jean-Pierre HUGOT(1), Se Hun GU(2), Carlos FELIU(3), Jacint VENTURA(4), Alexis RIBAS(5),Jérôme DORMION(6), Richard YANAGIHARA(2) and Violaine NICOLAS(1)
Communication présentée le 22 mai 2014
L’Hantavirus Nova (NVAV) a été identifié chez un spécimen de Talpa europaea capturé en Hongrie.L’analyse de 94 spécimens de taupes capturés en France a révélé la présence de NVAV chez 50% desindividus. Une étude populationnelle des taupes montre que les individus collectés entre Poitiers et Bordeaux sont génétiquement proches de l’espèce voisine T. occidentalis, jusqu’ici supposée êtrestrictement endémique dans la péninsule ibérique. Plusieurs hypothèses permettant d’expliquer cesobservations sont discutées : 1) la présence jusqu’ici ignorée de T. occidentalis dans le sud-ouest de la France ; 2) l’existence d’un ancien phénomène d’introgression mitochondriale entre les deuxespèces, ; 3) la présence d’une zone d’hybridation entre les deux espèces, produisant un phénotypeparticulier chez certains hybrides ; 4) l’existence d’une espèce nouvelle. NVAV n’ayant été détecté chezaucun des spécimens du Sud-Ouest, la question de l’existence d’un Hantavirus particulier dans cettepopulation et chez la taupe ibérique est posée.
Mots-Clés: Talpa europaea, Talpa occidentalis, Hantavirus, NVAV, phylogénétique, phylogéographie.
RÉSUMÉ
(1) UMR CNRS 7205, MNHN, 51, rue Buffon, 75231 Paris cedex, France.(2) Pacific Center for Emerging Infectious Diseases Research, John A. Burns School of Medicine, UNIVERSITY OF HAWAII AT MANOA, Honolulu, Hawaii, USA.(3) UNIV-BARCELONA, Facultad de Farmacia-Parasitología, Avda. Diagonal s/n 8028 – Barcelona, Espagne.(4) UNIVERSITAT AUTÒNOMA DE BARCELONA, Unitat de Zoologia, Departament de Biologia Animal, Biologia Vegetal i Ecologia, Facultat de Biociències, 08193 Cerdanyoladel Vallès, Espagne.
(5) Biodiversity Research Group, Faculty of Science, UDON THANI RAJABHAT UNIVERSITY, Udon thani 41000 Thailand.(6) TAUP’GREEN France, BP 19 92201, Neuilly sur Seine, France.
Nova hantavirus (NVAV) was first identified in a captured European mole (Talpa europaea) in Hungary.Analysis of lung tissues from 94 moles captured in France revealed NVAV in 50% of the animals. Basedon the genetic diversity of the cytochrome b mtDNA, moles collected in Poitiers and Bordeaux weremore closely related to the Iberian mole (T. occidentalis), a species previously assumed to berestricted to the Iberian Peninsula. Several hypotheses are discussed to explain these observations:1) the presence of hitherto unnoticed T. occidentalis in southwestern France; 2) the existence of anancient mitochondrial introgression phenomenon between the two Talpa species, producing a par-ticular phenotype in some hybrids; 3) the existence of a hybrid zone between the two species; and4) the existence of a new Talpa species. NVAV was not detected in the southwestern moles, whichraises the question of the possible presence of a particular Hantavirus species in this population and/orin the Iberian moles.
Key-Words: Talpa europaea, Talpa occidentalis, Hantavirus, NVAV, phylogenetics, phylogeography.
SUMMARY

COMMUNICATION
278 Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/
INTRODUCTION Hantaviruses (family Bunyaviridae), long known to be harbo-red by rodents (rats, mice, voles, etc.) of the Muridae andCricetidae families, have been detected in multiple species ofshrews (Soricidae) and moles (Talpidae) (Yanagihara et al.2014). More recently, highly divergent lineages of hantaviruseshave also been discovered in insectivorous bats (Gu et al. 2014b;Guo et al. 2013; Yanagihara et al. 2014; table 1). In-depth stu-dies of non-rodent hosts, which are heavily represented amongmicro-mammals all around the world, may provide a betterunderstanding about their evolutionary origins. Since some han-taviruses may infect and cause diseases in humans, such studiesmay also interest epidemiologists (Pontier & Fouchet, 2013;Reynes, 2013; Tordo et al. 2013).
Between October 2012 and March 2013, we sampled Europeanmoles, Talpa europaea, in various regions of France. The pur-pose of this investigation was:
1) to verify the presence of Nova hantavirus (NVAV), origi-nally described in a single specimen of T. europaea fromHungary (Kang et al. 2009);
2) to assess the prevalence and geneticvariability of NVAV; and
3) to compare NVAV genetic diversitywith mole population genetics.
The study revealed the presence ofNVAV in France and the existence of agenetic variability of the virus linked tothe localities where the moles werecaptured (Gu et al. 2014a). Analysis ofthe cytochrome b mtDNA producedan unexpected finding: that is, althoughonly one mole species (T. europaea)was considered to be present in France,some moles collected in the southwes-tern part of the country were geneticallymore closely related to T. occidentalis, adifferent species previously believed tobe endemic to the Iberian Peninsula.Details of these results are presented anddiscussed.
MATERIAL AND METHODS
Collecting moles
Approximately 350 moles were captu-red in different localities in France(figure 1) from October 2012 to March2013, using Putange traps and follo-wing a protocol approved by Frenchprofessional mole catchers (Dormion,2012).
Research of hantavirus
A subset of 94 moles captured either in the Golf of Ozoir-la-Ferrière(48.7700°N, 2.6800°E), or in the Beauvais agreement park called«Le Plan d'eau du Canada » (49°452698 N, 2°058234 E), were stu-died. We also analyzed 24 mole specimens collected betweenBordeaux and Poitiers and genetically different from T. europaea.After capture, moles were frozen at -20°C; for tissue dissection,moles were partially thawed and lung tissues were removed usingethanol-cleaned instruments and placed in RNAlater® RNAStabilization Reagent (Qiagen Inc., Valencia, CA).
Hantavirus detection and sequencing
Total RNA was extracted from mole lung tissues, using thePureLink Micro-to-Midi total RNA purification kit (Invitrogen,San Diego, CA), and cDNA was prepared using the SuperScriptIII First-Strand Synthesis System (Invitrogen) and randomhexamers. PCR was performed as described previously using oli-gonucleotide primers designed from NVAV and other sorico-morph-borne hantaviruses (Song et al. 2007; Arai et al. 2008)and DNA sequencing was performed using an ABI Prism377XL Genetic Analyzer (Applied Biosystems, Foster City, CA).
Figure 1. Map of collection localities.Map of France, showing localities where European moles were captured during October 2012 to March 2013.The results presented in this manuscript were obtained from specimens collected in the localities indicated inred (Blanquefort, St-Loubès, Sadirac, Ambarès, Arcins and Izon are small cities situated on either side of theGironde estuary, in the vicinity of Bordeaux).
Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/ 279
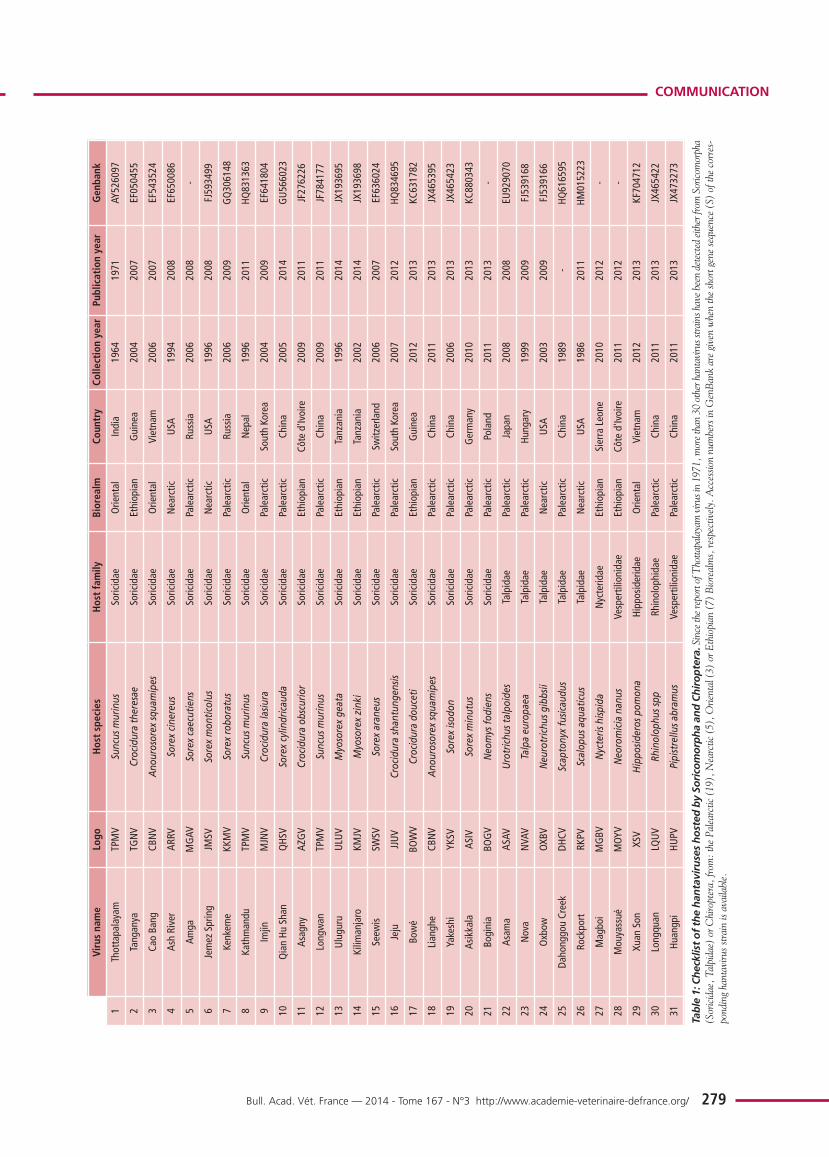
COMMUNICATIONVirus name
Logo
Host species
Host family
Biorealm
Country
Collection year
Publication year
Genbank
1Th
ottapa
layam
TPMV
Suncus murinus
Soric
idae
Orie
ntal
India
1964
1971
AY52
6097
2Tang
anya
TGNV
Crocidura theresae
Soric
idae
Ethiop
ian
Guine
a20
0420
07EF
0504
55
3Ca
o Ba
ngCB
NV
Anourosorex squamipes
Soric
idae
Orie
ntal
Vietna
m20
0620
07EF
5435
24
4As
h River
ARRV
Sorex cinereus
Soric
idae
Nea
rctic
USA
1994
2008
EF65
0086
5Am
gaMGAV
Sorex caecutiens
Soric
idae
Palearctic
Russia
2006
2008
-
6Jemez Spring
JMSV
Sorex monticolus
Soric
idae
Nea
rctic
USA
1996
2008
FJ59
3499
7Ke
nkem
eKK
MV
Sorex roboratus
Soric
idae
Palearctic
Russia
2006
2009
GQ30
6148
8Ka
thman
duTP
MV
Suncus murinus
Soric
idae
Orie
ntal
Nep
al19
9620
11HQ
8313
63
9Im
jinMJN
VCrocidura lasiura
Soric
idae
Palearctic
South Ko
rea
2004
2009
EF64
1804
10Qian Hu
Sha
nQHS
VSorex cylindricauda
Soric
idae
Palearctic
China
2005
2014
GU56
6023
11As
agny
AZGV
Crocidura obscurior
Soric
idae
Ethiop
ian
Côte d'Iv
oire
2009
2011
JF27
6226
12Lo
ngwan
TPMV
Suncus murinus
Soric
idae
Palearctic
China
2009
2011
JF78
4177
13Ulugu
ruULU
VMyosorex geata
Soric
idae
Ethiop
ian
Tanzan
ia19
9620
14JX19
3695
14Kilim
anjaro
KMJV
Myosorex zinki
Soric
idae
Ethiop
ian
Tanzan
ia20
0220
14JX19
3698
15Seew
isSW
SVSorex araneus
Soric
idae
Palearctic
Switz
erland
2006
2007
EF63
6024
16Jeju
JJUV
Crocidura shantungensis
Soric
idae
Palearctic
South Ko
rea
2007
2012
HQ83
4695
17Bo
wé
BOWV
Crocidura douceti
Soric
idae
Ethiop
ian
Guine
a20
1220
13KC
6317
82
18Lian
ghe
CBNV
Anourosorex squamipes
Soric
idae
Palearctic
China
2011
2013
JX46
5395
19Ya
keshi
YKSV
Sorex isodon
Soric
idae
Palearctic
China
2006
2013
JX46
5423
20As
ikka
laAS
IVSorex minutus
Soric
idae
Palearctic
German
y20
1020
13KC
8803
43
21Bo
ginia
BOGV
Neomys fodiens
Soric
idae
Palearctic
Poland
2011
2013
-
22As
ama
ASAV
Urotrichus talpoides
Talpidae
Palearctic
Japa
n20
0820
08EU
9290
70
23Nov
aNVA
VTalpa europaea
Talpidae
Palearctic
Hung
ary
1999
2009
FJ53
9168
24Oxb
owOX
BVNeurotrichus gibbsii
Talpidae
Nea
rctic
USA
2003
2009
FJ53
9166
25Da
hong
gou Creek
DHCV
Scaptonyx fusicaudus
Talpidae
Palearctic
China
1989
-HQ
6165
95
26Ro
ckpo
rtRK
PVScalopus aquaticus
Talpidae
Nea
rctic
USA
1986
20
11HM
0152
23
27Mag
boi
MGBV
Nycteris hispida
Nycterid
aeEthiop
ian
Sierra Leo
ne20
1020
12-
28Mou
yassué
MOY
VNeoromicia nanus
Vespertilionida
eEthiop
ian
Côte d'Iv
oire
2011
2012
-
29Xu
an Son
XSV
Hipposideros pom
ona
Hipp
osideridae
Orie
ntal
Vietna
m20
1220
13KF
7047
12
30Lo
ngqu
anLQ
UV
Rhinolophus spp
Rhinolop
hida
ePa
learctic
China
2011
2013
JX46
5422
31Hu
angp
iHU
PVPipistrellus abram
usVe
spertilionida
ePa
learctic
China
2011
2013
JX47
3273
Table 1: Checklist of the hantaviruses hosted by Soricomorpha and Chiroptera.Since the report of T
hottapalayam virus in 1971, m
ore than 30 other hantavirus strains have been detected either from
Soricom
orpha
(Soricidae, T
alpidae) or C
hiroptera, from
: the Palearctic (19), Nearctic (5), O
riental (3) or E
thiopian (7) Biorealms, respectively. Accession numbers in GenBank are given when the short gene sequence (S) of the corres-
ponding hantavirus strain is available.

COMMUNICATION
280 Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/
Mitochondrial DNA sequencing
DNA was extracted from ethanol-preserved tissues by theCTAB method (Winnepenninckx et al., 1993). The cytochromeb gene was amplified from 40 mole specimens using polymerasechain reaction (PCR) primers L14723 and H15915 (Ducroz etal. 2001; Nicolas et al. 2008).
Sequence alignment
Alignment of the coding region of the hantavirus S segment,or the cytochrome b of the moles was performed using CLUS-TAL-X automatic procedure (Thompson et al. 1994) thenimproved manually using SEAL v2.0a11 (Rambaut, 1996) andvalidated using the amino acid translation.
Phylogenetic analyses
Evolutionary relationships among sequences were estimated byconstructing phylogenetic trees using maximum parsimony(MP) and Bayesian Markov chain Monte Carlo phylogenetic ana-lyses (MCMC). MP analysis was performed with PAUP 4b10(Swofford, 2001) and Bayesian analysis with MrBayes 3.1.2(Huelsenbeck and Ronquist, 2001). The MP analysis was per-formed with tree-bisection-reconnection (TBR) branch swap-ping option and 10 random addition replicates. We estimated therobustness of internal nodes by 1,000 bootstrapping replicates(each with a single replication of random addition of taxa). Anequal weighting of character-state transformations was applied.In all MCMC analyses three heated chains and one single coldchain were employed, and runs were initiated with random trees.Two independent MCMC runs were conducted with six milliongenerations per run; trees (and parameters) sampled every 100generations. Stationarity was assessed by: examining the averagestandard deviation of split frequencies and, the Potential ScaleReduction Factor (Ronquist et al. 2005). For each run, the first25% of sampled trees were discarded as burn-in. A consensus treewas computed using the “halfcompat” option, equivalent to the50% majority rule and rooted using the “midpoint root” option.Proportion values of posterior probability of bipartition were usedfor evaluation of robustness of the nodes.
RESULTS
Prevalence of NVAV in moles
Hantavirus RNA was detected in 47 moles (50%) by RT-PCR.Of 36 and 28 moles captured in Ozoir-la-Ferrière on October18, 2012 and February 21, 2013, 15 and 14, respectively, werepositive, while 18 of 30 moles captured in Beauvais on March3, 2013 were positive (Gu et al. 2014a). NVAV was not detec-ted in the 24 southwestern moles.
Phylogenetic analysis of hantavirus found inmicro-mammals
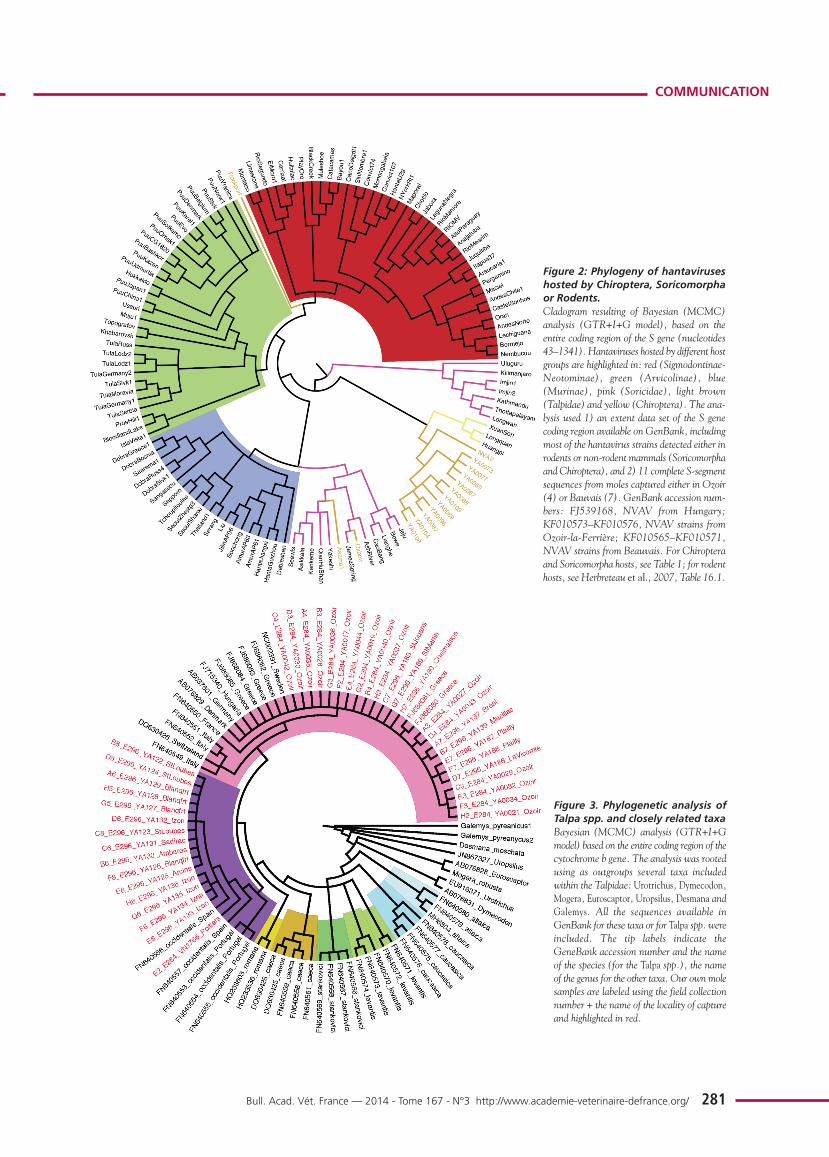
Phylogenetic analysis, using Bayesian methods (figure 2)showed that:
1) NVAV from France and prototype NVAV from Hungarygrouped together in a particular lineage;
2) NVAV strains from France segregated along geographic-spe-cific lineages;
3) NVAV clade was strongly associated with another clade,comprising hantaviruses detected in insectivorous bats;
4) other hantaviruses hosted by soricomorphs were distributedinto two separate clades: i) the most divergent included seven strains from shrews
captured in India, South Korea, China and Tanzania; ii) the second clade included strains from moles and shrews
captured in North America, China, Japan and Europe,and associated with hantaviruses hosted by Murinaerodents;
5) two other clades grouping hantaviruses hosted bySigmodontinae-Neotominae and Arvicolinae rodents, res-pectively;
6) a single mole-borne hantavirus, Rockport virus from the eas-tern mole in North America, was situated between theSigmodontinae and Arvicolinae clades. All the basal nodesof the cladogram had the maximum posterior probability ofbipartition value (= 1).
Genetic affinities within the Family Talpidae
Phylogenetic analysis, using Bayesian methods, of all availablesequences from Talpidae species (figure 3) showed that:
1) the genus Talpa may be considered a monophyletic group,in which each different species is recognized as a distinctclade;
2) T. europaea and T. occidentalis are sister taxa;
3) specimens captured in Brittany, Northern France and Saône-et-Loire clustered with strains identified as T. europaea andcollected from different European countries (Sweden,Denmark, Switzerland, Hungary, Germany, Italy andGreece);
4) specimens captured in the vicinity of Bordeaux, or Poitiers,clustered with specimens captured either from Spain orPortugal and identified as T. occidentalis, the Iberian mole.All the nodes of the cladogram corresponding to a particu-lar mole species had the maximum posterior probability ofbipartition value (= 1).
DISCUSSION
Mole infection
Prevalence of anti-viral antibodies and/or viral antigens inrodent species known to harbor disease-causing hantavirusesgenerally varies from less than 1% to more than 25%, depen-ding on seasonal factors, reservoir population density, geogra-phical area and ecological diversity (Lee et al.1978; Dobly et
Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/ 281
COMMUNICATION
Figure 2: Phylogeny of hantaviruseshosted by Chiroptera, Soricomorphaor Rodents.Cladogram resulting of Bayesian (MCMC)analysis (GTR+I+G model), based on theentire coding region of the S gene (nucleotides43–1341). Hantaviruses hosted by different hostgroups are highlighted in: red (Sigmodontinae-Neotominae), green (Arvicolinae), blue(Murinae), pink (Soricidae), light brown(Talpidae) and yellow (Chiroptera). The ana-lysis used 1) an extent data set of the S genecoding region available on GenBank, includingmost of the hantavirus strains detected either inrodents or non-rodent mammals (Soricomorphaand Chiroptera), and 2) 11 complete S-segmentsequences from moles captured either in Ozoir(4) or Bauvais (7). GenBank accession num-bers: FJ539168, NVAV from Hungary;KF010573–KF010576, NVAV strains fromOzoir-la-Ferrière; KF010565–KF010571,NVAV strains from Beauvais. For Chiropteraand Soricomorpha hosts, see Table 1; for rodenthosts, see Herbreteau et al., 2007, Table 16.1.
Figure 3. Phylogenetic analysis ofTalpa spp. and closely related taxaBayesian (MCMC) analysis (GTR+I+Gmodel) based on the entire coding region of thecytochrome b gene. The analysis was rootedusing as outgroups several taxa includedwithin the Talpidae: Urotrichus, Dymecodon,Mogera, Euroscaptor, Uropsilus, Desmana andGalemys. All the sequences available inGenBank for these taxa or for Talpa spp. wereincluded. The tip labels indicate theGeneBank accession number and the nameof the species (for the Talpa spp.), the nameof the genus for the other taxa. Our own molesamples are labeled using the field collectionnumber + the name of the locality of captureand highlighted in red.
COMMUNICATION
282 Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/
al. 2012). Recently, a similarly high prevalence of NVAVinfection, as evidenced by RT-PCR and confirmed by DNAsequencing, has been found in European moles captured in cen-tral Poland, suggesting that NVAV is widespread throughoutthe vast distribution of T. europaea (Gu et al. 2014a). The highprevalence of this hantavirus suggests that the mechanisms oftransmission between individuals are very efficient.
NVAV was not detected in the southwestern moles, but few spe-cimens (24) were available and the RNA quality was subopti-mal. Thus, future studies are warranted to ascertain if NVAVor NVAV-related hantaviruses are harbored by moles in sou-thwestern France. These investigations will be extended to molespecimens collected from different parts of the Iberian Peninsula.
Until now no human infection due to NVAV has been recor-ded. However, moles have a high propensity to occupy arablefields, deciduous woodland and permanent pastures, and han-tavirus are known to be able to survive for prolonged periods(12-15 days) in external environment (Kallio et al. 2006;Hardestam et al. 2007). Any unusual clinical syndromes, recor-ded among individuals reporting contact with European moles,should be thoroughly studied by physicians and public healthworkers.
Host and hantavirus coevolution
Hantaviruses were traditionally considered to have codiverged(co-speciatiated) with their rodent hosts (Herbreteau et al. 2006,2007; Hentonnen et al. 2008; Guo et al. 2013; Yanagihara et al.2014) and some evidence for such codivergence is apparenthere. In particular, rodent-borne hantaviruses clustered accor-ding to whether their hosts were members of the Murinae,
Arvicolinae or Sigmodontinae-Neotominae subfamilies.However, codivergence alone cannot explain the clustering ofhantaviruses hosted by Soricomorpha and Chiroptera, whichexist in several paraphyletic clades with no correspondence bet-ween the association of the viruses, their geographic originand/or host taxonomy (figure 2; Tordo et al. 2013).
Presence of genetically distinct moles inFrance
Identification of two genetically divergent mole lineages inFrance was unexpected. Until now, only T. europaea was consi-dered to be present in France. In the Iberian Peninsula, the situa-tion is quite different and two species have been recorded fora long time. T. europaea is present in Northeast Spain(Catalunya, Navarra and the Eastern part of Pais Vasco) whilethe closely related T. occidentalis is present in the Northwest andSouth of Spain and in Portugal (Palomo et al. 2007).
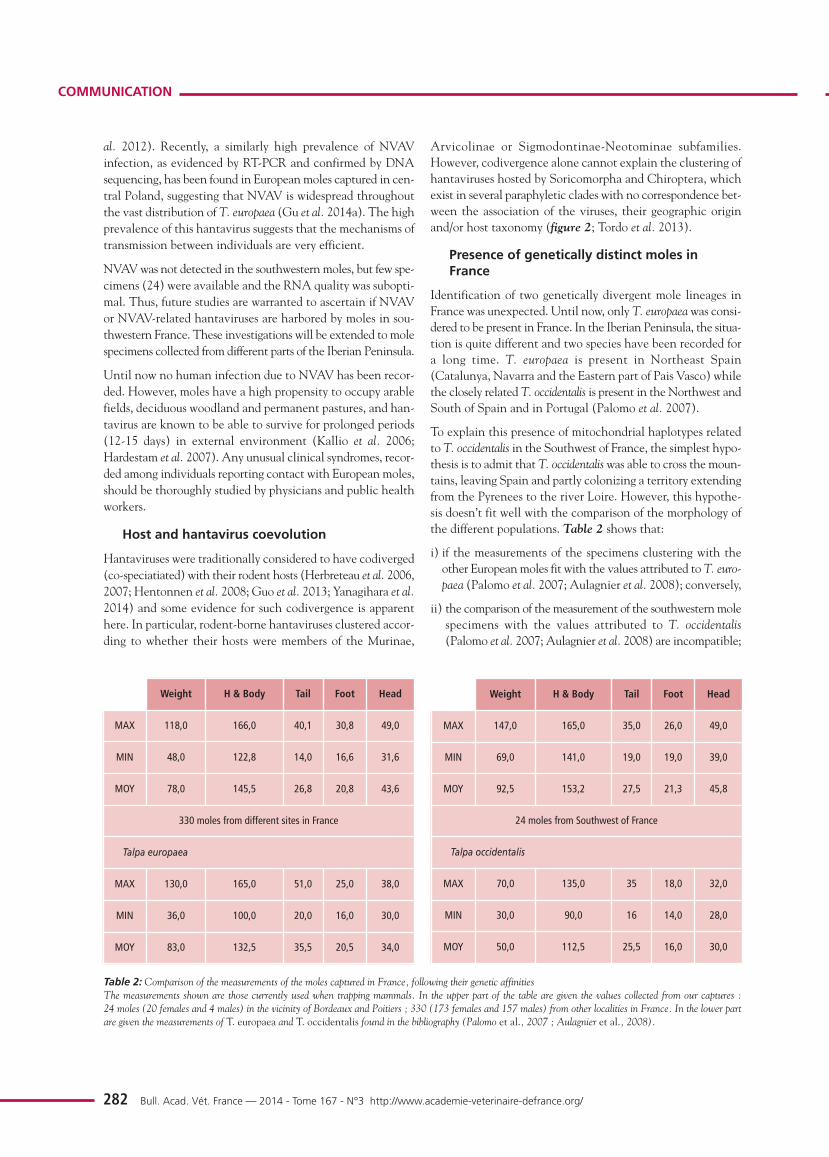
To explain this presence of mitochondrial haplotypes relatedto T. occidentalis in the Southwest of France, the simplest hypo-thesis is to admit that T. occidentaliswas able to cross the moun-tains, leaving Spain and partly colonizing a territory extendingfrom the Pyrenees to the river Loire. However, this hypothe-sis doesn’t fit well with the comparison of the morphology ofthe different populations. Table 2 shows that:
i) if the measurements of the specimens clustering with theother European moles fit with the values attributed to T. euro-paea (Palomo et al. 2007; Aulagnier et al. 2008); conversely,
ii) the comparison of the measurement of the southwestern molespecimens with the values attributed to T. occidentalis(Palomo et al. 2007; Aulagnier et al. 2008) are incompatible;
Weight H & Body Tail Foot Head
MAX 118,0 166,0 40,1 30,8 49,0
MIN 48,0 122,8 14,0 16,6 31,6
MOY 78,0 145,5 26,8 20,8 43,6
330 moles from different sites in France
Talpa europaea
MAX 130,0 165,0 51,0 25,0 38,0
MIN 36,0 100,0 20,0 16,0 30,0
MOY 83,0 132,5 35,5 20,5 34,0
Weight H & Body Tail Foot Head
MAX 147,0 165,0 35,0 26,0 49,0
MIN 69,0 141,0 19,0 19,0 39,0
MOY 92,5 153,2 27,5 21,3 45,8
24 moles from Southwest of France
Talpa occidentalis
MAX 70,0 135,0 35 18,0 32,0
MIN 30,0 90,0 16 14,0 28,0
MOY 50,0 112,5 25,5 16,0 30,0
Table 2: Comparison of the measurements of the moles captured in France, following their genetic affinities The measurements shown are those currently used when trapping mammals. In the upper part of the table are given the values collected from our captures : 24 moles (20 females and 4 males) in the vicinity of Bordeaux and Poitiers ; 330 (173 females and 157 males) from other localities in France. In the lower partare given the measurements of T. europaea and T. occidentalis found in the bibliography (Palomo et al., 2007 ; Aulagnier et al., 2008).

Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/ 283
COMMUNICATION
ACKNOWLEDGEMENTS
This work would have been impossible without the help of the professionals of the Taupgreen network (http://www.taupegreen.com/).
We especially thank: J.F. Barbot, F. Burgot, F. Cairey, M. Derosier, Y. Dupont, J.M. Georgeon, M. Giroud, J.L. Laclamette, E.J.Laffont, G. Moncel, B. Pradier, L. Quemener and R. Fons for their help in mole capture. Molecular analyses were performed at the
“Service de Systématique Moléculaire” (UMS 2700) of the MNHN.
This work was also supported in part by grant number R01AI075057 from the National Institute of Allergy and Infectious Diseases,National Institutes of Health.
iii) comparison between the two data sets identified in Franceshows that moles captured in the southwest reach thehighest size values recorded for T. europaea and are signi-ficantly heavier. Thus, moles from southwestern France maybe considered as giant specimens while T. occidentalis fromIberia may be considered as a dwarf species. These mor-phological results do not support the simplest hypothesis ofthe presence of T. occidentalis in France.
To interpret our observations, three alternative hypotheses couldbe raised:
1) an ancient phenomenon of mitochondrial introgressionoccurred between the two species;
2) existence of a hybrid zone between the two species; and,
3) existence of a new undescribed Talpa species in southwes-tern France.
To better understand the presence of mitochondrial haplotypesrelated to T. occidentalis in this part of France, it would be neces-sary to perform additional molecular analyses, especially ofmore specimens from various other localities in France and inthe Iberian Peninsula and also to sequence nuclear markers.These new investigations will be accompanied by concurrentresearch on the presence of NVAV, or another related hantavirusspecies, in moles genetically identified as T. occidentalis.
CONCLUSIONS
Our work confirms the presence of NVAV in France, in asso-ciation with its specific host, T. europaea. This result and thehigh prevalence observed in the analyzed specimens give addi-tional evidence for the probable distribution of NVAV throu-ghout the geographic range of T. europaea. Although NVAVinfection has not been recorded yet in humans, the presenceand abundance of this hantavirus increase the likelihood ofhuman exposure and should alert medical practitioners andpublic health workers to be vigilant for unusual clinical syn-dromes that may be caused by NVAV.
Hantaviruses are found in small mammals all around theWorld with the exception of the Australian and AntarticaBiorealms. An extended phylogenetic analysis of the hantavi-rus strains hosted by rodents, soricomorphs or bats suggests coe-volution between hantavirus and their hosts and implicates com-plex mechanisms including codivergence (cophylogeny), hostswitching and/or geographic isolation. The presence in Franceof two genetically different mole lineages begs the question ofthe taxonomic position of the southwestern moles and, also, ofthe potential circulation of a novel hantavirus species in theIberian mole (T. occidentalis). Further investigations are war-ranted to clarify this possibility.

COMMUNICATION
284 Bull. Acad. Vét. France — 2014 - Tome 167 - N°3 http://www.academie-veterinaire-defrance.org/
• Arai S, Ohdachi SD, Asakawa M, Kang HJ,Mocz G, Arikawa J, Okabe N, Yanagihara R.Molecular phylogeny of a newfound hantavi-rus in the Japanese shrew mole (Urotrichus tal-poides). Proc Natl Acad Sci USA. 2008;105(42):16296-301.
• Aulagnier S, Haffner P, Mitchell-Jones AJ,Moutou F, Zima J. Guide des mammifèresd’Europe, d’Afrique du Nord et du Moyen-Orient. Delachaux & Niestlé Eds, Paris, pp 272.2008.
• Dormion J. Le piégeage traditionnel des taupes.Editions Eugen Ulmer, Paris, pp. 67. 2012.Available from: http://www.taupegreen.com/methodes-eradication.htm
• Ducroz JF, Volobouev V, Granjon L. An assess-ment of the systematics of Arvicanthinerodents using mitochondrial DNA sequences:evolutionary and biogeographical implica-tions. J Mammal Evol. 2001; 8:173-206.
• Dobly A, Yzoard C, Cochez C, Ducoffre G,Aerts M, Roels S, Heyman P. Spatiotemporaldynamics of Puumala hantavirus in suburbanreservoir rodent populations. J Vector Ecol.2012; 37(2):276-83.
• Gu SH, Dormion J, Hugot J-P, Yanagihara R.High prevalence of hantavirus infection in theEuropean mole (Talpa europaea) in France.Epidemiol Infect. 2014a; 142(6):1167-71. doi:10.1017/S0950268813002197
• Gu SH, Lim BK, Kadjo B, Arai S, Kim JA,Nicolas V et al. Molecular phylogeny of han-taviruses harbored by insectivorous bats inCôte d'Ivoire and Vietnam. Viruses. 2014b;6(5):1897-910.
• Guo WP, Lin XD, Wang W, Tian JH, CongML, Zhang HL et al. Phylogeny and origins ofhantaviruses harbored by bats, insectivores, androdents. PLoS Pathog. 2013; 9(2):e1003159.
• Hardestam J, Simon M, Hedlund KO, VaheriA, Klingström J, Lundkvist Å. Ex vivo stabi-lity of the rodent-borne Hantaan virus incomparison to that of arthropod-borne mem-bers of the Bunyaviridae family. Applied andEnvironmental Microbiology, 2007; 73(8),2547-51.
• Henttonen H, Buchy P, Suputtamongkol Y,Jittapalapong S, Herbreteau V, Laakkonen J,Chaval Y, Galan M, Charbonnel N, MichauxJ, Cosson J-F, Morand S, Hugot J-P. Recent dis-coveries of new hantaviruses widen their rangeand question their origins. Ann NY Acad Sc.2008; 1149:84-9.
• Herbreteau V, Gonzalez JP, Hugot J-P.Implication of phylogenetic systematics ofrodent-borne hantaviruses on their distribution.Ann NY Acad Sc. 2006; 1081:39–56.
• Herbreteau V, Henttonen H, Yoshimatsu K,Gonzalez J-P, Suputtamongkol Y, Hugot J-P.Chapter 16: Hantavirus coevolution with theirrodent hosts. In: Tibayrenc M, ed. Encyclopediaof Infectious Diseases. Modern Methodologies.Wiley & Sons, 2007; 243-64.
• Huelsenbeck JP & Ronquist F. MRBAYES:Bayesian inference of phylogenetic trees.Bioinformatics. 2001; 17:754-5. Availablefrom: http://mrbayescsitfsuedu/:
• Kallio ER, Klingström J, Gustafsson E, ManniT, Vaheri A, Henttonen H et al. Prolonged sur-vival of Puumala hantavirus outside the host:Evidence for indirect transmission via theenvironment. J Gen Virol. 2006; 87(8), 2127-34.
• Kang HJ, Bennett SN, Sumibcay L, Arai S,Hope AG, Mocz G et al. Evolutionary insightsfrom a genetically divergent hantavirus har-bored by the European common mole (Talpaeuropaea). PLoS One. 2009; 4(7):e6149.
• Lee HW, Lee PW, Johnson KM. Isolation of theetiologic agent of Korean hemorrhagic fever.J Infect Dis. 1978; 137(3):298-308.
• Nicolas V, Mboumba JF, Verheyen E, Denys C, Lecompte E, Olayemi A et al.Phylogeographical structure and regional his-tory of Lemniscomys striatus (Rodentia:Muridae) in tropical Africa. J Biogeogr. 2008;35:2074-89.
• Palomo L, Gisbert J, Blanco JC. Atlas y LibroRojo de los Mamíferos Terrestres deEspaña.�Dirección General para laBiodiversidad-SECEM-SECEMU. Madrid, 588pp. 2007.
• Pontier D & Fouchet D. Animal health per-ceived in the light of ecology and evolution.Bull Acad Vét France. 2013; 166(4):354-63.
• Rambaut A. Se-Al. Sequence AlignmentEditor. Ver. 2.0a11. 1996 ; University ofOxford, Oxford, UK. Available from :http://tree.bio.ed.ac.uk/software/seal/
• Reynes JM. Hantavirus taxonomy and situationin France. Bull Acad Vét France. 2013;166(2):155-62.
• Ronquist F, Huelsenbeck JP, Van der Mark P.MrBayes 3.1 Manual. 2005. Available fromhttp://mrbayes.csit.fsu.edu/mb3.1_manual.pdf.
• Song J-W, Gu SH, Bennett SN, Arai S, PuorgerM, Hilbe M, Yanagihara R. Seewis virus, agenetically distinct hantavirus in the Eurasiancommon shrew (Sorex araneus). Virol J.2007;4:114.
• Swofford D. PAUP: phylogenetic analysis usingparsimony. Version 4b10 ed. Washington DC:Smithsonian Institution. Sinauer Associates,Sunderland, MA, USA, 2001.
• Thompson JD, Higgins DG, Gibson TJ. CLUS-TAL-W: improving the sensitivity of progres-sive multiple sequence alignment throughsequence weighting, position, specific gappenalties and weight matrix choice. NucleicAcids Res. 1994; 22:4673–80.
• Tordo N, Castel G, Filippone C, MarianneauP. Recent data on hantaviruses and perspectivesfor research Bull Acad Vét France. 2013;166(4):364-71.
• Winnepenninckx B, Backeljau T, De WachterR. Extraction of high molecular weight DNAfrom molluscs. Trends Genet. 1993; 9:407.
• Yanagihara R, Gu SH, Arai S, Kang HJ, SongJW. Hantaviruses: Rediscovery and new begin-nings. Virus Res. 2014 Jan 8. pii: S0168-1702(13)00484-X. doi: 10.1016/j.virusres.2013.12.038. [Epub ahead of print]
BIBLIOGRAPHIE