different growth sensitivity to climate of the conifer...
TRANSCRIPT

ORIGINAL PAPER
Different growth sensitivity to climate of the conifer Juniperusthurifera on both sides of the Mediterranean Sea
Lucía DeSoto & Filipa Varino & José P. Andrade &
Celia M. Gouveia & Filipe Campelo & Ricardo M. Trigo &
Cristina Nabais
Received: 15 October 2013 /Revised: 17 February 2014 /Accepted: 23 February 2014# ISB 2014
Abstract Mediterranean plants cope with cold wet wintersand dry hot summers, with a drought gradient from northwestto southeast. Limiting climatic conditions have become morepronounced in the last decades due to the warming trend andrainfall decrease. Juniperus thurifera L., a long-lived conifertree endemic to the western Mediterranean region, has adisjunct distribution in Europe and Africa, making it a suitablespecies to study sensitivity to climate in both sides of theMediterranean Basin. Tree-ring width chronologies were builtfor three J. thurifera stands at Spain (Europe) and three inMorocco (Africa) and correlated with monthly temperatureand precipitation. The temporal stability of climate-growthrelationships was assessed using moving correlations; thedrought effect on growth was calculated using the monthlystandardized precipitation-evapotranspiration index (SPEI) atdifferent temporal scales. In the wettest stands, increasingspring temperature and summer precipitation enhancedgrowth, while in the driest stands, growth was enhanced byhigher spring precipitation and lower summer temperature.The climate-growth correlations shifted during the twentiethcentury, especially since the 1970s. Particularly noticeable isthe recent negative correlation with previous autumn andwinter precipitation in the wettest stands of J. thurifera, prob-ably related with an effect of cloud cover or flooding oncarbon storage depletion for next year growth. The drieststands were affected by drought at long time scales, while
the wettest stands respond to drought at short time scales. Thisreveals a different strategy to cope with drought conditions,with populations from drier sites able to cope with shortperiods of water deficit.
Keywords Climate change . Dendrochronology . Juniper .
Mediterranean basin . Tree ring
Introduction
Mediterranean plants, and particularly those growing in con-tinental areas, cope with limiting climatic conditions charac-terized by cold wet winters and dry hot summers (Mitrakos1980). In general, the Mediterranean climate is influenced byboth maritime and continental air masses with significantorigin differences being under the influence of tropical andmid-latitude climate dynamics (Lionello et al. 2006). Despitethe large inter-annual variability, most of the precipitation fallsfrom October to March, with a peak between December andFebruary, when the mid-latitude cyclone belt reaches its south-ernmost position (e.g. Trigo 2006).
Recent modelling studies predict that during the 21st cen-tury the Mediterranean Basin will suffer from an increasingdrying trend due to both enhanced evaporation and decreasedprecipitation (e.g. Giorgi and Lionello 2008; Mariotti et al.2008). In fact, a tendency towards more frequent droughtevents has been already detected for large areas of the Med-iterranean region driven, to a large extent, by climate change(Hoerling et al. 2012; Sousa et al. 2011). Additionally, warmerdays have been rising dramatically, particularly near the Med-iterranean coast, exacerbating the dry episodes (Trenberthet al. 2007). A warmer winter can induce an earlier growingseason, but the decreasing precipitation associated with highertemperature during spring and summer will reduce water
Electronic supplementary material The online version of this article(doi:10.1007/s00484-014-0811-y) contains supplementary material,which is available to authorized users.
L. DeSoto (*) : J. P. Andrade : F. Campelo : C. NabaisCentre for Functional Ecology, University of Coimbra, Apdo. 3046,3001-401 Coimbra, Portugale-mail: [email protected]
F. Varino :C. M. Gouveia :R. M. TrigoInstituto Dom Luiz (IDL), University of Lisboa, Lisbon, Portugal
Int J BiometeorolDOI 10.1007/s00484-014-0811-y

availability and increase resource depletion (Corcuera et al.2004; Galiano et al. 2012; Piovesan and Schirone 2000).
Long-lived trees have to deal with climatic fluctuations ona long-term basis, witnessing and recording those fluctuationsin tree-ring width, a fingerprint of the adjustment of trees toclimate conditions. In fact, during the last decades, the effectsof climate change have been registered in the growth re-sponses of Mediterranean trees (e.g. Carrer et al. 2007; Jumpet al. 2006; Linares et al. 2009). Although the phenotypicplasticity may be enough to ensure the survival of a speciesacross a wide range of climatic conditions, the current rate ofclimate change may exceed the rate of evolutionary changes,genetic adaptation or migration in Mediterranean forest trees(Klein et al. 2013). Thus, it is increasingly important toinvestigate the response of trees to climate change conditions.Within this framework, tree species that occur on a broadgeographical range and along climatic gradients are importantmodels to understand the adjustment of trees to climaticconditions.
Geographical patterns in climate-growth relationships havebeen detected using tree-ring chronologies of Mediterraneantrees across wide distribution ranges (e.g. Carrer et al. 2007;DeSoto et al. 2012; Di Filippo et al. 2007). Juniperus thuriferaL. (Spanish juniper) is a long-lived conifer tree endemic to thewestern Mediterranean region with a disjunct distribution insouth western Europe and North Africa, being more abundantin North Central Spain and the High Atlas Mountains inMorocco (Gauquelin et al. 1999). The Strait of Gibraltar couldhave functioned as an effective barrier against gene flowbetween the African and European populations reflected inmorphological differences in shoots, leaves and cones(Boratyński et al. 2013; Terrab et al. 2008). Thus, J. thuriferais a valuable species to study the sensitivity to climate at abroader geographical scale, at both sides of the MediterraneanBasin. Regarding environmental conditions, both the Iberianand Moroccan regions are strongly influenced by the Atlanticflows and correlated with tele-connections as the NorthernAtlantic Oscillation (NAO) interactions (Born et al. 2010;Knippertz et al. 2003; Trigo 2004). The Iberian Peninsula ischaracterized by a northwest–southeast gradient of precipita-tion, warm temperature and summer drought (Esteban-Parraet al. 1998; Rodriguez-Puebla 1998). Morocco is mainlycharacterized by a semiarid climate, especially in the southof the Atlas Mountains region, while in the western coast, theclimate is more sub-humid as a result of Atlantic moisturetransport and orographic precipitation (Born et al. 2010;Knippertz et al. 2003). These differences in climate conditionsand geographical isolation may be reflected in a diversegrowth sensitivity and response to climate change ofJ. thurifera from both sides of the Mediterranean Basin. Wehypothesize that (1) the Iberian and North African populationsof J. thurifera have different climate-growth relationships, andconsequently, (2) their growth response to shifts in climatic
conditions along the 20th century is also different, namely, the(3) short- and long-term impact of water deficit on treegrowth.
Materials and methods
Study species
J. thurifera L. (Cupressaceae) is a long-lived conifer tree (upto 600 years, DeSoto et al. 2012). It is able to maintain itsphotosynthetic activity all year round (Gimeno et al. 2012),although secondary growth is restricted to the most favourableperiods in spring and early autumn (Camarero et al. 2010).The species is endemic to continental areas of the westernMediterranean at elevations ranging from 200 to 3,400 m andwas a dominant species during cold stages of the Pleistocene(Carrion et al. 2004). Its range is over a disjunct distribution insouth western Europe and North Africa (Terrab et al. 2008),being abundant in Spain (ca. 200,000 ha) and Morocco(30,000 ha), and sparse in the French and Italian Alps(1,200 ha), Pyrenees (300 ha), Corsica (500 ha) and Algeria(50 ha) (Gauquelin et al. 1999).
Study sites and sampling procedure
We selected three populations in the Iberian Peninsula (Lunain the Cantabrian Range, Soria and Monegros Desert) andthree in Morocco (Armd, Oukaimeden and Ourika, Fig. 1).Between November 2010 and June 2011, 20–26 trees withoutexternal signs of bole dieback were randomly selected in eachpopulation. Height and perimeter at breast height were mea-sured in the main stem (Table 1). We collected at least twocores per tree at ca. 1.3 m above ground using an incrementborer. Cores were dried and mechanically polished with aseries of successively finer grades of sandpaper until thexylem cellular structure was clearly visible under magnifica-tion. After visual cross-dating, tree-ring widths were measuredto the nearest 0.001 mm using a sliding-stage micrometre(LINTAB TM., Rinntech Inc., GE) interfaced with theTSAP-Win programme. The individual tree-ring series werestatistically compared with a site master chronology using theCOFECHA programme and checked for dating accuracy(Holmes 1983; Grissino-Mayer 2001). All series with missingor false rings were corrected when possible or otherwisediscarded.
Dendrochronological methods
The raw data from the ring-width series correctly synchro-nized within each population were standardized with the R-packages “dplR” (Bunn 2008) and “detrendeR” (Campeloet al. 2011) in the R environment (R Development Core Team
Int J Biometeorol

2010, http://cran.r-project.org). The raw series were first fit toa cubic smoothing spline with 55-year period, which wasflexible enough to preserve high-frequency climatic informa-tion by reducing the non-climatic variance such as age/size
trends (Cook and Peters 1981; Cook and Kairiukstis 1990).For each population, the obtained indices were averaged on ayear-by-year basis using a bi-weight robust mean, and themean series were subjected to autoregressive modelling to
a
b
c
Fig. 1 a Distribution range of J. thurifera (white dots, according toGBIF), CRU grid points selected (black dots) and sampling sites(crosses). b Climate diagrams of monthly precipitation (dashed line)and monthly mean temperature (solid line) of the sampling sites,
indicating annual precipitation, mean temperature (range) from 1901–2006 using CRU data and drought period (black area). c) Mean waterbalance (black line) and percentiles (grey lines)
Int J Biometeorol
obtain population residual chronologies of pre-whitenedgrowth indices.
For each population, several descriptive statistics werecalculated for the six populations using the raw tree-ringwidths (mean tree-ring width (MW), AC and msx) and theresidual chronologies (rbt, rwt, and EPS) (Cook andKairiukstis 1990; Fritts 1976; Table 2). The first-order auto-correlation (AC) expresses the correlation of the tree-ringwidth of a particular year with the tree-ring width of theprevious year, and mean sensitivity (msx) is the average ofchange from each tree-ring width measured to the followingtree-ring width. Mean between-trees correlation (rbt) is a mea-sure of the similarity in growth patterns among trees, andmean within-trees correlation (rwt) is a measure of the simi-larity in growth patterns among cores within a tree. Expressedpopulation signal (EPS) is a measure of the statistical qualityof a chronology based on a finite number of trees as comparedwith a hypothetically perfect chronology based on an infinitenumber of trees. The reliable period of the chronologies wasconsidered when EPS values were above 0.85, a thresholdwidely used in dendrochronological analysis (Wigley et al.1984). Thus, the reliable common period to compare allpopulations was from 1961 to 2009. Growth differences forthe common period of all populations were tested using gen-eralized linear models (see Appendix A1).
Climatic data and analysis
We used the TS 3.1 dataset produced by the Climate ResearchUnit (CRU, Mitchell and Jones 2005) that provides high-resolution (0.5°×0.5°) grids of monthly climatic variablesfor the period of 1901–2006. We used monthly mean
temperature and precipitation variables of the CRU grid pointsnearest the sampling sites (Fig. 1a). Due to the distance andaltitude differences between the study sites and the availablemeteorological stations, the CRU dataset was considered themost suitable database comparing to meteorological stationdatabases because the latter were less homogeneous and moredistant from the sampling sites (see Appendix A2 for details).Nonetheless, we have to take into account that CRU dataunderestimates and overestimates precipitation in the HighAtlas and in the southern part of the High Atlas region,respectively. This is consistent with some studies that ob-served this difference as a consequence of the poor orographicresolution in the CRU dataset (Adam et al. 2006; Born et al.2008). In this sense, Fink et al. (2010) have claimed thatprecipitation in the High Atlas Mountains may reach over800 mm per year, while CRU data of the nearest grid pointnever exceed 400 mm. Particularly, in our study area, theobserved annual precipitation ranged between 400 and500 mm (Alaoui Haroni et al. 2008 and http://www.marocmeteo.ma/fr/climat), being higher than the CRUvalues (370 mm).
We decomposed CRU climatic series of precipitation andtemperature for each site into seasonal and trend componentsto derive the long-term climatic patterns (see Appendix A3).Then, we computed moving medians of 30-year period for themonthly climatic data to detect the monthly climatic trends onthese variables over the century.
For each population, we computed Pearson correlationcoefficients between the residual chronology and monthlyprecipitation and mean monthly temperature of the commonperiod (1961–2006). A temporal window of 17 months wasselected to identify limiting climatic factors from July of the
Table 1 Study sites characteristics
Group Site Latitude(N)
Longitude(W)
Elevation(m a.s.l.)
Age(years)a
Perimeter(cm)
Height(m)
Spain Luna 42° 52′ 33″ 5° 50′ 10″ 1,380 113 100 7.0
(81–146) (58–170) (3.7–10)
Soria 41° 47′ 20″ 2° 49′ 37″ 1,200 120 88 5.4
(66–203) (48–210) (5.8–10.8)
Monegros 41° 48′ 39″ 0° 32′ 42″ 490 74 81 7.5
(44–94) (33–161) (4.2–15.1)
Morocco Ourika 31° 11′ 59″ 7° 52′ 56″ 2,600 198 287 7.5
(67–442) (125–480) (4.8–12.1)
Oukaimeden 31° 11′ 60″ 7° 54′ 00″ 2,550 293 226 6.5
(57–525) (110–709) (4.1–9.5)
Armd 31° 8′ 59″ 7° 57′ 26″ 2,100 170 222 6.2
(54–260) (116–358) (3.4–8.8)
Site name, geographical latitude and longitude, elevation, mean (range) values for tree age, perimeter at breast height and height are displayed. Sites’locations are displayed in Fig. 1a Age was estimated as the maximum number of rings counted at 1.3 m
Int J Biometeorol

previous year to November of the year of tree-ring formation.To analyse the instability of the climate-growth relationship,moving Pearson correlation coefficients were calculated be-tween these climatic variables and the population chronolo-gies considering 30-year intervals for the period 1905–2006 inthe Iberian populations and 1945–2006 in Moroccan popula-tions. For this analysis, we did not use the populations with theshortest tree-ring series because of the insufficient time periodto compute the moving correlations (see Appendix B1). Al-though we could only achieve EPS values above 0.82 forOukaimeden for the period from 1945 to 2009, we still usedthis period according to the dating reliability.
A potential bias may arise in the interpretation of the resultsof moving correlations because of the effect of tree age/size onthe response to climatic conditions. A previous study showedhigher sensitivity to summer drought in younger J. thuriferatrees (Rozas et al. 2009); hence, a reduction of climate sensi-tivity with time could be expected as trees age. To reduce thispotential bias, the population chronologies were built with apool of tree-ring series of different tree ages.
A composite analysis was performed to detect the directionof stronger climate-growth relationships. First, we determinedthe growth-anomalous years, i.e. the extreme growing yearswith values of the population residual chronologies above(under) the 75th (25th) percentiles, and the anomalies oftemperature and precipitation of these extreme growing years,i.e. the difference between the observed climatic data and themean for the period. Second, we calculated a subsequent10,000 random permutations of the data to get the 10 and90 % confidence intervals (see Appendix A3).
We used a recently developed multi-scalar drought index,the standardized precipitation-evapotranspiration index(SPEI; Vicente-Serrano et al. 2010), to analyse the effect ofdrought severity on tree growth in the studied populations.The SPEI is more suitable over semiarid regions than standarddrought indices, such as PDSI and standardized precipitationindex (SPI) at the global and regional scales, because it
combines precipitation and temperature data to describe dryconditions considering evapotranspiration processes (Vicente-Serrano et al. 2010; Vicente-Serrano et al. 2012; Vicente-Serrano et al. 2013). The SPEI data are calculated using themonthly difference between precipitation and potential evapo-transpiration (available at http://sac.csic.es/spei/database.html). Positive and negative SPEI values correspond,respectively, to wet and dry conditions. Similarly to thewidely used SPI, the SPEI can be computed at different timescales to characterize the duration and intensity of droughts. Insemiarid regions, at shorter time scales, dry periods are shortand occur at high frequency, while at longer time scalesdroughts last longer, but are less frequent (Vicente-Serranoet al. 2010; see Appendix A4). We computed monthly SPEIfor 24 different scales, from 1 month to the previous24 months, to be compared with radial growth at a widerange of time scales for the six studied populations similarlyto the approach used in previous studies (e.g. Vicente-Serranoet al. 2013). For the common period (1961–2006), we calcu-lated the correlation between the population residual chronol-ogies and SPEI for the different scales using Pearson coeffi-cients. Positive coefficients indicate better growing conditionswithout drought, whereas negative coefficients indicategrowth increment during dry conditions.
Results
Climatic characterization and trends
Monthly mean values of the CRU data (1901–2006) showedthat among the Iberian sampling sites, Luna was the moistestand coldest site and Monegros the warmest and driest(Fig. 1b). Maximum precipitation occurred in November andDecember in Luna, while in Soria and Monegros maximumoccurred in May. The sampling sites in the High Atlas Moun-tains in Morocco (Armd, Oukaimeden and Ourika) were drier
Table 2 Summary of the dendrochronological statistics calculated for the ring-width chronologies of each site for the common period (1961–2009)
Country Site Trees/radii Raw tree-ring data Population chronology
MW±SD (mm) msx AC rbt rwt EPS
Spain Luna 13/23 0.67±0.24a 0.26a 0.49a 0.41 0.60 0.92
Soria 13/28 0.69±0.25a 0.27a 0.58b 0.36 0.60 0.91
Monegros 12/18 1.50±0.84b 0.31b 0.62b 0.31 0.59 0.85
Morocco Ourika 10/14 1.71±0.62b 0.29ab 0.54ab 0.34 0.53 0.85
Oukaimeden 9/17 1.07±0.49c 0.35b 0.60b 0.34 0.58 0.86
Armd 9/16 0.97±0.49c 0.42c 0.58ab 0.38 0.58 0.88
Significant differences of least-square means among sampling sites are indicated with different letters (see Appendix A1 in Supplementary Material)
MWmean ring width, SD standard deviation, AC first-order autocorrelation, msx mean sensitivity, rbt mean between-trees correlation, rwt mean within-trees correlation, EPS expressed population signal
Int J Biometeorol

than Monegros with maximum precipitation from February toApril. The water balance was computed using precipitationand evapotranspiration data for every site and indicated thatMonegros and the High Atlas are under water deficit almostall year round (Fig. 1c).
Climatic conditions were not stationary during the lastcentury; particularly, since the 1970s, precipitation has beendecreasing in the Iberian Peninsula and Morocco (Fig. 2). Atthe Iberian sampling sites, in general, monthly temperatureincreased while precipitation decreased in winter and earlysummer. Nonetheless, an increase in May precipitation wasobserved in Luna. In the High Atlas Mountains, monthlytemperature showed two warmer periods, one from 1930 to1950 and a second since 1975. Precipitation has been decreas-ing in spring and late autumn but increasing in late winter.
Tree-growth parameters
Tree rings of Juniperus species are particularly difficult todistinguish because of the slow growth rates and high fre-quency of missing and wedging rings, which makes the datingprocess quite challenging, particularly in drier areas such as
Morocco (Wils et al. 2009). Although we were able to roughlyestimate the tree age for each population, with the Moroccanpopulations being older (ca. 500 years old) than the Iberianpopulations (ca. 200 years old), the reliable dating period onlycorresponded from 1945 to 2009 in the Moroccan populationsand from 1920 to 2009 in the Iberian populations (seeAppendix B1). To compare all populations, the reliable periodwas only considered from 1961 to 2009, with EPS valuesabove 0.85. Mean tree-ring width differed between popula-tions (χ5=43.51, P<0.001, 66 trees, Table 2, Fig. 3a). Ourikaand Monegros populations showed wider mean tree-ringwidth (ca. 1.6 mm) that significantly differed from Armdand Oukaimeden populations (ca. 1 mm), and Soria and Luna(ca. 0.7 mm). Mean sensitivity differed among populationsranging from 0.26 to 0.42 (χ5=52.65, P<0.001, 66 trees,Table 2), and AC slightly differed among populations andranged between 0.49 and 0.62 (χ5=11.37, P=0.045, 66 trees,Table 2, see Appendix B1 for details). The population chro-nologies from Morocco were highly correlated among them(r>0.66, P<0.001, n=49), and population chronology ofLuna correlated with Soria chronology (r=0.62, P<0.001,n=49), whereas non-significant correlations were found
Fig. 2 Moving mediananomalies for a 30-year period ofmonthly precipitation (a) andmean temperature (b) for eachsite. Y-axis corresponds to themiddle of the interval, and monthsare represented on the x-axis
Int J Biometeorol

between the chronologies of the Iberian and Moroccan popu-lation, and between Monegros and the other populations (seethe residual population chronologies in Fig. 3b).
Climate sensitivity
All the populations showed significant correlations betweenradial growth and climatic parameters in April, with Luna andSoria populations showing a positive correlation with temper-ature, with Monegros and all the Moroccan populations show-ing a positive correlation with precipitation (Table 3). Treegrowth in Luna and Soria was negatively correlated with winterand early spring precipitation and positively correlated withsummer precipitation (Table 3). Tree growth in Monegrosshowed a positive correlation with winter, spring and summerprecipitation (Table 3). In the Moroccan populations, growthwas positively correlated with winter precipitation (Ourika andArmd) and negatively correlated with temperature in spring(Armd) and summer (Ourika and Armd; Table 3).
The composite analysis between tree-ring width indexes andmonthly climatic data shows the correspondence between ex-treme growing years (wide or narrow tree rings) with anoma-lous climate variables (either positive or negative) during thecommon period (1961–2006, Fig. 4). In all the three Iberianpopulations, wider tree rings were associated with higher pre-cipitation and lower temperature in June (Fig. 4a–c). In Lunaand Soria, narrow rings were mainly associated with highprecipitation in winter or low precipitation in May and June(Fig. 4a, b), whereas wide (narrow) rings in Monegros wererelated to high (low)winter precipitation (Fig. 4c). InMoroccanpopulations, larger tree rings were associated with higher pre-cipitation in previous September and current April and withhigher temperature in previous November (Fig. 4d–f). Smallertree rings were mainly associated with lower precipitation inprevious November and current April and higher temperaturein spring and early summer (Fig. 4d–f).
The responses of growth to climate shifted during the lastcentury (Fig. 5). In Luna and Soria, the negative correlationwith winter and early spring precipitation only became
a b
Fig. 3 a Tree-ring width (mean±SE) series. b Population chronologiesfor the common period 1961–2009, of the six J. thurifera samplingpopulations belonging to Spain (Luna, Monegros and Soria) and
Morocco (Ourika, Oukaimeden and Armd). Site characteristics are shownin Table 1
Int J Biometeorol

significant since the 1970s–1980s (Fig. 5a, b). In Luna, thepositive correlation between growth and June precipitationwas stable during the last century, while in Soria it onlybecame significant since the 1970s (Fig. 5a, b). Both popula-tions showed a positive correlation with temperature in Au-gust but it was only significant until the 1940s–1950s(Fig. 5a, b). Luna population showed a temporal shift in thepositive correlations with temperature, with significant corre-lations in December from the 1920s to the 1940s, in Februaryfrom the 1950s to the 1970s and in April from the 1970sonwards (Fig. 5a). The negative correlation with Octobertemperature was only significant since the 1970s (Fig. 5a).In Soria, the negative correlation with June temperature wasonly significant since the 1980s (Fig. 5b). The temporal shiftsbetween growth and climate for the Moroccan populationswere only analysed for a shorter time scale because the reliabledating period was smaller (1945–2006). The positive correla-tion with the precipitation in April was only significant sincethe 1980s (Fig. 5c, d). In Armd, the positive correlation withJanuary precipitation was only significant until the 1980s,while the negative correlation with the temperature in Junewas more or less stable from the 1960s onwards (Fig. 5c, d).
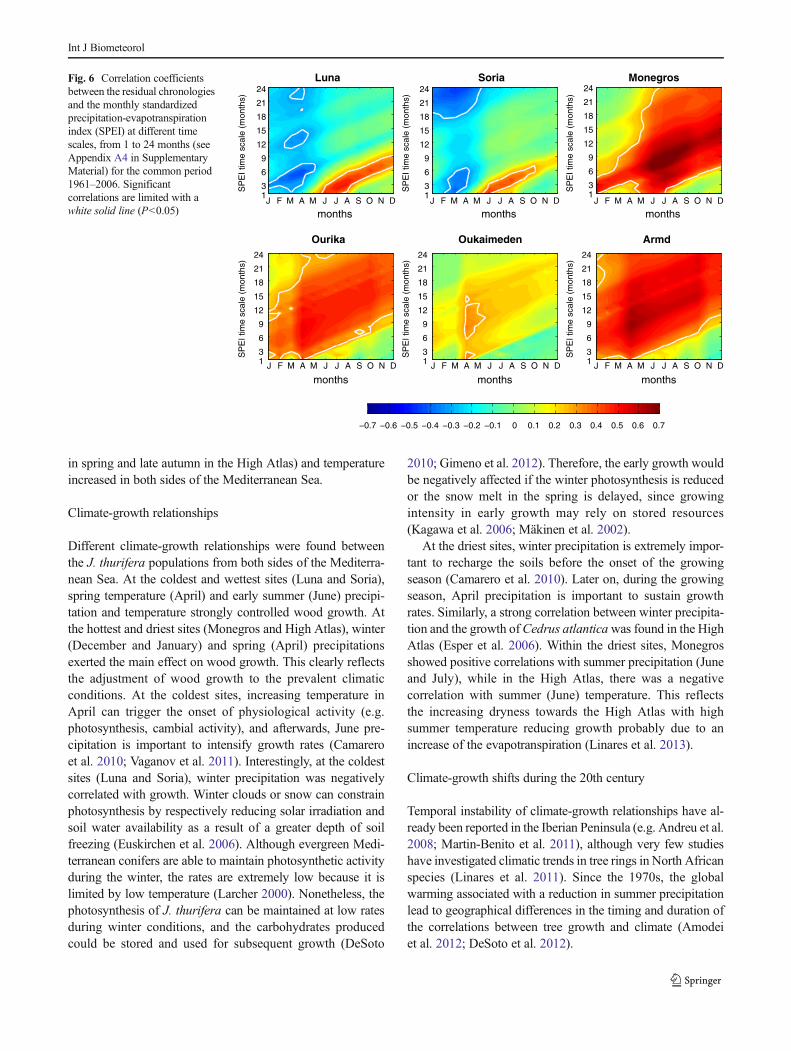
Two different patterns of correlation between SPEI andresidual chronologies were found. In wetter sites (Luna andSoria), a negative response to SPEI at a 6-month time scalewas observed during February to May, whereas a positiveresponse between 1- to 6-month time scale was observed fromJune to October (Fig 6). In the semiarid Iberian region(Monegros), growth was positively correlated with SPEI at6-month time scale during winter and between 1 to 21 monthsduring summer. In Moroccan populations, growth was posi-tively correlated with SPEI at different time scales, 6 to11 months in Oukaimedem but only during April and May,1 to 16 months in Ourika and 1 to 24 months in Armd.
Discussion
Climate differences and trends
Different climatic conditions were found among the samplingsites. In the Iberian Peninsula, Luna was the moistest andTa
ble3
Pearsoncorrelationvalues
betweenpopulatio
nradial-growth
indexesandtemperature
(T)andprecipitatio
n(P)variablesforthecommon
period
(1961–2006)
Country
Site
Variable
Previousyear
Current
year
Jul
Aug
Sep
Oct
Nov
Dec
Jan
Feb
Mar
Apr
May
Jun
Jul
Aug
Sep
Oct
Nov
Spain
Luna
T−0
.06
0.00
−0.13
−0.37
0.18
−0.04
−0.04
0.20
−0.05
0.42
−0.06
−0.37
−0.05
0.01
0.14
−0.09
0.12
P0.00
−0.06
0.17
0.27
−0.27
−0.29
−0.17
−0.23
−0.29
−0.01
0.19
0.43
0.35
0.15
−0.11
0.06
0.05
Soria
T0.08
−0.06
−0.10
−0.13
0.01
−0.02
−0.12
0.20
−0.01
0.35
−0.14
−0.33
−0.07
−0.08
0.07
0.01
0.14
P0.10
−0.09
0.11
0.09
−0.03
−0.20
−0.22
−0.34
−0.31
0.00
0.26
0.52
0.12
0.15
−0.19
0.03
−0.14
Monegros
T0.11
−0.18
−0.04
0.14
0.05
0.03
−0.01
0.16
−0.08
0.27
−0.07
−0.24
−0.19
−0.05
0.00
0.00
−0.04
P0.24
0.22
−0.15
−0.19
0.08
0.33
0.45
0.01
0.15
0.31
0.42
0.63
0.41
−0.08
−0.09
−0.02
0.09
Morocco
Ourika
T0.11
0.06
−0.02
−0.09
−0.15
−0.11
−0.20
−0.07
−0.28
−0.17
−0.08
−0.34
−0.03
−0.11
0.11
−0.02
0.18
P−0
.11
0.03
0.12
0.12
0.22
−0.06
0.30
0.27
−0.03
0.38
0.02
−0.13
0.14
0.02
−0.10
0.13
−0.07
Oukaimeden
T0.02
−0.05
−0.29
−0.14
−0.02
0.03
−0.19
−0.02
−0.25
−0.06
0.06
−0.18
0.13
0.04
0.12
0.03
0.10
P−0
.06
−0.10
0.20
0.07
0.20
−0.03
0.08
−0.01
−0.01
0.36
−0.11
−0.22
0.02
−0.07
0.00
−0.05
0.01
Arm
dT
0.11
0.04
−0.18
−0.06
−0.08
0.09
−0.03
0.03
−0.31
−0.19
−0.04
−0.33
−0.14
−0.08
−0.02
−0.03
0.15
P−0
.16
0.10
0.22
0.24
0.21
0.07
0.32
0.24
0.16
0.32
0.07
−0.08
0.03
0.08
−0.11
0.12
−0.19
Significant
coefficients(P<0.05)arein
bold
andP<0.01
arealso
initalic
�Fig. 4 Monthly composites calculated for the common period (1961–2006) between the ring-width indexes and the monthly climatic data fromgridded CRU database for the Iberian (a, b, c) and Moroccan (d, e, f)populations. The climate data corresponded tomean temperature and totalprecipitation over a 17-month window. Months before and during tree-ring formation are abbreviated by lower- and upper-case letters,respectively. Monthly composites of precipitation and temperatureanomalies of years with tree-ring growth lower (higher) than the 25th(75th) percentile are displayed in grey (black). Dotted lines represent the10th and 90th percentiles of 10,000 random samples. Observed valuesgreater or lower than the envelopes, built with the percentiles, indicatesignificant relationship
Int J Biometeorol

(a)
(b)
(c)
(d)
(e)
(f)
Int J Biometeorol

coldest site and Monegros the warmest and driest, accordingto the Iberian northwest–southeast drought gradient due to theincreasing temperature and decreasing precipitation (Esteban-Parra et al. 1998; Rodriguez-Puebla 1998). The precipitationpattern differed among sites. In Luna, the maximum precipi-tation occurred in November and December related to low-pressure systems that reach the western region of Iberia Pen-insula during the winter (Trigo 2006; Trigo et al. 2004).However, in Soria and Monegros, maximum precipitationoccurred in May as a consequence of spring and summerinstability (Ninyerola et al. 2003; Gestal-Souto et al. 2010).The High Atlas (Morocco) precipitation is strongly correlatedwith westerly flows and orographic sub-humid effects on the
southern slope of the mountains during the summer (Bornet al. 2010; Knippertz et al. 2003). Thus, the sampling sites inMorocco (Armd, Ourika and Oukaimeden) were the driestwith maximum precipitation from February to April, being,jointly with Monegros, under water deficit almost all yearround. In fact, the High Atlas Mountains and Monegros areclassified asmid-latitude dry semiarid climate according to theKöppen and Geiger climate classification, whereas Soria be-longs to marine-mild winter climate and Luna to coastalMediterranean climate (Kottek et al. 2006). In the last century,changes in climate at a global scale have been occurring in theIberian Peninsula andMorocco. Since the 1970s, precipitationhas decreased (in winter and early summer in the Iberian sites,
(a)
(b)
(c)
(d)
Fig. 5 Temporal shifts of therelationship between the ring-width indexes and the monthlyclimatic data from gridded CRUdatabase for the Iberian (a, b) andMoroccan (c, d) populations. Theclimate data corresponded tomean temperature and totalprecipitation over a 17-monthwindow and the period 1915–2006 (Iberia) and 1945–2006(Morocco). Months before andduring tree-ring formation areabbreviated by lower- and upper-case letters, respectively. Movingaverage correlations werecalculated for 30-year intervals inthe whole studied periods, andeach correlation corresponds tothe middle of the interval.Significant correlations aredisplayed as black circles(P<0.05)
Int J Biometeorol
in spring and late autumn in the High Atlas) and temperatureincreased in both sides of the Mediterranean Sea.
Climate-growth relationships
Different climate-growth relationships were found betweenthe J. thurifera populations from both sides of the Mediterra-nean Sea. At the coldest and wettest sites (Luna and Soria),spring temperature (April) and early summer (June) precipi-tation and temperature strongly controlled wood growth. Atthe hottest and driest sites (Monegros and High Atlas), winter(December and January) and spring (April) precipitationsexerted the main effect on wood growth. This clearly reflectsthe adjustment of wood growth to the prevalent climaticconditions. At the coldest sites, increasing temperature inApril can trigger the onset of physiological activity (e.g.photosynthesis, cambial activity), and afterwards, June pre-cipitation is important to intensify growth rates (Camareroet al. 2010; Vaganov et al. 2011). Interestingly, at the coldestsites (Luna and Soria), winter precipitation was negativelycorrelated with growth. Winter clouds or snow can constrainphotosynthesis by respectively reducing solar irradiation andsoil water availability as a result of a greater depth of soilfreezing (Euskirchen et al. 2006). Although evergreen Medi-terranean conifers are able to maintain photosynthetic activityduring the winter, the rates are extremely low because it islimited by low temperature (Larcher 2000). Nonetheless, thephotosynthesis of J. thurifera can be maintained at low ratesduring winter conditions, and the carbohydrates producedcould be stored and used for subsequent growth (DeSoto
2010; Gimeno et al. 2012). Therefore, the early growth wouldbe negatively affected if the winter photosynthesis is reducedor the snow melt in the spring is delayed, since growingintensity in early growth may rely on stored resources(Kagawa et al. 2006; Mäkinen et al. 2002).
At the driest sites, winter precipitation is extremely impor-tant to recharge the soils before the onset of the growingseason (Camarero et al. 2010). Later on, during the growingseason, April precipitation is important to sustain growthrates. Similarly, a strong correlation between winter precipita-tion and the growth ofCedrus atlanticawas found in the HighAtlas (Esper et al. 2006). Within the driest sites, Monegrosshowed positive correlations with summer precipitation (Juneand July), while in the High Atlas, there was a negativecorrelation with summer (June) temperature. This reflectsthe increasing dryness towards the High Atlas with highsummer temperature reducing growth probably due to anincrease of the evapotranspiration (Linares et al. 2013).
Climate-growth shifts during the 20th century
Temporal instability of climate-growth relationships have al-ready been reported in the Iberian Peninsula (e.g. Andreu et al.2008; Martin-Benito et al. 2011), although very few studieshave investigated climatic trends in tree rings in North Africanspecies (Linares et al. 2011). Since the 1970s, the globalwarming associated with a reduction in summer precipitationlead to geographical differences in the timing and duration ofthe correlations between tree growth and climate (Amodeiet al. 2012; DeSoto et al. 2012).
Luna
months
SP
EI t
ime
scal
e (m
onth
s)
J F M A M J J A S O N D13
6
9
12
15
18
21
24Soria
months
SP
EI t
ime
scal
e (m
onth
s)
J F M A M J J A S O N D13
6
9
12
15
18
21
24Monegros
months
SP
EI t
ime
scal
e (m
onth
s)
J F M A M J J A S O N D13
6
9
12
15
18
21
24
Ourika
months
SP
EI t
ime
scal
e (m
onth
s)
J F M A M J J A S O N D13
6
9
12
15
18
21
24
Oukaimeden
months
SP
EI t
ime
scal
e (m
onth
s)
J F M A M J J A S O N D13
6
9
12
15
18
21
24
Armd
months
SP
EI t
ime
scal
e (m
onth
s)
J F M A M J J A S O N D13
6
9
12
15
18
21
24
−0.7 −0.6 −0.5 −0.4 −0.3 −0.2 −0.1 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Fig. 6 Correlation coefficientsbetween the residual chronologiesand the monthly standardizedprecipitation-evapotranspirationindex (SPEI) at different timescales, from 1 to 24 months (seeAppendix A4 in SupplementaryMaterial) for the common period1961–2006. Significantcorrelations are limited with awhite solid line (P<0.05)
Int J Biometeorol

The growth of J. thurifera became more sensitive to winterprecipitation, with negative correlations, only in the northernpart of the Peninsula and especially since the 1970s (DeSotoet al. 2012). Negative responses to winter precipitation werealso observed in other conifer species from Mediterraneanmesic sites where the positive water balance in this seasonindicates the absence of water restrictions (Martin-Benito et al.2013; Rozas et al. 2011; Todaro et al. 2007; but see Boginoand Bravo 2008). In Luna and Soria, the negative correlationwith precipitation in autumn and winter has only becomesignificant since the 1970s–1980s. Recent studies show anincrease in total cloud cover during the 20th century in theIberian Peninsula (Sanchez-Lorenzo et al. 2012), although wedid not find a positive trend of cloud cover during the lastcentury (see Appendix B2). Autumn and winter cloudinessmay generate a reduction on sun irradiance limiting carbohy-drate production and reserves and thus affecting tree growth inthe next season (DeSoto 2010; Gimeno et al. 2012). However,in Luna and Soria, a negative correlation between growth andcloud cover was only found in March and April, respectively(see Appendix B2), as also observed in previous studies(Gimeno et al. 2012; Rozas and Olano 2013). An alternativeexplanation can be related with an increase in the frequency ofextreme climatic events (drought and floods) observed insome areas of the Mediterranean (Alpert et al. 2002). Extremewater regimes affect the mineral nutrition of trees by alteringthe microbial activity, the interaction with other plants and thecarbon balance (Kreuzwieser and Gessler 2010). Particularly,flooding cause anoxic conditions and thus oxygen deprivationin the root system of trees changing carbon metabolism andallocation (Kreuzwieser et al. 2004), exerting a negative effecton tree growth.
In Luna, growth sensitivity to temperature shifted along the20th century, showing positive correlations between the 1920sand 1940s with December, between the 1950s and 1970s withFebruary and since the 1970s with April. This can reflect thewarming trend and the limitations imposed by temperature incolder sites, such as Luna. Until the 1940s, warmer wintertemperature would probably increase the photosynthetic ratespromoting an increase of carbohydrate storage (DeSoto 2010;Körner 2003) that can be used in the following growing season.With the general warming trend along the 20th century, wintertemperature is probably no longer an important limiting factor,and the regulation of the temperature on the tree physiology onlybecomes significant within the growing season. Since the 1970s,the increasing April temperature could have favoured an earlierstart of the growing season in Luna because at high altitude treegrowth is mainly constrained by temperature particularly at thebeginning of the growing season (Carrer and Urbinati 2006).
Interestingly, until the 1940s–1950s, the temperature inAugust also showed a positive correlation with growth, whilethe negative correlation with temperature in June only becamesignificant since the 1980s, both in Luna and Soria. Again, this
is probably related with the warming trend in the IberianPeninsula, associated with a decrease of winter and early sum-mer precipitation along the 20th century, intensifying droughtconditions during the summer. It seems that since the 1980s,high temperature in June shortened the growing season lengthwith a consequent reduction of tree-ring width (Camarero et al.2010). Until the 1940s–1950s, there was probably a secondgrowth burst after the summer drought quiescent stage, indi-cating a less severe post-summer water deficit.
The negative effect of recurrent droughts and rising temper-ature since the 1970s was also observed in C. atlantica from theMiddleAtlas (Linares et al. 2013; Linares et al. 2011). Due to theshort reliable chronology of J. thurifera from the High Atlas, therange of climate-growth temporal shifts in Morocco is muchsmaller. Nonetheless, the recent positive correlation betweengrowth and April precipitation reinforces the importance ofspring precipitation.
Impact of drought
The correlation of SPEI with growth of J. thurifera changed atdifferent time scales in mesic and xeric populations. In north-ern mesic populations of the Iberian Peninsula, Luna andSoria, the high SPEI values in winter (and early spring) areassociated with low growth during 6 months. This result wasconsistent with the observed negative correlation with precip-itation, and thus, the effect of water surplus, cloudiness orsnow may compromise carbohydrates synthesis and short-term growing conditions. Summer drought (low SPEI values)exerted a negative effect that could last for 6 months mainlyaffecting latewood growth in the mesic populations (Martin-Benito et al. 2013).
In the Iberian driest population, Monegros, long-lastingwinters and summer droughts negatively affected growthindicating the importance of precipitation winter to refill watercontent in soil and the high relevance of dry summers inreducing the growing period. In the High Atlas Mountains,the impact of cumulative dry periods in winter and springcould last for more than 2 years in Armd and Ourika, indicat-ing that droughts recorded for 2 years can exert a negativeeffect on growth. In these drier populations, the reduction ofwater availability might penalize both the current and the nextgrowing year (Pasho et al. 2011).
At short time scales, dry periods occur with a high frequen-cy and short duration, while droughts become less frequentbut more prolonged at long time scales (Vicente-Serrano2006). Therefore, J. thurifera populations in semiarid andsub-humid regions respond to drought at long time scales,probably because they are able to withstand isolated periods ofwater deficit, whereas in humid regions, plants respond todrought at short time scales likely because of their low adap-tation to water shortages (Vicente-Serrano et al. 2013). This
Int J Biometeorol

result points out the high resilience to drought of Mediterra-nean plants from drier sites.
Conclusions
Tree growth is largely driven by the climatic conditions thataffect tree physiology and thus wood formation. As expected,growth responses of J. thurifera under dry climatic conditions,such as in Monegros and High Atlas, are mainly driven byprecipitation, whereas in more cold and humid climatic con-ditions, such as in Luna and Soria, temperature is a moreimportant driver. The adjustment of growth to prevalent cli-matic conditions depends on the plastic response of morpho-logical and physiological traits that ultimately depend on thegenetic framework. Thus, a species response to the climaticconditions can not only result from plastic adjustments butalso from genetic differences that already imply natural adap-tation. In the case of J. thurifera, the Strait of Gibraltar couldact as an effective barrier against gene flow between Africanand European populations (Terrab et al. 2008), and the differ-ent climate-growth relationships could be the consequence oflocal adaptation to these environments. However, the responseto climate is quite similar in the population ofMonegros, fromthe Iberian Peninsula, and in the Moroccan populations fromthe High Atlas, with similar climatic conditions but on differ-ent sides of the Mediterranean Sea.
Historical land use can also affect climate responses oftrees. J. thurifera woodlands are a consequence of historicalland use, mainly livestock browsing (Carrion et al. 2004;Gauquelin et al. 1999). In the Mediterranean Basin, thesewoodlands are currently changing into mixed stands becauseof the land-use abandonment (DeSoto et al. 2010). However,browsing pressure remains high in Morocco increasing soilerosion and degradation of the J. thurifera woodlandsresulting in a loss of natural vegetation cover (Barbero1990). The lack of vegetation could compromise the wateravailability, particularly in the High Atlas Mountains wheresurface runoff could exacerbate the effect of drought.
Taking into account the climatic changes especially notice-able since the 1970s, the general increase of temperature canbenefit wood growth in Luna and Soria because growth isquite responsive to temperature in spring (April). However, ifincreasing temperature in winter is responsible for an increasein cloudiness, or an increase of flooding episodes, this cannegatively affect wood growth. If winter and summer precip-itation is decreasing in the Iberian Peninsula, this will mainlyaffect tree growth in Monegros. Winter precipitation seems tobe increasing in the High Atlas, with strong benefits for thesoil recharge. Nonetheless, if spring precipitation is decreas-ing, this can have a negative impact on wood growth. Furtherresearch is needed to explore whether the genetic variation can
partially explain the climate response of a species and thus itsability to adjust growth to climate change.
Acknowledgments We especially thank M Alifriqui and MJ Silva fortheir help in field and laboratory work. Climatic data from Morocco andSpain were kindly provided by A Fink and R Schuster and by the SpanishAgencia Estatal de Meteorología (AEMet), respectively. We also thankthe comments by two anonymous referees that improved a previousversion of the manuscript. This work was supported by Fundação paraa Ciência e a Tecnologia (FCT) with the project MEDIATIC(PTDC/AAC-CLI/103361/2008). F Campelo and L DeSoto were sup-ported by postdoctoral scholarships from FCT (SFRH/BPD/47822/2008and SFRH/BPD/70632/2010, respectively) and F Varino and JPAndradeby two research fellowships of the project MEDIATIC.
References
Adam JC, Clark EA, Lettenmaier DP, Wood EF (2006) Correction ofglobal precipitation products for orographic effects. J Clim 19:15–38
Alaoui Haroni S, Alifriqui M, Simonneaux V (2008) Recent dynamics ofthe wet pastures at Oukaimeden plateau (High Atlas mountains,Morroco). Biodivers Conserv 18:167–189. doi:10.1007/s10531-008-9465-6
Alpert P, Baharad A, Ramis C et al (2002) The paradoxical increase ofMediterranean extreme daily rainfall in spite of decrease in totalvalues. Geophys Res Lett 29:29–32
Amodei T, Guibal F, Fady B (2012) Relationships between climate andradial growth in black pine (Pinus nigra Arnold ssp. salzmannii(Dunal) Franco) from the south of France. Ann For Sci. doi:10.1007/s13595-012-0237-9
Andreu L, Planells O, Gutiérrez E et al (2008) Climatic significance oftree-ring width and δ13C in a Spanish pine forest network. Tellus B60:771–781. doi:10.1111/j.1600-0889.2008.00370.x
Barbero M (1990) Changes and disturbances of forest ecosystems causedby human activities in the western part of the mediterranean basin.Vegetatio 7:257–173. doi:10.1007/BF00042952
Bogino SM, Bravo F (2008) Growth response of Pinus pinaster Ait. toclimatic variables in central Spanish forests. Ann For Sci 65:506.doi:10.1051/forest
Boratyński A, Jasińska AK, Marcysiak K et al (2013) Morphologicaldifferentiation supports the genetic pattern of the geographic struc-ture of Juniperus thurifera (Cupressaceae). Plant Syst Evol 299:773–784. doi:10.1007/s00606-013-0760-7
Born K, Christoph M, Fink AH et al (2008) Moroccan climate in thepresent and future: combined view from observational data andregional climate scenarios. In: Hötzl H (ed) Zereini F. Clim.Chang. Water Resour. Middle East North Africa, Springer BerlinHeidelberg, pp 29–45
Born K, Fink AH, Knippertz P (2010)Meteorological processes influenc-ing the weather and climate of Morocco. In: Speth P, Christoph M,Diekkrüger B (eds) Impacts Glob. Chang. Hydrol. Cycle WestNorthwest Africa. Springer, Heidelberg, Germany, pp 150–163
Bunn AG (2008) A dendrochronology program library in R (dplR).Dendrochronologia 26:115–124. doi:10.1016/j.dendro.2008.01.002
Camarero JJ, Olano JM, Parras A (2010) Plastic bimodal xylogenesis inconifers from continental Mediterranean climates. New Phytol 185:471–480. doi:10.1111/j.1469-8137.2009.03073.x
Campelo F, García-González I, Nabais C (2011) detrendeR—a graphicaluser interface to process and visualize tree-ring data using R.Dendrochronologia 30:57–60. doi:10.1016/j.dendro.2011.01.010
Int J Biometeorol

Carrer M, Urbinati C (2006) Long-term change in the sensitivity of tree-ring growth to climate forcing in Larix decidua. New Phytol 170:861–871. doi:10.1111/j.1469-8137.2006.01703.x
Carrer M, Nola P, Eduard JL et al (2007) Regional variability of climate?Growth relationships in Pinus cembra high elevation forests in theAlps. J Ecol 95:1072–1083. doi:10.1111/j.1365-2745.2007.01281.x
Carrion J, Yll E, Willis K, Sanchez P (2004) Holocene forest history ofthe eastern plateaux in the Segura Mountains (Murcia, southeasternSpain). Rev Palaeobot Palynol 132:219–236. doi:10.1016/j.revpalbo.2004.07.002
Cook ER, Kairiukstis L (1990) Methods of dendrochronology: applica-tions in the environmental sciences. Kluwer Academic Publishers.Dordrecht, The Netherlands
Cook ER, Peters K (1981) The smoothing spline: a new approach tostandardizing forest interior tree-ring width series for dendroclimaticstudies. Tree-Ring Bull 4:45–53
Corcuera L, Camarero JJ, Gil-Pelegrin E (2004) Effects of a severedrought on Quercus ilex radial growth and xylem anatomy. Trees-Struct Funct 18:83–92. doi:10.1007/s00468-003-0284-9
DeSoto L (2010) Global change effect on the dioecious tree Juniperusthurifera in the Iberian Peninsula. University of Valladolid, DoctoralDissertation, 196
DeSoto L, Olano JM, Rozas V, De la CruzM (2010) Release of Juniperusthuriferawoodlands from herbivore-mediated arrested succession inSpain. Appl Veg Sci 13:15–25. doi:10.1111/j.1654-109X.2009.01045.x
DeSoto L, Camarero JJ, Olano JM, Rozas V (2012) Geographicallystructured and temporally unstable growth responses of Juniperusthurifera to recent climate variability in the Iberian Peninsula. Eur JFor Res 131:905–917. doi:10.1007/s10342-011-0564-7
Di Filippo A, Biondi F, Čufar K et al (2007) Bioclimatology of beech(Fagus sylvatica L.) in the Eastern Alps: spatial and altitudinalclimatic signals identified through a tree-ring network. J Biogeogr34:1873–1892. doi:10.1111/j.1365-2699.2007.01747.x
Esper J, Büntgen U, Frank DC et al (2006) Multiple tree-ring parametersfrom Atlas cedar (Morocco) and their climatic signal. In: Heinrich I,Gärtner H, Monbaron M, Schleser G (eds) TRACE—Tree RingsArchaeol. Climatol. Ecol. Vol. 4 Proc. DENDROSYMPOSIUM2005. Schriften des Forschungszentrums Jülich, Reihe Umwelt,Fribourg, Switzerland, pp 46–55
Esteban-Parra MJ, Rodrigo FS, Castro-Diez Y (1998) Spatial and tem-poral patterns of precipitation in Spain for the period 1880–1992. IntJ Cl imato l 18:1557–1574. doi :10 .1002/ (SICI)1097-0088(19981130)18:14<1557::AID-JOC328>3.0.CO;2-J
Euskirchen ES, McGuire AD, Kicklighter DWet al (2006) Importance ofrecent shifts in soil thermal dynamics on growing season length,productivity, and carbon sequestration in terrestrial high-latitudeecosystems. Glob Chang Biol 12:731–750. doi:10.1111/j.1365-2486.2006.01113.x
Fink AH, ChristophM, BornK et al (2010) Climate. Impacts Glob ChangHydrol Cycle West Northwest Africa pp 54–58
Fritts HC (1976) Tree-rings and climate. Academic, LondonGaliano L, Martínez-Vilalta J, Sabaté S, Lloret F (2012) Determinants of
drought effects on crown condition and their relationship withdepletion of carbon reserves in a Mediterranean holm oak forest.Tree Physiol 32:478–489. doi:10.1093/treephys/tps025
Gauquelin T, Bertaudiere V, Montes N et al (1999) Endangered stands ofthuriferous juniper in the western Mediterranean basin: ecologicalstatus, conservation and management. Biodivers Conserv 8:1479–1498
Gestal-Souto L, Martinez A, Rios-Entenza A (2010) O papel das augasfreáticas nas interaccións terra-atmosfera, no clima e no reximepluviométrico da Península Ibérica. Revista Real AcademiaGalega de Ciencias 29:89–128
Gimeno TE, Camarero JJ, Granda E et al (2012) Enhanced growth ofJuniperus thurifera under a warmer climate is explained by a
positive carbon gain under cold and drought. Tree Physiol 32:326–336. doi:10.1093/treephys/tps011
Giorgi F, Lionello P (2008) Climate change projections for theMediterranean region. Glob Planet Change 63:90–104. doi:10.1016/j.gloplacha.2007.09.005
Grissino-Mayer HD (2001) Evaluating cross-dating accuracy: a manualand tutorial for the computer program COFECHA. Tree-Ring Res57:205–221
HoerlingM, Eischeid J, Perlwitz J et al (2012) On the increased frequencyof Mediterranean drought. J Clim 25:2146–2161. doi:10.1175/JCLI-D-11-00296.1
Holmes RL (1983) Computer-assisted quality control in tree-ring datingand measurement. Tree-Ring Bull 43:69–78
Jump AS, Hunt JM, Peñuelas J (2006) Rapid climate change-relatedgrowth decline at the southern range edge of Fagus sylvatica.Glob Chang Biol 12:2163–2174. doi:10.1111/j.1365-2486.2006.01250.x
Kagawa A, Sugimoto A, Maximov TC (2006) 13CO2 pulse-labelling ofphotoassimilates reveals carbon allocation within and between treerings. Plant Cell Environ 29:1571–1584. doi:10.1111/j.1365-3040.2006.01533.x
Klein T, Di Matteo G, Rotenberg E et al (2013) Differential ecophysio-logical response of a major Mediterranean pine species across aclimatic gradient. Tree Physiol 33:26–36. doi:10.1093/treephys/tps116
Knippertz P, Christoph M, Speth P (2003) Long-term precipitation vari-ability inMorocco and the link to the large-scale circulation in recentand future climates. Meteorol Atmos Phys 83:67–88. doi:10.1007/s00703-002-0561-y
Kottek M, Grieser J, Beck C et al (2006) World map of the Köppen-Geiger climate classification updated. Meteorol Zeitschrift 15:259–263. doi:10.1127/0941-2948/2006/0130
Kreuzwieser J, Gessler A (2010) Global climate change and tree nutrition:influence of water availability. Tree Physiol 1221–1234. doi:10.1093/treephys/tpq055
Kreuzwieser J, Papadopoulou E, Rennenberg H (2004) Interaction offlooding with carbon metabolism of forest trees. Plant Biol 6:299–306. doi:10.1055/s-2004-817882
Larcher W (2000) Temperature stress and survival ability ofMediterranean sclerophyllus. Plant Biosyst 134:279–295
Linares JC, Camarero JJ, Carreira JA (2009) Interacting effects of chang-es in climate and forest cover on mortality and growth of thesouthernmost European fir forests. Glob Ecol Biogeogr 18:485–497. doi:10.1111/j.1466-8238.2009.00465.x
Linares JC, Taïqui L, Camarero JJ (2011) Increasing drought sensitivityand decline of Atlas Cedar (Cedrus atlantica) in the MoroccanMiddle Atlas forests. Forests 2:777–796. doi:10.3390/f2030777
Linares JC, Taïqui L, Sangüesa-Barreda G et al (2013) Age-relateddrought sensitivity of Atlas cedar (Cedrus atlantica) in theMoroccan Middle Atlas forests. Dendrochronologia 31:88–96. doi:10.1016/j.dendro.2012.08.003
Lionello P, Malanotte-Rizzoli P, Boscolo R (2006) Mediterranean climatevariability. Elsevier, Amsterdam
Mäkinen H, Nöjd P, Kahle H-P et al (2002) Radial growth variation ofNorway spruce (Picea abies (L.) Karst.) across latitudinal andaltitudinal gradients in central and northern Europe. For EcolManage 171:243–259
Mariotti A, Zeng N, Yoon J-H et al (2008) Mediterranean water cyclechanges: transition to drier 21st century conditions in observationsand CMIP3 simulations. Environ Res Lett 3:044001. doi:10.1088/1748-9326/3/4/044001
Martin-Benito D, Kint V, del Río M et al (2011) Growth responses ofWest-Mediterranean Pinus nigra to climate change are modulated bycompetition and productivity: past trends and future perspectives.For Ecol Manage 262:1030–1040. doi:10.1016/j.foreco.2011.05.038
Int J Biometeorol

Martin-Benito D, BeeckmanH, Cañellas I (2013) Influence of drought ontree rings and tracheid features of Pinus nigra and Pinus sylvestris ina mesic Mediterranean forest. Eur J For Res 132:33–45. doi:10.1007/s10342-012-0652-3
Mitchell TD, Jones PD (2005) An improved method of constructing adatabase of monthly climate observations and associated high-resolution grids. Int J Climatol 25:693–712. doi:10.1002/joc.1181
Mitrakos KA (1980) A theory for Mediterranean plant life. Acta Oecol 1:245–252
Ninyerola M, Pons X, Roure JM et al. (2003) Atles Climàtics deCatalunya. CD-ROM. Servei Meteorològic de Catalunya yDepartament de Medi Ambient de la Generalitat de Catalunya.ISBN: 84-932860-5-2.
Pasho E, Camarero JJ, de Luis M, Vicente-Serrano SM (2011) Impacts ofdrought at different time scales on forest growth across a wideclimatic gradient in north-eastern Spain. Agric For Meteorol 151:1800–1811. doi:10.1016/j.agrformet.2011.07.018
Piovesan G, Schirone B (2000) Winter North Atlantic oscillation effectson the tree rings of the Italian beech (Fagus sylvatica L.). Int JBiometeorol 44:121–127
Rodriguez-Puebla C (1998) Spatial and temporal patterns of annualprecipitation variability over the Iberian Peninsula. Int J Climatol5:1097–1316. doi:10.1002/(SICI)1097-0088(19980315)18:3<299::AID-JOC247>3.0.CO;2-L
Rozas V, Olano JM (2013) Environmental heterogeneity andneighbourhood interference modulate the individual response ofJun i p e ru s t hu r i f e r a t r e e - r i n g g row th t o c l ima t e .Dendrochronologia 1–9. doi:10.1016/j.dendro.2012.09.001
Rozas V, DeSoto L, Olano JM (2009) Sex-specific, age-dependent sen-sitivity of tree-ring growth to climate in the dioecious tree Juniperusthurifera. New Phytol 182:687–697. doi:10.1111/j.1469-8137.2009.02770.x
Rozas V, Zas R, García-González I (2011) Contrasting effects of wateravailability on Pinus pinaster radial growth near the transitionbetween the Atlantic and Mediterranean biogeographical regionsin NW Spain. Eur J For Res 130:959–970. doi:10.1007/s10342-011-0494-4
Sanchez-Lorenzo A, Calbó J, Wild M (2012) Increasing cloud cover inthe 20th century: review and new findings in Spain. Clim Past 8:1199–1212. doi:10.5194/cp-8-1199-2012
Sousa PM, Trigo RM, Aizpurua P et al (2011) Trends and extremes ofdrought indices throughout the 20th century in the Mediterranean.Nat Hazards Earth Syst Sci 11:33–51. doi:10.5194/nhess-11-33-2011
Terrab A, Schönswetter P, Talavera S et al (2008) Range-widephylogeography of Juniperus thurifera L., a presumptive keystonespecies of western Mediterranean vegetation during cold stages of
the Pleistocene. Mol Phylogenet Evol 48:94–102. doi:10.1016/j.ympev.2008.03.018
Todaro L, Andreu L, Dalessandro C et al (2007) Response of Pinusleucodermis to climate and anthropogenic activity in the NationalPark of Pollino (Basilicata, Southern Italy). Biol Conserv 137:507–519. doi:10.1016/j.biocon.2007.03.010
Trenberth KE, Jones PD, Ambenje P et al (2007) Observations: surfaceand atmospheric climate. In: Solomon S, Qin D, Manning M (eds)Clim. Chang. 2007 Phys. Sci. Basis. Contrib. Work. Gr. I to FourthAssess. Rep. Intergov. Panel Clim. Chang. Cambridge UniversityPress, Cambridge, pp 235–336
Trigo R (2004) O Impacto da Oscilação do Atlântico Norte no caudal dosrios ibéricos Intenacionais. Física de la Tierra 16:185–196
Trigo IF (2006) Climatology and interannual variability of storm-tracks inthe Euro-Atlantic sector: a comparison between ERA-40 andNCEP/NCAR reanalyses. Clim Dyn 26:127–143. doi:10.1007/s00382-005-0065-9
Trigo RM, Pozo-Vázquez D, Osborn TJ et al (2004) North Atlanticoscillation influence on precipitation, river flow and water resourcesin the Iberian Peninsula. Int J Climatol 24:925–944. doi:10.1002/joc.1048
Vaganov EA, Anchukaitis KJ, Evans MN (2011) How well understoodare the processes that create dendroclimatic records? A mechanisticmodel of the climatic control on conifer tree-ring growth dynamics.In: Hughes MK, Swetnam TW, Diaz HF (eds) DendroclimatologyProg. Prospect, Dev. Paleoenviron. Res, pp 37–75
Vicente-Serrano SM (2006) Differences in spatial patterns of drought ondifferent time scales: an analysis of the Iberian Peninsula. WaterResour Manag 20:37–60. doi:10.1007/s11269-006-2974-8
Vicente-Serrano SM, Beguería S, López-Moreno JI (2010) A multiscalardrought index sensitive to global warming: the standardizedprecipitation-evapotranspiration index. J Clim 23:1696–1718. doi:10.1175/2009JCLI2909.1
Vicente-Serrano SM, Beguería S, Lorenzo-Lacruz J et al (2012)Performance of drought indices for ecological, agricultural, andhydrological applications. Earth Interact 16:1–27. doi:10.1175/2012EI000434.1
Vicente-Serrano SM, Gouveia C, Camarero JJ et al (2013) Response ofvegetation to drought time-scales across global land biomes. ProcNatl Acad Sci U S A 110:52–57. doi:10.1073/pnas.1207068110
Wigley TM, Briffa KR, Jones PD (1984) On the average value ofcorrelated time series, with applications in dendroclimatology andhydrometeorology. J Clim Appl Meteorol 23:201–213
Wils THG, Robertson I, Eshetu Z et al (2009) Periodicity of growth ringsin Juniperus procera from Ethiopia inferred from crossdating andradiocarbon dating. Dendrochronologia 27:45–58. doi:10.1016/j.dendro.2008.08.002
Int J Biometeorol