dietary cholesterol stimulates hepatic biosynthesis of ... · we recently defined a positive...
TRANSCRIPT

Dietary cholesterol stimulates hepatic biosynthesis of triglyceride and reduces oxidation of fatty acids in the rat
Thomas V. Fungwe, Lauren M. Cagen, George A. Cook, Henry G. Wilcox, and Murray Heimberg' Departments of Pharmacology and Medicine, University of Tennessee-Memphis, The Health Science Center, Memphis, TN 38163
Abstract Experiments were conducted in the intact rat and in the isolated, perfused rat liver to investigate the possibility that the increase in the concentration of hepatic triglyceride and in- crease in the secretion of the very low density lipoprotein (VLDL)-triglyceride (TG) resulting from addition of cholesterol to the diet are due to stimulation of synthesis of triglyceride, reduced fatty acid oxidation, or both. Male rats were fed for 7 days with either a cholesterol-free diet to which 5% (w/w) corn oil was added, or with the same diet supplemented with 0.5% cholesterol. Fed animals received [l-l4C]oleic acid via the tail vein, as a complex with rat serum, and were killed 2 h later. Feeding cholesterol for 7 days increased hepatic triglyceride and cholesteryl ester (CE) concentrations, moderately elevated free cholesterol, but did not affect phospholipid (PL) levels, as we had previously observed after a feeding period of 3 weeks. Incorpora- tion of [l-14C]oleic acid into hepatic and plasma triglyceride in- creased significantly (60 and 48%, respectively) with cholesterol feeding. Incorporation of [ 1- Wloleic acid into hepatic and plasma cholesteryl esters increased by 63 and 79'37, respectively, while incorporation into phospholipid was unaffected. Increas- ing the fat (corn oil) content of the diet to 20% (w/w) did not change these effects of dietary cholesterol. Studies using isolated, perfused rat livers were carried out in vitro after rats were fed the 5% corn oil diet for 3 weeks. [Perfusions lasted 4 h. The per- fusion medium contained 3% bovine serum albumin and 30% washed bovine erythrocytes in Krebs-Henseleit-HC03 buffer. [l-14C]oleic acid was infused at a constant rate (140 pmol/hr; 0.85 pCi/hr) after an initial pulse (60 pmol, 0.375 pCi).] Hepatic uptake of free fatty acid was not altered by dietary cholesterol. Output of all VLDL lipids was stimulated, as was incorporation of I4C into hepatic and VLDL TG and CE (but not PL), by dietary cholesterol. Conversely, formation of I4CO2 and perfu- sate 14C-labeled perchloric acid-soluble compounds (ketones) was reduced in livers of animals fed cholesterol. Furthermore, the activity of mitochondrial carnitine palmitoyltransferase, the rate-limiting enzyme of fatty acid oxidation, was reduced in the liver of intact rats fed cholesterol. Bll The increased level of triglyceride in liver and plasma may be secondary to reduced rates of fatty acid oxidation, to direct stimulation of the incorpo- ration of hepatic acyl-CoA into Z, or to both of these factors. The additional TG synthesized may facilitate transport of cholesteryl ester in the VLDL.- Fungwe, T. V., L. M. Cagen, G. A. Cook, H. G. Wilcox, and M. Heimberg. Dietary
cholesterol stimulates hepatic biosynthesis of triglyceride and reduces oxidation of fatty acids in the rat. J. Lipid Res. 1993. 34: 933-941.
Supplementary key words very low density lipoprotein oleic acid fatty acid metabolism carnitine palmitoyltransferase cholesteryl
ester
Our knowledge regarding a potential regulatory role of cholesterol on fatty acid (FA) metabolism is limited. Clearly, increasing the supply of FA to the liver stimulates esterification of FA to triglyceride (E) (1) and cholesteryl esters (CE) (2) and increases the secretion of the very low density lipoprotein (VLDL) (3, also review article, ref. 4). We have reported that cholesterol excess (5) or depletion (6) can stimulate or reduce, respectively, secretion of the VLDL by the liver. Conversely, the modulation of cholesterol metabolism by FA has been demonstrated in several ways. We reported previously that hepatic cholesterol synthesis, measured by incorporation of SH20
and by the activity of microsomal 3-hrd.oxy-3-methylglutaryl CoA reductase (HMGR), is stimulated in livers perfused with oleate (7, 8). This action of oleic acid, which has been confirmed by other workers (9), may be related to the role of free cholesterol (FC) and CE in the formation and secretion of VLDL (6, 10). Oleic acid also stimulates several other enzymes catalyzing steps in the formation of mevalonate, including cytosolic acetoacetyl CoA thiolase, acetoacetyl CoA synthase, and HMG-CoA synthase (2).
Abbreviations: FA, fatty acid; TG, triglyceride; FFA, free (unesterified) fatty acid; PL, phospholipid; FC, free cholesterol; CE, cholesteryl esters; VLDL, very low density lipoprotein; ACAT, acyl-Ca4 cholesterol acyltransferase; CPT, carnitine palmitoyltransferase; HMGR, 3-hydrmry-3- methylglutaryl CoA reductase; JlGAT, diacylglycerol acyltransferase. 'To whom requests for reprints should be addressed.
Journal of Lipid &search Volume 34, 1993 933
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from

Furthermore, impairment of hepatic FA oxidation at the level of carnitine palmitoyltransferase (CPT) has been reported to increase the activity of HMGR (11). Inhibi- tion of FA oxidation increases TG secretion by the liver and HMGR activity (12). Such actions, which would pro- vide more FA substrate for TG synthesis and increase the cholesterol-synthesizing capacity of the liver, should in- crease both the secretion of the VLDL-TG, and the hepatic content of TG and CE.
We recently defined a positive relationship between hepatic TG and CE levels and secretion rates of VLDL by livers isolated from animals fed varying quantities of cholesterol and perfused with a constant quantity of oleic acid (5). Dietary cholesterol stimulated secretion of VLDL-TG, PL, FC, CE, and apolipoprotein B in a dose- dependent manner. Consistent with our observation are the reports of Surette et al. (13), who demonstrated that n-3 fatty acids from fish oil were dependent on dietary cholesterol to increase plasma VLDL in the hamster, and the work of Ohtani et al. (14), who reported that VLDL secretion was stimulated in cultured hepatocytes har- vested from cholesterol-fed hamsters only when fat was in- cluded in the diet. The increased secretion of VLDL-TG provides a basis for the hypertriglyceridemia, concomi- tant with hypercholesterolemia seen with increased dietary cholesterol. Our data suggest that cholesterol might not only stimulate acyl-CoA cholesterol acyltrans- ferase (ACAT) and increase CE formation, but might also stimulate the synthesis of TG from endogenously and/or exogenously derived FA. Increase of total TG (5) may reflect decreased FA oxidation, a direct stimulatory effect on the rates of TG synthesis, increased de novo biosynthe- sis of FA, or a combination of these factors. As a first ap- proach to the evaluation of effects of dietary cholesterol on the biosynthesis of TG, the esterification of [ 1- 14C]oleic acid in the intact rat and by the isolated perfused rat liver was determined. We also investigated the effects of dietary cholesterol on FA oxidation by measurement of the ac- tivity of carnitine palmitoyltransferase in mitochondria from livers of intact rats, and ketogenesis and 14C02 production from [1-14C]oleic acid by the perfused liver. We observed that dietary cholesterol increased esterifica- tion of FFA to TG, as well as to CE, decreased the activity of hepatic mitochondrial CPT, and reduced oxidation of [14C]oleic acid to 14C02 and total ketones. A preliminary report of this work has appeared (15).
METHODS
Materials and reagents
Cholesterol (USP), cholesterol-free diet, and the scintil- lation fluid Ecolume were purchased from ICN Biochem., Inc (Costa Mesa, CA). Mazola corn oil (no cholesterol, 100% pure, food grade) was obtained locally. [l-l4C]oleic
acid was obtained from NEN (Boston, MA). L-[Methyl- 3H]carnitine hydrochloride was obtained from Amersham Corp. (Arlington Heights, IL). Silica gel-G thin-layer plates were purchased from Analtech Inc. (Newark, DE). All other chemicals and reagents were analytical grade.
Animals and diets
Male Sprague-Dawley rats (Harlan Labs, Madison, WI) weighing 150-175 g were fed ad libitum a cholesterol- free basal diet to which corn oil was added to constitute 5% (w/w), as described previously (5). The animals were housed under a normal light-dark cycle, with lights on from 0700 to 1900. After 1 week on the basal diet, the animals were randomly assigned to a group that was fed the same diet (control) or to a group fed the diet sup- plemented with 0.5 % cholesterol (w/w). The cholesterol was dissolved in the corn oil and mechanically mixed in the diet. The diets were stored at 4OC until used. The ex- perimental diet period was 7 or 21 days. In separate ex- periments, control and cholesterol-fed rats were both fed for 7 days on similar diets except that the corn oil content was increased to 20% (w/w). The composition of the high fat diet was 20% corn oil (fat), 51% carbohydrate (su- crose), 20% protein (casein); minerals, vitamins, and cel- lulose constituted 9%. Because of the differences in the overall composition of the two rat diets, only comparisons of the effect of cholesterol within each dietary fat group was possible. All the animals used in these studies were in a fed state at the initiation of the experiments.
Metabolism of [l-'4C]oleic acid in vivo
Serum from normal chow-fed rats was isolated and used to prepare a complex with [l-14C]oleic acid (6.4 x lo7 dpm/ml serum) (16). Rats fed diets as described above were injected, under light anesthesia with ethyl ether, with 1.8 pCi (20% fat group) or 2.4 pCi (5% fat group) [l-l4C]oleic acid/100 g body weight via a tail vein. After 2 h, during which the animals were conscious, the rats were anesthetized with pentobarbital (60 mg/kg, i.p.) and exsanguinated from the abdominal aorta. The livers were flushed with cold 0.9% NaCl, removed, and weighed. Aliquots of plasma and 1.0-g samples of liver and adipose tissue (epididymal fat pad) were removed for analysis of lipid content. Lipid fractions were separated by thin-layer chromatography (5), and incorporation of 14C into individual lipid fractions was measured by liquid scintillation counting directly without hydrolysis.
Metabolism of [ l-'4C]oleic acid by the isolated perfused rat liver
Livers for perfusions were obtained from rats fed for 21 days with diets as described above. The livers were surgi- cally isolated (4) and were perfused in vitro for 4 h, using a previously described recycling system (7). The livers were generally isolated between 0800 and 0900 h and
934 Journal of Lipid Research Volume 34, 1993
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from

were perfused initially for a period of 20 min without fatty acid or radioactive substrate to allow equilibration. The perfusion was then continued with a medium containing 3 g bovine serum albumin (BSA)/dl, 100 mg glucose/dl, washed bovine erythrocytes (30% v/v) in Krebs- Henseleit-HC03 buffer (pH 7.4). An oleate-BSA com- plex was prepared (6 g delipidated BSA and 1197 pmol oleate (7.5 pCi [l-14C]oleic acid)) and infused during the experiment at a rate of 11.7 mlih (140 pmol oleate and 0.85 pCi per h). At zero time, 5 ml of the radioactive complex was added (60 pmol oleate; 0.375 pCi). Under these con- ditions, the ambient concentration of FFA in the perfusate was maintained at 0.85 f 0.09 (n = 16) mM. Hepatic uptake of FFA was not affected by feeding cholesterol 28.9 * 0.03 pmol fatty acid/g per hr (n = IS)]. There were no significant differences between perfusions of the two diet groups in the specific radioactivity (dpm/pmol) of the circulating free fatty acids. Samples of perfusate were removed for analysis at the end of 2 and 4 h of perfu- sion. At the termination of the experiment, the livers were flushed with 60 ml of ice-cold, oxygenated 0.9% NaCl, blotted, and weighed. Samples of liver (1.0 g) were re- moved for analysis. Whole perfusate was centrifuged at low speed to sediment the erythrocytes; 60 ml of the cell- free perfusate was used to isolate nascent VLDL by ultracentrifugation. Lipids were extracted from perfusate, VLDL, and liver samples, and analyzed as described above. TG synthesis was estimated from incorporation of radioactivity from [ ‘4CIoleate.
As an indication of ketone body production, incorpora- tion of radioactivity from [ l-14C]oleic acid into perchloric acid-soluble products of the perfusion medium was deter- mined by liquid scintillation spectrometry (17). Radioac- tive 14C02 generated during the perfusions was trapped in 10% potassium hydroxide (KOH, w/v). The trapped “CO, was released with sulfuric acid, collected on filter paper strips saturated with hyamine hydroxide, and counted by liquid scintillation spectrometry (18).
Isolation of mitochondria and mitochondrial outer membranes and assay of carnitine palmitoyltransferase
Mitochondria were isolated from rat livers by the method of Johnson and Lardy (19). The final mitochon- drial pellet was resuspended in 20 mM phosphate buffer, pH 7, and mitochondrial outer membranes were isolated by the hypo-osmotic swellingldensity gradient centrifuga- tion technique of Parsons, Williams, and Chance (20). The outer carnitine palmitoyltransferase was measured as described previously (21). Each assay contained, in a total volume of 1 ml, 82 mM sucrose, 70 mM KCI, 70 mM im- idazole, 2 mM reduced glutathione, 1 mg of BSA, 0.5 mM L-carnitine (0.4 pCi of ~-[methyl-~H]carnitine), 1 pg of antimycin A, 2 mM ATP, 2 mM MgC12, 100 pM palmitoyl-CoA for optimum activity measurements or 40
pM palmitoyl-CoA for malonyl-CoA inhibition assays, and various concentrations of malonyl-CoA as indicated.
Statistics All data were analyzed using Student’s t-test for un-
paired samples. The results are expressed as means f SE; differences between means were considered to be significant at the probability level of 0.05.
RESULTS
Lack of effects of diet on weight and food consumption
The average daily consumption of the diet with 5% corn oil (with or without cholesterol) and the weight gained over the feeding periods were not significantly different between treatment groups. The lack of difference was also noted for animals fed similar diets for 3 weeks (5). When animals were maintained on the high fat diet (20% corn oil, with or without 0.5% cholesterol) for the 7-day period, the corresponding values were: food con- sumption per day, 20.8 * 1.8 and 23.0 k 2.1 g; average weight gained per day, 7.9 * 0.4 and 8.0 * 0.3 g; and final body weight, 265 * 9 and 258 * 2 g, for the 0.0 and 0.5 7% cholesterol-fed groups, respectively. The ratio of liver weight to final body weight in the high fat-fed group was unaffected by dietary cholesterol (0.040 * 0.001 vs. 0.046 * 0.002, respectively, for high fat p u p s * cholesterol). These values were not different from those for rats main- tained on the 5% fat diet.
Effects of dietary cholesterol and fat content on plasma and hepatic lipid concentrations
The concentrations of plasma and hepatic lipids were determined in rats fed diets containing either 5% or 20% corn oil for 1 week, with and without 0.5% cholesterol. Plasma lipid concentrations are shown in Table 1. Plasma levels of E and CE in animals fed cholesterol in the 5% corn oil diet were increased above those of rats not receiv- ing cholesterol, while plasma PL, FC, and FFA concentra- tions were not different among groups. As previously reported (5), feeding of inaemental amounts of cholesterol to rats for 3 weeks (with 5% corn oil), increased the con- centrations of all plasma, VLDL, and LDL lipids, but reduced HDL lipids. Interestingly, plasma concentrations of E, CE, and FFA were unaffected, but PL and FC were decreased, by dietary cholesterol in the group fed 20% corn oil.
The data shown in Table 2 indicate that cholesterol, fed with either the low fat or high fat diet, stimulated ac- cumulation of hepatic TG by approximately 100%. In rats receiving the 5% corn oil diet, cholesterol moderately elevated hepatic free cholesterol, and, as previously ob- served (5), increased CE substantially (13-fold). In the animals receiving the high fat diet, hepatic lipid classes
Fungwe et al. Cholesterol and triglyceride biosynthesis 935
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from

TABLE 1. Effects of dietary cholesterol and fat on plasma lipid concentrations
Diet TG PL FC CE FFA
pmol/dl
A. 5% Corn oil Cholesterol added
None (14) 67.9 f 6.0 183.9 f 8.1 53.0 f 2.8 162.9 f 7.4 55.9 + 3.1 0.5% (15) 94.5 f 6.4" 162.8 f 8.6 53.7 f 2.5 202.1 f 16.2" 54.5 f 4.7
B. 20% Corn oil Cholesterol added
None (14) 42.9 f 7.7 155.8 f 3.5 47.2 f 0.9 131.8 f 3.3 41.8 f 2.8 0.5% (15) 41.2 f 4.0 134.9 f 5.4" 37.6 f 1.6" 124.0 f 4.2 41.8 f 2.6
Data are means f SEM. Figures in parentheses indicate number of animals in each group. Animals received
"Different from "None" of the corresponding dietary group, P < 0.05. the diets containing either 5 % or 20% (w/w) corn oil f cholesterol for 7 days.
were similarly increased by cholesterol feeding. The addi- tion of cholesterol to either the 5 or 20% fat diet did not affect total hepatic PL concentration.
Effects of dietary cholesterol and fat on the metabolism of [1-"C]oleic acid in the intact rat
The metabolism of [l-14C]oleic acid in vivo was inves- tigated to determine whether feeding of cholesterol stimu- lates the synthesis of TG from exogenous fatty acids. The incorporation of [l-14C]oleic acid into plasma lipids is summarized in Table 3. When cholesterol was added to the 5% corn oil diet, the net incorporation of 14C into plasma triglyceride increased by 48%, while that into plasma cholesteryl ester was increased 7-fold (Table 3A). The incorporation of 14C into PL was unaffected, and plasma FFA radioactivity was similar in both groups. When rats were fed the 20% fat diet, incorporation of [l-l4C]oleic acid was increased only in plasma CE (Table 3B).
In the animals fed the 5% corn oil diet containing 0.5% cholesterol for 7 days, the incorporation of [l-14C]oleic acid into hepatic TG was stimulated about 6070, and into
hepatic CE by 9-fold, over the 0.0% cholesterol fed group (Table 4A). Total incorporation into hepatic lipids was in- creased by 62 % in the cholesterol-fed group. Incorpora- tion of I4C into hepatic PL was unaffected by dietary cholesterol. Increasing the fat content of the diet to 20% corn oil (Table 4B) did not alter the effect of cholesterol on synthesis of TG and CE. Incorporation of I4C into adi- pose tissue (epididymal fat pad) lipids was unaffected by dietary cholesterol. These findings indicate that hepatic synthesis of TG, as well as CE, from [l-14C]oleic acid is augmented with dietary cholesterol.
Effects of dietary cholesterol on the metabolism of [1-"C]oleic acid by the isolated perfused rat liver
The effect of 0.5% dietary cholesterol on the net secre- tion of VLDL lipids is shown in Fig. 1. The rates of lipid secretion are similar to those previously reported by us for livers from rats similarly fed (5). The effects of cholesterol feeding on the incorporation of [ l-14C]oleic acid into hepatic and VLDL-PL and CE by perfused livers are shown in Fig. 2, while those for TG are shown in Fig. 3. During the 4-hr perfusion, livers from cholesterol-fed rats
TABLE 2. Effects of dietary cholesterol and fat on hepatic lipid concentrations in vivo
Diet TG PL FC CE
pmol/g l ivn
A. 5% Com oil Cholesterol added
3.60 f 0.11 0.88 f 0.08 None (14) 5.28 f 0.52 26.36 f 1.32 0.5% (15) 10.35 f 0.91" 25.16 f 1.37 4.28 f 0.10" 11.49 f 1.16"
B. 20% Corn oil Cholesterol added
None (14) 15.19 f 0.79 29.80 f 1.11 3.94 + 0.08 3.12 f 0.24 20.16 f 1.14"
Data are means f SEM. Figures in parentheses indicate number of animals in each group. Animals received
"Different from "None" of the corresponding group, P < 0.05.
0.5% (15) 34.37 f 1.52" 27.94 f 1.00 4.37 f 0.13"
the diets containing either 5% or 20% (w/w) corn oil + cholesterol for 7 days.
936 Journal of Lipid Research Volume 34, 1993
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from
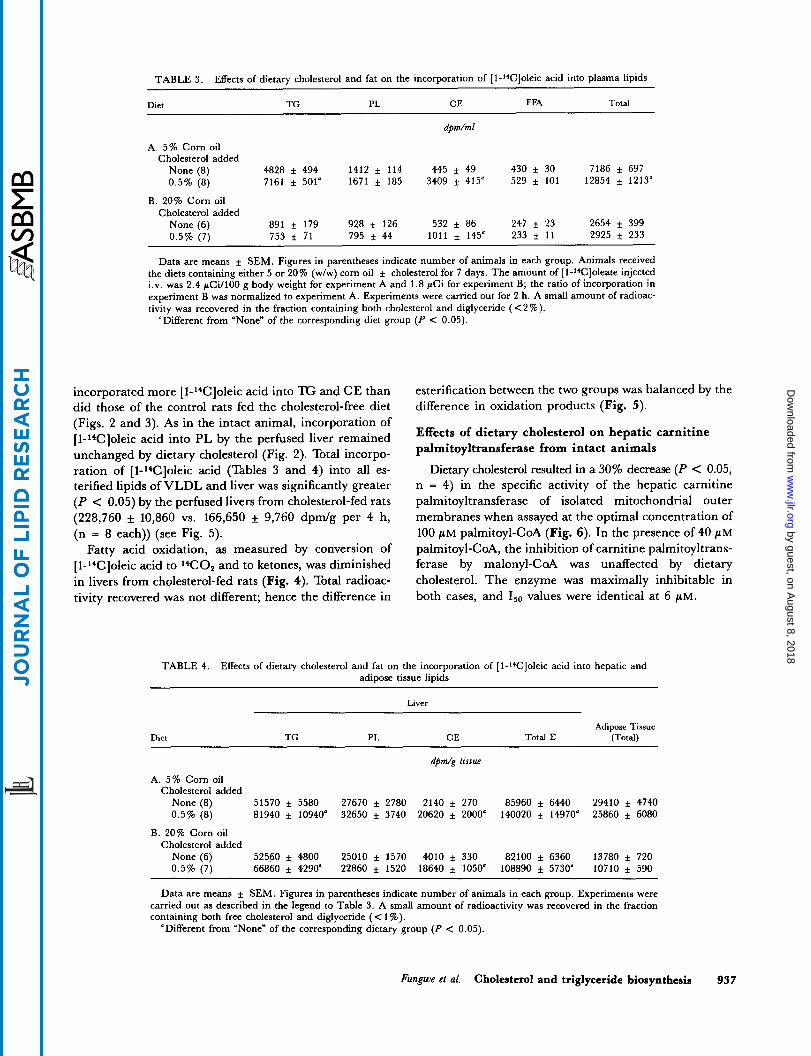
TABLE 3. Effects of dietary cholesterol and fat on the incorporation of [l-l*C]oleic acid into plasma lipids
Dirt TG PL CE FFA Total
dpm/ml
A. 5% Corn oil Cholesterol added
None (8) 4828 f 494 1412 i 114 445 f 49 430 f 30 7186 f 697 0.5% (8) 7161 i 501" 1671 f 185 3409 4 415" 529 f 101 12854 f 1213n
B. 20% Corn oil Cholesterol added
None (6) 891 i 179 928 i 126 532 f 86 247 i 23 2654 i 399 0.5% (7) 753 f 71 795 f 44 1011 i 145" 233 i 11 2925 f 233
Data are means i SEM. Figures in parentheses indicate number of animals in each group. Animals received the diets containing either 5 or 20% (w/w) corn oil f cholesterol for 7 days. The amount of [1-"Cloleate injected i.v. was 2.4 pCi/lOO g body weight for experiment A and 1.8 pCi for experiment B; the ratio of incorporation in experiment B was normalized to experiment A. Experiments were carried out for 2 h. A small amount of radioac- tivity was recovered in the fraction containing both cholesterol and diglyceride (<2%).
"Different from "None" of the corresponding diet group (P < 0.05).
incorporated more [l-l4C]oleic acid into TG and CE than did those of the control rats fed the cholesterol-free diet (Figs. 2 and 3). As in the intact animal, incorporation of [l-'4C]oleic acid into PL by the perfused liver remained unchanged by dietary cholesterol (Fig. 2). Total incorpo- ration of [l-'+C]oleic acid (Tables 3 and 4) into all es- terified lipids of VLDL and liver was significantly greater (P < 0.05) by the perfused livers from cholesterol-fed rats (228,760 * 10,860 vs. 166,650 * 9,760 d p d g per 4 h, (n = 8 each)) (see Fig. 5).
Fatty acid oxidation, as measured by conversion of [1-'4C]oleic acid to '4C02 and to ketones, was diminished in livers from cholesterol-fed rats (Fig. 4). Total radioac- tivity recovered was not different; hence the difference in
esterification between the two groups was balanced by the difference in oxidation products (Fig. 5).
Effects of dietary cholesterol on hepatic carnitine palmitoyltransferase from intact animals
Dietary cholesterol resulted in a 30% decrease (P < 0.05, n = 4) in the specific activity of the hepatic carnitine palmitoyltransferase of isolated mitochondrial outer membranes when assayed at the optimal concentration of 100 p M palmitoyl-CoA (Fig. 6). In the presence of 40 p M palmitoyl-CoA, the inhibition of carnitine palmitoyltrans- ferase by malonyl-CoA was unaffected by dietary cholesterol. The enzyme was maximally inhibitable in both cases, and 150 values were identical at 6 pM.
TABLE 4. Effects of dietary cholesterol and fat on the incorporation of [l-1*C]oleic acid into hepatic and adipose tissue lipids
Diet
Liver
TG PL CE Adipose Tissue
Total E (Total)
d p d g tissue
A. 5% Corn oil Cholesterol added
None (8) 51570 f 5580 27670 f 2780 2140 f 270 85960 f 6440 29410 f 4740 0.5% (8) 81940 f 10940" 32650 i 3740 20620 f 2000" 140020 f 14970" 25860 f 6080
B. 20% Corn oil Cholesterol added
None (6) 52560 f 4800 25010 f 1570 4010 330 82100 f 6360 13780 f 720 0.5% (7) 66860 i 4290" 22860 f 1520 18640 f 1050" 108890 i 5730" 10710 f 590
Data are means f SEM. Figures in parentheses indicate number of animals in each group. Experiments were carried out as described in the legend to Table 3. A small amount of radioactivity was recovered in the fraction containing both free cholesterol and diglyceride (< 1 %).
"Different from "None" of the corresponding dietary group (P < 0.05).
Fungwe et al. Cholesterol and triglyceride biosynthesis 937
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from

1 .o
0.8
L r L QJ c 0.6
M
0
3.
.
.- -
. - E 0.4
0.2
0.0
* % Cholesterol (diet) W None(n=S)
0.5 (n=8) (* p<o.ns)
*
TG PL FC CE
Lipid Class
Fig. 1. Effects of dietary cholesterol on the secretion of VLDL lipids by the isolated perfused rat liver. See text for experimental details. Bars represent means SE; ‘P < 0.05 versus control value.
DISCUSSION
The objectives of this study were to determine whether the increased accumulation of hepatic T G and the hyper- triglyceridemia (5) that accompanies inclusion of 0.5 % cholesterol in the diet of the rat result from increased rates of esterification of exogenous FFA, decreased FFA oxida- tion, or both. When synthesis of T G exceeds the forma- tion and secretion of VLDL, TG accumulates in the liver (22). Several conditions may potentially lead to increased TG synthesis; increased availability of exogenous fatty acids from the diet or derived from adipose tissue (23), stimulation of fatty acid synthesis (24), increased channel- ing of fatty acids into hepatic triglycerides (24), decreased oxidation of fatty acids, increased uptake of fatty acids by the liver, or some combination of these factors. We exa- mined some of these conditions as potential mechanisms to account for the accumulation of T G in the liver with feeding of cholesterol. Because perfused livers from cholesterol-fed rats secrete more VLDL lipids and apoB than livers from control rats, inability to secrete lipoproteins by these livers at normal rates is not a contributing factor to TG accumulation in the liver (5). However, an inability to assemble VLDL at rates comparable to increased TG and C E synthesis may certainly be a factor. In the present study no differences were observed in the uptake of FFA by the perfused livers, suggesting that the rate of uptake is not a factor contributing to TG accumulating in the livers of cholesterol-fed rats. Nevertheless, an increase in the availability of FA for esterification is one means that could account for the increase in TG and C E synthesis resulting in an increase in TG output and accumulation of TG in the liver.
We anticipated that feeding of cholesterol would in- crease the flux of substrate into CE. However, we also ob- served that incorporation of [l-l4C]oleic acid into TG was enhanced in livers of cholesterol-fed rats in vivo and in livers isolated and perfused in vitro. Dietary cholesterol may stimulate biosynthesis of TG through increased ac- tivity of glycerophosphate acyltransferase (EC 2.3.1.15), which catalyzes the first committed step in glycerolipid synthesis, phosphatidate phosphohydrolase (EC 3.1.3.4), which controls the rate of diglyceride production and di- acylglyceride acyltransferase (EC 2.3.1.20) (DGAT), which may regulate the channeling of diglyceride into triglyceride. As changes in P L synthesis were not ob- served, the data suggest that dietary cholesterol may aug- ment DGAT activity, or that P L synthesis is maximal un-
15000 -
T T
2oooo 1 T
15000
10000
5000
0 VLDL LIVER TOTAL
Fig. 2. Effects of cholesterol feeding (5% corn oil) for 3 weeks on the incorporation of [l-’4C]oleate into liver and VLDL PL and C E by the isolated perfused rat liver. Livers were isolated from control (0.0% cholesterol diet, None) or cholesterol-fed rats (0.5% cholesterol) and per- fused as described in the text. Incorporation of [l-’4C]oleate into PL and CE was measured after the 4 h of perfusion in VLDL isolated from the medium, or in the livers. Values are presented as means & SEM with the number of experiments given in parentheses. “Total” refers to sum of VLDL liver data; *P < 0.05 versus control value.
938 Journal of Lipid Research Volume 34, 1993
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from
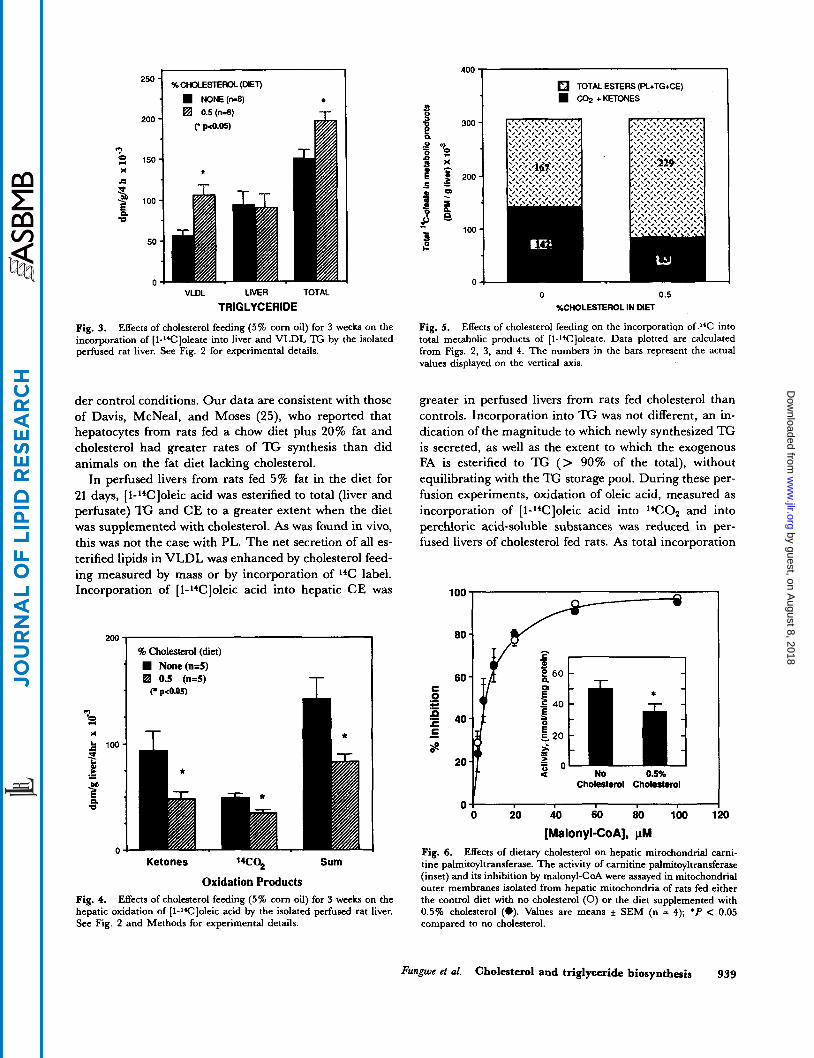
250
.
200
? 150
X
G
j 100 P U
50
0
%CHOLESTEROL (DIET)
NONE(n-8)
VLDL LIVER TOTAL
TRIGLYCERIDE
Fig. 3. Effects of cholesterol feeding (5% corn oil) for 3 weeks on the incorporation of [1-Wloleate into liver and VLDL 'E by the isolated perfused rat liver. See Fig. 2 for experimental details.
der control conditions. Our data are consistent with those of Davis, McNeal, and Moses (25), who reported that hepatocytes from rats fed a chow diet plus 20% fat and cholesterol had greater rates of TG synthesis than did animals on the fat diet lacking cholesterol.
In perfused livers from rats fed 5% fat in the diet for 21 days, [l-l*C]oleic acid was esterified to total (liver and perfusate) TG and CE to a greater extent when the diet was supplemented with cholesterol. As was found in vivo, this was not the case with PL. The net secretion of all es- terified lipids in VLDL was enhanced by cholesterol feed- ing measured by mass or by incorporation of 14C label. Incorporation of [l-W]oleic acid into hepatic CE was
96 Cholesterol (diet) None(n=5)
0 0.5 ( n 3 ) (* p<o.o5) T
Ketones ' 4 q Sum
Oxidation Products Fig. 4. Effects of cholesterol feeding (5% corn oil) for 3 weeks on the hepatic oxidation of [l-"C]oleic acid by the isolated perfused rat liver. See Fig. 2 and Methods for experimental details.
TOTAL ESTERS (PLtTG+CE) C o p +KETONES
0 0.5 %CHOLESTEROL IN MET
Fig. 5. Effects of cholesterol feeding on the incorporatign of l'C into total metabolic products of [l-Wloleate. Data plotted are calculated from Figs. 2, 3, and 4. The numbers in the bars represent the actual values displayed on the vertical axis.
greater in perfused livers from rats fed cholesterol than controls. Incorporation into TG was not different, an in- dication of the magnitude to which newly synthesized TG is secreted, as well as the extent to which the exogenous FA is esterified to E (> 90% of the total), without equilibrating with the TG storage pool. During these per- fusion experiments, oxidation of oleic acid, measured as incorporation of [l-i4C]oleic acid into '+GO2 and into perchloric acid-soluble substances was reduced in per- fused livers of cholesterol fed rats. As total incorporation
loo] 80
2 - No 0.5% Cholesterol Cholesterol
" l i o 40 60 so IQO 1 0
[Malonyl--A], pM Fig. 6. Effects of dietary cholesterol on hepatic mirochondrial carni- tine palmitoyltransferase. The activity of carnitine palmitoyltransferase (inset) and its inhibition by malonyl-CoA were assayed in mitochondrial outer membranes isolated from hepatic mitochondria of rats fed either the control diet with no cholesterol (0) or the diet supplemented with 0.5% cholesterol (0). Values are means f SEM (n = 4); 'P < 0.05 compared to no cholesterol.
Fungwe et al. Cholesterol and triglyceride biosynthesis 939
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from

into all esterified and oxidation products was similar in both groups of animals, the differences observed were due to the manner in which the exogenously supplied fatty acid was metabolized in the presence or absence of dietary cholesterol.
The data reported in this study also demonstrate that cholesterol feeding reduced the activity of carnitine palmitoyltransferase (CPT). Impairment of FA oxidation at the level of CPT has been reported to stimulate cholesterol synthesis by the liver and increase esterifica- tion of fatty acids to TG (12, 26), suggesting that, under conditions where substrate supply is available, lowering CPT activity may lead to cholesterol and TG accumula- tion in vivo. These data support the observations that in- creasing the supply of FFA to the liver stimulates TG syn- thesis and VLDL secretion, in addition to stimulating cholesterol synthesis and the activity of HMGR (7-9). Decreasing the activity of CFT, which is normally the rate-limiting enzyme in the hepatic fatty acid oxidation pathway, would be expected to increase substantially the quantity of FA esterified, even if cholesterol did not cause any modification of other pathways. Jansen, Van der Liden, and Hulsmann (11) cite their unpublished observa- tions that CPT activity is reduced in the cholesterol-fed guinea pig. Consistent with that is our observation (Fig. 6) that cholesterol feeding diminishes the activity of CPT without changing its sensitivity to inhibition by malonyl- CoA. These two aspects are usually regulated in tandem in changing physiological and pathophysiological states (21). We observed that the activity of CPT was increased by 0.1% dietary lovastatin, but the sensitivity of CPT to inhibition by malonyl-CoA was unaffected (27). By in- hibiting HMGR, lovastatin may have acted to lower hepatic membrane cholesterol content, producing effects that were opposite to those of cholesterol feeding. Kashfi, Dory, and Cook (28) observed that cholesterol loading of cultured macrophages also depressed CPT activity. Clearly, the data on FA oxidation obtained from the per- fused liver experiments complement observations made in the intact rat, suggesting that cholesterol feeding dimin- ishes the oxidation of fatty acids.
The increase in plasma and hepatic TG ( 5 ) we observed is consistent with published reports in guinea pigs fed cholesterol in chow diets containing corn or coconut oil (29) and in rabbits receiving coconut oil- or olive oil- based diets (30). However, the hypertriglyceridemia ob- served with coconut oil, but not olive oil, in the latter study was attributed more to decreased lipoprotein lipase activity than to increased VLDL secretion (30). It is clear from our study that when the polyunsaturated vegetable oil (corn oil) is the dietary source of fatty acids for energy and for membrane structure, cholesterol stimulates TG synthesis and VLDL secretion. Even when the corn oil content is increased from 5 to 2096, the effects of cholesterol on TG synthesis are still evident.
In light of their previous studies (31-33) and those of others (34), Yamamoto et al. (31) proposed a sequence of biochemical adjustments of the hepatocyte to high sucrose diets. These include: u) increased glycolysis, providing substrate for synthesis of long chain fatty acids; b) in- creased energy production from glycolysis and oxidation of the resulting acetyl-CoA via the Krebs cycle; c) in- creased formation of malonyl-CoA due to elevated flux of citrate to fatty acids; d) suppression of FA oxidation via increased ATP synthesis and inhibition of CPT by malonyl-CoA; e) increase in FA uptake, stimulating TG synthesis due to increased availability of FA substrate; 8 greater transfer of TG to cytoplasmic lipid droplets and VLDL particles in the endoplasmic reticulum and, there- fore; h) greater deposition of TG in the liver and increased secretion of TG-rich lipoproteins into plasma. In this study we used a diet similar to that of Yamamoto et al. (31) except that cholesterol was added to the diet of the ex- perimental group. Therefore, the above sequence of events probably holds true in our current study for both control and experimental animals, but the final outcome was enhanced by cholesterol in the diet.
Dietary cholesterol failed to stimulate hepatic forma- tion of phospholipid in vivo or in vitro. This observation is in agreement with that of Driscoll et al. (29) in the cholesterol-fed guinea pig. This may reflect selective stimulation of DGAT activity or the fact that DGAT, which regulates the channeling of diglyceride into triglyceride, is saturated at much higher levels of FA flux through the liver than is the activity of choline- phosphotransferase, the key enzyme of de novo phos- phatidylcholine synthesis.
In summary, the accumulation of TG in the liver, con- comitant with an increase in the secretion of V L D L - E , that occurs when cholesterol is fed in the diet to rats results from increase in the synthesis of TG. Reduction in oxidation of fatty acids may contribute, in part, to the in- creased TG synthesis. The increase in the incorporation of FA into CE and TG may reflect increased availability of FA substrate because of reduced rates of FA oxidation, or increased capacity for enzymatic synthesis of CE and TG, or both of these pathways. An increase in the availa- bility of TG resulting from the increase in synthesis may be required for the formation of VLDL as a vehicle for outward transport of excess CE from the liver. I
The authors are grateful to Mss. Poonam Kumar, Jewel Donald- son, and Jeanine Weakley for the excellent technical assistance provided during the course of this study. This research was sup- ported by grants R01-HL-27850, HL-39228, and HL-40929 from the National Institutes of Health, U.S. Public Health Serv- ice, and by Biomedical Research Support Grant-90-02. This work was completed while T.V.F. was a Postdoctoral trainee sup- ported by training grant T32-HL-07641 from the National Insti- tutes of Health. Manuscript received I1 September 1992 and in reuzied form I 4 January 1993.
940 Journal of Lipid Research Volume 34, 1993
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from

1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
REFERENCES
Heimberg, M., H. G. Wilcox, G. D. Dewey, W. F. Wood- side, K. J. Breen, and C. Soler-Argilaga. 1974. Studies on the regulation of secretion of very low density lipoprotein, and on ketogenesis by the liver. In Regulation of Hepatic Metabolism. F. Lundquist, and N. Tygstrup editors. Munksgaard, Copenhagen. 119. Salam, W. S., H. G. Wilcox, L. M. Cagen, and M. Heim- berg. 1989. Stimulation of hepatic cholesterol biosynthesis by fatty acids: effect of oleate on cytoplasmic acetoacetyl- CoA thiolase, acetoacetyl-CoA synthetase and hydroxy- methylglutaryl-CoA synthase. Biochem. J. 258: 563-568. Heimberg, M., J. Olubadewo, and H. G. Wilcox. 1985. Plasma lipoproteins and regulation of hepatic metabolism of fatty acids in altered thyroid states. Endocr. Rev. 6: 590-607. Grundy, S. M., and M. A. Denke. 1990. Dietary influences on serum lipids and lipoproteins. J. Lipid Res. 31: 1149-1172. Fungwe, T. V., L. M. Cagen, H. G. Wilcox, and M. Heim- berg. 1992. Regulation of hepatic secretion of very low den- sity lipoprotein by dietary cholesterol. J. Lipid Res. 33:
Khan, B., H. G. Wilcox, and M. Heimberg. 1989. Cholesterol is required for secretion of very-low-density lipoprotein by rat liver. Biochem. J. 259: 807-816. Goh, E. H., and M. Heimberg. 1977. Effects of free fatty acids on the activity of hepatic microsomal 3-hydroxy-3- methylglutaryl coenzyme A reductase and on secretion of triglyceride and cholesterol by liver. J. Biol. Chem. 252:
Goh, E. H., and M. Heimberg. 1979. Relationship between activity of hepatic microsomal 3-hydroxy-3-methyglutaryl coenzyme A reductase and secretion of very low density lipoprotein cholesterol by the isolated perfused liver and in the intact rat. Biochem. J. 184: 1-6. Takashi, I., and J. A. Ontko. 1981. Increased secretion of very low density lipoprotein triglyceride following inhibi- tion of long chain fatty acid oxidation in isolated rat liver. J. Biol. Chem. 256: 10247-10255. Cianflone, K. M., Z. Yasruel, M. A. Rodriquez, D. Vas, and A. D. Sniderman. 1990. Regulation of apoB secretion from HepG2 cells: evidence for a critical role for cholesteryl ester synthesis in the response to fatty acid challenge. J. Lipid Res. 31: 2045-2055. Jansen, H., H. N. Van der Liden, and C. W. Hulsmann. 1990. Inhibition of carnitine palmitoyltransferase leads to induction of 3-hydroxymethylglutaryl coenzyme A reduc- tase activity in rat liver. Biochim. Biophys. Acta. 1044:
Fukada, N., and J. A. Ontko. 1984. Interactions between fatty acid synthesis, oxidation, and esterification in the production of triglyceride-rich lipoproteins by the liver. J. Lipid Res. 25: 831-842. Surette, M. E., J. Whelan, G-P. Lu, K. S. Broughton, and J. E. Kinsella. 1992. Dependence on dietary cholesterol for n-3 polyunsaturated fatty acid-induced changes in plasma cholesterol in the Syrian hamster. J. Lipid Res. 33: 263-271. Ohatani, H., K. Hayashi, Y. Hirata, S. Dojo, K. Nakashima, E. Nishio, H. Kurushima, M. Saeki, and G. Kajiyama. 1990. Effects of dietary cholesterol and fatty acids on plasma cholesterol level and hepatic lipoprotein metabolism. J. Lipid Res. 31: 1413-1422. Wilcox, H. G., T. V. Fungwe, and M. Heimberg. 1992. Dietary cholesterol stimulates triglyceride biosynthesis in the intact rat and in the isolated perfused rat liver. FASEB J. 6: A1206.
179-191.
2822-2826.
390-393.
16. Soler-Argilaga, C., H. G. Wilcox, and M. Heimberg. 1976. The effect of sex on the quantity and properties of the very low density lipoprotein secreted by the liver in vitr0.J. Lipid
17. Stakkestad, J. A., and J. Bremer. 1982. The metabolism of fatty acid in hepatocytes isolated from triiodothyronine- treated rats. Biochim. Biophys. Acta. 711: 90-100.
18. Keyes, W. G., H. G. Wilcox, and M. Heimberg. 1981. For- mation of the very low density lipoprotein and metabolism of [l-l*C]oleate by perfused livers from rats treated with tri- iodothyronine or propylthiouracil. Metabolism. 30: 135-145.
19. Johnson, K., and H. Lardy. 1967. Isolation of liver or kid- ney mitochondria. Methods Emymol. 10: 94-96.
20. Parsons, D. F., G. R. Williams, and B. Chance. 1966. Characteristics of isolated and purified preparations of the outer and inner membranes of mitochondria. Ann. N Y Acad. Sci. 137: 643-666. Cook, G. A,, and M. S. Gamble. 1987. Regulation of carni- tine palmitoyltransferase by insulin results in decreased ac- tivity and decreased apparent Ki values for malonyl-CoA. J. Biol. Chem. 262: 2050-2055.
22. Woodside, W. F., and M. Heimberg. 1978. The metabolism of oleic acid by the perfused rat liver in experimental dia- betes induced by anti-insulin serum. Metab. Clin. Exp. 27:
23. Fex, G. 1970. The metabolism of liver and plasma lipids after partial hepatectomy. Biochim. Biophys. Acta. 202: 415-425.
24. Grove, C. D., and D. A., Hems. 1978. Fatty acid synthesis in regenerating liver of the rat. Biochem. J. 170: 1-8.
25. Davis, A. R., M. M. McNeal, and L. R. Moses. 1982. In- trahepatic assembly of very low density lipoprotein: compe- tition by cholesterol esters for the hydrophobic core. J. Biol.
26. Bergstrom, J. D., G. A. Wong, P. A. Edwards, and J. Ed- mond. 1984. The regulation of acetyl-coA synthetase ac- tivity by modulators of cholesterol synthesis in vivo and the utilization of acetoacetate for cholesterogenesis. J. Biol.
27. Cook, G. A., B. Khan, and M. Heimberg. 1988. Feeding of lovastatin to rats increases the activity of the hepatic mitochondrial outer carnitine palmitoyltransferase. Bi- ochem. Biophys. Res. Commun. 150: 1077-1082.
28. Kashfi, K., L. Dory, and G. A. Cook. 1991. Effects of cholesterol loading of mouse macrophages on carnitine palmitoyltransferase activity and sensitivity to inhibition by malonyl-CoA. Biochem. Biophys. Res. Commun. 177: 1121-1126.
29. Driscoll, D. M., T. Mazzone, T. Matsushima, and G. S. Getz. 1990. Apoprotein E biosynthesis in the cholesterol-fed guinea pig. Arteriosclemsis. 10: 31-39.
30. Van Heek, M., and D. B. Zilversmit. 1991. Mechanisms of hypertriglyceridemia in coconut oil/cholesterol-fed rabbit. Increased secretion and decreased catabolism of very low density lipoprotein. Arteriosclm Thmmb. 11: 918-927. Yamamoto, M., I. Yamamoto, Y. Tanaka, and J. Ontko. 1987. Fatty acid metabolism and lipid secretion by perfused livers from rats fed laboratory stock and sucrose-rich diets. J. Lipid Res. 28: 1156-1165.
32. Ontko, J. A. 1972. Metabolism of free fatty acids in isolated liver cells. J. Biol. Chem. 247: 1788-1800.
33. Zaleski, J., and J. A. Ontko. 1985. Reciprocal effects of energy utilization on palmitate oxidation and esterification in hepatocytes of fed rats. Biochim. Biophys. Acta. 826:
34. McGarry, J. D., G. F. Leatherman, and D. W. Foster. 1978. Carnitine palmitoyltransferase I. Site of inhibition of hepatic fatty acid oxidation by malonyl-CoA. J. Biol. Chem.
Res. 17: 139-145.
21.
1763-1 7 77.
Chem. 257: 2634-2640.
Chem. 259: 14548-14553.
31.
134-142.
253: 4128-4136.
Fungwe et al. Cholesterol and triglyceride biosynthesis 941
by guest, on August 8, 2018
ww
w.jlr.org
Dow
nloaded from