distinct transcriptional pathways regulate basal and activated
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, May 2003, p. 3377–3391 Vol. 23, No. 100270-7306/03/$08.00�0 DOI: 10.1128/MCB.23.10.3377–3391.2003
Distinct Transcriptional Pathways Regulate Basal and Activated MajorHistocompatibility Complex Class I Expression
T. Kevin Howcroft,* Aparna Raval, Jocelyn D. Weissman, Anne Gegonne, and Dinah S. SingerExperimental Immunology Branch, National Cancer Institute, Bethesda, Maryland 20892-1360
Received 14 January 2003/Accepted 25 February 2003
Transcription of major histocompatibility complex (MHC) class I genes is regulated by both tissue-specific(basal) and hormone/cytokine (activated) mechanisms. Although promoter-proximal regulatory elements havebeen characterized extensively, the role of the core promoter in mediating regulation has been largely unde-fined. We report here that the class I core promoter consists of distinct elements that are differentially utilizedin basal and activated transcription pathways. These pathways recruit distinct transcription factor complexesto the core promoter elements and target distinct transcription initiation sites. Class I transcription initiatesat four major sites within the core promoter and is clustered in two distinct regions: “upstream” (�14 and�18) and “downstream” (�12 and �1). Basal transcription initiates predominantly from the upstream startsite region and is completely dependent upon the general transcription factor TAF1 (TAFII250). Activatedtranscription initiates predominantly from the downstream region and is TAF1 (TAFII250) independent. USF1augments transcription initiating through the upstream start sites and is dependent on TAF1 (TAFII250), afinding consistent with its role in regulating basal class I transcription. In contrast, transcription activated bythe interferon mediator CIITA is independent of TAF1 (TAFII250) and focuses initiation on the downstreamstart sites. Thus, basal and activated transcriptions of an MHC class I gene target distinct core promoterdomains, nucleate distinct transcription initiation complexes and initiate at distinct sites within the promoter.We propose that transcription initiation at the core promoter is a dynamic process in which the mechanismsof core promoter function differ depending on the cellular environment.
Major histocompatibility complex (MHC) class I genes, likemost typical housekeeping genes, are constitutively active in alltissues. However, unlike housekeeping genes, the relative lev-els of class I expression vary dramatically among differenttissues. The highest levels of expression occur in tissues andcells of the immune system; the lowest levels are observed inthe nervous system and germ line cells (18, 33, 51). Thus,although the class I promoter is thought to be constitutivelyaccessible to the general transcription machinery, it is alsosubject to diverse tissue-specific regulatory influences. To-gether, the constitutive and tissue-specific regulatory mecha-nisms determine the basal level of class I expression in anytissue at any given time.
MHC class I expression is also dynamically modulated in thepresence of certain cytokines, hormones, and other inflamma-tory agents. For example, interferon (IFN) increases class Itranscription, whereas thyroid-stimulating hormone (TSH) re-presses it (11, 17, 52). Thus, class I expression is regulated bytwo distinct pathways. The basal pathway regulates homeo-static expression and establishes the tissue-specific “set-point”level of class I expression in any given tissue. In contrast, themodulated pathways dynamically regulate, either specificallyactivating (activated pathway) or repressing (repressed path-way), class I expression in response to transiently expressedcytokines and hormones.
The upstream DNA elements regulating basal and modu-lated expression of the MHC class I gene, PD1, have been
intensively investigated. All regulatory elements necessary toconfer normal patterns of class I expression are containedwithin about 1 kb upstream of the coding sequence (14, 50).Distinct domains regulate basal transcription and dynamicallyactivated transcription: one that is located between �800 and�700 bp is responsible for tissue-specific expression, and an-other one located between �500 and �50 bp is responsible forboth activated and basal expression (23, 25, 37, 41, 64). Amongthe elements that regulate basal class I expression is a canon-ical E-box (at positions �314 to �309) recognized by thetranscription factor USF (23). USF consists of two familymembers, USF1 and USF2 (23, 54). Both are ubiquitouslyexpressed; their expression is not known to be altered by hor-mone/cytokine stimulation, and therefore they are consideredto contribute to basal class I expression.
The modulatory domain contains both elements that sup-port basal expression and dynamically modulated class I tran-scription in response to cytokines, hormones, and inflamma-tory agents. Examples of the latter include enhancer A (enhA), an IFN-stimulated response element, and a compositeRF-X/cyclic AMP response element (CRE) that modulateclass I expression by binding inducible trans-acting factors (2,11, 18, 19, 50, 59). The CRE mediates both induction of tran-scription by gamma IFN (IFN-�) and repression by TSH (43,52).
The IFN-� mediator, CIITA, is a non-DNA-binding coacti-vator that interacts with constitutively expressed RF-X andATF trans-acting factors already bound at the RF-X/CRE site(27, 42). CIITA contributes to class I expression in B lympho-cytes, dendritic cells, and macrophages, cell types in whichCIITA is constitutively expressed. In addition, IFN-� induces
* Corresponding author. Mailing address: Experimental Immunol-ogy Branch, NCI, Bldg. 10, Rm. 4B-17, NIH, 10 Center Dr., MSC 1360,Bethesda, MD 20892-1360. Phone: (301) 496-9097. Fax: (301) 480-8499. E-mail: [email protected].
3377
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

CIITA in many other cell types, resulting in their activatedclass I expression (20, 38, 43).
MHC class I gene expression depends upon the proper in-tegration of regulatory signals from these upstream domainswith the appropriate general transcription machinery at thecore promoter. However, a detailed analysis of the MHC classI core promoter and its contribution to these diverse regulatorypathways is lacking. The present study was undertaken to beginto characterize the core promoter and define the mechanism(s)by which the basal and activated pathways are integrated at theMHC class I core promoter to achieve appropriate levels oftranscription.
The core promoter is defined as the minimal length of DNAsufficient to direct accurate transcription initiation by RNApolymerase (Pol) II (9). Thus, the core promoter region reg-ulates three fundamental steps in transcription. First, it pro-vides a docking site for general transcription factors (GTFs)capable of recruiting RNA Pol II and associated factors re-quired for basal transcription. Second, through the assembledGTFs and RNA Pol II, it serves as a molecular platform tointegrate regulatory signals delivered by upstream silencer andenhancer elements to appropriately adjust the level of pro-moter activity. Third, a core promoter determines the startsite(s) of transcription (3, 16, 21, 44, 45, 55, 60).
Analysis of a number of cellular and viral promoters has ledto the identification of a small number of core promoter ele-ments. The two most common and best-characterized corepromoter elements are the TATAA box and Inr motifs. TheTATAA box is an A/T-rich sequence located about 30 basesupstream of initiation (7). The Inr is a pyrimidine-rich elementthat generally spans the site of transcript initiation (29, 30, 36,56, 57, 65). Consensus sequences for both TATAA and Inrelements have been defined, although considerable sequencevariation occurs among promoters (36, 53). Different core pro-moters may contain one, both, or neither element. For exam-ple, whereas the TdT promoter has only an Inr (56), the AdMLhas both TATAA and Inr motifs (1). In Drosophila melano-gaster, a recent analysis found an equal distribution of thesevarious structures (31). Additional elements, including aTFIIB recognition element (BRE) and a downstream pro-moter element (DPE), are also found in many core promoterregions (5, 6, 9, 32). Surprisingly, many promoters do not havea recognizable homolog of any of these elements, suggestingthat additional promoter elements remain to be identified (31).A number of such promoters, many of them regulating house-keeping genes, are located within CpG islands (15).
In contrast to the modulatory and tissue-specific domains,the class I core promoter region is relatively less characterized.By sequence homology, three elements are identified in theclass I core promoter region: TATAA- and Inr-like motifs anda CA/GT-rich region (S-box). The S-box sequence is homolo-gous to those recognized by the Sp1 family of transcriptionfactors in other promoters (10, 13, 35). The relative functionalimportance of these elements in MHC class I transcription hasnot been established. Nor has the site(s) of transcription initi-ation been determined.
Here we report that the class I core promoter is a complexstructure, in which no single element subserves all transcrip-tion functions. Rather, basal and activated transcription targetdistinct regions of the core promoter and have distinct core
promoter element requirements. Transcription initiates atmultiple sites, within two general domains: basal transcriptioninitiates largely upstream of �6 bp, whereas activated tran-scription predominantly initiates downstream of �6. Further-more, basal transcription is dependent upon Sp1 binding to theCA/GT-rich region of the S-box. In contrast, activated tran-scription is neither dependent upon the CA/GT rich region norSp1 binding. Basal and activated transcription also differ intheir requirement for a TAF1 (TAFII250)-containing initiationcomplex. Thus, basal and activated modes of class I transcrip-tion represent distinct molecular pathways that engage differ-ent components of the core promoter. These findings suggestthat the complex structure of the MHC class I core promoterallows it to uniquely integrate cell type and/or dynamic up-stream regulatory signals to direct appropriate levels of tran-scription.
MATERIALS AND METHODS
Cell lines and cultivation. The HeLa epithelial, baby hamster kidney (BHK)and tsBN462 cell lines were grown in Dulbecco modified Eagles medium sup-plemented with 10% fetal bovine serum, 2 mM L-glutamine, 20 mM HEPES (pH7.2), and gentamicin sulfate (10 �g/ml). Jurkat (T-cell) and M12 (B cell) lineswere grown in RPMI 1640 supplemented with 10% fetal bovine serum, 2 mML-glutamine, 55 �M �-mercaptoethanol, 100 �M minimal essential amino acids,1 mM sodium pyruvate, 20 mM HEPES (pH 7.2), and gentamicin sulfate (10�g/ml). Cell lines were maintained in a humidified incubator at 37°C in 7% CO2,except tsBN462 which were maintained at 32°C in 7% CO2.
Plasmids and cloning strategies. The MHC class I promoter used in thesestudies derived from the swine class I gene, PD1 (14, 49). The PD1 promotertruncation series, ligated to the chloramphenicol acetyltransferase (CAT) re-porter, was previously described (24, 25). To generate �50CAT, �313CAT wasdigested with NarI and NdeI (New England Biolabs), followed by Klenow fill-inand blunt-end ligation. The NarI site is at position �50 within the class Ipromoter, and the NdeI site is located immediately 5� of the class I promotersequences in the CAT reporter. The class I promoter sequences, extending fromthe 5� XbaI site to the HindIII at position �1, were excised from the CAT 5�truncation series and were ligated into the NheI/HindIII sites in the multiplecloning region of the pGL2B luciferase expression vector. Synthetic double-stranded oligonucleotides were inserted into the HindIII site of pSV0CAT togenerate the InrWT core promoter and derivative mutant reporter constructs(24). The sense strand sequences of the oligonucleotides synthesized (from �30to �14) are illustrated in Fig. 4A. The NarI/NcoI promoter fragments of the Inrcore promoter series were subsequently cloned into NarI/NcoI digested�416CAT to generate the �416Inr series. The NarI site is at position �50 withinthe class I promoter, and the NcoI site is located within the CAT coding region.The mammalian expression vector Flag-CIITA wild type (WT) has been previ-ously described (43); CIITA WT was cloned into the baculovirus transfer vectorPVL1293 at the EcoRI site.
Transfections. Transient transfections were performed by using a constantamount of DNA (5 �g). At 24 h prior to transfection 106 HeLa or BHK cellswere seeded in 100-mm tissue culture dishes. Transfections utilized standardcalcium phosphate precipitation as previously described (25). The medium wasreplaced 24 h after transfection with fresh medium, and cells were harvestedafter an additional 24 h. Temperature-sensitive tsBN462 cells were left at 32°Cfor 24 h after transfection and then shifted to 39°C (restrictive temperature) orleft at 32°C (permissive temperature) for an additional 24 h (63). HeLa cells andL cells were maintained at 37°C for 48 h after transfection. Reporter activity wascorrected by cotransfecting an internal control plasmid control, either pSV2LUC(200 ng) or CMV–�-Gal (50 ng). Jurkat and M12 lymphocytes were transfectedby electroporation (250 V, 975 �F) by using Bio-Rad gene pulser II electropo-rator. All CAT enzyme assays were measured in the linear range; control 14C-labeled chloramphenicol values ranged between 20 and 80% among the differentexperiments. Luciferase and �-galactosidase determinations were made by usinga Monolight 2010 luminometer (Analytical Luminescence Laboratory).
Preparation of recombinant CIITA protein. Flag-CIITA WT or the mutantswere expressed in Sf9 cells by using baculovirus-mediated transfection accordingto the manufacturer’s protocol (using the Pharmingen kit). Recombinant Flag-
3378 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

CIITA (rCIITA) was immunoprecipitated by using anti-Flag M2 agarose beads(Sigma) and eluted with 100 �g of Flag peptide/ml (43).
Isolation of RNA and 5�RACE. The PD1 transgenic mouse strain used for invivo start site analysis, CAT.516, was previously described (37). The PD1, stablytransfected, murine L-cell line, 93B2, was previously described (49). Total RNAfrom transgenic spleen or 93B2 cell line was isolated by using RNA STAT-60(Tel-Test, Inc.). Transcription start sites, utilized by transgenic splenocytes, weredetermined by using the SMART RACE (rapid amplification of cDNA ends)cDNA amplification kit (Clontech) with 1 mg of total splenic RNA; RACE-Ready cDNA was subsequently PCR amplified by using the SMARTII and CATgene-specific primers (GGTGGTATATCCAGTGATTTTTTTCTCCAT). Am-plified products were cloned into the TA cloning vector (Invitrogen) and used totransform DH5� competent bacteria (Invitrogen). Ampicillin-resistant colonieswere screened for the presence of the CAT sequence by using the CAT primer.DNA prepared from positive colonies was sequenced by using the ABI Prism dyeterminator cycle sequencing ready reaction kit (Perkin-Elmer). Analysis of invivo start sites utilized in the 93B2 PD1 stably transfected L-cell line was done byprimer extension as previously described (62).
In vitro transcription and coupled primer extension. In vitro transcriptionreaction mixtures contained 2 �g of class I CAT reporter construct, 6 mMMgCl2, 0.8 mM deoxynucleoside triphosphates, and 30 U of HeLa nuclearextract (Promega) in 20 mM HEPES (pH 7.9), 100 mM KCl, 0.2 mM EDTA, 0.5mM dithiothreitol, and 20% glycerol in a total of 25 �l was incubated at 20°C for60 min. Analysis of the in vitro-transcribed RNA was done by primer extensionas previously described (62).
RESULTS
A minimal MHC class I core promoter is transcriptionallyactive in vivo. To determine the functional boundaries of theclass I promoter, we compared the activity of an extended�416WT class I promoter construct that terminates at �416bp and contains a series of upstream enhancer elements withthe promoter-proximal �50WT construct, truncated at �50bp. Both promoter segments have a common 3� terminus at�14 and were ligated to a CAT reporter (Fig. 1A). The con-structs were transiently transfected into HeLa epithelial cells,and their activities were compared. Surprisingly, �50WT re-tained promoter activity that was significantly higher than thepromoter-less control (pSVO) and was less than twofold re-duced compared to �416WT (Fig. 1A). Similar results wereobtained when a luciferase reporter was substituted for theCAT reporter gene (data not shown). These data indicate thatthe class I core promoter is contained within a 64-bp segment,between bp �50 and bp �14, and remains active in the absenceof upstream elements.
MHC class I transcription initiates at multiple start sites.The studies described above demonstrate that the MHC classI core promoter, unlike most described core promoters, istranscriptionally active. To determine that the core promoterinitiates transcription properly, the transcript initiation profilegenerated by the minimal promoter construct (�50WT) wascompared to that of the extended WT class I promoter(�416WT) in in vitro transcription assays with HeLa nuclearextract. With either template, a spectrum of transcripts wasobserved initiating from two distinct regions within the corepromoter (Fig. 1B, shadowed ellipses): an upstream regiondefined by the A�18 and A�14 start sites, and a downstreamregion defined by the A�1 and A�12 start sites (Fig. 1B, lanes3 and 5). These start sites were not observed in controls con-taining either HeLa nuclear extract alone (Fig. 1B, lane 1) orMHC class I promoter DNA template in the absence of HeLanuclear extract (Fig. 1B, lanes 2 and 4). In the presence of 10�g of �-amanitin/ml, class I transcripts are inhibited by �70%,
indicating that these are RNA Pol II-derived transcripts (datanot shown). These data indicate that MHC class I transcriptscan initiate at multiple sites in the core promoter region. Fur-thermore, the same pattern was observed in both the extended�416WT and core promoter �50WT class I constructs. Thus,the absence of upstream sequences does not alter transcriptioninitiation sites from the core promoter. Further, under theseconditions of in vitro basal transcription, the A�14 site was themost frequently utilized site.
To verify that the transcription initiation sites observed in invitro transcription reflected in vivo usage, we examined startsite usage in a murine fibroblast cell line containing a stablyintegrated full-length MHC class I gene, PD1, that has beenshown to direct normal expression of class I in transgenic mice(14, 49). Primer extension analysis of RNA derived from thiscell line also revealed multiple initiation sites, a finding con-sistent with the in vitro transcription data. As observed in invitro transcription, the dominant start site was at the A�14position, with additional weaker sites at positions �18, �1, and�12. In addition, weak initiation sites mapped to positions �3,�5, �5, and �7 that were not routinely observed in vitro (Fig.1C). In murine fibroblasts and in vitro in HeLa nuclear ex-tract—both conditions of largely basal transcription—tran-scription initiation from the bp �14 site predominates. Takentogether, these data indicate that both in vivo and in vitro,transcription initiates within the core promoter but at multiplesites. These findings demonstrate that (i) in vitro transcriptiondoes not generate aberrant initiation sites and (ii) DNA se-quences outside of the core promoter (bp �50 to �14) do notdetectably contribute to the pattern of basal transcription ini-tiation.
Mutational analysis of the class I core promoter region. Thesequence of the 64-bp class I core promoter is shown in Fig.2A. To define the constituent DNA sequence element(s) re-quired for core promoter function, we generated constructscontaining mutations spanning the core promoter (Fig. 2A,mutated sequences are listed underneath each sequence) anddetermined their effects on transcription initiation in vitro andpromoter activity in vivo.
Two of the mutations, mut T (bp �30 to �26) and mut I (bp�3 to �7), encompassed sequences of the core promoter withhomology to known promoter elements: the 5� TCTAA se-quence is orthologous to the TATAA box of the mouse class Igenes, whereas the 3� CTCAGCTT sequence is homologous toa consensus Inr sequence. Both mut T and mut I supportedtranscription and used the same initiation sites as the WTpromoter in vitro (Fig. 2B). However, the relative usage ofthese in vitro transcription start sites differed. Most notablewas a consistent reduction in the usage of A�18, A�1, andA�12 start sites relative to A�14 observed with mut T (Fig.2B, compare lanes 3 and 5). This difference was observedregardless of the extent of 5� upstream sequences (data notshown). In contrast, the frequency of start sites utilized by mutI was indistinguishable from that of the WT core promoter(Fig. 2B, compare lanes 3 and 7). [The increase in the mut IA�12 start site, seen in Fig. 2B, was not routinely observed.]Neither mut T nor mut I affected transcript initiation at the�14 start site (Fig. 2B, compare lanes 3, 5, and 7). Mutationsupstream of bp �30 had no effect on promoter activity (datanot shown). Taken together, these results extend our previous
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3379
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 1. Identification of a minimal MHC class I core promoter region. (A) HeLa cells were transiently transfected with MHC class I promoterconstructs �416WT, �50WT, and promoterless pSV0. The �50WT extends upstream from positions �14 to �50, whereas �416WT extendsupstream from positions �14 to �416 and contains USF and CRE regulatory elements. Class I promoter and pSV0 control constructs,diagrammed on the left, are all ligated to the CAT reporter. Data are expressed as relative percentages of acetylation corrected to an internaltransfection control, pSV2LUC. The black triangle represents the core promoter region. Error bars indicate the standard error. (B) In vitrotranscriptional start sites utilized by �416WT and �50WT constructs were determined by primer extension of in vitro-transcribed RNA. HeLaextract alone control is shown in the lane 1. Background bands present in the HeLa extract alone control are indicated by solid black circles.DNA-only controls for �416WT and �50WT are shown in lanes 2 and 4, respectively. The arrows along the left indicate specific transcriptionalstart sites in lanes 3 and 5, in the presence of HeLa nuclear extract. Shaded ellipses representing upstream and downstream transcription start siteregions (where the major start sites, indicated by arrows, were observed to originate) are provided on the left for orientation. The scan encompassesthe length of the �50WT class I sequences. A sequence ladder for the class I promoter used in these studies (shown on the right) was generatedby using the same primer used in primer extension analysis of in vitro-synthesized RNA and was used to determine the precise start sites. Minorstart sites were also inconsistently observed at other sites upon longer exposure. (C) RNA was prepared from a fibroblast cell line (93B2) containinga stably integrated PD1 MHC class I gene. Increasing amounts of total RNA, 3 and 10 �g (lanes 1 and 2) were analyzed by primer extension todetermine start site utilization in vivo. Shadowed ellipses representing the upstream and downstream transcription regions where the major startsites originate in in vitro transcription (indicated by arrows) are provided on the left for orientation. The �3 and �5 start sites, observed in vivowith 93B2 RNA, would have been obscured by a nonspecific band generated by HeLa nuclear extract in in vitro transcription reactions (see Fig.1B). A sequence ladder of the class I promoter is also included.
3380 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
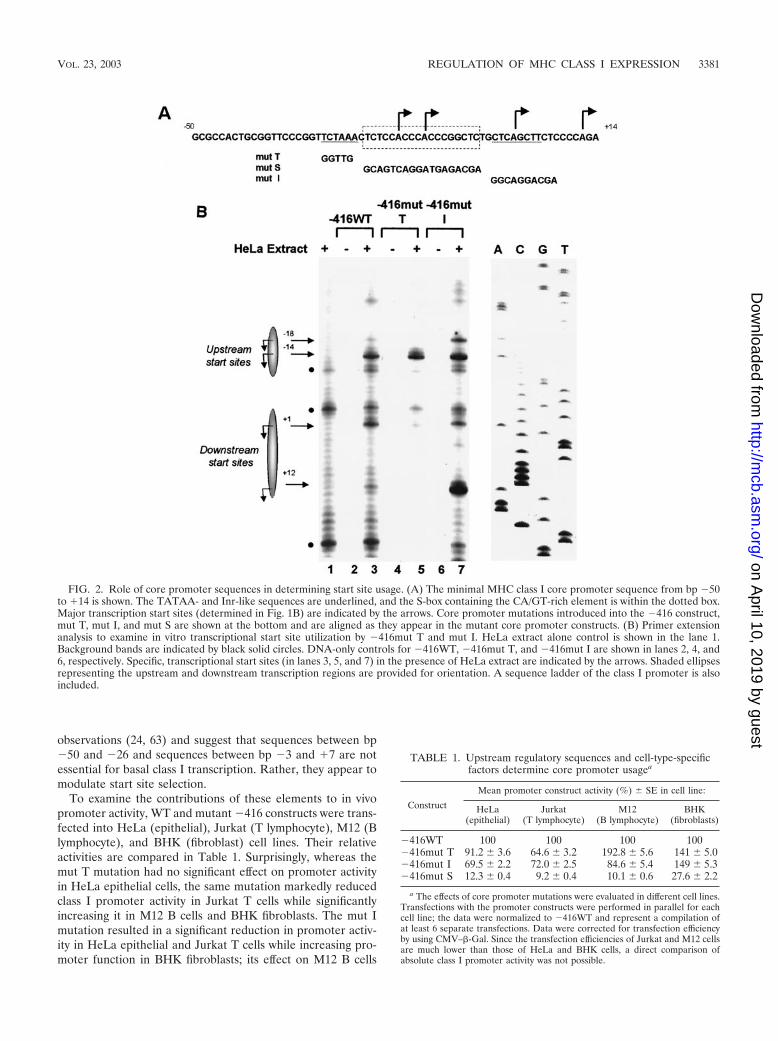
observations (24, 63) and suggest that sequences between bp�50 and �26 and sequences between bp �3 and �7 are notessential for basal class I transcription. Rather, they appear tomodulate start site selection.
To examine the contributions of these elements to in vivopromoter activity, WT and mutant �416 constructs were trans-fected into HeLa (epithelial), Jurkat (T lymphocyte), M12 (Blymphocyte), and BHK (fibroblast) cell lines. Their relativeactivities are compared in Table 1. Surprisingly, whereas themut T mutation had no significant effect on promoter activityin HeLa epithelial cells, the same mutation markedly reducedclass I promoter activity in Jurkat T cells while significantlyincreasing it in M12 B cells and BHK fibroblasts. The mut Imutation resulted in a significant reduction in promoter activ-ity in HeLa epithelial and Jurkat T cells while increasing pro-moter function in BHK fibroblasts; its effect on M12 B cells
FIG. 2. Role of core promoter sequences in determining start site usage. (A) The minimal MHC class I core promoter sequence from bp �50to �14 is shown. The TATAA- and Inr-like sequences are underlined, and the S-box containing the CA/GT-rich element is within the dotted box.Major transcription start sites (determined in Fig. 1B) are indicated by the arrows. Core promoter mutations introduced into the �416 construct,mut T, mut I, and mut S are shown at the bottom and are aligned as they appear in the mutant core promoter constructs. (B) Primer extensionanalysis to examine in vitro transcriptional start site utilization by �416mut T and mut I. HeLa extract alone control is shown in the lane 1.Background bands are indicated by black solid circles. DNA-only controls for �416WT, �416mut T, and �416mut I are shown in lanes 2, 4, and6, respectively. Specific, transcriptional start sites (in lanes 3, 5, and 7) in the presence of HeLa extract are indicated by the arrows. Shaded ellipsesrepresenting the upstream and downstream transcription regions are provided for orientation. A sequence ladder of the class I promoter is alsoincluded.
TABLE 1. Upstream regulatory sequences and cell-type-specificfactors determine core promoter usagea
Construct
Mean promoter construct activity (%) SE in cell line:
HeLa(epithelial)
Jurkat(T lymphocyte)
M12(B lymphocyte)
BHK(fibroblasts)
�416WT 100 100 100 100�416mut T 91.2 3.6 64.6 3.2 192.8 5.6 141 5.0�416mut I 69.5 2.2 72.0 2.5 84.6 5.4 149 5.3�416mut S 12.3 0.4 9.2 0.4 10.1 0.6 27.6 2.2
a The effects of core promoter mutations were evaluated in different cell lines.Transfections with the promoter constructs were performed in parallel for eachcell line; the data were normalized to �416WT and represent a compilation ofat least 6 separate transfections. Data were corrected for transfection efficiencyby using CMV–�-Gal. Since the transfection efficiencies of Jurkat and M12 cellsare much lower than those of HeLa and BHK cells, a direct comparison ofabsolute class I promoter activity was not possible.
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3381
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

was intermediate (Table 1). Additional mutations upstream of�30 bp had no effect on basal promoter function in HeLa cells(data not shown).
These data suggest that (i) the class I core promoter is notstrictly dependent on the TATAA-like or Inr-like elements and(ii) these sequences function in a cell-type-specific fashion tomodulate promoter activity. Further, since neither mutationeliminates promoter function, these data suggest that the classI promoter contains an alternative element(s) responsible forrecruiting RNA Pol II for basal transcription.
Mutating the S-box greatly reduces MHC class I basal tran-scription. We next characterized the effect of mutating the18-bp region (S-box) located between bp �23 and �6 (Fig. 2A,mut S). In contrast to the mut T and mut I mutations, mut Sdramatically reduced promoter activity both in vivo (Table 1)(24) and in vitro (Fig. 3A). In mut S, initiations at the upstreamA-14 site were markedly reduced relative to those at the down-stream �1 or �12 sites (Fig. 3A, compare lanes 2 and 3). Thus,a critical sequence(s) required for basal MHC class I expres-sion resides within the S-box.
The minimal sequence(s) within the S-box responsible forclass I core promoter transcription was determined by gener-ating a series of overlapping 4-bp mutations spanning the S-box region and assessing their activities in in vitro transcriptionreactions (Fig. 3B). Like the original Mut S, mutations span-ning the central CA/GT-rich region of the S-box (M1 to M3)reduced in vitro transcription at all start sites (Fig. 3C, com-pare lanes 1 and 2 to 4). Mutant M4 had slightly reducedrelative �1 and �12 start site usage (Fig. 3C, lane 5). The most3� mutation, M5, had no effect on start site usage (Fig. 3C, lane6). These data indicate that the CA/GT-rich sequence betweenbp �21 and �12 is critical for class I basal promoter activity.
A specific complex binds the CA/GT-rich S-box sequence. Inorder to identify transcription factors that interact with theS-box, the 64-bp core promoter region was used as a probe ingel shift analysis. Two principal complexes, indicated by thearrowheads, were generated with the WT class I core promoterprobe and HeLa nuclear extract. (Fig. 4A, lane 2). Both com-plexes were competed for by increasing unlabeled WT com-petitor oligonucleotide (Fig. 4A, lanes 3 to 5). In contrast, amut S oligonucleotide only competed the faster-migratingcomplex, indicated by the open arrowhead (Fig. 4A, lanes 6 to8). When used directly as a probe in gel shift analysis, the mutS oligonucleotide failed to generate the slower-migrating S-boxspecific complex (data not shown). These data indicate that theslower-migrating complex, indicated by the solid arrowhead,
FIG. 3. The S-box is essential for basal MHC class I transcription.(A) Mutation of the S-box in the extended �416 class I promoterconstruct (�416mut S, Fig. 2A) dramatically reduced transcriptioninitiations in vitro. In vitro transcription reactions of �416WT and�416mut S constructs were analyzed by primer extension analysis.HeLa extract alone control is shown in the lane 1. Specific, transcrip-tional start sites generated by �416WT and �416mut S, in lanes 2 and3, respectively, are indicated by the arrows. Additional, less-utilizedstart sites are observed throughout the WT core promoter region uponthe longer exposure time required to observe initiations in �416mut S.(B) Scanning mutations of 4 bp across the S-box region were intro-duced into the �416 promoter construct to map the minimal S-boxregion required for basal transcription. Brackets indicate the 4-bp
regions, M1 to M5, that were individually introduced into the �416promoter construct. The mutated sequences appear at the bottom.(C) The effect of the S-box scanning mutants (M1 to M5) on in vitrotranscription of the extended �416 promoter in HeLa extract wasexamined by primer extension analysis. Transcription start sites gen-erated by the WT promoter are shown in lane 1, and those generatedby mutants M1 to M5 are shown in lanes 2 to 6, respectively. Thearrowheads indicate major transcriptional start sites, and the solidcircle indicates a background band observed in HeLa extract control(not shown). Shadowed ellipses representing the upstream and down-stream transcription regions where the major start sites were observedto originate (indicated by arrows) are provided for orientation.
3382 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 4. The transcription factor Sp1 binds CA/GT-rich S-box element. (A) Gel shift analysis by using the class I core promoter region (�50to �14), shown in Fig. 2A, as a probe to identify specific core promoter binding complexes. Lane 1 contains probe alone in the absence of HeLaextract. Lane 2 shows two complexes formed in the presence of HeLa extract, indicated by the arrowheads. Lanes 3 to 5 and lanes 6 to 8 representincreasing (102-, 103-, and 104-fold) amounts of unlabeled WT or mut S oligonucleotide competitors, respectively. The solid arrowhead indicatesthe S-box specific complex; the open arrowhead indicates a nonspecific single-strand binding complex. (B) Mapping the specific S-box bindingcomplex by using the M1 to M5 panel of mutant oligonucleotides detailed in Fig. 3B. Lane 1 contains the core promoter probe in the absence ofHeLa extract. Lane 2 shows the two complexes formed in the presence of HeLa extract, indicated by the arrowheads. Lanes 3 and 4 and lanes 5and 6 represent increasing unlabeled WT or mut S oligonucleotide competitors, respectively. Increasing unlabeled, mutant oligonucleotidecompetitors, M1 to M5, are shown in lanes 7 to 16, respectively. The S-box-specific complex is indicated by the solid arrowhead. (C) Antibodysupershift analysis of the specific S-box binding complex. Lane 1 contains core promoter (�50 to �14) probe in the absence of HeLa extract. Lane2 contains complexes formed in the presence of HeLa extract, indicated by the arrowheads; the solid arrowhead is the S-box-specific complex. Lane3 contains control antisera; lanes 4 to 7 contain antisera specific for Sp1, Sp2, Sp3, and Sp4, respectively. The supershifted complex, in the presenceof anti-Sp1 antisera, is indicated by the solid circle. (D) Purified Sp1 protein (pSp1) binds to the S-box region of the core promoter. Lane 1 containsprobe alone. Lane 2 contains the complex formed in the presence of pSp1, indicated by the solid arrowhead. Lane 3 contains specific anti-Sp1antiserum against Sp1; the supershifted complex is indicated by the solid circle. Lanes 4 and 5 contain WT and mut S unlabeled competitoroligonucleotides, respectively.
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3383
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

specifically interacts with S-box sequences contained in theclass I core promoter region.
In order to determine the precise binding site within theS-box, we utilized the panel of derivative oligonucleotides con-taining the overlapping 4-bp mutations across the S-box region,presented in Fig. 3B, as unlabeled competitors in gel shiftanalysis (Fig. 4B). Whereas an unlabeled WT oligonucleotidecould compete complex binding to the WT core promoterprobe, the scanning mutations M1, M2, and M3 were unable todo so (Fig. 4B, lanes 7 to 12). In contrast, mutations M4 andM5 were able to inhibit complex formation, although some-what less efficiently than WT (Fig. 4B, lanes 13 to 16). Muta-tions outside the S-box (mut T) efficiently competed for theS-box specific complex (data not shown). This pattern of com-petition parallels the effect of these mutations on in vitrotranscription (Fig. 3C). Taken together, these data indicatethat a specific binding complex interacts with the central CA/GT-rich sequences, bp �21 to �12, of the S-box and suggestthat it is necessary for transcription.
The transcription factor Sp1 binds the S-box. CA/GT-richsequences, similar to the class I core promoter element de-scribed above, have been reported to be Sp1-binding sites inother promoters (10, 13, 35). To determine whether Sp1 is acomponent of the S-box-specific complex, we added anti-Sp1antisera to the gel shift reaction (Fig. 4C). Addition of anti-Sp1antisera resulted in a supershift of the slower-migrating S-boxcomplex (Fig. 4C, lane 4, solid circle). Control antisera had noeffect (Fig. 4C, lane 3). Antisera to other Sp1 family members,including Sp2, Sp3, or Sp4, also had no effect on the S-boxcomplex (Fig. 4C, lanes 5 to 7). Thus, the S-box-specific com-plex contains Sp1.
The faster-migrating complex was unaffected by the additionof anti-Sp1 antisera (Fig. 4C, open arrowhead). This complexwas composed of single-stranded DNA binding factors since itcould be effectively competed for by unlabeled sense or anti-sense single-stranded oligonucleotides (data not shown); theidentity of these factors was not pursued. The slower-migratingcomplex, containing Sp1, was not competed for by increasingunlabeled single-stranded competitor oligonucleotides (datanot shown).
Purified Sp1 protein (pSp1) generated a complex with thecore promoter probe that migrated with the same mobility asthe endogenous Sp1-containing complex derived from HeLaextract (Fig. 4D and data not shown). Similar to the Sp1-containing complex observed with HeLa extracts, pSp1 wassupershifted by anti-Sp1 antisera (Fig. 4D, lane 3) and wascompeted for by WT but not by mut S oligonucleotides, re-spectively (Fig. 4D, lanes 4 and 5). These data demonstratethat Sp1 specifically interacts with the CA/GT-rich sequencesin the class I core promoter.
The CA/GT-rich sequence is required for MHC class I ex-pression in vivo. The above data suggest a positive correlationbetween Sp1 binding and MHC class I promoter activity. Con-sistent with this correlation, we have demonstrated previouslythat Sp1 activates class I promoter activity in Schneider Dro-sophila cells, which lack endogenous Sp1 (24). Therefore, wedetermined whether the MHC core promoter mutant,�416Mut S, which does not bind Sp1 and does not supporttranscription in vitro, affects class I promoter activity in vivo.We examined basal class I promoter activity of �416Mut S in
transiently transfected HeLa (epithelial), Jurkat T and M12 B(lymphocyte), and BHK (fibroblast) cell lines (Table 1). Thefour cell lines utilized in this analysis all expressed similaramounts of Sp1 protein and generated comparable Sp1-depen-dent complex formation with the WT core promoter probe ingel shift analyses (data not shown). Mutation of the Sp1-bind-ing site led to a decrease of 75 to 90% in class I promoteractivity in all cell lines (Table 1). These results suggest thatSp1, binding to the core promoter, recruits a transcriptionalcomplex critical for MHC class I basal expression.
Basal transcription in vivo initiates in the upstream startsite region. The results described above demonstrate that basalclass I transcription initiates at multiple sites and that it isdirected by an Sp1-dependent mechanism. Although muta-tions that compromise Sp1 binding reduce all in vitro transcrip-tion initiations, they have a greater effect on upstream (A�18and A�14) than downstream (A�1 and A�12) start site re-gions (Fig. 3C). This observation raises the question ofwhether the upstream and downstream start site regions arefunctionally distinct in vivo. To investigate this question, weintroduced a mutation in the class I core promoter region thatwould distinguish upstream from downstream initiations, whilepreserving the overall structural and spatial integrity of thecore promoter, as well as the Sp1-binding site. Two nucleotidesin the core promoter region were mutated (underlined nucle-otides in �416Mut�6, Fig. 5A), resulting in the creation of anATG at the �6 position, in the context of a strong consensusKozak sequence, interposed between upstream start sites (i.e.,A�18 and A�14) and downstream start sites (i.e., A�1 andA�12) (Fig. 5A, �416Mut�6). Since this ATG is out of framewith respect to the downstream CAT reporter gene, any tran-scription starting upstream of �6 would not generate mRNAencoding CAT reporter enzyme in transient-transfection as-says. On the other hand, transcripts initiating downstream of�6 in the �416Mut�6 construct should have no effect onsubsequent translation of the CAT gene product. Thus, the�416Mut�6 distinguishes upstream from downstream tran-scription initiations.
Transient transfection of 416Mut�6 into HeLa epithelialcells resulted in greatly reduced levels of CAT reporter enzymeactivity relative to the �416WT construct, indicating that basaltranscription initiates primarily from the upstream start siteregion (Fig. 5A). This result is consistent with the observationthat the �14 start site is the preferred start site in basal in vitrotranscription in HeLa extracts (Fig. 1B). As a control, the A-6and T-5 residues were reversed (�416Con�6), thereby elimi-nating the out-of-frame ATG, and activity was fully restored(Fig. 5A). This control also demonstrates that the nucleotidechanges at these positions do not affect in vivo promoter func-tion. As an additional control, the class I core promoter, con-taining the ATG�6 mutation, was subcloned in frame with theluciferase reporter. Consistent with the interpretation thatbasal promoter activity initiates predominantly upstream of�6, the ATG�6 mutation, which is out of frame for CAT anddoes not generate CAT enzyme activity, does generate lucif-erase activity when present in-frame with the luciferase gene(Fig. 5B). The ATG�6 mutant does not affect class I corepromoter activity nonspecifically, since �416Mut�6 was as ac-tive as the WT class I promoter (�416WT) in in vitro tran-scription (Fig. 5C). Although some downstream initiation is
3384 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
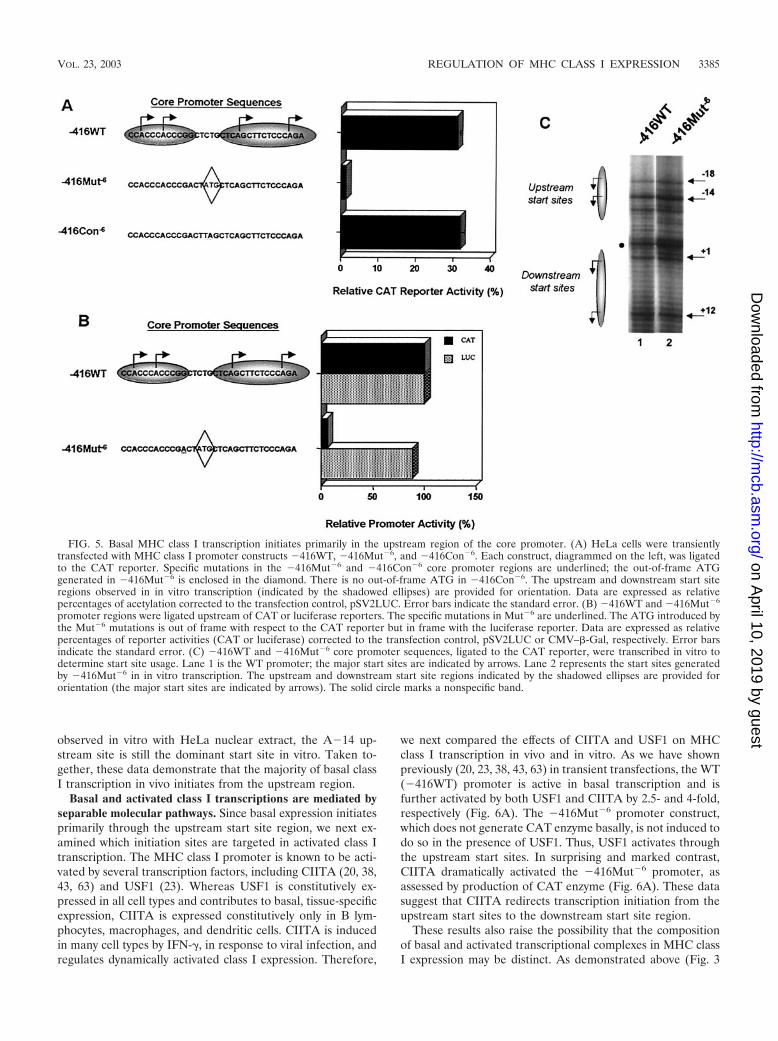
observed in vitro with HeLa nuclear extract, the A�14 up-stream site is still the dominant start site in vitro. Taken to-gether, these data demonstrate that the majority of basal classI transcription in vivo initiates from the upstream region.
Basal and activated class I transcriptions are mediated byseparable molecular pathways. Since basal expression initiatesprimarily through the upstream start site region, we next ex-amined which initiation sites are targeted in activated class Itranscription. The MHC class I promoter is known to be acti-vated by several transcription factors, including CIITA (20, 38,43, 63) and USF1 (23). Whereas USF1 is constitutively ex-pressed in all cell types and contributes to basal, tissue-specificexpression, CIITA is expressed constitutively only in B lym-phocytes, macrophages, and dendritic cells. CIITA is inducedin many cell types by IFN-�, in response to viral infection, andregulates dynamically activated class I expression. Therefore,
we next compared the effects of CIITA and USF1 on MHCclass I transcription in vivo and in vitro. As we have shownpreviously (20, 23, 38, 43, 63) in transient transfections, the WT(�416WT) promoter is active in basal transcription and isfurther activated by both USF1 and CIITA by 2.5- and 4-fold,respectively (Fig. 6A). The �416Mut�6 promoter construct,which does not generate CAT enzyme basally, is not induced todo so in the presence of USF1. Thus, USF1 activates throughthe upstream start sites. In surprising and marked contrast,CIITA dramatically activated the �416Mut�6 promoter, asassessed by production of CAT enzyme (Fig. 6A). These datasuggest that CIITA redirects transcription initiation from theupstream start sites to the downstream start site region.
These results also raise the possibility that the compositionof basal and activated transcriptional complexes in MHC classI expression may be distinct. As demonstrated above (Fig. 3
FIG. 5. Basal MHC class I transcription initiates primarily in the upstream region of the core promoter. (A) HeLa cells were transientlytransfected with MHC class I promoter constructs �416WT, �416Mut�6, and �416Con�6. Each construct, diagrammed on the left, was ligatedto the CAT reporter. Specific mutations in the �416Mut�6 and �416Con�6 core promoter regions are underlined; the out-of-frame ATGgenerated in �416Mut�6 is enclosed in the diamond. There is no out-of-frame ATG in �416Con�6. The upstream and downstream start siteregions observed in in vitro transcription (indicated by the shadowed ellipses) are provided for orientation. Data are expressed as relativepercentages of acetylation corrected to the transfection control, pSV2LUC. Error bars indicate the standard error. (B) �416WT and �416Mut�6
promoter regions were ligated upstream of CAT or luciferase reporters. The specific mutations in Mut�6 are underlined. The ATG introduced bythe Mut�6 mutations is out of frame with respect to the CAT reporter but in frame with the luciferase reporter. Data are expressed as relativepercentages of reporter activities (CAT or luciferase) corrected to the transfection control, pSV2LUC or CMV–�-Gal, respectively. Error barsindicate the standard error. (C) �416WT and �416Mut�6 core promoter sequences, ligated to the CAT reporter, were transcribed in vitro todetermine start site usage. Lane 1 is the WT promoter; the major start sites are indicated by arrows. Lane 2 represents the start sites generatedby �416Mut�6 in in vitro transcription. The upstream and downstream start site regions indicated by the shadowed ellipses are provided fororientation (the major start sites are indicated by arrows). The solid circle marks a nonspecific band.
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3385
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
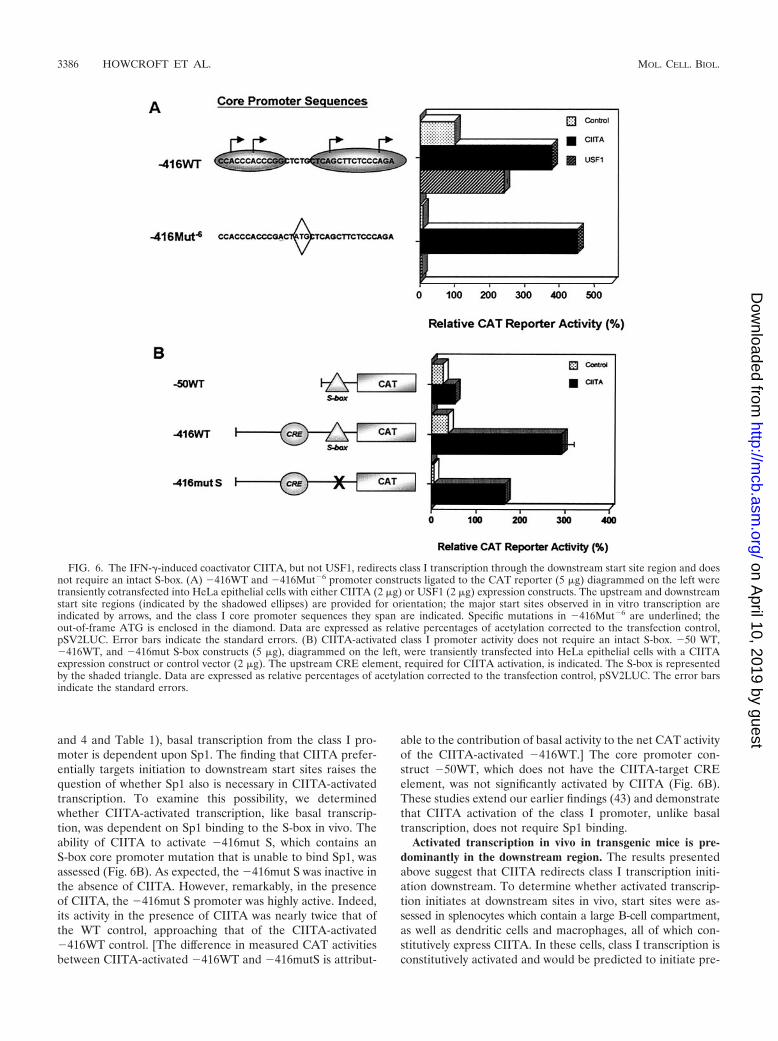
and 4 and Table 1), basal transcription from the class I pro-moter is dependent upon Sp1. The finding that CIITA prefer-entially targets initiation to downstream start sites raises thequestion of whether Sp1 also is necessary in CIITA-activatedtranscription. To examine this possibility, we determinedwhether CIITA-activated transcription, like basal transcrip-tion, was dependent on Sp1 binding to the S-box in vivo. Theability of CIITA to activate �416mut S, which contains anS-box core promoter mutation that is unable to bind Sp1, wasassessed (Fig. 6B). As expected, the �416mut S was inactive inthe absence of CIITA. However, remarkably, in the presenceof CIITA, the �416mut S promoter was highly active. Indeed,its activity in the presence of CIITA was nearly twice that ofthe WT control, approaching that of the CIITA-activated�416WT control. [The difference in measured CAT activitiesbetween CIITA-activated �416WT and �416mutS is attribut-
able to the contribution of basal activity to the net CAT activityof the CIITA-activated �416WT.] The core promoter con-struct �50WT, which does not have the CIITA-target CREelement, was not significantly activated by CIITA (Fig. 6B).These studies extend our earlier findings (43) and demonstratethat CIITA activation of the class I promoter, unlike basaltranscription, does not require Sp1 binding.
Activated transcription in vivo in transgenic mice is pre-dominantly in the downstream region. The results presentedabove suggest that CIITA redirects class I transcription initi-ation downstream. To determine whether activated transcrip-tion initiates at downstream sites in vivo, start sites were as-sessed in splenocytes which contain a large B-cell compartment,as well as dendritic cells and macrophages, all of which con-stitutively express CIITA. In these cells, class I transcription isconstitutively activated and would be predicted to initiate pre-
FIG. 6. The IFN-�-induced coactivator CIITA, but not USF1, redirects class I transcription through the downstream start site region and doesnot require an intact S-box. (A) �416WT and �416Mut�6 promoter constructs ligated to the CAT reporter (5 �g) diagrammed on the left weretransiently cotransfected into HeLa epithelial cells with either CIITA (2 �g) or USF1 (2 �g) expression constructs. The upstream and downstreamstart site regions (indicated by the shadowed ellipses) are provided for orientation; the major start sites observed in in vitro transcription areindicated by arrows, and the class I core promoter sequences they span are indicated. Specific mutations in �416Mut�6 are underlined; theout-of-frame ATG is enclosed in the diamond. Data are expressed as relative percentages of acetylation corrected to the transfection control,pSV2LUC. Error bars indicate the standard errors. (B) CIITA-activated class I promoter activity does not require an intact S-box. �50 WT,�416WT, and �416mut S-box constructs (5 �g), diagrammed on the left, were transiently transfected into HeLa epithelial cells with a CIITAexpression construct or control vector (2 �g). The upstream CRE element, required for CIITA activation, is indicated. The S-box is representedby the shaded triangle. Data are expressed as relative percentages of acetylation corrected to the transfection control, pSV2LUC. The error barsindicate the standard errors.
3386 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

dominantly at the downstream start site region. The MHCcontains multiple class I genes, all of which are highly homol-ogous, in both their coding and upstream sequences, making itdifficult to unambiguously define the start site of any one en-dogenous gene. To circumvent this problem, we examinedtranscription of a hybrid transgene containing 516 bp of thePD1 class I promoter sequence (positions �516 to �45, in-cluding the ATG of the first exon) upstream of the CATreporter gene (Fig. 7A). In previous studies, we demonstratedthat the in vivo pattern of expression of this hybrid transgeneparallels that of the endogenous murine H-2 class I genes (37).To identify transcript initiation sites, we performed 5�RACEanalysis with a CAT-specific primer that distinguished PD1promoter transgene transcripts from endogenous MHC class Ipromoter directed transcripts. The 5�RACE products weregenerated from splenic RNA and cloned. Critical to this anal-ysis, subsequent sequencing of appropriate clones allowed un-ambiguous identification of transcription initiation start site(s)generated by the class I transgene. As shown in Fig. 7A, mul-tiple specific start sites were detected, all of which were locatedin the downstream start site region of the class I core pro-moter. (It should be noted that the �2 and �4 start sites thatpredominate in vivo would not have been detected in vitro, dueto a strong nonspecific band present in the HeLa nuclearextract [see Fig. 1B]; the �5 and �7 start sites were observedoccasionally.) These data demonstrate that in a population ofcells where class I expression is activated, most transcriptioninitiates at the downstream sites.
We next determined whether IFN-� could actively alterMHC class I promoter initiation start site usage. Nuclear ex-tracts from either control or IFN-�-treated HeLa cells wereused to direct in vitro transcription of the bp �516 to �45promoter segment (the same one present in the transgene)(Fig. 7B). The overall pattern of start site selection paralleledthat observed in vivo and did not differ between the two nu-clear extracts. However, consistent with the in vivo pattern, the
FIG. 7. Mapping basal and activated MHC class I initiation sites invivo. (A) Results of 5�RACE analysis of �516CAT transgenic spleenRNA to map class I start sites utilized by splenocytes in vivo. Thetransgene, shown at the top of the figure, extends from �516 to �45(�516CAT) and is ligated to the CAT reporter gene. The translationinitiation codon of the PD1 class I gene is shown relative to the CATreporter gene; the location of the CAT gene-specific primer used in5�RACE is indicated by the left arrowhead. 5�RACE products derivedfrom �516CAT transgenic spleen RNA were generated and cloned.More than 300 clones were screened by hybridization; 16 containedclass I sequences. All 16 were sequenced to determine the precise
transcription initiation sites. Initiation sites, and their relative usageare indicated by the position and size of the right-hand arrows. Therelative frequency with which individual sites are used is also indicatedat the bottom of the figure and is aligned to the sequence above.Shaded ellipses representing the upstream and downstream transcrip-tion regions where the major start sites originate (indicated by arrows)is provided for orientation. The gray, broken arrows represent theupstream start sites that contribute to basal expression, in vitro and invivo, as determined by primer extension. (B) Nuclear extracts fromcontrol and IFN-�-treated HeLa cells were used in in vitro transcrip-tion reactions with a class I reporter construct �516CAT (�516 to�45). Primer extension of in vitro-transcribed RNA was used to iden-tify specific start sites. In vitro generated transcripts generated byIFN-�-treated HeLa nuclear extract are shown in lane 1; control HeLaextract is shown in lane 2. (C) Purified rCIITA or control lysate frombaculovirus-infected cells were added to in vitro transcription reactionsby using the class I promoter construct �416WT; specific start siteswere detected by using primer extension analysis. Class I-specific tran-script in the presence of HeLa extract without added baculovirus lysateis shown in the lane 1. Additions of control lysate and purified rCIITAare shown in lanes 2 and 3, respectively. The principal transcriptionstart sites, detected in Fig. 1B, are indicated by the arrows. Shadedellipses representing the upstream and downstream transcription re-gions where the major start sites originate (indicated by arrows) areprovided on the left for orientation. A sequence ladder of the class Ipromoter is also included.
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3387
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
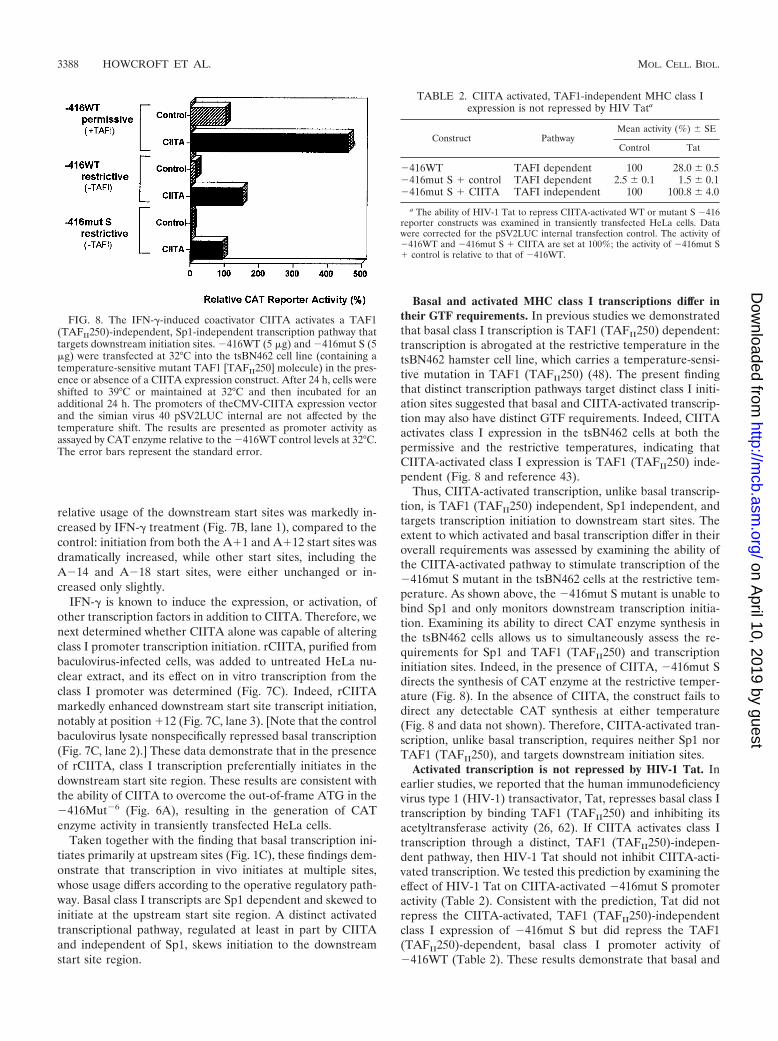
relative usage of the downstream start sites was markedly in-creased by IFN-� treatment (Fig. 7B, lane 1), compared to thecontrol: initiation from both the A�1 and A�12 start sites wasdramatically increased, while other start sites, including theA�14 and A�18 start sites, were either unchanged or in-creased only slightly.
IFN-� is known to induce the expression, or activation, ofother transcription factors in addition to CIITA. Therefore, wenext determined whether CIITA alone was capable of alteringclass I promoter transcription initiation. rCIITA, purified frombaculovirus-infected cells, was added to untreated HeLa nu-clear extract, and its effect on in vitro transcription from theclass I promoter was determined (Fig. 7C). Indeed, rCIITAmarkedly enhanced downstream start site transcript initiation,notably at position �12 (Fig. 7C, lane 3). [Note that the controlbaculovirus lysate nonspecifically repressed basal transcription(Fig. 7C, lane 2).] These data demonstrate that in the presenceof rCIITA, class I transcription preferentially initiates in thedownstream start site region. These results are consistent withthe ability of CIITA to overcome the out-of-frame ATG in the�416Mut�6 (Fig. 6A), resulting in the generation of CATenzyme activity in transiently transfected HeLa cells.
Taken together with the finding that basal transcription ini-tiates primarily at upstream sites (Fig. 1C), these findings dem-onstrate that transcription in vivo initiates at multiple sites,whose usage differs according to the operative regulatory path-way. Basal class I transcripts are Sp1 dependent and skewed toinitiate at the upstream start site region. A distinct activatedtranscriptional pathway, regulated at least in part by CIITAand independent of Sp1, skews initiation to the downstreamstart site region.
Basal and activated MHC class I transcriptions differ intheir GTF requirements. In previous studies we demonstratedthat basal class I transcription is TAF1 (TAFII250) dependent:transcription is abrogated at the restrictive temperature in thetsBN462 hamster cell line, which carries a temperature-sensi-tive mutation in TAF1 (TAFII250) (48). The present findingthat distinct transcription pathways target distinct class I initi-ation sites suggested that basal and CIITA-activated transcrip-tion may also have distinct GTF requirements. Indeed, CIITAactivates class I expression in the tsBN462 cells at both thepermissive and the restrictive temperatures, indicating thatCIITA-activated class I expression is TAF1 (TAFII250) inde-pendent (Fig. 8 and reference 43).
Thus, CIITA-activated transcription, unlike basal transcrip-tion, is TAF1 (TAFII250) independent, Sp1 independent, andtargets transcription initiation to downstream start sites. Theextent to which activated and basal transcription differ in theiroverall requirements was assessed by examining the ability ofthe CIITA-activated pathway to stimulate transcription of the�416mut S mutant in the tsBN462 cells at the restrictive tem-perature. As shown above, the �416mut S mutant is unable tobind Sp1 and only monitors downstream transcription initia-tion. Examining its ability to direct CAT enzyme synthesis inthe tsBN462 cells allows us to simultaneously assess the re-quirements for Sp1 and TAF1 (TAFII250) and transcriptioninitiation sites. Indeed, in the presence of CIITA, �416mut Sdirects the synthesis of CAT enzyme at the restrictive temper-ature (Fig. 8). In the absence of CIITA, the construct fails todirect any detectable CAT synthesis at either temperature(Fig. 8 and data not shown). Therefore, CIITA-activated tran-scription, unlike basal transcription, requires neither Sp1 norTAF1 (TAFII250), and targets downstream initiation sites.
Activated transcription is not repressed by HIV-1 Tat. Inearlier studies, we reported that the human immunodeficiencyvirus type 1 (HIV-1) transactivator, Tat, represses basal class Itranscription by binding TAF1 (TAFII250) and inhibiting itsacetyltransferase activity (26, 62). If CIITA activates class Itranscription through a distinct, TAF1 (TAFII250)-indepen-dent pathway, then HIV-1 Tat should not inhibit CIITA-acti-vated transcription. We tested this prediction by examining theeffect of HIV-1 Tat on CIITA-activated �416mut S promoteractivity (Table 2). Consistent with the prediction, Tat did notrepress the CIITA-activated, TAF1 (TAFII250)-independentclass I expression of �416mut S but did repress the TAF1(TAFII250)-dependent, basal class I promoter activity of�416WT (Table 2). These results demonstrate that basal and
FIG. 8. The IFN-�-induced coactivator CIITA activates a TAF1(TAFII250)-independent, Sp1-independent transcription pathway thattargets downstream initiation sites. �416WT (5 �g) and �416mut S (5�g) were transfected at 32°C into the tsBN462 cell line (containing atemperature-sensitive mutant TAF1 [TAFII250] molecule) in the pres-ence or absence of a CIITA expression construct. After 24 h, cells wereshifted to 39°C or maintained at 32°C and then incubated for anadditional 24 h. The promoters of theCMV-CIITA expression vectorand the simian virus 40 pSV2LUC internal are not affected by thetemperature shift. The results are presented as promoter activity asassayed by CAT enzyme relative to the �416WT control levels at 32°C.The error bars represent the standard error.
TABLE 2. CIITA activated, TAF1-independent MHC class Iexpression is not repressed by HIV Tata
Construct PathwayMean activity (%) SE
Control Tat
�416WT TAFI dependent 100 28.0 0.5�416mut S � control TAFI dependent 2.5 0.1 1.5 0.1�416mut S � CIITA TAFI independent 100 100.8 4.0
a The ability of HIV-1 Tat to repress CIITA-activated WT or mutant S �416reporter constructs was examined in transiently transfected HeLa cells. Datawere corrected for the pSV2LUC internal transfection control. The activity of�416WT and �416mut S � CIITA are set at 100%; the activity of �416mut S� control is relative to that of �416WT.
3388 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

activated transcription represent distinct regulatory pathwaysthat utilize distinct promoter elements, depend on distincttranscription factors, and assemble distinct preinitiation com-plexes.
DISCUSSION
The present studies demonstrate that activated and basaltranscription target distinct sites within the class I core pro-moter and have distinct activator and GTF requirements.Basal transcription initiates primarily at upstream sites withinthe core promoter, requires the presence of an intact Sp1-binding element and is TAF1 (TAFII250) dependent. Inmarked contrast, activated transcription, as defined by theIFN-�-induced coactivator CIITA, initiates transcription pri-marily at downstream sites, is unaffected by mutations thatdisable Sp1 binding, and is TAF1 (TAFII250) independent.These studies demonstrate that basal and activated modes ofMHC class I transcription are regulated by distinct pathwaysthat converge on a single core promoter.
Consistent with a role in integrating multiple transcriptionalpathways, the MHC class I core promoter is complex, both inits organization and function. The core promoter is encom-passed within a 65-bp DNA fragment (bp �50 to �14): thissegment is sufficient to mediate basal transcription and is nec-essary for activated transcription. Transcription initiation oc-curs at multiple but largely nonoverlapping sites in both basaland activated transcription. Although sequences homologousto canonical TATA (bp �30 to �25) and Inr elements (bp �3to �5) occur in this segment and contribute to overall pro-moter activity, neither of these is necessary for either transcrip-tional pathway. Two Inr-like sequences also occur within thecentral S-box, overlapping both sites of initiation of basal tran-scription and Sp1-binding sites. These sequences may functionas Inr elements in basal transcription but are not necessary foractivated transcription. Thus, no single element within the corepromoter is necessary for all transcription.
The promoter of the MHC class I gene, PD1, occurs withina CpG island (40). The interval between bp �300 and �300contains 41 CpG and 53 GpC motifs. Such CpG islands aregenerally undermethylated and contain promoters for a varietyof genes, commonly housekeeping genes. The GpC motifs of-ten serve as binding sites for Sp1, which may serve both torecruit general transcription factors and to maintain the hy-pomethylated state. Promoters within CpG islands often donot have canonical TATA or Inr sequences and initiate atmultiple sites. Consistent with the suggestion that the class Ipromoter is a CpG island promoter is the lack of canonicalpromoter elements. The TATA-like and Inr-like elements arenot necessary for transcription, and no TBP binding has beenobserved to the TATA-like element (K. Howcroft and D.Singer, unpublished observations). No sequences with homol-ogy to either DPE or DCE elements are found within the corepromoter. A classical TFIIB-binding site, BRE, occurs at bp�51 to �46, but there is no evidence that it functions as aregulatory element; neither TFIIB nor the TATA-binding pro-tein have been observed to bind the BRE, either alone or incombination (Z. Sercan and D. S. Singer, unpublished data).Further, factors that have been reported to bind to the Inr,such as TFII I, USF (46, 47), and YY1 (34, 58) do not bind to
the core promoter (T. K. Howcroft, J. D. Weissman, and D. S.Singer, unpublished results). Thus, the class I core promoterstructurally resembles other CpG island promoters.
Through its complex structure, the class I core promoterdynamically integrates disparate regulatory signals. As demon-strated in the present study, the multiplicity of transcriptionstart sites within the class I core promoter reflects differentialstart site usage for activated and basal transcription. Basaltranscription is focused on initiation sites upstream of bp �6,primarily at bp �14 and �18, within the S-box. In contrast,activated transcription initiates downstream of �6 bp, primar-ily between bp �4 and �12. This differential usage correlateswith promoter element specificity, activator factor require-ments, and general transcription factor dependence. Thus,basal transcription depends on the S-box element, on the Sp1activator binding to the S-box and on the general transcriptionfactor, TAF1. Activated transcription requires none of these.Such core promoter element specificity, linked with differentialtranscription factor usage, has been termed “combinatorialregulation” and has been reported in a number of systems (55).In Drosophila, it has been demonstrated that enhancers arespecific either for promoters with TATA or with DPE ele-ments (8). Similarly, in the mouse TdT gene, promoter activitydepends on the interaction between enhancer elements andthe native Inr, which cannot be replaced by a TATA element.However, artificial introduction of both Sp1 and TATA ele-ments into the TdT promoter restores activity (12). Thus, thereis regulation through appropriate enhancer/promoter elementcombinations. However, in these examples, regulation is static:a single promoter uses one enhancer-promoter element com-bination.
In marked contrast, the class I promoter has at least twodistinct enhancer-promoter element combinations and displaysdynamic combinatorial regulation under different physiologicalconditions. Basal and activated transcription invoke nonover-lapping combinations within the same core promoter. Only theyeast HIS3 promoter is known to have a similar dynamic com-binatorial regulation. This promoter has two TATAA ele-ments: a downstream canonical one and an upstream variant.At low levels of transcription, the variant element is usedpreferentially, whereas at high levels of transcription, the ca-nonical TATA is used (28). In contrast to the class I promoter,the differential usage of HIS3 TATA elements does not reflectdifferential activator protein function.
Differential start site usage in the class I core promoterdepends on both activator and general transcription factorfunction. Basal transcription is regulated by tissue-specific butconstitutively expressed activators, such as Sp1 and USF. ClassI basal transcription in all cell types depends on Sp1 binding tothe S-box, and in particular to the central 10-bp CA/GT-richsequence, where basal transcription initiates. We speculatethat Sp1 focuses basal transcription to this upstream region byrecruiting the transcription machinery, maintaining an under-methylated state around the promoter, or both. Sp1 is knownto recruit RNA Pol II through its interaction with the TAF4(TAFII110) subunit of TFIID (61). The constitutively ex-pressed USF also activates through the basal pathway. In ad-dition to the upstream E-box element, USF-enhanced tran-scription requires an intact S-box (data not shown), requiresTAF1 (63), and targets the upstream start sites. We suggest
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3389
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

that the class I S-box, through recruitment of the appropriatepreinitiation complex (PIC), integrates the constitutive tissue-specific signals of the basal transcription pathway.
Although the S-box is the major core promoter sequencerequired for basal transcription, it is not involved in activatedtranscription. CIITA-activated transcription is unaffected bymutation of the S-box. Based on our findings, we propose thathormone- or cytokine-regulated activators, such as CIITA,function through an alternative transcription pathway withpromoter element requirements distinct from those of basaltranscription. We further suggest that activated transcriptionpathways recruit novel PIC(s) that have components distinctfrom those assembled under basal conditions, which in turn,differentially target the various core promoter elements.
What is the significance of having distinct transcriptionalpathways to express MHC class I genes? We speculate that theCIITA-activated transcription pathway is an adaptive strategyfor continued MHC class I expression under pathologicallyadverse conditions. For example, many viruses can activelyrepress MHC class I basal transcription in order to preventdetection and destruction by MHC class I-restricted cytotoxicT lymphocytes (4, 22, 39). Previous studies from our lab havedemonstrated that HIV-1 Tat specifically interacts with theTAF1 (TAFII250) subunit of TFIID and inhibits its acetyl-transferase activity (62). Since MHC class I expression is TAF1(TAFII250) dependent, this process results in repression ofbasal transcription and reduced cell surface expression ofMHC class I (24, 26). In the present study we demonstrate thatCIITA, which has intrinsic acetyltransferase activity (43), acti-vates an alternate TAF1 (TAFII250)-independent pathway ofMHC class I expression that is resistant to Tat repression.Thus, the TAF1 (TAFII250)-independent pathway activated byCIITA bypasses the Tat repressed basal pathway and promotesclass I expression in HIV-1-infected cells.
In conclusion, the present studies have defined two distincttranscriptional pathways that regulate MHC class I gene ex-pression. We propose that proper class I expression is achievedby the appropriate integration of these two pathways. We spec-ulate that this system has the further selective advantage ofensuring continued immune surveillance in the face of intra-cellular pathogens.
ACKNOWLEDGMENTS
We gratefully acknowledge Joshua Meyers and Stacey McLaughlinfor their technical assistance and John Brady, David Levens, andDanny Reinberg for helpful discussion and critical review of the manu-script.
REFERENCES
1. Aso, T., J. W. Conaway, and R. C. Conaway. 1994. Role of core promoterstructure in assembly of the RNA polymerase II ormattedpreinitiation com-plex: a common pathway for formation of preinitiation intermediates atmany TATA and TATA-less promoters. J. Biol. Chem. 269:26575–26583.
2. Baldwin, A. S., Jr., and P. A. Sharp. 1987. Binding of a nuclear factor to aregulatory sequence in the promoter of the mouse H-2Kb class I majorhistocompatibility gene. Mol. Cell. Biol. 7:305–313.
3. Berk, A. J. 1999. Activation of RNA polymerase II transcription. Curr. Opin.Cell Biol. 11:330–335.
4. Brodsky, F. M. 1999. Stealth, sabotage, and exploitation. Immunol. Rev.168:5–11.
5. Burke, T. W., and J. T. Kadonaga. 1997. The downstream core promoterelement, DPE, is conserved from Drosophila to humans and is recognized byTAFII60 of Drosophila. Genes Dev. 11:3020–3031.
6. Burke, T. W., and J. T. Kadonaga. 1996. Drosophila TFIID binds to a
conserved downstream basal promoter element that is present in manyTATA-box-deficient promoters. Genes Dev. 10:711–724.
7. Burley, S. K. 1996. The TATA box binding protein. Curr. Opin. Struct. Biol.6:69–75.
8. Butler, J. E., and J. T. Kadonaga. 2001. Enhancer-promoter specificity me-diated by DPE or TATA core promoter motifs. Genes Dev. 15:2515–2519.
9. Butler, J. E., and J. T. Kadonaga. 2002. The RNA polymerase II corepromoter: a key component in the regulation of gene expression. Genes Dev.16:2583–2592.
10. Chen, L. I., T. Nishinaka, K. Kwan, I. Kitabayashi, K. Yokoyama, Y. H. Fu,S. Grunwald, and R. Chiu. 1994. The retinoblastoma gene product RBstimulates Sp1-mediated transcription by liberating Sp1 from a negativeregulator. Mol. Cell. Biol. 14:4380–4389.
11. David-Watine, B., A. Israel, and P. Kourilsky. 1990. The regulation andexpression of MHC class I genes. Immunol. Today 11:286–292.
12. Emami, K. H., W. W. Navarre, and S. T. Smale. 1995. Core promoterspecificities of the Sp1 and VP16 transcriptional activation domains. Mol.Cell. Biol. 15:5906–5916.
13. Fischer, K. D., A. Haese, and J. Nowock. 1993. Cooperation of GATA-1 andSp1 can result in synergistic transcriptional activation or interference. J. Biol.Chem. 268:23915–23923.
14. Frels, W. I., J. A. Bluestone, R. J. Hodes, M. R. Capecchi, and D. S. Singer.1985. Expression of a microinjected porcine class I major histocompatibilitycomplex gene in transgenic mice. Science 228:577–580.
15. Gardiner-Garden, M., and M. Frommer. 1987. CpG islands in vertebrategenomes. J. Mol. Biol. 196:261–282.
16. Gill, G. 2001. Regulation of the initiation of eukaryotic transcription. EssaysBiochem. 37:33–43.
17. Girdlestone, J. 1995. Regulation of HLA class I loci by interferons. Immu-nobiology 193:229–237.
18. Girdlestone, J. 1996. Transcriptional regulation of MHC class I genes. Eur.J. Immunogenet. 23:395–413.
19. Gobin, S. J., V. Keijsers, M. van Zutphen, and P. J. van den Elsen. 1998. Therole of enhancer A in the locus-specific transactivation of classical andnonclassical HLA class I genes by nuclear factor B. J. Immunol. 161:2276–2283.
20. Gobin, S. J., A. Peijnenburg, V. Keijsers, and P. J. van den Elsen. 1997. Sitealpha is crucial for two routes of IFN gamma-induced MHC class I trans-activation: the ISRE-mediated route and a novel pathway involving CIITA.Immunity 6:601–611.
21. Hampsey, M. 1998. Molecular genetics of the RNA polymerase II generaltranscriptional machinery. Microbiol. Mol. Biol. Rev. 62:465–503.
22. Howcroft, T. K. 2000. HIV-1: a molecular toolkit for analysis of majorhistocompatibility complex class I expression. Eur. J. Clin. Investig. 30:657–659.
23. Howcroft, T. K., C. Murphy, J. D. Weissman, S. J. Huber, M. Sawadogo, andD. S. Singer. 1999. Upstream stimulatory factor regulates major histocom-patibility complex class I gene expression: the U2�E4 splice variant abro-gates E-box activity. Mol. Cell. Biol. 19:4788–4797.
24. Howcroft, T. K., L. A. Palmer, J. Brown, B. Rellahan, F. Kashanchi, J. N.Brady, and D. S. Singer. 1995. HIV Tat represses transcription throughSp1-like elements in the basal promoter. Immunity 3:127–138.
25. Howcroft, T. K., J. C. Richardson, and D. S. Singer. 1993. MHC class I geneexpression is negatively regulated by the proto-oncogene, c-jun. EMBO J.12:3163–3169.
26. Howcroft, T. K., K. Strebel, M. A. Martin, and D. S. Singer. 1993. Repressionof MHC class I gene promoter activity by two-exon Tat of HIV. Science260:1320–1322.
27. Ishiguro, N., G. D. Brown, and D. Meruelo. 1997. Activation transcriptionfactor 1 involvement in the regulation of murine H-2Dd expression. J. Biol.Chem. 272:15993–16001.
28. Iyer, V., and K. Struhl. 1995. Mechanism of differential utilization of the his3TR and TC TATA elements. Mol. Cell. Biol. 15:7059–7066.
29. Kaufmann, J., K. Ahrens, R. Koop, S. T. Smale, and R. Muller. 1998.CIF150, a human cofactor for transcription factor IID-dependent initiatorfunction. Mol. Cell. Biol. 18:233–239.
30. Kaufmann, J., and S. T. Smale. 1994. Direct recognition of initiator elementsby a component of the transcription factor IID complex. Genes Dev. 8:821–829.
31. Kutach, A. K., and J. T. Kadonaga. 2000. The downstream promoter ele-ment DPE appears to be as widely used as the TATA box in Drosophila corepromoters. Mol. Cell. Biol. 20:4754–4764.
32. Lagrange, T., A. N. Kapanidis, H. Tang, D. Reinberg, and R. H. Ebright.1998. New core promoter element in RNA polymerase II-dependent tran-scription: sequence-specific DNA binding by transcription factor IIB. GenesDev. 12:34–44.
33. Le Bouteiller, P. 1994. HLA class I chromosomal region, genes, and prod-ucts: facts and questions. Crit. Rev. Immunol. 14:89–129.
34. Lee, J. S., K. M. Galvin, and Y. Shi. 1993. Evidence for physical interactionbetween the zinc-finger transcription factors YY1 and Sp1. Proc. Natl. Acad.Sci. USA 90:6145–6149.
35. Liang, Y., D. F. Robinson, G. C. Kujoth, and W. E. Fahl. 1996. Functional
3390 HOWCROFT ET AL. MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

analysis of the SIS proximal element and its activating factors: regulatedtranscription of the c-SIS/PDGF-B gene in human erythroleukemia cells.Oncogene 13:863–871.
36. Lo, K., and S. T. Smale. 1996. Generality of a functional initiator consensussequence. Gene 182:13–22.
37. Maguire, J. E., W. I. Frels, J. C. Richardson, J. D. Weissman, and D. S.Singer. 1992. In vivo function of regulatory DNA sequence elements of amajor histocompatibility complex class I gene. Mol. Cell. Biol. 12:3078–3086.
38. Martin, B. K., K. C. Chin, J. C. Olsen, C. A. Skinner, A. Dey, K. Ozato, andJ. P. Ting. 1997. Induction of MHC class I expression by the MHC class IItransactivator CIITA. Immunity 6:591–600.
39. Maudsley, D. J., and J. D. Pound. 1991. Modulation of MHC antigen ex-pression by viruses and oncogenes. Immunol. Today 12:429–431.
40. McQueen, H. A., V. H. Clark, A. P. Bird, M. Yerle, and A. L. Archibald. 1997.CpG islands of the pig. Genome Res. 7:924–931.
41. Murphy, C., D. Nikodem, K. Howcroft, J. D. Weissman, and D. S. Singer.1996. Active repression of major histocompatibility complex class I genes ina human neuroblastoma cell line. J. Biol. Chem. 271:30992–30999.
42. Nagarajan, U. M., A. Peijnenburg, S. J. Gobin, J. M. Boss, and P. J. van denelsen. 2000. Novel mutations within the RFX-B gene and partial rescue of,M. H. C., and related genes through exogenous class II transactivator inRFX-B-deficient cells. J. Immunol. 164:3666–3674.
43. Raval, A., T. K. Howcroft, J. D. Weissman, S. Kirshner, X. S. Zhu, K.Yokoyama, J. Ting, and D. S. Singer. 2001. Transcriptional coactivator,CIITA, is an acetyltransferase that bypasses a promoter requirement forTAFII250. Mol. Cell 7:105–115.
44. Reinberg, D., G. Orphanides, R. Ebright, S. Akoulitchev, J. Carcamo, H.Cho, P. Cortes, R. Drapkin, O. Flores, I. Ha, J. A. Inostroza, S. Kim, T. K.Kim, P. Kumar, T. Lagrange, G. LeRoy, H. Lu, D. M. Ma, E. Maldonado, A.Merino, F. Mermelstein, I. Olave, M. Sheldon, R. Shiekhattar, L. Zawel, etal. 1998. The RNA polymerase II general transcription factors: past, present,and future. Cold Spring Harbor Symp. Quant. Biol. 63:83–103.
45. Roeder, R. G. 1996. The role of general initiation factors in transcription byRNA polymerase II. Trends Biochem. Sci. 21:327–335.
46. Roy, A. L., S. Malik, M. Meisterernst, and R. G. Roeder. 1993. An alternativepathway for transcription initiation involving TFII-I. Nature 365:355–359.
47. Roy, A. L., M. Meisterernst, P. Pognonec, and R. G. Roeder. 1991. Cooper-ative interaction of an initiator-binding transcription initiation factor and thehelix-loop-helix activator USF. Nature 354:245–248.
48. Sekiguchi, T., Y. Nohiro, Y. Nakamura, N. Hisamoto, and T. Nishimoto.1991. The human CCG1 gene, essential for progression of the G1 phase,encodes a 210-kilodalton nuclear DNA-binding protein. Mol. Cell. Biol.11:3317–3325.
49. Singer, D. S., R. D. Camerini-Otero, M. L. Satz, B. Osborne, D. Sachs, andS. Rudikoff. 1982. Characterization of a porcine genomic clone encoding amajor histocompatibility antigen: expression in mouse L cells. Proc. Natl.Acad. Sci. USA 79:1403–1407.
50. Singer, D. S., and R. Ehrlich. 1988. Identification of regulatory elementsassociated with a class I MHC gene. Curr. Top. Microbiol. Immunol. 137:148–154.
51. Singer, D. S., and J. E. Maguire. 1990. Regulation of the expression of classI MHC genes. Crit. Rev. Immunol. 10:235–257.
52. Singer, D. S., E. Mozes, S. Kirshner, and L. D. Kohn. 1997. Role of MHCclass I molecules in autoimmune disease. Crit. Rev. Immunol. 17:463–468.
53. Singer, V. L., C. R. Wobbe, and K. Struhl. 1990. A wide variety of DNAsequences can functionally replace a yeast TATA element for transcriptionalactivation. Genes Dev. 4:636–645.
54. Sirito, M., Q. Lin, J. M. Deng, R. R. Behringer, and M. Sawadogo. 1998.Overlapping roles and asymmetrical cross-regulation of the USF proteins inmice. Proc. Natl. Acad. Sci. USA 95:3758–3763.
55. Smale, S. T. 2001. Core promoters: active contributors to combinatorial generegulation. Genes Dev. 15:2503–2508.
56. Smale, S. T., and D. Baltimore. 1989. The “initiator” as a transcriptioncontrol element. Cell 57:103–113.
57. Smale, S. T., A. Jain, J. Kaufmann, K. H. Emami, K. Lo, and I. P. Garraway.1998. The initiator element: a paradigm for core promoter heterogeneitywithin metazoan protein-coding genes. Cold Spring Harbor Symp. Quant.Biol. 63:21–31.
58. Usheva, A., and T. Shenk. 1996. YY1 transcriptional initiator: protein inter-actions and association with a DNA site containing unpaired strands. Proc.Natl. Acad. Sci. USA 93:13571–13576.
59. van den Elsen, P. J., and S. J. Gobin. 1999. The common regulatory pathwayof MHC class I and class II transactivation. Microbes Infect. 1:887–892.
60. Verrijzer, C. P., and R. Tjian. 1996. TAFs mediate transcriptional activationand promoter selectivity. Trends Biochem. Sci. 21:338–342.
61. Weinzierl, R. O., B. D. Dynlacht, and R. Tjian. 1993. Largest subunit ofDrosophila transcription factor IID directs assembly of a complex containingTBP and a coactivator. Nature 362:511–517.
62. Weissman, J. D., J. A. Brown, T. K. Howcroft, J. Hwang, A. Chawla, P. A.Roche, L. Schiltz, Y. Nakatani, and D. S. Singer. 1998. HIV-1 tat bindsTAFII250 and represses TAFII250-dependent transcription of major histo-compatibility class I genes. Proc. Natl. Acad. Sci. USA 95:11601–11606.
63. Weissman, J. D., T. K. Howcroft, and D. S. Singer. 2000. TAFII250-inde-pendent transcription can be conferred on a TAFII250-dependent basalpromoter by upstream activators. J. Biol. Chem. 275:10160–10167.
64. Weissman, J. D., and D. S. Singer. 1991. A complex regulatory DNA elementassociated with a major histocompatibility complex class I gene consists ofboth a silencer and an enhancer. Mol. Cell. Biol. 11:4217–4227.
65. Zenzie-Gregory, B., A. Khachi, I. P. Garraway, and S. T. Smale. 1993.Mechanism of initiator-mediated transcription: evidence for a functionalinteraction between the TATA-binding protein and DNA in the absence ofa specific recognition sequence. Mol. Cell. Biol. 13:3841–3849.
VOL. 23, 2003 REGULATION OF MHC CLASS I EXPRESSION 3391
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from