heterologous expression of plant vacuolar pyrophosphatase in
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 91, pp. 6128-6132, June 1994Plant Biology
Heterologous expression of plant vacuolar pyrophosphatase in yeastdemonstrates sufficiency of the substrate-binding subunit forproton transport
(bopsw iwa/proton pump/Sacckrmycs cenrea/manbraeW pobte)
EUGENE J. KIM, Rui-GUANG ZHEN, AND PHILIP A. REA*Plant Science Institute, Department of Biology, University of Pennsylvania, Philadelphia, PA 19104
Communicated by Emanuel Epstein, March 3, 1994 (received for review November 28, 1993)
ABSTRACT Themembre bou the vaclofplantceils onta two ee proton pu . These are thevacuolar H+-ATPase (EC 3.6.1.3), an ezme mo to aleukaryotes, and a vacuolar -transocatingp yu(EC3.6.1.1), whichIsuupltsbtotherwIkownin only a few ea. Alto the s ate-bin g subunit of the vacolar H+-pyrophosphatase has beenIdentified and pi and cDNAs encoding It have beenIsolated and r ied, the miml uit coPeent inpyrophosphate PPI)energed H tra on Is not known.Here we address this qu and show that heterologousexpresDson of the cDNA (AVP) e ing the b i gsubunit of the vacuoar H-pyrophosphatase from the vascu-lar plant Arbidepa i the yeast Saccharoeaycn cere-viu results in the roducion of vu y lid func-tonal enzyme active in PPI-dependent H+ transocatln. Sincethe heterlogously pmp is to I I omthe native plant w respect to PPI hyrysis, Htransocaton, activation by p s , and selcie inhbtionby calum and 1,1-dposphonates, it is concluded that all ofthe known catalytic funcions of the enzyme map to the onesubunit encoded by AVP.
The vacuole of plant cells is unusual in possessing twoelectrogenic HW-translocating phosphohydrolases. In addi-tion to the vacuolar-type H+-ATPase (V-ATPase; EC3.6.1.3), common to the endomembranes of animal, fungal,and plant cells (1, 2), the vacuolar membrane of plant cellscontains a HW-translocating pyrophosphatase (V-PPase; EC3.6.1.1), which exclusively utilizes pyrophosphate (PPi) asenergy source (3, 4). The V-PPase, like the V-ATPase, isabundant and ubiquitous in the plant vacuolar membrane andcontributes substantially to the establishment of the trans-tonoplast H+ electrochemical potential difference (2, 4) re-sponsible for the energization ofa broad range ofH+-coupledand electrically coupled transport processes.
Progress toward understanding the structure and function ofthe V-PPase, which belongs to a new category of ion trans-locases (3), has been rapid, but the minimal unit competent inPPi-dependent H+ translocation is still not known. While it isclear that the intrinsic, Mr 64,500-66,800, substrate-bindingsubunit catalyzes K+-activated PPi hydrolysis (4), it remainsto be determined if this polypeptide, alone, is sufficient forPPi-energized transmembrane H+ transport. On the one hand,it has been argued that because cDNAs encoding the sub-strate-binding subunit of the V-PPase ftom Arabidopsisthaliana (5) and Beta vulgaris (Y. Kim, E.J.K., and P.A.R.,unpublished results), designated AVP and BVP, respectively,specify a transmembrane polypeptide satisfying the minimumstructural requirements of a PPi-dependent H+ translocase-
direct interaction with substrate and continuity across thephospholipid bilayer-additional subunits need not be impli-cated (3). On the other hand, direct tests ofthis proposal usingconventional biochemical procedures, such as reconstitutionofpurifiedpreparations orradiation inactivation oftheenzymein isolated vacuolar membrane vesicles, have yielded discrep-ant estimates of functional size and variable subunit compo-sitions (4). It was in recognition of the difficulty of reachingfirm conclusions by traditional biochemical methods that theindependent strategy described here of heterologously ex-pressing cDNAs encoding the substrate-binding subunit andtesting for PPi-dependent H+ translocation was adopted. If,and only if, the substrate-binding subunit is the sole polypep-tide of the transport-competent pump will P~i-energized H+pumping be generated de novo when cDNAs encoding thissubunit are expressed.The yeast Saccharomyces cerevisiae was chosen for het-
erologous expression of the V-PPase for several reasons: (i)It is amenable to rigorous molecular genetic mniulation. (ii)Several finctional eukaryotic membrane proteins have al-ready been expressed in this organism (or Schizosaccharo-myces pombe). (iii) It is vacuolate and the vacuolar mem-brane in yeast is probably the default destination for heter-ologously expressed membrane proteins (6-8). (iv) It lacks anendogenous V-PPase. Though active in V-ATPase-catalyzedH+ translocation, vacuolar membrane vesicles isolated fromS. cerevisiae neither mediate PPi-dependent H+ tranloca-tion nor contain polypeptides with immunological cross-reactivity toward the V-PPase from plants (3). Thus, Sac-charomyces represents a highly manipulable null backgroundbiased in favor of the vacuolar localization of heterologouslyexpressed V-PPase. Two major experimental objectivesmight therefore be fulfilled using this system: (i) a direct testof the sufficiency of the substrate-binding subunit of theV-PPase for PPi-dependent H+ translocation, and (ii) theprovision of a molecularly manipulable system for site-directed mutagenesis of the transport-competent enzyme forelucidation of the structural requirements of PPi-energizedtransmembrane H+ translocation.
Here, we report expression of the cDNA (AVP) encodingthe substrate-binding subunit of the V-PPase from the vas-cular plant A. thaliana in the yeast S. cerevisiae to yieldvacuolar membrane-localized enzyme active in K+-stimulated, PPi-energized H+ translocation. In so doing, wedemonstrate the sufficiency of the polypeptide encoded byAVP for both scalar and vectorial V-PPase function.
MATERIALS AND METHODSMaterials. S. cerevisiae strain AACY1 and aminomethyl-
enediphosphonate were kindly provided by T. H. Stevens
Abbreviations: V-ATPase, vacuolar HW-translocating ATPase; V-PPase, vacuolar HW-translocating pyrophosphatase.*To whom reprint requests should be addressed.
6128
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 91 (1994) 6129
(University of Oregon, Eugene) and A. A. Baykov (MoscowState University, Russian Federation), respectively. SolublePPase, purified from S. cerevisiae, was purchased fromSigma.
Constructin of Expression Vector. The entire readingframe ofthe cDNA encoding the substrate-binding subunit ofthe V-PPase from A. thaliana (AVP; ref. 5) was amplified byPCR using Vent polymerase (New England Biolabs) accord-ing to the manufacturer's recommendations. The PCR prim-ers (corresponding to positions 106-122 and 2403-2418 in thesequence of AVP; ref. 5) were engineered to contain SnaBIand Xba I restriction sites at their 5' termini for directionalcloning purposes, and the resulting 2.3-kb, PCR-amplifledfragment was ligated into the multiple cloning site of PvuH-Xba I, double-digested pYES2 expression vector (Invi-trogen) to generate a construct containing AVP insertedbetween the yeast GAL] promoter and CYCI terminationsequences of the plasmid (see Fig. 1). Most of the PCR-amplified AVP insert was then replaced by the original cDNAto mie sequence fidelity and yield a construct in whichthe original cDNA, lacking only the 5' untranslated sequence,was inserted directionally. The sequence of the segment ofAVP that was not replaced by the original cDNA (25 bp) wasdetermined by the dideoxynucleotide chain terminationmethod (9) using custom-synthesized oligonucleotide prim-ers and found to match exactly the sequence of the originalcDNA clone (5).
Yeast Transformation. Ura- strains of S. cerevisiae weretransformed with the final construct, designated pYES2-AVP, by the LiOAc/PEG method (11). The transformantswere selected for uracil prototrophy by plating on AHCmedium [0.17% (wt/vol) yeast nitrogen base without aminoacids/0.5% (wt/vol) ammonium sulfate/1% (wt/vol) acid-hydrolyzed casein/0.002% (wt/vol) adenine hemisulfate/2%(wt/vol) glucose/50 mM Mes-Tris, pH 5.5]. The resultingUra+ colonies were picked and grown for 2 days in AHCmedium containing 2% glucose ("glucose-grown cells") or2% (wt/vol) galactose supplemented with 0.2% (wt/vol)fructose ("galactose-grown cells"). Fructose, rather thanglucose, was routinely employed as supplementary carbonsource for galactose induction because preliminary experi-ments had shown that the same concentration of glucose(0.2%) diminishes cellular V-PPase polypeptide content ver-sus fructose-supplemented controls.
Preparation of Whole-Celi Lysates. For the initial screeningof V-PPase expression, crude whole-cell lysates were pre-pared from galactose-grown or glucose-grown pYES2-AVP-transformed cells and galactose-grown untransformed cells.Cells were grown in 10-ml volumes of AHC medium, col-lected by centrifugation, and lysed by the method of Kane etal. (12) except that 1 mM phenylmethanesulfonyl fluoridewas added to the lysis buffer [8 M urea/5% (wt/vol) SDS/4%(vol/vol) 2-mercaptoethanol/50 mM Tris HCl, pH 6.8] im-mediately before use. Aliquots of the lysates were subjectedto SDS/PAGE, electroblotted to nitrocellulose, and probedwith antibody raised against keyhole limpet hemocyanin-conjugated V-PPase synthetic peptide.
Preparation of Microsomes and Vacuolar Membrane Vesi-cles. Vacuolar membrane vesicles were isolated from etio-lated hypocotyls of Vigna radiata (mung bean) as describedby Rea and Turner (13). For the routine preparation of yeastmicrosomes enriched for the V-PPase, cell cultures weregrown by diluting 200 ml of stationary phase cells into 2 litersof minimal medium supplemented with the appropriate car-bon source. The cells were grown for 24 h at 30°C to an OD600of %0.8 and collected by centrifugation. After washing withdistilled water, the cells were pelleted, resuspended in 10mMdithiothreitol/100 mM Tris-HCI, pH 9.4, and incubated for 20min at 300C with gentle shaking. The cells were pelleted,resuspended in 50 ml of YP medium [1% (wt/vol) yeast
extract/2% (wt/vol) Bacto Peptone] containing 0.7 M sorbi-tol, 1% glucose or 1% galactose, and 100 mM Tris-HCl (pH7.5), and converted to spheroplasts by the addition of 75 mgZymolyase 20T (ICN). The suspension was incubated for 60min at 300C with gentle shaking, centrifuged through acushion of 1.4 M sorbitol, and resuspended in 15-20 cellvolumes of ice-cold homogenization medium [0.15 M KCI/10% (wt/vol) glycerol, 1.5% (wt/vol) polyvinylpyrrolidone/5mM EGTA/50 mM Tris-ascorbate, pH 7.5] containing 1 mMphenylmethanesulfonyl fluoride and leupeptin at 1 pg/ml.After lysis by homogenization in a 40-ml Dounce homoge-nizer, the crude lysate was cleared of cell debris and unbro-ken cells by centrifugation at 650 x g for 5 min, homogenizedagain in the same medium, and recentrifuged. The superna-tants from both low-speed centrifugations were pooled andcentrifuged at 100,000 x g for 35 min. The pellet ("crudemicrosomes") was resuspended in suspension medium (1.1M glycerol/2 mM dithiothreitol/1 mM EGTA/5 mM Tris-Mes, pH 7.6), layered onto a discontinuous sucrose densitygradient consisting of 10%6 (wt/wt) and 30% sucrose andcentrifuged at 100,000 x gfor2 h. Partially purified, V-PPase-enriched microsomes were withdrawn from the 10/30% in-terface, diluted more than 10-fold with suspension medium,and pelleted by centrifugation at 100,000 x g for 35 min. Theresulting membrane pellet was washed one more time withsuspension medium, resuspended in suspension medium, andused immediately or stored at -85°C. Inclusion of KCl (0.15M) in the homogenization medium was found to minimizecontamination of the final microsome preparation with en-dogenous soluble PPase.
Intact yeast vacuoles were isolated by flotation centrifu-gation on Ficoll step gradients as described by Roberts et al.(10). To diminish proteolysis during membrane isolation, thisprocedure was shortened for some of the experiments bysubjecting the spheroplast homogenate to only one cycle offlotation centrifugation. The spheroplast homogenate wasoverlaid with buffer B [10 mM Mes-Tris, pH 6.9/0.5 mMMgCl2/8% (wt/vol) Ficoll 400; ref. 10] containing 1 mMEGTA and centrifuged at 52,000 x g for 30 min. Aftercentrifugation, the white layer at the top of the tube, whichwas enriched for vacuoles, was vesiculated for the transportassays or prepared directly for electrophoresis.Northern Analyses. Total RNA was isolated from galac-
tose-grown untransformed cells, galactose-grown trans-formed cells, and glucose-grown transformed cells as de-scribed by Zitomer et al. (14) with extraction buffer contain-ing 10 mM Tris HCl (pH 8.0), 0.1 M LiCl, and 1 mM EDTA.Contaminating DNA was removed by the addition ofRNase-free DNase, after which SDS was added to a final concen-tration of0.5% before deproteinization oftheRNA by phenolextraction. The samples were size-fractionated by electro-phoresis on 1.2% (wt/vol) agarose gels containing 0.63 Mformaldehyde and blotted onto nylon membrane filters (Ny-tran45; Schleicher & Schuell). The filters were prehybrid-ized and hybridized in the same solution containing random-primed AVP cDNA as probe as detailed (5). The filters werewashed and exposed to Kodak X-Omat AR x-ray film for 12h at -850C.SDS/PAGE and Western Analyses. One-dimensional SDS/
PAGE was performed on concave exponential gradient gelsafter delipidation of the samples (15). The electrophoresedsamples were electroblotted to 0.45-jm nitrocellulose filtersusing a semidry electroblotting apparatus in 10% (vol/vol)methanol/10 mM Caps-NaOH, pH 11.0, for 30 min at acurrent density of 2.5 mA/cm2. Transferred protein wasreversibly stained with Ponceau S, blocked, and incubatedwith V-PPase antibody as described (16). The rabbit poly-clonal antibody employed in these studies (PABHKi) wasraised against keyhole limpet hemocyanin-conjugated syn-thetic peptide with the sequence HKAAVIGDTIGDPLK,
Plant Biology: Kim et aL
Proc. Nati. Acad. Sci. USA 91 (1994)
corresponding to positions 720-734 of the deduced aminoacid sequence of the polypeptide encoded by AVP (5).PPase and Protein Assays. PPi hydrolysis and PPi-
dependent H+ translocation were assayed as detailed by Reaand Turner (13). Protein was estimated by a modification ofthe method of Peterson (17).
RESULTSTrformaton of Yeast with pYES2-AVP Expression Vec-
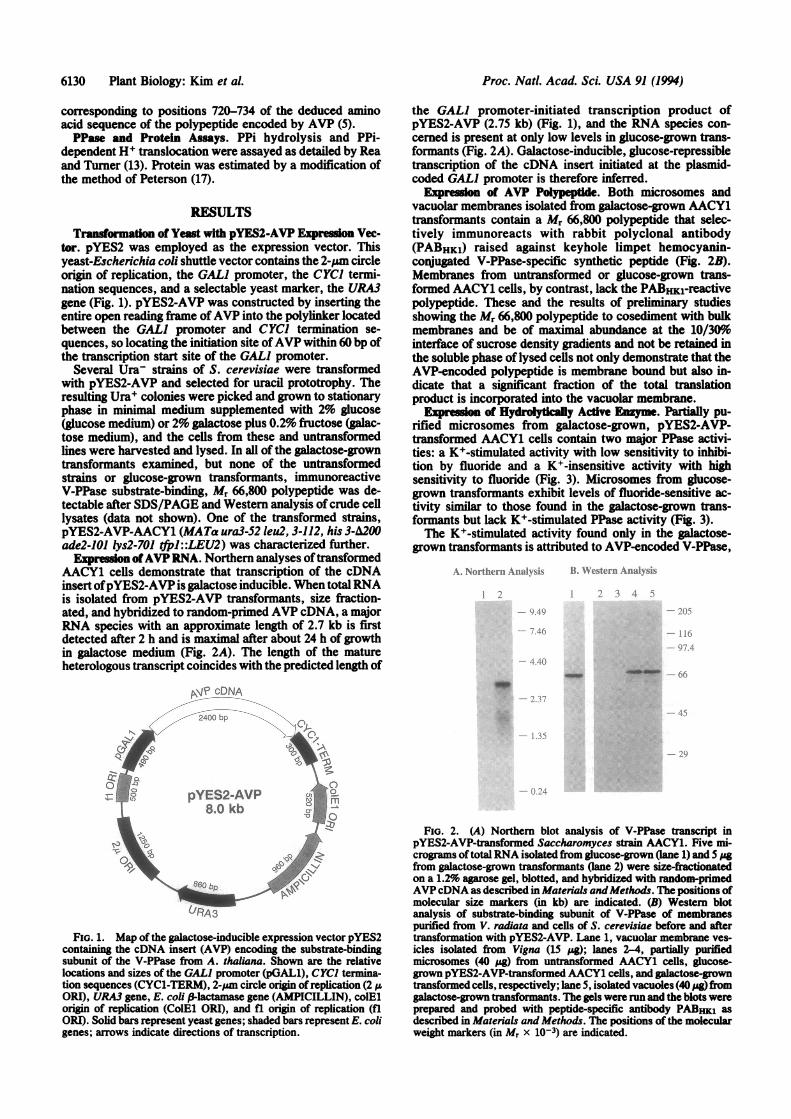
tor. pYES2 was employed as the expression vector. Thisyeast-Escherichia coli shuttle vector contains the 2-pam circleorigin of replication, the GAL) promoter, the CYCI termi-nation sequences, and a selectable yeast marker, the URA3gene (Fig. 1). pYES2-AVP was constructed by inserting theentire open reading frame ofAVP into the polylinker locatedbetween the GAL) promoter and CYCI termination se-quences, so locating the initiation site ofAVP within 60 bp ofthe transcription start site of the GAL) promoter.
Several Ura- strains of S. cerevisiae were transformedwith pYES2-AVP and selected for uracil prototrophy. Theresulting Ura+ colonies were picked and grown to stationaryphase in minimal medium supplemented with 2% glucose(glucose medium) or 2% galactose plus 0.2% fructose (galac-tose medium), and the cells from these and untransformedlines were harvested and lysed. In all of the galactose-growntransformants examined, but none of the untransformedstrains or glucose-grown transformants, immunoreactiveV-PPase substrate-binding, Mr 66,800 polypeptide was de-tectable after SDS/PAGE and Western analysis of crude celllysates (data not shown). One of the transformed strains,pYES2-AVP-AACY1 (MATa ura3-52 leu2, 3-112, his 3-A200ade2-101 lys2-701 (fpl::LEU2) was characterized further.
Expression ofAVPRNA. Northern analyses oftransformedAACY1 cells demonstrate that transcription of the cDNAinsert ofpYES2-AVP is galactose inducible. When totalRNAis isolated from pYES2-AVP transformants, size fraction-ated, and hybridized to random-primed AVP cDNA, a majorRNA species with an approximate length of 2.7 kb is firstdetected after 2 h and is maximal after about 24 h of growthin galactose medium (Fig. 2A). The length of the matureheterologous transcript coincides with the predicted length of
rpN' cDNA
the GAL) promoter-initiated transcription product ofpYES2-AVP (2.75 kb) (Fig. 1), and the RNA species con-cerned is present at only low levels in glucose-grown trans-formants (Fig. 2A). Galactose-inducible, glucose-repressibletranscription of the cDNA insert initiated at the plasmid-coded GAL) promoter is therefore inferred.
Expression of AVP Polypeptide. Both microsomes andvacuolar membranes isolated from galactose-grown AACY1transformants contain a Mr 66,800 polypeptide that selec-tively immunoreacts with rabbit polyclonal antibody(PABHK1) raised against keyhole limpet hemocyanin-conjugated V-PPase-specific synthetic peptide (Fig. 2B).Membranes from untransformed or glucose-grown trans-formed AACY1 cells, by contrast, lack the PABHK1-reactivepolypeptide. These and the results of preliminary studiesshowing the Mr 66,800 polypeptide to cosediment with bulkmembranes and be of maximal abundance at the 10/30%interface of sucrose density gradients and not be retained inthe soluble phase of lysed cells not only demonstrate that theAVP-encoded polypeptide is membrane bound but also in-dicate that a significant fraction of the total translationproduct is incorporated into the vacuolar membrane.
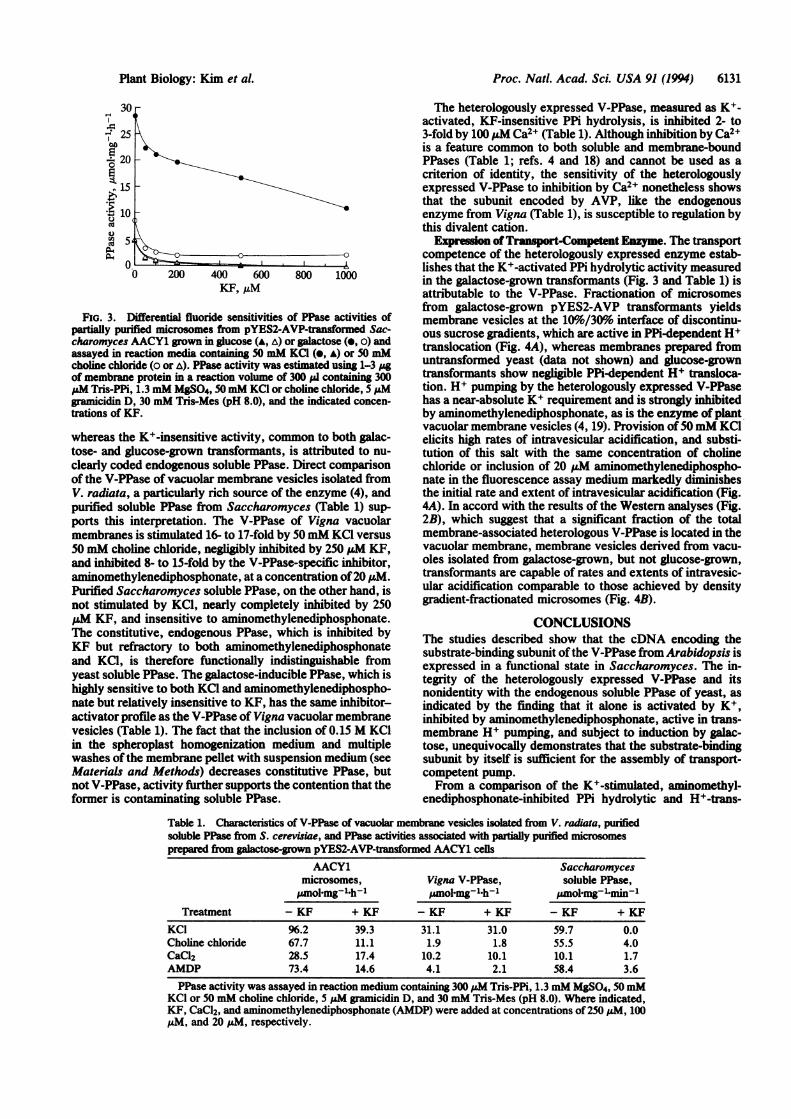
WI pra11IMof Hydroytically Active Enzyme. Partially pu-rified microsomes from galactose-grown, pYES2-AVP-transformed AACY1 cells contain two major PPase activi-ties: a K+-stimulated activity with low sensitivity to inhibi-tion by fluoride and a K+-insensitive activity with highsensitivity to fluoride (Fig. 3). Microsomes from glucose-grown transformants exhibit levels of fluoride-sensitive ac-tivity similar to those found in the galactose-grown trans-formants but lack K+-stimulated PPase activity (Fig. 3).The K+-stimulated activity found only in the galactose-
grown transformants is attributed to AVP-encoded V-PPase,
A. Northern Anah sis
1i
W1 .I
-p
2400 bp
pYES2-AVP8.0 kb
URA3
FIG. 1. Map of the galactose-inducible expression vector pYES2containing the cDNA insert (AVP) encoding the substrate-bindingsubunit of the V-PPase from A. thaliana. Shown are the relativelocations and sizes of the GAL] promoter (pGAL1), CYC) termina-tion sequences (CYCl-TERM), 2-pim circle origin ofreplication (2 IAORI), URA3 gene, E. coli P-lactamase gene (AMPICILLIN), colElorigin of replication (CoIEl ORI), and fl origin of replication (flORI). Solid bars represent yeast genes; shaded bars represent E. coligenes; arrows indicate directions of transcription.
FiG. 2. (A) Northern blot analysis of V-PPase transcript inpYES2-AVP-transformed Saccharomyces strain AACY1. Five mi-crograms of total RNA isolated from glucose-grown (lane 1) and 5 pgfrom galactose-grown transformants (lane 2) were size-frationatedon a 1.2% agarose gel, blotted, and hybridized with random-primedAVPcDNA as described in Materials andMethods. The positions ofmolecular size markers (in kb) are indicated. (B) Western blotanalysis of substrate-binding subunit of V-PPase of membranespurified from V. radiata and cells of S. cerevisiae before and aftertransformation with pYES2-AVP. Lane 1, vacuolar membrane ves-icles isolated from Vigna (15 pig); lanes 2-4, partially purifiedmicrosomes (40 pug) from untransformed AACY1 cells, glucose-grown pYES2-AVP-transformed AACY1 cells, and galactose-growntransformed cells, respectively; lane 5, isolated vacuoles (40 pg fromgalactose-grown transformants. The gels were run and the blots wereprepared and probed with peptide-specific antibody PABHKI asdescribed in Materials and Methods. The positions of the molecularweight markers (in Mr x 10-3) are indicated.
6130 Plant Biology: Kim et al.
B. W\esternAl.-IulN-sis
-- i1ll _ TIo
4. -dNWMW4NPP* (I f.
_ +..iz
Proc. Nati. Acad. Sci. USA 91 (1994) 6131
400 600KF, AM
FIG. 3. Differential fluoride sensitivities of PPase activities ofpartially purified microsomes from pYES2-AVP-tranormed Sac-charomyces AACY1 grown in glucose (A, a) or galactose (e, o) andassayed in reaction media containing 50 mM KCl (e, A) or 50 mMcholine chloride (o or A). PPase activity was estimated using 1-3 pgof membrane protein in a reaction volume of 300 pA containing 300pM Tris-PPi, 1.3 mM MgSO4, 50 mM KCI or choline chloride, 5 pMgramicidin D, 30 mM Tris-Mes (pH 8.0), and the indicated concen-trations of KF.
whereas the K+-insensitive activity, common to both galac-tose- and glucose-grown transformants, is attributed to nu-
clearly coded endogenous soluble PPase. Direct comparisonof the V-PPase of vacuolar membrane vesicles isolated fromV. radiata, a particularly rich source of the enzyme (4), andpurified soluble PPase from Saccharomyces (Table 1) sup-ports this interpretation. The V-PPase of Vigna vacuolarmembranes is stimulated 16- to 17-fold by 50 mM KCI versus50 mM choline chloride, negligibly inhibited by 250 pM KF,and inhibited 8- to 15-fold by the V-PPase-specific inhibitor,aminomethylenediphosphonate, at a concentration of20 IAM.Purified Saccharomyces soluble PPase, on the other hand, isnot stimulated by KCI, nearly completely inhibited by 250MM KF, and insensitive to aminomethylenediphosphonate.The constitutive, endogenous PPase, which is inhibited byKF but refractory to both aminomethylenediphosphonateand KCI, is therefore functionally indistinguishable fromyeast soluble PPase. The galactose-inducible PPase, which ishighly sensitive to both KCI and aminomethylenediphospho-nate but relatively insensitive to KF, has the same inhibitor-activator profile as the V-PPase of Vigna vacuolar membranevesicles (Table 1). The fact that the inclusion of 0.15 M KCIin the spheroplast homogenization medium and multiplewashes of the membrane pellet with suspension medium (seeMaterials and Methods) decreases constitutive PPase, butnot V-PPase, activity further supports the contention that theformer is contaminating soluble PPase.
The heterologously expressed V-PPase, measured as K+-activated, KF-insensitive PPi hydrolysis, is inhibited 2- to3-fold by 100 .uM Ca2+ (Table 1). Although inhibition by Ca2+is a feature common to both soluble and membrane-boundPPases (Table 1; refs. 4 and 18) and cannot be used as acriterion of identity, the sensitivity of the heterologouslyexpressed V-PPase to inhibition by Ca2+ nonetheless showsthat the subunit encoded by AVP, like the endogenousenzyme from Vigna (Table 1), is susceptible to regulation bythis divalent cation.
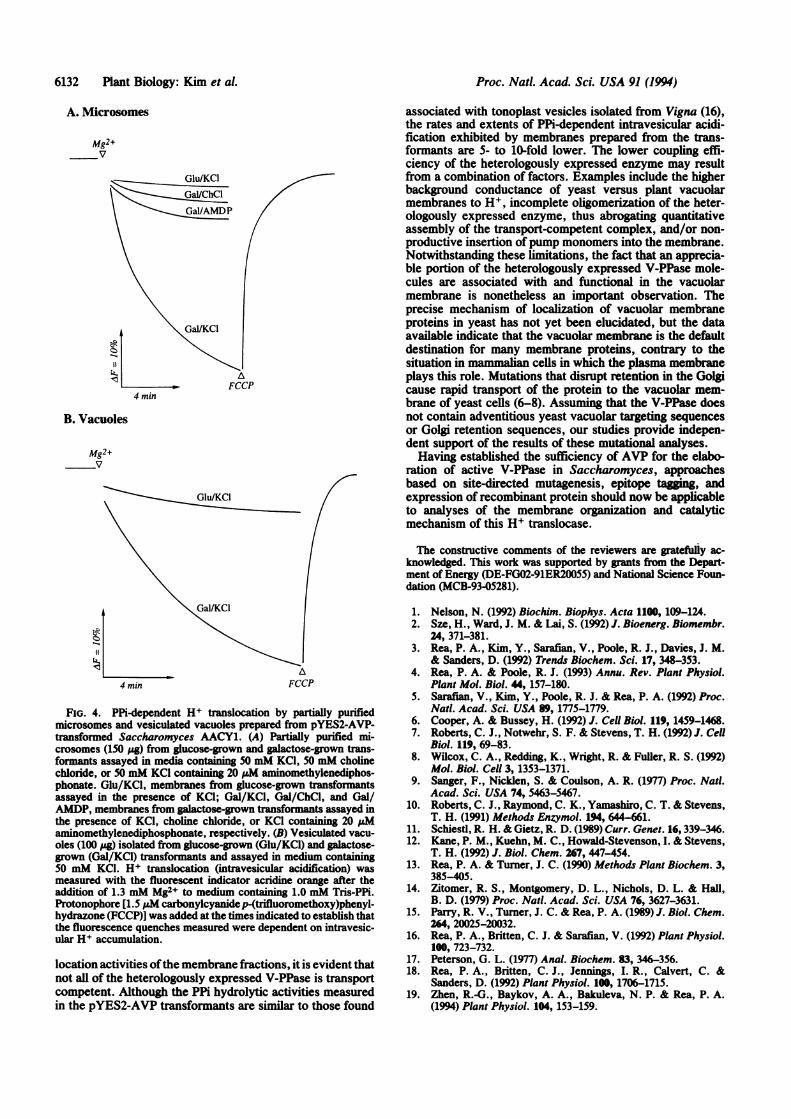
Expressin ofTranspot-Competent Enzyme. The transportcompetence of the heterologously expressed enzyme estab-lishes that the K+-activated PPi hydrolytic activity measuredin the galactose-grown transformants (Fig. 3 and Table 1) isattributable to the V-PPase. Fractionation of microsomesfrom galactose-grown pYES2-AVP transformants yieldsmembrane vesicles at the 10%6/30%o interface of discontinu-ous sucrose gradients, which are active in PPi-dependent H+translocation (Fig. 4A), whereas membranes prepared fromuntransformed yeast (data not shown) and glucose-growntransformants show negligible PPi-dependent H+ transloca-tion. H+ pumping by the heterologously expressed V-PPasehas a near-absolute K+ requirement and is strongly inhibitedby aminomethylenediphosphonate, as is the enzyme ofplantvacuolar membrane vesicles (4, 19). Provision of50mM KClelicits high rates of intravesicular acidification, and substi-tution of this salt with the same concentration of cholinechloride or inclusion of 20 ,uM aminomethylenediphospho-nate in the fluorescence assay medium markedly diminishesthe initial rate and extent of intravesicular acidification (Fig.4A). In accord with the results of the Western analyses (Fig.2B), which suggest that a significant fraction of the totalmembrane-associated heterologous V-PPase is located in thevacuolar membrane, membrane vesicles derived from vacu-oles isolated from galactose-grown, but not glucose-grown,transformants are capable of rates and extents of intravesic-ular acidification comparable to those achieved by densitygradient-fractionated microsomes (Fig. 4B).
CONCLUSIONSThe studies described show that the cDNA encoding thesubstrate-binding subunit ofthe V-PPase from Arabidopsis isexpressed in a functional state in Saccharomyces. The in-tegrity of the heterologously expressed V-PPase and itsnonidentity with the endogenous soluble PPase of yeast, asindicated by the finding that it alone is activated by K+,inhibited by aminomethylenediphosphonate, active in trans-membrane H+ pumping, and subject to induction by galac-tose, unequivocally demonstrates that the substrate-bindingsubunit by itself is sufficient for the assembly of transport-competent pump.From a comparison of the K+-stimulated, aminomethyl-
enediphosphonate-inhibited PPi hydrolytic and H+-trans-
Table 1. Characteristics of V-PPase of vacuolar membrane vesicles isolated from V. radiata, purifiedsoluble PPase from S. cerevisiae, and PPase activities associated with partially purified microsomesprepare from galactose-grown pYES2-AVP-transformed AACY1 cells
AACY1 Saccharomycesmicrosomes, Vigna V-PPase, soluble PPase,zmol-mg-1h-1 smol-mg-'h+- umol-mg-1min-1
Treatment - KF + KF - KF + KF - KF + KFKCI 96.2 39.3 31.1 31.0 59.7 0.0Choline chloride 67.7 11.1 1.9 1.8 55.5 4.0CaC12 28.5 17.4 10.2 10.1 10.1 1.7AMDP 73.4 14.6 4.1 2.1 58.4 3.6PPase activity was assayed in reaction medium containing 300 pAM Tris-PPi, 1.3 mM MgSO4, 50mM
KCI or 50 mM choline chloride, 5 pM gramicidin D, and 30 mM Tris-Mes (pH 8.0). Where indicated,KF, CaCl2, and aminomethylenediphosphonate (AMDP) were added at concentrations of 250 pM, 100,uM, and 20 IAM, respectively.
Plant Biology: Kim et al.
Proc. Natl. Acad. Sci. USA 91 (1994)
A. Microsomes
Mg2+v
4 mmi
B. Vacuoles
Mg2+v
GluIKCI
4 min
A
FCCP
FIG. 4. PPi-dependent H+ translocation by partially purifiedmicrosomes and vesiculated vacuoles prepared from pYES2-AVP-transformed Saccharomyces AACY1. (A) Partially purified mi-crosomes (150 ptg) from glucose-grown and galactose-grown trans-formants assayed in media con ing 50 mM KCl, 50 mM cholinechloride, or 50 mM KC1 containing 20 gsM aminomethylenediphos-
phonate. Glu/KCl, membranes from glucose-grown transformantsassayed in the presence of KCl; Gal/KCl, Gal/ChCl, and Gal/AMDP, membranes from galactose-grown transformants assayed inthe presence of KCl, choline chloride, or KCI containing 20 pMaminomethylenediphosphonate, respectively. (B) Vesiculated vacu-oles (100 Ag) isolated from glucose-grown (Glu/KCl) and galactose-grown (Gal/KCl) transformants and assayed in medium containing50 mM KCl. H+ translocation (intravesicular acidification) wasmeasured with the fluorescent indicator acridine orange after theaddition of 1.3 mM Mg2+ to medium containing 1.0 mM Tris-PPi.Protonophore [1.5 pM carbonylcyanidep-(trifluoromethoxy)phenyl-hydrazone (FCCP)J was added at the times indicated to establish thatthe fluorescence quenches measured were dependent on intravesic-ular H+ accumulation.
location activities ofthe membrane fractions, it is evident thatnot all of the heterologously expressed V-PPase is transportcompetent. Although the PPi hydrolytic activities measuredin the pYES2-AVP transformants are similar to those found
associated with tonoplast vesicles isolated from Vigna (16),the rates and extents of PPi-dependent intravesicular acidi-fication exhibited by membranes prepared from the trans-formants are 5- to 10-fold lower. The lower coupling effi-ciency of the heterologously expressed enzyme may resultfrom a combination of factors. Examples include the higherbackground conductance of yeast versus plant vacuolarmembranes to H+, incomplete oligomerization of the heter-ologously expressed enzyme, thus abrogating quantitativeassembly of the transport-competent complex, and/or non-productive insertion ofpump monomers into the membrane.Notwithstanding these limitations, the fact that an apprecia-ble portion of the heterologously expressed V-PPase mole-cules are associated with and functional in the vacuolarmembrane is nonetheless an important observation. Theprecise mechanism of localization of vacuolar membraneproteins in yeast has not yet been elucidated, but the dataavailable indicate that the vacuolar membrane is the defaultdestination for many membrane proteins, contrary to thesituation in mammalian cells in which the plasma membraneplays this role. Mutations that disrupt retention in the Golgicause rapid transport of the protein to the vacuolar mem-brane of yeast cells (6-8). Assuming that the V-PPase doesnot contain adventitious yeast vacuolar targeting sequencesor Golgi retention sequences, our studies provide indepen-dent support of the results of these mutational analyses.Having established the sufficiency of AVP for the elabo-
ration of active V-PPase in Saccharomyces, approachesbased on site-directed mutagenesis, epitope tagging, andexpression of recombinant protein should now be applicableto analyses of the membrane organization and catalyticmechanism of this H+ translocase.
The constructive comments of the reviewers are gratefully ac-knowledged. This work was supported by grants firom the Depart-ment of Energy (DE-FG02-91ER20055) and National Science Foun-dation (MCB-93-05281).
1. Nelson, N. (1992) Biochim. Biophys. Acta 1100, 109-124.2. Sze, H., Ward, J. M. & Lai, S. (1992) J. Bioenerg. Biomembr.
24, 371-381.3. Rea, P. A., Kim, Y., Sarafian, V., Poole, R. J., Davies, J. M.
& Sanders, D. (1992) Trends Biochem. Sci. 17, 348-353.4. Rea, P. A. & Poole, R. J. (1993) Annu. Rev. Plant Physiol.
Plant Mol. Biol. 44, 157-180.5. Saraflan, V., Kim, Y., Poole, R. J. & Rea, P. A. (1992) Proc.
Natl. Acad. Sci. USA 89, 1775-1779.6. Cooper, A. & Bussey, H. (1992) J. Cell Biol. 119, 1459-1468.7. Roberts, C. J., Notwehr, S. F. & Stevens, T. H. (1992) J. Cell
Biol. 119, 69-83.8. Wilcox, C. A., Redding, K., Wright, R. & Fuller, R. S. (1992)
Mol. Biol. Cell 3, 1353-1371.9. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl.
Acad. Sci. USA 74, 5463-5467.10. Roberts, C. J., Raymond, C. K., Yamashiro, C. T. & Stevens,
T. H. (1991) Methods Enzymol. 194, 644-661.11. Schiestl, R. H. & Gietz, R. D. (1989) Curr. Genet. 16, 339-346.12. Kane, P. M., Kuehn, M. C., Howald-Stevenson, I. & Stevens,
T. H. (1992) J. Biol. Chem. 267, 447-454.13. Rea, P. A. & Turner, J. C. (1990) Methods Plant Biochem. 3,
385-405.14. Zitomer, R. S., Montgomery, D. L., Nichols, D. L. & Hall,
B. D. (1979) Proc. Natl. Acad. Sci. USA 76, 3627-3631.15. Parry, R. V., Turner, J. C. & Rea, P. A. (1989) J. Biol. Chem.
264, 20025-20032.16. Rea, P. A., Britten, C. J. & Sarafian, V. (1992) Plant Physiol.
100, 723-732.17. Peterson, G. L. (1977) Anal. Biochem. 83, 346-356.18. Rea, P. A., Britten, C. J., Jennings, I. R., Calvert, C. &
Sanders, D. (1992) Plant Physiol. 100, 1706-1715.19. Zhen, R.-G., Baykov, A. A., Bakuleva, N. P. & Rea, P. A.
(1994) Plant Physiol. 104, 153-159.
6132 Plant Biology: Kim et al.