developmental toxicity testingfrom animal to … · developmental toxicity testingfrom animal...
TRANSCRIPT

--~--------------------------
Developmental Toxicity Testing fromAnimal to-wards Embryonie Stem CellsCristian Pellizzer; Susanne Bremer and Thomas HartungECVAM (European Centre for the Validation of Alternative Methods), IHCP (Institute for Health and Consumer Protection),J.R.C. (Joint Research Centre), I-Ispra
SummaryDevelopmental toxicology is the study of undesirable effectson the development of the organism, which may result fromexposure before conception, from the period of prenataldevelopment, or postnatally during the time of sexualmaturation. The principal manifestations of developmentaltoxicity include: embryolethality, malformations, growthretardation, and functional impairment.In 2001, the European Commission published the futurechemieals policy entitled "White Paper: Strategy for a FutureChemieals Policy". The new regulation requires a toxicologicalevaluation with strong emphasis on reproductive toxicity, byusing in vitro methods, especially for those chemieals marketedat more than I ton per year. For this reason, the establishmentof in vitro models capable of detecting major undesirablemanifestations in the fetus, are urgently required. The aim of thepresent review is to explore the capacity of existing in vitrosystems, based on embryonie stem (ES) cells, to identifyembryotoxicity with a focus on specific effects such as terato-genicity and growth retardation. In addition, we discuss thepossibility to adapt the mouse ES cells based tests to humanES cells, avoiding inter-species variations in developmentaltoxicity studies and address related ethical issues. Consideringthe different manifestations of developmental toxicity, only abattery of in vitro tests will provide the necessary informationfor regulatory developmental toxicity assessment.
Zusammenfassung: Entwicklungstoxizitätstestung: Vom Tier-modell zu embryonalen StammzellenIn der Entwicklungstoxikologie werden nachteilige Effekte aufdie Entwicklung eines Organismus studiert, die sich vor der Be-fruchtung, während der pränatalen Entwicklung oder postnatalin der Zeit bis zur Geschlechtsreife auswirken können. Diewichtigsten durch Entwicklungstoxizität bedingten Symptomesind: Embryoletalität, Missbildungen, vermindertes Wachstumoder Funktionsstörungen.Die Europäische Kommission veröffentlichte 2001 die zu-künftige Chemikalienpolitik "White Paper: Strategy for aFuture Chemieals Policy". Diese Regelungen fordern einetoxikologische Beurteilung mit Schwerpunkt auf Reproduktion-stoxizität mittels in vitro Methoden, besonders für Chemikalien,deren jährliche Produktion ein Volumen von 1 Tonne über-schreitet. Aus diesem Grund wird dringend die Entwicklung vonin vitro Modellen benötigt, die die wichtigsten unerwünschtenSymptome im Fötus abdecken. Die vorliegende Review-Arbeituntersucht die Fähigkeit von in vitro Systemen - basierend aufembryonalen Stammzellen -, Embryotoxizität nachzuweisen mitFokus auf spezifische Effekte wie Teratogenität und Wachs-tumsstörungen. Zusätzlich werden aus ethischer Sicht dieMöglichkeiten der Adaptierung von Mausstammzell-Tests aufhumane Stammzellen diskutiert, um Interspezies- Variationen inEntwicklungstoxikologischen Studien zu vermeiden. In Anbe-tracht der unterschiedlichen Erscheinungsbilder einer Ent-wicklungstoxizität kann allerdings nur eine Batterie von invitro Tests die nötigen Informationen zur regulatorischen Be-wertung bezüglich Entwicklungstoxizität liefern.
Keywords: embryonie stem cells, teratogenicity, developmental toxicity, alternatives to animals experiments, malformations,growth retardation
1 Introduction
Developmental toxicology is an importantarea of toxicology. Rosano et al. (2000)showed that birth defects are the princi-ple cause of infant mortality as assessedin 36 countries. Children born with majorbirth defects are normally affected bymental, physical and social disorders
(Rosano et al., 2000; MacLeod, 1993;Yoon et al., 1997). To date, significantprogress has been made in order todetermine causes of birth defects. Birthdefects is a term used to indicate thosemalformations observable at birth (con-genital) or thereafter. Birth defect indi-cates structural or anatomical defects, butit can also inc1ude physiological or func-
Received 21 February 2005; received in final form and accepted tor publication 9 May 2005
ALTEX 22, 2/05
tional and behavioural defects. For themajority of those, the etiology is un-known, for some it is only poorly estab-lished. The causes of birth defects arevarious and can be summarised as genet-ic causes (autosomal genetic diseases,cytogenetic; 15-20%), environmentalcauses (maternal conditions and infec-tions, mechanical problems, chemicalsldrugs/radiation; 10%), preconception ex-posures (unknown percentage) and un-
47

PELLIZZER ET AL. m.....-------------~-
known causes (65%) (Brent and Beck-man, 1990).Wi1son 1977 considered chemical
agents responsible for 4-6% of the totalbirth defects. Nevertheless, in order toreduce risks of deviant development, it isa worthwhile goal to identify thosechemical-agents present in the environ-ment and in consumer products to whichthe gravid mother and her conceptus arepotentiallyexposed.The sciences of birth defects "terato-
logy", or the process of induction of ma1-formation "teratogenesis", from the Greekword root "teras" meaning malformationor monstrosity. Three fundamental princi-ples of teratogenesis govern the chemicalpotential or the capability to inducecongenital malformations in human orin other animal species. ParaphrasingKamofsky (1965), these principles canbe illustrated by the axiom: "A terato-genic response depends upon the admin-istration of a specific treatment of a par-ticular dose to a genetically susceptiblespecies when the embryos are in a sus-ceptible stage of developrnent,"As assumed by Wilson 1973, the terato-
genesis depends on the genotype of theconceptus, the way in which this interactswith environmental factors, the develop-mental stage and the mechanism (patho-genesis) as well. In addition, manifesta-tion of adverse development increase inincidence and degree as dosage increasesfrom no-effect to totally lethallevel.
The four main manifestations of devel-opmental toxicity are malformations,growth retardation, embryolethality andfunctional impairment (Wilson, 1973)(see Fig.1).MalformationsMost types of malformations are observ-able in the majority of all species ofanimals, but specific malformations ortheir incidence are species-dependent.For example, eye defects, exencephaly,polydactyly, and eleft palate are mostfrequent in mice (Flynn, 1968; Kalter,1968). In rabbits it is easy to observelimb defects, umbilical hernians, andcraniofacial defects more frequentlythan other types of malformations (Chaiand Degenhardt, 1962; Staples and Holt-kamp, 1963). Nonetheless, some chemi-cals, capable of inducing malformations,may also produce other elasses of devel-opmental toxicity effects, consequentlyan agent that causes developmental toxiceffects may not necessary be consideredas teratogenic.Growth retardationAnother aspect to be considered in devel-opmental toxicity testing is intrauterinegrowth retardation (IUGR). A generallyaccepted definition for IUGR is: "Birthweight less than the tenth percentile forgestational age" (Lubchenco et al.,1963). An alternative definition is: "Birthweight more than two standard devia-tions below the mean for gestational age,corresponding to approximately the third
Fig. 1: Principal manifestations in developmental toxicity.Embryolethality, malformations, growth retardation, and functional impairment, are themost important manifestations in developmental toxicity to be considered in an in vitrotest strategy. Some other reproductive parameters should be taken into account.
48
percentile of the intrauterine growthcurves" (Gruenwald, 1966). Numerousagents are known to cause IUGR. In alarge human study, the incidence ofIUGR was 5.3% and perinatal mortalityamong these infants occurred three timesmore often than normal (Low and Gal-braith, 1974). In Callan and Witter(1990) 86% of perinatal deaths werefound in the IUGR group. In addition, upto 20% of spontaneous abortions exhibitsevere embryonie growth retardation.IUGR is common in infants that exhibitsevere and multiple malformations(Warkany, 1971). In addition, congenitalinfections, neonatal deaths, and longterm neurological and intellectualdeficits have been noted more often ininfants affected by IUGR (Miller, 1981).EmbryolethalityDeath of the offspring is another aspectof developmental toxicity. Embryonieand early fetal death occurs in one outof every two pregnancies in humans(Shepard and Fantel, 1979). Fetal deathis often associated with congenital mal-formations. The causes of mortality maybe due to direct effects to the conceptus(alcohol, anti cancer agents, lithium,methyl mercury and retinoids analogues)or indirect (maternally mediated) (Kalter,1980). Maternally mediated effects aregenerally associated with: maternalhomeostatic changes due to diabetes andphenylketonuria; and maternal toxicityresulting from alcohol abuse, use ofaminopterin and trimethadione (Khera,1985, 1987). Some teratogens causeprimarily morphological modificationsand in some situations this may lead toembryonie death. In humans, correlationbetween malformations and death hasbeen demonstrated (Haas and Schotten-feld, 1979).Functional impairmentRecently, parameters such as motorability, sociability, emotional, and learn-ing capacity are considered as effects inthe developmental toxicity field. Indeed,some evidence indicates that specifictypes of behavioral alterations may resultafter exposure to certain agents, duringcritical periods in fetogenesis. Theseabnormalities are the result of modi-fication occurring during development ofspecific neurotransmitter systems, afterchemieals insult (Leonard, 1981). Many
ALTEX 22, 2/05

~ PELLIZZER ET AL.---~----------------------------------------
compounds are well-known causes offunctional impairment, for example as-pirin (Okamoto et al., 1986), ethoxyetha-nol (Nelson et al., 1981) and hydrox-yurea (Vorhees et al., 1979). In recentyears, additional behavioural tests havebecome constituents of developmentaltoxicity assessment, especially in thefield of pharmacology.In addition to the classes described
above, there are some other parametersto consider in developmental toxicitystudies (see Fig. 1). These are reproduc-tive parameters that may have relevanceto humans. Male-mediated effects areone of those parameters, e.g. when de-fects to offspring result from sperm cells.For example, several studies in the pro-genies that came from male rabbits treat-ed with colchicin or thalidomide, showedsevere malformations (Chang, 1944; Lut-wak-Mann, 1964). According to a reviewproposed by Davis et al. (1992) somechemical agents were classified accord-ing to their capacity to cause male-medi-ated adverse reproductive outcomes.Another parameter is the developmen-
tal variation. Minor aberrations in struc-ture and variations in ossification arerelated to this factor. These effects takeplace more often than malformations,and the comrnon consequences to thefetus are delay in growth, minor changesin structure and alteration in the normaldifferentiation.The influence of matemal toxicity is
another factor to be taken into account interatology assessment. It becomes obvi-ous that toxicity to the embryo may bemodified or influenced by toxicity to themother. Nevertheless, it is poorly or notyet understood why, and how toxicityto the mother has direct effects on fetusdevelopment.Some pharmacokinetic considerations
are essential in teratology assessment.Pregnancy itself alters pharmocokineticsthrough physiological changes. For thisreason, the study of pharmocokineticproperties of chemieals or drugs be-comes essential, in order to quantify thereal amount of exposure to the fetus andthe resulting toxicological effect.The aim of the present review is to give
an overview regarding the use of in vitrosystems, based on mouse embryoniestern cells, in developmental toxicity
ALTEX 22, 2/05
testing. The application of these methodsto identify phenomena such as terato-genicity and growth retardation isdiscussed. In order to avoid inter-speciesvariations, the possible use of humanembryonie stern cells in developmentaltoxicity studies is explored and relatedethical issues are taken into account.
2 Animal models used indevelopmental toxicity studies
Mouse, rat and rabbit are the generallyaccepted species of laboratory animals.However in recent years, other specieshas been proposed, such as hamster,guinea pig, dog, swine, ferret, andvarious species of primates. Neverthe-less, the results obtained from testingthese species in developmental toxicitystudies, demonstrate that few provedhuman teratogens. Following the advicemade by Wilson (1978) a critical pointto be considered for safety evaluation, isthe presence, in the model used, of thematemal-placenta-embryo relation. Inaddition Wilson (1975) advised the useof animal models able to metabolise,distribute and transfer the compoundacross the placenta in ways sirnilar tohumans.In one large comparative study
(Schardein, 1983), the rabbit and the pri-mate were considered the most suitablespecies for general screening. In Schard-ein and Keller (1989) some other animalmodel were evaluated and rat, mouse,hamster, several species of primate andrabbit showed quite reliable ability toidentify human developmental toxicants,in descending order. To date, no singlespecies has been identified as mostpromising animal model for develop-mental toxicity testing.
The embryotoxic and in particular theteratogenic potential of a substance istested using the Prenatal DevelopmentalToxicity Study (OECD, Organisationfor Economic Co-operation and Devel-opment, Test Guidelines 414, Anon,2001b). Other OECD guidelines areused, in order to evaluate effects on pre-/postnatal development and on fertility(see Tab. 1). Such studies, especiallythe two- or multigeneration study, arecost- and time-consuming. They con-sume up to 3,200 anirnals per substance.
3 Future European chemiealspolicy
Recently the European Comrnission re-ported to the Council and the EuropeanParliament the statistics on the number ofanimals used for experimental and otherscientific purposes (Anon, 2005). In2002, it has been estimated, that in theEuropean Members States, 10.7 millionanimals have been sacrificed. In toxico-logical and other safety evaluation, thenumber of animals used represents9.93% of the total and about 12.1% ofthese animals are used for reproductiveand developmental toxicity tests.In 2001, the European Comrnission
published a policy statement on futurechemieals regulation and risk reductionentitled "White Paper: Strategy for aFuture Chemieals Policy" (Anon, 2001a).The new chemieals policy airns to finalisethe risk assessments within a 12-15 yearperiod for all chemieals marketed at morethan 1 ton per year. For this reason, itbecomes urgent to identify data on thehazardous properties of these chemicals.In addition, for those chemieals not testedin the past it will be necessary to re-evaluate their toxicological potential.
Tab. 1. OECD guidelines used to evaluate pre-/postnatal and fertility effects.
OECD Guideline Description References
OECD TG415: One-Generation Reproduction Toxicity Study Anon, 1983
OECD TG416: Two-Generation Reproduction Toxicity Study Anon,2001c
OECD TG421: Reproduction/ Anon,2001dDevelopmental Toxicity Screening Study
OECD TG422: Combined Repeated Dose Toxicity Study Anon, 1996with the Reproduction/ DevelopmentalToxicity Screening Study
49

PELLIZZER ET AL. m-...-------------~-
Therefore, the European Union is inthe process of introducing a new systemfor the Registration, Evaluation andAuthorisation of new and existing Chem-icals (REACH) in order to harmonise thetesting requirements for new and existingchernicals. The White Paper and the draftlegislatiorf suggest that in the first phasetesting should be restricted to in vitrotests.
4 In vitro model fordevelopmental toxicityassessment
In vitro systems for developmental toxi-cology testing can be subdivided intothree different c1asses: cell cultures (e.g.Embryonie Stern Cells Test (EST)),organ cultures (e.g. Micromass assay),and embryo cultures (e.g. Whole embryoculture). Brown (1987) and Brown et al.(1995) presented an extensive overviewof the existing in vitro systems.Ease of performance and reduced or
no animal use are the most importantadvantages of cell cultures and for thisreason, these models not requiring theuse of experimental animals arefavoured. Systems like embryo culturesare complex systems which provide agreater understanding of the develop-mental mechanisms involved. However,these models are more laborious andanimal consurning.Other alternatives to animal models
are the organ culture systems. They needprimary animal material, and they re-quire specific technical infrastructures,which make the standardisation of themodel more difficult. Limb buds, digits,palatal shelves, lung, intestine, and sexorgans are examples of such organ cul-ture systems (Faustman, 1988).It was estimated that several millions
of animals would have to be sacrificedaccording to the needs of the WhitePaper, developmental toxicology con-suming up to two thirds of these (van derJagt et al., 2004).1t becomes obvious thatnew predictive screening methods for de-velopmental toxicity testing are urgentlyrequired, especially since this field willmost probably consume the highestnumbers of animals.
50
Numerous workshops have been heldin order to evaluate the most promisingalternative tests. An ECVAM (EuropeanCentre for the Validation of AlternativeMethods) workshop was held in 1994(Brown et al., 1995) and the participantssuggested the use of Xenopus embryos,mammalian embryo as well as micro-mass cultures and murine embryoniestern (ES) cells. However, in May 2000the Interagency Coordinating Commit-tee on the Validation of AlternativeMethods (ICCVAM) after evaluating theFrog Embryo Teratogenesis Assay(Xenopus (FETAX)), which is consid-ered a teratogenesis assay, reported thatFETAX is not sufficiently validated oroptimised for regulatory applications(Anon, 2000).In addition, the participants of a
workshop held in 1981 on in vitroteratogenesis, agreed that only a batteryof test will provide relevant informationin teratogenic assessment (Kimmel etal., 1982). Indeed, taking into accountthe comp1exity of embryo genesis, it isc1ear that a single in vitro model is notable to substitute in vivo testing. There-fore, only a strategy of in vitro tests willbe able to cover the complete range ofmechanisms of development and theirinteraction (Bremer and Hartung, 2004).
5 Development of in vitrosystems by using embryonicstem cells for developmentaltoxicity testing
In order to detect chemical effects duringthe process of differentiation, several ap-proaches based on embryonie stern cellshave been used. The EST (Spielmann etal., 1997) is one of the in vitro validatedsystems and it is based on ES cells,which permit the c1assification of cherni-cals in strong, weak, and non-embryo-toxic. Nevertheless, for regulatory accep-tance, EST needs to be improved. Duringan ECVAM workshop held in January2003, a group of experts have reviewedthe EST and they advised ECVAM toinc1ude supplementary endpoints ableto identify effects on nervous system(Strubing et al., 1995), skeletal system(zur Nieden et al., 2003) and cardio-vas-
cular system (Boheler et al., 2002). Theability of the ES cells to differentiateinto a wide range of tissues, which mightbe the target of a teratogen, provides theunique opportunity to use them in terato-genesis studies. Furthermore, the expertsrecommended to add metabolic compe-tences to the EST as well as the other validated embryotoxicity tests.In the last few years, teratogenic ef-
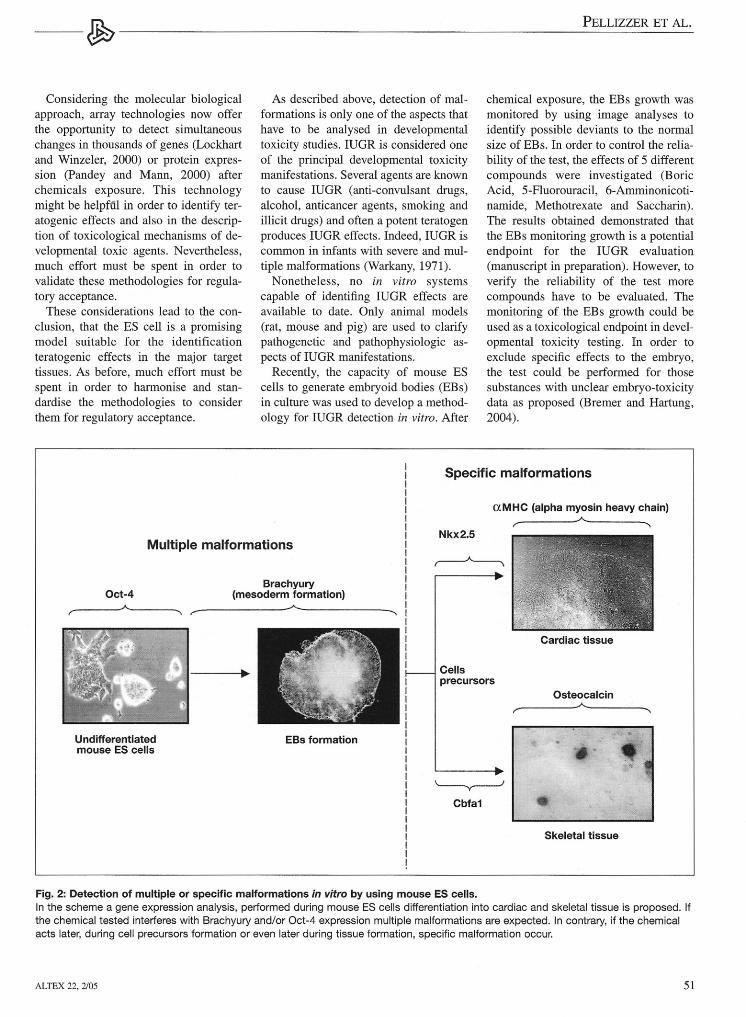
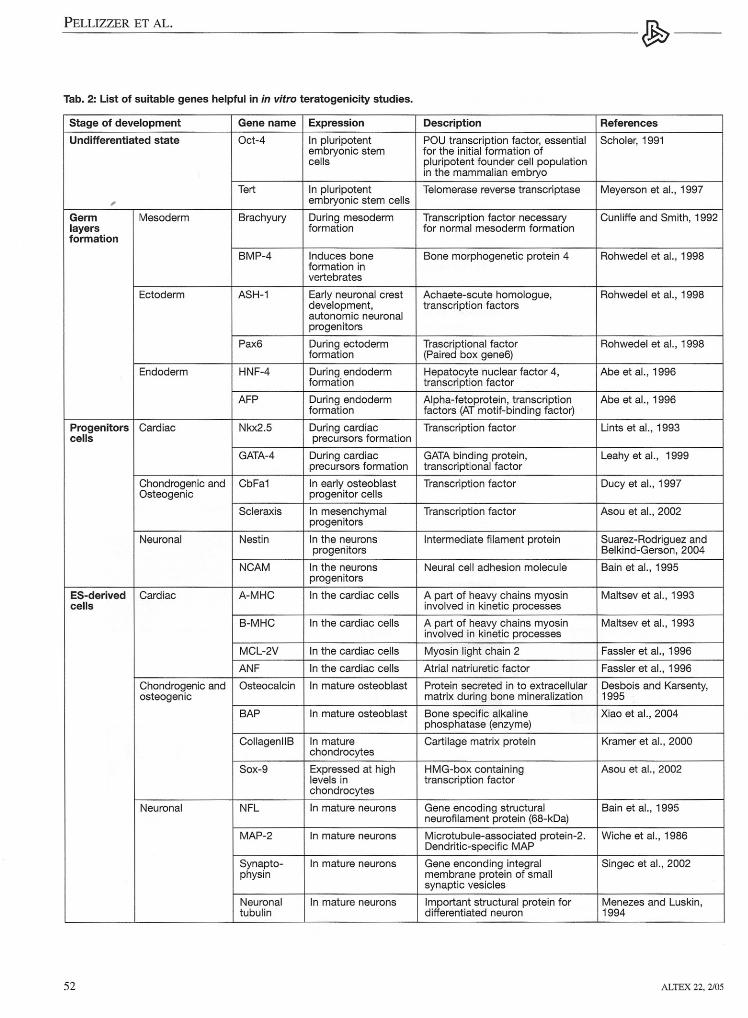
fects of chemieals occurring duringES cells differentiation have been anal-ysed by using molecular biological andimmunological endpoints. Cultivation oftransgenic ES cells with green fiuores-cence pro tein (GFP) driven by a specifictissue gene promoter is another elegantmethod used to detect modification inthe normal ES differentiation (Metzgeret al.; 1996, Kolossov et al.; 1998,Bremer et al., 2001; Paparella et al.,2002). In Wobus et al. (1997), Bigot etal. (1999), zur Nieden et al. (2001),Bremer et al. (2001), Schmidt et al.(2001) and Seiler et al. (2002) a molecu-lar approach was used in order to moni-tor changes in the expression of specifictissue genes, during cardiac ES cells dif-ferentiation after chemieals exposure. Inthese studies only tissue specific mark-ers were analysed. In Seiler et al. (2002,2004) an immunologica1 approach wasproposed based on intracellular stainingand fiow cytometry to detect changes insarcomeric myosin heavy chain (MHC)and alpha-actinin, during ES cells car-diac differentiation. In order to obtainmore detai1ed information at which de-velopmental stage the chemical is active,a different approach was proposed re-cently (Pellizzer et al., 2004a, 2004b)(see Fig. 2). The described method aimsto analyse the expression of genes in-volved in early and later stages of car-diac and skeletal ES cells differentiation,in order to detect multiple of specificmalformations after chemieals exposure.The in viva data of the chemieals testedwere correctly confirmed by the in vitrotests using the mentioned approach,demonstrating the usefulness of theES system in teratogenicity studies. Asuitable list of genes which may be use-ful in the identification of teratogenicityeffects in the principal target tissues aresummarised in Table 2.
ALTEX 22, 2/05
m...... PELLIZZER ET AL.-~-------------
Considering the molecular biologicalapproach, array technologies now offerthe opportunity to detect simultaneouschanges in thousands of genes (Lockhartand Winzeler, 2000) or protein expres-sion (Pandey and Mann, 2000) afterchemieals exposure. This technologymight be helpf 1 in order to identify ter-atogenic effects and also in the descrip-tion of toxicological mechanisms of de-velopmental toxic agents. Nevertheless,much effort must be spent in order tovalidate these methodologies for regula-tory acceptance.These considerations lead to the con-
clusion, that the ES cell is a promisingmodel suitable for the identificationteratogenic effects in the major targettissues. As before, much effort must bespent in order to harmonise and stan-dardise the methodologies to considerthem for regulatory acceptance.
As described above, detection of mal-formations is only one of the aspects thathave to be analysed in developmentaltoxicity studies. IUGR is considered oneof the principal developmental toxicitymanifestations. Several agents are knownto cause IUGR (anti-convulsant drugs,a1cohol, anticancer agents, smoking andillicit drugs) and often a potent teratogenproduces IUGR effects. Indeed, IUGR iscommon in infants with severe and mul-tiple malformations (Warkany, 1971).Nonetheless, no in vitro systems
capable of identifing IUGR effects areavailable to date. Only animal models(rat, mouse and pig) are used to clarifypathogenetic and pathophysiologic as-pects of IUGR manifestations.Recently, the capacity of mouse ES
cells to generate embryoid bodies (EBs)in culture was used to develop a method-ology for IUGR detection in vitro. After
chemical exposure, the EBs growth wasmonitored by using image analyses toidentify possible deviants to the normalsize of EBs. In order to control the relia-bility of the test, the effects of 5 differentcompounds were investigated (BoricAcid, 5-Fluorouracil, 6-Amrninonicoti-narnide, Methotrexate and Saccharin).The results obtained demonstrated thatthe EBs monitoring growth is a potentialendpoint for the IUGR evaluation(manuscript in preparation). However, toverify the reliability of the test morecompounds have to be evaluated. Themonitoring of the EBs growth could beused as a toxicological endpoint in devel-opmental toxicity testing. In order toexclude specific effects to the embryo,the test could be performed for thosesubstances with unclear embryo-toxicitydata as proposed (Bremer and Hartung,2004).
Specific malformations
Multiple malformations
Oct-4Brachyury
(mesoderm formation)
Undifferentiatedmouse ES cells
Cellsprecursors
EBs formation
'--y---l
Cbfa1
aMHC (alpha myosin heavy chain)
Cardiac tissue
Osteocalcin
.'Skeletal tissue
Fig. 2: Detection of multiple or specific malformations in vitro by using mouse ES cells.In the scheme a gene expression analysis, performed during mouse ES cells differentiation into cardiac and skeletal tissue is proposed. Ifthe chemical tested interferes with Brachyury and/or Oct-4 expression multiple malformations are expected. In contrary, if the chemicalacts later, during cell precursors formation or even later during tissue formation, specific malformation occur.
ALTEX 22, 2/05 51
PELLIZZER ET AL. m-..----------------------------------------~---
Tab. 2: List of suitable genes helpful in in vltro teratogenicity studies.
Stage of development Gene name Expression Description References
Undifferentiated state Oct-4 In pluripotent POU transcription factor, essential Scholer, 1991embryonic stem for the initial formation ofcells pluripotent founder cell population
in the mammalian embryo
Tert In pluripotent Telomerase reverse transcriptase Meyerson et al., 1997, embryonic stem cells
Germ Mesoderm Brachyury During mesoderm Transcription factor necessary Cunliffe and Smith, 1992layers formation for normal mesoderm formationformation
BMP-4 Induces bone Bone morphogenetic protein 4 Rohwedel et al., 1998formation invertebrates
Ectoderm ASH-1 Early neuronal crest Achaete-scute homologue, Rohwedel et al., 1998development, transcription factorsautonomic neuronalprogenitors
Pax6 During ectoderm Trascriptional factor Rohwedel et al., 1998formation (Paired box gene6)
Endoderm HNF-4 During endoderm Hepatocyte nuclear factor 4, Abe et al., 1996formation transcription factor
AFP During endoderm Alpha-fetoprotein, transcription Abe et al., 1996formation factors (AT motif-binding factor)
Progenitors Cardiac Nkx2.5 During cardiac Transcription factor Unts et al., 1993ceUs precursors formation
GATA-4 During cardiac GATA binding protein, Leahy et al., 1999precursors formation transcriptional factor
Chondrogenic and CbFa1 In early osteoblast Transcription factor Ducy et al., 1997Osteogenic progenitor cells
Scleraxis In mesenchym al Transcription factor Asou et al., 2002progenitors
Neuronal Nestin In the neurons Intermediate filament protein Suarez-Rodriguez andprogenitors Belkind-Gerson, 2004
NCAM In the neurons Neural cell adhesion molecule Bain et al., 1995progenitors
ES-derived Cardiac A-MHC In the cardiac cells Apart of heavy chains myosin Maltsev et al., 1993ceUs involved in kinetic processes
B-MHC In the cardiac cells Apart of heavy chains myosin Maltsev et al., 1993involved in kinetic processes
MCL-2V In the cardiac cells Myosin light chain 2 Fassler et al., 1996
ANF In the cardiac cells Atrial natriuretic factor Fassler et al., 1996
Chondrogenic and Osteocalcin In mature osteoblast Protein secreted in to extracellular Desbois and Karsenty,osteogenic matrix during bone mineralization 1995
BAP In mature osteoblast Bone specific alkaline Xiao et al., 2004phosphatase (enzyme)
CollagenllB In mature Cartilage matrix protein Kramer et al., 2000chondrocytes
Sox-9 Expressed at high HMG-box containing Asou et al., 2002levels in transcription factorchondrocytes
Neuronal NFL In mature neurons Gene encoding structural Bain et al., 1995neurofilament protein (68-kDa)
MAP-2 In mature neurons Microtubule-associated protein-2. Wiche et al., 1986Dendritic-specific MAP
Synapto- In mature neurons Gene enconding integral Singec et al., 2002physin membrane protein of small
synaptic vesicles
Neuronal In mature neurons Important structural protein for Menezes and Luskin,tubulin differentiated neuron 1994
52 ALTEX 22, 2/05

~ PELLIZZER ET AL.-----~------------------------------------------------------------
6 Humanisation of mouseEScell existing model andethical aspects
An advantage of the use of the ES cellsfor developmental toxicity testing is theavailability of human ES (hES) cells(Thomson et al~ 1998). In fact, the exist-ing test based on murine ES cells couldbe adapted to hES cells. Not all mam-malian species are equally susceptible orsensitive to the toxic influence of achemical and for this reason a humanisedsystem should be more predictive. In-deed, a chemical can be teratogenic inone species but not in other species.Human models will avoid interspeciesvariations, which are a crucial factor indevelopmental toxicity testing, hence noextrapolations to human might be neces-sary any more.Recent publications have demonstrat-
ed that hES cells are able to produceEBs, expressing genes involved in allthree germ layers formation and indifferentiation of several cell types(Schuldiner et al., 2000). In addition, theability of hES cells to differentiate invitro into neuronal, skin, adrenal andkeratinocyte (Reubinoff et al. , 2000;Zhang et al., 2001; Green et al., 2003),blood, endothelial, kidney, bone, muscle,heart (Kaufman et al., 2001; Kehat et al.,2002,2003; Levenberg et al., 2002; Xu etal., 2002; He et al., 2003), pancreasand liver cells (Schuldiner et al., 2000,Assady et al., 2001; Eiges and Ben-venisty, 2002) has been demonstrated.
Moreover, several studies have shownthe capacity of hES to generate cardio-myocytes (Kehat et al., 2002; Kehat etal., 2003; He et al. , 2003; Xu et al., 2002)electrophysiologically similar to normalhuman cardiomyocytes, and foetal ven-tricular myocytes (Mummery et al. ,2003). In the validated EST cardiomy-ocytes differentiation represents an end-point and hES cells could substitutemouse cells in the test.This capability of the hES cells to dif-
ferentiate into all germ layer derivativespromises to provide a system able toidentify possible effects on a wide rangeof target tissues. In addition, the systemsbased on hES will avoid interspecies dif-ferences.Moreover, the use of hES in research
will also offer the chance to study humandevelopment, providing more preciseinformation concerning events that cannotbe studied directly in the human fetus.Using this information, the pre-ventionand the treatment of unusual humandevelopment might become possible. Inaddition, the reduction of risks related todrug exposure should be achievable.The optimisation and the standardisa-
tion of hES cell in vitro models are themost important goals. However, at themoment hES cell research representsan ethical issue. In order to respect theethical and moral value of the human em-bryo, many European countries have tak-en legislative actions (see Tab. 3). At thesame time the European Communitysupports the expansion of funding for
hES cell research. Indeed, in the contestof the 6th Research Framework Pro-gramme (FP6), adopted in 2002, theEuropean Union (EU) provided financialsupport for hES cell research, especiallyto fight major human diseases.In order to protect the human embryo,
In 2003 guidelines on EU-funded hEScell research were adopted by the Com-mission (Anon, 2003a). The aim of theCommission proposal was not to createethical principles that must be respectedby the EU Member States, but to set upconditions for EU FP6 research, in whichstern cells from human supernumeraryembryos are involved.The most important topics regulate
by the Commission proposal are thefollowing:• "EU will not fund human embryoniestern cell research where it is forbiddenby a Member State";
• "Human embryonie stern cells canonly be derived from supernumeraryembryos that are donated for researchby parents and that were created before27 June 2002, the date of the adoptionof the Framework Programme";
• "Potential research project partnersapplying for EU funding must seekethical advice at national or locallevelin Member States where the researchwill take place, even in countrieswhere obtaining such ethical advice isnot mandatory";
• "Research will be funded only whenit is demonstrated that it meets particu-larly important research objectives";
Country
Tab. 3: Regulation of the use of human embryonic stern cells in the European Member States (update September 2003).
Italy, Luxembourg, Portugal
Austria, Spain, France, Ireland
Austria, Denmark, Germany, Spain, Finland,France, Greece, Ireland, Netherlands, Portugal
No specific regulation concerning human ESC research----------------------------~Legal veto tor the derivation of ES cells from human embryos----~--~----------------~Legal veto for the production of human embryos for research aims.Veto for the derivation of ES cells from human embryos, by law or by ratificationof the Convention of the Council of Europe on Human Rights andBiomedicine (Oviedo on April 4, 1997)
Policy adopted
Belgium, Denmark, Finland, Greece,Netherlands, Sweden, United Kingdom
Germany
Legal permission for the derivation of hES cells from supernumerary embryos
Legal veto for the derivation of ES cells from human embryos.Legal permission for the import of hES cell lines, which were createdbefore January 2002
Belgium, United Kingdom
Compiled from Anon, 2001e and Anon, 2003b
ALTEX 22, 2/05
Legal perm iss ion for the production of human embryos for ES cells research
53

PELLIZZER ET AL. m....----------------------------------------~---
• "Research will be funded only whenthere is no adequate alternative avail-able. In particular, it must demonstrat-ed that one cannot use existing embry-onic or adult stern celllines";
• "Traceability of stern cells will berequired".The emjiloyment of hES cells in the re-
search field is encouraged by the EU. Infact, within FP6 the EU sponsors an Inte-grated Project titled "ReProTect", with aduration of 5 years and a budgetof 9.1 rnillions of EURO. 35 differentpartners from acadernia, SMEs, govern-mental institutions and others are in-volved. The principal aim of ReProTectis to create new in vitro models anduse existing in vitro models in orderto set up a test strategy that will provideinformation on the chemical hazardsto the mammalian reproductive cyc1e.In ReProTect the investigation of thepotential of hES cells for embryotoxicitytesting is adressed.
7 Conclusions
The main goal of the present review is topoint out the potential of the ES cells inin vitro hazard assessment. Many studieshave demonstrated the usefulness ofES cells in the identification of develop-mental toxicity effects of chemicals.Nevertheless, in order to replace theuse of animals in laboratory studies, it isnecessary to consider many aspectsregarding the use of in vitro systems.Metabolism and biotransformation ingeneral, are one of the most irnportanttopics. Due the complexity of thewhole organism, not all aspects can bernirnicked in vitro, but the creation ofhuman metabolism based-models wouldbe a potent tool that could support the EScells and other in vitro systems. Inaddition, the understanding of the toxico-logical mechanisms of congenital defectsand genetic and environmental factors insuch mechanisms is a crucial topic indevelopmental studies. For this reason, itbecomes obvious that both studies ofbiological mechanisms of developmentaltoxicants, their toxicological assessmentand exposure to pregnant women will beessential in the future.
54
ReferencesAbe, K., Niwa, H., Iwase, K. et al.
(1996). Endoderm-specific geneexpression in embryonie stern cellsdifferentiated to embryoid bodies.Exp. Cell. Res. 229, 27-34.
Anon (2005). The Fourth report on thestatistics on the number of animalsused for experimental and other seien-tific purposes in the Member States ofthe European Union. http://www.nca-nl.org/EnglishIDocs/EU%20stats.pdf(access 01/2005).
Anon (2003a). IP/03/969, Brussels,9 July 2003. European Comrnissionproposes strict ethical guidelines onEU funding of human embryonie sterncell research. http://europa.eu.int/rapid/pressReleasesAction.do? refer-ence=IP/03/969&format=HTML&aged=O&language=N&guiLanguage=en(access 01/2005).
Anon (2003b). EUROPEAN COMMIS-SION Directorate General: RE-SEARCH. Survey on opinions fromNational Ethics Comrnittees or sirnilarbodies, public debate and national leg-islation in relation to human embryon-ic stern cell research and use. Volume Iin EU Member States. europa.eu.int/commlresearch! biosociety/pdf/cata-logue_stem_cells.pdf (access 0112005).
Anon (2001a). White Paper on a Strategyfor a Future Chernicals Policy. Brus-sels: Commission of the EuropeanCommunities. http://europa.eu.int/commlenviroment/chernicals/whitepa-per.htm (accessed 6/2003).
Anon (200lb). OECD Guidelines fortesting of chemicals, Guideline 414:Prenatal developmental toxicity study.http://www.sourceoecd.org/content/templates/co/co_main_oecdguid.htm?comm=oecdguid (accessed 30/2003).
Anon (200lc). OECD guideline for test-ing of chemieals 416: two-generationreproduction toxicity study. Updateguideline, adopted 22nd January 2001.
Anon (2001d). OECD Guidelines fortesting of chernicals, Guideline 421:Two-generation reproduction toxicitystudy. Updated guideline, adopted 22ndJanuary 2001.
Anon (2001e). European Science Foun-dation Policy Briefing. Human sterncell research: scientific uncertainties
and ethical dilemmas. www.esf.org/medias/ESPB14.pdf (access 01/2005).
Anon (2000). Frog Embryo Teratogene-sis Assay-Xenopus (FETAX). http://icc vam.niehs. nih. gov /methods/fetax.htm.
Anon (1996). OECD Guidelines for test-ing of chernicals, Guideline 422: Com-bined repeated dose toxicity study withthe reproductionldevelopmental toxici-ty screening test Original Guideline,adopted 22nd March 1996.
Anon (1983). OECD Guidelines for test-ing of chernicals, Guideline 415: One-generation reproduction toxicity study.Original Guideline, adopted 26th May1983.
Asou, Y., Nifuji, A., Tsuji, K. et al.(2002).Coordinated expression ofsc1eraxis and Sox9 genes during em-bryonic development of tendons andcartilage. J. Orthop. Res. 20, 827-833.
Assady, S., Maor, G., Amit, M. et al.(2001). Insulin production by humanembryonie stern cells. Diabetes 50,1691-1697.
Bain, G., Kitchens, D., Yao, M. et al.(1995). Embryonie stern cells expressneuronal properties in vitro. Dev. Biol.168,342-357.
Bigot, K., de Lange, J., Archer, G. et al.(1999). The relative semi-quantitativeexpression as useful toxicological end-point for identification of embryotox-ic-teratogenic substances. Toxicol. 1nVitro 13, 619-623.
Boheler, K. R., Czyz, J., Tweedie, D. etal. (2002). Differentiation of pluripo-tent embryonie stern cells into car-diomyocytes. Cire. Res. 91,189-201.
Bremer, S. and Hartung, T. (2004). Theuse of embryonie stern cells for theregulatory developmental toxicity test-ing in vitro - The current status of testdevelopment. Curr. Pharm. Des. 10,2733-2747.
Bremer, S., Worth, A. P., Paparella, M. etal. (2001). Establishment of an in vitroreporter gene assay for developmentalcardiac toxicity. Toxicol. 1n Vitro 15,215-223.
Brent, R. L. and Beckman, D. A. (1990).Environmental teratogens. Bull. N YAcad. Med. 66, 123-163.
Brown, N. A., Spielmann, H., Bechter, R.et al. (1995). Screening Chernicals for
ALTEX 22, 2/05

~ PELLIZZER ET AL.--~--------------------------
Reproductive Toxicity: The CurrentAlternatives. ATIA 23,868-882.
Brown, N. A. (1987). Teratogenicitytesting in vitro: status of validationstudies. Areh. Toxieol. Suppl. 11, 105-114.
Callan, N. A. and Witter, F. R. (1990).Intrauterine growth retardation: char-acteristics, risk factors and gestationalage. Int. J. Gynaeeol. Obstet. 33, 215-220.
Chai, C. K. and Degenhardt, K. H.(1962). Developmental anomaties ininbred rabbits. J. Hered. 53,174-182.
Chang, M. C. (1944). Artificial produc-tion of monstrosities in the rabbit.Nature 154, 150.
Cunliffe, V. and Smith, 1. C. (1992).Ectopic mesoderm formation in Xeno-pus embryos caused by widespreadexpression of a Brachyury homologue.Nature 358, 427-430.
Davis, D. L., Friedler, G., Mattison, D.and Morris, R. (1992). Male-mediatedteratogenesis and other reproductiveeffects: biologic and epidemiologicfindings and a plea for clinical re-search. Reprod. Toxicol. 6,289-292.
Desbois, C. and Karsenty, G. (1995).Osteocalcin cluster: implications forfunctional studies. 1. Cello Bioehern.57, 379-383.
Ducy, P., Zhang, R., Geoffroy, V. et al.(1997). Osf2/Cbfal: a transcriptionalactivator of osteoblast differentiation.Cell 89, 747-754.
Eiges, R. and Benvenisty, N. (2002). Amolecular view on pluripotent sterncells. FEBS Lett. 529, 135-141.
Fassler, R., Rohwedel, J., Maltsev, V. etal. (1996). Differentiation and integrityof cardiac muscle cells are impaired inthe absence of beta 1 integrin. J. CelloSei. 109,2989-2999.
Faustman, E. M. (1988). Short-term testsfor teratogens. Mutat. Res. 205, 355-384.
Flynn, R. 1. (1968). Exencephalia: its oc-currence in untreated mice. Seience160,898-899.
Green, H., Easley, K. and Iuchi, S.(2003). Marker succession during thedevelopment of keratinocytes fromcultured human embryonie stern cells.Proc. Natl. Acad. Sei. USA. 100,15625-15630.
ALTEX 22, 2/05
Gruenwald, P. (1966). Growth of thehuman fetus. I. Normal growth and itsvariation. Am. J. Obstet. Gynecol. 94,1112-1119.
Haas, J. F. and Schottenfeld, D. (1979).Risks to the offspring from parentaloccupational exposures. J. Occup.Med. 21, 607-6l3.
He, J. Q., Ma, Y., Lee, Y. et al. (2003).Human embryonie stern cells developinto multiple types of cardiacmy-ocytes: action potential characteriza-tion. Cire. Res. 93, 32-39.
Kalter, H. (1980). The relation betweencongenital malformation and prenatalmortality in experimental animals.In I. H. Porter and E. Hook (eds),Human Embryonie and Fetal Death(29-44). New York: Academic Press.
Kalter, H. (1968). Sporadic congenitalmalformations of newborn inbredmice. Teratology 1,193-199.
Karnofsky, D. A. (1965). Drugs asteratogens in animals and man. Annu.Rev. Pharmaeol. 10,447-472.
Kaufman, D. S., Hanson, E. T., Lewis, R.L. et al. (2001). Hematopoietic colony-forrning cells derived from human em-bryonic stern cells. Proc. Natl. Aead.Sei. USA. 98, 10716-10721.
Kehat, 1., Amit, M., Gepstein, A. et al.(2003). Development of cardiomy-ocytes from human ES cells. Meth. En-zymol. 365,461-473.
Kehat, 1., Gepstein, A., Spira, A. et al.(2002). High-resolution electrophysio-logical assessment ofhuman embryon-ic stern cell-derived cardiomyocytes: anovel in vitro model for the study ofconduction. Cire. Res. 91, 659-661..
Khera, K. S. (1985). Maternal toxicity:a possible etiological factor in embryo-fetal deaths and fetal malformations ofrodent-rabbit species. Teratology 31,129-153.
Khera, K. S. (1987). Maternal toxicity inhumans and animals: effects on fetaldevelopment and criteria for detection.Teratog. Careinog. Mutagen. 7, 287-295.
Kimmei, G. L., Smith, K., Kochhar, D.M. and Pratt, R. M. (1982). Overviewof In Vitro Teratogenicity Testing: as-pects of validation and application toscreening. Teratog. Careinog. Muta-gen. 2,221-229.
Kolossov, E., Fleischmann, B. K., Liu, Q.et al. (1998). Functional characteristicsof ES cell-derived cardiac precursorcells identified by tissue-specific ex-pression of the green fiuorescent pro-tein. J. CelloBiol. 143,2045-2056.
Kramer, 1., Hegert, c., Guan, K. et al.(2000). Embryonie stern cell-derivedchondrogenic differentiation in vitro:activation by BMP-2 and BMP-4.Meeh. Dev. 92, 193-205.
Leahy, A., Xiong, 1. w., Kuhnert, F. andStuhlmann, H. (1999). Use of de-velopmental marker genes to definetemporal and spatial patterns ofdifferentiation during embryoid bodyformation. J. Exp. Zool. 284,67-81.
Levenberg, S., Golub, J. S., Amit, M. etal. (2002). Endothelial cells derivedfrom human embryonie stern cells.Proc. Natl. Acad. Sci. USA 99, 4391-4396.
Leonard, B. E. (1981). Effect of psy-chotropic drugs administered topregnant rats on the behaviour of theoffspring. Neuropharmacology 20,1237-1242.
Lints, T. J., Parsons, L. M., Hartley, L. etal. (1993). Nkx-2.5: a novel murinehomeobox gene expressed in earlyheart progenitor cells and theirmyogenic descendants. Development119,419-431.
Lockhart, D. J. and Winzeler, E. A.(2000). Genomics, gene expressionand DNA arrays. Nature 405,827-836.
Low, 1. A. and Galbraith, R. S. (1974).Pregnancy characteristics of intrauter-ine growth retardation. Obstet. Gy-neeol. 44, 122-126.
Lubchenco, L. 0., Handman, C.,Dressler, M. and Boyd, E. (1963).Intaruterine growth as estimatedfrom liveborn birth-weight data at 24to 42 weeks of gestation. Pediatries32, 793-800.
Lutwak-Mann, C. (1964). Observation ofprogeny of thalidomide-treated malerabbits. Br. Med. 1. 1, 1090-1091.
MacLeod, P. M. (1993). Implication ofthe child with malformations for thefamily and society. In R. E Stevenson,1. G HaI and R. M Goodman (eds),Human malformations and relatedanomalies (Vol. I, 187-196). OxfordMonographs on Medical Genetics No.
55

PELLIZZER ET AL. ~-------------~-
27, Oxford University Press.Maltsev, V. A., Rohwedel, J., Hescheler,J. and Wobus, A. M. (1993). Embryon-ic stern cells differentiate in vitrointo cardiomyocytes representing si-nusnodal, atrial and ventricular celltypes. Meeh. Dev. 44,41-50.
Meyersoif M., Counter, C. M., Eaton, E.N. et al. (1997). hEST2, the putativehuman telomerase catalytic subunitgene, is up-regulated in tumor cellsand during immortalization. Cell 90,785-795.
Menezes, J. R and Luskin, M. B. (1994).Expression of neuron-specific tubulindefines a novel population in theproliferative layers of the developingte1encephalon. J. Neurasei. 1, 5399-5416.
Metzger, J. M., Lin, W. I. and Samuel-son, L. C. (1996). Vital staining ofcardiac myocytes during embryoniestern cell cardiogenesis in vitro. Cire.Res. 78, 547-552.
Miller, H. C. (1981). Intrauterine growthretardation. An unrnet challenge. Am.J. Dis. Child. 135, 944-948.
Mummery, C., Ward-van Oostwaard, D.,Doevendans, P. et al. (2003). Differen-tiation of human embryonie stern cellsto cardiornyocytes: role of coculturewith visceral endoderm-like cells. Cir-eulation 107, 2733-2740.
Nelson, B. K., Brightwell, W. S., Setzer,J. V. et al. (1981). Ethoxyethanol be-havioral teratology in rats. Neuratoxi-eology 2,231-249.
Okamoto, M., Kihara, T. and Tanimura,T. (1986). Developmental toxicity ofprenatal aspirin exposure to rats. Tera-tology 34, 451.
Pandey, A. and Mann, M. (2000). Pro-teornics to study genes and genomes.Nature 405,837-846.
Paparella, M., Kolossov, E., Fleis-chmann, B. K. et aL (2002). The use ofquantitative image analysis in the as-sessment of in vitro embryotoxicityendpoints based on a novel embryoniestern cell clone with endoderm-relatedGFP expression. Toxieol. In Vitro 16,589-597.
Pellizzer, c., Adler, S., Corvi, R. et al.(2004a). Monitoring of teratogeniceffects in vitro by analysing a selectedgene expression pattern. Toxieol. In
56
Vitra 18, 325-335.Pellizzer, C., Bello, E., Adler, S. et al.(2004b). Detection of tissue specificeffects by Methotrexate on differenti-ating mouse embryonie stern cells.Birth Defeets Res. B. Dev. Reprad.Toxieol. 71,331-341.
Reubinoff, B. E., Pera, M. F., Fong, C. Y.et al. (2000). Embryonie stern celllines frorn human blastocysts: sornaticdifferentiation in vitro. Nat. Bioteeh-nol. 18,399-404.
Rohwedel, J., Guan, K., Zuschratter, W.et al. (1998). Loss of beta1 integrinfunction results in a retardation ofrnyogenic, but an acceleration ofneuronal, differentiation of ernbryonicstern cells in vitro. Dev. Biol. 201, 167-184.
Rosano, A., Botto, L. D., Botting, B. andMastroiacovo, P. (2000). Infant mortal-ity and congenital anomalies from1950 to 1994: an international perspec-tive. J. Epidemiol. Community. Health54, 660-666.
Schardein, J. L. and Keller, K. A. (1989).Potential human developrnental toxi-cants and the role of animal testing intheir identification and characteriza-tion. Crit. Rev. Toxieol. 19,251-339.
Schardein, J. L. (1983). Teartogenic riskassessrnent. Past, present and future. InH. Kalter (ed), Issues and Reviews inTeratology (Vol. 1, 181-214). NewYork: Plenum.
Schmidt, M. M., Guan, K. and Wobus, A.M. (2001). Lithium influences differ-entiation and tissue-specific geneexpression of mouse ernbryonic stern(ES) cells in vitro. Int. J. Dev. Biol. 45,421-429.
Schuldiner, M., Yanuka, 0., Itskovitz-El-dor, J. et al. (2000). Effects of eightgrowth factors on the differentiation ofcells derived from human ernbryonicstern cells. Proe. Natl. Aead. Sei. USA97, 11307-11312.
Scholer, H. R. (1991). Octamania: thePOU factors in murine developrnent.Trends in Genet. 7, 323-329.
Seiler, A., Visan, A., Buesen, R. et al.(2004). Irnprovernent of an in vitrostern cell assay for developmental tox-icity: the use of molecular endpoints inthe ernbryonic stern cell test. Reprod.Toxieol. 18,231-240.
Seiler, A., Visan, A., Pohl, I. et al. (2002).Irnproving the embryonie stern celltest (EST) by establishing molecularendpoints of tissue specific develop-ment using rnurine ernbryonic sterncells (D3 cells). ALTEX 19 Suppl. 1,55-63.
Shepard, T. H. and Fantel, A. G. (1979).Embryonie and early fetal loss. Clin.Perinatol. 6,219-243.
Singec, I., Knoth, R., Ditter, M. et al.(2002). Synaptic vesicle pro teinsynaptoporin is differently expressedby subpopulations of mouse hip-pocampal neurons. J. Comp. Neurol.452,139-153.
Spielmann, H., Poh, I., Doering, B. et al.(1997). The Embryonie Stern CellsTest (EST), an in vitro ernbryotoxicitytest using two permanent cells line:3T3 fibrob1asts and ernbryonic sterncells. Toxieol. In Vitra 10, 119-127.
Staples, R. E. and Holtkamp, D. E.(1963). Effects of parental Thalido-rnide treatment on gestation and fetaldeve1opment. Exp. Mol. Pathol. 26,81-106.
Strubing, c., Ahnert-Hi1ger, G., Shan, J.et aL (1995). Differentiation ofpluripotent ernbryonic stern cells intothe neuronallineage in vitro gives riseto mature inhibitory and excitatoryneurons. Meeh. Dev. 53,275-287.
Suarez-Rodriguez, R. and Belkind-Gerson, J. (2004). Cultured nestin-positive cells frorn postnatal mousesmall bowe1 differentiate ex vivo intoneurons, glia, and srnooth muscle.Stem Cells. 22, 1373-1385.
Thomson, J. A., Itskovitz-Eldor, J.,Shapiro, S. S. et al. (1998). Embryoniestern cell lines derived frorn humanblastocysts. Seienee 282,1145-1147.
van der Jagt, K., Munn, S., Terslev, J.and de Bruijn, J. (2004). Alternativeapproaches can reduce the use of testanimals under REACH. http://ihcp.jrc.cec.eu.intIDOCUMENTATION/ECBI20041 Reducing jhe jise ofjesr ani-mals_under_REACH_IHCP _report.pdf(access 0112005).
Vorhees, C. v.. Butcher, R. E., Brunner,R. L. and Sobotka, T. J. (1979). Adevelopmental test battery for neu-robehavioral toxicity in rats: a pre-liminary analysis using monosodium
ALTEX 22, 2/05

~ PELLIZZER ET AL.-~-------------
glutamate calcium carrageenan, andhydroxyurea. Toxicol. Appl. Pharma-col. 50,267-282.
Warkany, J. (1971). Syndromes. Am. J.Dis. Child. 121, 365-370.
Wiche, G., Herrmann, H., Dalton, J. M.et al. (1986). Mo1ecu1ar aspects ofMAP-1 and 1.V1AP-2:miereheterogene-ity, in vitro localization and distribu-tion in neuronal and nonneuronal cells.Ann. N. Y. Acad. Sei. 466, 180-198.
Wilson, J. G. (1978). Review of in vitrosystems with potential for use interatogenicity screening. J. Environ.Pathol. Toxicol. 2, 149-167.
Wi1son, J. G. (1977). Embryotoxicity ofdrugs in man. In Plenum PublishingCorp (ed), Handbook of teratology,(Vo111., 309-355). New York.
Wi1son, J. G. (1975). Reproduction andteratogenesis: current methods andsuggested improvements. J. Assoc. Off.Anal. Chem. 584,657-667.
Wilson, J. G. (1973). Environment andbirth defects. New York: AcademicPress.
Wobus, A. M., Kaomei, G., Shan, J. et al.(1997). Retinoic acid acce1eratesembryonie stern cell-derived cardiacdifferentiation and enhances develop-ment of ventricular cardiomyocytes.J. Mol. Cell. Cardiol. 29, 1525-1539.
Xiao, Y., Haase, H., Young, W. G. andBartold ,P. M. (2004). Developmentand transplantation of a mineralizedmatrix forrned by osteoblasts in vitrofor bone regeneration. Cell Transplant.13,15-25.
Xu, C., Police, S., Rao, N. and Carpenter,M. K. (2002). Characterization andenrichment of cardiomyocytes derivedfrom human embryonie stern cells.Circ. Res. 91,501-508.
Yoon, P. W., Olney, R. S., Khoury, M. J.et al. (1997). Contribution of birthdefects and genetic diseases to pedi-atric hospitalizations. A population-based study. Arch. Pediatr. Adolesc.Med. 151, 1096-1103.
Zhang, S. C., Wernig, M., Duncan, I. D.et al. (2001). In vitro differentiation oftransplantab1e neural precursors from
human embryonie stern cells. Nat.Biotechnol. 19, 1129-1133.
zur Nieden, N. 1., Kempka, G. and Ahr,H. J. (2003). In vitro differentiationof embryonie stern cells into mineral-ized osteoblasts. Differentiation 71,18-27.
zur Nieden, N. 1., Ruf, L. J., Kempka, G.et al. (2001). Molecular markers inembryonie stern cells. Toxicol. In Vitro15,455-461.
Correspondence toProf. Dr. Dr. Thomas HartungECVAM (European Center for theValidation of Alternative Methods)IHCP (Institute for Health andConsumer Protection)Joint Research CentreVia E. Fermi n. 11-21020 Ispra (VA)Italyphone:+39-033278-5939fax: +39-033278-6297e-mail: [email protected]