directed formation of deletions and duplications … · biology department, university of utah,...
TRANSCRIPT

Copyright 0 1985 by the Genetics Society of America
DIRECTED FORMATION O F DELETIONS AND DUPLICATIONS USING Mud(Ap, lac)
KELLY T . HUGHES AND JOHN R. ROTH
Biology Department, University of Utah, Salt Lake City, Utah 84112
Manuscript received June 15, 1984 Revised copy accepted October 1, 1984
ABSTRACT
A genetic procedure is directed for the isolation of chromosomal deletions and duplications with predetermined endpoints. These rearrangements are generated in transduction crosses using a mixture of P22-transducing phage lysates grown on two strains, each carrying a Mud-lac insertion. The formation of duplications and deletions was demonstrated in the his operon using inser- tions of Mudl-8 (a transposition-defectiveMud-lac phage). This technique was also used to make larger chromosomal duplications between Mud 1-8 insertions in the thr and leu biosynthetic operons and between Mud insertions in the thr and pjrB operons. Genetic evidence is presented that strongly suggests that inheritance of a single Mud prophage by P22-mediated crosses requires two transduced fragments; each carrying part of the Mud prophage. The two frag- ments must be involved in three recombinational exchanges; one exchange joins the donor Mud fragments and two exchanges occur between the com- posite fragment and the recipient chromosome, one on either side of the complete donor Mud element. Since duplications only occur between Mud insertions in the same orientation on the chromosome, the method of dupli- cation formation provides a simple means, of determining the orientation of Mudl-8 on the chromosome and, therefore, the direction of transcription of the gene into which Mud is inserted. This method was also used to construct recombinants between a Mudl-8 prophage and Casadaban’s protein fusion vec- tor Mud2 and, thereby, isolate Mud2-8, a Mud derivative containing the pro- tein fusion ability of Mud2 and the defective transposition functions of Mudl- 8.
E have previously described the isolation of a conditionally transposition- W defective derivative of CASADABAN and COHEN’S (1 979) specialized trans- ducing phage, Mud l(Ap, lac). The derivative, Mudl-8, transposes only in the presence of an amber suppressor mutation (HUGHES and ROTH 1984). In a strain with an amber suppressor, transposition of Mud 1-8 occurs during zygotic induction or at elevated temperatures due to the expression of a temperature- sensitive repressor protein. Thus, in the presence of an amber suppressor mutation, Mudl-8 is phenotypically similar to the original Mudl. In strains that lack an amber suppressor, Mudl-8 rarely transposes, even under zygotic induction conditions or at elevated temperatures.
In Salmonella typhimurium, the Mudl-8 may be handled as a standard trans- poson (HOLLEY and FOSTER 1982; ROSENFELD and BRENCHLEY 1980) and
Genetics 109 263-282 February, 1985.

2 64 K . T . HUGHES AND J, R. ROTH
transduced from strain to strain by P22-mediated transduction (HOLLEY and FOSTER 1982). Transductants that inherit Mudl-8 can be selected using the ampicillin resistance (Ap') conferred by Mudl -8 (CASADABAN and COHEN 1979). If the transductional recipient carries an amber suppressor, the majority of the Ap' transductants are due to transposition of Mudl-8 to new sites throughout the chromosome. If the recipient lacks an amber suppressor, then Mud18 is inherited primarily by standard recombination events. This results in inheritance of Mudl-8 in the original insertion site of the donor mutation, and the donor insertion phenotype is expressed by the recombinant.
In this report we provide genetic evidence that P22-mediated transduction of Mud requires two transduced fragments that join and are then inherited by homologous recombination. P22 can package 44 kb of chromosomal DNA into a transducing fragment (S. CASJENS, unpublished results). Since the Mud ge- nome is 38 kb in length (O'CONNOR and MALAMY 1983), it can be packaged into a single transducing phage particle at a low frequency; however, genetic evidence presented here demonstrates that either this event is very rare or the regions flanking Mud are insufficient to assure inheritance of Mud by homol- ogous recombination between the chromosome and a single transduced frag- ment. Successful transduction of Mudl-8 into a suppressor-free strain requires that two transducing phage particles carrying overlapping fragments of the Mud18 genome enter the same recipient, and these fragments must recombine to form a larger composite fragment. The composite fragment carries enough flanking chromosomal DNA to allow integration of Mud into the recipient chromosome by homologous recombination. Whether or not these two frag- ments recombine before or after they recombine with the recipient chromo- some has not been determined. We describe how recombination between Mud fragments can be used to generate chromosomal deletions and duplications with predetermined endpoints. In addition, we describe how the formation of chromosomal rearrangements can be used to determine the direction of tran- scription of genes to which Mud was fused.
Recently, a new Mud derivative, Mud2, has been constructed which is similar to Mudl except that it forms hybrid P-galactosidase fusion proteins if inserted into a coding sequence in the correct orientation and reading frame (CASA- DABAN and CHOU 1984). By requiring recombination between transducing fragments carrying Mud2 and the transposition-defective Mud 1-8, we have constructed a derivative of Mud2 (MudZ-8) which forms protein fusions and is conditionally defective in transposition.
MATERIALS AND METHODS
Bacterial strains: All strains used in this study and their sources are listed in Table 1 . All S. typhimurzum strains are derived from strain LT2.
Media: T h e E medium of VOCEL and BONNER (1956), supplemented with 0.2% glucose, was used as minimal medium. Difco nutrient broth (NB, 8 g/liter), with NaCl added (5 g/liter), was used as rich medium. Difco agar was added at a final concentration of 1.5% for solid media. Auxotrophic supplements were included in media at final concentrations described by DAVIS, BOTSTEIN and ROTH ( 1 980). Ampicillin was added to a final concentration of 30 rg/ml in NB and 15 rg/ml in E media.

DIRECTED CHROMOSOME REARRANGEMENTS 2 65
TABLE 1
List of strains
Strains Genotype Source"
S . typhimurium LT2
SL1657
T r 7 6 10 TT7692 TT7693 TT7696 TT8268 TT8269 TT8388 TT8370 TT837 1 TT9534 TT9537 TT9543 TT9544 TT9686 TT9687 TT9688 TT9689 TT9738 TT9744 TT9756 TT9757 TT9760 TT9761 TT9762 TT9765 TT9802 TT9803
TT9805 T T 9 8 10 TT98 1 1 TT9868 TT9869 TT9870 TT987 1 TT9872
TT9873
TT9874 TT9880
~ ~ 9 8 0 4
Wild type Lab collection his-22 Lab collection his-63 Lab collection hisC 12 I Lab collection his1 125 Lab collection hspLT6 hspS29 (rLT- mLT+ rs- ms+) HI-b nml- H2-
e, n, x Fels2- gulE550 ilv-452 metA22 MetE55I trpB2 strA 120 xyl-404 JlaA66
B. A. D. STOCKER
zeb-609: :Tn I O , supD50I D. BIEK hisD9953::Mud 1-8 hisF9954: :Mud 1-8 hisE9957::Mud 1-8 leuAI 177::MudI-8 leuA I 1 79::Mud 1-8 zeb-609: :TnlO recA 1 (F' 128 zzf1066::Mud 1-8) thr-458::Mudl-8 thr-469::Mudl-8 pyrB2691: :Mud 1-8 pyrB2694: :Mudl -8 DUP438[(thr-469)* Mud 1-8*(pyrB2694)] DUP4?9[(thr-471)*Mudl-8* (pyrB2694)I hsiC1264::MudI-8 hsiCl265: :Mud 1-8 his01 242 hsiCl264: :Mud 1-8 his01242 hsiCl265: :Mud 1-8
hsiC1294: : M u d 2 4 his-63 hsiCl264: :Mud1-8 his-63 hsiC1265::MudI-8 his-63 hsiC1288::Mud2-8 h isc l21 hsiC1264: :Mud 1-8 hisCl2l hsiC1265::Mudl-8 hisC 121 hsiCI288: :Mud2-8 hisCI2I hsiCl264: : Mud2-8 zeb-609: :TnlO supD5Ol hisCI.21 hsiC1264: : M u d 2 4 zeb-609: :TnlO hisC I2 I hsiCl265: : Mud2-8 zeb-609: :Tn I 0 supD50 I hisCl21 hsiC1265: :Mud2-8 zeb-609: :TnlO hisCl2I hsiC1288: : M u d 2 4 zeb-609: :TnlO supD501 hisC121 hsiC1288: : M u d 2 4 zeb-609: :Tn 10 DUP442[ (hisD9953)* Mud I -8*(hisF9954)] DEL443[(hisD9953)* Mud I -8*(hisF9954)] DUP444[(thr-469)*MudI-8*(leuA I I77)] DUP445[(thr-458)*Mudl-8*(leuAI I79)] DUP444[(thr-469)*Mudl-8*(leuAI 177 pyr-
DUP445[(thr-458)*Mudl-8*(leuA 1 I 7 9 pyr-
his01 242 hsiC1288 :Mud2-8 hsiC1264:: Mud 1-8 zeb-609: :Tn I0 supD501
h~iC1288: :Mud2-8
A685: :Tn 1 O ) ]
A685: :Tn I O ) ]

266 K. T. HUGHES AND J. R. ROTH
TABLE 1-Continued
Strains Genotype Source"
TT988 1 hsiC1264::Mud 1-8 zeb-609::TnlO TT9882 hsiC1265: :Mud 1-8 zeb-609: :TnlO supD501 TT9883 hsiC1265: : Mud 1-8 zeb-609: :TnlO TT9888 hsiC1288: :Mud14 zeb-609: :TnlO supD5Ol TT9889 hsiCl288: :Mud 1-8 reb-609: :TnlO TT9909 his01 242 hsiCl294: :Mud 1-8
E. coli MAL3 15 Mud2 c(Ts) (Ap, lac) mal- strA araD139 DEL M. CASADABAN via
(proAB-lac) X111 DEL(ara-leu) 7679 T . SILHAVY
a Unless indicated otherwise all strains were constructed during the course of this work.
Transductional methods: For all transductional crosses, the high frequency generalized transduc- ing mutant of bacteriophage P22 (HT105/1 int-201) was used (SANDERSON and ROTH 1983). Selective plates were spread directly with 2 X 10' cells and 108-109 phage. Transductants were purified and phage-free clones were isolated by streaking nonselectively onto green indicator plates (CHAN et al. 1972). Phage-free clones were then checked for phage sensitivity by cross-streaking with P22 H5 (a clear plaque mutant) phage. PI-vir phage lysates were prepared, and transductions were performed according to the method of SILHAVY, BERMAN and ENQUIST (1984). P22 lysates were titered according to the method of DAVIS, BOTSTEIN and ROTH (1980). Phage from different stocks were mixed in appropriate proportions to give equal numbers of phage from the different stocks in the final mixture.
Characterization of deletions of the his operon generated by Mudl-8: T o generate deletions, trans- ductional crosses were performed using as donors a mixture of two P22 lysates grown on different his::Mud 1-8 mutants. Recombinants were screened for possession of the expected deletions by testing whether or not they could serve as donors to repair mutations located in the region between the sites of the two Mudl-8 insertions. For example, when P22 grown on a hisD::Mudl-8 mutant was mixed with P22 grown on a hisF::Mudl-8 mutant and used to transduce LT2 to Ap', four types of Ap' transductants were obtained (see Figure 1): (1) parental hisF insertion mutants which were His- Hol+ (the ability to grow on the histidine biosynthetic intermediate histidinol (Hol) require only the expression of a functional hisD gene); (2) parental hisD insertion mutants which were His- Hol-; (3) strains with duplications of the genetic material between the points of insertion, which were His+ Hol+ (see RESULTS) and (4) strains with deletions of the genetic material between the points of insertion, which were His- Hol-. In this cross both the deletion-bearing recombinants and parental HisD- recombinants had identical phenotypes. These were distinguished by the ability to donate genetic material between the points of insertion in a standard genetic cross. P22 was grown on individual His- Hol- recombinants and used to transduce a hisBH deletion mutant (his- 22) to His+. The inability to transduce his-22 to His+ is evidence that the material between the points of insertion (including the hisCBHA genes) had been deleted. In mixed crosses using hisD and hi& insertion mutants the his-22 mutant was used to screen for deletions of the intervening genetic material as described before. In mixed crosses using hisF and hisE insertion mutants, a his1 point mutant was used to screen for deletions of the intervening genetic material.
Determination of genetic instability: Genetic duplications are unstable, undergoing homologous recombination and subsequent loss of one of the two copies. In the case of duplications generated by Mudl-8, the copies were not tandem but were separated by a Mudl-8 insertion. Segregation of one of the copies by homologous recombination results in the loss of the internal Mudl-8 insertion and, hence, yields Ap' segregants. Genetic instability of these duplications was measured by analyzing the frequency of appearance of Ap' clones. A culture of a putative duplicated strain was grown overnight in the absence of ampicillin to full density (2 X 10' cells/ml), diluted IO7- fold and plated for single colonies onto NB plates. After overnight incubation, these clones were replica-plated to NB and NB + Ap to determine the frequency of Ap' segregants. The number of Ap' segregants obtained was 1-5% of the total for small duplications (ca. 1-10 kb) and up to 30% for large duplications (up to 1000 kb).

DIRECTED CHROMOSOME REARRANGEMENTS 267
Introduction of Mud2 into S. typhimurium: The E. coli transducing phage Plvir was grown on an E. coli strain lysogenic for Mud2 (MAL315, Table 1). The phage lysate grown on MAL315 was used to transduce a S. typhimurium restriction-deficient, modification-proficient galE recipient (SL1657, Table 1 ) to Apr. Salmonella galE mutants are sensitive to infection by P1 when grown in media lacking galactose (ORNELLAS and STOCKER 1974) and sensitive to P22 when grown in media containing galactose. When introduced into Salmonella by P1 transduction, the Mud2 phage underwent zygotic induction and transposed into the Salmonella chromosome. P22 was then grown on a Salmonella Mud2 lysogen and used to introduce the Mud2 prophage into other S. typhimurium strains.
Genetic nomenclature: The basic genetic nomenclature used is that described by DEMEREC et al. (1966). Some of the his mutations are given hsi allele numbers. This is an extension of the his allele nomenclature. All his alleles up to his-9999 have been used. All subsequent mutants isolated in the his operon are given hsi allele numbers starting with hsi-1.
The genetic nomenclature used for chromosomal rearrangements is that described by Schmid (1 98 1 and SCHMID and ROTH ( 1 983a). Each deletion or duplication isolated is given a chromosomal rearrangement number regardless of whether or not the nature of the rearrangement is known. This is followed by a detailed description of the genotype in brackets adjacent to the rearrange- ment number. When the rearrangement is a result of homologous recombination between Mud prophages or other transposons, then as asterisk is used at the junction between the sequences that recombined to form the rearrangement. For example, the strain DEL983[(hisD)*Mudl- 8(hisF)] is a deletion caused by homologous recombination between the Mudl-8 insertions in hisD and hisF. The hybrid Mud 1-8 exists at the join point of the deletion. The endpoints of this deletion are the points of insertion of the original Mud insertions. The same nomenclature is used for duplications. For example, DUP984[(hisD)*Mudl-t?*(hisF)] is a duplication of the genetic material between the Mudl-8 insertions in hisD and hisF as a result of homologous recombination between the two Mud insertions. The hybrid Mudl-8 exists at the join point of the duplication. When genetic differences exist between the two copies of the duplicated material, the differences are given. DUP985[(thr)*Mudl-8*(leu)] is a Mud generated duplication between the thr and leu ope- rons. DUP986[(thr)*Mudl-8*(leu pyrA::TnlO)] is the same duplication except that one of the duplicated segments harbors the pyrA::TnlO mutation.
In RESULTS, Mud insertion mutations have been given orientation designations “A” or “B.” We define the orientation of a Mud insertion as follows. If Mud is inserted such that expression of the lac genes would require clockwise transcription, the insertion is designated as having orientation A; this designation is given regardless of whetheror not the lac genes are, in fact, being expressed. Conversely, orientation B is assigned to inserts that would require counterclockwise transcription of lac.
Construction of double mutants carrying both the hisC12 1(Am) mutation, and each of the hisC::Mud insertion mutations: T o be certain that the Mud2-8 phage does, in fact, form protein fusions, we wanted to measure how an upstream amber mutation affects the expression of different Mudl-8 and Mud2-8 insertions in the same gene. In constructing these double-mutant strains, we expected recombinational events between the point of insertion of Mud and the closely linked amber mu- tation to be a rare class of the total recombinants. If a strain carrying the hisC amber mutation is transduced to Ap‘ with P22 grown on a hisC::Mud insertion mutant, the majority of Ap‘ trans- ductants will inherit the wild-type portion of the hisC gene covering the amber mutation. Many transductants would have to be screened by backcrosses to determine which retained the amber mutation. Moreover, duplication events that are ordinarily rare can represent a larger percentage of the putative double-mutant transductants. We, therefore, set up a transductional cross to screen for the desired class of double-mutant recombinants. Phage P22 was grown on each of the Mud insertion mutants and used to transduce a his-63 deletion mutant to Apr. The his-63 deletion removes the his promoter-operator region plus the first two structural genes of the his operon, hisG and hisD. It also removes a portion of the hisC gene which includes the hisCl2l amber mutation site. Recombination between the endpoint of his-63 and the Mud insertion points was rare among the total Ap‘ transductants. However, recombinants were easily detected if X-gal was included in the transduction plates. Since the his-63 deletion removes the his promoter, a his-63, hisC::Mud double-mutant recombinant was Lac- and thus produced a white colony when grown in the presence of X-gal. Putative double mutants were tested for growth on histidinol, which

268 K . T . HUGHES AND J. R. ROTH
requires an intact hisD gene. lnability to grow on histidinol showed that the transductants retained the his-6? deletion. The putative double mutants were also transduced to His+. If 100% of the His+ transductants were Ap', then the Mud insertion had recombined into the original insertion site in hisC and had not transposed to some other location in the chromosome. The his-63 hisC::Mud double mutants were then transduced to Hol' with P22 phage grown on a hisCl2l amber mutant strain. Since the hisCZ2l mutation lies within the his-63 deletion each Hol+ trans- ductant must inherit the hisCZ2l mutation. Two classes of Hol+Ap' transductants were obtained in these crosses. The first class was the desired hisC12Z(Am) hisC::Mud double mutants. The second class of HOP transductants were those in which a duplication event of the his region had occurred in the cell prior to transduction (ANDERSON and ROTH 1981). One of the his regions was trans- duced to HOP repairing both the his-63 and the hisC::Mud mutations. Thus, one of the his copies had only the hisCl2I mutation, whereas the other retained the mutant his region of the parent. These partial diploid transductants had the desired Hol+ Ap' phenotype; however, they segregated Aps clones at high frequency, whereas the desired hisClZZ hisC::Muddouble mutants did not.
Biochemical techniques: The preparation of cell extracts has been previously described (HUGHES et al. 1983). Immunoprecipitation was done using Staphylococcus aureus cells (Calbiochem) which were prepared before use by pelleting and resuspending in 50 mM Tris-CI, 1 mM EDTA, pH 6.8, with 0.2% sodium dodecyl sulfate (SDS), and then boiling the suspension for 5 min. S. aureus was washed four times with an equal volume of RIPA buffer (BRUGGE and ERIKSON 1977) (0.15 M
NaCI, 10 mM Tris-HCI, pH 7.4, 5 mM EDTA, 1 % Triton X-100 (Sigma), 1 % deoxycholate (Sigma) and 0.1 % SDS). &galactosidase was immunoprecipitated as follows. 10 PI of @-galactosidase anti- body was added to 100 yl of crude extract, and the mixture was incubated for 1 hr on ice. After 1 hr, 100 yl of the washed S. aureus suspension was added to the reaction mixture and incubated for an additional 15 min on ice. The antibody complexes were then pelleted and washed twice with RIPA buffer. The pellet was resuspended in 20-yl sample-loading buffer (AMES 1974), boiled for 5 min and microfuged for 15 min, and 10 yl of the supernatant were loaded onto an SDS polyacrylamide gel (5% acrylamide stacking gel, 8% acrylamide separating gel), prepared and electrophoresed as described by AWES (1974).
RESULTS
Deletion and duplication mutations generated by homologous recombination be- tween Mud 1-8 fragments: When Mudl -8 is transduced into a suppressor-carrying recipient in standard P22 transductional crosses, inheritance of Mud 1-8 occurs primarily by transposition of Mudl -8 to new sites throughout the chromosome (HUGHES and ROTH 1984). When the same cross is done using a recipient that lacks an amber suppressor mutation, inheritance of Mudl -8 occurs primarily by homologous recombination of the original insertion mutation into the re- cipient chromosome. During the initial characterization of Mudl -8 , it was ob- served that P22 transduction of Mudl -8 yielded a ten- to 100-fold higher number of Ap' transductants in recipients with an amber suppressor than in strains lacking an amber suppressor. Thus, even when recombination systems are intact, inheritance by recombination seemed inefficient. This suggested some barrier to recombinational inheritance.
The large size of the Mudl -8 prophage appears to cause this impaired re- combinational inheritance. Bacteriophage P22 is 44 kb in length and Mud is 38 kb in length. Therefore, P22 is just capable of packaging a complete Mud prophage. However, inheritance of the Mud prophage by a wild-type (no sup- pressor) recipient requires homologous recombination between the recipient chromosome and the homologous chromosomal DNA flanking the Mud ge- nome in the donor-transducing fragment. T o account for the low frequency

DIRECTED CHROMOSOME REARRANGEMENTS 269
of recombinational inheritance of Mudl-8, we hypothesized that the Mudl -8 prophage was so large that very few if any single P22-transducing particles could carry the entire Mud sequence and also include sufficient flanking host sequence to allow recombination. If single P22 fragments cannot transduce Mud 1-8 fusions, then recombinational inheritance of the Mud 1-8 fusion must occur by recombination between two transduced fragments that simultaneously infect the recipient cell, each carrying part of Mudl-8. The frequency of Mudl -8 inheritance is consistent with the requirement for two fragments and, in fact, shows a second-order dependence on donor phage concentration (K. T . HUGHES and J. R. ROTH, unpublished results).
The transducing phage used in this study, P22 HT105/1, packages chro- mosomal DNA at a high frequency; approximately half of the phage particles in a lysate carry bacterial DNA (SCHMIEGER 1972; EBEL-TSIPIS, BOTSTEIN and Fox 1972; TYE, HUBERMAN and BOTSTEIN 1974). The high frequency of transducing fragments is thought to be due to a reduced specificity for the phage’s selection of packaging start sites. Although packaging sites in the mu- tant may not be completely random, clearly many more sites are recognized than is the case for wild-type P22. The two-fragment transduction of Mudl -8 requires that two transduced fragments carrying different parts of the Mud prophage enter a cell simultaneously. These fragments must overlap such that, when they recombine, a hybrid fragment is generated that includes a complete Mud prophage and substantial flanking chromosomal sequence that can recom- bine with the host chromosome. The frequency at which this occurs depends on the randomness in the distribution of endpoints at which P22 HT105/1 packages chromosomal DNA. If there is a random distribution of packaging endpoints for P22 HT105/1, then the probability of introducing two Mud- carrying fragments that overlap which can recombine to regenerate an intact Mud prophage is high. Evidence will be presented that suggests that the dis- tribution of endpoints for packaging of transducing fragments by P22 HTlO5/ 1 may not be completely random in all regions of the chromosome.
Since inheritance of Mud 1-8 requires two fragments, some striking predic- tions can be made. If lysates of two different his::Mud insertions are mixed, some of the fragments that recombine to form a complete M u d l - 8 in the recipient will be derived from different parental insertions. In addition to parental insertion types, some recombinants should carry deletions of the ma- terial between the sites of the two donor Mudl -8 prophages; others will carry duplications of this region. With equal input of phage on the donor insertions and a random distribution of fragment endpoints, one would predict that the four recombinant types (parent I, parent 11, deletion and duplication) ought to occur with equal frequency. The formation of these predicted classes is described in Figure 1; their actual detection will be described.
The predictions were borne out when a transduction cross was performed using as donors a mixture containing approximately equal numbers of phage particles from lysates grown on two different his::Mudl-8 insertions in the same orientation, one in the hisD gene and the other in the hisF gene (see Figure 1). If M u d 1 8 genomes recombine, then pieces originating from the same or

270 K. T . HUGHES AND J. R. ROTH
Donor strains his P 0 G D’& ‘ D C B H A F I € A r 3
Deletion formation
‘ h i s P 0 G D C 6 H A F I € ’
‘ F I E \
Donor fragments (AxB)
Recipient chromosome
Deletion recombinant His-, stable Ampr
Duplication formation
Donor fragments (BxA)
r his P O G D C 8 H A F I E
’ D C B H A F I E \
(COPY I 1 (COPY I1 )
Recipient chromosome
Duplication recombinant His+,
unstable Ampr
FIGURE 1 .-Recombination between different Mud fragments. Recombination events between different Mud-transducing fragments leading to the formation of hybrid deletion and duplication recombinants. Donor strain A is TT7692, a Lac+ Mudl-8 insertion in the hisD gene. Donor strain B is TT7693, a Lac+ Mudl-8 insertion in the hisF gene. T h e insertions are in the same orientation in the chromosome (orientation B, see MATERIALS AND METHODS).
different Mudl-8 insertions could join and be inserted into the chromosome by standard recombination. The expected recombinants from this experiment are shown in Figure 1. When the mixed lysate was used to transduce a wild- type strain to Ap’, four recombinant classes were possible. Two transducing fragments originating from a single insertion mutant in the hisD or hisF gene could infect the same cell and recombine to regenerate the parental Mudl-8

DIRECTED CHROMOSOME REARRANGEMENTS 271
TABLE 2
Deletions and duplications in the his operona generated by Mudl-8
Parental inser No. of Ap' tion recombi- No. of deletion No. of duplica
Donor lysates* transductants nants recombinants tion recombinants
hisD::Mudl-8 B 40 15 2 2 3 X (9 = hisD)
h i sF:Mudl -8 B * (6 = hisF) hisD::Mudl-8 B 21 21 0 0
X (12 = hisD) hisE::Mudl-8 A (9 = hi&) h i sF:Mudl -8 B 19 19 0 0
X (4 = hisF) hisE::Mudl-8 A (1 5 = hisE)
The recipient in all crosses was LT2. The donors were TT7692(hisD9953::Mudl-8, Lac+),
* The designation A or B signifies the orientation of the Mud insertion on the chromosome (see TT7693 (hisF9954)::Mud 1-8, Lac") and TT76996(hisE9957:MudI-8, Lac-).
MATERIALS AND METHODS).
prophage, which can then recombine into the recipient chromosome yielding the parental-type hisD or hisF insertion mutant transductants. The order of recombinational events between the Mud-carrying fragments and the chro- mosome is not known but should not alter the final outcome. Alternatively, and at an equal frequency, two transducing fragments, each originating from a different parental insertion mutant, could infect the same cell and recombine to generate a hybrid Mud 1-8 prophage which can then recombine into the recipient chromosome yielding deletions and duplications of the region be- tween the hisD and hisF Mudl-8 insertions (Fig. l).
The results of the mixed lysate transduction experiment are shown in Table 2. In the first cross, mixed lysates were prepared from hisD::Mudl-8 and hisF::Mudl-8 donors in which both insertions were Lac+ and thus in the same orientation on the chromosome (orientation B). Of 40 Ap' transductants, 15 were parental insertion types [hisD::Mudl-8 (9) or ItisF::Mudl-8 (S)], three were duplications of the material between the insertion sites and 22 were deletion recombinants. Thus, although one predicted 1 : 1 : 1 : 1, the cross yielded nine parental to six parental to 22 deletion to three duplication. The under rep- resentation of duplication transductants and overrepresentation of deletion transductants in this cross are not fully understood. However, a likely expla- nation is that the endpoint distribution of transduced fragments is not random. If nonrandom packaging occurred and half-fragments required for duplication formation frequently showed minimal overlap in the Mud 1-8 homology, du- plications might form less readily; this would also account for the excess dele- tion types. The distribution of recombinant types is more nearly equal in a cross described later, suggesting that the uneven distribution seen in the his region is a property of the region and not inherent in the nature of the cross.
The three his+ duplication transductants segregated Aps clones at a fre-

272 K . T . HUGHES AND J. R. ROTH
01 100 1 2 Chromosome
thr PYrA ara leu (minutes)
I I I I 1 I I I \ location
PABC DCBAP - FIGURE 2.-The threonine and leucine biosynthetic operons. A diagram of the region of the
chromosome in which the thr and leu biosynthetic operons are located. The thr gene is transcribed clockwise and the leu gene is transcribed counterclockwise.
quency of 4% after overnight incubation in nonselective medium, demonstrat- ing genetic instability of the duplications (see MATERIALS AND METHODS). The His+ phenotype of the duplications was due to complementation between the two copies of the his operon. This is possible due to two additional internal promoters in the his operon; one between the hisC and hisB genes and the other between the hisF and hisl genes (SCHMID and ROTH 1983b). Thus, copy I (in Figure 1) can provide G, D, C, B, H and A activities from the main his promoter; copy I1 can provide B, H , A, F, I and E activities by virtue of the two internal promoters. The ability to distinguish between Hol- transductants that were parental hisD::Mudl-8 insertions and those that resulted from dele- tions involved transductional recombination tests described in MATERIALS AND
The rearrangements were only generated if the two Mudl-8 insertions were in the same orientation on the chromosome: insertions oriented in opposite directions on the chromosome yielded only parental Mud insertion transduc- tant types (Table 2). This was expected since recombination between Mud prophages in opposite orientations and subsequent integration into the chro- mosome would result in a broken chromosome, and such a recipient would not be expected to survive.
Generation of large chromosomal duplications using Mud 1-8 homology: The method used in generating small duplications within the his operon between Mud 1-8 insertions should be able to generate duplications of larger segments of the host chromosome. This possibility was tested by using Mudl-8 insertions in the thr and leu operons (Figure 2). The intervening region makes up about 3% of the chromosome. These insertions were used primarily because the orientation of transcription of these two operons was already known: the thr operon is transcribed clockwise and the leu operon counterclockwise (SAND- ERSON and HARTMAN 1978). Thus, we could deduce the orientation on the chromosome of Mud insertions in those operons. A Lac+ insertion in thr would be in the same orientation on the chromosome as a Lac- insertion in leu and vice versa. Since Mud insertions must be in the same chromosomal orientation to generate duplications, we could predict which insertions in the thr and leu operons could be used to generate duplications. Duplications formed by recom- bination between appropriately oriented thr::Mudl-8 and leu::Mudl-8 insertion fragments should result in unstable Ap' prototrophs (Figure 3).
Single and mixed P22 lysates grown on strains harboring Mudl-8 insertions in both orientations were used to transduce LT2 to Apr. The results of these crosses are shown in Table 3. In the transductions involving insertions in the same chromosomal orientation, approximately 25% of the Ap' transductants
METHODS.
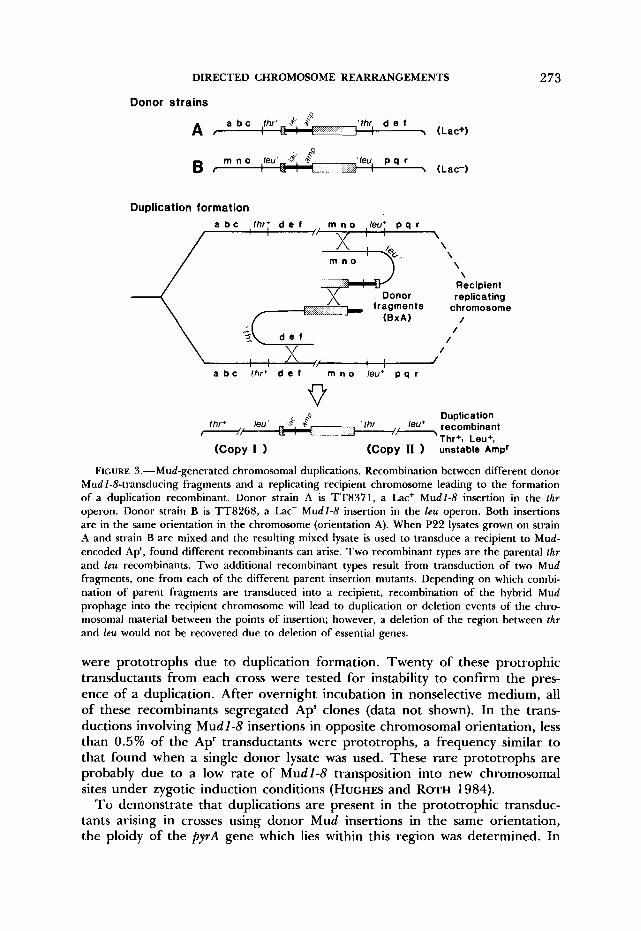
DIRECTED CHROMOSOME REARRANGEMENTS 273
Donor strains
'thr d e f A t I -, (Lac+)
'leu, p q r I \ (Lac-)
Duplication formation
\ \ \
Recipient replicating
\
/ /
/ v /
I , I 1
a b c thr+ d e f m n o leu' p q r
Duplication thr leu* recombinant leu' ,&,
U 1 '- T h + . Leu+. thr+/ . . . . - - - ,
(COPY I 1 (Copy 11 unstable Amp'
FIGURE 3.-Mud-generated chromosomal duplications. Recombination between different donor Mudl-8-transducing fragments and a replicating recipient chromosome leading to the formation of a duplication recombinant. Donor strain A is TT8371, a Lac+ Mudl-8 insertion in the thr operon. Donor strain B is TT8268, a Lac- Mudl-8 insertion in the leu operon. Both insertions are in the same orientation in the chromosome (orientation A). When P22 lysates grown on strain A and strain B are mixed and the resulting mixed lysate is used to transduce a recipient to Mud- encoded Ap', found different recombinants can arise. T w o recombinant types are the parental thr and leu recombinants. Two additional recombinant types result from transduction of two Mud fragments, one from each of the different parent insertion mutants. Depending on which combi- nation of parent fragments are transduced into a recipient, recombination of the hybrid Mud prophage into the recipient chromosome will lead to duplication or deletion events of the chro- mosomal material between the points of insertion; however, a deletion of the region between thr and leu would not be recovered due to deletion of essential genes.
were prototrophs due to duplication formation. Twenty of these protrophic transductants from each cross were tested for instability to confirm the pres- ence of a duplication. After overnight incubation in nonselective medium, all of these recombinants segregated Aps clones (data not shown). In the trans- ductions involving Mud 1-8 insertions in opposite chromosomal orientation, less than 0.5% of the Ap' transductants were prototrophs, a frequency similar to that found when a single donor lysate was used. These rare prototrophs are probably due to a low rate of Mudl-8 transposition into new chromosomal sites under zygotic induction conditions (HUGHES and ROTH 1984).
To demonstrate that duplications are present in the prototrophic transduc- tants arising in crosses using donor Mud insertions in the same orientation, the ploidy of the pyrA gene which lies within this region was determined. In

274 K. T. HUGHES AND J. R. ROTH
TABLE 3
Duplications between the leucine and threonine biosynthetic operonsa generated by Mud 1-8
No. of Ap' No. of leucine No. of threonine No. of Donor($* transductants auxotrophs auxotrophs prototrophs
leuA1177::Mudl-8 A leuA1179:Mudl-8 B thr-469::MudI-8 A thr-458::Mudl-8 B leuA1177 A
X thr-469 A leuAl177 A
thr-458 B leuA1179 B
X thr-469 A leuA1179 B
X thr-458 B
X
677 389 518 374
518
488
351
439
677 388
0 0
185
294
210
233
0 0
517 373
199
193
139
106
0 1' I' 1'
134
1"
2'
100
The recipient in all crosses was LT2. The donors were TT8268 (leuAl177::Mudl-d, Lac-), TT8269 (leuAl179::Mudl-8, Lac+), TT8370 (thr-458::Mudl-8, Lac-), TT8371 (thr-469::Mudl-8, Lac+).
The designation A or B signifies the orientation of the Mud insertion on the chromosome (see MATERIALS A N D METHODS).
' There is a low background frequency at which the Mudl-8 phage transposes under zygotic induction conditions (HUGHES and ROTH 1984).
two experiments, prototrophic Ap' transductants from mixed lysate crosses were purified and transduced to tetracycline resistance with P22 grown on a strain carrying a pyrA::TnlO insertion mutation. Each cross yielded Tc' Ap' protrophic transductants as expected if the PyrA gene were duplicated. Four Ap' Tc' colonies from one cross (TT8268 X TT8371) and three from the other (TT8269 X TT8370) were grown overnight nonselectively to allow segregation of the duplication and loss of the Mudl-8 located at the join point (see Figure 4). Each culture yielded Aps segregants at a frequency of 9-30%. Approximately 50% of the Aps segregants in each case were Tc', Pyr-, whereas the other 50% were Tc', Pyr+ (Table 4). This showed that the Ap' prototrophic transductants carried a duplication of chromosomal material between the sites of the two donor Mud insertions.
The appearance of prototrophic recombinants at a frequency of 25% (Table 3) strongly supports the notion that the P22 transduction of Mudl-8 is a two- fragment process. If the deletion recombinants were lost due to the deletion of essential genes, then the frequency of duplication recombinants should have been 33% of the total. This was fairly close to the actual frequency of 25% that we observed.
Determination of the orientation of transcription of the pyrB operon: The ability of Mudl-8 insertions in the same orientation on the chromosome to form

DIRECTED CHROMOSOME REARRANGEMENTS 275
Merodiploid recombinant Tet' PyrA+
thr+ +
Amps thr+ pyrA::TnlO leu+ thr+ pyrA+ leu+ Haploid - - segregant
Tet' PyrA- types
TetS PyrA+
FIGURE 4.-Duplication segregation. Segregation of Mudld-generated duplication between the thr and leu operons. P22 grown on a strain carrying a pyrA::TnlO insertion was used to transduce the duplicated recipient to tetracycline resistance. Since the pyrA::TnlO is integrated into either one of the two duplicated regions, all of the Tc' transductants are Pyr+ due to the presence of a pyrA+ gene in the other duplicated segment. When selection for ampicillin resistance is removed, ampicillin-sensitive clones arise in which homologous recombination between the duplicated region has occurred. This results in loss of Mudl-8 and one of the two pyrA alleles. Recombination event I results in an Ap" PyrA- Tc' haploid segregant; recombination event I1 results in an Ap' PyrA+ Tc" haploid segregant.
duplications provides a simple method for determining the direction of tran- scription of virtually any gene. If one has a Mudl-8 insertion in a gene of interest, then a transductional testcross can be done with a mixed lysate grown on the Mudld insertion mutant of interest and an insertion mutant in a reference gene whose transcription orientation is already known. When this mixed lysate is used to transduce a recipient to Ap', duplication recombinants will only arise if both Mud insertions are oriented in the same direction. Thus, if the cross yields duplications, the orientation of the Mud prophages is the same.
The Mud 2-8 duplication formation method was used to determine the tran-

276 K. T . HUGHES AND J. R. ROTH
TABLE 4
Segregation of thr-leu duplications ~~
Segregant phenotypes" Total segregants
Duplication tested (Ap') PyrA- PyrA+
DCJP444[(thr-469)*Mudl-8*(leuA 1 I77 pyrA685: :TnlO)] 1 818 377 44 1 2 856 414 442 3 91 1 403 508 4 949 54 1 408
Total 3534 1735 1799
DUP445[(thr-458)*Mud1-8* (LeuAI 179 pyrA685::TnlO)l 1 433 174 259 2 234 128 106 3 455 237 218
Total 1122 539 583
All PyrA- segregants were Tc'; all PyrA+ segregants were Tc".
scription orientation of the pyrB operon. P22-transducing lysates were prepared on a Lac+ and on a Lac- pyrB::Mudl-8 insertion mutant. Phage grown on the pyrB insertion mutants were mixed (individually) with P22 lysates from a Lac+ thr::Mudl-8 insertion mutant. These mixed lysates were used to transduce LT2 to Apr. The resulting transductants from each cross were screened for proto- trophic (duplication) recombinants. The results in Table 5 show that only prototrophic recombinants were produced from the Lac- pyrB::Mudl-8 inser- tion. All of these prototrophic recombinants segregated Aps clones at high frequencies, demonstrating that they carry duplications. These results show that transcription of the pyrB gene is counterclockwise, opposite to that of the thr operon. When this experiment was first done, the direction of transcription of the pyrB operon was unclear (SANDERSON and HARTMAN 1978). Since then, the direction of transcription of the pyrB gene in Salmonella has been deter- mined independently (SANDERSON and ROTH 1983) and is in agreement with our results.
Construction of a conditionally transposition-defective derivative of Mud2 (Ap', lac): We have demonstrated that fragments of Mudl-8 from different donors can recombine to generate an intact Mud phage. Mudl-8 and Mud2 are iden- tical except for the end of Mud carrying the lac operon and the mutations resulting in the transposition defect of Mudl-8. We set out to introduce the mutations in Mudl-8 into Mud2 by isolating duplications that require recom- bination within the Mud prophages.
P22 grown on a Lac+ thr::Mud2 insertion mutant was mixed with P22 grown on a Lac- pyrB::Mudl-8 insertion mutant and used to transduce LT2 to Ap' on NB + ampicillin plates at 44". Duplications from this cross result in the formation of a hybrid Mud recombinant at the duplication join point with the appropriate halves from the two different Mud prophages. When Mud2 infects recipients at 44" it can not lysogenize due to relief of repression resulting in

DIRECTED CHROMOSOME REARRANGEMENTS 277
TABLE 5
Transcription orientation of the pyrB gene
Donor(s)
pyrB26Yl::Mudl-8 B pyrB2694::Mud1-8 A thr770Y: : Mud 1-8 A pyrB26YI B
+ thr7709 A pyrB2694 A
+ thr770Y A
~~
No. of Ap' No. of uracil No. of threonine No. of transductants auxotrophs auxotrophs prototrophs
306 303 0 3 210 210 0 0 164 0 163 1
215 1 1 1 104 0
284 124 186 26
The recipient in all crosses was LT2. The donors were TT8371 (thr-46Y::Mudl-8, Lac+), TT9534 (pyrB2691::Mudl-B, Lac+) and TT9537 (pyrB2694::Mudl-8, Lac-).
expression of the wild-type transposition genes and the kil gene (BUKHARI 1975; GIPHART-GASSLER et al. 1979; GIPHART-GASSLER and VAN DE PUTTE 1979; WAGGONER et al. 1981). Both of these properties are absent in the Mudl-8 prophage. Thus, Mud2 could not have lysogenized at 44", and none of the Ap' transductants at 44" should be due to inheritance of the parental Mud2 prophages. Only two types of prophages were expected among the Ap' transductants, Mudl-8 prophages from the pyrB::Mudl-8 donor or a hybrid prophage resulting from the recombination of the protein fusion half of Mud2 with the transposition-defective half of Mudl-8. The latter class would be duplication recombinants and thus genetically unstable. Of 19 Ap' transduc- tants obtained in this cross, three were prototrophic and genetically unstable. These three transductants should have contained hybrid Mud2-Mud 1-8 pro- phages. One of these, which we have designated Mud2-8, was screened for its ability to generate @-galactosidase protein fusions. The transposition-defective nature of this phage has been determined and is reported elsewhere (HUGHES and ROTH 1984). The observation that duplications generated from this cross arose through recombination of the two Mud prophages supports the two- fragment model for P22 transduction of Mud.
Demonstration that Mud2-8 forms protein fusions: Proof that protein fusions are formed by Mud2-8 was obtained by both genetic and biochemical tech- niques. First, many Lac+ Mudld and Mud2-8 insertion mutants in the his operon were isolated. Of 65 his::Mudl-8 insertion mutants screened for pro- duction of @-galactosidase on medium containing X-gal, 37 were Lac+ (blue) and 28 were Lac- (white). However, of 154 his::Mud2-8 insertion mutants isolated, only 25 were Lac+. The frequency of Lac+ his::Mud2-8 insertions was about the frequency observed for protein fusions generated with the original Mud2 (CASADABAN and CHOU 1984). Approximately half of the random inser- tions of Mud into the his operon would have been expected to be in the proper transcriptional orientation to express the lac operon from the his promoter, but only one in three of these would be in the correct translational reading

278 K. T. HUGHES AND J. R. ROTH
TABLE 6
8-Galactosidase activities for operon fusion and protein &ion Mud prophages in the hisC (hsiC) gene of S. typhimurium
Upstream amber Amber &Galactosidase Strain mutation Mud fusion suppressor specific activity
TT988 1 TT9880 TT9803 TT9802 TT9883 TT9882 TT9805 TT9804 TT9889 TT9888 TT98 1 1 TT98 10 LT2"
None None
hisCl2l hisCl2l
None None
hisCI2I hisCl2l
None None
hisc l21 hisCl2l
None
hsiC1264: : M u d 1 4 hsiC1264::MudI-8 hsiC1264: : M u d l - 8 hsiCl264: :Mud 1-8 hsiC1265: : M u d 1 4 hsiCl265: :Mud 1-8 hsiCl265: :Mud 1-8 hsiC1265: : M u d 1 4 hsiC1288: : M u d 2 4 hsiC1288: :Mud2-8 hsiC1288: : M u d 2 4 hsiC1288: :Mud2-8
(No fusion)
None sup D None supD None supD None supD None supD None sup0 None
52.9 164
114 5.15
15.3 47.5
31.7 37.4
2.34
125 0.148
0.257 30.3
' A s a control, wild-type strain LT2, which has no fusion and does not have a lac operon, was assayed.
h i s D h i s C ( h s i C ) h i s 8
MI264 hsic1265
W P l ~ hsic1294 kCl288 " " I ... ...
63 , . . . 2558 ...
2624 .,.
822 ...
... 2630
... 2236
. .. 538 129 ... ,
FIGURE 5.-Deletion map of the hisC gene. A genetic map of the hisC gene including the Mudl- 8 (hisC1264 and hsiC1265) and M u d 2 4 (hsiC1294 and hsiC1288) insertions. The location of the hisCIPl(Am) allele is also shown. Some of the his insertions are given hsi allele numbers; this is an extension of the his genetic nomenclature (see MATERIALS AND METHODS).
frame. Thus, we expected Lac+ insertions using a protein fusion Mud to occur at a frequency of about l/s of the total insertions, which is very close to the frequency we observed using Mud2-8. For the operon fusion prophage Mudl- 8, we expected 50% of the total insertions to be Lac' which is also very close to the frequency we observed. The locations of the Mud insertions in the his operon were determined by complementation analysis and deletion mapping (data not shown). Mud2-8 insertions in the hisC gene were used for further tests. These were chosen because two hisC::Mudl-8 and one hisC::Mud2-8 in- sertions mapped in the same deletion interval in the hisC gene, allowing direct comparison of these Mud18 and Mud2-8 insertions (Figure 5) .

DIRECTED CHROMOSOME REARRANGEMENTS 279
A B C D E F G H wvqnnra(nm I
- 200K
- 68K 4 !
c FIGURE B.--SDS gel of hisC-lac fusions. SDS polyacrylamide gel electrophoresis of proteins
precipitated with antibody to @-galactosidase from strains harboring Mudl-8 and Mud2-8 fusions in the hisC gene. The Mud2-8 insertion mutants, TT9874 (lane A) and TT9909 (lane B), have a normal &galactosidase band (1 2 1 kd) and a larger band (1 43 and 127 kd, respectively) correspond- ing to a fusion of @-galactosidase to the upstream portion of the hisC gene. The fusion in TT9874 maps further downstream than the fusion in TT9909 (see Figure 5 ) and thus results in a larger fusion protein. In contrast, Mudl-8 operon fusions, TT9688 (lane C) and TT9689 (lane D), which map in the same deletion interval as the Mud2-8 protein fusion in TT9874 (lane A), produce only normal-sized @-galactosidase bands (121 kd). A strain with no fusion, his01242 (lane E) produces no &galactosidase band as does a control in which no extract was added (lane F). Authentic 8- galactosidase (1 16 kd, Sigma) which was not antibody precipitated is shown in lane G.
Direct biochemical proof that Mud2-8 forms protein fusions were obtained by precipitating the presumed hybrid &galactosidase protein produced in hisC::MudZ-8 and hisC::Mud2-8 insertion mutants using antibody to @-galacto- sidase and comparing the mobility of these proteins with wild-type /3-galacto- sidase on SDS polyacrylamide gels. To maximize fusion protein production, strains were constructed that contain the hisC::Mud fusion and the his regula- tory mutation his01242. T h e p-galactosidase proteins produced in these strains are shown in Figure 6. The hisC::Mudl8 insertion mutants produced a 121- kilodalton (kd)-sized &galactosidase band, whereas the hisC::Mud2-8 insertion mutants produced both the 121-kd &galactosidase band and a larger fusion band (127 kd for hsiCZ294::Mud2-8 and 143 kd for hsiCl288::Mud2-8). The 143-kd fusion protein corresponds to the Mud2-8 insertion that mapped fur- ther downstream in the hisC gene and consequently resulted in a larger fusion protein. The presence of the smaller 121-kd &galactosidase band in the fusion strains was probably due to the instability of the fused protein which was subject to proteolytic degradation to a more stable, normal-sized @-galactosid- ase. This phenomenon has also been observed for the original Mud2 phage (CASADABAN and CHOU 1984). The 121-kd @-galactosidase band in each fusion was larger than wild-type &galactosidase (1 16 kd). This is because both Mudl

280 K. T. HUGHES AND J. R. ROTH
and Mud2 fusion vectors have a hybrid lac2 gene that is fused to other DNA sequences (CASADABAN and COHEN 1979; CASADABAN and CHOU 1984).
Formation of protein fusions by Mud2-8 was also demonstrated genetically. Double mutants were constructed which placed the hisCl21 amber mutation in front of the hisC::Mudl-8 insertions and the hisC::Mud2-8 insertion which maps in the same deletion interval in the hisC gene (Figure 5). If the amber mutation were upstream of a Mud operon fusion in the same gene, then we expected the amber mutation to decrease @-galactosidase expression due to a polar effect on transcription of the lac operon. However, if the amber mutation were upstream of a Mud protein fusion in the same gene, then we expected no expression of &galactosidase because the amber mutation would prevent translation of the lac2 gene.
We examined the effect of a hisC amber mutation on the expression of @- galactosidase when placed upstream of a hisC::Mud 1-8 (operon fusion) or a hisC::Mud2-8 (protein fusion) insertion. The results of this experiment are shown in Table 6. When the hisCl2l amber mutation was upstream of the operon fusion insertions hsiC1264::Mudl-8 or hsiCl265::Mudl-8, the expres- sion of @-galactosidase was reduced about eight-fold compared to isogenic in- sertion mutants which do not have the upstream amber mutation. When the hisCl2l amber mutation was upstream of the hsiC1288::Mud2-8 fusion the level of @-galactosidase was essentially zero. When an amber suppressor was introduced into the hisCl21(Am) hisC::Mud2-8 strain, @-galactosidase activity was restored to almost the level of the isogenic hisC::Mud2-8 insertion strain without an upstream amber mutation (Table 6). Introduction of an amber suppressor into the hisCl2I hsiC::Mudl-8 double-mutant strains resulted in relief of polarity; the levels of @-galactosidase in these strains were also restored to levels similar to the isogenic insertion mutant without an upstream amber mutation. Even without the hisC121 mutation, all of the fusion strains showed a three-fold increase in @-galactosidase expression in the presence of an amber suppressor mutation (Table 6). This was due to a slight derepression of the his operator control region in the presence of the amber suppressor caused by read-through of the termination codon for the leader peptide (JOHNSTON et al. 1980). These results are consistent with the Mud2-8 insertion resulting in a protein fusion of @-galactosidase with the upstream portion of hi&.
DISCUSSION
The data presented here demonstrate that P22-mediated transduction of the Mud prophage by homologous recombination involves two simultaneously transduced fragments. The requirement that the two fragments recombine provides a way to select for recombination between Mud prophages. This fact can be used to construct directed duplications and deletions and to determine the orientation of any Mud insertion in the chromosome. Deletions and dupli- cations are standard tools in genetic analysis. Deletions have been used in genetic mapping of genes and operons; duplications have been very useful in complementation analysis and dominance studies of mutations in genes throughout the chromosome. An advantage of the technique using the Mud

DIRECTED CHROMOSOME REARRANGEMENTS 281
transposon is that the rearrangements are constructed in a single transductional cross and make up a major fraction of the total transductants. In the cases presented here the duplications are readily distinguished by the prototrophic phenotype associated with the duplication event. The methods described here have been successfully applied to many other regions of the Salmonella chro- mosome by several people in this lab (B. COOKSON, D. HILLYARD, R. JETER and D. ROOF, unpublished results).
A genetic technique similar to the one reported here has been reported for determining transcription orientation in E. coli (MACNEIL 198 1). The tech- nique in E. coli requires the construction of Mu-Mud double lysogens and can be employed to construct deletions and duplications by recombination events between the Mu and Mud homology. Its disadvantage is the difficulty of con- structing the double lysogens. The method reported here is simpler because prior strain construction is unnecessary.
The methods described here may also be useful in genetic analysis of phage Mu. P22-mediated transduction of Mud prophages requires recombination be- tween two Mud phages. Use of P22 to transduce singly marked prophages provides a means of forcing Mu-Mu recombination. We have used this method to construct the Mud2-8 prophage which forms protein fusions to the lac2 protein and is conditionally transposition defective. We expect Mud2-8 to be a useful tool in the construction of protein fusions to @-galactosidase.
We thank MOLLY SCHMID for the idea that two fragments might be required for transduction of MUD1 by p22. This work was supported by Public Health Service grant GM 23408 from the National lnstitutes of Health. K. T. H. was supported by predoctoral training grant T32-GM 07464-07 from the National Institutes of Health.
LITERATURE CITED
AMES, G. F., 1974 Resolution of bacterial proteins by polyacrylamide gel electrophoresis on slabs. J. Biol. Chem. 249: 634-644.
ANDERSON, P. and J. ROTH, 1981 Spontaneous tandem genetic duplications in Salmonella typhi- murium arise by unequal recombination between rRNA (rm) cistrons. Proc. Natl. Acad. Sci.
Identification of a transformation-specific antigen induced
USA 7 8 3113-3117.
BRUGGE, J. S. and R. L. ERIKSON, 1977
BUKHARI, A. I., 1975
CASADABAN, M. J. and J. CHOU, 1984 In vivo formation of gene fusions encoding hybrid 8- galactosidase proteins in one step with a transposable Mu-lac transducing phage. Proc. Natl. Acad. Sci. USA 81: 535-539.
CASADABAN, M. J. and S. N. COHEN, 1979 Lactose genes fused to exogenous promoters in one step using a Mu-lac bacteriophage: in vivo probe for transcriptional control sequences. Proc. Natl. Acad. Sci. USA 76: 4530-4533.
Specialized transduction of tetracycline resistance by phage P22 in Salmonella typhimurium 11. Properties of a high fre- quency transducing lysate. Virology 5 0 883-898.
DAVIS, R. W., D. BOTSTEIN and J. R. ROTH, 1980 Advanced Bacterial Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
DEMEREC, M., E. ADELBERG, A. J. CLARK and P. E. HARTMAN, 1966 A proposal for a uniform nomenclature in bacterial genetics. Genetics 5 4 6 1-76.
by an avian sarcoma virus. Nature (Lond.) 269: 346-348. Reversal of mutator phage Mu integration. J. Mol. Biol. 9 6 87-99.
CHAN, R. K., D. BOTSTEIN, T. WATANABE and Y. OGATA, 1972

282
EBEL-TSIPIS, J., D. BOTSTEIN and M. S. Fox, 1972 Generalized transduction by phage P22 in Salmonella typhimurium. I. Molecular origin of transducing DNA. J. Mol. Biol. 71: 433-448.
GIPHART-GASSLER, M., T. GOOSEN, A. VAN MEETEREN, C. WIJFFELMAN and P. VAN DE PUTTE, 1979 Properties of the recombinant plasmid pGPl containing part of the early region of bacteriophage Mu. Cold Spring Harbor Symp. Quant. Biol. 43: 1179-1 185.
GIPHART-GASSLER, M. and P. VAN DE PUTTE, 1979 Thermo-inducible expression of cloned early genes of bacteriophage Mu. Gene 7: 33-50.
Bacteriophage P22 as a vector for Mu mutagenesis in Salmonella typhimurium: isolation of nad-lac and pnc-lac gene fusions. J. Bacteriol. 152 959- 962.
6-Aminoni-
K. T. HUGHES AND J. R. ROTH
HOLLEY, E. A. and J. W. FOSTER, 1982
HUGHES, K. T., B. T. COOKSON, D. LADIKA, B. M. OLIVERA and J. R. .ROTH, 1983 cotinamide-resistant mutants of Salmonella tyfhimvrium. J. Bacteriol. 154: 1 126-1 136.
HUGHES, K. T. and J. R. ROTH, 1984 Conditionally transpositiondefective derivative of Mu dl(Amp Lac). J. Bacteriol. 159 130-137.
JOHNSTON, H. M., W. M. BARNES, F. G. CHUMLEY, L. Boss1 and J. R. ROTH, 1980 Model for the regulation of the histidine operon of Salmonella. Proc. Natl. Acad. Sci. USA 77: 508-512.
MACNEIL, D., 1981 General method, using Mu-Mud1 dilysogens, to determine the direction of transcription of and generate deletions in the glnA region of Escherichia coli. J. Bacteriol. 146
O'CONNOR, M. B. and M. H. MALAMY, 1983 A new insertion sequence, IS121, is found on the Mu dII(Ap lac) bacteriophage and the Escherichia coli K-12 chromosome. J. Bacteriol. 156
ORNELLAS, E. P. and B. A. D. STOCKER, 1974 Relation of lipopolysaccharide character to P1
ROSENFELD, S. A. and J. E. BRENCHLEY, 1980 Bacteriophage P1 as a vehicle for Mu mutagenesis
SANDERSON, K. E. and P. E. HARTMAN, 1978 Linkage map of Salmonella typhimurium, edition V.
SANDERSON, K. E. and J. R. ROTH, 1983 Linkage map of Salmonella typhimurium, edition VI.
SCHMID, M. B., 198 1 Chromosome rearrangements in Salmonella typhimurium. Ph.D. Thesis,
SCHMID, M. B. and J. R. ROTH, 1983a Genetic methods for analysis and manipulation of inversion
SCHMID, M. B. and J. R. ROTH, 1983b Internal promoters of the his operon in Salmonella
SCHMIEGER, H., 1972 Phage P22-mutants with increased or decreased transduction abilities. Mol.
SILHAVY, T. J., M. L. BERMAN and L. W. ENQUIST, 1984 Experiments with Gene Fusions. Cold
TYE, B.-K., J. A. HUBERMAN and D. BOTSTEIN, 1974 Non-random circubr permutation of phage
VOGEL, H. J. and D. M. BONNER, 1956 Acetylornithase of Escherichia coli: partial purification
WAGGONER, B., M. PATO, A. TOUSSAINT and M. FAELEN, 1981 Replication of mini-Mu prophage
260-268.
669-679.
sensitivity in Salmonella typhimurium. Virology 6 0 491-502.
in Salmonella typhimurium. J. Bacteriol. 144: 848-85 1.
Microbiol. Rev. 4 2 47 1-5 19.
Microbiol. Rev. 47: 410-453.
University of Utah, Salt Lake City, Utah.
mutations in bacteria. Genetics 105 51 7-537.
typhimurium. J. Bacteriol. 153: 11 14-1 119.
Gen. Genet. 119 75-88.
Spring Harbor Laboratory, Cold Spring Harbor, New York.
P22 DNA. J. Mol. Biol. 85: 501-532.
and some properties. J. Biol. Chem. 218: 97-106.
DNA. Virology 113: 379-387.
Communicating editor: E. JONES