dissemination of invasive salmonella via - pnas.org · dissemination of invasive salmonella via...
TRANSCRIPT

Dissemination of invasive Salmonella viabacterial-induced extrusion of mucosal epitheliaLeigh A. Knodlera,1, Bruce A. Vallanceb, Jean Cellia, Seth Winfreea, Bryan Hansenc, Marinieve Monterob,and Olivia Steele-Mortimera
aLaboratory of Intracellular Parasites and cResearch Technologies Branch, Microscopy Unit, Rocky Mountain Laboratories, National Institute of Allergy andInfectious Diseases, National Institutes of Health, Hamilton, MT 59840; and bDivision of Gastroenterology, Department of Paediatrics, University of BritishColumbia, British Columbia Children’s Hospital, Vancouver, BC, Canada V6H 3V4
Edited by Pascale Cossart, Institut Pasteur, Paris, France, and approved August 24, 2010 (received for review May 5, 2010)
Salmonella enterica is an intracellular bacterial pathogen that residesand proliferateswithin amembrane-bound vacuole in epithelial cellsof the gut and gallbladder. Although essential to disease, how Sal-monella escapes from its intracellular niche and spreads to secondarycells within the same host, or to a new host, is not known. Here, wedemonstrate that a subpopulation of Salmonella hyperreplicating inthe cytosol of epithelial cells serves as a reservoir for dissemination.These bacteria are transcriptionally distinct from intravacuolar Sal-monella. They are induced for the invasion-associated type III secre-tion system and possessflagella; hence, they are primed for invasion.Epithelial cells ladenwith these cytosolic bacteria are extruded out ofthe monolayer, releasing invasion-primed and -competent Salmo-nella into the lumen. This extrusion mechanism is morphologicallysimilar to the process of cell shedding required for turnover of theintestinal epithelium. In contrast to the homeostatic mechanism,however, bacterial-induced extrusion is accompanied by an inflam-matory cell death characterizedby caspase-1 activationand the apicalreleaseof IL-18, an important cytokine regulatorofgut inflammation.Although epithelial extrusion is obviously beneficial to Salmonellafor completion of its life cycle, it also provides amechanistic explana-tion for themucosal inflammation that is triggeredduringSalmonellainfection of the gastrointestinal and biliary tracts.
caspase-1 | epithelial cells | flagella | IL-18 | type III secretion
Intracellular pathogens reside either within a membrane-boundvacuole or freely within the host cell cytosol. However, regardless
of their specific lifestyle, three distinct steps are common to theirinfectious cycle: the ability to enter host cells, their establishment ofan intracellular niche, and their escape from the host cell (1). Entryand intracellular survival are critical virulence stages for thesepathogens, but exit from the infected cell is essential for dissemi-nation and transmission to other hosts. Although considerableprogress has been made in elucidating the first two facets of thiscycle, themechanism bywhich intracellular pathogens escape fromhost cells has been comparably neglected.The Gram-negative bacterium Salmonella enterica causes a wide
range of food- and water-borne diseases ranging from self-limitinggastroenteritis to systemic typhoid fever in both humans and ani-mals. In enteric infections, Salmonella preferentially targets thesingle layer of polarized columnar epithelial cells lining the surfaceof the gastrointestinal tract (2–4), triggering an extensive in-flammatory response. After invading epithelial cells from the apicalside, Salmonella resides and replicates within a membrane-boundvacuole, known as the Salmonella-containing vacuole (SCV).Symptomatic and asymptomatic infections are characterized bythe fecal shedding of bacteria (5, 6), suggesting that Salmonellaescapes from its intracellular niche back into the gut lumen as partof its infectious cycle. Here, we report that Salmonella exits frompolarized epithelia by coopting a mechanism normally used by thehost to remove senescent cells from the mucosal epithelium.
Results and DiscussionWT Salmonella Hyperreplicates in the Cytosol of Epithelial Cells.Human colonic epithelial cells (C2BBe1, a subclone of Caco-2)
grown on filters were used as a model polarized monolayer to ex-amine the infectious cycle of Salmonella. Confocal microscopyanalysis of infectedmonolayers revealed two distinct populations ofproliferating bacteria following the onset of replication ≥4 h post-infection (p.i.) (Fig. 1A and Fig. S1A). Compared with an averagedoubling time of ≥95 min for the total population (Fig. S1A), somebacteria were replicating at a much faster rate, with a doubling timeof ∼20 min (Movie S1). There was a temporal increase in the in-cidence of these “hyperreplicating” Salmonella (defined as >50bacteria per cell) (Fig. 1A andB). By 10 h p.i., 11± 4.2%of infectedcells contained hyperreplicating bacteria (Fig. 1A). A similar phe-notype was previously described for a Salmonella sifA mutant,which hyperreplicates in the host cell cytosol because of a defect inmaintaining vacuolar integrity (7). We therefore assessed whetherthe hyperreplicating WT Salmonella we observed in polarized epi-thelial cells are also free in the cytosol. Confocal microscopy in-dicated that many of these bacteria were not in a lysosome-associated membrane protein 1 (LAMP1)-positive compartment(Fig. 1B and Fig. S1B), and thus not in a mature SCV (8). Selectivemembrane permeabilization followed by immunostaining withpolyclonal anti-Salmonella LPS antibody revealed that at least one-third of the hyperreplicating bacteria are cytosolic (Fig. 1C and Fig.S2). These experiments demonstrate that WT Salmonella can rep-licate in a vacuole and the cytosol in epithelial cells, but they pro-liferate more efficiently in the cytosolic environment (7, 9, 10).
Cytosolic Salmonella Are Invasion-Primed.We hypothesized that thetwo distinct intracellular environments, intravacuolar and cyto-solic, would differentially influence the expression of bacterialvirulence genes. To assess this, we used a plasmid-derived tran-scriptional fusion assay based on destabilized GFP(LVA). Pro-moters were selected from well-characterized genes in each of thethree type III secretion systems (T3SSs): PfliC-gfp[LVA] (flagel-lar T3SS), PprgH-gfp[LVA] (T3SS1), and PssaG-gfp[LVA](T3SS2) (11). The number of fluorescent bacteria was monitoredwith time. Under the infection conditions used here, efficientinvasion requires both T3SS1 and flagellar-based motility (11–13)(Fig. S1A). This single-cell assay confirmed that both T3SS1(PprgH-gfp[LVA]) and flagella (PfliC-gfp[LVA]) were rapidlydown-regulated after bacterial internalization (11, 14, 15) (Fig.S1C). Surprisingly, these virulence factors were not completelyinactivated (Fig. 2 and Fig. S1C). At 10 h p.i., ∼6% of infectedcells contained fluorescent PprgH-GFP[LVA] bacteria (3.8 ±1.2% of the total bacterial population) (Fig. 2 and S1C). Strik-
Author contributions: L.A.K., B.A.V., J.C., S.W., andO.S.-M. designed research; L.A.K., B.A.V.,J.C., S.W., B.H., and M.M. performed research; L.A.K., B.A.V., J.C., and S.W. analyzed data;and L.A.K. and O.S.-M. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1006098107/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1006098107 PNAS | October 12, 2010 | vol. 107 | no. 41 | 17733–17738
MICRO
BIOLO
GY
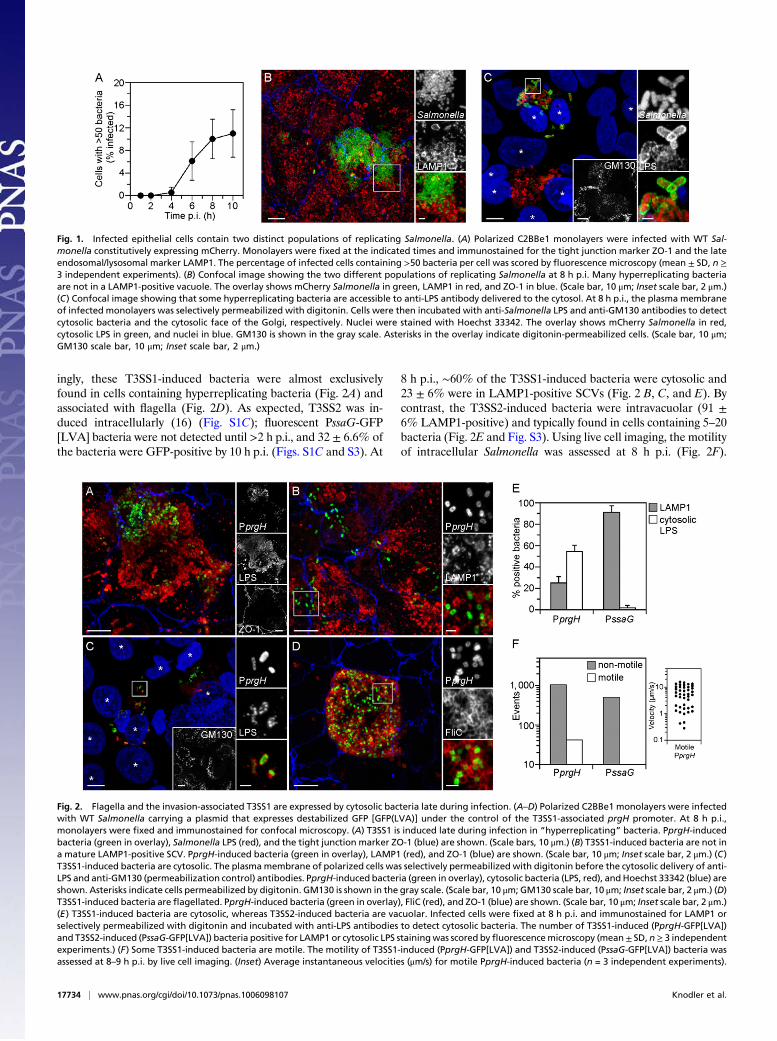
ingly, these T3SS1-induced bacteria were almost exclusivelyfound in cells containing hyperreplicating bacteria (Fig. 2A) andassociated with flagella (Fig. 2D). As expected, T3SS2 was in-duced intracellularly (16) (Fig. S1C); fluorescent PssaG-GFP[LVA] bacteria were not detected until >2 h p.i., and 32 ± 6.6% ofthe bacteria were GFP-positive by 10 h p.i. (Figs. S1C and S3). At
8 h p.i., ∼60% of the T3SS1-induced bacteria were cytosolic and23 ± 6% were in LAMP1-positive SCVs (Fig. 2 B, C, and E). Bycontrast, the T3SS2-induced bacteria were intravacuolar (91 ±6% LAMP1-positive) and typically found in cells containing 5–20bacteria (Fig. 2E and Fig. S3). Using live cell imaging, the motilityof intracellular Salmonella was assessed at 8 h p.i. (Fig. 2F).
Fig. 1. Infected epithelial cells contain two distinct populations of replicating Salmonella. (A) Polarized C2BBe1 monolayers were infected with WT Sal-monella constitutively expressing mCherry. Monolayers were fixed at the indicated times and immunostained for the tight junction marker ZO-1 and the lateendosomal/lysosomal marker LAMP1. The percentage of infected cells containing >50 bacteria per cell was scored by fluorescence microscopy (mean ± SD, n ≥3 independent experiments). (B) Confocal image showing the two different populations of replicating Salmonella at 8 h p.i. Many hyperreplicating bacteriaare not in a LAMP1-positive vacuole. The overlay shows mCherry Salmonella in green, LAMP1 in red, and ZO-1 in blue. (Scale bar, 10 μm; Inset scale bar, 2 μm.)(C) Confocal image showing that some hyperreplicating bacteria are accessible to anti-LPS antibody delivered to the cytosol. At 8 h p.i., the plasma membraneof infected monolayers was selectively permeabilized with digitonin. Cells were then incubated with anti-Salmonella LPS and anti-GM130 antibodies to detectcytosolic bacteria and the cytosolic face of the Golgi, respectively. Nuclei were stained with Hoechst 33342. The overlay shows mCherry Salmonella in red,cytosolic LPS in green, and nuclei in blue. GM130 is shown in the gray scale. Asterisks in the overlay indicate digitonin-permeabilized cells. (Scale bar, 10 μm;GM130 scale bar, 10 μm; Inset scale bar, 2 μm.)
Fig. 2. Flagella and the invasion-associated T3SS1 are expressed by cytosolic bacteria late during infection. (A–D) Polarized C2BBe1 monolayers were infectedwith WT Salmonella carrying a plasmid that expresses destabilized GFP [GFP(LVA)] under the control of the T3SS1-associated prgH promoter. At 8 h p.i.,monolayers were fixed and immunostained for confocal microscopy. (A) T3SS1 is induced late during infection in “hyperreplicating” bacteria. PprgH-inducedbacteria (green in overlay), Salmonella LPS (red), and the tight junction marker ZO-1 (blue) are shown. (Scale bars, 10 μm.) (B) T3SS1-induced bacteria are not ina mature LAMP1-positive SCV. PprgH-induced bacteria (green in overlay), LAMP1 (red), and ZO-1 (blue) are shown. (Scale bar, 10 μm; Inset scale bar, 2 μm.) (C)T3SS1-induced bacteria are cytosolic. The plasmamembrane of polarized cells was selectively permeabilized with digitonin before the cytosolic delivery of anti-LPS and anti-GM130 (permeabilization control) antibodies. PprgH-induced bacteria (green in overlay), cytosolic bacteria (LPS, red), andHoechst 33342 (blue) areshown. Asterisks indicate cells permeabilized by digitonin. GM130 is shown in the gray scale. (Scale bar, 10 μm; GM130 scale bar, 10 μm; Inset scale bar, 2 μm.) (D)T3SS1-induced bacteria are flagellated. PprgH-induced bacteria (green in overlay), FliC (red), and ZO-1 (blue) are shown. (Scale bar, 10 μm; Inset scale bar, 2 μm.)(E) T3SS1-induced bacteria are cytosolic, whereas T3SS2-induced bacteria are vacuolar. Infected cells were fixed at 8 h p.i. and immunostained for LAMP1 orselectively permeabilized with digitonin and incubated with anti-LPS antibodies to detect cytosolic bacteria. The number of T3SS1-induced (PprgH-GFP[LVA])and T3SS2-induced (PssaG-GFP[LVA]) bacteria positive for LAMP1 or cytosolic LPS stainingwas scored byfluorescencemicroscopy (mean± SD, n≥ 3 independentexperiments.) (F) Some T3SS1-induced bacteria are motile. The motility of T3SS1-induced (PprgH-GFP[LVA]) and T3SS2-induced (PssaG-GFP[LVA]) bacteria wasassessed at 8–9 h p.i. by live cell imaging. (Inset) Average instantaneous velocities (μm/s) for motile PprgH-induced bacteria (n = 3 independent experiments).
17734 | www.pnas.org/cgi/doi/10.1073/pnas.1006098107 Knodler et al.

Consistent with flagellin (FliC) expression (Fig. 2D), a subset ofT3SS1-induced bacteria was motile (Fig. 2F and Movie S2). Thispopulation moved at speeds consistent with flagellar-based mo-tility (4–15 μm/s) (17), whereas T3SS2-induced bacteria wereimmobile (Fig. 2F and Movie S2). Hence, there are at least twotranscriptionally distinct intracellular populations of replicatingbacteria in epithelial cells: T3SS2-induced intravacuolar bacteriaand T3SS1-induced flagellated bacteria that are cytosolic.
Invasion-Primed Salmonella Are Released into the Lumen byExtruding Cells. Coincident with the onset and kinetics of hyper-replication, bacteria-laden cells extruding toward the apical sidewere observed by EM (Fig. 3 A and B). Bacteria in these extrudedcells were not surrounded by a vacuolar membrane [Fig. 3B (ii)],in agreement with the confocal microscopy data for hyper-replicating bacteria (Figs. 1C and 2E). By contrast, SCV mem-branes were readily apparent around replicating bacteria in cellswithin the monolayer (Fig. 3C). To examine whether extrusion of
infected epithelial cells occurs in vivo, we used a mouse model inwhich Salmonella rapidly breaches the intestinal barrier andspreads systemically to various tissues, including the mucosalepithelium lining the gallbladder. In this murine infection model,Salmonella causes inflammation similar to cholecystitis seen inhumans during acute typhoid fever (18). We observed both freebacteria and bacteria-laden epithelial cells in the gallbladder lu-men (Fig. 3 D and E). In agreement with our observations,sloughing of enterocytes laden with Salmonella from villus tips hasalso been reported in rabbit ileal loop studies (2). Therefore,extrusion of Salmonella-infected epithelial cells is evident in bothenteric and systemic infections.Salmonella-associated extrusion resembles cell extrusion in-
volved in the rapid turnover of polarized epithelial cells in the gut.This process occurs when neighboring cells contract to pusha dying cell out of the monolayer and is characterized by re-organization of adherens and tight junctions to maintain the in-tegrity of the epithelial monolayer (19–22). Here, we observedsimilar actin contractile rings and tight junction “rosettes” at thebase of extruding Salmonella-infected cells (Fig. S4 and MovieS3), suggesting that the Salmonella-associated and homeostaticextrusion events occur via similar cell biological processes.The incidence of extrusion was significantly increased on in-
fection: 10± 2.7%of cells containing Salmonella showed evidenceof extrusion at 10 h p.i. compared with only 0.85 ± 0.89% of un-infected cells (Fig. 4A). Furthermore, cells containing T3SS1-induced Salmonella were more likely to be extruded than thosecontaining T3SS2-induced bacteria (20 ± 4.7% vs. 2.4 ± 0.9%;Fig. 4 A and B). In infected gallbladders, T3SS1-induced bacteriawere found within epithelial cells lining the gallbladder at 4 d p.i.and did not immunostain for FliC (Fig. S5). By 5 d p.i., flagellatedT3SS1-induced bacteria were predominantly found within cellsthat had been sloughed into the lumen or were free in the lumen(Fig. 4C and Fig. S5). Altogether, these data imply a strong cor-relation between T3SS1 induction and Salmonella-induced ex-trusion in vitro and in vivo. To address the invasion competenceof these bacteria, we developed a secondary infection assay(Fig. S6A). Consistent with the onset of cell extrusion from thepolarized monolayer, significant numbers of bacteria could berecovered from a naive population of epithelial cells from 6 h p.i.and increasing thereafter. This demonstrates that Salmonella re-leased from extruded cells are invasion-primed and -competent.
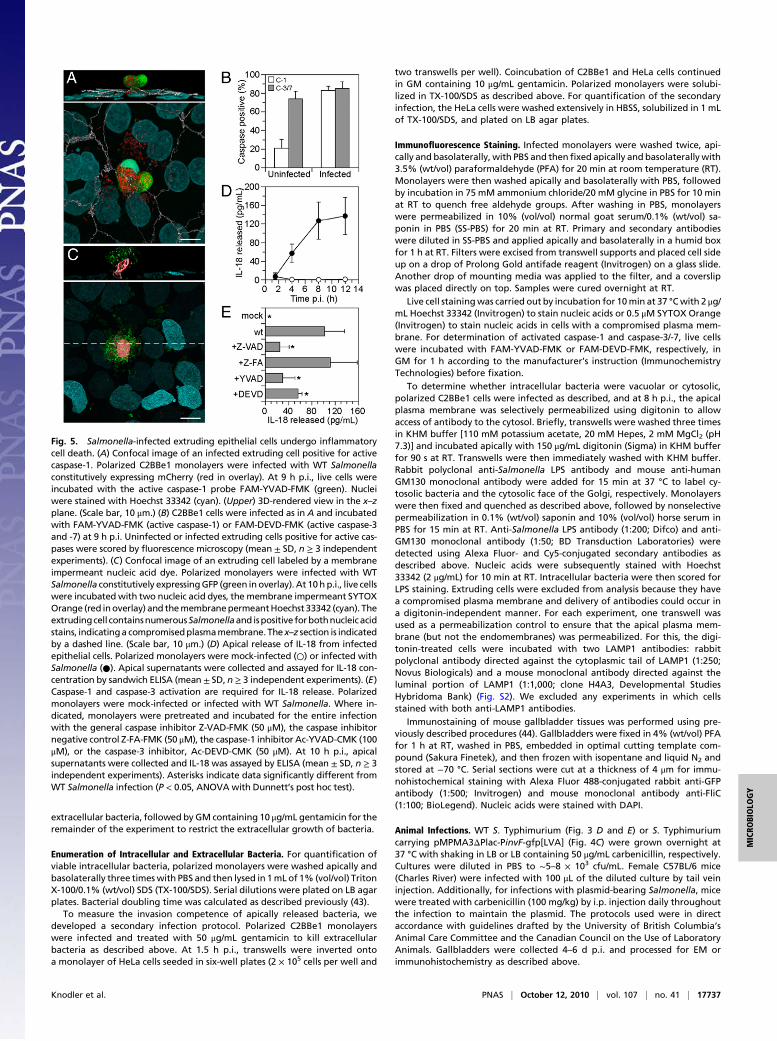
Extruding Cells Undergo Inflammatory Cell Death. During gut ho-meostasis, epithelial cells are shed into the lumen as a result ofanoikis, a form of apoptosis characterized by activated caspase-2,-3, and -9 but not caspase-1 or -8 (23–26). Because the Salmonella-laden extruding cells also showed structural features typical of celldeath both in vivo and in vitro (Fig. 3B,D, andE), we next assayedfor caspase activity. Epithelial monolayers were incubated withcell-permeable fluorescent probes that bind irreversibly to acti-vated caspases. Uninfected extruding cells were positive for activecaspase-3/-7 (74± 8.1%) but not for active caspase-1 (21 ± 9.4%),as previously described (27) (Fig. 5B). By contrast, the majority ofinfected extruding cells were positive for both active caspase-1(83 ± 4.5%) and caspase-3/-7 (85 ± 7.3%) (Fig. 5 A and B). Ho-meostatic extrusion of epithelial cells is not prevented by generalcaspase inhibition (21). Addition of a caspase-1 inhibitor did notblock, but significantly decreased, extrusion of infected cells(Fig. 4A), suggesting that the signal for homeostatic and bacterial-induced extrusion precedes caspase activation.Caspase-1–dependent programmed cell death, also known as
pyroptosis, is characterized by pore formation in the plasmamembrane, followed by cell swelling and lysis, and the proteolyticprocessing of proinflammatory cytokines, leading to the secretionof mature active IL-1β and IL-18 (28). To monitor plasmamembrane integrity, infected cells were incubated with SYTOXOrange nucleic acid dye, which can only enter cells with a com-
Fig. 3. Apical extrusion of infected cells frommucosal epithelium in vitro andin vivo. (A) SEM of the apical surface of an infected monolayer. PolarizedC2BBe1 cells were infectedwithWT Salmonella, and at 10 h p.i., samples werefixed and processed for SEM. (i) Extruding cell is bacteria-laden. Microvilli onthe apical surface of the underlying monolayer are evident. (Scale bar, 2 μm.)(ii) Inset from i. (Scale bar, 0.5 μm.) (B) Transmission EM (TEM) of an extrudedcell. Cells were infected as for A, and samples were fixed and processed forTEM. (i) Extruded cell isfilledwith bacteria and is adjacent to the apical surfaceof the monolayer, distinguished by microvilli (MV). (Scale bar, 2 μm.) (ii) Insetfrom i showing the absence of a vacuolar membrane around Salmonella inextruded cells. N, epithelial cell nucleus. (Scale bar, 0.5 μm.) (C) TEM of aninfected C2BBe1 cell that was not extruding. Cells were infected as for A, andsamples were fixed and processed for TEM. Salmonella are clearly surroundedby a vacuolar membrane (arrowheads). (Scale bar, 0.5 μm.) (D and E) EM ofinfected gallbladders. Gallbladders from C57BL/6 mice were collected 6 d p.i.andprocessed for TEM.Apical extrusion of Salmonella-infected epithelial cellsinto the lumen (L) is evident. Some bacteria are also free in the lumen. (Scalebars, 2 μm.) (E) Infected cell is undergoing extrusion into the luminal space (L),with an obvious constriction site formed by the junctional complexes ofneighboring cells (arrowheads). Microvilli (MV) on the apical surface of themucosal epithelium are indicated.
Knodler et al. PNAS | October 12, 2010 | vol. 107 | no. 41 | 17735
MICRO
BIOLO
GY

promised plasma membrane, and Hoechst 33342, a cell-perme-able nuclear stain. Salmonella-infected extruding cells were pos-itive for both dyes, indicating plasma membrane rupture, whereasneighboring cells within the monolayer stained only with Hoechst33342 (Fig. 5C). We next quantified the apical and basolateralrelease of cytokines from infected C2BBe1 cells. Although IL-1βrelease was below the limits of detection (0.8 pg/mL), we observeda steady temporal increase in the release of IL-18, which was re-stricted to the apical side of monolayers (Fig. 5D) and dependenton caspase-1 and -3 (Fig. 5E). Collectively, these data highlighta clear difference in the activated cell death program betweenSalmonella-induced and homeostatic extrusion.We have demonstrated that extruding Salmonella-infected cells
undergo inflammatory cell death. A complex but still largely un-explained pathological feature of salmonelloses is an overwhelminginflammatory response. We propose that bacterial-induced ex-trusion provides one mechanistic explanation for the pathogenesisof mucosal inflammation during Salmonella infections of the in-testine and gallbladder. In support of our data, activation of IL-18 inporcine intestinal mucosa has been reported for S. enterica serovarCholeraesuis infections (29). Interestingly, in inflammatory boweldiseases, chronic inflammation is also associated with elevatedIL-18 levels (30–32). Therefore, caspase-1–dependent IL-18 pro-duction by intestinal epithelial cells might prove to be a mediatorof mucosal inflammation associated with both autoimmune dis-orders (33, 34) and bacterial infections.Conceptually, an increased turnover of mucosal epithelium
provides the host with an ideal defense mechanism against in-fection. Indeed, it has been implicated as a protective mechanismin the gut against both bacteria and parasites (35, 36).However, webelieve that Salmonella takes advantage of this process as a uniquemeans of bacterial egress. Critical to this is the finding thata population of Salmonella is cytosolic and expresses the virulencegenes required for invasion. We hypothesize that a vacuole mat-uration defect leads to the cytosolic release of a small, but signif-icant, fraction of bacteria. The nutrient-rich cytosol supportsa high bacterial replication rate and reprograms virulence geneexpression toward invasion. The cytosolic load of bacteria issensed by the host cell, leading to inflammatory cell death and
extrusion, releasing the invasion-primed Salmonella into the lu-men of the gastrointestinal and biliary tracts. Escape into the lu-men allows Salmonella to infect secondary cells rapidly, and mayalso contribute to host-to-host transmission. Thus, by subvertinga host-dependent cell turnover event, Salmonella completes itsinfectious cycle (Fig. S6B). Given the prevalence of mucosal-dwelling pathogens, other pathogens may also use this host cellprocess as an exit strategy.
Materials and MethodsBacterial Strains and Plasmids. WT S. enterica serovar Typhimurium(S. Typhimurium) SL1344 (37) and ΔSPI2::kan (38), ΔSPI1::kan (15), and flgB::Tn10mutants (11)havebeendescribedpreviously. For constitutiveexpressionofGFP or mCherry, WT S. Typhimurium was electroporated with pFPV25.1 (39) orpFPV-mCherry (40), respectively. S. Typhimurium carrying the destabilized GFP(GFP[LVA]) (41) reporter plasmids pMPMA3ΔPlac-PprgH-gfp[LVA], pMPMA3Δ-Plac-PinvF-gfp[LVA], pMPMA3ΔPlac-PfliC-gfp[LVA], or pMPMA3ΔPlac-PssaG-gfp[LVA] (11) were used to analyze intracellular virulence gene expression.
Infection of Cultured Epithelial Cells with Salmonella. Cell lines were obtainedfrom theAmerican Type Culture Collection and used at a passage number≤13for all experiments. C2BBe1 human colorectal adenocarcinoma cells (CRL-2012), a clone of Caco-2, were maintained in DMEM (Mediatech) containing10 μg/mL human transferrin (Sigma) and 10% (vol/vol) heat-inactivated FCS(Gibco). HeLa human cervical adenocarcinoma cells were maintained inEagle’s modified Eagle medium (Mediatech) containing 10% (vol/vol) heat-inactivated FCS. Polarized monolayers were established by seeding 2 × 105
C2BBe1 cells in basal seeding medium containing MITO+ serum extender(Becton Dickinson) on BIOCOAT fibrillar collagen 24-well inserts with a 1-μmmembrane pore size (Becton Dickinson). After 24 h, the seeding medium wasreplaced with enterocyte differentiation medium containing MITO+ serumextender. Cells were incubated in differentiation medium for a total of 3 d,replaced with fresh medium each day, until the transepithelial electrical re-sistance was ≥250 Ω.cm2, as measured using a Millicell Electrical ResistanceSystem (Millipore). The medium was changed to DMEM containing 10% (vol/vol) heat-inactivated FCS (growth medium, GM) before infection.
Preparation of invasive Salmonella and infection of HeLa cells at a multi-plicity of infection (MOI) of ∼50 were as previously described (42). Polarizedmonolayers were infected apically at an MOI of ∼50–100 for 10 min and thenwashed three times apically and twice basolaterally in HBSS (Mediatech). Cellswere incubated in antibiotic-free GM until 30 min p.i. Thereafter, GM con-taining 50 μg/mL gentamicin (Sigma) was added for 1 h to kill any remaining
Fig. 4. Extruding epithelial cells contain invasion-primed Salmonella. (A) Quantification of extrusion. Polarized C2BBe1 monolayers were infected withSalmonella constitutively expressing mCherry or carrying destabilized GFP reporters for T3SS1 (PprgH-GFP[LVA]) or T3SS2 (PssaG-GFP[LVA]). At 10 h p.i.,monolayers were fixed and immunostained for ZO-1. DNA was stained with Hoechst 33342. Where indicated (+YVAD), 100 μM Ac-YVAD-CMK was addedbefore, and maintained throughout, the infection. Infected (mCherry bacteria) cells, or cells containing PprgH-positive or PssaG-positive bacteria, showingsigns of extrusion were scored by fluorescence microscopy (mean ± SD, n ≥ 4 independent experiments). Asterisks indicate significantly different frominfected cells (P < 0.05, ANOVA with Dunnett’s post hoc test). (B) Confocal image showing an extruding cell containing T3SS1-induced bacteria. PolarizedC2BBe1 monolayers infected with WT Salmonella carrying PprgH-GFP[LVA] were fixed and immunostained at 10 h p.i. DNA was stained with Hoechst 33342.PprgH-induced bacteria (green in overlay), apical plasma membrane marker, villin (red), DNA (cyan), and tight junction marker ZO-3 (gray) are shown. The x–zsection is indicated by a dashed line. (Scale bar, 10 μm.) (C) Fluorescence microscopy image showing that epithelial cells containing T3SS1-induced, flagellatedbacteria are extruded into the gallbladder lumen (L). C57BL/6 mice were infected with Salmonella carrying a plasmid that expresses destabilized GFP underthe control of the T3SS1-associated invF promoter (PinvF-GFP[LVA]). Mice were killed 5–6 d p.i., and gallbladders processed for immunostaining. DNA wasstained with DAPI. PinvF-induced bacteria (green in overlay), FliC (red), and DNA (cyan) are shown. The arrowhead indicates an extruded cell containingnumerous T3SS1-induced, flagellated bacteria. (Scale bar, 10 μm; Inset scale bar, 2 μm.)
17736 | www.pnas.org/cgi/doi/10.1073/pnas.1006098107 Knodler et al.
extracellular bacteria, followed by GM containing 10 μg/mL gentamicin for theremainder of the experiment to restrict the extracellular growth of bacteria.
Enumeration of Intracellular and Extracellular Bacteria. For quantification ofviable intracellular bacteria, polarized monolayers were washed apically andbasolaterally three timeswith PBS and then lysed in 1mLof 1% (vol/vol) TritonX-100/0.1% (wt/vol) SDS (TX-100/SDS). Serial dilutions were plated on LB agarplates. Bacterial doubling time was calculated as described previously (43).
To measure the invasion competence of apically released bacteria, wedeveloped a secondary infection protocol. Polarized C2BBe1 monolayerswere infected and treated with 50 μg/mL gentamicin to kill extracellularbacteria as described above. At 1.5 h p.i., transwells were inverted ontoa monolayer of HeLa cells seeded in six-well plates (2 × 105 cells per well and
two transwells per well). Coincubation of C2BBe1 and HeLa cells continuedin GM containing 10 μg/mL gentamicin. Polarized monolayers were solubi-lized in TX-100/SDS as described above. For quantification of the secondaryinfection, the HeLa cells were washed extensively in HBSS, solubilized in 1 mLof TX-100/SDS, and plated on LB agar plates.
Immunofluorescence Staining. Infected monolayers were washed twice, api-cally and basolaterally, with PBS and thenfixed apically and basolaterallywith3.5% (wt/vol) paraformaldehyde (PFA) for 20 min at room temperature (RT).Monolayers were then washed apically and basolaterally with PBS, followedby incubation in 75 mM ammonium chloride/20 mM glycine in PBS for 10 minat RT to quench free aldehyde groups. After washing in PBS, monolayerswere permeabilized in 10% (vol/vol) normal goat serum/0.1% (wt/vol) sa-ponin in PBS (SS-PBS) for 20 min at RT. Primary and secondary antibodieswere diluted in SS-PBS and applied apically and basolaterally in a humid boxfor 1 h at RT. Filters were excised from transwell supports and placed cell sideup on a drop of Prolong Gold antifade reagent (Invitrogen) on a glass slide.Another drop of mounting media was applied to the filter, and a coverslipwas placed directly on top. Samples were cured overnight at RT.
Live cell stainingwas carried out by incubation for 10min at 37 °Cwith 2 μg/mL Hoechst 33342 (Invitrogen) to stain nucleic acids or 0.5 μM SYTOX Orange(Invitrogen) to stain nucleic acids in cells with a compromised plasma mem-brane. For determination of activated caspase-1 and caspase-3/-7, live cellswere incubated with FAM-YVAD-FMK or FAM-DEVD-FMK, respectively, inGM for 1 h according to the manufacturer’s instruction (ImmunochemistryTechnologies) before fixation.
To determine whether intracellular bacteria were vacuolar or cytosolic,polarized C2BBe1 cells were infected as described, and at 8 h p.i., the apicalplasma membrane was selectively permeabilized using digitonin to allowaccess of antibody to the cytosol. Briefly, transwells were washed three timesin KHM buffer [110 mM potassium acetate, 20 mM Hepes, 2 mM MgCl2 (pH7.3)] and incubated apically with 150 μg/mL digitonin (Sigma) in KHM bufferfor 90 s at RT. Transwells were then immediately washed with KHM buffer.Rabbit polyclonal anti-Salmonella LPS antibody and mouse anti-humanGM130 monoclonal antibody were added for 15 min at 37 °C to label cy-tosolic bacteria and the cytosolic face of the Golgi, respectively. Monolayerswere then fixed and quenched as described above, followed by nonselectivepermeabilization in 0.1% (wt/vol) saponin and 10% (vol/vol) horse serum inPBS for 15 min at RT. Anti-Salmonella LPS antibody (1:200; Difco) and anti-GM130 monoclonal antibody (1:50; BD Transduction Laboratories) weredetected using Alexa Fluor- and Cy5-conjugated secondary antibodies asdescribed above. Nucleic acids were subsequently stained with Hoechst33342 (2 μg/mL) for 10 min at RT. Intracellular bacteria were then scored forLPS staining. Extruding cells were excluded from analysis because they havea compromised plasma membrane and delivery of antibodies could occur ina digitonin-independent manner. For each experiment, one transwell wasused as a permeabilization control to ensure that the apical plasma mem-brane (but not the endomembranes) was permeabilized. For this, the digi-tonin-treated cells were incubated with two LAMP1 antibodies: rabbitpolyclonal antibody directed against the cytoplasmic tail of LAMP1 (1:250;Novus Biologicals) and a mouse monoclonal antibody directed against theluminal portion of LAMP1 (1:1,000; clone H4A3, Developmental StudiesHybridoma Bank) (Fig. S2). We excluded any experiments in which cellsstained with both anti-LAMP1 antibodies.
Immunostaining of mouse gallbladder tissues was performed using pre-viously described procedures (44). Gallbladders were fixed in 4% (wt/vol) PFAfor 1 h at RT, washed in PBS, embedded in optimal cutting template com-pound (Sakura Finetek), and then frozen with isopentane and liquid N2 andstored at −70 °C. Serial sections were cut at a thickness of 4 μm for immu-nohistochemical staining with Alexa Fluor 488-conjugated rabbit anti-GFPantibody (1:500; Invitrogen) and mouse monoclonal antibody anti-FliC(1:100; BioLegend). Nucleic acids were stained with DAPI.
Animal Infections. WT S. Typhimurium (Fig. 3 D and E) or S. Typhimuriumcarrying pMPMA3ΔPlac-PinvF-gfp[LVA] (Fig. 4C) were grown overnight at37 °C with shaking in LB or LB containing 50 μg/mL carbenicillin, respectively.Cultures were diluted in PBS to ∼5–8 × 103 cfu/mL. Female C57BL/6 mice(Charles River) were infected with 100 μL of the diluted culture by tail veininjection. Additionally, for infections with plasmid-bearing Salmonella, micewere treated with carbenicillin (100 mg/kg) by i.p. injection daily throughoutthe infection to maintain the plasmid. The protocols used were in directaccordance with guidelines drafted by the University of British Columbia’sAnimal Care Committee and the Canadian Council on the Use of LaboratoryAnimals. Gallbladders were collected 4–6 d p.i. and processed for EM orimmunohistochemistry as described above.
Fig. 5. Salmonella-infected extruding epithelial cells undergo inflammatorycell death. (A) Confocal image of an infected extruding cell positive for activecaspase-1. Polarized C2BBe1 monolayers were infected with WT Salmonellaconstitutively expressing mCherry (red in overlay). At 9 h p.i., live cells wereincubated with the active caspase-1 probe FAM-YVAD-FMK (green). Nucleiwere stained with Hoechst 33342 (cyan). (Upper) 3D-rendered view in the x–zplane. (Scale bar, 10 μm.) (B) C2BBe1 cells were infected as in A and incubatedwith FAM-YVAD-FMK (active caspase-1) or FAM-DEVD-FMK (active caspase-3and -7) at 9 h p.i. Uninfected or infected extruding cells positive for active cas-pases were scored by fluorescence microscopy (mean ± SD, n ≥ 3 independentexperiments). (C) Confocal image of an extruding cell labeled by a membraneimpermeant nucleic acid dye. Polarized monolayers were infected with WTSalmonella constitutively expressingGFP (green inoverlay). At 10h p.i., live cellswere incubated with two nucleic acid dyes, the membrane impermeant SYTOXOrange (red inoverlay) and themembranepermeantHoechst 33342 (cyan). Theextrudingcell containsnumerousSalmonellaand ispositive forbothnucleicacidstains, indicatinga compromisedplasmamembrane. The x–z section is indicatedby a dashed line. (Scale bar, 10 μm.) (D) Apical release of IL-18 from infectedepithelial cells. Polarized monolayers were mock-infected (○) or infected withSalmonella (●). Apical supernatants were collected and assayed for IL-18 con-centration by sandwich ELISA (mean ± SD, n ≥ 3 independent experiments). (E)Caspase-1 and caspase-3 activation are required for IL-18 release. Polarizedmonolayers were mock-infected or infected with WT Salmonella. Where in-dicated, monolayers were pretreated and incubated for the entire infectionwith the general caspase inhibitor Z-VAD-FMK (50 μM), the caspase inhibitornegative control Z-FA-FMK (50 μM), the caspase-1 inhibitor Ac-YVAD-CMK (100μM), or the caspase-3 inhibitor, Ac-DEVD-CMK (50 μM). At 10 h p.i., apicalsupernatants were collected and IL-18 was assayed by ELISA (mean ± SD, n ≥ 3independent experiments). Asterisks indicate data significantly different fromWT Salmonella infection (P < 0.05, ANOVAwith Dunnett’s post hoc test).
Knodler et al. PNAS | October 12, 2010 | vol. 107 | no. 41 | 17737
MICRO
BIOLO
GY

Information on reagents, quantification of cytokine release, fluorescencemicroscopy, determination of bacterial velocities, and EM is provided in SIMaterials and Methods.
ACKNOWLEDGMENTS. We thank Caixia Ma and Tina Huang for their experttechnical assistance; Anita Mora for graphics assistance; the Genomics CoreFacility at Rocky Mountain Laboratories for DNA sequence analysis; and ReyCarabeo, Ed Miao, Staffan Svärd, and members of the Steele–Mortimer lab-
oratory for discussion and critique of this manuscript. This research was sup-ported by the Intramural Research Program of the National Institute ofAllergy and Infectious Diseases, National Institutes of Health (O.S.-M. andJ.C.) and by grants from the Canadian Institutes of Health Research and theCrohn’s and Colitis Foundation (to B.A.V.). M.M. was supported by the Cana-dian Institute of Gastroenterology/Crohn’s and Colitis Foundation of Canada/Canadian Institutes of Health Research Fellowship. B.A.V. is the Canada Re-search Chair in Pediatric Gastroenterology and the Children with Intestinaland Liver Disorders (CHILD) Foundation Research Scholar.
1. Hybiske K, Stephens RS (2008) Exit strategies of intracellular pathogens. Nat RevMicrobiol 6:99–110.
2. Wallis TS, et al. (1986) The nature and role of mucosal damage in relation to Salmonellatyphimurium-induced fluid secretion in the rabbit ileum. J Med Microbiol 22:39–49.
3. Kent TH, Formal SB, Labrec EH (1966) Salmonella gastroenteritis in rhesus monkeys.Arch Pathol 82:272–279.
4. Santos RL, et al. (2001) Animal models of Salmonella infections: Enteritis versustyphoid fever. Microbes Infect 3:1335–1344.
5. Vogelsang TM, Bøe J (1948) Temporary and chronic carriers of Salmonella typhi andSalmonella paratyphi B. J Hyg (Lond) 46:252–261.
6. Buchwald DS, Blaser MJ (1984) A review of human salmonellosis: II. Duration ofexcretion following infection with nontyphi Salmonella. Rev Infect Dis 6:345–356.
7. Beuzón CR, Salcedo SP, Holden DW (2002) Growth and killing of a Salmonella entericaserovar Typhimurium sifA mutant strain in the cytosol of different host cell lines.Microbiology 148:2705–2715.
8. Drecktrah D, Knodler LA, Howe D, Steele-Mortimer O (2007) Salmonella trafficking isdefined by continuous dynamic interactions with the endolysosomal system. Traffic 8:212–225.
9. Beuzón CR, et al. (2000) Salmonella maintains the integrity of its intracellular vacuolethrough the action of SifA. EMBO J 19:3235–3249.
10. Paz I, et al. (2010) Galectin-3, a marker for vacuole lysis by invasive pathogens. CellMicrobiol 12:530–544.
11. Ibarra JA, et al. (2010) Induction of Salmonella pathogenicity island 1 under differentgrowth conditions can affect Salmonella-host cell interactions in vitro. Microbiology156:1120–1133.
12. Galán JE, Ginocchio C, Costeas P (1992) Molecular and functional characterization ofthe Salmonella invasion gene invA: Homology of InvA to members of a new proteinfamily. J Bacteriol 174:4338–4349.
13. Van Asten FJ, Hendriks HG, Koninkx JF, Van der Zeijst BA, Gaastra W (2000)Inactivation of the flagellin gene of Salmonella enterica serotype enteritidis stronglyreduces invasion into differentiated Caco-2 cells. FEMS Microbiol Lett 185:175–179.
14. Boddicker JD, Jones BD (2004) Lon protease activity causes down-regulation ofSalmonella pathogenicity island 1 invasion gene expression after infection of epithelialcells. Infect Immun 72:2002–2013.
15. Drecktrah D, Knodler LA, Ireland R, Steele-Mortimer O (2006) The mechanism ofSalmonella entry determines the vacuolar environment and intracellular geneexpression. Traffic 7:39–51.
16. Cirillo DM, Valdivia RH, Monack DM, Falkow S (1998) Macrophage-dependentinduction of the Salmonella pathogenicity island 2 type III secretion system and itsrole in intracellular survival. Mol Microbiol 30:175–188.
17. Minamino T, Imae Y, Oosawa F, Kobayashi Y, Oosawa K (2003) Effect of intracellularpH on rotational speed of bacterial flagellar motors. J Bacteriol 185:1190–1194.
18. Menendez A, et al. (2009) Salmonella infection of gallbladder epithelial cells driveslocal inflammation and injury in a model of acute typhoid fever. J Infect Dis 200:1703–1713.
19. Madara JL (1990) Maintenance of the macromolecular barrier at cell extrusion sites inintestinal epithelium: Physiological rearrangement of tight junctions. J Membr Biol116:177–184.
20. Mayhew TM, Myklebust R, Whybrow A, Jenkins R (1999) Epithelial integrity, celldeath and cell loss in mammalian small intestine. Histol Histopathol 14:257–267.
21. Rosenblatt J, Raff MC, Cramer LP (2001) An epithelial cell destined for apoptosissignals its neighbors to extrude it by an actin- and myosin-dependent mechanism.Curr Biol 11:1847–1857.
22. Baron DA, Miller DH (1990) Extrusion of colonic epithelial cells in vitro. J Electron
Microsc Tech 16:15–24.23. Papaconstantinou HT, et al. (2000) Prevention of mucosal atrophy: Role of glutamine
and caspases in apoptosis in intestinal epithelial cells. J Gastrointest Surg 4:416–423.24. Grossmann J, et al. (2002) Induction of apoptosis before shedding of human intestinal
epithelial cells. Am J Gastroenterol 97:1421–1428.25. Fouquet S, et al. (2004) Early loss of E-cadherin from cell-cell contacts is involved in the
onset of Anoikis in enterocytes. J Biol Chem 279:43061–43069.26. Bullen TF, et al. (2006) Characterization of epithelial cell shedding from human small
intestine. Lab Invest 86:1052–1063.27. Slattum G, McGee KM, Rosenblatt J (2009) P115 RhoGEF and microtubules decide the
direction apoptotic cells extrude from an epithelium. J Cell Biol 186:693–702.28. Bergsbaken T, Fink SL, Cookson BT (2009) Pyroptosis: Host cell death and
inflammation. Nat Rev Microbiol 7:99–109.29. Foss DL, Zilliox MJ, Murtaugh MP (2001) Bacterially induced activation of interleukin-
18 in porcine intestinal mucosa. Vet Immunol Immunopathol 78:263–277.30. Monteleone G, et al. (1999) Bioactive IL-18 expression is up-regulated in Crohn’s
disease. J Immunol 163:143–147.31. Salvati VM, et al. (2002) Interleukin 18 and associated markers of T helper cell type 1
activity in coeliac disease. Gut 50:186–190.32. Wiercinska-Drapalo A, Flisiak R, Jaroszewicz J, Prokopowicz D (2005) Plasma
interleukin-18 reflects severity of ulcerative colitis. World J Gastroenterol 11:605–608.33. Dupaul-Chicoine J, et al. (2010) Control of intestinal homeostasis, colitis, and colitis-
associated colorectal cancer by the inflammatory caspases. Immunity 32:367–378.34. Zaki MH, et al. (2010) The NLRP3 inflammasome protects against loss of epithelial
integrity and mortality during experimental colitis. Immunity 32:379–391.35. Cliffe LJ, et al. (2005) Accelerated intestinal epithelial cell turnover: A new mechanism
of parasite expulsion. Science 308:1463–1465.36. Sherman MP, Petrak K (2005) Lactoferrin-enhanced anoikis: A defense against
neonatal necrotizing enterocolitis. Med Hypotheses 65:478–482.37. Hoiseth SK, Stocker BA (1981) Aromatic-dependent Salmonella typhimurium are non-
virulent and effective as live vaccines. Nature 291:238–239.38. Knodler LA, et al. (2003) Salmonella type III effectors PipB and PipB2 are targeted to
detergent-resistant microdomains on internal host cell membranes.Mol Microbiol 49:685–704.
39. Valdivia RH, Falkow S (1996) Bacterial genetics by flow cytometry: Rapid isolation of
Salmonella typhimurium acid-inducible promoters by differential fluorescence induc-
tion. Mol Microbiol 22:367–378.40. Drecktrah D, et al. (2008) Dynamic behavior of Salmonella-induced membrane
tubules in epithelial cells. Traffic 9:2117–2129.41. Andersen JB, et al. (1998) Newunstable variants of greenfluorescent protein for studies
of transient gene expression in bacteria. Appl Environ Microbiol 64:2240–2246.42. Knodler LA, Bertero M, Yip C, Strynadka NC, Steele-Mortimer O (2006) Structure-
based mutagenesis of SigE verifies the importance of hydrophobic and electrostatic
residues in type III chaperone function. Mol Microbiol 62:928–940.43. Chong A, et al. (2008) The early phagosomal stage of Francisella tularensis determines
optimal phagosomal escape and Francisella pathogenicity island protein expression.Infect Immun 76:5488–5499.
44. Khan MA, et al. (2006) Toll-like receptor 4 contributes to colitis development but not
to host defense during Citrobacter rodentium infection in mice. Infect Immun 74:
2522–2536.
17738 | www.pnas.org/cgi/doi/10.1073/pnas.1006098107 Knodler et al.