developments in bioremediation of soils and sediments polluted … · 2015-09-22 · developments...
TRANSCRIPT

Developments in bioremediation of soils and sediments polluted withmetals and radionuclides – 1. Microbial processes and mechanisms affectingbioremediation of metal contamination and influencing metaltoxicity and transport
Henry H. Tabak1,*, Piet Lens2, Eric D. van Hullebusch2,4,5 & Winnie Dejonghe31US EPA, ORD, National Risk Management Research Laboratory, 26 West Martin Luther King Drive,Cincinnati, OH, USA; 2Sub-Department of Environmental Technology, Wageningen University, Wageningen,The Netherlands; 3Flemish Institute for Technological Research, VITO, Belgium; 4Universite de Marne-la-Vallee, Laboratoire des Geomateriaux, 77454 Marne La Vallee, Cedex, 2 – France; 5Universite de Limoges,Laboratoire des Sciences de l’Eau et de l’Environnement, 87060 Limoges Cedex, France (*author for cor-respondence, e-mail: [email protected])
1. Introduction
1.1. Bioremediation Technologies
Bioremediation technology uses microorganismsto reduce, eliminate, contain and transform tobenign products, contaminants present in soils,sediments, water or air. The last 15 years haveseen an increase in the types of contaminants towhich bioremediation is being applied, includingsolvents, explosives, polycyclic aromatic hydro-carbons (PAHs), and polychlorinated biphenyls(PCBs). Now, microbial processes are beginningto be used in the cleanup of radioactive andmetallic contaminants.
Bioremediation is an alternative to traditionalremediation technologies such as landfilling orincineration. Bioremediation depends on the pres-ence of the appropriate microorganisms in thecorrect amounts and combinations and on theappropriate environmental conditions. Althoughprokaryotes – Bacteria and Archaea – are usuallythe agents responsible for most bioremediationstrategies, eukaryotes such as fungi and algae alsocan transform and degrade contaminants. Micro-organisms already living in contaminated envi-ronments are often well-adapted to survival in thepresence of existing contaminants and to the tem-perature, pH, and oxidation–reduction potentialof the site. These indigenous microbes tend to uti-lize the nutrients and electron acceptors that areavailable in situ, provided liquid water is present.
The bulk of subsurface microbial populations areassociated with both microorganisms and dis-solved substances, including contaminants andtheir breakdown products.
Bioremediation works by either transformingor degrading contaminants to non-hazardous orless hazardous chemicals. These processes arecalled, respectively, biotransformation andbiodegradation. Biotransformation is any alter-ation of the molecular or atomic structure of acompound by microorganisms. Biodegradation isthe breaking down of organic substance bymicroorganisms into smaller organic or inorganiccomponents.
Unfortunately, metals and radionuclides can-not be biodegraded. However, microorganismscan interact with these contaminants and trans-form them from one chemical form to another bychanging their oxidation state through theaddition of (reduction) or removing of (oxida-tion) electrons. In some bioremediation strategies,the solubility of the transformed metal or radio-nuclide increases, thus increasing the mobility ofthe contaminant and allowing it to more easily beflushed from the environment. In other strategies,the opposite will occur, and the transformedmetal or radionuclide may precipitate out ofsolution, leading to immobilization. Both kindsof transformations present opportunities forbioremediation of metal and radionuclides in theenvironment – either to immobilize them in placeor to accelerate their removal.
Reviews in Environmental Science and Bio/Technology (2005) 4:115–156 � Springer 2005DOI 10.1007/s11157-005-2169-4

1.2. Metal contamination
The challenge posed by metal contaminants liesin the permanence of their nuclear structures:stable isotopes are virtually indestructible. Theymay be redistributed, however, as well as inter-converted among forms of varying chemical andphysical properties; as such metal mobility andmutability are the sources of both problemsand of many promising solutions.
Most heavy metals exist naturally in theearth’s crust at trace concentrations of just a fewparts per million (Bodek et al. 1988), sufficient toprovide local biota with trace nutrients, but toolow to cause toxicity. Disposal of wastes frommetal excavation and processing have concen-trated these metals to dangerous levels in somesoils and sediments, endangering both wildlifeand people and motivating efforts to detoxify thesoils. EPA standards rarely require that metalcontamination be reduced to background levels,however, with preliminary remediation goals formetals varying from 0.2 to 63,000 ppm based en-tirely on projected risks (Smucker 1996).
Metal contamination of soil is especiallyproblematic because of the strong adsorption ofmany metals to the surfaces of soil particles. Dueto the difficulty of desorbing metal contaminants,some traditional remediation methods simplyimmobilize metals in contaminated soils, forexample, by the addition of cement or chemicalfixatives, by capping with asphalt, or by in-situvitrification. Alternatively, soils are often isolatedby excavation and confinement in hazardouswaste facilities (Schleck 1990; Trezek 1990). Al-though rapid in effect, both of these options areexpensive ($30–$300 m)3) and destroy the soil’sfuture productivity (Cunningham et al. 1995).
Soil washing and pump-and-treat technologiesambitiously attempt to remove contaminatingmetals from soils by pumping water or other sol-vents into soil and extracting the liquid down-gradient from the contamination. Metals arethen precipitated from the liquid, which is recy-cled if possible. The success of these methods isseverely limited by the slow desorption kineticsof adsorbed metals, with the result that addi-tional agents are often used to promote metaltransfer to the aqueous phase. Typical additivesare acids, chelates, and reductants, which im-
prove cost effectiveness but may introduce fur-ther harmful chemicals (Boyle 1993; EPA 1991).
A primary strategy of bioremediation is theuse of similar metal-mobilizing agents in conjunc-tion with soil washing, with the advantage thatthey pose no known environmental threat them-selves. Biopolymers have been discovered thatbind metals with high affinity and travel relativelyunimpeded through porous medium. Certainmicroorganisms transform strongly adsorbingmetal species into more soluble forms, and plantsare being recruited that act as self-containedpump-and-treat systems. Other methods employenzymatic ativities to transform metal species intovolatile, less toxic, or insoluble forms. Techniquesfor soil bioremediation are usually designed to beused in-situ, lowering costs; they avoid the use oftoxic chemicals, and in nearly all cases, the soilstructure and potential for productivity arepreserved. Although the understanding of theprocesses discussed below is far from complete,intense research in bioremediation is rapidlyelucidating the mechanisms involved.
2. Microbial processes affecting bioremediation
of metals and radionuclides
Bioremediation of metals and radionuclides relieson a complex interplay of biological, chemicaland physical processes. A fundamental mechanis-tic understanding of coupling microbial metabo-lism, chemical reactions and contaminanttransport is beginning to develop as well as howthese activities could work together to bioremedi-ate metal and radionuclide pollution
Microbes exist in complex biogeochemicalmatrices in subsurface sediments and soils. Theirinteractions with metals and radionuclides areinfluenced by a number of environmental factors,including solution chemistry, sorptive reactivesurfaces, and the presence or absence of organicligands and reductants (Figure 1).
Microorganisms can interact with metals andradionuclides via many mechanisms some ofwhich may be used as the basis of potentialbioremediation strategies. The major types ofinteraction are summarized in Figure 2. In addi-tion to the mechanisms outlined, accumulationmof metals by plants (phytoremediation) warrants
g p g
116

attention as an additional established route for thebioremediation of metal contamination. The stud-ies of Salt, Raskin and co-workers (Raskin et al.1997; Salt et al. 1995; Salt et al. 1998) provide adetailed description of use of plants for (1) phy-toextraction; the use of metal-accumulating plantsto remove toxic metals from soil; (2)rhizofiltration; the use of plant roots to removetoxic metals from polluted waters; and (3) phyto-stabilization; the use of plants to eliminate the bio-availability of toxic metals in soils. Recently,comprehensive reviews were published on phyto-remediation of metal pollution in soils (Chaneyet al. 1997; Saxena et al. 1999; Vangronsveld 2000;van der Lelie et al. 2001; Reeves & Barker 2001;Schwitzguebel et al. 2002; Pulford & Watson 2003).
Iron cycling and associated changes in solid-phase chemistry have dramatic implications forthe mobility and bioavailability of heavy metalsand radionuclides. Coupled flow and waterchemistry control the rate and solid phase prod-ucts of iron hydroxide reduction and providecritical information in assessing the reactivity of
reduced environments toward metal and radionu-clide contaminants.
Bioremediation of soils, sediments, and watercontaminated with metals and radionuclides canbe achieved through biologically mediated chan-ges in the oxidation state (speciation) of thosecontaminants – biotransformation. Changes inspeciation can alter the solubility of metals andradionuclides, and therefore their transport prop-erties and toxicity. The latter two characteristicscan determine bioavailability (see van Hullebuschet al. 2005 for further details). Resistance by sub-surface microorganisms to the toxicity of heavymetals is critical for the bioremediation of con-taminated subsurface sites. Remedial action de-pends on actively metabolizing microbes, andthese microbes might be inhibited by high con-centrations of toxic heavy metals.
There are at least three types of microbialprocesses that can influence toxicity and trans-port of metals and radionuclides: biotransforma-tion, biosorption and bioaccumulation, anddegradation or synthesis of organic ligands that
( g y , y)
Figure 1. Abiotic and biotic mechanisms influence the fate of metals (M) in subsurface environments. From left to right: Organicmaterial produced by microorganisms can act as ligands (L) and complex with metals, facilitating transport. These ligands can alsobe degraded by microbes to relase the metal. Microbes can also directly adsorp/desorb, takeup/excrete, and oxidize/reduce metals.Oxidation and reduction will change the valence state of the metal either up or down (Mn±x). Abiotically, metals can form solu-tion complexes or can complex with the surface of clays (kaolinite–polymer complexes). These mineral complexes can sorb metalseither directly to the mineral or to organic or iron oxide coatings. Metals can also be directly oxidized or reduced to differencevalence states by the ambient redox conditions or by organic reductants (Lred) or oxidants (Lox). Changing the valence state ofmetals will affect their sorption, mobility, precipitation, and toxicity (image courtesy of S. Fendorf, Standford University).
117

affect the solubility of the contaminants. Eachoffers the potential for bioremediation of metallicand radioactive contaminants in the environment.
2.1. Biotransformation
Metal-reducing microorganisms can reduce awide variety of multivalent metals that pose envi-ronmental problems. The heavy metals and ra-dionuclides subject to enzymatic reduction bymicrobes include but are not limited to uranium(U), technetium (Tc), and chromium (Cr). Directenzymatic reduction involves use of the oxidizedforms of these contaminants as electron accep-tors. The oxidized forms of U, Tc, and Cr arehighly soluble in aqueous media and are gener-ally very mobile in aerobic ground water, whilethe reduced species are highly insoluble and oftenprecipitate from solution. Direct enzymaticreduction of soluble U(VI), Tc(VII), and Cr(VI)
to insoluble species has been documented and isillustrated in Figure 3.
Extracellular precipitation of metals andradionuclides has been demonstrated in anumber of microbial isolates. For example, theprecipitation of uranium on the cell surface ofthe bacterium Shewanella is shown in Figure 4.Metal-reducing organisms reduce uranyl carbon-ate, which is exceedingly soluble in carbonate-bearing ground water, to highly insoluble U(IV),which precipitates from solution as the uraniumoxide mineral uraninite.
Significant advances have been made in under-standing the mechanisms of reduction of Fe(III),U(VI) and Tc(VI) in the subsurface bacteriumGeobacter sulfurreducens, using the tools of bio-chemistry and molecular biology. A surface-bound c-type cytochrome (mass 40 kDa) that isinvolved in the transfer of electrons to extracellu-lar insoluble Fe(III) oxides has been identified
Figure 2. Metal–microbe interactions impacting bioremediation.
118

and characterized. This protein is not required forthe reduction of U(VI), suggesting that the mech-anisms of Fe(III) and U(VI) reduction may bedistinct. Another c-type cytochrome (9.6 kDa),found in the periplasm, appears to be requiredfor U(VI) reduction (Figure 5). This protein isable to reduce U(VI) in vitro, and a mutantunable to synthesize the protein was unable toreduce U(VI) efficiently. Surprisingly, Tc(VII) isreduced by yet another mechanism, a periplasmicNi/Fe-containing hydrogenase that uses hydrogenas the electron donor for metal reduction. More-over, efficient indirect mechanisms may be impor-tant in immobilizing Tc in sediments whereinbiologically reduced Fe(III) or U(IV) is able totransfer electrons directly to Tc(VII).
A wide range of bacteria reduces the highlysoluble chromate ion to Cr(III), which underappropriate conditions precipitates as Cr(OH)3.A number of Cr(VI)-reducing microorganismshave been isolated from chromate-contaminatedwaters, oils, and sediments, including Arthrobact-er sp., Pseudomonas aeruginosa S128, someanaerobic-reducing bacteria, and even several al-gae. Laboratory experiments with Hanford Sitesediments showed that Cr(VI) concentration inwater decreased significantly (>66%) in amonth-long incubation in the presence of nitrateand added dilute molasses as an electron donor.Thus, the addition of molasses to vadose zonesediments shows potential to decrease the trans-port of chromium and nitrate into underlyingaquifers.
Although some microorganisms can enzymati-cally reduce heavy metals and radionuclides di-rectly, indirect reduction of soluble contaminantsmay be possible in sedimentary and subsurfaceenvironments, although this has not been demon-strated under natural conditions, to date. Thisindirect immobilization could be accomplishedby metal-reducing or sulfate-reducing bacteria.One approach would be to couple the oxidationof organic compounds or hydrogen to the reduc-tion of iron [Fe(III)], manganese [Mn(IV)], orsulfur [S(IV)] in the form of sulfate, [SO4
2)]. Ir-on(III) can be biologically reduced to Fe(II),Mn(IV) to Mn(III), and S(VI) (sulfate) to S(II)(hydrogen sulfide, H2S). The reduced productmight then, in turn, chemically reduce metals orradionuclides to yield separate or multicompo-nent insoluble species (see van Hullebusch et al.2005 for details).
The most reactive of these reduced speciesare ferrous iron [Fe(II)] and H2S. The latter is
Figure 3. Direct enzymatic reduction of soluble heavy metals and radionuclides by metal-reducing bacteria. Non-hazardousorganic compounds, such as lactate or acetate, provide electrons used by these microorganisms. Note, however, that if complexed,the reduced species may become mobile.
Figure 4. Microbes can play an important role in immobiliz-ing radionuclides. This image shows a cross section of thebacterium Shewanella with uraninite precipitated on the cellsurface (image courtesy of S. Fendorf, Standford University).
119

generated by the enzymatic activity of iron-reducing and some fermentative bacteria, canreduce multivalent metals such as uranium,chromium, and technetium (Figure 6a). The re-duced forms of these metals are insoluble andcan either precipitate as reduced oxide orhydroxide minerals, or coprecipitate with Fe(III)minerals that form during the reoxidation ofFe(II). In coprecipitation, elements are incorpo-rated in metal oxide minerals as they precipitatefrom solution.
Sulfate-reducing bacteria also may be stimu-lated to produce a chemically reactive redox bar-rier (Figure 6b). Hydrogen sulfide generated bysulfate-reducing bacteria could chemically reduce
the contaminant to a form that would be stablefor extended periods of time. A study of biofilmsin a zinc and lead mine is a good example of indi-rect immobilization of heavy metals by sulfate-reducing bacteria. Sulfide produced by sulfate re-duces in the film scavenged zinc and other toxicmetals. X-ray fluorescence microbeam analysis re-vealed that zinc and small amounts of arsenic andselenium were extracted from ground water andconcentrated in biofilms in zinc sulphide precipi-tates (Figure 7) (Courtesy of K.M. Kenner,Arrgonne National laboratory and K.M. Ban-field, University of California, Berkeley). Thus,microbial formation of sulfide deposits drasticallydecreased the migration of contaminant metals.
Figure 5. Transmission electron micrograph (TEM) showing extracellular and periplasmic U(IV) precipitates formed by enzymaticreduction of U(VI) by the subsurface bacterium Geobacter sulfurreducens. (TEM image obtained by S. Glasauer, Guelph University).
Figure 6. Indirect mobilization of heavy metals and radionuclides by (a) metal-reducing and (b) sulfate-reducing bacteria.
120

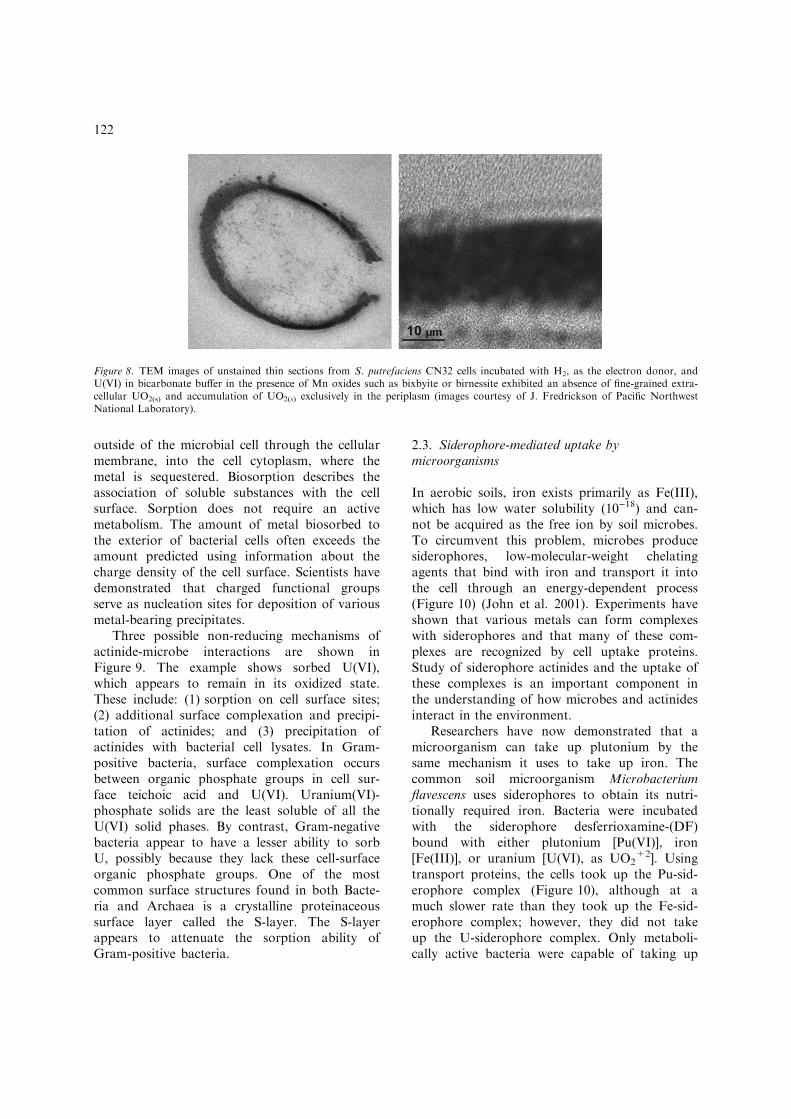
Manganese (III/IV) oxides, which are com-mon mineral phases in many soils and sediments,are also electron acceptors for metal-reducingbacteria. Manganese oxides are also relativelystrong oxidants and can oxidize insoluble, re-duced contaminants such as the mineral uraninite(UO2), a common product of microbial uraniumreduction. Differences in the solubility of oxi-dized Mn (insoluble) and U (soluble) challengebacterium predictions of their biogeochemicalbehaviour during in situ bioreduction. Resultsfrom laboratory experiments with the subsurfacebacterium Shewanella putrefaciens CN32 quanti-tatively reduced U(VI) to U(IV), in the form of asolid UO2. The UO2 was observed in regionsexternal to the cell as well as in the periplasm,the region between the inner and outer mem-brane of the cell, in the presence of the Mn oxi-des, the reduced U resided exclusively in theperiplasm of the bacterial cells (Figure 8) (Cour-tesy of J. Fredrickson, of Pacific NorthwestNational Laboratory). These results indicate thatthe presence on Mn(III/IV) oxides may impedethe in situ biological reduction of U(VI) insubsoils and sediments. However, the accumula-tion of U(IV) in the periplasm indicates that the
cell may physically protect reduced U from oxi-dation by Mn oxides, suggesting that extensivereduction of soil Mn oxides may not be requiredfor reductive biomobilization of U(IV).
Natural organic matter (NOM) may play arole in the reduction of contaminants such asCr(VI) and U(VI) in subsurface environments.NOM consists of a mixture of organic com-pounds with different structures and functionalgroups. These groups include aromatic and phe-nolic moieties, carboxylic and heteroaliphatic hy-droxyl functional groups, and free radicals. In thepresence of a metal-reducing bacterium, NOMeffectively mediated the transfer of electrons forthe reduction of Fe(III), Cr(VI), and U(VI), al-though the reduction rate varied among differentNOM samples and among contaminants.
2.2. Bioaccumulation and biosorption
Microorganisms can physically remove heavymetals and radionuclides from solution throughassociation of these contaminants with biomass.Bioaccumulation is the retention and concentra-tion of a substance within an organism. In bioac-cumulation, solutes are transported from the
g 0 p y fl ( )
Figure 7. X-ray fluorescence trace metal microanalysis of sulfate-reducing bacteria. Results of electron (bottom left) and X-ray (topright) microprobe analysis of specific biomineralized zinc sulfide precipitates. The sensitivity of the X-ray microprobe enables iden-tification of arsenic and selenium constituents in the zinc sulfide precipitate on the surface of a sulfate-reducing bacterium.
121
outside of the microbial cell through the cellularmembrane, into the cell cytoplasm, where themetal is sequestered. Biosorption describes theassociation of soluble substances with the cellsurface. Sorption does not require an activemetabolism. The amount of metal biosorbed tothe exterior of bacterial cells often exceeds theamount predicted using information about thecharge density of the cell surface. Scientists havedemonstrated that charged functional groupsserve as nucleation sites for deposition of variousmetal-bearing precipitates.
Three possible non-reducing mechanisms ofactinide-microbe interactions are shown inFigure 9. The example shows sorbed U(VI),which appears to remain in its oxidized state.These include: (1) sorption on cell surface sites;(2) additional surface complexation and precipi-tation of actinides; and (3) precipitation ofactinides with bacterial cell lysates. In Gram-positive bacteria, surface complexation occursbetween organic phosphate groups in cell sur-face teichoic acid and U(VI). Uranium(VI)-phosphate solids are the least soluble of all theU(VI) solid phases. By contrast, Gram-negativebacteria appear to have a lesser ability to sorbU, possibly because they lack these cell-surfaceorganic phosphate groups. One of the mostcommon surface structures found in both Bacte-ria and Archaea is a crystalline proteinaceoussurface layer called the S-layer. The S-layerappears to attenuate the sorption ability ofGram-positive bacteria.
2.3. Siderophore-mediated uptake bymicroorganisms
In aerobic soils, iron exists primarily as Fe(III),which has low water solubility (10)18) and can-not be acquired as the free ion by soil microbes.To circumvent this problem, microbes producesiderophores, low-molecular-weight chelatingagents that bind with iron and transport it intothe cell through an energy-dependent process(Figure 10) (John et al. 2001). Experiments haveshown that various metals can form complexeswith siderophores and that many of these com-plexes are recognized by cell uptake proteins.Study of siderophore actinides and the uptake ofthese complexes is an important component inthe understanding of how microbes and actinidesinteract in the environment.
Researchers have now demonstrated that amicroorganism can take up plutonium by thesame mechanism it uses to take up iron. Thecommon soil microorganism Microbacteriumflavescens uses siderophores to obtain its nutri-tionally required iron. Bacteria were incubatedwith the siderophore desferrioxamine-(DF)bound with either plutonium [Pu(VI)], iron[Fe(III)], or uranium [U(VI), as UO2
+2]. Usingtransport proteins, the cells took up the Pu-sid-erophore complex (Figure 10), although at amuch slower rate than they took up the Fe-sid-erophore complex; however, they did not takeup the U-siderophore complex. Only metaboli-cally active bacteria were capable of taking up
Figure 8. TEM images of unstained thin sections from S. putrefaciens CN32 cells incubated with H2, as the electron donor, andU(VI) in bicarbonate buffer in the presence of Mn oxides such as bixbyite or birnessite exhibited an absence of fine-grained extra-cellular UO2(s) and accumulation of UO2(s) exclusively in the periplasm (images courtesy of J. Fredrickson of Pacific NorthwestNational Laboratory).
122

the Pu siderophore complexes, just as with Fe-siderophore uptake. These discoveries couldhave wide-ranging implications for future biom-ediation efforts and for more accurate predic-tions of how plutonium and other actinidesbehave in the environment. Siderophore-medi-ated uptake and transport could be an impor-tant pathway for environmental mobility and Puentry into the food chain.
2.4. Microbes and synthetic organic chelators
Organic complexing agents can have a profoundeffect on the mobility of metals and radionuclidesin subsurface environments. Synthetic chelatorssuch as EDTA and NTA can form stable, solu-ble complexes with heavy metals and radionuc-lides. These chelators were commonly used ascleaning agents during industrial processing of
y gy)
Figure 9. Three possible non-reductive mechanisms of bacterial cell surface interaction with U(VI) (Courtesy of H. Nitzche andT. Hazen, Lawrence Berkeley National Laboratory, and S. Clark, Wash. State University).
Figure 10. Siderophore-mediated Pu accumulation by Microbacterium flavescens (John et al. 2001).
123

nuclear fuels and were sometimes co-disposedwith metals and radionuclides. Metal–chelatecomplexes have entered the environment andmay migrate in ground water. However, themigration of these complexes can be reduced bythe biodegradation of the organic ligand (Fig-ure 11). The resulting free metal ions are likely toabsorb to mineral surfaces or to form oxide min-eral precipitates that would be less mobile inground water. The degradation of organic chela-tors associated with metal or radionuclide con-taminants, then, might achieve a desirableimmobilization of contaminants in place.
Some of these chelators can be degraded bynaturally occurring microorganisms. A number ofEDTA- and NTA-degrading organisms have beenisolated and identified. In one study, microbialdegradation of EDTA by the environmental iso-late BNC1 was influenced by the complex metal.Cobalt(II)–EDTA, cobalt (III)–EDTA, and nick-el(II)–EDTA complexes were not degraded,whereas copper (II)–EDTA and zinc–EDTAcomplexes were. The genes and enzymes responsi-ble for EDTA and NTA degradation have beenidentified, and the genes have been cloned and se-quenced. All the genes necessary to code for deg-radation of EDTA and NTA occur together in a‘‘gene cluster’’. Elucidating the genes and en-zymes responsible for EDTA and NTA biodegra-dation will provide an understanding of theenvironmental and physiological controls on che-late degradation in bacteria, and provide geneprobes for monitoring this process in the environ-ment. Such fundamental research on the mecha-nisms of enzymatic degradation of syntheticchelators is expected to provide useful informa-tion for developing bioremediation strategies.
Metal-reducing bacteria also sometimes actu-ally promote the mobilization of insoluble formsof some heavy metals and radionuclices. It hasbeen demonstrated that metal-reducing bacteriacan solubilize PuO2, which is insoluble, in the
presence of the synthetic chelator NTA. It isthought that the bacteria reduced the insolublePu(IV) to Pu(III), which was then complexed byNTA. This process may provide a means ofmobilizing Pu from contaminated soils and sedi-ments, and could be a step in the removal of thishighly toxic radionuclide from the environment.However, this approach has not been tested inthe field.
Organic acids formed by the metabolic activ-ity of microorganisms can lower the pH of thesystem to values that interfere with the electro-static forces that hold heavy metals and radio-nuclides on the surface of iron or manganeseoxide minerals. Displacement of cations byhydrogen ions may lead to the solubilization ofthe surface-associated metal or radionuclide. Insome cases, the organic metabolites also serve ascomplexing agents that can form soluble metal–ligand complexes. These complexing agents(which include dicarboxylic acids, phenolic com-pounds, ketogluconic acids, and salicylic acids)have been shown to promote the dissolution of awide range of heavy metals and radionuclides,including PuO2. Therefore, biogenic productionof complexing agents can accelerate the move-ment of metals in soils and sediments.
2.5. Microbial metabolism of iron reducingbacteria
2.5.1. Dissimilatory iron reductionIron is extremely abundant in the Earth’s crust,primarily in the form of insoluble Fe(III) oxides.The reduction potential of Fe(III)/Fe(II) is elec-tropositive (Table 1). A number of microorgan-isms are able to couple oxidation of hydrogen ororganic compounds to the reduction of Fe(III)and gain energy for growth. The use of iron orother metals as terminal electron acceptors iscalled dissimilatory metal reduction. (Not all dis-similatory metal reduction, however, is linked toenergy conservation.) Geological and microbio-logical evidence suggests that Fe(III) reductionwas a very early form of respiration on Earth.
A phylogenetically diverse group of Bacteriaand Archaea is known to conserve energy to sup-port growth by oxidizing hydrogen or organiccompounds (including contaminants such asaromatic hydrocarbons) with the reduction of
y)
Figure 11. Immobilization of heavy metals by enzymatic deg-radation of organic chelators, such as EDTA and NTA.
124

Fe(III). Such a group includes species from suchgenera as Geobacter, Desulfuromonas, Pelobacter,Shewanella, Ferrimonas, Geovibrio, Geothrix, andothers. These organisms have a broad spectrumof other metabolic capabilities as well. Many dis-similatory metal reducers such as Geobacter spe-cies can reduce soluble U(VI) to insoluble U(IV)(Figure 12).
Dissimilatory metal-reducing microorganismsmight prevent migration of uranium in groundwater by precipitation and immobilization in thesubsurface. When a simple organic compoundsuch as acetate is added to the subsurface, aero-bic microorganisms quickly consume availabledissolved oxygen and nitrate. Then dissimilatory
metal-reducing microorganisms begin to metabo-lize acetate, oxidizing it to CO2 while reducingavailable metals. While Fe(III) is generally themost abundant metal electron acceptor in thesubsurface, dissimilatory metal-reducing microor-ganisms can also simultaneously reduce U(VI) toU(IV), precipitating it out of ground water. Thishas been demonstrated conclusively in laboratorystudies, and is the basis for new strategies ofin situ biomediation.
2.5.2. MechanicsMechanisms for ion reduction appear to varyamong dissimilatory metal-reducing microorgan-isms (Figure 13). Some species use strategies to
Table 1. Microbially significant half-reaction reduction potentials
Transformation Reaction Eh, Volts (@ pH 7)
O2 depletion 0.5O2+2H+ fi H2O 0.82
Denitrification NO3)+6H++5e) fi 0.5N2+3H2O 0.71
Mn reduction, Mn(IV) to Mn(II) MnO2+4H++2e) fi Mn2++2H2O 0.54
Fe reduction, Fe(III) to Fe(II) Fe(OH)3+3H++e) fi Fe2++3H2O 0.01
Sulfate reduction, S(VI) to S(-II) SO42)+10H++8e) fi H2S+4H2O )0.22
Methane generation, C(IV) to C(-IV) HCO)3+9H++8e) fi CH4+3H2O )0.26
H2 generation, H(I) to H(0) H++e+ fi 0.5H2 )0.41
see Stumm and Morgan (1996).
Figure 12. Geobacter sulfurreducens growing with insoluble Mn(IV) oxides as the electron acceptor (Image courtesy of D. Lovley,University Massachusetts).
125

overcome the need for direct contact with Fe(III)oxides. For example, Shewanella oneidensis is aversatile microbe that can use oxygen, nitrate,uranium, manganese, and iron as electron accep-tors. This bacterium appears to release quinonesinto the culture medium during growth that serveas electron shuttles between the bacterium andthe Fe(III) oxide. Shewanella alga and Ceothrixfermentans (in addition to producing electronshuttles) solubilize Fe(III) during growth, pre-sumably by releasing one or more Fe(III)-com-plexing compounds called chelators. By contrast,Geobacter metallireducens does not release elec-tron shuttles and is highly adapted to contactwith the solid Fe(III) oxide. When growing oninsoluble Fe(III) or Mn(IV) oxides, this microor-ganism produces flagella and uses chemotaxis tofind the electron acceptor. Pili are also producedunder these conditions, presumably to attach toinsoluble oxides.
Although dissimilatory metal reducers are ofobvious importance to developing strategies forbiomediation of organic contaminants as well asmetals and radionuclides, this process can be
slow. One idea for stimulating their activity inaquifer sediments is to add humic acids or otherquinone-containing compounds to which Fe(III)-reducing microbes can transfer electrons. Elec-tron shuttling via extracellular quinones mayaccelerate the rate and extent of biomediation.
2.5.3. Exploring the diversity of iron (III)-reducing bacteria in subsurface sedimentsIron(III)-reducing bacteria are thought to cata-lyze a large number of sedimentary processesthat have important impacts on biomediation.Many new strains of iron-reducing microorgan-isms have now been isolated from uranium-con-taminated subsurface sediments, expanding ourknowledge of the diversity of this environment.Gene-sequencing methods have been used toclassify these isolates, which include Gram-posi-tive genera (Clostridium, Camoccus) that werenot closely related to any previously character-ized pure cultures of Fe(III)-reducing bacteria.
The Clostridia are well-studied anaerobic bac-teria that have been isolated from sedimentssince the origin of environmental microbiology.
Figure 13. Mechanisms of Fe(III) reduction by dissimilatory metal-reducing bacteria (c = chelator) (Image courtesy of D. Lovleyand ASM News).
126

However, all of the Clostridia isolated previouslywere fermentative organisms incapable of respira-tion. In contrast, many of the Clostridium strainsdescribed from contaminated subsurface sedi-ments were shown to conserve energy for growthby coupling the respiration of Fe(III) oxideminerals to the oxidation of organic acids (ace-tate or lactate). Several of the bacterial isolateswere also shown to reduce U(VI). Although theirenvironmental significance remains to be ex-plored, these newly isolated FeRBs could play animportant role in subsurface biomediation.
3. Microbial mechanisms involved in
bioremediation of metal and radionuclide
contaminated soils and sediments
3.1. Biopolymers and their use in bioremediationof metal contamination
Microbially produced macromolecules with me-tal-binding properties include cyclodextrins, exo-polysaccharides and amphipathic moleculestermed ‘‘biosurfactants.’’ Through combinationsof carboxyl, phosphoryl, and hydroxyl groups,these molecules complex divalent cations,increasing their aqueous solubility to enhance theefficiency of soil washing. Biopolymers may beinjected with washing solvents or produced with-in contaminated soil by metal-tolerant microor-ganisms. The properties of these molecules arealso potentially alterable by genetic engineeringto produce structures with higher metal specifici-ties and complexing strengths.
3.1.1. BiosurfactantsComposed of polar, often phosphate-based‘‘heads’’ and non-polar lipid ‘‘tails’’, biosurfac-tants combine complexation activity with physi-cal sequestration of the complexed ions (Miller,1995). A ‘‘surfactant,’’ or surface-active agent,diminishes the attractive forces among the mole-cules of a liquid at the liquid’s surface. Becausethe free energy of the surface is thereby lowered,surfactant molecules tend to concentrate at suchsurfaces, including soil–water interfaces (TinocoJr. et al. 1985). This characteristic directly bene-fits the surfactant’s bioremedial purpose by con-centrating the molecules near sorbed metals,
enhancing rates of attainment of complexationequilibrium. At a sufficiently high-concentrationof 1–200 mg/L, called the critical micelle concen-tration (CMC), biosurfactant molecules begin toaggregate into micelles (spherical bilayers),sequestering the chelated ions within the parti-cles. These particles usually measure less than50 nm in diameter and are thus expected toavoid filtration effects in most soils. Adsorptionmay still impede their progress, however (Miller1995).
The most thoroughly studied bio-surfactant isthe anionic rhamnolipid of Pseudomonas aerugi-nos (Figure 14). The rhamnolipid forms com-plexes with Cd2+, Zn2+ and Pb2+ in aqueoussolution, with stability constants (Log K) of 6.5,6.6 and 5.4, respectively (Herman et al. 1995).These are far lower than the metal–EDTA con-stants (b=105.0 to 106.0) suggesting that the bio-surfactant should compete successfully with soilorganic matter for metal complexation.
The rhamnolipid’s metal-mobilizing capabilityhas been investigated in a sandy loam soil of min-imal (0.11%) organic matter, showing that leaddesorption is the rhamnolipid’s greatest strength.In soil samples with adsorbed cadmium, zinc andlead (total 3.4 mmol kg)1), 12.5 mM rhamnolipdenhanced lead desorption relative to an electro-lyte control solution, while 80 mM rhamnolipidwas needed to improve cadmium and zinc desorp-tion. The variable effectiveness is likely to be dueto properties of the metals: lead is a softer Lewisacid than cadmium or zinc. Therefore, it tends toform more stable complexes and is less affectedby ion-exchange processes. These characteristicsare evident in both the notorious difficulty of leadremoval from soil and in the measured metal–rhamnolipid stability constants. While a fairlyconcentrated (110–180 mM) electrolyte solutionreleased the harder acids by cation exchange,
Figure 14. Rhamnolipid structure of Pseudomonas aeruginosaATCC 9027. For C18 monorhamnolipid, m+n=10; for C20,m+n=12; for C22, m+n=12; and for C24, m+n=12. (Rep-rinted from Zhang & Miller, 1994, with permission. Copy-right 1994 American Society for Microbiology).
127

rhamnolipid complexation was required to mobi-lize soil-sorbed lead. If rhamnolipids are generallyeffective in mobilizing soft Lewis acids, they willprovide a much-needed tool in soil remediation(Herman et al. 1995).
After complex formation and presumablyafter subsurface transport and recovery, therhamnolipid can be recovered and recycled:acidification of a solution containing a cad-mium–rhamnolipid complex precipitates therhamnolipid, releasing nearly all of the metalinto solution. The rhamnolipid may then be sep-arated by centrifugation, redissolved at neutralpH and reused, while the metal is recoveredfrom the supernatant by alkali precipitation orcations exchange (Tan et al. 1994).
One limitation to rhamnolipid use is the highconcentration needed to release metals from soil,due to strong adsorption of the rhamnolipid it-self. Rhamnolipid-soil sorption is, however,greatly diminished in solutions of low ionicstrength, suggesting that ‘‘cation bridging’’ be-tween the anionic head group and cations sorbedto the soil occurs. Efforts are currently underwayto improve the efficiency of rhamnolipid treat-ment (Herman et al. 1995).
Remarkably, the P. aeruginosa rhamnolipidcan also promote the transport of entire microor-ganisms through a solid matrix, in this case aporous sand. Experimentation and modelinghave shown that the rhamnolipid, used at150–1000 mg l)1, acts by decreasing adsorptionof the cells to the particle surfaces, either byincreasing the negative charge density of the sur-faces and thus the repulsion between cells andsurfaces, by dissolving extracellular polymersused in adhesion, or by competing with themicroorganisms for sorption sites. Because in-situbioremediation will sometimes require the deliv-ery of microorganisms to sites of contaminationand because most bacteria have low mobility insoils, this rhamnolipid property may becomequite valuable (Bai et al. 1997).
3.1.2. CyclodextrinsCyclodextrins are cyclic oligosaccharides (Fig-ure 15) formed during bacterial degradation ofplant starches (Wang & Brusseau 1993). The cen-tral ‘‘cavity’’ of relatively low polarity allows for-mation of 1:1 inclusion complexes with non-polarorganic molecules, while the hydrophilic outer
ring enables the complex to remain water-soluble.The addition of carboxylate groups to the outerring has given the cyclodextrins metal-complexingability as well: carboxymethyl-b-cyclodextrin, orCMCD, strongly complexes Cd2+ with a constantof 103.66 for 1 g/l CMCD (Wang & Brusseau 1995)and the presence of phenanthrene as a model or-ganic contaminant does not reduce this metal-complexing capacity (Brusseau et al. 1997).
Like biosurfactants, cyclodextrins do lowerthe surface tension of water, but they do nothave a critical micelle concentration (Wang &Brusseau 1993). Cyclodextrins are small enoughto avoid pore exclusion in soils and experimentshave shown no measurable adsorption of thecyclodextrin to sandy subsoils and surface soils(Brusseau et al. 1994), indicating that cyclodex-trin mobility in soils should be good. Further,cyclodextrins are generally non-toxic (Wang &Brusseau 1995).
In column experiments, mixtures of carboxyl-ated and non-carboxylated cyclodextrins dramat-ically improved cadmium, nickel, strontium andphenanthrene elution from soils in comparison todilute electrolyte solutions, removing approxi-mately 90% of the metals and 86% of the phen-anthrene within 30 pore volumes (Figure 16).This ability of cyclodextrins to mobilize metaland organic contaminants simultaneously is asignificant attribute, suggesting that use of cyclo-dextrins may greatly enhance pump-and-treatremediation of sites with mixed metal-organiccontaminants.
3.1.3. ExopolysaccharidesExopolysaccharides are relatively large molecules,frequently with molecular weight of 106 daltonsor more, with anionic character due to acidicfunctional groups. Exopolysaccharides are typi-cally highly hydrated and flexible in solution, andas metals are bound, they may fold into morecompact structures. Some polysaccharides aggre-gate into large flocs in the presence of metals,possibly due to formation of insoluble metalhydrolysis products that form nuclei for furtherprecipitation. While useful in the recovery ofmetals from waters, this reduced solubility great-ly diminishes such a polymer’s mobility and bio-remedial effectiveness in soils (Brierley 1990).A great diversity of microorganisms producesfree and/or capsular exopolysaccharides, and
128

Figure 15. Structure of hydroxypropyl-b-cyclodextrin (Reprinted from Wang & Brusseau (1993), with permission. Copyright 1993American Chemical Society).
Figure 16. Elution of a southern Arizona surface soil (0.68% organic carbon, 88.5% sand, 4.3% silt, 10.2% clay, pH 7.5) contam-inanted with cadmium (1.6 mg/g soil), nickel, strontium and phenanthrene (7.1 lg/g soil) using CMCD (carboxymethyl-b-cylodex-trin) or 0.01 M KNO3. Phenanthrene elution is not shown (Reprinted from Brusseau et al. 1997, with permission. Copyright 1997American Chemcial Society).
129

most of these are known to possess metal-bind-ing activity. The emulsan of Acinetobacter RAG-1, for example, binds up to 240 lg UO2 per mg.Other exopolysaccharides produced by Pseudo-monas, Arthrobacter and Klebsiella have boundper mg polymer, up to 96 lg uranium, 3.3 lgcadmium and 22 lg copper, respectively (Miller1995).
Recently, the first demonstration has beenmade that a expolysaccharide can desorb a metalbound to a mineral surface and transport thecomplexed metal through a porous medium. Thisstructurally undefined polysaccharide, producedby a Gram negative soil isolate Strain 9702-M4,actively complexed Cd2+ and sorbed only weaklyto sand particle surfaces. Most importantly, thepolysaccharide released 57% of the (approxi-mately 0.08 ppm) cadmium adsorbed to sandparticles in 1500 pore volumes, compared to non-detectable cadmium release in polymer-free con-trols. The polymer-facilitated cadmium transportis an encouraging result, but not necessarily ageneral one; in particular, it waits testing in morefinely textured soils (Chen et al. 1995).
3.2. Microbial leaching
Bacteria that leach metals from ores are chemo-lithotrophic, or ‘‘rock-eating,’’ meaning that theyobtain energy from the oxidation of inorganicsubstances coupled to the respiration (reduction)of oxygen. Chemolithotrophs of the genera
Thiobacillus and Leptospirillum, the most impor-tant in microbial leaching, flourish in metal sul-fide deposits exposed to air and water. Theyoxidize iron and sulfides, generating sulfuric acidand releasing associated metals into aqueoussolution (Brierley 1982). A model of the indirectleaching mechanism of Thiobacillus ferrooxidansconsists of several steps (Figure 17): the bacteriabind themselves to a metal sulfide substrate withextracellular polymers, forming biofilm of one ortwo cell layers. The exopolymers hold 0.5–5%iron, needed to overcome the repulsion betweennegatively charged sulfide minerals and anioniccell surface macro-molecules (Blake et al. 1994).The bacteria oxidize the bound iron with molecu-lar oxygen, forming Fe (III) hexahydrates at lowpH, and the Fe(III) hexahydrates in turn oxidizemineral-surface sulfides to soluble thiosulfates.Thiosulfates may be further oxidized by T. ferr-oxidans or by commensal bacteria that cannotoxidize sulfide directly, ultimately producing sul-furic acid (Sand et al. 1995).
Thiobacilli are widely exploited: the miningrecovery of metals from low grade ores throughprocesses known as ‘‘heap leaching’’ and ‘‘dumpleaching ‘‘ in which excavated ores are moundedtogether, often on an impermeable slope and irri-gated with acidified water to promote indige-nous, microbial activity. The leachate is collectedat the bottom for metal recovery (Brierly 1982).The success of microbiological mining has in-spired work in recovering metals from other
Figure 17. A model for the indirect leaching attack mechanism as catalyzed by a metal-sulfide-attached cell of Thiobacillus ferroox-idans, exaggerating the exopolymer layer to show its importance (Adapted from Sand et al. 1995).
130

substrates as well, notably metal contaminatedsewage sludge. Such sludges, generated in abun-dance by municipal water treatment plants, canbe spread onto agricultural land for decomposi-tion if their metal contents do not exceed regula-tory standards. Unfortunately, approximately50% of the sludges in the USA and Canada docontain metals, predominantly as metal sulfides,in excess of the standards. Attempts at chemicaldissolution of sludge metals, usually by additionof acids have proven costly and ineffective(Couillard & Mercier 1991).
To solve the sludge problem, a continuousflow bioreactor system has been developedfor sludge treatment, using T. ferrooxidans tocatalyze metal dissolution and using industrialFeSO4 Æ7H2O as an energy source for the bacteria.With an hydraulic residence time of 18 h andsludge volume of 30 L, the following metalsolubilizations have been achieved; Cu, 91% of2500 ppm; Zn, 94% of 1800 ppm; Mn, 93% of380 ppm; Ni, 67% of 23 ppm and Cd, 67%of 16 ppm. After treatment, therefore the sludgeseasily met the Quebec limits of 1000 ppm Cu,2500 ppm Zn, 1500 ppm Mn, 180 ppm, Ni and15 ppm Cd. The biological leaching method re-duces the necessary acid input to 15% and theincubation time to 10% of those required inbatch-type chemical leaching methods (Couillard& Mercier 1991; Couillard & Mercier 1993). Infurther experiments, leaching of Pb was greatlyimproved when the FeSO4 Æ7H2O substrate wasreplaced with FeCl2 presumably due to the lowsolubility of lead sulfate. The bacterial populationadapted well to the FeCl, but not to the substitu-tion of HCl for H2SO4 in initial acidification ofthe system, due to either chloride intolerance or toa sulfate requirement. Further research is neededto optimize lead solublization by these methods,but the leaching technique for the other metalsappears ready for implementation (Mercier et al.1996).
In the past decade, the ability of Thiobacillito remove trace metals from soils and sedimentshas been studied (Tichy et al. 1998). Operationalconditions studied include soil sediment charac-teristics (Loser et al. 2004), initial pH (Chen &Lin 2001a), percentage of inoculum, retentiontime, initial solid content (Chen & Lin 2000)and nutrition (Chen & Lin 2001b). When insol-uble elemental sulfur is used as substrate in the
bioleaching process, the microbial oxidation ofthe sulfur by Thiobacilli is believed to take placeby the adsorption and growth of bacteria onthe surface of the sulfur particles (Boseker1997). The adsorption of bacteria to the solidsubstrate and plays a vital role in the bioleach-ing process the sulfur oxidation rate (Chen &Lin 2001b). Suspension leaching is not economi-cally feasible for treating large amounts of sedi-ment. An alternative is solid bed leaching, asdone with ores. In a solid bed, the density andparticle size distribution of the solid matrix areimportant factors regulating mass transport, sul-fur oxidation and microbial activity. Improvingthe physicochemical properties and the structureof dredged sediments by pretreatment withplants enabled leaching of heavy metals in labo-ratory percolator systems with nearly the sameefficiency as suspension leaching (Loser et al.2001). Nevertheless, good performance in thelaboratory does not guarantee similar efficiencyon a large scale since important process param-eters can be carefully controlled in the labora-tory and fluctuations are usually avoided. Someinfluences on process operation under practicalconditions, such as the effects of solid inhomo-geneity, channeling, or pH and temperaturegradients can only be studied reliably on a pilotscale. To be applicable in a commercial plant,the process must be scaled up without loss ofefficiency (Seidel et al. 2004).
3.3. Dissimilatory metal reduction
Respiratory microorganisms obtain energy fromthe enzymatically controlled oxidation of electrondonors, such as glucose, acetate, or H2, coupledto the reduction of electron donors, such as O2,NO3
), Fe(III) and SO42) (Table 2). Recently,
several toxic metals and metalloids have joinedthe list of terminal electron acceptors: As(V) asarsenate, Se(VI) as selenate and U(VI) as uranylacetate (Lovley et al. 1991; Ahmann et al. 1994;Oremland et al. 1994). In addition chromium(VI)and plutonium(IV) appear to be reduced by respi-ratory enzymes, although respiratory growth withthese acceptors has not been proven (Lovley andPhillips 1994; Rusin et al. 1994). The importanceof these reductions for bioremediation lies in theprofound physical and chemical changes that themetals exhibit following reduction, creating
p py g y gy y)
131

opportunities for detoxification or for separationof the metallic compounds from soil matrices.
3.3.1. Selenium reductionSelenium contamination has gained particularattention in the Central Valley of California andother areas of the western United States wherenaturally selenium-rich soils are irrigated for agri-culture. Excess water is drained from the fieldsand carried to evaporation ponds, some in eco-logically sensitive areas such as the KestersonWildlife Refuge, where the selenium has becomeconcentrated enough to cause toxicity, deformi-ties, reproductive problems and death in water-fowl (Oremland et al. 1989; Macy 1994). Theselenium problem has inspired investigations intotwo bioremedial strategies: microbial reduction ofsoluble selenate into insoluble elemental selenium,discussed below, and microbial methylation ofinorganic species to form volatile products, con-sidered later in this chapter (see 3.4.1).
Selenium is a trace metalloid with a redoxchemistry similar to that of sulfur, possessing pri-mary oxidation states of VI, IV, O and –II. Sele-nate (SeO4
2)) is the form found most commonlyin soil porewaters (Frankenberger Jr. & Karlson1994a). Because the reduction potential of sele-nate (Se(VI)) to elemental selenium is sufficientlyhigh, researchers hypothesized that selenatereduction coupled to hydrogen or acetate oxida-tion could generate enough energy to support
anaerobic microbial metabolism (Table 2). Threeselenate-respiring bacterial strains have sincebeen isolated from selenate-rich-sediments: SeS,SES-3 and Thauera selenatis (Oremland et al.1994, 1989; Macy et al. 1993). Bacillus and Mi-crobacterium species have also been identifiedthat reduce, but do not necessarily respire, sele-nate (Combs et al. 1996).
Selenate and nitrate respirations proceedsimultaneously in T. selenatis. For bioremedialpurposes, the presence of nitrate is desirable,since nitrate respiration induces the nitrite reduc-tase that catalyzes selenate reduction to elementalselenium (DeMoll-Decker & Macy 1993). SES-3,in contrast, prefers nitrate as an electron accep-tor and will not respire selenate until nitrate isdepleted (Steinberg et al. 1992). The agriculturaldrainage waters contain about 50 mg/L nitrate,150 times the selenate concentration, making thisan important consideration (Macy et al. 1993).
As the mobile, highly toxic Se(VI) is the dom-inant species found in the Central Valley’s SanJoaquin River and the various evaporationponds (Oremland et al. 1989; Thompson-Eagle etal. 1989), microbial reduction has the potential toremove selenate from soil porewaters and immo-bilize it as Se(0) an insoluble, much less bioavail-able form. Studies of selenate reduction inevaporation pond surface sediment slurries haveshown that anaerobic conditions, indigenousmicroorganisms are able to remove up to 99.7%
Table 2. Free energies of electron acceptor reduction coupled to H2 oxidation
Reaction DG* (kcal/mol e))
1/4 O(g) + 1/2 H2 fi 1/2 H2O )23.55a
1/5NO3) + 1/5H+ +1/2H2 fi 1/10N2(g) + 1/5H2O )20.66a
1/2MnO2(s) + H+ + 1/2 H2 fi 1/2Mn2+ + H2O )22.48b
1/2UO22+ + 1/2 H2 fi H+ + 1/2UO2(s) )18.89c
1/2SeO42) +1/2H+ + 1/2H2 fi 1/2HSeO3
) + 1/2H2O )15.53b
1/2 CrO42) + 5/3H+ + 1/2 H2 fi 3/2Cr3) + 4/3H2O )10.76b
Fe(OH)3(s) + 2H+ + 1/2H2 fi Fe2+ + 3H2O )10.49b
1/4HSeO3) + 1/4H+ + 1/2 H2 fi 1/4Se + 3/4H2O )8.93b
1/2H2AsO4) + 1/2H+ + 1/2H2 fi 1/2H2AsO3 + 1/2H2O )5.51d
1/4SO42)+1/4H++1/2 H2 fi 1/4 HS)+1/2H2O )0.10a
PH2= 1066 atm; PN2
= 0.78 atm; PO2= 0.21 atm; pH = 7.0.
[NO�2 ]=[Mn2+]=[UO2þ2 ]=[SeO2�
4 ]=[HSeO�3 ] =[CrO2�4 ]=[Cr3+]=[Fe2+]=[HSeO�3 ]=[H2AsO�4 ] =[H2AsO3]=[SO2�
4 ]=[HS)]=[10)6] M.a Calculated from Zehnder & Stumm 1988.b Calculated from Morel & Hering 1993.c Calculated from Rusin et al. 1994.d Calculated from Vanysek 1995.
132

of the added selenate (approx. 300 lM) fromporewater within 7 days of incubation, withaddition of lactate, acetate and H2 stimulatingreduction rates (Oremland et al. 1989). In pondsediments, selenate reduction occurs primarilynear the surface, in the same region as denitrifi-cation and vertically above sulfate reduction.Areal rates of selenate removal from pond sedi-ment porewaters have been estimated at up to300lmol/m2/day (Figure 18).
Bioremediation of selenate contamination bymicrobial reduction and precipitation has at-tracted many advocates; nevertheless, the long-term stability of elemental selenium in oxic soilsis questioned (Frankenberger Jr. and Karlson,1994b). Because selenite and selenate are solubleand relatively mobile, aqueous extraction fromsoils may be a better alternative. Microbialreduction might then be used to recover seleniumfrom the aqueous phase. This technique has beenextremely successful in various wastewaters, con-sistently removing 95–98% of the Se (Lawsonand Macy, 1995; Macy et al. 1993)
3.3.2. Uranium reductionUranium contamination of soils and groundwa-ters has resulted from the release of wastes frommining, extraction from ores, nuclear fuel repro-cessing and ammunitions manufacture. Uranium
exists in most oxygenated wastes as the U(VI)uranyl ion UO2
2+, in reducing environmentssuch as groundwater (Macaskie, 1991). WhileU(VI) is soluble and immobile, U(IV) is mobile,and highly insoluble in most aqueous solutions(Lovley & Phillips 1992a, b).
Several years ago, the iron-respiring microor-ganisms Strain GS-15 and Alteromonas purefac-iens (later renamed Geobacter metallireducens andShewanella purefaciens, respectively) were shownto respire uranium as well as iron. The group ofuranium-reducing microorganisms now includesseveral sulfate-reducing bacteria, and mechanisticinvestigation of Desulfovibrio vulgaris has shownthat the respiratory electron transport proteincytochrome c, catalyzes the U(VI) reduction(Lovley et al. 1993b). Sulfate-reducing bacteriado not appear to obtain energy for growth fromthe process, however (Lovley et al. 1993a).
Microbial uranium reduction, clearly applica-ble to precipitation of uranium from wastewa-ters, has also found a role in uranium removalfrom soils. Because bicarbonate ions form verystrong, soluble complexes with U(VI), investiga-tors used a 100 mM solution to extract uraniumfrom contaminated soils, uranium ores and milltailings. The bicarbonate was not able to removethe uranium completely (20–94% recovery) al-though the extracted uranium might have been
Figure 18. Rates of reduction and porewater concentrations of sulfate, nitrate and selenate in an evaporation pond core from theSan Joaquin Valley. (a) Sulfate reduction; (b) denitrification; (c) selenate reduction. Values indicate the mean of 3 samples (Rep-rinted from Oremland et al. (1990), with permission. Copyright 1990 American Chemical Society).
133

the most dangerous fraction, as unrelated studieshave shown the bicarbonate removes a muchhigher percentage of soil uranium than is bio-available (Sheppard & Evenden 1992). To thisextract, kept anaerobic under N2ACO2 (80:20),H2 was added as an electron donor and Desulf-ovibrio desulfuricansas as the catalyst. The cellsreduced and precipitated 75–100% of the ura-nium from the leachate within 24 h, yielding avery pure, highly concentrated uraninite solid.The reduction was somewhat inhibited by dis-solved organic matter in the extract, as evidencedby a yellow color, but peroxide addition to oxi-dize the organic matter eliminated the color andthe inhibition (Phillips et al. 1995). A remarkablefeature of this microbial U(VI) precipitation isthat the Desulfovibrio cells can be freeze-driedand stored under air for up to six months with-out loss of uranium-reducing activity (Lovley &Phillips 1992b).
3.3.3. Plutonium reductionPlutonium, obtained from irradiation of uraniumand assembled into nuclear weapons, is an exten-sive soil contaminant near the production sites atHanford, WA and Rocky Flats, CO (Zorpette1996). Plutonium is also a product of nuclear en-ergy generation, although much of it can bereprocessed (Macaskie 1991). Like uranium, plu-tonium is a redox-active metal, with oxidationstates of III to VI known to exist in aqueoussolution. Oxides and hydroxides of Pu(IV), withextremely low solubilities estimated at ksp=1056.8
to 1057.8, appear to predominate in contaminatedsoils (Rusin et al. 1994).
Pu (III) hydroxide, while still quite insolubleat Ksp=1022.6, is more soluble than Pu(IV) com-pounds. Because the reduction of Pu(IV), as hy-drous PuO2(s) to Pu3+ is theorectically capable ofsupporting bacterial growth, with a reduction po-tential similar to that of Fe(III), researchers haveinvestigated the potential for plutonium reduc-tion and dissolution by iron-reducing Bacillusstrains. Experiments showed that cultures of B.circulans and B. polymyxa did indeed dissolve so-lid plutonium, amounting to 85–91% of the hy-drous PuO2(s) added, or 1.25 lmol Pu, in thepresence of 50 mM nitrilotriacetic acid (NTA).Reduction was the most likely means of dissolu-tion, although this was not conclusively demon-strated: plutonium-dependent growth of the
bacteria, indicating energy gain from the reduc-tion, was also not shown. With further develop-ment, including the use of biodegradablecomplexing agents in place of NTA, microbialreduction could provide a new pathway for in-situ plutonium remediation (Rusin et al. 1994).
3.3.4. Arsenic reductionMicrobial arsenic reduction, like plutoniumreduction, transforms a relatively immobile spe-cies into a more mobile one and thus has the po-tential to operate in conjunction with soilwashing to mobilize soil-bound arsenic. Inor-ganic arsenic in neutral aqueous solution occursas As(V), arsenate (H2AsO4), and As(III), arse-nite (H3AsO3). As(-III), occurring as arsine(AsH3), is a volatile reactive species formed bycertain fungi and bacteria (Cheng & Focht 1979).Arsenate adsorbs strongly to iron and manganeseoxyhydroxides and is thus very difficult to re-move from soils and sediments. Arsenite, al-though more toxic, is more mobile, suggestingthat reduction may promote arsenic mobilization(Bodek et al. 1988; Kuhn & Sigg 1993).
Recently, several species of arsenate-respiringbacteria have been isolated from arsenic-rich sub-strates, including Strain MIT-13 (renamed Geo-spirillum arsenophius) (Ahmann et al. 1994). Itsclose relatives that also reduce selenate, areStrain SES-3 (Laverman et al. 1995) Desulfoto-maculum auripigmentum (Newman et al. 1997)and Chrysiogenes arsenatis (Macy et al. 1996).Arsenic reduction is rapid in these microorgan-isms, measured at 2–5 mmol As-ml culture1 Æ day1
in both G. arsenophilus (Ahmann et al. 1994) andin Strain SES-3 (Laverman et al. 1995). Therespiratory action of G. arsenophilus is able todissolve arsenic from contaminated sediments(approximately 1000 ppm As) and other arsenicalsolids (Ahmann et al. 1997), and continuous-flowpilot-scale studies are in progress to maximizethe arsenic-mobilizing potentials of the microor-ganism and others indigenous to contaminatedsoils.
3.3.5. Chromium reductionCr(III) found at trace concentrations of4–90 ppm in pristine areas, is the most stableand abundant form of chromium in most soils.
134

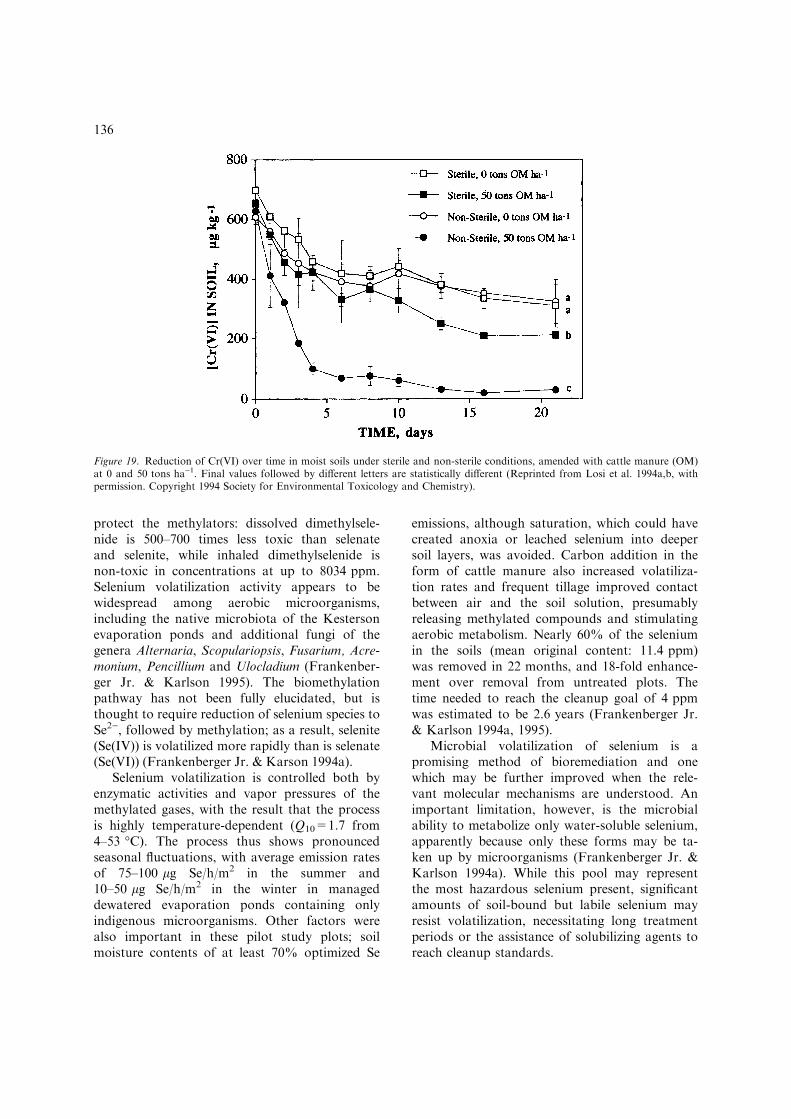
Kinetically it is quite inert, forming long-livedaqueous complexes with ammonia, sulfates, ha-lides and organic acids. At slightly acidic to alka-line pH, however, ionic Cr(III) species tend toprecipitate as amorphous Cr(OH3) if Fe3+ ispresent, with the result that much of the Cr(III)in soils is insoluble and immobile (Krishnamur-thy & Wilkens 1994). Although Cr(III) is anessential trace nutrient, it does not readily enterliving cells. Industrially-produced and -dis-charged Cr(VI), in contrast occurs as soluble,highly toxic and carcinogenic chromate and di-chromate. Hexavalent chromium is freely takenup by cells and reduced intracellularly to Cr(III),which appears to mutagenize. Because Cr(III) issignificantlly less toxic and less mobile thanCr(VI)-contaminated soil, numerous studies havefocused on reducing Cr(VI) in-situ, by chemical(e.g., ferrous iron) or biological means. Theintent is thus to ‘‘fix’’ the chromium in the soilin a harmless form. This approach requirescareful management, since reoxidation of Cr(III)to Cr(VI) is catalyzed by manganese oxidesand by molecular oxygen. The oxidation appearsto be quite slow, primarily limited by solubilityand mobility of the Cr(III)-laden soil is stillquite possible under oxidizing conditions. Thetwo essential requirements for safe chromiumremediation by reduction, therefore, are (1) apermanent reducing environment and (2) immo-bilization, by precipitation or by complexationwith degradation-resistant organic polymers, ofthe reduced chromium (Bartlett 1991; Krishna-murthy & Wilkens 1994; Losi et al. 1994a).
Bioremediation efforts to date have focusedon the study of Cr(VI)-reducing microorganisms.Numerous such bacteria have been isolated,including species of Bacillus, Desulfovibrio,Achromobacter, Aeromonas, Escherichia, Enterob-acter and Pseudomonas (Lovley & Phillips 1994;Turick et al. 1996). Mechanisms of chromatereduction in anaerobic bacteria appear to involverespiratory electron-transport enzymes (Bopp &Ehrlich 1988; Shen & Wang 1993), with convinc-ing evidence for catalysis by cytochrome c3 inDesulfovibrio vulgaris (Lovley & Phillips 1994).While it is thermodynamically possible for chro-mate reduction to generate enough energy tosupport respiration (Table 2), true chromate res-piration remains to be discovered (Lovley & Phil-lips 1994; Shen & Wang 1993; Turick et al.
1996). In aerobic chromate-reducers, reductionmay be fortuitously catalyzed by enzymes thathave other natural substrates (Cervantes 1991).
The presence of reactive electron donors ap-pears to be the single greatest factor promotingchromate reduction both chemical and biological,in soils. In one illustrative set of experiments,sterile soils alone reduced almost half of the800 lg/kg Cr(VI) present within 8 days. Additionof organic matter (cattle manure) improved thereduction to approximately 63%, and nativemicrobial activity in organic-amended samplesresulted in over 96% reduction, emphasizing theeffectiveness of biological catalysis (Figure 19).Studies simulating field conditions have sup-ported these results, confirming (1) the ubiquityof indigenous Cr-reducing microorganisms in avariety of soil, both contaminated and clean, (2)the importance of irrigation to maintain reducingconditions and (3) the proportionality betweenorganic matter loading and Cr(VI) reduction(Cifuentes et al. 1996; Losi et al. 1994a, b), sug-gesting that Cr(VI) bioremediation by reductionin soil holds great promise in cases where reoxi-dation can be permanently prevented.
3.4. Volatilization
3.4.1. Selenium volatilizationMicrobial transformation of selenium into vola-tile methylated species, principally dimethylsele-nide (CH3)2Se with smaller amounts ofdimethyldiselenide (CH3)2(Se)2 and dimethylsele-none (CH3)2 (SeO2), removes selenium from soiland disperses it into the atmosphere. Airbornedimethylselenide (DMSe) reacts with hydroxylradicals and ozone to yield oxidized productswhich are then thought to become associatedwith particles or aerosols that have residencetimes of several days, during which they may bedispersed over great distances (Frankenberger Jr.& Karlson 1994a). Microbial volatilization wasfirst recognized as an important part of the sele-nium biogeochemical cycle in 1987, and it is nowestimated that soils and plants emit 1700–3300 metric tons of selenium into the atmospherein the Northern hemisphere each year (Cooke &Bruland 1987; Frankenberger Jr. & Karlson1995).
Selenium methylation produces substances ofgreatly reduced toxicity and is thus thought to
135
protect the methylators: dissolved dimethylsele-nide is 500–700 times less toxic than selenateand selenite, while inhaled dimethylselenide isnon-toxic in concentrations at up to 8034 ppm.Selenium volatilization activity appears to bewidespread among aerobic microorganisms,including the native microbiota of the Kestersonevaporation ponds and additional fungi of thegenera Alternaria, Scopulariopsis, Fusarium, Acre-monium, Pencillium and Ulocladium (Frankenber-ger Jr. & Karlson 1995). The biomethylationpathway has not been fully elucidated, but isthought to require reduction of selenium species toSe2), followed by methylation; as a result, selenite(Se(IV)) is volatilized more rapidly than is selenate(Se(VI)) (Frankenberger Jr. & Karson 1994a).
Selenium volatilization is controlled both byenzymatic activities and vapor pressures of themethylated gases, with the result that the processis highly temperature-dependent (Q10=1.7 from4–53 �C). The process thus shows pronouncedseasonal fluctuations, with average emission ratesof 75–100 lg Se/h/m2 in the summer and10–50 lg Se/h/m2 in the winter in manageddewatered evaporation ponds containing onlyindigenous microorganisms. Other factors werealso important in these pilot study plots; soilmoisture contents of at least 70% optimized Se
emissions, although saturation, which could havecreated anoxia or leached selenium into deepersoil layers, was avoided. Carbon addition in theform of cattle manure also increased volatiliza-tion rates and frequent tillage improved contactbetween air and the soil solution, presumablyreleasing methylated compounds and stimulatingaerobic metabolism. Nearly 60% of the seleniumin the soils (mean original content: 11.4 ppm)was removed in 22 months, and 18-fold enhance-ment over removal from untreated plots. Thetime needed to reach the cleanup goal of 4 ppmwas estimated to be 2.6 years (Frankenberger Jr.& Karlson 1994a, 1995).
Microbial volatilization of selenium is apromising method of bioremediation and onewhich may be further improved when the rele-vant molecular mechanisms are understood. Animportant limitation, however, is the microbialability to metabolize only water-soluble selenium,apparently because only these forms may be ta-ken up by microorganisms (Frankenberger Jr. &Karlson 1994a). While this pool may representthe most hazardous selenium present, significantamounts of soil-bound but labile selenium mayresist volatilization, necessitating long treatmentperiods or the assistance of solubilizing agents toreach cleanup standards.
Figure 19. Reduction of Cr(VI) over time in moist soils under sterile and non-sterile conditions, amended with cattle manure (OM)at 0 and 50 tons ha)1. Final values followed by different letters are statistically different (Reprinted from Losi et al. 1994a,b, withpermission. Copyright 1994 Society for Environmental Toxicology and Chemistry).
136

3.4.2. Mercury volatilizationMercury resistance systems have been found inall bacterial groups tested and consistently in-clude an operon of six genes: merR, a regulatorygene: merD, a second regulator, merP and merT,encoding proteins that transport mercuric ionsinto the cell; merA, encoding a mercuric reduc-tase that transforms Hg2+ to volatile Hg0 andmerB, encoding an organomercurial lyase thatcleaves Hg from organomercury compounds(Figure 20). The bioremedial potential of thissystem lies in the ability of the bacteria to re-move highly toxic mercuric ions from a sub-strate, releasing them as elemental mercury ofgreatly reduced toxicity that can then be trappedor dispersed into the atmosphere. The system isquite well understood and although it is ubiqui-tous among bacteria, it is showing greatest prom-ise within transgenic plants for mercury removalfrom soils (Rugh et al. 1996; Silver 1994).
4. Metal-microbe interactions and their impact
on bioremediation of metal contaminated soils
and sediments
Estimates of the global market for the cleanupand prevention of metal contamination vary, butconservative calculations suggest that the currentmarket for metal bioremediation may rise to$200 billion in the US alone by 2005. The emerg-ing market for the clean up of radioactive con-tamination in the US may already be worth asmuch as $300 billion (McCullough et al. 1999).
Unfortunately, existing chemical techniques arenot always economical or cost effective for theremediation of water or land contaminated withmetals and radionuclides. Current strategies forland contaminated with metals include the use of‘‘dig and dump’’ approaches that only move theproblem to another site, are expensive andimpractical for large volumes of soil or sediment.Likewise soil washing, which removes the small-est particles that bind most of the metals, is use-ful but can be prohibitively expensive for somesites. ‘‘Pump and treat’’ technologies rely on theremoval of metals from the site in an aqueousphase which is treated ex situ (e.g. above land).These approaches can cut down on excavationcosts but are still expensive, and metal removalcan be inefficient. A potentially economical alter-native is to develop biotechnological approachesthat could be used in the sediment or soil (insitu) to either extract the metals or stabilise themin forms that are immobile or non-toxic.
In addition to developing biotechnologies totreat contaminated land, there is also considerableinterest in more effective techniques that can beused to treat water contaminated with metalsfrom a range of industrial processes. Problemsinherent in currently used chemical approaches in-clude a lack of specificity associated with some ionexchange resins, or the generation of large quanti-ties of poorly settling sludge through treatmentwith alkali or flocculating agents. This sectiongives an overview of metal–microbe interactions,and describes how they could be harnessed toclean up metal contaminated, soil and sediments.
Figure 20. Genes and polypeptides of mercury resistance systems from Gram-negative Serratia sp. Aa, length of gene products inamino acids; P/O, promoter/operator region (Adapted from Ji & Silver 1995, with permission of the Society for Industrial Micro-biology).
137

4.1. Metal and radionuclide contamination
The US EPA (http://www.epa.gov) lists fourmain groups of priority compounds which in-clude polychlorinated biphenyls (PCBs), baseneutrals and acids (e.g. phenol and naphthalene)and volatile organic compounds (VOCs), includ-ing carbon tetrachloride and BTEX contami-nants. Trace metals that are recoverable anddissolved are also listed; these include antimony,chromium, mercury, silver, arsenic, copper, nick-el, thallium, beryllium, lead, selenium, zinc andcadmium. In addition to these toxic metals, ra-dionuclides are priority contaminants at nuclearinstallations worldwide, and will require treat-ment. For example, in the US Department ofEnergy’s 120 sites contains 1.7 trillion gallons ofcontaminated ground water and 40 million cubicmetres of contaminated soil and debris. Morethan 50% of the sites are contaminated with ra-dionuclides, with the priority radionuclides beingcesium-137, plutonium-239, strontium-90, techne-tium-99 and uranium-238 and uranium-235, inaddition to toxic heavy metals including chro-mium, lead and mercury (McCullough et al.1999). The US DOE Natural and AcceleratedBioremediation (NABIR) program is notable inits support of emerging biotechnological ap-proaches for dealing with this legacy waste. InEurope, the EU Water Framework Directive(2000/60/EC) is the principal driver for environ-mental legislation, and also recognizes metalsamongst the 32 substances on the priority list.These include lead, nickel, mercury cadmium andtin and their compounds.
4.2. Biosorption
The term biosorption is used to describe themetabolism-independent sorption of heavy metalsand radionuclides to biomass. It encompassesboth adsorption; the accumulation of substancesat a surface or interface and absorption; the al-most uniform penetration of atoms or moleculesof one phase forming a solution with a secondphase (Gadd & White 1989). Both living anddead biomass are capable of biosorption and li-gands involved in metal binding include car-boxyl, amine, hydroxyl, phosphate andsulfhydryl groups. Biosorption of metals hasbeen reviewed extensively (McHale & McHale
1994; Tobin et al. 1994; Volesky & Holan 1995;Beveridge et al. 1997a,b; Lloyd & Macaskie2000). Biosorption is generally rapid and unaf-fected over modest temperature ranges and inmany cases can be described by isotherm modelssuch as the Langmuir and Freundlich isotherms(Volesky & Holan 1995). Gadd and White(1989), however, noted that more complex inter-actions are difficult to model because the adsorp-tion of solutes by solids is affected by factorsincluding diffusion, heterogeneity of the surfaceand pH. An additional isotherm, the Brunauer–Emmett–Teller (BET) isotherm, which assumesmultilayer binding at constant energy has alsobeen used to describe metal biosorption (deRome & Gold 1991; Andres et al. 1993). Thismodel assumes that one layer need not necessar-ily be completely filled before another is com-menced. Further insight is offered by Andreset al. (1993), who summarize that each adsorp-tion layer of the BET model can be reduced toLangmuir behavior with homogeneous surfaceenergy, in contrast to the adsorption energyrequirements of the Freundlich isotherm. Otherstudies have used complex multistage kinetic ap-proaches to model biosorption (Weidemann et al.1981; Treen-Sears et al. 1984).
Ultimately, however, the amount of residualmetal remaining in solution at equilibrium isgoverned by the stability constant of the metal–ligand complex (Macaskie 1991), and the onlyway to change the equilibrium position is tomodify the binding ligand to one which has agreater binding affinity for the given metal, or totransform the metal from a poorly sorbing spe-cies to one which has a higher ligand-bindingaffinity, e.g. by a change of metal valence. Alter-natively some new studies have applied the toolsof molecular biology to enhance metal sorptionand do warrant attention. For example, a mousemetallothionein was targeted to the outer mem-brane of a metal-resistant Ralstonia eutropha iso-late (Valls et al. 2000). The engineered strainaccumulated more Cd2+ than its wild type coun-terpart, and offered tobacco plants some protec-tion from Cd2+ when inoculated intocontaminated soil (Valls et al. 2000). A surfacedisplay technique has also been used successfullyto generate ZnO binding peptides fused to fim-brae on the surface of cells of Escherichia coli(Kjaergaard et al. 2000). Finally, Gram-positive
138

bacteria (Staphylococci) have also been engi-neered to produce surface exposed peptides, ableto bind Hg2+ and Cd2+ (Samuelson et al. 2000).Enhanced uptake of cadmium and mercury byEscherichia coli expressing a metal-binding motifhas also been reported (Pazirandeh et al. 1998).
Future studies could usefully compare theperformance of such engineered systems withother more traditional biosorbants. However, ithas been noted that despite the relatively longtime during which biosorption has been studied,and considerable early interest, there has been re-duced interest in commercialising this type oftechnology, and the authors are aware of no cur-rent commercial applications of this type oftechnology.
4.3. Metabolism-dependent bioaccumulation
Energy-dependent metal uptake has been demon-strated for most physiologically important metalions, and some toxic metals and radionuclidesenter the cell as chemical ‘‘surrogates’’ usingthese transport systems. Monovalent cationtransport, for example K+ uptake, is linked tothe plasma membrane-bound H+-ATPase via themembrane potential, and is, therefore, affectedby factors that inhibit cell energy metabolism.These include the absence of substrate, anaerobi-osis, incubation at low temperatures and thepresence of respiratory inhibitors such as cyanide(White & Gadd 1987). The requirement for met-abolically active cells may, therefore, limit thepractical application of this mode of metal up-take to the treatment of metals and radionuclideswith low toxicity and radioactivity. For example,in the study of White and Gadd (1997), increas-ing metal concentrations inhibited H+ pumping,potentially deenergizing the cell membrane andreducing cation uptake. Although the presence ofmultiple transport mechanisms of differing affini-ties may cause added complication, metal influxfrequently conforms to Michaelis–Menten kinet-ics (Borst-Pauwels 1981).Once in the cell, metals may be sequestered bycysteine-rich metallothioneins (Higham et al.1984; Turner & Robinson 1995) or, in the case offungi, compartmentalized into the vacuole (Gadd& White 1989; Okorov et al. 1977). In this con-text, it should be emphasized that the uptake ofhigher mass radionuclides, e.g. the actinides into
microbial cells has been reported sporadicallyand remains poorly characterised (Lloyd & Ma-caskie 2002; Lloyd & Macaskie 2000). Metabo-lism-dependent bioaccumulation of metals bymicroorganisms has not been used commerciallyfor bioremediation purposes.
4.4. Enzymatically catalysed biotransformation
Microorganisms can catalyze the transformationof toxic metals to less soluble or more volatileforms. For example, the microbial reduction ofCr(VI) to Cr(III), Se(VI) to Se(0), V(V) to V(III),Au(III) to Au(0), Pd(II) to Pd(0), U(VI) toU(IV) and Np(V) to Np(IV) results in metal pre-cipitation under physiological conditions and hasbeen reviewed in detail recently (Lloyd 2003). Inmany cases the high valence metal can be used asan electron acceptor under anoxic conditions. Agood example here is the reduction of solubleU(VI) to insoluble U(VI) by Fe(III)-reducingbacteria (Lovley et al. 1991), which has been har-nessed recently to remediate sediments contami-nated with uranium in situ. Metal detoxificationis also possible through biotransformations oftoxic metals. The bioreduction of Hg(II) to rela-tively non-toxic Hg(0) is perhaps the best studiedexample. Finally, in addition to redox biotrans-formations, biomethylation may also increase thevolatility of metals, with the methylation of mer-cury, cadmium, lead, tin, selenium and telluriumrecorded (Diels et al. 1995). These mechanismscould also offer potential use for the in situ reme-diation of contaminated soil (Losi & Franken-berger 1997).
As mentioned above, microbes have evolvedsophisticated approaches to deal with toxic me-tal (Bruins et al. 2000), often involving redoxtransformations of the toxic metal. Perhaps thebest studied metal resistance system is encodedby genes of the mer or mercury resistance oper-on (Hobman et al. 2000). Recent studies haveconfirmed the biotechnological potential of thiswidespread resistance determinant. Hg(II) isbound in the periplasm of Gram-negative bacte-ria by the MerP protein, transported into thecell via the MerT transporter, and detoxified byreduction to relatively non-toxic volatile elemen-tal mercury by an intracellular mercuric reduc-tase (MerA). Mer proteins are expressed underthe regulation of the activator protein, MerR,
139

which binds Hg(II) and activates gene expres-sion. Mercury-resistant bacteria and the proteinsthat they encode have been used recently for thebioremediation of Hg-contaminated water andin the development of biosensors for bioavail-able concentrations of Hg(II) (Bontidean et al.2002). Such sensors may prove very useful inidentifying the need for metal remediation,which will be dictated in many cases by theconcentration of bioavailable toxic metals in agiven soil or water matrix, and also in definingthe end point for bioremediation efforts. Focus-ing on Hg biosensors, several different compo-nents of the mer system have been used inprototype sensors, including the NADPH-depen-dent mercuric reductase (MerA) in an enzyme-linked biosensor (Eccles et al. 1997), the merregulatory region in a whole cell biosensor (Gei-selhart et al. 1991), and the MerR protein in acapacitance biosensor (Bontidean et al. 2000).
4.5. Biomineralization via microbially generatedligands
Microorganisms are able to precipitate metalsand radionuclides as carbonates and hydroxidesvia plasmid-borne resistance mechanisms, where-by proton influx countercurrent (antiport) to me-tal efflux results in localized alkalinization at thecell surface (Diels et al. 1995; Van Roy et al.1997). Alternatively, metals can precipitate withenzymatically generated ligands, e.g. sulfide(Barnes et al. 1991), or phosphate (Macaskieet al. 1992). The concentration of residual freemetal at equilibrium is governed by the solubilityproduct of the metal complex (e.g. 10)20 to 10)30
for the sulfides and phosphates, higher for thecarbonates). Most of the metal should be re-moved from solution if an excess of ligand issupplied. This is difficult to achieve using chemi-cal precipitation methods in dilute solutions; theadvantage of microbial ligand generation is thathigh concentrations of ligand are achieved in jux-taposition to the cell surface, that can also pro-vide nucleation foci for the rapid onset of metalprecipitation; effectively the metals are concen-trated ‘‘uphill’’ against a concentration gradient.This was demonstrated using the gamma isotope241Am supplied at an input concentration ofapproximately 2.5 parts per billion; approxi-mately 95% of the metal was removed as bio-
mass-bound phosphate (Macaskie et al. 1994),with the use of gamma-counting permittingdetection at levels below those of most analyticalmethods. In many cases, the production of the li-gand can also be fine-tuned by the application ofMichaelis–Menten kinetics.
Sulfide precipitation, catalysed by a mixed cul-ture of sulfate-reducing bacteria, has been uti-lized first to treat water co-contaminated bysulfate and zinc (Barnes et al. 1991) and also,soil leachate contaminated with sulfate alongsidemetal and radionuclides (Kearney et al. 1996).Ethanol was used as the electron donor for thereduction of sulfate to sulfide in both examples.A later study has confirmed the potential of inte-grating the action of sulfur cycling bacteria(White et al. 1998). In the first step of a two-stage process, sulfur-oxidising bacteria were usedto leach metals from contaminated soil via thegeneration of sulfuric acid, and in the secondstep the metals were stripped from solution in ananaerobic bioreactor containing sulfate-reducingbacteria (Kearney et al. 1996). The ubiquitousdistribution of sulfate-reducing bacteria in acid,neutral and alkali environments (Postgate 1979),suggests that they have the potential to treat avariety of effluents, while the ability of theorganisms to metabolize a wide range of electrondonors, may also allow co-treatment of otherorganic contaminants. Recent work by PaquesBV of the Netherlands have confirmed the poten-tial of sulfate-reducing bacteria to treat metalwaste in ex situ bioreactors for the treatment ofa wide range of metal-contaminated waters.
Bioprecipitation of metal phosphates viahydrolysis of stored polyphosphate by Acineto-bacter spp. is dependent upon alternating aerobic(polyphosphate synthesis) and anaerobic (poly-phosphate hydrolysis and phosphate release)periods (Boswell et al. 2001). This obligately aer-obic organism is fairly restricted in the range ofcarbon sources utilized, but the preferred sub-strates (acetate and ethanol) are widely andcheaply available. The best-documented organismfor metal phosphate biomineralization, a Citro-bacter sp., (now reclassified as a Serratia sp. onthe basis of molecular methods, the presence ofthe phoN phosphatase gene and the productionof pink pigment under some conditions(Pattanapipitpaisal et al. 2002) grows well oncheaply available substrates and viable cells are
140

not required for metal uptake since this relies onhydrolytic cleavage of a supplied organic phos-phate donor (Macaskie et al. 1992). The expenseof adding organic phosphate was calculated to bethe single factor which limited the economic via-bility of this approach (Roig et al. 1995); more-over organophosphorus compounds are oftenhighly toxic. A possible alternative phosphatedonor, tributyl phosphate (TBP), is used widelyas a cheap solvent and plasticizer but its degra-dation is more difficult because this compound isa phosphate triester, with three cleavage eventsper molecule required to liberate one molecule ofphosphate. TBP is biodegradable; this activitywas unstable (Thomas & Macaskie 1998) butbiogenic phosphate from TBP hydrolysis washarnessed to the removal of uranium from solu-tion in a flow-through system (Thomas & Macas-kie 1996). The enzymatic activity responsible forTBP hydrolysis remains obscure but a recent re-port of TBP hydrolysis by enzymatic degradationwas studied using cell free extracts of Serratiaodorifera (Berne 2004).
As an alternative to TBP and glycerol 2-phos-phate the possibility exists for the harnessing ofphytic acid degrading organisms to metal accu-mulation. Phytic acid, widely and cheaply avail-able as a natural plant product, accounts for upto 50% of the total organic P in soils (Rodriguez& Fraga 1999). The additional advantage of phy-tic acid is that it contains 6 phosphates/molecule(c.f. 1 phosphate/molecule in TBP and G2P).Several phytases have been cloned and character-ized, including one from the related organism E.coli (Greiner et al. 1993) with the phytase reac-tion mechanism described. Preliminary studieshave suggested that E. coli phytase can be ap-plied to metal removal (Patersen–Beedle et al.unpublished).
4.6. Microbially enhanced chemisorption of heavymetals
Microbially enhanced chemisorption of heavymetals (MECHM) is a generic term to describe aclass of reactions whereby microbial cells firstprecipitate a biomineral of one metal (‘primingdeposit’). The priming deposit then acts as anucleation focus, or ‘host crystal’ for the sub-sequent deposition of the metal of interest (‘tar-get’ metal), acting to promote and accelerate
target metal precipitation reactions (Macaskie etal. 1996). The priming deposit is made initiallyby the sulfide or phosphate biomineralizationroutes described above.
With addition of Fe as the precipitant metalto sulfate-reducing bacteria producing H2S, cell-bound FeS then acts as a sorbent for ‘target’metals (Ellwood et al. 1992; Watson & Ellwood1994, 1988). The use of FeS is notable becausethis provides the mechanism for rapid biomassseparation from the liquor via the magnetic prop-erties of Fe in a high-gradient magnetic separator(Ellwood et al. 1992; Watson & Ellwood 1994;Watson & Ellwood 1988).
Metal phosphate can also be used as thepriming deposit, in two ways. LaPO4 pre-depos-ited onto metal-accumulating Citrobacter sp. viabiogenic phosphate release can be used as the‘priming’ deposit for subsequent deposition ofactinide phosphates (Macaskie et al. 1994). Also,by use of a priming deposit with an appropriate,well-defined cystalline lattice, the target metalcan be intercalated within the ‘host’ lattice, effec-tively by a mechanism of bioinorganic ion-ex-change, well-established as a chemical process(Clearfield 1988; Pham-Thi & Columban 1985;Pozas-Tormo et al. 1987) and best-characterizedbiologically in the case of Ni2+ removal into bio-mass-bound HUO2PO4ÆnH2O (Bonthrone et al.1996). Within the context of nuclear waste reme-diation, the approach of MECHM, using bio-genic hydrogen uranyl phosphate as the ‘host’matrix, and using either intercalative ion ex-change or co-crystallisation, has been applied tothe removal of Cs, Co and Sr from ‘surrogate’solutions and the isotopes 137Cs, 60Co and 90Srfrom real nuclear wastes in South Korea (Pater-son–Beedle et al. unpublished).
4.7. Metal remediation through biodegradation ofassociated organic compounds
Citrate has found use as a chelating agent indecontamination operations forming highly solu-ble metal–citrate complexes, which can be de-graded by microorganisms resulting insubsequent re-precipitation of metals (Francis1994). Early work suggested that the type ofcomplex formed between the metal and citricacid plays an important role in determining its
141

biodegradability, with the binuclear uranium–cit-rate complex being recalcitrant to microbial deg-radation (Gadd & White 1989), but subsequentphotodegradation was possible for these prob-lematic complexes. This opened up the way for amulti-stage process for the biodegradation ofmixed metal–citrate complexes via microbiologi-cal and photochemical steps (Francis 1994). Cul-tures of Pseudomonas aeruginosa and P. putidawere also able to grow using a range of metal–citrate complexes as carbon, with metal precipita-tion promoted by the addition of inorganic phos-phate (Thomas et al. 2000). A unique aspect ofthis study was the use of P. putida to treat Ni–citrate waste generated by cleaning a bioinorgan-ic ion exchange column previously used to re-move Ni. Another industrial chelating agentethylenediaminetetraacetate (EDTA) also formsvery strong complexes with di- and trivalent met-als that are degraded by a mixed microbial popu-lation (Thomas et al. 1998) and by bacterialstrain DSM 9103 (Satroutdinov et al. 2000). Thebiodegradation of toxic organotin compoundsused as biocides and antifouling agents has alsoreceived recent attention (e.g. the degradation oftributyl tin to di- and monobutyl tin reviewed inGadd (2000)).
4.8. In-situ versus ex-situ remediation
Although most of the processes described thusfar have been developed primarily for ex situtreatment of contaminated water in situ remedi-ation offers the potential for low cost treatmentof groundwater contamination with metals. Exsitu remediation strategies such as pump andtreat systems remain a method of choice formetal-contaminated sites and can be effective atremoving significant quantities of contaminantmetals from heavily contaminated (and highlyconcentrated) areas. For low contaminant con-centration scenarios in situ immobilization ofcontaminants has been proposed as an attractivepotential treatment option (Lovley et al. 1991;Abdelouas et al. 1998a, b, 1999; Finneran et al.2002).
In situ immobilization of contaminant metalsrelies on the stimulation of anaerobic microbialcommunities within the subsurface. Anaerobicmicroorganisms enzymatically reduce contami-nant metals such as uranium, chromium, techne-
tium, vanadium and metalloids such as seleniumthereby converting these metals to less solubleforms (Lloyd et al. 2000; Lovley & Coates 2000;Anderson & Lovley 2002; Carpentier et al. 2003;Ortiz-Bernard et al. 2004). Stimulation of metalreduction within the subsurface offers a relativelysimple method to remove metal contaminantsfrom groundwater, thereby limiting or greatlyreducing further transport of contaminants down-gradient (Lovley et al. 1991; Lovley 1993; Ander-son & Lovley 2002). Efforts to understand andcontrol in situ metal reduction are currently activeareas of research within the U.S. Department ofEnergy (U.S. Department of Energy, O.o.B.a.E.R2004) particularly in light of the recent focus onthe potential ecological impacts of ex situ remedi-ation practice (Whicker et al. 2004).
A key issue in the development of in situ bio-remediation systems for metal contaminants isthe identification of target groups of microorgan-isms within the subsurface whose known physiol-ogy has the potential to affect contaminant metalmobility. Both Fe(III)- and sulfate-reducingorganisms are known to enzymatically reducecontaminant metals such as U(VI), Cr(VI),Co(III) and Tc(VII) in laboratory cultures (Lov-ley et al. 1991; Lovley & Phillips 1992a, b; Gorby& Lovley 1992; Lovley 1993; Caccavo Jr. et al.1994; Lloyd & Macaskie 1996; Gorby & Bolton1998; Tebo & Obraztsova 1998; Lloyd et al. 2000)but for remediation purposes it is necessary todemonstrate that the appropriate microorganismsare present within the contaminated subsurface.Members of the Geobacteraceae (d-proteobacte-ria) are of particular note as these organisms havebeen identified as a dominant group within sedi-ments upon the stimulation of Fe(III)-reducingconditions (Snoeyenbos-West et al. 2000; Finner-an et al. 2002; Holmes et al. 2002; Andersonet al. 2003) and in contaminated sediment whereFe(III) reduction is a dominant process (Ander-son et al. 1998; Roling et al. 2001; Cummingset al. 2003). Geobacteraceae were also recently de-tected within the groundwater of an uranium-contaminated aquifer during a test of stimulatedin situ U(VI) reduction (Anderson et al. 2003).Geobacteraceae, while best known as dissimila-tory Fe(III)-reducing bacteria, also couple theoxidation of simple organics to the reduction ofcontaminant metals such as uranium, chromium,technetium and vanadium (Chen & Hao 1998;
142

Lloyd et al. 2000; Anderson & Lovley 2002; Or-itz-Bernad et al. 2004). Additionally, the productof dissimilatory Fe(III) reduction, Fe(II), is a po-tential abiotic reductant for contaminant metalssuch as Tc(VII), V(V) and Cr(VI) (Fendorf et al.2000; Lloyd et al. 2000; Oritz-Bernad et al. 2004).The demonstrated enrichment of Geobacteraceaewithin an aquifer during an in situ trial of stimu-lated uranium reduction and the potential abioticbenefits of in situ Fe(II) production suggest Geob-acteraceae to be a target group of microorgan-isms within the subsurface for removing metalsfrom groundwater. Further investigation of stim-ulated subsurface metal reduction will likely re-veal other groups of bacteria, such as sulfatereducers, that could also play an important rolein contaminant metal reduction and stabilizationwithin subsurface environments.
5. Role of biofilms in bioremediation of metal
polluted soils and sediments
5.1. Biofilms in natural environment
Bacteria are widespread in soil and aquatic envi-ronments and are predominantly found in bio-film communities. A biofilm may be defined as agelatinous extracellular polymeric substance(EPS) of biological origin containing a consor-tium of bacteria, fungi, and algae. Biofilms arepresent at the solid–aqueous interface throughoutthe aquatic environment (Lappin-Scott et al.1993) and typically at the sediment–water inter-face (Davison et al. 1997; Forstner, 2004). Bio-films form when bacterial consortia attach tomineral surfaces and produce films of hydratedextracellular polymers (Jackson et al. 1999). Oncebiofilms form, the bacterial-mineral micro-aggre-gates create complex interfaces with the sur-rounding aqueous solution. The biofilms may actas an ‘‘insulating layer’’ between the solution andthe mineral surface or form ‘‘micro-environ-ments’’ in which the local solution conditions aredifferent than those in the bulk solution (Glazeret al. 2002). Reactive functional groups, such ascarboxyl, hydroxyl, amino and phosphoryl moie-ties, present on the bacterial surfaces and exo-polysaccharide matrix, can provide a large arrayof binding sites for metals (Flemming & Leis2002). For instance, autotrophic biofilms have
been observed to sequester metals at 4–5 ordersof magnitude above the concentration in the sur-rounding waters (Liehr et al. 1994). Biofilms maysimultaneously block surface sites on the under-lying substrate (Templeton et al. 2003a,b). Inaddition, bacterial activity may catalyze thetrans-formation of toxic metals into less (ormore) toxic species or enhance the dissolution ofthe underlying mineral substrate and bulk solu-tion surrounding of the biofilm (Beyenal et al.2004; Labrenz & Banfield 2004). All of thesecombined interactions may strongly affect themechanisms and kinetics of metal sorption reac-tions in soils and aquatic environments (Newman& Banfield 2002; Gadd 2004).
However, biofilms that form on solid phasesin natural contaminated environment sites maydiffer significantly in form and function fromthose found in other environments described todate. So far biofilms have been mainly used toremove inorganic contaminants from aqueoussystems (industrial wastewaters) (Diels et al.2003, Remoudaki et al. 2003; Wagner-Dobler2003; van Hullebusch et al. 2003; Bender et al.2004, Bender & Phillips 2004). However, theybeen also shown to influence the mobility ofinorganic contaminants in subsurface environ-ment such as soils and sediments. Ecological suc-cess of biofilms and their broad array ofmicrobial activities suggest that these microbialecosystems might be useful to bioremediation ofenvironmental pollutants as described in thefollowing subsection.
5.2. Biofilm and metals/radionuclidesimmobilization
Only the main mechanisms involved in theimmobilization of metal and radionuclides, asshown Figure 21, will be described in the follow-ing subsection.
5.2.1. Passive processes: biosorption of metals bybiofilmsMicroorganisms adsorb and concentrate manymetallic cations through electrostatic interactionswith anionic carboxyl and phosphoryl groups inthe cell wall. Biosorption consists of severalmechanisms, mainly ion exchange, chelation,adsorption, and diffusion through cell walls andmembranes. Biosorption encompasses those
143

physico-chemical mechanisms by which metalspecies and radionuclides are removed from anaqueous solution. This phenomenon is oftenattributed to the binding of metals onto the bac-terial cell surface (White et al. 1995; Cox et al.1999). In addition, however, the EPS that sur-round cells are also very reactive and can readilybind metals such as iron (Konhauser et al. 1993,1994). A growing body of literature shows evi-dence that EPS bind and concentrate a range ofinorganic contaminants. EPS is a highly hydratedmaterial (�99% water) possessing an extremelyporous fibrillar structure that renders it highlyadsorptive. The EPS consist of polysaccharides,proteins, lectins, lipids, nucleic acids, etc... Thestructure of the EPS-matrix is considerablystronger in the presence of different cations,which interact with exposed carboxyl groups onthe EPS. EPS can bind a wide variety of metals,including Pb, Sr, Zn, Cd, Co, Cu, Mn, Mg, Fe,Ag, and Ni (Ferris et al. 1989). The sorptioncapacity of biofilms can be attributed to thecomplex or chelate formation of the EPS-matrixwith different cations and the uptake of cations
by algae and bacteria living in the biofilms. Thebinding capacity of a given EPS-matrix also de-pends on its physical state (gel versus slime ver-sus dissolved state) and the pH of the water. Inorder to reach stronger binding of the metal ions,the gel structure and higher pHs (basic condi-tions) are favourable. Ferris et al. (1989) re-ported that near neutral pH, microbial biofilmsconcentrated metals up to 12 orders of magni-tude higher than observed under acidic condi-tions. Neutral to alkaline microenvironments arecommonly produced within microbial mats andbiofilms through such processes as photosynthe-sis and sulfate reduction (Krumbein 1979).
5.2.2. Microbial mineralizationMinerals formed in association within biofilmsare commonly referred to as biominerals, e.g.sulfate, phosphate, carbonate, sulfide or silicateions (Schultze-Lam et al. 1996). Given the abilityof EPS to concentrate various metals, it is notsurprising that bacteria have been implicated in awide variety of biomineralization processes. As aresult of cellular metabolism, microorganisms
Figure 21. Biofilm–metal interactions which have an impact on bioremediation of metal contaminated soils and sediments (Mod-ified from van Hullebusch et al. 2003).
144

alter the chemical microenvironment around thecell, modulating the pH, as well as the concentra-tion of a variety of organic and inorganic sol-utes. This can induce the large-scale precipitationof authigenic minerals in natural environments(Geesey et al. 1989; Beveridge et al. 1997a,b;Douglas and Beveridge, 1998; Langley & Beve-ridge 1999). For instance, stromatolites, made ofcalcium carbonate precipitates, are intimatelyassociated with biofilms that comprise an abun-dant mix of cyanobacteria, bacteria and diatoms(Douglas & Beveridge 1998; Arp et al. 2001).Sometimes nanomineral phases can even form di-rectly in the bacterial cytoplasm as has been de-scribed for acanthite (Ag2S) (Klaus et al. 1999)or amorphous iron hydroxide (Fortin 2004). Thetolerance of biofilms to high metal concentra-tions may also be due to their ability to precipi-tate insoluble metal salts outside the cells assulfides, phosphates, carbonates, oxides orhydroxides (Bender et al. 1995; Douglas & Beve-ridge 1998).
5.2.3. Oxidation and reductionInorganic contaminant entrapment and adsorptionby iron and manganese oxides.A microbial biofilm consortium catalyses oxida-tion of Fe(II) in acidic and near neutral solutionsto produce iron (Fe(III)) hydroxides (Brownet al. 1998b, 1999). Iron oxide precipitates associ-ated with biofilm EPS were identified as ferrihy-drite in both pH conditions but the precipitatemorphology and minor elemental compositionwas pH dependent (Ferris et al. 1989). Investiga-tion of biofilm consortia from groundwater byTransmission Electronic Microscopy (TEM)showed that the accumulation of iron on the cellsurfaces was stepwise and that iron precipitateswere found at the cell surfaces and throughoutthe biofilm matrix (Brown et al. 1998a). Bacterialcell walls and EPS are generally negativelycharged, which allows positively charged metalions to be adsorbed. These metal ions may thanact as nucleation sites for the deposition of moreiron from solution as shown by Brown et al.(1998a). Many investigations report that thesorptive capacities of Fe/Mn oxides for metalsare increased in the presence of microorganisms(Haack & Warren 2003; Nelson et al. 2002). Theiron oxide present in the biofilm can adsorb zinc,lead and cadmium in the biofilm (Baldi et al.
2001). Lack et al. (2002a, b) showed that theanaerobic bio-oxidation of 10 mM Fe(II) by De-chlorosoma species and precipitation of Fe(III)oxides by these microorganisms resulted in rapidadsorption and removal of 55 lM uranium and81 lM cobalt from solution.
Metal reductive precipitation. Bioreduction isgenerally recognized as an important metabolicprocess controlling the fate and transport of hea-vy metals and radionuclides, as well as the overall geochemistry of subsurface and sedimentaryenvironments. Bioreduction of multivalent metalscan convert dissolved, oxidized forms of multi-valent heavy metals and radionuclides (Cr, Mo,Se, U, Pd, Tc, Au), such as U(VI) and Cr(VI), toreduced forms that readily precipitate from solu-tion. The mobility of most of these elements inthe environment is strongly dependent on theirchemical oxidation state. Under oxidizing condi-tions, they occur as highly soluble and mobileions, however, under reducing conditions theyusually form insoluble phases (Abdelouas et al.2000). A recent review gives a detailed under-standing of some of these transformations at amolecular level (Lloyd 2003). For instance,molybdenum (VI) reduction by D. desulfuricansin the presence of sulfide resulted in the extracel-lular precipitation of the mineral molybdeniteMoS2(S) (Tucker et al. 1997). Lloyd et al. (2001)show that SRB reduce Tc(VII) and this metalprecipitates as TcO2 at the periphery of the cell.Microbial reduction of toxic Se(VI) and Se(IV)to the colloidal elemental selenium [Se(0)] can beused to remove Se from wastewater (Lloyd et al.2001; Hockin & Gadd 2003). Elemental seleniumhas not been localized on the bacterial cells(Lloyd et al. 2001) but was released in the bulksolution. SRB biofilms have been shown to re-duce Cr(VI) to Cr(III), which precipitates asinsoluble hydroxides (Cr(III)) (Smith & Gadd2000).
5.2.4. Microbial precipitationSulfide precipitation. Sulfate-reducing bacteria(SRB) are the important physiological group forsulfide production. SRB are commonly used forthe bioremediation of metal-contaminated soilor water. White et al. (1998), for instance,described an integrated microbial process for the
145

bioremediation of soil contaminated with toxicmetals using microbially catalyzed reactions. Inthis process, bioleaching of Cd, Co, Cr, Cu,Mn, Ni, and Zn via sulfuric acid produced bysulfur-oxidizing bacteria was followed by precip-itation of the leached metals as insoluble sulfidesby the action of SRB. Microorganisms havebeen used in the remediation of metal pollutionin very specific cases. The most successful hasbeen the reduction and the eventual precipita-tion of soluble metal sulphates as insoluble sul-fides in liquid wastes by the use of themicroorganisms that are part of the sulfur cycle,especially sulfate-reducing bacteria that use sul-fate as their ultimate electron acceptor to pro-duce hydrogen sulfide, which binds with themetals to give metal sulfide. This procedure isused nowadays not only for the treatment ofsurface waters but also for massive cleaning upof underground water and is available in manyformats, even as commercial large-scale treat-ment plants. Bioprecipitation of metals bymicrobiologically produced sulfide (SRB) is thenan effective way to immobilise a wide range ofmetals, including Zn, Cu, Pb, Ni and Cd. Themetals precipitate as highly insoluble sulfides(entrapment, nucleation and crystallisation ofinsoluble sulfides), e.g. ZnS, CdS, CuS, CoS,NiS and FeS (White & Gadd 1998; White &Gadd 2000; Labrenz et al. 2000; Wang et al.2001; Drzyzga et al. 2002; Valls & de Lorenzo,2002; Utgikar et al. 2002; White et al. 2003;Krumholz et al. 2003). Sulfide precipitation is anefficient means of removing toxic metals fromsolution, while immobilisation in a biofilm isalso an affective method to reduce the mobilityof the metals, which is of value in bioremedia-tion of anoxic sediment and wetlands (Whiteet al. 1998; Kaksonen et al. 2003; Labrenz &Banfield 2004). Sharma et al. (2000) studied ahighly cadmium resistant Klebsiella planticolastrain that is able to induce the precipitation ofcadmium sulfide. This strain is a good candidatefor the accelerated bioremediation of systemscontaminated by high levels of cadmium. Eden-born (2004) showed that the addition of bothpolylactic acid (PLA) and gypsum in con-structed wetlands and reactive barrier walls isrequired for stimulating indigenous bacteria tolower redox potential, raise pH, and enhance thebacterial sulfate reduction activity.
Phosphate precipitation. Microorganisms can pro-duce phosphate that precipitates metals as phos-phates and it is effective for a range of metals. Amixture of accumulative and chemisorptive mech-anisms contribute to the overall process. Biologi-cally produced metal precipitates as phosphates iseffective for a range of metals and radionuclides(Renninger et al. 2001; Basnakova et al. 1998, SeeMacaskie et al. 2004 for a detailed review). A Cit-robacter sp. is able to accumulate the uranyl ion(UO2
2+) via precipitation with phosphate liber-ated from organic phosphate substrates by phos-phatase activity. Accumulation of uranium asHUO2PO4 by Citrobacter sp. was observed in thepresence of excess inorganic phosphorus (Rennin-ger et al. 2001). Cell-bound HUO2PO4 facilitatedalso Ni2+ removal by intercalative ion exchangeinto the polycrystalline lattice and promoted alsozirconium removal (Basnakova et al. 1998).
Carbonate precipitation. Bacterial calcification isanother example for the precipitation of inor-ganic material by biofilms. The formation of cal-cium carbonates particles in microbial mats andother aggregates was suggested to be, at least inpart, because of heterotrophic bacteria (Hammes& Verstraete 2002). Recently, interest in bacteri-ally associated calcium carbonate precipitationhas grown due to strategic bioremediation ap-proaches involving solid phase capture of radio-nuclide and trace element contaminants bycarbonates in groundwater systems (Curti 1999;Warren et al. 2001; Fujita et al. 2004). Fe(III)oxides are commonly thought of as the mostimportant sorbents responsible for sequestrationof many metals and radionuclides in soils andgroundwater systems (Warren & Haack 2001).However, in subsurface anoxic aquifers, Fe(III)(hydr)oxides can be dissolved through microbialreactions involving reduction of Fe (III) to Fe(II). Such dissolution is accompanied by releaseof adsorbed or coprecipitated metals or radio-nuclides. Hence, if bacteria can promote precipi-tation of other geochemically reactive, nonredox-sensitive solid phases such as carbonates,then the bacteria may provide an important bio-remediation tool for solid phase sequestration ofradionuclide or trace element contaminants inanoxic environments. Microbial degradation ofurea was investigated as a potential geochemicalcatalyst for Ca carbonate precipitation and asso-
146

ciated solid phase capture of common groundwa-ter contaminants (Sr, UO2, Cu) in laboratorybatch experiments (Warren et al. 2001). Bacterialdegradation of urea increased pH and promotedCa carbonate precipitation in both bacterial con-trol and contaminant treatments. Associated so-lid phase capture of Sr was highly effective,capturing 95% of the 1 mM Sr added within24 h (Warren et al. 2001). Because bacterial ure-olysis can generate high rates of calcite precipita-tion, the application of this approach ispromising for remediation of Sr contamination inenvironments where calcite is stable over the longterm (Fujita et al. 2004).
5.3. In-situ utilization of biofilms forbioremediation
As a process for immobilizing metals, precipita-tion of metals as sulfides has already achievedlarge-scale application. Although much research iscurrently taking place in metal bioadsorption andthe microbial reductive precipitation of metal andradionuclides contaminants, the fact is that thesetechniques are not yet ready for immediate use inextensive cleaning up operations. Table 3 givessome examples of bacterial strains isolated fromthe environment that can be used for the bioreme-diation of contaminated subsurface environmentthrough microbial reductive precipitation process.However, understanding the bacterially mediatedprecipitation processes at a mechanistic level isstill a requisite for properly applying these tech-nologies to complex systems.
Most bacteria (>90%) in subsurface or sedi-mentary environments are attached to particulatephases. Knowledge of the arrangement of sur-face-associated microorganisms is necessary inorder to more fully understand the factors (e.g.,electron donor and acceptor availability, pH, en-zyme kinetics, diffusional limitations) that con-trol the biogeochemistry of redox-sensitiveminerals. This knowledge will allow to determinethe role of bacteria in influencing or controllingthe fate and transport of heavy metals and radio-nuclides in contaminated environments. The cur-rent understanding of sorption and accumulationof heavy metals and radionuclides by bacteria at-tached to mineral surfaces compromises our abil-ity to predict microbiological immobilization of
contaminants in subsurface and sedimentaryenvironments.
6. Summary
The bioremediation of metal-contaminated soilsis a multi-faceted enterprise that applies a widevariety of biological activities, from single mole-cules to entire macroorganisms, to the substantialproblem of detoxifying hazardous land. Severaltechniques attempt to remove the tightly bound,slowly desorbing metals; complexation and facili-tated transport by bio-surfactants, cyclodextrinsand exopolysaccharides; acid leaching by thechemolithotrophic activity of Thiobacillus; reduc-tions of arsenic and plutonium to more solublespecies and reduction of mercury to volatile Hg0,methylation of selenium to volatile dimethylsele-nide; and extraction of cadmium, zinc, lead, sele-nium and other metals into living plant tissue.The reductions of selenium and chromium, incontrast, have as their goal the detoxification andimmobilization of the metals in-situ, with theunderstanding that continuous careful manage-ment will be required to maintain reducing con-ditions in such environments.
An important question in each of these meth-ods is the source of the bioremedial organisms.Some of the more unusual activities, such as plu-tonium reduction, are known to be catalyzed byonly a few microorganisms and it may be neces-sary to introduce specific microbes for in-situtreatment. Distributing bacterial cells throughsoils is itself a significant undertaking, althoughprogress is being made in this area (Bai et al.1997) and the survival and activity of non-nativemicrobes in a new community is never assured.In several cases, including chromate reduction,selenate reduction, selenium volatilization andmercury volatilization, the desired activities areconveniently ubiquitous, indigenous to contami-nated areas and easily stimulated by hydrogen,water, warmth, and various forms of organicmatter. In-situ treatments requiring whole-cellactivities thus appear most straightforward whennative microorganisms can be recruited. Inphytoremediation, in contrast, native plants donot often appear to be the most effective; becauseof the comparative ease of plant cultivation,
147

however, maintaining the desired organismsposes little difficulty.
All attempts at restoration of contaminatedland face the similar, internally conflicting goalsof speed, cost efficiency, and generation of aclean, productive patch of earth. The earliest‘‘brute-force’’ methods of solidification/stabiliza-
tion, excavation and landfilling, and cappingwere markedly constrained by the first two goals,at the expense of the third. Developing chemicaltechnology increased the efficiency of soil wash-ing and pump-and-treat methods, greatlyimproving the fate of the treated land but stillrequiring the addition of further hazardous
Table 3. Studies on metal–microbe interaction in soils and sediments that have impact on bioremediation of metal contamination
Contaminant Comments References
Uranium Mine
sediment
Uranium Desulfosporosinus sp. Reduced U(VI) to U(IV). This organism may
be stimulated with addition of sulfate and nutrients uranium.
Suzuki et al. 2003
Soil and
sediment
Bio-oxidation
of Fe(II)
Anaerobic Bio-oxidation of 10 mM Fe(II) and precipitation of
Fe(III) oxides by Dechlorosoma sp. Resulted in rapid adsorption
and removal of 55 lM uranium and 81 lM cobalt. The results of
this study demonstrate the potential of this novel approach for
stabilization and immobilization of heavy metals and radionuclides
in the environment
Lack et al. 2002a, b
Sediment Selenium
reduction
A selenite respiring bacterium, Bacillus selenitrieducens, produced
significant levels of Se()II) and is also able to reduce selenite
through Se(0) to Se ()II). The microorganism showed a great
potential to immobilize selenium in anaerobic environments
Herbel et al. 2003
Sediment Chromate
reduction
Consortium of marine sulfate-reducing bacteria was shown to
reduce chromate (CrO2�4 ) into Cr3+. These indigenous SRB might
have potential application in bioremediation of metal contami-
nated sediments.
Cheung and Gu 2003
Subsurface
environment
Uranium
immobilization
Immobilisation of hexavalent uranium [U(VI)] by biofilm of the
sulfate-reducing bacterium Desulfovibrio desulfuricans G20. The
hexavalent uranium was immobilized following enzymatic or
chemical reduction reaction. The chemical reduction of uranium
was due to the presence of dissolved sulfide species.
Beyenal et al. 2004
Soils and
sediments
Cadmium Highly cadmium resistant Klebsiella planticola strain that is able to
induce the precipitation of cadmium sulfide. This strain is a good
candidate for the accelerated bioremediation of systems contami-
nated by high levels of cadmium
Sharma et al. 2000
Subsurface
sediment
Uranium Stimulation of dissimilatory metal reduction activity of an enrich-
ment of microorganisms closely related to Pseudomonas and Des-
ulfosporisinus sp. which promote the reductive precipitation of
uranium
Nevin et al. 2003
Selenite polluted
site
Selenium Ralstonia metallidurans CH34 can reduce selenite to elemental red
selenium, that is highly insoluble
Roux et al. 2001
Subsurface
sediment
Cobalt Immobilization of cobalt by sulfide precipitates Krumholz et al. 2003
Soil Mercury Microbial reduction of soluble ionic mercury, Hg(II), to volatile
Hg(0). The reduced Hg(0) can then flux out of the environment and
be diluted in the atmosphere
Hobman et al. 2000
Subsurface
environments
Uranium Biofilm of the sulfate reducing bacterium Desulfovibrio desulfuri-
cans attached to hematite (a-Fe2O3) surface can accumulates both
U(VI) and U(IV) and thus can limit the transport of uranium in
subsurface environments
Neal et al. 2004
148

substances. Research in bioremediation is nowopening the virtually infinite possibilities inherentin enzymatic systems. Because biological tech-niques are being developed to cost as little or lessthan the best available conventional methods, tooperate in a timely manner, employ biodegrad-able and (with the exception of metal-laden phy-toremedial shoots) non-toxic substances, and toyield functional soils and sediments, bioremedia-tion may soon provide the tools to restore metal-contaminated soils rapidly, inexpensively andpermanently.
Driven by the realization that large areas ofland contaminated with metals and radionuclidescannot be economically remediated using conven-tional chemical approaches, significant resourceshave become available for the area of bioremedia-tion of metal contamination. Supported bygenomics-enabled studies ongoing in many labora-tories worldwide (Lovley 2003) we can expect thisresearch area to develop further, delivering morerobust techniques for the bioremediation of metalcontaminated waters, soils and sediments. Excit-ing developments in the use of microorganisms forthe recycling of metal wastes, with the formationof novel biominerals with unique properties arealso predicted for the near future.
References
Abdelouas A, Lu YL, Lutze W & Nuttall HE (1998a)Reduction of U(VI) to U(IV) by indigenous bacteria incontaminated ground water. J. Contam. Hydrol. 35: 217–233
Abdelouas A, Lutze W & Nuttall EH (1998b) Chemicalreactions of uranium in groundwater at a mill tailings site.J. Contam. Hydrol. 34: 343–361
Abdelouas A, Lutze W & Nuttall HE (1999) Uraniumcontamination in the subsurface: characterization and reme-diation In: Burns PC & Finch R (Eds.) Uranium: Mineral-ogy, Geochemistry and the Environment (pp 433–473). vol.38 Mineralogical Society of America, Washington DC, USA
Abdelouas A, Lutze W, Gong W, Nuttall EH, Strietelmeier BA& Travis BJ (2000) Biological reduction of uranium ingroundwater and subsurface soil. Sci. Total Environ. 250:21–35
Ahmann D, Roberts AL, Krumholz HF & Morel FMM (1994)Microbe grows by reducing arsenic. Nature 351: 750
Ahmann D, Krumholz LR, Hemond HF, Lovley DR & MorelFMM 1997. Microbial Mobilization of arsenic from sedi-ments of the Aberjona Watershed. Environ. Sci. Technol. 31:2923–2930.
Anderson RT & Lovley DR (2002) Microbial redox interac-tions with uranium: an environmental perspective. In: Livens
FR & Keith-Roach M (Eds) Microbiology and Radioactivity(pp. 205–223). Elsevier
Anderson RT, Rooney-Varga J, Gaw CV & Lovley DR (1998)Anaerobic benzene oxidation in the Fe(III)-reduction zone ofpetroleum-contamianted aquifers. Environ. Sci. Technol.32: 1222–1229
Anderson RT, Vrionis HA, Ortiz-Bernad I, Resch CT, LongPE, Dayvault R, Karp K, Marutzky S, Metzler DR, PeacockA, White DC, Lowe M & Lovley DR (2003) Stimulating thein situ activity of Geobacter species to remove uranium fromthe groundwater of a uranium-contaminated aquifer. Appl.Environ. Microbiol. 69: 5884–5891
Andres Y, MacCordick HJ & Hubert JC (1993) Adsorption ofseveral actinide (Th, U) and lanthanide (La, Eu, Yb) ions byMycobacterium smegmatis. Appl. Microbiol. Biotechnol.39: 413–417
Arp G, Reimer A & Reitner J (2001) Photosynthesis-inducedbiofilm calcification and calcium concentrations in Phanero-zoic oceans. Science 292: 1701–1704
Bai G, Brusseau ML & Miller RM (1997) Influence of arhamnolipid biosurfactant on the transport of bacteriathrough a sandy soil. Appl. Environ. Microbiol. 63: 1866–1873
Baldi F, Minacci A, Pepi M & Scozzafava A (2001) Gelsequestration of heavy metals by Klebsiella oxytoca isolatedfrom iron mat. FEMS Microbiol. Ecol. 36: 169–174
Barkay T & Schaefer J (2001) Metal and radionuclidebioremediation: issues, considerations and potentials. Curr.Opin. Microbiol. 4: 318–323
Barnes LJ, Janssen FJ, Sherren J, Versteegh JH, Koch RO &Scheeren PJH (1991) A new process for the microbialremoval of sulphate and heavy metal from contaminatedwaters extracted by a geohydrological control system. Chem.Eng. Res. Des. 69: 184–186
Bartlett RJ (1991) Chromium cycling in soils and water: links,gaps and methods. Environ. Health Perspect. 92: 17–24
Basnokova G, Spencer AJ, Palsgard E & GrimeGW& Macas-kie LE (1998) Identification of the nickel uranyl phosphatedeposits on Citrobacter cells by electron microscopy withelectron probe X ray microanalysis and by proton induced Xray emission analysis. Environ. Sci. Technol. 32: 760–765
Bender J & Phillips P (2004) Microbial mats for multipleapplications in aquaculture and bioremediation. Biores.Technol. 94: 229–238
Bender J., Lee RF& Phillips P (1995) A review of the uptake andtransformation of metals metalloids by microbial mats andtheir use in bioremediation. J. Ind. Microbial. 14: 114–118
Bender J, Lee RF, Sheppard M, Bulski K, Phillips P, Yeboah Y& Chee Wah R (2004) A waste effluent treatment systembased on microbial mats for black sea bass Centrapristisstrata recycled-water aquaculture. Aqua. Eng. 31: 73–82
Berne C (2004) Etude des Mecanismes de degradation duphosphate de tributyle par des bacteries. PhD. Universite dela Mediterranee, Aix-Marseille France
Beveridge TJ, Makin SA, Kadurugamuwa JL & Li Z (1997a)Interactions between biofilms and the environment. FEMSMicrobiol. Rev. 20: 291–303
Beveridge TJ, Hughes MN, Lee H, Leung KT, Poole RK,Savvaidis I, Silver S & Trevors JT (1997b) Metal–microbeinteractions: contemporary approaches. Adv. Microbial.Physiol 38: 177–243
BeyenalH, SaniRK, PeytonBM,DohnalkovaAC,Amonette JE& Lewandowski Z (2004) Uranium immobilization by sulfate-reducing biofilms. Environ. Sci. Technol. 38: 2067–2074
149

Beynal H & Lewandowski Z (2004) Dynamics of lead immobi-lization in sulfate reducing biofilms. Wat. Res. 38: 2726–2736
Blake RC, Shute EA & Howard GT (1994) Solubilization ofminerals by bacteria: electrophoretic mobility of Thiobacillusferrooxidans in the presence of iron. Microbiol. 60:3349–3357
Bodek ILyman WJReehl WFRosenblatt DH. (1988) Environ-mental Inorganic Chemistry: Properties, Processes and Esti-mation Methods. Pergamon Press, Inc, Elmsford, NY
Bolton H, Xun L & Girvin DC (2000) Biodegradation ofsynthetic chelating agents In: Lovley D (Eds.) EnvironmentalMicrobe-Metal Interactions (pp 363–383). ASM Press,Washington, D.C.
Bonthrone KM, Basnakova G, Lin F & Macaskie LE (1996)Bioaccumulation of nickel byintercalation into polycrystal-line hydrogen uranyl phosphate deposited via an enzymaticmechanism. Nat. Biotechnol. 14: 635–638
Bontidean I, Csoregi E, Corbisier P, Lloyd JR & Brown NL(2002) Bacterial metal- responsive elements and their use inbiosensors for monitoring of heavy metals In: Sankar B(Eds.) The Handbook of Heavy Metals in the Environment(pp 647–680). Marcel Dekker Inc, New York
Bontidean I, Lloyd JR, Hobman JL, Wilson JR, Csoregi E,Mattiasson B & Brown NL (2000) Bacterial metal-resistanceproteins and their use in biosensors for the detection ofbioavailable heavy metals. J. Inorg. Biochem. 79: 225–229
Bopp L & Ehrlich H (1988) Chromate resistance and reductionin Pseudomonas fluorescens strain LB300. Arch. Microbiol.150: 426–431
Borst-Pauwels GWFH (1981) Ion transport in yeast. Biochim.Biophys. Acta 650: 88–127
Boseker K (1997) Bioleaching: metal solubilization by micro-organisms. FEMS Microbiol. Rev. 20: 591–604
Boswell CD, Dick RE, Eccles H & Macaskie LE (2001)Phosphate uptake and release by Acinetobacter johnsonii incontinuous culture and coupling of phosphate release toheavy metal accumulation. J. Ind. Microbiol. Biotechnol. 26:333–340
Boyle V (1993) Soils washing In: Pratt M (Eds.) RemedialProcesses for Contaminated Land (pp 33–53). Institution ofChemical Engineers, Warwicksjhire
Brierley CL (1982) Microbiological Mining. Sci. Am. 247:44–53
Brierley CL (1990) Biomediation of metal-contaminated surfaceand ground waters. Geomicrobiology 8: 201–223
Brown DA, Beveridge TJ, Keevil CW & Sherriff BL (1998a)Evaluation of microscopic techniques to observe iron pre-cipitation in a natural microbial biofilm. FEMS Microbiol.Ecol. 26: 297–310
Brown DA, Sawicki JA & Scheriff BL (1998b) Alteration ofmicrobially precipitated iron oxides and hydroxides. Am.Mineral. 83: 1419–1425
Brown DA, Scheriff BL, Sawicki JA & Sparling R (1999)Precipitation of iron minerals by a natural microbialconsortium. Geochim. Cosmochim. Acta 63: 2163–2169
Bruins MR, Kapil S & Oehme FW (2000) Microbial resistanceto metals in the environment. Ecotoxicol. Environ. Saf. 45:198–207
Brusseau ML, Wang X & Hu Q (1994) Enhanced transport oflow-polarity organic compounds through soil by cyclodex-trin. Environ. Sci. Technol. 31: 952–956
Brusseau ML, Wang X & Wang WZ (1997) Simultaneouselution of heavy metals and organic compounds from soil bycyclodextrin. Environ. Sci. Technol. 31: 1087–1092
Caccavo F Jr, Lonergan DJ, Lovley DR, Davis M, Stolz JF &McInerney MJ (1994) Geobacter sulfurreducens sp. nov., ahydrogen and acetate-oxidizing dissimilatory metal reducingmicroorganism. Appl. Environ. Microbiol. 60: 3752–3759
Carpentier W, Sandra K, De Smet I, Brige A, De Smet L & VanBeeumen J (2003) Microbial reduction and precipitation ofvanadium by Shewanella oneidensis. Appl. Environ. Micro-biol. 69: 3636–3639
Cervantes C (1991) Bacterial interactions with chromate.Antonie van Leeuwenhoek 59: 229–233
Chaney RL (1997) Phytoremediation of soil metals. Cur. Opin.Biotechnol. 8: 370–384
Chen JM & Hao OJ (1998) Microbial chromium(VI) reduction.Crit. Rev. Environ. Sci. Technol. 28: 219–251
Chen SY & Lin JG (2000) Influence of solid content onbioleaching of heavy metals from contaminated sediment byThiobacillus spp. J. Chem. Technol. Biotechnol. 75: 649–656
Chen SY & Lin JG (2001a) Bioleaching of heavy metals fromsediment: significance of pH. Chemosphere 443: 1093–1102
Chen SY & Lin JG (2001b) Effect of substrate concentration onbioleaching of metal- contaminated sediment. J. Hazard.Mater. 82: 77–89
Chen SY, Lin J-G & Lee CY (2003) Effects of ferric ion onbioleaching of heavy metals from contaminated sediment.Wat. Sci. Technol. 48: 151–158
Chen S-Y & Lin J-G (2004) Bioleaching of heavy metals romcontaminated sediment by indigenous sulfur-oxidizing bac-teria in an air-lift bioreactor: effects of sulfur concentration.Wat. Res. 38: 3205–3214
Chen JH, Lion LW, Ghiorse WC & Shuler ML (1995)Mobilization of adsorbed cadmium and lead in aquifermaterial by bacterial extracellular polymers. Wat. Res. 29:421–430
Cheng CN & Focht D (1979) Production of arsine andmethylarsines in soil and in culture. Appl. Environ. Micro-biol. 38: 494–498
Cheung KH & Gu JD (2003) Reduction of chromate (CrO42))
by an enrichment consortium and an isolate of marinesulfate-reducing bacteria. Chemosphere 52: 1523–1529
Cifuentes FR, Lindemann WC & Barton LL (1996) Chromiumsorption and reduction in soil with implications to bioreme-diation. Soil Sci. 161: 233–24
Combs GF Jr, Garbisu C, Lee BC, Yee A, Carlson DE, SmithNR, Magyarosy AC, Leighton T & Buchanan BB (1996)Bioavailability of selenium accumulated by selenite-reducingbacteria. Biol. Trace Elem. Res. 52: 209–225
Cooke T & Bruland K (1987) Aquatic chemistry of selenium:evidence of biomethylation. Environ. Sci. Technol. 21:1214–1219
Couillard D & Mercier G (1991) Optimum residence time (inCSTR and airlift reactors) for bacterial leaching of metalsfrom anaerobic sewage sludge. Wat. Res. 25: 211–218
Couillard D & Mercier G (1993) Removal of metals and fate ofN and P in the bacterial leaching of aerobically digestedsewage sludge. Wat. Res. 27: 1227–1235
Cox JS, Scott Smith D, Warren LA & Ferris FG (1999)Characterizing heterogeneous bacterial surface functionalgroups using discrete affinity spectra for proton binding.Environ. Sci. Technol. 33: 4514–4521
Cummings DE, Snoeyenbos-West OL, Newby DT, NiggemyerAM, Lovley DR, Achenbach LA & Rosenzweig RF (2003)Diversity of Geobacteraceae species inhabiting metal-pol-luted freshwater lake sediments ascertained by 16S rDNAanalyses. Microb. Ecol. 46: 257–269
150

Cunningham SD, Berti WR & Huang JW (1995) Phytoremedi-ation of contaminated soils. Trends Biotechnol. 13: 393–397
Curti E (1999) Coprecipitation of radionuclides with calcite:estimation of partition coefficients based on a review oflaboratory investigations and geochemical data. Appl. Geo-chem. 14: 433–445
Daly MJ (2000) Engineering radiation-resistant bacteria forenvironmental biotechnology. Curr. Opin. Biotechnol.11: 280–285
Davison W, Fones GR & Grime GW (1997) Dissolved metalsin surface sediment and a microbial mat at 100lm resolution.Nature 387: 885–888
DeMoll-Decker HM & Macy JM (1993) The periplasmic nitritereductase of Thanera selenatis may catalyze the reduction ofselenite to elemental selenium. Arch. Microbiol. 160: 241–247
deRome L & Gadd GM (1991) Use of pelleted and immobilizedyeast and fungal biomass for heavy metal and radionucliderecovery. J. Ind. Microbiol 7: 97–104
Diels L, Dong Q, van der Lelie D, Baeyens W & Mergeay M(1995) The czc operon of Alcaligenes eutrophus CH34: fromresistance mechanism to the removal of heavy metals. J. Ind.Microbiol. 14: 142–153
Diels L, Spaans H, Van Roy S, Hooyberghs L, Ryngaert A,Wouters H, Walter E, Winters J, Macaskie L, Finlay J,Pernfuss B,Woebking H, Pumpel T & TsezosM (2003) Heavymetals removal by sand filters inoculated with metal sorbingand precipitating bacteria. Hydrometallurgy 71: 235–241
Douglas S & Beveridge TJ (1998) Mineral formation bybacteria in natural microbial communities. FEMS Microbiol.Ecol. 26: 79–88
Drzyzga O, El Mamouni R, Agathos SN & Gottschal J (2002)Dehalogenation of chlorinated ethenes and immobilization ofnickel in anaerobic sediment columns under sulfidogenicconditions. Environ. Sci. Technol. 36: 2630–2635
Eccles H, Garnham GW, Lowe CR & Bruce NC (1997)Biosensors for detecting metal ions capable of being reducedby reductase enzymes. Biotechnol. Adv. 15: 189–189
Edenborn HM (2004) Use of poly(lactic acid) amendments topromote the bacterial fixation of metals in zinc smeltertailings. Biores. Technol. 92: 111–119
Ellwood DC, Hill MJ & Watson JHP (1992) Pollution controlusing microorganisms and metal separation In: Fry JC, GaddGM, Herbert RA, Jones CW & Watson-Craik A (Eds.)Microbial Control of Polution (pp 89–112). Vol. 48Cam-bridge University Press, Cambridge, U.K
Fendorf S, Wielinga BW & Hansel CM (2000) Chromiumtransformations in natural environments: the role of biolog-ical and abiological processes in chromium(VI) reduction.Intl. Geol. Rev. 42: 691–701
Ferris FG, Schultze , Witten T, Fyfe WS & Beveridge TJ (1989)Metal interactions with microbial biofilms in acidic andneutral pH environments. Appl. Environ. Microbiol. 55:1249–1257
Finneran KT, Anderson RT, Nevin KP & Lovley DR (2002)Potential for bioremediation of uranium-contaminated aqui-fers with microbial U(VI) reduction. Soil Sed. Comtam.11: 339–357
Flemming H-C, Leis A (2002) Sorption properties of biofilms.In: Flemming H-C (Ed) Biofilms. In: Bitton G. (Ed)Encyclopedia of Environmental Microbiology, vol. 5, (pp.2958–2967)
Forstner U (2004) Sediment dynamics and pollutant mobility inrivers: An interdisciplinary approach. Lakes Reservoirs Res.Manag. 9: 25–40
Fortin D (2004) Geochemistry. What biogenic minerals tell us.Science 303: 618–1619
Francis AJ (1994) Microbial transformations of radioactivewastes and environmental restoration through bioremedia-tion. J. Alloys Comp 213/214: 226–231
Francis AJ & Dodge CJ (1998) Remediation of soils and wastescontaminated with uranium and toxic metals. Environ. Sci.Technol. 32: 3993–3998
Frankenberger WT Jr. & Karlson U (1994a) Microbialvolatilization of selenium from soils and sediments In:Frankenberger WT Jr. & Benson S (Eds.) Selenium in theEnvironment (pp 369–387). Marcel Dekler, Inc., New York
Frankenberger WT Jr. & Karlson U (1994b) Soil managementfactors affecting volatilization of selenium from dewateredsediments. Geomicrobiol. J. 12: 265–278
Frankenberger WT Jr & Karlson U (1995) Volitilization ofselenium from a dewatered seleniferous sediment: a fieldstudy. J. Indust. Microbiol. 14: 226–232
Fredrickson JK, Zachara JM, Kennedy DW, Lin C, Duff MC,Hunter DB & Dohnalkova A (2002) Influence of Mn oxideson the reduction of uranium (VI) by the metal-reducingbacterium Shewanella putrefaciens. Geochem. Vosmochim.Acta 66: 3247–3262
Fredrickson JK, et al. (2000) Reduction of Fe(III), Cr(VI),U(VI), and Tc(VII) by Deinococcus radiodurans. Appl.Environ. Microbiol. 66: 2006–2011
Fujita Y, Redden GD, Ingram JC, Cortez MM, Ferris FG &Smith RW (2004) Strontium incorporation into calcitegenerated by bacterial ureolysis. Geochim. Cosmochim. Acta68: 3261–3270
Gadd GM (2004) Microbial influence on metal mobility andapplication for bioremediation. Geoderma 122: 109–119
Gadd GM (2000) Microbial interactions with tributyltincompounds; detoxification, accumulation, and environmen-tal fate. Sci. Total. Environ. 258: 119–127
Gadd GM & White C (1989) Heavy metal and radionuclideaccumulation and toxicity in fungi and yeasts In: Poole RK& Gadd GM (Eds.) In Metal–Microbe Interactions. Societyfor General Microbiology (pp 19–38). IRL Press, Oxford,England
Geesey GG, Lang L, Jolley JG, Hankins MR, Iwaoka T &Griffiths PR (1989) Binding of metal ion by extracellularpolymers of biofilm bacteria. Water Sci. Technol. 20: 161–165
Geiselhart L, Osgood M & Holmes DS (1991) Ann. NY Acad.Sci. 646: 53–60
Glazer BT, Cary SC, Hohmann L & Luther GW III (2002)In situ sulfur speciation using Au/Hg microelectrode as anaid to microbial characterisation of an intertidal saltmarsh microbial mat In: Taillefert M & Rozan T (Eds.)Environmental Electrochemistry: Analyses of Trace ElementBiogeochemistry (pp 283–305). American Chemical Soci-ety Symposium Series; American Chemical Society, Wash-ington D.C
Gorby YA & Lovley DR (1992) Enzymatic uranium precipi-tation. Environ. Sci. Technol. 26: 205–207
Gorby YACF and Bolton H (1998) Microbial reduction ofcobalt(III)EDTA in the presence and absence of manga-nese(IV) oxide. Environ. Sci. Technol. 32: 244–250
Greiner R, Konierny & Jany KD (1993) Purification andcharacterization of two phytases from Eschericha Coli. Arch.Biochem. Biophys. 303: 107–113
Haack EA & Warren LA (2003) Biofilm hydrous manganeseoxyhydroxides and metal dynamics in acid rock drainage.Environ. Sci. Technol. 37: 4138–4147
151

Hammes F & Verstraete W (2002) Key roles of pH and calciummetabolism in microbial carbonate precipitation. Re/ViewsEnviron. Sci. Bio/Technol. 1: 3–7
Herbel MJ, Switzer Blum J, Borglin SE & Oremland RS (2003)Reduction of elemental selenium to selenide: experimentswith anoxic sediments and bacteria that respire Se- oxya-nions. Geomicrobiol. J. 20: 587–602
Herman DC, Artiola JF & Miller RM (1995) Removal ofcadmium, lead and zinc from soil by a rhamnolipid biosurf-actant. Environ. Sci. Technol. 29: 2280–2285
Higham DP, Sadler PJ & Scawen MD (1984) Cadmium-resistant Pseudomonas putida synthesizes novel cadmiumbinding proteins. Science 225: 1043–1046
Hinck ML, Ferguson J & Puhakka J (1997) Resistance ofEDTA and DTPA to aerobic biodegraduation. Water Sci.Technol. 35: 25–31
Hobman JL, Wilson JR & Brown NL (2000) Microbialmercury reduction In: Lovley DR (Eds.) EnvironmentalMicrobe–Metal Interactions (pp 177–197). ASM Press,Washington, D.C
Hockin SL & Gadd GM (2003) Linked redox precipitation ofsulfur and selenium under anaerobic conditions by sulfate-reducing bacterial biofilms. Appl. Environ. Microbiol. 69:7063–7072
Holmes DE, Finneran KT & Lovley DR (2002) Enrichment ofGeobacteraceae associated with stimulation of dissimilatorymetal reduction in uranium-contaminated aquifer sediments.Appl. Environ. Microbiol. 68: 2300–2306
Jackson M, West M & Leppard GG (1999) Accumulation ofheavy metals by individually analysed bacterial cells andassociated nonliving material in polluted lake sediments.Environ. Sci. Technol. 33: 3795–3801
Ji G & Silver S (1995) Bacterial resistance mechanisms forheavy metals of environmental Citrobacter concern.J. Indust. Microbiol. 14: 61–75
John SG (2001) Siderophore mediated plutonium accumulationby Microbacterium flavescens (JG-9). Environ. Sci. Technol.35: 2942–2948
Kaksonen AH, Riekkola-Vanhanen ML & Puhakka JA(2003) Optimization of metal sulfide precipitation influidised-bed treatment of acidic wastewater. Water Res.37: 255–266
Kearney T, Eccles H, Graves D & Gonzalez A (1996) Presentedat the Proceedings of the 18th Annual Conference of theNational Low-Level Waste Management Program, May 20–22, Salt Lake City, Utah, USA
Kjaergaard K, Sorenson JK, Schembri MA & Klemm P (2000)Sequestration of zinc oxide by fimbrial designer chelators.Appl. Environ. Microbiol. 66: 10–14
Klaus T, Joeger R, Olsson E & Granqvist CG (1999) Silver-based crystalline nanoparticles, microbially fabricated. Proc.Natl. Acad. Sci. USA 96: 13611–13614
Konhauser KO, Fyfe WS, Ferris FG & Beveridge TJ (1993)Metal sorption and mineral precipitation by bacteria in twoAmazonian river systems: Rio Solimoes and Rio Negro,Brazil. Geology 21: 1103–1106
Konhauser KO, Schultze-Lam S, Ferris FG, Fyfe WS, Longs-taffe FJ & Beveridge TJ (1994) Mineral precipitation byepilithic biofims in the Speed River, Ontario, Canada. Appl.Environ. Microbiol. 60: 549–553
Kostka JE (2002) Growth of iron(III)-reducing bacteria in clayminerals as the sole electron acceptor and comparison ofgrowth yields on a variety of oxidized iron forms. Appl.Environ. Microbiol. 68: 6256–6262
Krishnamurthy S & Wilkens MM (1994) Environmentalchemistry of chromium. Northeast. Geol. 16: 14–17
Krumbein WE (1979) Calcification by bacteria and algae.In: Trudinger PA & Swaine DJ (Eds) BiogeochemicalCycling of Mineral- Forming Elements. Elsevier Science,New York
Krumholz LR, Elias DA & Suflita JM (2003) Immobilization ofcobalt by sulfate-reducing bacteria in subsurface sediments.Geomicrobiol. J. 20: 61–72
Kuhn A & Sigg L (1993) Arsenic cycling in eutrophic LakeGreifen, Switzerland: influence of seasonal redox processes.Limnol. Oceanogr. 38: 1052–1055
Labrenz M & Banfield JF (2004) Sulfate reducing bacteria-dominated biofilms that precipitate ZnS in a subsurfacecircumneutral-pH mine drainage system. Microb. Ecol. 47:205–217
Labrenz M, Druschel GK, Thomsen-Ebert T, Gilbert B, WelchSA, Kemner KM, Logan GA, Summons RE, De Stasio G,Bond PL, Lai B, Kelly SD & Banfield JF (2000) Formationof sphalerite (ZnS) deposits in natural biofilms of sulfate-reducing bacteria. Science 290: 1744–1747
Lack JG, Chaudhuri SK, Kely SD, Kemmer KM, O’ConnorSM & Coates JD (2002) Immobilization of radionuclides andheavy metals through anaerobic bio-oxidation of Fe(II).Appl. Environ. Microbiol. 68: 2704–2710
Lack JG, et al. (2002) Immobilization of radionuclides andheavy metals through anaerobic bio-oxidation of Fe(III) byDechlorosoma suillum. Microbiol. Ecol. 43: 424–431
Langley S & Beveridge TJ (1999) Effect of O-side-chain-lipopolysaccharide chemistry on metal binding. Appl. Envi-ron. Microbiol. 65: 489–498
Lappin-Scott HM, Jass J & Costerton JW (1993) Microbialbiofilm formation and characterization In: Lappin-Scott HM& Costerton JW (Eds.) Microbial Films: Formation andControl (pp 1–73). SAB Press, London, UK
Laverman AM, Blum JS, Schaefer JK, Phillips EJP, Lovley DR& Oremland RS (1995) Growth of strain SES-3 with arsenateand other diverse electron acceptors. Appl. Environ. Micro-biol. 61: 3556–3561
Lawson S & Macy J (1995) Bioremediation of selenite in oilrefinery wastewater. Appl.Microbiol. Biotechnol. 43: 762–765
Liehr SK, Chen HJ & Lin SH (1994) Metals removal by algalbiofilms. Water Sci. Technol. 30: 59–60
Lloyd JR (2003) Microbial reduction of metals and radionuc-lides. FEMS Microbiol. Rev. 27: 411–425
Lloyd JR & Macaskie LE (1996) A novel phosphorImagerbased technique for monitoring the microbial reduction oftechnetium. Appl. Environ. Microbiol. 62: 578–582
Lloyd JR &Macaskie LE (2000) Bioremediation of radioactivemetals In: Lovley DR (Eds.) Environmental Microbe–MetalInteractions (pp 277–327). ASM Press, Washington, D.C
Lloyd JR & Lovley DR (2001) Microbial detoxification ofheavy metals and radionuclides. Curr. Opin. Biotechnol. 12:248–253
Lloyd JR & Macaskie LE 2002. The biochemical basis ofradionuclide-microbe Interaction. In: FR Livens and MKeith-Roach (ed.), Microbiology and Radioactivity. Elsevier
Lloyd JR, Sole VA, Van Praagh CV & Lovley DR (2000)Direct and Fe(II)-mediated reduction of technetium byFe(III)-reducing bacteria. Appl. Environ. Microbiol. 66:3743–3749
Lloyd JR, Mabbett AN, Williams DR & Macaskie LE (2001)Metal reduction by sulfate-reducing bacteria: physiologicaldiversity and metal specificity. Hydrometallurgy 59: 327–337
152

Loser C, Seidel H, Hoffman P & Zehnsdorf A (2001) Reme-diation of heavy metal-contaminated sediments by solid-bedbioleaching. Environ. Geol. 40: 643–650
Losi ME & Frankenberger WT (1997) Bioremediation ofselenium in soil and water. Soil Sci. 162: 692–702
Losi ME, Amrhein C & Frankenberger WT Jr (1994a) Factorsaffecting chemical and biological reduction of hexavalentchromium in soil. Environ. Toxicol. Chem. 13: 1727–1735
Losi ME, Amrhein C & Frankenberger WT Jr. (1994b)Bioremediation of chromate-contaminated groundwater byreduction and precipitation in surface soils. J. Environ. Qual.23: 1141–1150
Lovley DR (1993) Dissimilatory metal reduction. Ann. Rev.Microbiol. 47: 263–290
Lovley D (2000) Environmental Microbe–Metal Interactions.ASM Press, Washington, D.C. 395
Lovley DR (2001a) Reduction of iron and humics insubsurface environments In: Frederickson JK & Fletcher M(Eds.) Subsurface Microbiology and Biogeochemistry (pp193–217). Wiley-Liss, Inc., New York
Lovley DR (2001b) Biomediation – anerobes to the rescue.Science 293: 1444–1446
Lovley D (2002) Dissimilatory metal reduction: from early lifeto biomediation. ASM News 68: 231–237
Lovley DR (2003) Cleaning up with genomics: Applyingmolecular biology to bioremediation. Nat. Rev. Microbiol.1: 36–44
Lovley D & Phillips EJ (1992a) Reduction of uranium byDesulfovibrio desulfuricans. Appl. Environ. Microbiol 58:850–856
Lovley DR & Phillips EJP (1992b) Bioremediation of uraniumcontamination with enzymatic uranium reduction. Environ.Sci. Technol. 26: 2228–2234
Lovley DR & Blunt-Harris EL (1999) Role of humics-boundiron as an electron transfer agent in dissimilatory Fe(III)reduction. Appl. Environ. Microbiol. 65: 4252–4254
Lovley DR & Coates JD (2000) Novel forms of anaerobicrespiration of environmental relevance. Curr. Opin. Micro-biol. 3: 252–256
Lovley DR, Phillips EJP, Gorby YA & Landa E (1991)Microbial reduction of uranium. Nature 350: 413–416
Lovley DR, Roden EE, Phillips EJP & Woodward JC (1993a)Enzymatic iron and uranium reduction by sulfate reducingbacteria. Marine Geol. 113: 41–53
Lovley DR, Widman PK, Woodward JC & Phillips EJP(1993b) Reduction of uranium by cytochrome c3 of Desul-fovbrio vulgaris. Appl. Environ. Microbiol. 59: 3572–3576
Lovley DR & Phillips EJP (1994) Reduction of chromate byDesulfovibrio vulgaris and its cytochrome. Appl. Environ.Microbiol. 60: 726–728
Macaskie LE (1991) The application of biotechnology to thetreatment of wastes produce from nuclear fuel cycle: biodeg-radation and bioaccumulation as a means of treating radio-nuclide-containing streams. Crit. Rev. Biotechnol. 11: 41–112
Macaskie LE, Empson RM, Cheetham AK, Grey CP &Skarnulis AJ (1992) Uranium bioaccumulation by a Citrob-acter sp. as a result of enzymically-mediated growth ofpolycrystalline HUO2PO4. Science 257: 782–784
Macaskie LE, Jeong BC & Tolley MR (1994) Enzymically-accelerated biomineralization of heavy metals: application tothe removal of americium and plutonium from aqueousflows. FEMS Microbiol. Rev. 14: 351–379
Macaskie LE, Lloyd JR, Thomas RAP & Tolley MR (1996)The use of microorganisms for the remediation of solutions
contaminated with actinide elements, other radionuclides,and organic contaminants generated by nuclear fuel cycleactivities.. Nucl. Energy 35: 257–271
Macaskie LE, Yong P & Paterson-Beedle M (2004) Bacterialprecipitation of metal phosphates In: Valsami-jones E (Eds.)Phosphorus in Environmental Technology; Principles andApplications (pp 547–579). IWA publishing, London, UK
Macaskie LE (1991) The application biotechnology to thetreatment of wastes produced from the nuclear fuel cycle:biodegradation and bioaccumulation as a means of treatingradionuclide-containing streams. Crit. Rev. Biotechnol. 11:41–112
Macy J (1994) Biochemistry of selenium metabolism byThauera selenatis gen.nov. sp. Nov and use of the organismfor bioremediation of selenium oxyanion in San JoaquinValley drainage water In: Frankenberger WT Jr. & Benson S(Eds.) Selenium in the Environment (pp 421–444). MarcelDekker, Inc., New York, NY
Macy J, Lawson S & DeMoll-Decker H (1993) Bioremediationof selenium oxyanions in San Joaquin drainage water usingThauera selentis in a biological reactor system. Appl.Microbiol. 40: 588–594
Macy J, Nunan K, Hagen K, Dixon D, Harbour P, Cahill M &Sly L (1996) Chryioenes arsenatis gen. nov., sp. Nov., a newarsenate-respiring bacterium isolated from gold mine waste-water. Int. J. Syst. Bacteriol. 46: 1153–1157
McCullough J, Hazen TC, Benson SM, Metting FB &Palmisano AC (1999) Bioremediation of Metals and Radio-nuclides. What Is It and How it Works. Lawrence BerkeleyNational Laboratory, Berkeley, Cailfornia
McHale AP & McHale S (1994) Microbial biosorption ofmetals: potential in the treatment of metal pollution.Biotechnol. Adv. 12: 647–652
Mercier G, Chartier M & Couillard D (1996) Strategies tomaximize the microbial leaching of lead from metal-contam-inated aquatic sediments. Wat. Res. 30: 2452–2464
Miller RM (1995) Biosurfactant-facilitated remediation ofmetal-contaminated soils. Environ. Health Perspect.103(Suppl. 1), 59–62
Morel FMM & Hering JG (1993) Principles and Applicationsof Aquatic Chemistry. John Wiley & Sons, Inc, New York,NY
Neal AL, Amonette JE, Peyton BM & Geesey GG (2004)Uranium complexes formed at hematite surfaces colonizedby sulfate-reducing bacteria. Environ. Sci. Technol. 38:3019–3027
Nelson YM, Lion LW, Shuler ML & Ghiorse WC (2002) Effectof oxide formation mechanisms on lead adsorption bybiogenic manganese hydroxides, iron hydroxides, and theirmixtures. Environ. Sci. Technol. 36: 421–425
Neu MP (2000) Siderophore-mediated chemistry and microbialuptake of plutonium. Los Alamos Sci. 26: 416–417
Neu MP, Matonic JH, Ruggiero CE, Scott BL & Angew(2000b) The first structural characterization of plutonium(IV)complexed by a siderophore: Single crystal structure of PuDesferrioxamine E. Angewandte Chemie Intl. Edit. 39: 1442–1444
Nevin KP, Finneran KT & Lovley DR (2003) Microorganismsassociated with Uranium bioremediation in a high-salinitysubsurface sediment. Appl. Environ. Microbiol. 69: 3672–3675
Newman DK & Banfield JF (2002) Geomicrobiology: howmolecular-scale interactions underpin biogeochemicalsystems. Science 296: 1071–1077
153

Newman D, Kennedy E, Coates J, Ahmann D, Ellis D, LovleyD & Morel F. (1997) Dissimilatory As(V) and Se(VI)reduction in Desulfotomaculum auripigmentum, sp. Nov.Arch. Microbiol. 168: 380–388
North NN, Dollhopf SL, Petrie L, Istok JD, Balkwill DL &Kostka JE (2004) Change in bacterial community structureduring in situ biostimulation of subsurface sediment cocon-taminated with uranium and nitrate. Appl. Environ. Micro-biol. 70: 4911–4920
Okorov LA, Lichko LP, Kodomtseva VM, Kholodenko VP,Titovsky VT & Kulaev IS (1977) Energy-dependent transportof manganese into yeast cells and distribution of accumulatedions. Eur. J. Biochem. 75: 373–377
Oremland R, Blum JS, Culbertson C, Visscher P, Miller L,Dowdle P & Strohmaier F (1994) Isolation, growth andmetabolism of an obligately anaerobic, selenate-respiringbacterium, strain SES-3. Appl. Environ. Microbiol. 60: 3011–3019
Oremland R, Steinberg N, Maest A, Miller L & Hollibaugh J(1990) Measurement of removal of in situ rates of selenateremoval by dissimilatory bacterial reduction in sediments.Environ. Sci. Technol. 24: 1157–1164
Oremland RS, Hollibaugh JT, Maest AS, Presser TS, Miller LG& Culbertson CW (1989) Selenate reduction to elementalselenium by anaerobic bacteria in sediments and culture:biogeochemical significance of a novel, sulphate-independentrespiration. Appl. Environ. Microbiol. 55: 2333–2343
Ortiz-Bernad I, Anderson RT, Vrionis HA & Lovley DR (2004)Vanadium respiration byGeobacter metallireducens: a novelstrategy for the in situ removal of vanadium from ground-water. Appl. Environ. Microbiol. 70: 3091–3095
Paterson-Beedle M & Macaskie LE (2003) Presented at the IBS2003 Biohydrometallurgy: A sustainablee Technology inEvolution, Athens, Greece, September 14–17th
Pattanapipitpaisal P, Mabbett AN, Finlay JA, Beswick AJ,Paterson-Beedle M, Wright EAJ, Tolley MR, Badar U,Ahmed N, Hobman JL, Brown NL & Macaskie LE (2002)Reduction of Cr(VI) and bioaccumulation of chromium byGram positive and Gram negative microorganisms notpreviously exposed to Cr-stress. Environ. Technol. 23:731–745
Payne RB, et al. (2002) Uranium reduction by Desulfiovbriodesulfuricans strain G20 and a cytochrome c3 mutant. Appl.Environ. Microbiol. 68: 3129–3132
Pazirandeh M, Wells BM & Ryan RL (1998) Development ofbacterium-based heavy metal biosorbants : enhanced uptakeof cadmium and mercury by Escherichia coli expressing ametal-binding motif. Appl. Environ. Microbiol. 64: 4068–4072
Phillips EJP, Landa ER & Lovley DR (1995) Remediation ofuranium contaminated soils withbicarbonate extraction andmicrobial U(VI) reduction. J. Indust. Microbiol. 14: 203–20
Postgate JR (1979) The Sulphate Reducing Bacteria. Cam-bridge University Press, Cambridge, U.K
Pozas-Tormo R, Moreno-Real L, Martinez L & Bruque-Gomez S (1987) Intercalation of lanthanides into H3OUO2-
PO4Æ3H2O and C4H9NH3UO2PO4Æ3H2O. Inorg. Chem. 26:1442–1445
Pulford ID & Watson C (2003) Phytoremediation of heavymetyal-contaminated land by trees: a review. Environ. Int.29: 539–560
Raskin I, Smith RD & Salt DE (1997) Phytoremediation ofmetals: Using plants to remove pollutants from the environ-ment. Curr. Opin. Biotechnol. 8: 221–226
Reeves RD & Baker AJM (2001) Metal-accumulating plantsIn: Raskin I & Ensley BD (Eds.) Phytoremediation of ToxicMetals: Using Plants to Clean-up the Environment (pp 193–221). John Wiley and Sons, New York
Remoudaki E, Hatzikioseyian A, Kousi P & Tsezos M (2003)The mechanism of metals precipitation by biologicallygenerated alkalinity in biofilm reactors. Water Res. 37:3843–3854
Renninger N, McMahon KD, Knopp R, Nitsche H, Clark DS& Keasling JD (2001) Uranyl precipitation by biomass froman enhanced biological phosphorus removal reactor. Biodeg-radation 12: 401–410
Rodriguez H & Fraga R (1999) Phosphate solubilizing bacteriaand their role in plant growth promotion. Biotechnol. Adv.17: 319–339
Roig MG, Kennedy JF & Macaskie LE (1995) BiologicalRehabilitation of Metal Bearing Wastewaters. Finalreport EC contract EV5V-CT93–0251. Programme: Envi-ronment
Roling WFM, van Breukelen BM, Braster M, Lin B & vanVerseveld HW (2001) Relationships between microbial com-munity structure and hydrochemistry in a landfill leachate-polluted aquifer. Appl. Environ. Microbiol. 67:4619–4629
Roux M, Sarret G, Pignot-Paintrand I, Fontecave M & Coves J(2001) Mobilization of selinite by Ralstonia metalliduransCH34. Appl. Environ. Microbiol. 67: 769–773
Rugh CL, Wilde HD, Stack NM, Thompson DM, SummersAO & Meagher RB (1996) Mercuric ion reduction andresistance in transgenic Arabidopsis thaliana plants expressinga modified bacteria merA gene. Proc. Natl. Acad. Sci. 93:3182–3187
Rusin P, Quintana L, Brainard J, Strietelmeier B, Tait C,Ekberg S, Palmer P, Newton T & Clark D (1994) Solubili-zation of plutonium hydrous oxide by non-reducing bacteria.Environ. Sci. Technol. 28: 1686–1690
Salt DE, Blaylock M, Kumar NPBA, Dushenkov V, EnsleyBD, Chet I & Raskin I (1995) Phytoremediation: a novelstrategy for the removal of toxic metals from the environ-ment using plants. Biotechnol. 13: 468–474
Salt DE, Smith RD & Raskin I (1998) Phytoremediation. Ann.Rev. Plant Phys. Plant Mol. Biol. 49: 643–668
Samuelson P, Wernerus H, Svedberg M & Stahl S (2000)Staphylococcal surface display of metal-binding polyhistidylpeptides. Appl. Environ. Microbiol. 66: 1243–1248
Sand W, Gerke T, Hallmann R & Schippers A (1995) Sulfurchemistry, biofilm and the indirect attack mechanism-acrit-ical evaluation of bacterial leaching. Appl. Microbiol. Bio-technol. 43: 961–966
Satroutdinov AD, Dedyukhina EG, Chistyakova TI, WitschelM, Minkevich IG, Eroshin VK & Egli T (2000) Degrada-tion of metal–EDTA complexes by resting cells of thebacterial strain DSM 9103. Environ. Sci. Technol. 34: 1715–1720
Saxena P, Krishma Raj S, Dan T, Perras M & Vettakkoruma-kankav N (1999) Phytoremediation of heavy metalcontaminated and polluted soils In: Prasad MNV & HagenerJ (Eds.) Heavy Metal Stress in Plants; from Moleculesto Ecosystems (pp 305–329). Springer-Verlag, Berlin,Heidelberg
Schleck DS (1990) In situ treatment: When does it apply?Superfund ’90: Proceedings of the National Conference: Nov.26–28 (pp. 677–680), Environmental Protection Agency,Washington, D.C
154

Schultze-Lam S, Fortin D, Davis BS & Beveridge TJ (1996)Mineralization of bacterial surfaces. Chem. Geol. 132:171–181
Schwitzguebel JP, van der Lelie D, Baker A, Glass D &Vangrosweld J (2002) Phytoremediation: european andamerican trends, successes, obstacles and needs. J. SoilsSediments 2: 91–99
Seidel H, Loser C, Zensdorf A, Hoffman P & Scmerold R(2004) Bioremediation process for sediments contaminatedby heavy metals: feasibility study on a pilot scale. Environ.Sci. Technol. 38: 1582–1588
Sharma PK, Balkwill DL, Frenkel A & Vairavamurthy MA(2000) New Klebsiella planticola strain (Cd-1) grows anaero-bically at high cadmium concentrations and precipitatescadmium sulfide. Appl. Environ. Microbiol. 66: 3083–3087
Shen H & Wang YT (1993) Characterization of enzymaticreduction of hexavalent chromium by Escherichia coli ATCC33456. Appl. Environ. Microbiol. 59: 3771–3777
Sheppard SC & Evenden WG (1992) Bioavailability indices foruranium: effect of Concentration in eleven soils. Arch.Environ. Contam. Toxicol. 23: 117–124
Silver S (1994) Exploiting heavy metal resistance systems inbioremediation. Res. Microbiol. 145: 61–67
Smith WL & Gadd GM (2000) Reduction and precipitation ofchromate by mixed culture sulfate- reducing bacterialbiofilms. J. Appl. Microbiol. 88: 983–991
Smucker SJ (1996) Region 9 preliminary remediation goals;http://www.epa.gov/region9/waste/sfund/prg/index.html:U.S
Snoeyenbos-West O, Nevin KP, Anderson RT & Lovley DR(2000) Enrichment of Geobacter species in response tostimulation of Fe(III) reduction in sandy aquifer sediments.Microbiol. Ecol. 39: 153–167
Steinberg N, Blum J, Hochstein L & Oremland R (1992) Nitrateis a preferred electron acceptor for growth of freshwaterselenate-respiring bacteria. Appl. Environ. Microbiol. 501:426–428
Stumm W & Morgan JJ (1996) Aquatic Chemistry: ChemicalEquilibria and Rates in Natural Waters, 3. J. Wiley & Sons,New York, 1022 pp
Suzuki Y, Kelly SD, Kemner KM & Banfield JF (2003)Microbial populations stimulated for hexavalent uraniumreduction in uranium mine sediment. Appl. Environ. Micro-biol. 69: 1337–1346
Tan H, Champion JT, Artiola JF, Brusseau ML & Miller RM(1994) Complexation of cadmium by a rhamnolipid biosurf-actant. Environ. Sci. Technol. 28: 2402–1413
Tebo BM & Obraztsova AY (1998) Sulfate-reducing bacteriumgrows with Cr(VI), U(VI), Mn(IV), and Fe(III) as electronacceptors. FEMS Micro. Lett. 162: 193–198
Templeton AS, Trainor TP, Spormann AM & Brown Jr. GE(2003) Selenium speciation and partitioning within Burk-holderia cepacia biofilms formed on – Al2O3 surfaces.Geochim. Cosmochim. Acta 67: 3547–3557
Templeton AS, Trainor TP, Spormann AM, Newville M,Sutton SR, Dohnalkova A, Gorby Y & Brown Jr GE (2003)Sorption versus biomineralization of Pb(II) within Burk-holderia cepacia biofilms. Environ. Sci. Technol. 37: 300–307
Thomas RAP, Beswick AJ, Basnakova G, Moller R &Macaskie LE (2000) Growth of naturally-occurring microbialisolates in metal–citrate medium and bioremediation ofmetal–citrate wastes. J. Chem. Technol. Biotechnol.75: 187–195
Thomas RAP &Macaskie LE (1996) Biodegradation of tributylphosphate by naturally occuring microbial isolates and
coupling to the removal of uranium from aqueous solution.Environ. Sci. Technol. 30: 2371–2375
Thomas RAP & Macaskie LE (1998) The effect of growthconditions on the biodegradation of tributyl phosphate andpotential for the remediation of acid mine drainage waters bya naturally-occurring mixed microbial culture. Appl. Micro-biol. Biotechnol. 49: 202–209
Thomas RAP, Lawlor K, Bailey M & Macaskie LE (1998) Thebiodegradation of metal–EDTA complexes by an enrichedmicrobial population. Appl. Environ. Microbiol. 64: 1319–1322
Thompson-Eagle E, Frankenberger WT Jr. & Karlson U (1989)Volatilization of selenium by Alternaria alternata. Appl.Environ. Microbiol. 55: 1406–1413
Tichy R, Rulkens WH, Grutenhuis JTC, Nydl V, Cuypers C &Fajtl J (1998) Bioleaching of metals from soils or sediments.Water Sci. Technol. 37: 119–127
Tobin JM, White C & Gadd GM (1994) Metal accumulation byfungi: applications in environmental biotechnology. J. Ind.Microbiol. 13: 126–130
Treen-Sears ME, Volesky B & Neufeld RJ (1984) Ion exchange/complexation of the uranyl ion by Rhizopus biosorbent.Biotechnol. Bioeng. 26: 1323–1329
Trezek GJ (1990) Heavy metal-contaminated soil remediationat high throughput; Superfund ’90 (pp. 673–676): Proceed-ings of the National Conference, Nov. 26–28, EnvironmentalProtection Agency, Washington, D.C
Tucker MD, Barton LL & Thomson BM (1997) Reduction andimmobilization of molybdenum by Desulfovibrio desulfuri-cans. J. Environ. Qual. 26: 1146–1152
Turick CE, Apel WA & Carmiol NS (1996) Isolation ofhexavalent chromium-reducing anaerobes from hexavalentchromium-contaminated and noncontaminated environ-ments. Appl. Microbiol. Biotechnol. 44: 683–688
Turner JS & Robinson NJ (1995) Cyanobacterial metallothi-oneins: Biochemistry and molecular genetics. J. Ind. Micro-biol. 14: 119–125
Tsai LJ, Yu KC, Chen SF & Kung PY (2003) Effect oftemperature on removal of heavy metals from contami-nated river sediments via bioleaching. Water Res. 37: 2449–2457
U.S. Department of Energy, GJO (1999) Final site observa-tional work plan for the UMTRA project Old Rifle siteGJO-99-88-TAR
U.S. Department of Energy, O. o. B. a. ER (2004) posting date.Natural and Accelerated Bioremediation Research (NABIR)program. [Online.]
Utgikar VP, Harmon SM, Chaudhary N, Tabak HH, GovindR & Haines JR (2002) Inhibition of sulfate reducing bacteriaby metal sulfide formation in bioremediation of acid mine-drainage. Environ. Toxicol. 17: 40–48
Valls M & de Lorenzo V (2002) Exploiting the genetic andbiochemical capacities of bacteria for remediation of heavymetal pollution. FEMS Microbiol. Rev. 26: 327–338
Valls M., Atrian S, de Lorenzo V & Fernandez LA (2000)Engineering a mouse metallothionein on the cell surface ofRalstonia eutropha CH34 for immobilization of heavy metalsin soil. Nat. Biotechnol. 18: 661–665
Van der Lelie D, Schwitzguebel JP, Glass DJ & Baker A (2001)Assessing phytoremediation’s progress in the United Statesand Europe. Environ. Sci. Technol. 35(21), 447A–452A
Van Hullebusch ED, Zandvoort MH & Lens PNL (2003) Metalimmobilisation by biofilms : Mechanisms and analyticaltools. Re/Views Environ. Sci. Bio/Technol. 2: 9–33
155

Van Hullebusch ED, Lens PNL & Tabak HH (2005)Developments in bioremediation of soils and sedimentspolluted with metals and radionuclides. 3. Influence ofChemical Speciation and bioavailability on contaminantsimmobilization/mobilization bio-processes. Re/Views in En-vironmental Science & Bio/Technology, 4: 185–212
Vangronsveld J (2000) In-Situ inactivation and phytoremedi-ation of metal and metalloid contaminated soils In: Wise J,Cichon E, Inyang H & Stotmeister U (Eds.) Bioremediationof Contaminated Soils (pp 859–894). Marcel Dekker Inc.,New York
Van Roy S, Peys K, Dresselaers T & Diels L (1997) The use ofan Alcaligenes eutrophus biofilm in a membrane bioreactorfor heavy metal recovery. Res. Microbiol. 148: 526–528
Volesky B & Holan ZR (1995) Biosorption of heavy metals.Biotechnol. Prog. 11: 235–250
Wagner-Dobler I (2003) Pilot plant for bioremediation ofmercury-containing industrial wastewater. Appl. Microbiol.Biotechnol 62: 124–133
Wang CL, Clark DS & Keasling JD (2001) Analysis of anengineered sulfate reduction pathway and cadmium precip-itation on the cell surface. Biotechnol. Bioeng. 75: 285–291
Wang P, Mori T, Toda K & Ohtake H (1990) Membrane-associated chromate activity from Enterobacter cloacae.J. Bacteriol. 172: 1670–1672
Wang X & Brusseau ML (1993) Solubilization of some low-polarity organic compounds by hydroxypropyl-Beta-cyclo-dextrin. Environ. Sci. Technol. 27: 2821–2825
Wang X & Brusseau ML (1995) Simultaneous complexation oforganic compounds and heavy metals by a modified cyclo-dextrin. Environ. Sci. Technol. 29: 2632–2635
Warren LA & Haack EA (2001) Biogeochemical controls onmetal behaviour in freshwater environments. Earth Sci. Rev.54: 261–320
Warren LA, Maurice PA, Parmar N & Ferris FG (2001)Microbially mediated calcium carbonate precipitation: Impli-cations for interpreting calcite precipitation and for solid-phase capture of inorganic contaminants. Geomicrobiol.J. 18: 93–115
Watanabe ME (1997) Phytoremediation on the brink ofcommercialization. Environ. Sci . Technol. 31(4), 182A–186A
Watson JHP & Ellwood DC (1988) Presented at the Proc. Int.Conf on Control of Environmental Problems from MetalMines, Roros, Norway
Watson JHP & Ellwood DC (1994) Biomagnetic separation andextraction process for heavy metals from solution. MineralsEng. 7: 1017–1028
Weidemann DP, Tanner RD, Strandberg GW & Shumate SE(1981) Modelling the rate of transfer of uranyl ions ontomicrobial cells. Enzyme Microbial. Technol. 3: 33–40
Whicker FW, Hinton TG, MacDonell MM, Pinder JE &Habegger LJ (2004) Avoiding destructive remediation atDOE sites. Science 303: 1615–1616
White C & Gadd GM (1998) Accumulation and effects ofcadmium on sulfate reducing bacterial biofilms. Microbiol-ogy 144: 1407–1415
White C, Dennis JS & Gadd GM (2003) A mathematicalprocess model for cadmium precipitation by sulfate-reducingbacterial biofilms. Biodegradation 14: 139–151
White C, Sharman AK & Gadd GM (1998) An integratedmicrobial process for the bioremediation of soil contami-nated with toxic metals. Nat. Biotechnol. 16: 572–575
White C, Wilkinson SC & Gadd GM (1995) The role ofmicroorganisms in biosorption of toxic metals and radionuc-lides. Int. Biodeter. Biodegrad. 35: 17–40
White C & Gadd GM (1987) Inhibition of H+ efflux and K+
uptake and induction of K+ efflux in yeast by heavy metals.Tox. Assess. 2: 437–447
White C & Gadd GM (2000) Copper accumulation by sulfate-reducing bacterial biofilms. FEMS Microbiol. Lett. 183:313–318
Wu L, Mantgem PJvan & Guo X (1996) Effects of forage plantand field legume species on soil selenium redistribution,leaching and bioextraction in soils contaminated by agricul-tural drain water sediment. Arch. Environ. Contam. Toxicol.31: 329–338
Yong P & Macaskie LE (1995) Enhancement of uraniumbioaccumulation by a Citrobacter sp. via enzymatically-mediated growth of polycrystalline NH4UO2PO4. J. Chem.Technol. Biotechnol. 63: 101–108
Zehnder AJB & Stumm W (1988). Geochemistry and biogeo-chemistry of anaerobic habitats. In: Zehnder AJB (Ed)Anaerobic Microbiol (pp 1–38). Wiley, New York
Zorpette G (1996). Hanford’s nuclear wasteland. Sci. Am. May:88–97
156