diiferential efect of oleic and palmitic acid on lipid accumulation
TRANSCRIPT

GASTROENTEROLOGY jgh_5733 830..840
Differential effect of oleic and palmitic acid on lipidaccumulation and apoptosis in cultured hepatocytesMatteo Ricchi,* Maria Rosaria Odoardi,* Lucia Carulli,* Claudia Anzivino,* Stefano Ballestri,*Adriano Pinetti,† Luca Isaia Fantoni,‡ Fabio Marra,§ Marco Bertolotti,* Sebastiano Banni,¶
Amedeo Lonardo,* Nicola Carulli* and Paola Loria*
*Dipartimento Integrato di Medicina Interna, Endocrinologia, Metabolismo & Geriatria – Università degli Studi di Modena e Reggio Emilia,†Dipartimento di Chimica, Università di Modena e Reggio Emilia, ‡Dipartimento di Scienze Biomediche, Sezione di Chimica Biologica, Universitàdegli Studi di Modena e Reggio Emilia, Modena, §Dipartimento di Medicina Interna and Center for Research, Transfer, and High EducationDENOTHE, Università di Firenze, Firenze, and ¶Dipartimento di Biologia Sperimentale, Università degli Studi di Cagliari, Cagliari, Italy
AbstractBackground and Aim: Studies have shown monounsaturated oleic acid to be less toxicthan palmitic acid and to prevent/attenuate palmitic acid hepatocites toxicity in steatosismodels in vitro. However, to what degree these effects are mediated by steatosis extent isunknown.Methods: We evaluated whether steatosis per se is associated with hepatocytes apoptosisand determined the role of oleic and palmitic acid, the most abundant fatty acids in westerndiets, on triglyceride accumulation and apoptosis in an in vitro model of steatosis inducedin three hepatocytic cell lines (HepG2, HuH7, WRL68). The impact of incubation for 24 hwith oleic (0.66 and 1.32 mM) and palmitic acid (0.33 and 0.66 mM), alone or combined(molar ratio 2 : 1) on steatosis, apoptosis, and insulin signalling, was evaluated.Results: Concurrent with PPARg and SREBP-1 gene activation, steatosis extent was largerwhen cells were treated with oleic than with palmitic acid; the latter fatty acid wasassociated with increased PPARa expression. Cell apoptosis was inversely proportional tosteatosis deposition. Moreover, palmitic, but not oleic acid, impaired insulin signalling.Despite the higher amount of fat resulting from incubation of the two fatty acids combined,the apoptosis rate and impaired insulin signalling were lower than in cells treated withpalmitic acid alone, indicating a protective effect of oleic acid.Conclusions: Oleic acid is more steatogenic but less apoptotic than palmitic acid inhepatocityc cell cultures. These data may provide a biological basis for clinical findings ondietary patterns and pathogenetic models of nonalcoholic fatty liver disease.
Key words
apoptosis, cell culture techniques, fatty acids,saturated and insaturated, fatty liver, insulinresistance.
Accepted for publication 21 October 2008.
Correspondence
Nicola Carulli and Paola Loria, Università degliStudi di Modena, Dipartimento di MedicinaInterna, Endocrinologia, Metabolismo eGeriatria, Nuovo Ospedale CivileSant’Agostino Estense di Baggiovara, ViaGiardini Baggiovara, Modena 41100, Italy.Email: [email protected]
Partly supported by grants from MIURMinistero Istruzione Università e RicercaScientifica -PRIN 2004061213_001.
Abbreviations:
DAPI, 4′,6-diamidino-2-phenylindoledihydrochloride; FA, Fatty Acid; FFA, FreeFatty Acids; IR, Insulin Resistance; NEFA,Non esterified fatty acids; OA, Oleic Acid;PA, Palmitic Acid; T2D, Type 2 Diabetes;TG, Triglycerides; VLDL, Very low densitylipoproteins.
IntroductionFatty acids (FAs) are major components of biological cell mem-branes that play important roles in intracellular signalling and asprecursors for ligands that bind to nuclear receptors.1,2 FAs repre-sent vital energy stores, but high-fat diets are associated with thedevelopment of obesity and type 2 diabetes (T2D).3,4 Several linesof evidence indicate the importance of both quantitative and quali-tative (e.g. saturated vs unsaturated) changes in dietary FAs asrelevant mechanisms for the development of nonalcoholic fattyliver disease (NAFLD) both in rodent models and in humans.5–9
Finally, in obese NAFLD patients, the decreased unsaturated/
saturated fatty acid ratio in serum, fat and liver tissue might have apathogenetic role in the disease.10,11 Increased free fatty acids (FFA)levels are linked with the pathogenesis of insulin resistance (IR),which is considered a major determinant in the pathogenesis ofNAFLD. Peripheral IR results in increased concentrations of circu-lating FFAs due to unopposed antilipolytic insulin action.Addition-ally, accumulation of FFA results in an impaired post-receptorinsulin signalling12 contributing to IR and causing a further increasein FFA.
The hepatocytes are not a physiological site of lipid storage anddevelopment of steatosis is associated with cellular dysfunctionand apoptosis.13,14 This phenomenon, which also occurs in the
doi:10.1111/j.1440-1746.2008.05733.x
830 Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

kidney, pancreas and heart, is referred to as lipotoxicity and isbelieved to play a significant role in the pathogenesis of tissuedamage.15 In fact, the extent of apoptosis correlates with the sever-ity of steatohepatitis and the stage of fibrosis in human NAFLD.16
Lipotoxicity also contributes to decreased insulin sensitivity,perhaps by promoting the accumulation of fat-derived metabolitesthat inhibit insulin signalling and action.3,17
Fatty acids are chemically classified as saturated and unsatur-ated (monounsaturated and polyunsaturated) and their structureaffects their biological effects. This is of particular relevance forthe most abundant FAs present in the diet and in serum, namelypalmitic acid (PA), a saturated FA, and oleic acid (OA), a monoun-saturated FA.18 In spite of the amount of information available forthe pathogenesis of triglyceride accumulation, little is known onthe metabolic effects of PA and OA as determinants of the patho-physiology of NAFLD.
Studies on the effect of FAs-induced steatosis on cellular apo-ptosis have demonstrated that PA and OA mixtures-induced ste-atosis is associated with apoptosis in hepatocyte cell cultures.19
Moreover, similar to other cell lines, monounsaturated FAs wereless toxic also in hepatocytes20 and were able to prevent/attenuatePA toxicity.21 However, it remains unknown to what extent theseeffects were mediated by degree/type of lipid accumulated in thehepatocytes.
Aim of the present study was to evaluate the role of OA and PA,the major FA present in western diets, on steatogenesis, cell sur-vival, FAs composition, gene expression and insulin signalling inan in vitro model of steatosis. Given that a priori it could not beruled out that specific hepatocytic cell lines might differ in theirsusceptibility to steatosis/apoptosis, we carried out experiments inthree different cell lines.
Methods
Hepatocyte cell cultures
Three cell lines with different characteristics were used: (i) HepG2cells are derived from a well differentiated human hepatoblastomacell line that retain many characteristics of normal differentiatedquiescent hepatocytes, and are p53 wild-type;22,23 (ii) WRL-68cells, a fetal liver cell line;24 and (iii) p53-mutated HuH7 cells,derived from a differentiated hepatocellular carcinoma.25 HepG2and WRL68 cells were purchased from Istituto ZooprofilatticoSperimentale (Brescia, Italy) and HuH7 cells from JapaneseCancer Research Resources Bank (Osaka, Japan).
Long-chain FAs, palmitic (16:0) and oleic (18:1) were providedas sodium salts (Sigma-Aldrich, Milan, Italy). Palmitic acid andOA were dissolved in MetOH 99% (stock solution 100 mM).Stock solutions were kept at -20°C before the experiments. Solu-tions and reagents used for cell cultures were from GIBCO LifeTechnologies Ltd (Grand Island, NY, USA).
Protocol of the study—induction andevaluation of steatosis
Steatosis was induced by a slight modification of previouslydescribed methods.26,27 HepG2, WRL-68 and HuH-7 cell cultureswere incubated with phenol red-free medium containing 10% ofcharcoal stripped fetal bovine serum (FBS; Cambrex, East
Rutherford, NJ, USA), 1% bovine serum albumin (BSA), supple-mented with FFA (oleic and palmitic acid alone or in association)at the following final concentrations: a) PA: 0.33 mM and0.66 mM; b) OA: 0.66 mM and 1.32 mM and c) mixtures of OAand PA (ratio 2:1) at two different final concentrations: PA0.33 mM + OA 0.66 mM (final fatty acids concentration 1 mM)and PA 0.66 mM + OA 1.32 mM (final fatty acids concentration2 mM). Control cell cultures were incubated with plain medium orwith medium added with the vehicle in which fatty acids weredissolved. After 24 h of incubation with PA and OA alone or inassociation, the extent of steatosis, apoptosis and gene expressionwere evaluated as detailed above. In agreement with previousstudies,19,27 we found 24 h to be optimal incubation time. At 12 hor less the TG accumulation was low and no detectable variationsin gene expression could be shown (data not shown). A moreprolonged incubation time (e.g. 48 h and 72 h) while not providinga significant advantage in terms of intracellular TG accumulationwas associated with a significant decrease in cell viability at higherFFA concentrations in our and others’ experience (data notshown,28).
After fixation with formaldehyde, neutral lipids were stainedusing 0.5% Oil-Red-O (Sigma-Aldrich) in isopropanol for 30 minand nuclei were stained with hematoxylin.
For the electron microscopy pictures cells were detached fromthe substrate with trypsin, fixed in 2.5% glutaraldeyde for 1 h andthan post-fixed in tyrode 1% OsO4 (osmium tetraoxyde) for 1 h.Cells were then dehydrated in progressive concentrations ofethanol, dried in propylene oxide and embedded in Durcupan’sresin. Sections 50 nm were cut on an Ultratatome (Reichert-Jung,Wetzlar, Germany), placed on mesh copper grids (S162, ASSING,Rome, Italy) and stained with 7% uranyl acetate and 2.66% leadcitrate. Micro-photographs were taken by an electron microscope(JEOL, model 2011, Peabody, MA, USA) at 200 kV.
Intracellular triglyceride content was evaluated after lysis of thecells with NaOH 0.3 N. Triglyceride concentration (mg/dL) wasdetermined by standard technique with an automatic analyzer(Roche, Milan, Italy) and normalised by protein content (mg/mL).
Total intracellular lipid content was evaluated by Nile Red stain-ing (Adipored, Cambrex); briefly, cells were grown in 96 black-plates and treated with FAs. At the end of incubation cells werewashed twice with phosphate buffered saline (PBS) and incubatedwith Adipored for 10 min. Fluorescence was evaluated as previ-ously described.29
Evaluation of apoptosis
Apoptosis, assessed with DAPI (4’,6-diamidino-2-phenylindoledihydrochloride, Sigma-Aldrich) staining and caspases 3/7 activ-ity (Promega, Milan, Italy) were evaluated as previouslydescribed.29
Measurement of the fatty acid composition
Lipids were extracted from cells using the method of Folch et al.30
Aliquots were mildly saponified as previously described31 in orderto obtain free fatty acids for high pressure liquid chromatography(HPLC) analysis. Separation of FAs was carried out with a Agilent1100 HPLC system (Agilent, Palo Alto, CA, USA) equipped witha diode array detector. A C-18 Inertsil 5 ODS-2 Chrompack
M Ricchi et al. Fatty acids and apoptosis in hepatocytes
831Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

column (Chrompack International BV, Middleburg, The Nether-lands), 5 mm particle size, 150 ¥ 4.6 mm, was used with a mobilephase of CH 3 CN/H2O/CH 3 COOH (70/30/0.12, v/v/v) at a flowrate of 1.5 mL/min. Unsaturated fatty acids were detected at200 nm. Spectra (195–315 nm) of the eluate were obtained every1.28 s and were electronically stored.32 These spectra were takento confirm the identification of the HPLC peaks.
Analysis of saturated FAs and further confirmation of unsatur-ated fatty acids was carried out by GC assay of FA methyl esters.
Free fatty acids obtained as described above were methylated bythe addition of 14% BF3/CH3OH at room temperature and imme-diately extracted into a solvent consisting of n-hexane and water(4:3 ratio). After centrifugation to separate the two phases, thehexane phase was saved and the aqueous phase was furtherextracted by another round of hexane. The two hexane collectionswere combined, dried, and redissolved in 500 mL of n-hexane.33
The gas chromatograph (Model 6890, Agilent) was equippedwith split ratio of 20:1 injection port, a flame ionization detector(FID), an autosampler (Model 7673, Agilent), a 100 m HP-88fused capillary column (Agilent), and an Agilent ChemStationsoftware system. The injector and detector temperatures were setat 250°C and 280°C respectively. Hydrogen served as carrier gas(1 mL/min), and the FID gases were H2 (30 mL/min), N2 (30 mL/min), and purified air (300 mL/min). The temperature programwas as follows: initial temperature was 120°C, programmed at10°C/min to 210°C and 5°C/min to 230°C, then programmed at25°C/min to 250°C and held for 2 min.
Determination of cellular mRNA level
In HepG2 cell line total RNA was isolated using the RNeasy lipidtissue kit (Qiagen, Milan, Italy) and quantified/checked with RNANano LabChip (Agilent, Milan, Italy). About 1 mg of total RNAwas reverse-transcribed with the high capacity cDNA Archive Kit(Applied Biosystems, Monza Italy). TaqMan polymerase chainreactions (PCR) were performed on cDNA samples using theTaqMan Universal PCR Master Mix (Applied Biosystems)according to PRISM 7900 HT Sequence Detection Systems.
The TaqMan strategies for each gene have been developed asAssay-on-Demand by Applied Biosystems. Gene expression pro-filing was achieved using the comparative cycle threshold (CT)method of relative quantification (the calibrator samples were non-treated cells, with 18S RNA used as endogenous control). Data areexpressed as log2 of the relative quantity (RQ) defined also as ‘foldinduction versus the controls’.
Western blot analysis of cellular proteins
Confluent, serum-starved (12 h) HepG2 were treated with fattyacid, after stimulation with 100 nM of insulin for 15′, the cellswere quickly placed on ice, and washed with ice-cold PBS. Themono-layer was lysed in RIPA buffer (20 mM Tris·HCl, pH 7.4,150 mM NaCl, 5 mM EDTA, 1% Nonidet P-40, 1 mM Na3VO4,1 mM phenyl methyl sulfonyl fluoride, 0.01% protease inhibitorcocktail [Protease Inhibitor Cocktail, Sigma-Aldrich]) and trans-ferred to micro-centrifuge tubes. Insoluble proteins were discardedby centrifugation at 12 000 rpm at 4°C. Protein concentration inthe supernatant was measured in duplicate using a commerciallyavailable assay (Pierce, Rockford, IL, USA). Equal amounts of
total cellular proteins were separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and analyzed byWestern blot using primary antibodies as indicated. Detection wasperformed using a chemiluminescent substrate (ECL, Amersham,IL, USA). Primary antibodies were rabbit Phospho-Akt (Ser-473)Antibody (Cell Signalling Technology, Danvers, MA, USA) andmouse Anti-b-Actin antibody (Sigma-Aldrich).
Statistical analysis
Results were expressed as mean � standard error (SE). All datarepresent a minimum of three experiments conducted in triplicate,unless otherwise specified. The significance of differences wasassessed by Student’s t-test for independent data. Linear regres-sion analysis was performed by the least square method. Thedifferences between slope values were evaluated by the analysis ofvariance. Significance was accepted at the P < 0.05 level. Statisti-cal analysis was performed with the aid of SPSS statistical soft-ware (version 14.0 for Windows, SPSS Inc., Chicago, IL, USA).
Results
OA and PA differentially affect triglycerideaccumulation in hepatocytic cell lines
We first analyzed the development of lipid accumulation in an invitro model of hepatic steatosis. HepG2 were exposed to increas-ing concentrations of OA or PA, or to a combination of the twofatty acids at a 2:1 ratio in favour of OA. After 24 h, lipid accu-mulation was evident in all cells exposed to FA, as indicated bystaining with Nile Red staining (Fig. 1). The degree of fat accu-mulation was roughly proportional to the concentration of FA to
Figure 1 Effect of different fatty acids on lipid accumulation in HepG2cells. After 24 h of incubation (for details see materials and methods)intracellular lipid droplets were measured by Nile Red. Data areexpressed as means � standard error of Relative Fluorescence Units(RFLU) per mg of protein. Three experiments conducted in duplicate.*P < 0.01 vs control; **P < 0.01 vs palmitic acid (PA) 0.33 mM;***P < 0.01 vs PA 0.66 mM; ****P < 0.01 vs oleic acid (OA) 0.66 mM;*****P < 0.01 vs PO1 mM.
Fatty acids and apoptosis in hepatocytes M Ricchi et al.
832 Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd
which cells were exposed. Similar findings were obtained forWRL68 and HuH7 cell lines (data not shown). Finally, electronmicroscopy pictures displayed a massive increase in the numberand size of lipid droplets into the cytosol of HepG2 cells incubatedwith FAs as compared to controls (Fig. 2).
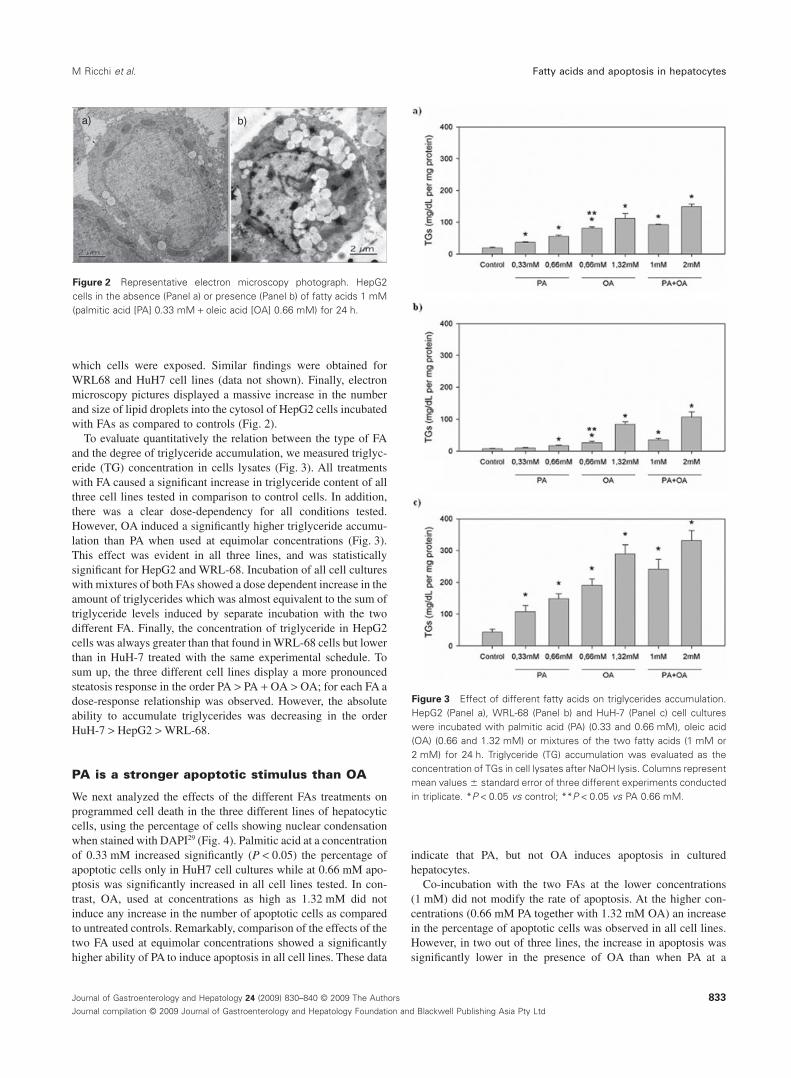
To evaluate quantitatively the relation between the type of FAand the degree of triglyceride accumulation, we measured triglyc-eride (TG) concentration in cells lysates (Fig. 3). All treatmentswith FA caused a significant increase in triglyceride content of allthree cell lines tested in comparison to control cells. In addition,there was a clear dose-dependency for all conditions tested.However, OA induced a significantly higher triglyceride accumu-lation than PA when used at equimolar concentrations (Fig. 3).This effect was evident in all three lines, and was statisticallysignificant for HepG2 and WRL-68. Incubation of all cell cultureswith mixtures of both FAs showed a dose dependent increase in theamount of triglycerides which was almost equivalent to the sum oftriglyceride levels induced by separate incubation with the twodifferent FA. Finally, the concentration of triglyceride in HepG2cells was always greater than that found in WRL-68 cells but lowerthan in HuH-7 treated with the same experimental schedule. Tosum up, the three different cell lines display a more pronouncedsteatosis response in the order PA > PA + OA > OA; for each FA adose-response relationship was observed. However, the absoluteability to accumulate triglycerides was decreasing in the orderHuH-7 > HepG2 > WRL-68.
PA is a stronger apoptotic stimulus than OA
We next analyzed the effects of the different FAs treatments onprogrammed cell death in the three different lines of hepatocyticcells, using the percentage of cells showing nuclear condensationwhen stained with DAPI29 (Fig. 4). Palmitic acid at a concentrationof 0.33 mM increased significantly (P < 0.05) the percentage ofapoptotic cells only in HuH7 cell cultures while at 0.66 mM apo-ptosis was significantly increased in all cell lines tested. In con-trast, OA, used at concentrations as high as 1.32 mM did notinduce any increase in the number of apoptotic cells as comparedto untreated controls. Remarkably, comparison of the effects of thetwo FA used at equimolar concentrations showed a significantlyhigher ability of PA to induce apoptosis in all cell lines. These data
indicate that PA, but not OA induces apoptosis in culturedhepatocytes.
Co-incubation with the two FAs at the lower concentrations(1 mM) did not modify the rate of apoptosis. At the higher con-centrations (0.66 mM PA together with 1.32 mM OA) an increasein the percentage of apoptotic cells was observed in all cell lines.However, in two out of three lines, the increase in apoptosis wassignificantly lower in the presence of OA than when PA at a
a) b)
Figure 2 Representative electron microscopy photograph. HepG2cells in the absence (Panel a) or presence (Panel b) of fatty acids 1 mM(palmitic acid [PA] 0.33 mM + oleic acid [OA] 0.66 mM) for 24 h.
Figure 3 Effect of different fatty acids on triglycerides accumulation.HepG2 (Panel a), WRL-68 (Panel b) and HuH-7 (Panel c) cell cultureswere incubated with palmitic acid (PA) (0.33 and 0.66 mM), oleic acid(OA) (0.66 and 1.32 mM) or mixtures of the two fatty acids (1 mM or2 mM) for 24 h. Triglyceride (TG) accumulation was evaluated as theconcentration of TGs in cell lysates after NaOH lysis. Columns representmean values � standard error of three different experiments conductedin triplicate. *P < 0.05 vs control; **P < 0.05 vs PA 0.66 mM.
M Ricchi et al. Fatty acids and apoptosis in hepatocytes
833Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

concentration of 0.66 mM was used alone. These data are stronglysuggestive of a protective effect of OA on apoptosis inducedby PA.
To add further evidence to the differential effects of the two fattyacids on apoptosis, we measured the activity of caspases 3/7, thatbelong to the group of the execution caspases, activated once thecell death program is fully operative (Fig. 5). In cells exposed toPA, the activity of caspases 3/7 increased in a dose-dependentfashion. In contrast, OA did not have any effects on caspase
activity except in HepG2, where a significant increase wasobserved. However, it should be stressed that the increase inresponse to OA was only 1–2 fold as compared to control, whilePA induced an 8–16 fold increase. As a result, caspases 3/7 activityin HepG2 exposed to OA was always significantly lower than incells treated with PA.
In co-incubation experiments, the addition of OA to PAdecreased caspases activity in comparison to cells treated with PAalone in HepG2 and in WRL68 but not in HuH7 cell line. Thus,data on caspases activity are closely similar to those obtained withDAPI staining.
Figure 4 Effect of fatty acids on apoptosis rate. In HepG2 (Panel a),WRL-68 (Panel b) and HuH-7 (Panel c) apoptosis was quantified byassessing the characteristic nuclear changes of apoptosis using thenuclear binding dye 4’,6-diamidino-2-phenylindole dihydrochloride(DAPI) and counting cell numbers under a fluorescent microscopy.Columns represent mean values � standard error of three differentexperiments conducted in triplicate. *P < 0.05 vs control; **P < 0.05 vspalmitic acid 0.66 mM.
Figure 5 Effect of fatty acids on caspases 3/7 activity. In HepG2 (Panela), WRL-68 (Panel b) and HuH-7 (Panel c) caspases 3/7 activity wasquantified measuring the fluorescence from Z-DEVD-R110 cleavage andnormalised by protein content in parallel plates. Columns representmean values � standard error of three different experiments conductedin triplicate. *P < 0.05 vs control; **P < 0.05 vs palmitic acid (PA)0.66 mM; ***P < 0.05 vs PA 0.33 mM.
Fatty acids and apoptosis in hepatocytes M Ricchi et al.
834 Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

PA and OA are associated with different FAprofiles in HepG2 cells
We hypothesized that the mechanisms of cell toxicity due to palm-itic acid could be ascribed to different fatty acid profiles and to theesterified/unesterified ratio. To test this hypothesis, we comparedindividual and total fatty acid composition obtained by HPLC withthe results of TG content assessed enzymatically.
The absolute FAs composition in steatotic HepG2 cultures afterexposure to different treatments is shown in Table 1. All treatmentsinduced significant changes in the concentration of various FAs.PA and OA concentration increased after incubation with therespective FA. In addition, while PA incubation was associatedwith an increase 16:1, 18:1c11 and 14:0, the incubation with OAinduced an increase of 18:0, 16:0 and 14:0. As expected, theincubation with the two FAs induced a combined pattern and wasalso associated with a reduction in the concentration of 16:1 ascompared to PA alone. The putative metabolic fate of FAs added tothe medium based on results shown in Table 1 is depicted inFigure 6.
Total FA content was proportional to the moles of fatty acidsadded to the medium (data not shown). Contrasting with data onTG content, we found that OA and PA at equimolar doses induceda comparable accumulation of total fatty acids. This strongly sug-gests that after incubation with PA the concentration of TG is lower(Fig. 1) than total FA concentration due to the presence of a con-sistent FFA not being detected in the total TG assay. It could beindirectly estimated that the percentage of FFAs following incu-bation with PA is in the order of magnitude of 15% (vs 0–1%following incubation with OA).
FAs structure and not TG content is the keydeterminant of apoptosis
To better define the relationship between the degree of steatosisand induction of apoptosis, we plotted the intracellular TG con-centration with the activity of caspases 3/7 (Fig. 7). While theseparameters were significantly correlated for both FA, the slope ofthe curve obtained with PA was significantly steeper than the oneobserved for OA. It is of relevance that the regression for com-bined treatments fall in between PA and OA, again suggesting aprotective effect of OA on PA induced cytotoxicity. Similar rela-tionships were observed also plotting TG versus percentage ofapoptosis evaluated by DAPI staining (data not shown). These dataconfirm that for a given amount of intracellular triglycerides, thedegree of apoptosis depends on the FA used for the induction ofsteatosis being far higher for PA, followed by their association andby OA.
Gene expression
The expression of PPARalpha, gamma and SREBP-1 constantlyincreased following exposure to FAs. However, as shown inFigure 8, PA but not OA induced a significant increase in PPARaexpression vs controls (P < 0.05). In contrast, incubation ofHepG2 with OA but not with PA was associated, at both concen-trations used, with an increase in the expression of PPARg vscontrols (P < 0.05). In addition OA at the highest concentrationincreased the expression of SREBP-1 vs controls (P < 0.05). When T
ab
le1
Fatt
yac
idpr
ofile
ofH
epG
2ce
llsin
cuba
ted
with
palm
itic
acid
(PA
),ol
eic
acid
(OA
)or
PAan
dO
Aco
mbi
ned
20:5
22:6
16:1
20:4
18:2
22:4
18:1
18:1
c11
12:0
14:0
15:0
16:0
18:0
20:0
22:0
Tota
lFA
s
cont
rol
1.27
45.
139
31.3
468.
966
16.4
007.
276
80.8
4763
.996
1.56
710
.308
2.65
499
.095
23.9
491.
500
1.67
435
5.99
0S
D0.
065
0.60
33.
168
0.39
20.
207
0.34
515
.220
10.6
170.
158
2.16
90.
082
20.1
594.
812
0.21
30.
018
58.2
28P
0,3
1.17
64.
653
47.1
59*
8.18
017
.143
6.69
685
.252
74.4
851.
584
12.0
532.
791
150.
288*
25.6
941.
771
1.64
644
0.57
0S
D0.
043
0.27
41.
939
0.29
10.
207
0.03
65.
729
0.58
60.
178
0.06
70.
653
20.8
914.
244
0.88
50.
388
36.4
10P
0,6
1.32
75.
336
68.9
36*
10.1
3821
.924
8.03
799
.727
97.6
34*
1.85
915
.323
*3.
405
223.
582*
28.5
331.
814
1.78
658
4.36
0*S
D0.
033
0.41
53.
297
1.05
40.
833
0.35
39.
266
5.62
70.
441
1.11
20.
431
40.3
113.
536
0.17
30.
289
69.1
71O
0,6
1.36
25.
218
30.6
5110
.061
18.4
848.
441
291.
330*
66.2
241.
314
12.0
833.
404
104.
335
22.1
651.
616
1.77
257
8.46
0*S
D0.
208
0.53
92.
227
1.15
13.
109
1.17
43.
547
3.26
50.
504
1.79
00.
581
8.48
41.
211
0.12
00.
195
28.1
04O
1,3
1.41
56.
240
35.7
5611
.745
20.5
8811
.683
594.
991*
92.4
851.
542
17.6
34*
3.44
715
2.01
7*37
.363
*2.
120
2.30
399
1.33
0*S
D0.
054
1.24
612
.192
1.94
83.
826
3.45
929
.215
2.59
40.
181
2.34
40.
300
5.29
61.
287
0.23
80.
172
64.3
52P
O1
1.16
75.
061
36.0
389.
466
19.6
368.
433
293.
478*
70.8
541.
625
13.5
053.
084
169.
168*
27.5
141.
676
1.80
666
2.51
0*S
D0.
100
0.31
93.
719
0.43
32.
014
1.80
835
.181
21.1
760.
070
3.05
20.
363
18.8
113.
065
0.15
40.
209
90.4
73P
O2
1.43
65.
445
49.4
72*
10.3
0123
.122
9.59
453
2.03
8*86
.541
*2.
659*
16.6
87*
3.60
128
0.68
2*33
.767
*1.
480
2.15
410
58.9
80*
SD
0.02
50.
263
10.3
010.
662
6.80
20.
610
35.3
335.
623
0.15
41.
022
0.01
927
.117
4.27
20.
319
0.20
292
.724
*P<
0.05
vsco
ntro
l.D
ata
are
expr
esse
das
nmol
es/m
gof
prot
eins
.
M Ricchi et al. Fatty acids and apoptosis in hepatocytes
835Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

the effect of two equimolar doses of PA and OA were compared,the difference did not reach statistical significance.
PA inhibits signaling downstream of theinsulin receptor
Peripheral IR, is caused, at least in part, by blunted transmissionof the signal along the enzymatic cascade downstream of theinsulin receptor. A critical step in this pathway is represented by
Figure 6 Putative metabolic pathway compatible with changes in fattyacid profile observed after adding palmitic acid (PA) and oleic acid (OA)to cell cultures base on findings shown in Table 1. PA (16:0) increasesthe concentration of 16:1, 18:1c11 presumably by stimulating delta 9desaturation of PA yielding 16:1, further elongation (18:1c11). Theincrease of 14:0 may be ascribed to peroxisomal beta oxidation of PA.OA (18:1c9) increases 18:0, 16:0 and 14:0. 18:0 increase might be dueto a partial inhibition of SCD1 with a consequent increase of 16:0 and14:0 as products of peroxisomal beta oxidation of 18:0.
Figure 7 Relationship between extent of triglyceride (TG) accumula-tion and caspases 3/7 activity. Regression lines represent the relation-ship between the two parameters after incubation of HepG2 cellsrespectively with palmitic acid (PA; �), PA + oleic acid (OA; �) and OA(�). Single points are the mean of three experiments that have inparallel evaluated TGs content and caspases activity. The slope valuesof PA vs PA + OA (t = 6.805; P < 0.0001)and OA vs PA + OA (t = -5.353;P < 0.0001) regression lines are significantly different.
Figure 8 Effect of palmitic acid (PA) and oleic acid (OA) on PPARg,SREBP-1 and PPARa gene expression. Expression was evaluated byreal time-polymerase chain reaction (PCR; for details see materials andmethods). Data are reported as mean � standard error of log2(RQ) nor-malized to the controls. *P < 0.05 vs control.
Fatty acids and apoptosis in hepatocytes M Ricchi et al.
836 Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

phosphorylation and activation of the serine/threonine kinaseAkt, a central regulator of glucose uptake, anabolic metabolismand antiapoptotic signals.34 We compared the effects of steatosisinduced by the different FAs, alone or in association, on insulin-induced phosphorylation of Akt (Fig. 9). Insulin caused a rapidand robust phosphorylation of Akt on Ser473, an activation-specific residue, in HepG2 cells. Steatosis induced by PA wasassociated with a marked decrease of insulin-induced Akt phos-phorylation, while in cells treated with OA Akt phosphorylationwas maintained. Interestingly, in cells exposed to both FA, anintermediate level of phosphorylation was detected. No differ-ences were found among different fatty acid treatments in theactivity of ERK1-2 (data not shown). Taken together, these datademonstrate that PA and OA differentially affect the early signal-ing pathways downstream of the insulin receptor.
DiscussionIn this in vitro model of steatogenesis, three different lines ofhepatocytes were exposed to increasing doses of FAs forcomparison purposes. The effects of administering OA, PA ortheir combination on TG accumulation, cell survival, FAscomposition, gene expression and insulin signalling wereexplored. Data showed that while the extent of steatosis was moresevere in cells treated with OA than in those exposed to PA,opposite effects of the two FAs were found on cell apoptosis. Thegreater steatosis extent observed following exposure to OA wasassociated with prominent PPARg and SREBP-1 activation. Incontrast, PA supplementation was associated with PPARa activa-tion. Moreover, PA, but not OA, impaired insulin signalling.Despite the higher steatosis extent resulting from incubation withthe two FAs combined, apoptosis rate and insulin signallingimpairment were lower than in cells treated with PA alone, indi-cating a protective effect of OA.
As expected, in our study the accumulation of TGs was propor-tional to the concentration of FAs in the culture medium. However,OA was a more powerful trigger for TG accumulation than PAirrespective of the cell line we used. This is a novel finding forhepatocytes and may represent a general property of OA, whichhas been previously reported to induce a more marked fat
accumulation also in other cell types, such as islet b cells.35 It istempting to associate the more effective steatogenic property ofOA with the specific pattern of gene expression (PPAR-gammaand SREBP-1) shown in the present study. In contrast, the lessersteatosis extent and increased apoptosis are due to PPAR-alphaactivation which results in enhanced b-oxidation and thus oxida-tive stress.36
The most intriguing finding of the present study is the dissocia-tion between the effects on steatosis and apoptosis of the differentFAs. Steatosis per se induces cell apoptosis.14,15 Other studies haveindicated a greater pro-apoptotic effect of PA as compared to OA,but few studies have examined the relationship between steatosisextent and apoptosis in hepatic cell cultures.20,28 The finding thatOA is less efficient in the induction of apoptosis is in agreementwith the recent work by Mahli et al. who, however, at variancewith our study, found no differences in the steatogenic effect ofOA and PA.20 This discrepancy might probably be accounted forby differences in the experimental conditions notably including thelower (0.2 mM in Mahli’s vs 0.66 mM in our study) FA concen-trations used.19 In this study, we have shown a correlation betweenTG accumulation and apoptosis with both OA and PA. However,the slope of this function was markedly different comparing thetwo FAs indicating that, for a given amount of TG accumulation,the effects on apoptosis were dramatically enhanced when cellswere treated with PA (Fig. 6).
A large number of molecular mechanisms have been implicatedin PA-associated apoptosis: ceramide production, NO synthesis,suppression of antiapoptotic factors such as Bcl-2,15 reactiveoxygen species generation,37 endoplasmic reticulum stress;37,38
nuclear factor-kB activation39 and decreased synthesis of cardio-lipin.40 In hepatocytes, steatosis may enhance the expression ofinflammatory cytokines, such as TNF-a, and increase apoptosisvia Fas receptor activation.16,19,27 A JNK stimulated mitochon-drial20 and a reticulum stress-mediated9,21 apoptotic pathways havebeen also reported. Death receptors are cell surface proteins, whichcan trigger apoptosis when bound by their ligands. Ribeiro &Cortez-Pinto41 observed enhanced expression of the death receptorFas in alcoholic hepatitis and tumor necrosis factor (TNF)receptor-1 in NASH. Others have identified enhanced Fas expres-sion in alcoholic and nonalcoholic steatohepatitis16,42 Thus steato-hepatitis may sensitize hepatocytes to extracellular death ligands(e.g. Fas ligand, tumor necrosis factor-alpha) promoting an extrin-sic pathway to apoptosis.19,27 NF-kB is a transcription factor thatcan upregulate both death receptors and ligands.43,44 NF-kB acti-vation in Kupffer cells or infiltrating monocytes is proinflamma-tory and induces the expression of death ligands such as TNFa.
FFAs can directly activate the IKK-k/ NF-kB pathway in hepa-tocytes via a lysosomal, cathepsin B– dependent mechanism.27
This pathway involves the translocation of Bax to lysosomes withsubsequent lysosomal destabilization and release of the cysteineprotease, cathepsin B, into the cytosol. This subsequently leads tothe activation of NF-kB, via IKK-k, and a subsequent increase inthe expression of TNFa apoptosis has increasingly been linked toinflammation and hepatic fibrogenesis.45–47
More recently, Gores’ group48 reported OA to sensitise hepato-cytes in vitro to to the death ligand TRAIL. Oleic acid led toupregulation of the cognate TRAIL receptor death receptor 5whose expression was enhanced in steatotic human liver samples.DR5 was responsible for FFA sensitisation to TRAIL killing medi-
Figure 9 Effect of fatty acids (FAs) on Akt phosphorylation. Akt phos-phorylation was evaluated in confluent, serum-starved (12 h) HepG2were treated with FAs, after stimulation with 100 nM of insulin for 15′and analyzed by Western blot using Phospho-Akt (Ser-473) Antibody.
M Ricchi et al. Fatty acids and apoptosis in hepatocytes
837Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

ated by a JNK dependent mechanism. Moreover, as previouslyreported, if Kupffer cells are activated they produce death ligands,including Fas ligand, and TNFa which, in turn, induce apoptosisin hepatocytes expressing death receptors,49 this might be also acondition in NASH patients. Given that inhibiting apoptosis maywell prevent the feared consequences of chronic liver disease,cirrhosis, portal hypertension, and liver failure thus drugs arebeing developed to block apoptosis.50
The cells’ ability to readily incorporate FAs into cytoplasmicTGs might serve as a protection against their pro-apoptoticeffects.51 The finding that OA resulted in a greater steatosis extentimplies that this FA is more readily incorporated into TG andtherefore is associated with less apoptosis than PA which was lesssteatogenic. Along these lines, the cytoprotective effect of OA invitro could thus be explained by this FA’s ability to promotechannelling of PA into TG synthesis, as demonstrated in culturedfibroblasts.51 The toxic effect of PA on hepatocytic cell culturescould be, at least, in part related to the amount of FFA associatedwith incubation with this FA. However, given that this explanationis based on in vitro indirect data, our hypothesis needs to beverified by further studies given that a recent report disproves theparadigm that FFAs are increased in human NAFLD.52 In additionto PA channelling into TG synthesis, changes in the composition ofthe intracellular FAs pool may represent an alternative strategy toprotect cells from PA-dependent apoptosis. The addition of PA tothe medium resulted in an increased proportion of the less dam-aging palmitoleic acid indicating the attempt to ‘detoxify’ PA. Thisdetoxification might be achieved through increased steroyl-CoAdesaturase 1 (D9) activity (Fig. 8), which protects fromPA-induced apoptosis.53
Our data provide in vitro evidence for the principle that TGaccumulation might be a defence mechanism against the toxicityof excess FFAs. Yamaguchi et al. recently reached the same con-clusion in the obese mouse model, wherein inhibited triglyceridesynthesis was associated with improved hepatic steatosis thoughto the expenses of worsened hepatic inflammatory and fibroticchanges.54 Observations that simple steatosis is rarely progressivein humans55–57 further confirm that FAs stored as inert triglyceridesare harmless unless associated to another injuring factor, such asIR, or oxidative stress, that may switch on the inflammatorycascade. Familial heterozygous hypobetalipoproteinemia repre-sents a naturally occurring model of human simple steatosis due togenetically impaired capacity to export VLDL from the hepato-cytes into the bloodstream. Of interest, in the absence of IR,steatosis in these individuals, although massive, does not appear tobe progressive.58 Studies have shown that PA impairs insulin sig-nalling via increased JNK activity leading, in turn, to phosphory-lation of insulin receptor substrate-1 and 2 at the inhibitory siteseventually leading to IR.59 Increased circulating FFAs concentra-tions represent an early correlate of peripheral IR, which may bedemonstrated to occur before the development of obesity inexperimental condition.12 In this connection, our data have shownthat PA administration, in itself, impaired insulin signalling whileOA partially restored it. This finding implies that the steatosisextent is not the only, nor perhaps the major, determinant ofimpaired insulin action. Indeed, rat models7,9 and human stud-ies5,6,8 consistently support the concentrations and the chemicalstructure of FAs to be fundamental players in the developmentand progression of NAFLD. In human NAFLD, FAs that are
incorporated in hepatic TG are mostly derived from the circulatingNEFA pool and thus reflect the adipose tissue FAs composition.60
Individuals consuming western-type high-fat diets show a serum/tissue predominance of PA with a relative reduction in OA.6,10,11 Inprinciple, characterization of FAs profile in the serum/liver mayassist in predicting those individuals with progressive/severeNAFLD.20 Our data also highlight that variations in the desaturaseactivity might have a role in the type of FAs stored in hepatic TGand thus in the pathogenesis of human NAFLD. Conversely, PAdeprivation and OA enrichment in diet might be a goal to prevent/treat the disease.
In conclusion, while a limitation of the current study is the useof three hepatocyte cultures of different origin but no data onprimary hepatocyte culture, our study showing that OA is moresteatogenic but less damaging than PA in hepatocyte cell culturesmay provide a biological clue useful to a better understanding ofanimal models of NAFLD. As far as human NAFLD is concerned,our data might suggest two major clinical implications/researchhypothesis. First, for the diagnosis of NASH, steatosis extentevaluated histologically might not be as relevant as its chemicalcomposition. Second, in agreement with recent studies envisagingfor saturated FAs a role of ‘second hit’,61 the chemical compositionof steatosis might help differentiate those non-progressive (‘inert’)forms of NAFLD from those that, due to their enrichment insaturated FAs, are at a substantial risk of progression. Thesehypotheses are worthy being tested in specific studies.
AcknowledgmentsWe thank Dorval Ganazzi for the assistance in statistical analysis.We are also grateful to Dr. Cristiana Bertolani and Nadia Navari,coworkers of Prof. Marra, for their contribution.
Part of these data have been presented in abstract form at theEuropean Association for the Study of the Liver, Wien, April 2006.
References1 Chawla A, Repa JJ, Evans RM, Mangelsdorf DJ. Nuclear receptors
and lipid physiology: opening the x-files. Science 2001; 294:1866–70.
2 Clarke SD. The multi-dimensional regulation of gene expression byfatty acids: polyunsaturated fats as nutrient sensors. Curr. Opin.Lipidol. 2004; 15: 13–18.
3 McGarry JD. Banting lecture 2001: dysregulation of fatty acidmetabolism in the etiology of type 2 diabetes. Diabetes 2002; 51:7–18.
4 Haslam DW, James WP. Obesity. Lancet 2005; 366: 1197–209.5 Nehra V, Angulo P, Buchman AL, Lindor KD. Nutritional and
metabolic considerations in the etiology of non-alcoholicsteatohepatitis. Dig. Dis. Sci. 2001; 46: 2347–52.
6 Musso G, Gambino R, De Michieli F et al. Dietary habits and theirrelations to insulin resistance and postprandial lipemia innonalcoholic steatohepatitis. Hepatology 2003; 37: 909–16.
7 Hernandez R, Martinez-Lara E, Canuelo A et al. Steatosis recoveryafter treatment with a balanced sunflower or olive oil-based diet:involvement of perisinusoidal stellate cells. World J. Gastroenterol.2005; 11: 7480–5.
8 Cortez-Pinto H, Jesus L, Barros H, Lopes C, Moura MC,Camilo ME. How different is the dietary pattern in non-alcoholicsteatohepatitis patients? Clin. Nutr. 2006; 25: 816–23.
Fatty acids and apoptosis in hepatocytes M Ricchi et al.
838 Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

9 Wang D, Wei Y, Pagliassotti MJ. Saturated fatty acids promoteendoplasmic reticulum stress and liver injury in rats with hepaticsteatosis. Endocrinology 2006; 147: 943–51.
10 De Almeida IT, Cortez-Pinto H, Fidalgo G, Rodrigues D,Camilo ME. Plasma total and free fatty acids composition in humannon-alcoholic steatohepatitis. Clin. Nutr. 2002; 21: 219–23.
11 Videla LA, Rodrigo R, Araya J, Poniachik J. Oxidative stress anddepletion of hepatic long-chain polyunsaturated fatty acids maycontribute to nonalcoholic fatty liver disease. Free Radic. Biol. Med.2004; 37: 1499–507.
12 Samuel VT, Liu ZX, Qu X et al. Mechanism of hepatic insulinresistance in non-alcoholic fatty liver disease. J. Biol. Chem. 2004;279: 32345–53.
13 Browning JD, Horton JD. Molecular mediators of hepatic steatosisand liver injury. J. Clin. Invest. 2004; 114: 147–52.
14 Canbay A, Gieseler RK, Gores GJ, Gerken G. The relationshipbetween apoptosis and non-alcoholic fatty liver disease: anevolutionary cornerstone turned pathogenic. Z. Gastroenterol. 2005;43: 211–17.
15 Unger RH, Orci L. Lipoapoptosis: its mechanism and its diseases.Biochim. Biophys. Acta 2002; 1585: 202–12.
16 Feldstein AE, Canbay A, Angulo P et al. Hepatocyte apoptosis andfas expression are prominent features of human nonalcoholicsteatohepatitis. Gastroenterology 2003; 125: 437–43.
17 Schmitz-Peiffer C, Craig DL, Biden TJ. Ceramide generation issufficient to account for the inhibition of the insulin-stimulated PKBpathway in C2C12 skeletal muscle cells pretreated with palmitate. J.Biol. Chem. 1999; 274: 24202–10.
18 Baylin A, Kabagambe EK, Siles X, Campos H. Adipose tissuebiomarkers of fatty acid intake. Am. J. Clin. Nutr. 2002; 76:750–7.
19 Feldstein AE, Canbay A, Guicciardi ME, Higuchi H, Bronk SF,Gores GJ. Diet associated hepatic steatosis sensitizes to Fasmediated liver injury in mice. J. Hepatol. 2003; 39: 978–83.
20 Malhi H, Bronk SF, Werneburg NW, Gores GJ. Free fatty acidsinduce JNK-dependent hepatocyte lipoapoptosis. J. Biol. Chem.2006; 281: 12093–101.
21 Wei Y, Wang D, Topczewski F, Pagliassotti MJ. Saturated fatty acidsinduce endoplasmic reticulum stress and apoptosis independently ofceramide in liver cells. Am. J. Physiol. Endocrinol. Metab. 2006;291: E275–81.
22 Javitt NB. Hep G2 cells as a resource for metabolic studies:lipoprotein, cholesterol, and bile acids. FASEB J. 1990; 4:161–8.
23 Dashti N, Wolfbauer G. Secretion of lipids, apolipoproteins, andlipoproteins by human hepatoma cell line, HepG2: effects of oleicacid and insulin. J. Lipid Res. 1987; 28: 423–36.
24 Gutierrez-Ruiz MC, Bucio L, Souza V, Gomez JJ, Campos C,Carabez A. Expression of some hepatocyte-like functional propertiesof WRL-68 cells in culture. In Vitro Cell. Dev. Biol. Anim. 1994;30A: 366–71.
25 Nakabayashi H, Taketa K, Yamane T, Miyazaki M, Miyano K,Sato J. Phenotypical stability of a human hepatoma cell line, HuH-7,in long-term culture with chemically defined medium. Gann 1984;75: 151–8.
26 Okamoto Y, Tanaka S, Haga Y. Enhanced GLUT2 gene expressionin an oleic acid-induced in vitro fatty liver model. Hepatol. Res.2002; 23: 138–44.
27 Feldstein AE, Werneburg NW, Canbay A et al. Free fatty acidspromote hepatic lipotoxicity by stimulating TNF-alpha expressionvia a lysosomal pathway. Hepatology 2004; 40: 185–94.
28 Gomez-Lechon MJ, Donato MT, Martinez-Romero A, Jimenez N,Castell JV, O’Connor JE. A human hepatocellular in vitro model toinvestigate steatosis. Chem. Biol. Interact. 2007; 165: 106–16.
29 Ricchi M, Bertolotti M, Anzivino C et al. 17Beta-estradiol preventscytotoxicity from hydrophobic bile acids in HepG2 and WRL-68 cellcultures. J. Gastroenterol. Hepatol. 2006; 21: 894–901.
30 Folch J, Lees M, Sloane Stanley GH. A simple method for theisolation and purification of total lipides from animal tissues. J. Biol.Chem. 1957; 226: 497–509.
31 Banni S, Carta G, Contini MS et al. Characterization of conjugateddiene fatty acids in milk, dairy products, and lamb tissues. J. Nutr.Biochem. 1996; 7: 150–5.
32 Melis MP, Angioni E, Carta G et al. Characterization of conjugatedlinoleic acid and its metabolites by RP-HPLC with diode arraydetector. Eur. J. Lipid Sci. Technol. 2001; 103: 617–21.
33 Ip C, Banni S, Angioni E et al. Conjugated linoleic acid-enrichedbutter fat alters mammary gland morphogenesis and reduces cancerrisk in rats. J. Nutr. 1999; 129: 2135–42.
34 Whiteman EL, Cho H, Birnbaum MJ. Role of Akt/protein kinase Bin metabolism. Trends Endocrinol. Metab. 2002; 13: 444–51.
35 Cnop M, Hannaert JC, Hoorens A, Eizirik DL, Pipeleers DG. Inverserelationship between cytotoxicity of free fatty acids in pancreaticislet cells and cellular triglyceride accumulation. Diabetes 2001; 50:1771–7.
36 Li Z, Berk M, McIntyre TM, Gores GJ, Feldstein AE. Thelysosomal-mitochondrial axis in free fatty acid-induced hepaticlipotoxicity. Hepatology 2008; 47: 1495–503.
37 Listenberger LL, Ory DS, Schaffer JE. Palmitate-induced apoptosiscan occur through a ceramide-independent pathway. J. Biol. Chem.2001; 276: 14890–5.
38 Karaskov E, Scott C, Zhang L, Teodoro T, Ravazzola M, Volchuk A.Chronic palmitate but not oleate exposure induces endoplasmicreticulum stress, which may contribute to INS-1 pancreatic beta-cellapoptosis. Endocrinology 2006; 147: 3398–407.
39 Rakatzi I, Mueller H, Ritzeler O, Tennagels N, Eckel J.Adiponectin counteracts cytokine- and fatty acid-induced apoptosisin the pancreatic beta-cell line INS-1. Diabetologia 2004; 47:249–58.
40 Ostrander DB, Sparagna GC, Amoscato AA, McMillin JB,Dowhan W. Decreased cardiolipin synthesis corresponds withcytochrome c release in palmitate-induced cardiomyocyte apoptosis.J. Biol. Chem. 2001; 276: 38061–7.
41 Ribeiro PS, Cortez-Pinto H, Sola S et al. Hepatocyte apoptosis,expression of death receptors and activation of NfkappaB in the liverof non-alcoholic and alcoholic steatohepatitis patients. Am. J.Gastroenterol. 2004; 99: 1718–19.
42 Natori S, Rust C, Stadheim LM et al. Hepatocyte apoptosis is apathologic feature of human alcoholic hepatitis. J. Hepatol. 2001;34: 248–53.
43 Ghosh S, May MJ, Kopp EB. NF-kappa B and Rel proteins:evolutionarily conserved mediators of immune responses. Annu. Rev.Immunol. 1998; 16: 225–60.
44 Green DR. Death and NF-kappaB in T cell activation: life at theedge. Mol. Cell 2003; 11: 551–2.
45 Faouzi S, Burckhardt BE, Hanson JC et al. Anti-Fas induces hepaticchemokines and promotes inflammation by an NfkappaB-independent, caspase-3-dependent pathway. J. Biol. Chem. 2001;276: 49077–82.
46 Canbay A, Guicciardi ME, Higuchi H et al. Cathepsin B inactivationattenuates hepatic injury and fibrosis during cholestasis. J. Clin.Invest. 2003; 112: 152–9.
47 Canbay A, Taimr P, Torok N et al. Apoptotic body engulfment by ahuman stellate cell line is profibrogenic. Lab. Invest. 2003; 83:655–63.
48 Malhi H, Barreyro FJ, Isomoto H, Bronk SF, Gores GJ. Free fattyacids sensitise hepatocytes TRAIL mediated cytotoxicity. Gut 2007;56: 1124–31.
M Ricchi et al. Fatty acids and apoptosis in hepatocytes
839Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd

49 Canbay A, Feldstein AE, Higuchi H et al. Kupffer cell engulfmentof apoptotic bodies stimulates death ligand and cytokine expression.Hepatology 2003; 38: 1188–98.
50 Feldstein A, Gores GJ. Steatohepatitis and apoptosis: therapeuticimplications. Am. J. Gastroenterol. 2004; 99: 1718–19.
51 Listenberger LL, Han X, Lewis SE et al. Triglyceride accumulationprotects against fatty acid-induced lipotoxicity. Proc. Natl. Acad. Sci.USA 2003; 100: 3077–82.
52 Puri P, Baillie RA, Wiest MM et al. A lipidomic analysis ofnonalcoholic fatty liver disease. Hepatology 2007; 46: 1081–90.
53 Warensjo E, Ohrvall M, Vessby B. Fatty acid composition andestimated desaturase activities are associated with obesity andlifestyle variables in men and women. Nutr. Metab. Cardiovasc. Dis.2006; 16: 128–36.
54 Yamaguchi K, Yang L, McCall S et al. Inhibiting triglyceridesynthesis improves hepatic steatosis but exacerbates liver damageand fibrosis in obese mice with nonalcoholic steatohepatitis.Hepatology 2007; 45: 1366–74.
55 Matteoni CA, Younossi ZM, Gramlich T, Boparai N, Liu YC,McCullough AJ. Nonalcoholic fatty liver disease: a spectrum ofclinical and pathological severity. Gastroenterology 1999; 116:1413–19.
56 Harrison SA, Neuschwander-Tetri BA. Nonalcoholic fatty liverdisease and nonalcoholic steatohepatitis. Clin. Liver Dis. 2004; 8:861–79.
57 Adams LA, Lymp JF, St Sauver J et al. The natural history ofnonalcoholic fatty liver disease: a population-based cohort study.Gastroenterology 2005; 129: 113–21.
58 Lonardo A, Lombardini S, Scaglioni F et al. Hepatic steatosis andinsulin resistance: does etiology make a difference? J. Hepatol.2006; 44: 190–6.
59 Solinas G, Naugler W, Galimi F, Lee MS, Karin M. Saturated fattyacids inhibit induction of insulin gene transcription by JNK-mediatedphosphorylation of insulin-receptor substrates. Proc. Natl. Acad. Sci.USA 2006; 103: 16454–9.
60 Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD,Parks EJ. Sources of fatty acids stored in liver and secreted vialipoproteins in patients with nonalcoholic fatty liver disease. J. Clin.Invest. 2005; 115: 1343–51.
61 Gentile CL, Pagliassotti MJ. The role of fatty acids in thedevelopment and progression of nonalcoholic fatty liver disease.J. Nutr. Biochem. 2008; 19: 567–76.
Fatty acids and apoptosis in hepatocytes M Ricchi et al.
840 Journal of Gastroenterology and Hepatology 24 (2009) 830–840 © 2009 The Authors
Journal compilation © 2009 Journal of Gastroenterology and Hepatology Foundation and Blackwell Publishing Asia Pty Ltd
