der einfluß von stärke auf die aktivität der pyrophosphatase aus isolierten chloroplasten
TRANSCRIPT

Planta (Berl.) 120, 155--162 (1974) �9 by Springer-Verlag 1974
Der Einflu$ yon St/irke auf die Aktivit/it der Pyrophosphatase aus isolierten Chloroplasten*
Brig i t t e K l e m m e und Gfinter J acob i
Lehrstuhl f/ir Biochemie der Pflanze, Pflanzenphysiologisches Institut, Universit~t D-3400 GSttingen, Untere Karspiile 2, Federal Republic of Germany
Eingegangen am 9. Juli / 4. August 1974
The Inf luence of S ta rch on the A c t i v i t y of P y r o p h o s p h a t a s e f rom I so la ted Spinach Chloroplasts
Sum~nary. The activity of the inorganic pyrophosphatase from isolated spinach (Spinacia oleracea L.) chloroplasts is strongly dependent upon the addition of magnesium ions. Since the complex of the bivalent ion with inorganic pyrophosphate is the real substrate, a definite Mg2+/Na~PeOT-ratio is required for maximum activity. When the activity was measured in par- ticle-free extracts from chloroplasts, this ratio was shown to be approximately 3. However, an increase up to 10 was observed in the presence of thylakoid membranes. Furthermore, the kinetics in the presence of broken chloroplasts becomes sigmoidal.
The altered kinetics have been shown to be due to starch located in the thylakoids. The inhibitory effect is caused by amylose alone but not by amylopectin. Detailed kinetic analysis of the inhibition showed no influence of amylose on the Hill-coefficient. Since ethylenedia- minetetracetic acid was shown to have similar effects as amylose, starch might regulate the pyrophosphatase activity by binding Mg2+-ions.
Einleitung Das V o r k o m m e n yon P y r o p h o s p h a t a s e n (PPasen) in hSheren Pf lanzen wurde
wiederhol t durch Akt iv i t i t t smessungen in Homogena t en yon ganzen B1/fttern und auch in E x t r a k t e n von isol ier ten Chloroplas ten nachgewiesen (Naganna et al.,
1955; Wessels und Bal tscheffsky, 1960; La tzko uud Gibbs, 1968; K a r n und Moudr ianakis , 1969; S immons und But ler , 1969; Burke, 1970; E1-Badry und Bassham, 1970; Gould und Winget , 1973;). Ki i rz l ich gelang es ers tmalig, eine spezifisch in den Chloroptas ten lokal is ier te Py rophospha t a se yon einer , ,cytoplas- ma t i schen" zu t r ennen und die Exis tenz yon mindes tens zwei I soenzymen mi t unterschiedl ichen kinet ischen Charak te r i s t ika zu beweisen (Klemme uud Jacobi , 1974). Ffir die Ak t iv i t s is t bei beiden E n z y m e n der Zusatz yon Mg ~+- Ionen erforderl ich. Da aber fiir die Max ima lak t iv i t i i t der gere in ig ten Enzympri~- p a r a t e ein Verhs von zugesetz tem Mg z+ zu P y r o p h o s p h a t (PP) yon e twa 2 - - 3 zu 1 bes teht , is t vor a l lem eine Wechse lwi rkung des b iva len ten Ions mi t dem Sub- s t r a t gegeben.
Insbesondere durch Unte r suchungen mi t P y r o p h o s p h a t a s e n aus t ier ischen Geweben und aus Mikroorganismen wurde der Beweis e rbracht , dab das eigent-
* Abki~rzungen: EDTA = ~thylendiamintetraessigs~ure (Na-Salz) ; Pa ~ anorganisches Phos- phat; PP=Na4PeOT; PP-ase=alkalische Pyrophosphatase; Tris=Tris(hydroxymethyl) aminomcthan.
11 Planta (Berl.), Vol. 120

156 B. Klemme und G. Jacobi
liche Subs t r a t fiir die Py rophospha t a se der K o m p l e x zwischen P y r o p h o s p h a t und einem zweiwert igen Metal l ion ist (Naganna et al., 1955; Dixon and Webb , 1964; Josse, 1966; R a p o p o r t et al., 1972). Diese Ta t sache wird vor al lem ffir den Akt iv i - t i i t snachweis des E n z y m s in R o h c x t r a k t e n und in F r a k t i o n e n isol ier ter Organelle bedeutsam, in denen durch eine Komplex ie rung der Ionen durch andere Zell- bes tandte i le die appa ren te Mg2+-Konzentrat ion ve rminde r t wird. E in Beispiel hierfi ir wird in der vor l iegenden Arbe i t durch vergle ichende Akt iv i tg t smessungen mi~ isol ier ten Chloroplas ten und Chlorop las tenex t rak ten aufgezeigt und ein Beweis fiir die N a t u r der komplex ie renden K o m p o n e n t e erbracht .
Material und Methoden Die Untersuchungen wurden mit isolierten Chloroplasten aus Spinacia oleracea durch-
gefiihrt. Das Pflanzenmaterial wurde unter konstanten Bedingungen in einem halbklimati- sierten Raum bei 15 ~ C im Kurztag angezogen (Postius and Jacobi, 1971). Die Isolation der Chloroplasten erfolgte aus 4--6 Wochen alten Pflanzen nach Avron (1960). Ffir die Messungen warden die einmal gewaschenen Chloroplasten, die etwa 10--20% der Gesamteinheiten an PPase enthielten, direkt eingesetzt und mit der Aktivitg~t eines partikelfreien Chloroplasten- extraktes verglichen.
Fiir die Herstellung des partikelfreien Extraktes warden die gewaschenen Chloroplasten in einem hypotonischen Medium (0.05 Tris-ttC1 Puffer, Ph 8,2) aufgenommen und 10 rain beschallt (20 kHz). Der nach Zentrifugation (30 min, 40000• gewonnene klare ?Jberstand enthielt die gesamte PPase-Aktivit~t und wurde als ,,partikelfreier" Extrakt verwendet.
Die Aktivit~t der PPase wurde bei 37 ~ C in einem 1 ml Reaktionsgemisch mit der folgenden Zusammensetzung bestimmt: 60 mM Tris-HC1, pH 8,2; 1 mM PP; 0--20 mM MgCl~ und wenn nicht anders angegeben, 15 mg Chloroplastenprotein bzw. 3 mg Protein des partikelfreien Enzyms. Nach 3 rain Vorinkubation wurde die Reaktion durch Zugabe yon PP gestartet und nach 3 rain dureh Zugabe yon 0,5 ml HCIO 4 (10 % ) abgestoppt. In dem proteinfreien Uberstand wurde das aus PP freigesetzte Pa nach der Methode von Taussky und Shorr (1953) bestimmt.
Die nach der Methode yon Whelau (1955) aus Chloroplasten isolierte und gereinigte Amy- lose wurde in 0,8 :N HC1 bei 80 ~ C im Wasserbad hydrolysiert. Die Amylose-Konzentration zu Beginn der Hydrolyse betrug 2,5 mg/ml. :Nach 0--200 min Inkubationszeit wurde die Hydro- lyse durch Zugabe yon festem Na~CO 3 gestoppt und die freigesetzte Glucose photometrisch mit ttexokinase und Glucose-6-Phosphat-Dehydrogenase bestimmt (Bergmeyer, 1970). Ein Aliquot des jeweiligen Ansatzes wurde dem Reaktionsgemisch zur Bestimmung der PPase- Aktiviti~t zugestzt.
Die St~rkemenge wurde mit K J naeh der yon Whelau (1955) beschriebenen Methode bestimmt.
Protein wurde nach Lowry et al. (1951) mit t~inderserumalbumin als Standard gemessen.
Ergebnisse 1. Abhiingigkeit der PPase-Aktivitiit von der Mg2+-Konzentration
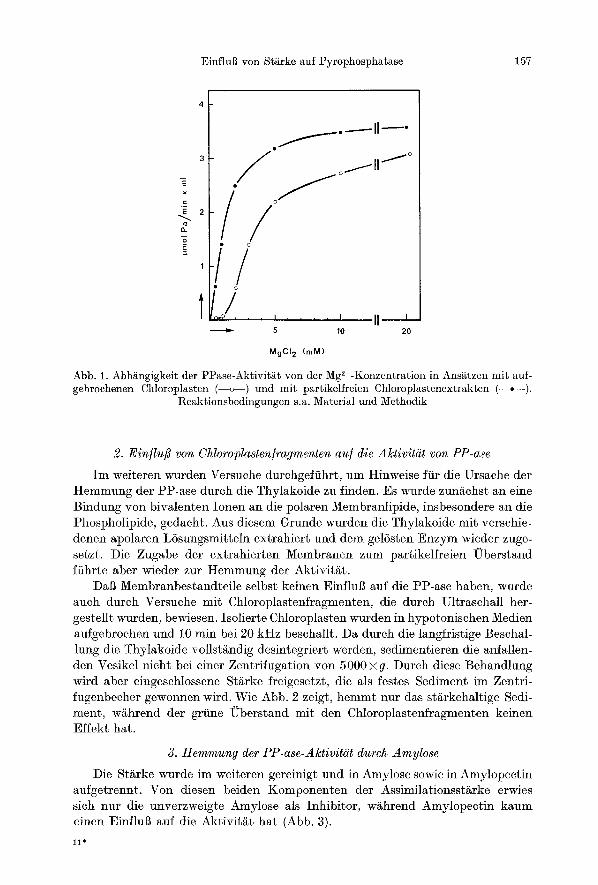
Die Abh~ngigke i t der PP-ase A k t i v i t ~ t yon der Mg2+-Konzentrat ion mi t Chloropl~sten und mi t dem par t ike l f re ien E x t r a k t ist in Abb. 1 dargestel l t . W~hrend die Max ima lak t iv i t~ t im E x t r a k t bei e inem 3 ~ 5 f a c h e n UberschuB an Mg 2+ re la t iv zu Pyrophosph~ t e rmi t t e l t wurde, ist das Verh~ltnis e twa 10:1, wenn im Ansatz n o c h T h y l a k o i d e en tha l t en sind. Bei der Messung mi t dem E x t r a k t folgt die Si~ttigung e inem hyperbo l i schen Verlauf im Sinne einer einfachen Michael is -Menten-Kinet ik , w~thrend un te r Zusatz yon Thylako iden eine sigmoide S/~ttigungskurve e rha l ten wird. Auf die kinet ische I n t e r p r e t a t i o n wird wel ter un ten eingegangen.
Einflul~ yon St~rke auf Pyrophosphatase 157
•
-6 E
, / o
./o oJ / j / / /
10 20 �9 , 5
MgCI 2 ( raM)
Abb. 1. Abh~ngigkeit der PP~se-Aktivit~Lt yon der Mg2+-Konzentration in Ans~tzen mit auf- gebrochenen Chloroplgsten (--o--) und mit partikelfreien Chloroplastenextrakten (--.--).
Regktionsbedingungen s.a. Material und Methodik
2. Ein/lufi yon Chloroplasten/ragmenten au[ die Aktivitdit von PP-ase
I m weiteren wurden Versuehe durchgeftihrt, um Hinweise ffir die Ursaehe der Hemmung der PP-ase durch die Thylakoide zu linden. Es wurde zun~Lchst an eine Bindung yon bivalenten Ionen an die polaren Membranlipide, insbesondere an die Phospholipide, gedacht. Aus diesem Grunde wurden die Thylakoide mit verschie- denen apolaren LSsungsmitteln extrahiert und dem gelSsten Enzym wieder zuge- setzt. Die Zugabe der extrahierten Membranen zum paI~ikelfreien Uberstand f~hrte aber wieder zur Hemmung der Aktivit~t.
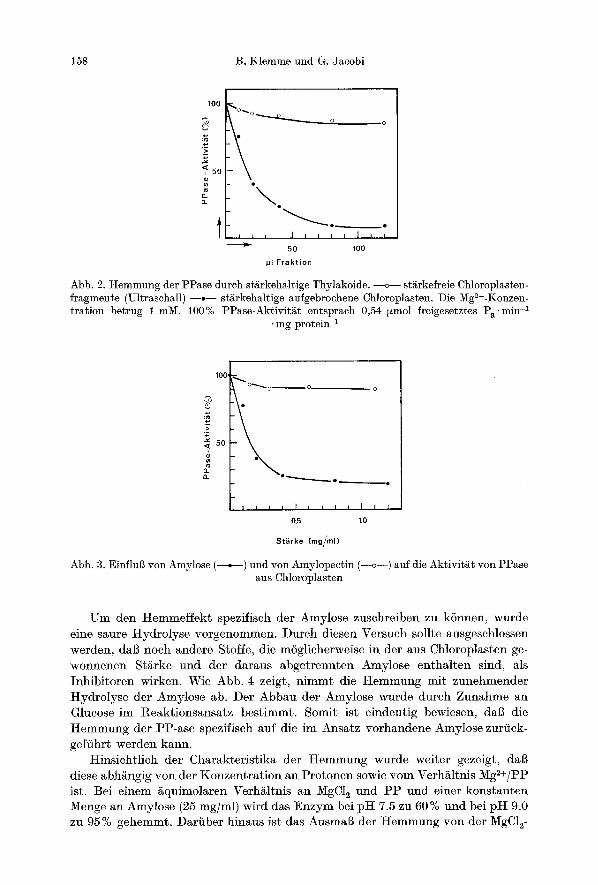
Dai~ Membranbestandteile selbst keinen Einflul3 auf die PP-ase haben, wurde auch dutch Versuche mit Chloroplastenfragmenten, die durch Ultrasehall her- gestellt wurden, bewiesen. Isolierte Chloroplasten wurden in hypotonisehen Medien aufgebrochen und 10 rain bei 20 kHz beschallt. Da dureh die langfristige Besehal- lung die Thylakoide vollst/tndig desintegriert werden, sedimentieren die anfallen- den Vesikel nicht bei einer Zentrifugation yon 5000 • g. Durch diese Behandlung wird aber eingeschlossene St~rke freigesetzt, die als festes Sediment im Zentri- fugenbeeher gewonnen wird. Wie Abb. 2 zeigt, hemmt nur das stSzkehaltige Sedi- ment, w~hrend der grfine Uberstand mit den Chloroplastenfragmenten keinen Effekt hat.
3. Hemmung der PP-ase-Alctivitiit dutch Amylose
Die St~rke wurde im weiteren gereinigt und in Amylose sowie in Amylopeetin aufgetrennt. Von diesen beiden Komioonenten der Assimilationssts erwies sich nur die unverzweigte Amylose als Inhibitor, ws Amylopeetin kaum einen EinfluI] auf die Aktivits hat (Abb. 3).
11"
158 B. Klemme und G. Jacobi
100
,_> ..~ ~, 5 o
'~ ~ ~ ~ o o
\ o
\ . , J ~ i I i i i i I i i
50 100
~JI F r a k t i o n
Abb. 2. Hemmung der PPase durch st/irkehaltige Thylakoide. --o-- st~rkefreie Chloroplasten- fragmente (Ultraschall) - - - - - st/~rkehaltige aufgebrochene Chloroplasten. Die Mg2+-Konzen - tration betrug 1 raM. 100% PPase-Aktivit~t entsprach 0,54 ~zmol freigesetztes Pa'min-1
�9 protei~ ~
' iO0
> .g,
5 0
a . 2__ ~ . _ _
I i , , I i i i T I I I
0.5 1.0
St~rke (mg /m l )
Abb. 3. Einflul~ yon Amylose (--.--) und yon Amylopectin (--o--) auf die Aktivit~t yon PPase aus Chloropl~sten
Um den Hemmeffekt spezifisch der Amylose zuschreiben zu kSnnen, wurde eine saure Hydrolyse vorgenommen. Durch diesen Versuch sollte ausgeschlossen werden, dab noch andere Stoffe, die mSglicherweise in der aus Chloroplasten ge- wonnenen St~trke und der daraus abgetrennten Amylose enthalten sind, als Inhibitoren wirken. Wit Abb. 4 zeigt, nimmt die Hemmung mit zunehmender Hydrolyse der Amylose ab. Der Abbau der Amylose wurde durch Zunahme an Glucose im Reaktionsansatz bestimmt. Somit ist eindeutig bewiesen, d~l] die Hemmung der PP-ase spezifisch auf die im Ansatz vorhandene Amylose zurack- gefiihrt werden kann.
Hinsichtlich der Charakteristika der Hemmung wurde welter gezeigt, dab diese abh~ngig yon der Konzentration an Protonen sowie vom Verh~ltnis Mg2+/PP ist. Bei einem ~quimolaren Verhgltnis an MgC12 und PP und einer konstanten Menge an Amylose (25 mg/ml) wird das Enzym bei pH 7.5 zu 60% und bei pH 9,0 zu 95 % gehemmt. Darfiber hinaus ist das AusmaB der Hemmung yon der MgCl~-

Einflug von Stiirke auf Pyrophosphatase 159
5 100
_ �9 ~ ~ �9
N 4 o ~
/ 50 E t 2 o E
5
:[ 0 ~ " I I I I I I , I I
100 200
Hydrolysezeit (rain)
Abb. 4. Beziehung zwischen Hydrolysegrad von Amylose (--*--) und Ausm~B der I-Iemmung ( ~ ) auf die Aktivitgt yon PPase aus Chloroplasten
0.3 . ~ "
5 10 15 20
MgCI 2 (mM)
Abb. 5. Si~ttigungskurven ffir Mg 'a+ mit partikelfreier PPase in Abh~ngigkeit yon der zuge- setzte n lVIenge an Amylose. Ohne Zusatz �9 ,0,10 mg Amylose/ml - - , - - , 0,25 mg Amylose/
ml - - . - -
K0nzentration im Reaktionsansatz abh~ngig. So betr~gt bei pH 8,2 die I-Iemmung 80% bei einem MgC12/PP-Verhi~ltnis yon 1, aber nur 20% bei einem Verh<nis yon 20.
4. Kinetisehe Analyse der Hemmung dutch Amylose Werden die Reaktionsraten (v) bei 1 mM PP als Funktion der Konzentration
yon Mg ~+ gemessen, so verstgrkt sich die Sigmoidit&t des Kurvenverlaufes bei Erh6hung der Menge an zugesetzter Amylose (Abb. 5). Die in Abb. 6 dargestellte doppelt reziproke Auftragung nach Lineweaver-Burk zeig~ weiterhin, d~6 die Naximalgeschwindigkeit aber nicht veriindert wird.
Der Sigmoide KurvenverlauI lfir die S&ttigung mit MgC12 lie6e sich ~uch dahin- gehend interpretieren, dMt Amylose ein negativer Effektor ffiir das Substrat am Enzym ist. Da sich die Steigung der Geraden yon n• = 1.25 in einem Hill-Dia-

160 B. Klemme und G. Jacobi
10
I I I I I i I 0,5
M g C I 2
Abb. 6. Lineweaver-Burk-Diagr~mm zu Abb. 5
gramm (log Mg 2+ gegen log [V(/Vm~x-V)] jedoch nieht ~ndert, kann eine Inter- aktion zwischen Amylose und Substrat am Enzym ausgesehlossen werden.
Als Alternative muB daher angenommen werden, dab Amylose Mg ~+ komple- xiert und somit die Konzentration des Substrates, das als Mg-PP-Komplex wirkt, vermindert. Werm jedoeh die Hemmung der Aktivit/~t yon PP-ase allein auf die Chelierung des Cosubstrates Mg 2+ dureh Amylose beruht, so mfiSte aueh jeder andere Chelator eine/~hnliehe Wirkung auf die Aktivit/~t haben. Als Modellver- such wurde daher die Wirkung yon EDTA getestet, das in gleicher Weise wie Amylose mit steigender Konzentration die Sigmoidit/~t der Ss mit Mg 2+ verst/irkt. Aneh bier wird die maximale Reaktionsrate nieht ver/indert.
Diskussion
Zu den wesentliehsten Leistungen der Pflanze z/~hlt die in den Chloroplasten lokalisierte Biosynthese yon St/~rke. Die energieabh~ngige Polymerisation wird dutch die Aktivierung yon Glucose-l-Phosphat mit ATP eingeleitet, wobei ein Molekfi ! Pyrophosphat freigesetzt wird, Die Funktion der Pyrophosphatase wird vor allem in Hinbliek auf eine Gleichgewiehtsverschiebung in Richtung der Bio- synthese durch Verminderung der Konzentration eines der Reaktionsprodukte diskutiert. Gleiehzeitig wird auch der endogene Spiegel an anorganischem Ortho- phosphat erh6ht, so dub fiber diese Reaktion aueh ein Substrat ffir die Photo- synthesephosphorylierung angelietert wird.
Es stellt sieh zwangsl/~ufig die Frage, ob ein derartig zentrales Enzym, das eine Mittlerrolle zwisehen Biosynthese und Energiestoffweehsel spielt, mSglieher- weise aueh einer metabolisehen Regulation unterliegt. Im Gegensatz zu einigen bakteriellen PP-asen (Klemme, 1972) als aueh der PP-ase aus S&ugetierleber (Horn, et al., 1971) unterliegen jedoeh die bisher untersuchten PP-asen aus h6heren Pflanzen keiner Mlosterisehen Regulation. Eine mSgliche Beeinflussung der Akti- vit/~t dureh endogene Anderungen der Mg~+-Konzentration wurden abet bereits yon Bueke (1970) und EIBadry und Bassham (1970) diskutiert. Da beliehtete

EinfluB yon Sti~rke auf Pyrophosphatase 161
Thy lako ide ak t iv K a t i o n e n anfnehmen (Dilley und Vernon, 1965 ; Bassam et al., 1968), kann es zn einer Verminderung der Mg2+-Konzenfrat ion an der Oberfl/~che der Membranen kommen, was sieh sekund/~r auf die PP-ase auswirkt .
Mit der in dieser Arbe i t nachgewiesenen H e m m u n g der PP-ase du tch Amylose erg ib t sich ein wei terer Fak to r , der m6glicherweise anch in vivo eine l~olle spielen kann. Wie die Ergebnisse zeigen, fungier t die zwischen den T h y l a k o i d m e m b r a n e n l iegende St/~rke als Komplexb i ldne r fiir Mg 2+ und d a m i t ind i rek t Ms Hemmsto f f f/Jr die Pyrophospha tase . Es ka im somit spekul ier t werden, dab dureh eine lokale Vera rmung an Mg 2+ die Biosynthese von Sti~rke zu einem selbst regul ierenden Sys tem wird.
Mit Unterstfitzung der Deutschen Forschungsgemeinschaft. Ffir die technische Assistenz danken wir Frl. Marion Elbrecht.
Literatur
Avron, M.: Photophosphorylation by swiss-chard chloroplasts. Biochim. biophys. Acta (Amst.) 40, 257--272 (1960)
Bassham, J .A. , Kirk, M.: In Shibata, K., Takamiya, A., Jagendorf, A.T., Puller, 1%. C., edit.: Comparative biochemistry and biophysics of photosynthesis, p. 365. Tokye: Uni- versity of Tokyo Press 1968
Bergmeyer, U. : Methoden der enzymatischen Analyse, Bd. II , S. 1163. Weinheim: Verlag Chemie 1970
Bucke, C. : The distribution and properties of alkaline inorganic pyrophosphatase from higher plants. Phytochem. 9, 1303--1309 (1970)
Dilley, 1%. A., Vernon, L. P. : Ion and water transport processes related to the light dependent shrinkage of spinach chloroplasts. Arch. Biochem. Biophys. 111, 365--375 (1965)
Dixon, 3/[., Webb, E. C. : The enzymes. S. 75. New York: Academic Press 1964 El-Badry, A. M., Bassham, J. A.: Chloroplast inorganic pyrophosphatase. Biochim. biophys.
Acta (Amst.) 197, 308--316 (1970) Gould, J .M., Winger, G.D.: A membrane-bound alkaline inorganic pyrophosphatase in
isolated spinach chloroplasts. Arch. Biochem. Biophys. 154, 606--613 (1973) Horn, A., B6rnig, I-I., Thiele, G. : Allosteric properties of the Mg++-dependent inorganic pyro-
phosphatase in mouse liver cytoplasm. Europ. J. Biochem. 2, 243--249 (1967) Josse, J. : Constitutive inorganic pyrophosphatase of Escherichia coll. I I Nature and binding
of active substrate and the role of magnesium. J. biol. Chem. 241, 1948--1957 (1966) Karu, A.E. , Moudrianakis, E .N. : Fractionation and comparative studies of enzymes in
aqueous extracts of spinach chloroplasts. Arch. Biochem. Biophys. 129, 655--671 (1969) Klemme, B., Jacobi, G. : Separation and characterization of two inorganic pyrophosphatases
from spinach leaves. Planta (Berl.) 120, 147-153 (1974) Klemme, J.-tt . : Allosterisch regulierte und nicht-regulierte Pyrophosphatasen aus photo-
trophen Bakterien. I-Iabilitafionsschrift, Universiti/t G5tfingen (1972) Latzko, E., Gibbs, M. : Distribution and activity of enzymes of the reductive pentose phosphate
cycle in spinach leaves and in chloroplasts isolated by different methods. Z. Pflanzen- physiol. 59, 184--194 (1968)
Lowry, O. It., l~osebrough, N. J., Parr, A. L., Randall, R. J. : Protein measurement with the folin phenol reagent. J. biol. Chem. 193, 265--175 (1951)
Postius, S., Jacobi, G. : Dark starvation and chloroplast function. Planta (Bert.) 99, 222--229 (1971)
Naganna, B., Venugopol, B., Sripatbi, C.E.: Occurrence of alkaline pyrophosphatase in vegetable tissues. Biochem. J. 60, 224--225 (1955)
Rapoport, T. A., I-IShne, W. E., I~eich, J. G., Heitmann, P., Rapoport, S. M. : A kinetic model for the action of the inorganic pyrophosphatase from baker's yeast. The activating influence of magnesium ions. Europ. J. Biochem. 26, 237--246 (1972)

162 B. Klemme und G. Jacobi
Simmons, S., Butler, L. G. : Alkaline inorganic pyrophosphatase of maize leaves. Biochim. biophys. Acta (Amst.) 172, 150--157 (1969)
Taussky, H.H. , Shorr, E.: A mieroeolorimetric method for the determination of inorganic phosphorus. J. biol. :Chem, 202, 675--685 (1953)
Wessels, J. S. C., Baltscheffsky, H. : Adenosine triphosphatase activity in chloroplasts. Acta chem. scand. 14, 233--246 (1960)
Whelan, W. J. : Starch, glycogen ffuctosanes and similar polysaccharides. In: K. Peach, M. V. Tracey, Moderne Methoden dcr Pflanzenanalyse, Bd. II , S. 145ff. Berlin-Heidelberg- New York: Springer 1955