degradation ofhydrogen sulfide xanthomonas strain dy44 ...aem.asm.org/content/58/4/1183.full.pdf ·...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1992, p. 1183-11890099-2240/92/041183-07$02.00/0Copyright © 1992, American Society for Microbiology
Degradation of Hydrogen Sulfide by Xanthomonas sp. Strain DY44Isolated from Peat
KYEOUNG-SUK CHO, MITSUYO HIRAI, AND MAKOTO SHODA*Research Laboratory ofResources Utilization, Tokyo Institute of Technology, 4259 Nagatsuta, Midori-ku,
Yokohama 227, Japan
Received 4 October 1991/Accepted 8 January 1992
Xanthomonas sp. strain DY44, capable of degrading H2S, was isolated from dimethyl disulfide-acclimatedpeat. This bacterium removed H2S either as a single gas or in the presence of the sulfur-containing compoundsmethanethiol, dimethyl sulfide, and dimethyl disulfide. The maximum specific H2S removal rate, obtained inthe late stationary phase, was 3.92 mmol g of dry cells-' h-1 (6.7 x 10-16 mol cell-' h-1) at pH 7 and 30°Cthrough a batch experiment in a basal mineral medium. Since Xanthomonas sp. strain DY44 exhibited no
autotrophic growth with H2S, the H2S removal was judged not to be a consequence of chemolithotrophicactivity. By using X-ray photoelectron spectroscopy, the metabolic product of H2S oxidation was determinedto be polysulfide, which has properties very similar to those of elemental sulfur. Autoclaved cells (120°C, 20min) did not show H2S degradation, but cells killed by -y-irradiation and cell extracts both oxidized H2S,suggesting the existence of a heat-labile intracellular enzymatic system for H2S oxidation. When Xanthomonassp. strain DY44 was inoculated into fibrous peat, this strain degraded H2S without lag time, suggesting that itwill be a good candidate for maintaining high H2S removability in the treatment of exhaust gases.
Hydrogen sulfide (H2S) is one of the main malodorouscompounds occurring in exhaust gases. This compound isoften evolved in the highest concentration together withsulfur-containing volatile compounds such as methanethiol(MT), dimethyl sulfide (DMS), and dimethyl disulfide(DMDS) from wastewater and night soil treatment plants. Toremove H2S, physicochemical methods are generally used.However, biological methods, using microbial activity forH2S removal, have drawn attention since they are moreefficient and more economical than physicochemical meth-ods if proper operational conditions are maintained. Anumber of microbial processes for H2S removal have beenproposed that are based on oxidation by thiobacilli and othersulfur microorganisms. Thiobacillus spp. have been used tooxidize H2S to sulfate in liquid (19) and in packed towersystems such as peat biofilters (4, 5, 21, 23). Mixed culturesof bacteria from the Beggiatoa and Thiothrix genera (14) andthe photosynthetic bacterium Chlorobium thiosulfatophilumhave been proposed for use in the oxidation of H2S toelemental sulfur from the gas stream (6). Most of them are
autotrophic bacteria, but these bacteria are often difficult tohandle, mainly because their growth rates are significantlylower than those of heterotrophic bacteria. In applyingphototrophic microorganisms, the supply of solar or artificialenergy is a limiting factor for growth.
This paper describes some characteristics of a new H2S-degrading heterotrophic bacterium, a Xanthomonas sp.,isolated from DMDS-acclimated peat; the advantageousfeatures of this bacterium over autotrophic bacteria as an
H2S remover are also discussed.
MATERIALS AND METHODSMedia and cultivation conditions. A nutrient medium was
used for cultivation of the isolate and contained the following(in grams per liter): meat extract, 3.0; peptone, 15.0; yeastextract, 3.0; Na2HPO4. 12H20, 2.0; and NaCl, 3.0. For
* Corresponding author.
batch- and continuous-supply H2S experiments, a basalmineral medium, containing 2.0 g of K2HPO4, 2.0 g ofKH2PO4, 2.0 g of KH2PO4, 0.4 g of NH4CI, 0.2 g ofMgCl2 6H20, and 0.01 g of FeSO4. 7H20 (each per liter),was used. The pH was adjusted to 7 by using 2 N NaOH inboth the nutrient and basal media.
Isolation and identification of the bacterium. A heterotro-phic bacterium was isolated by chance in the process ofisolating the autotrophic bacterium Thiobacillus thioparusDW44 on thiosulfate-agar medium from DMDS-acclimatedpeat (3, 4). The heterotrophic bacterium was purified byrepeated transfer of the cells to fresh nutrient medium andwas labeled DY44. Identification of strain DY44 was carriedout by the National Collections of Industrial and MarineBacteria Limited, Torry Research Station, Aberdeen,United Kingdom.H2S removal in batch system. Cells grown in 100 ml of the
nutrient medium for 4 days at 30°C were harvested bycentrifugation (7,500 x g, 10 min), washed in sterilized,distilled water, and resuspended in 100 ml of sterilizedwater. One milliliter of the cell suspension (about 1010 cells)was added to 99 ml of the basal mineral medium in a 500-mlflask. One milliliter of an autoclaved cell suspension (120°Cfor 20 min) and 1 ml of -y-irradiated cells (60Co for 1 h at 1megarad h-1) were added to similar flasks. As a control, 1 mlof sterilized, distilled water was added to a fourth flask. Theflasks were tightly sealed with butyl rubber stoppers, H2Sgas from a cylinder containing 20,000 ,ul of H2S liter-' in N2(Takachiho Co. Inc., Tokyo, Japan) was injected into eachflask through the stopper to give a total H2S amount of 10,umol per flask (the aqueous H2S concentration at 10 ,umolwas 44.9 ,uM), and the flasks were shaken at 120 rpm at 30°C.The headspace of each flask was sampled periodically byusing a gastight syringe, and the H2S concentration was
determined by gas chromatography (model GC-14 with a
flame photometric detector; Shimadzu Co. Ltd., Kyoto,Japan). The aqueous H2S concentration was calculated fromthe gas concentration by using Henry's constant (13). Details
1183
Vol. 58, No. 4
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
of experimental and analytical methods were describedpreviously (5, 9).H2S removal by cell extract. Cells harvested in the manner
described above were suspended in 10 mM Tris-HCl buffer(pH 8) containing 0.2 M NaCl, 100 ,uM EDTA, and 10 ,uMphenylmethylsulfonyl fluoride and passed twice through achilled French pressure cell at 1,000 kg cm-2. After DNAwas digested by the addition of DNase, intact cells wereremoved by centrifugation at 10,000 x g for 10 min. Thesupernatant was centrifuged at 30,000 x g for 1 h at 4°C andused as the crude cell extract. One milliliter of the cellextract and 9 ml of 10 mM Tris-HCl buffer (pH 7.4; finalprotein concentration, 0.87 mg ml-') were put into a 123-mlvial bottle which was then sealed with a butyl rubberstopper. A second 123-ml vial bottle containing 10 ml of 10mM Tris-HCl buffer without cell extract was used as acontrol. H2S gas was injected to give total H2S amounts of 4,umol per bottle (aqueous H2S concentration at 4 ,umol, 166.9FM), and the H2S concentration was measured periodicallyas described above.Growth pattern and H2S removal rates in different growth
phases. Cells grown in nutrient-agar medium were suspendedin 10 ml of sterilized water, and 1 ml of the suspension (about1010 cells) was inoculated into 99 ml of fresh nutrient mediumin each of eight 500-ml flasks. The flasks were incubated at30°C, and the growth of strain DY44 was monitored bymeasuring dry weight after the whole cells in one flask weredried at 110°C for 4 h at each cultivation time. The pH of theculture broth was also measured. H2S removal rates after 1and 4 days of cultivation time were determined as follows.The cells harvested by centrifugation at each time werewashed and resuspended in 10 ml of distilled water. The cellsuspension was diluted with distilled water at differentdilution ratios, with 1 ml of each differently diluted suspen-sion being added to 99 ml of the basal mineral medium (pH7) in a 500-ml flask. H2S removal by the bacterial suspensionwas determined for each flask by the same procedures asdescribed above. The specific removal rate (millimoles ofH2S per gram of dry cells per hour) was calculated from thetotal amount of H2S removed in both the gas and liquidphases divided by the dry cell mass.
Experiment with a continuous supply of H2S in the basalmineral medium. Figure la shows the bubbling system forthe experiment using a continuous supply of H2S. Tenmilliliters of cell suspension (about 1011 cells; dry weight ofcells, 0.2 g) was added to a 300-ml glass bubbling bottlecontaining 240 ml of the basal mineral medium. About 200 to250 [LI of H2S gas liter-', which was generated by thedilution of H2S gas from a cylinder (20,000 ,ul liter-') with airfrom a compressor, was supplied into the bottle at a flow rateof 100 ml min-1 at 30°C for 16 days. During the supply ofH2S gas, the inlet and outlet concentrations of H2S wereintermittently measured; the culture broth was also sampled,and the pH and the viable cell count on the nutrient-agarmedium were determined.H2S removal in the presence of organic compounds. H2S
removal by the isolate in the presence of organic compoundswas studied in batch and continuous systems. In the batchexperiment, glucose, maltose, and acetate were added to thebasal mineral medium to give a final concentration of 2 gliter-1, and H2S removal was monitored in the same manneras described above. In the continuous experiment, 1 ml ofcell suspension (about 1010 cells) was added to a 300-mlbottle containing 199 ml of the basal mineral medium sup-plemented with 2 g of yeast extract liter-', and about 20 Rl
1
13
I | 10 (a)FIG. 1. Schematic diagram of bubbling system (a) and laborato-
ry-scale peat biofilter (b). 1, air; 2, regulator; 3, pressure gauge; 4,H2S cylinder; 5, control valve; 6, flow meter; 7, three-way cock; 8,air filter (0.2-,um-pore-size membrane); 9, bubbling bottle; 10, con-ical flask; 11, thermostat; 12, glass column; 13, peat.
of H2S liter-1 was supplied into the bottle at a flow rate of200 ml min-' at 30°C.
Identification of the product of removed H2S. One milliliterof culture broth obtained from the continuous-supply exper-iment described above was filtered through a 0.45-,um-pore-size membrane. Anionic sulfur compounds in the filtratewere analyzed by ion chromatography (HIC-6A; Shimadzu).The rest of the culture broth was lyophilized (FD-1; TokyoRikakikai Co. Ltd., Tokyo, Japan) and used for analysis ofsulfur compounds by X-ray photoelectron spectroscopy(XPS) (ESCA750; Shimadzu). Non-H2S-supplied culturebroth was used as a reference. To observe the differencebetween the two, a mixture of the lyophilized culture brothsupplied with H2S (1.25 g) and elemental sulfur (1 g) was alsosubjected to analysis by XPS.H2S removal in the presence of other sulfur-containing
gases. Two milliliters of cell suspension prepared as de-scribed above was added to 98 ml of the basal mineralmedium in each of four 500-ml flasks. The flasks were tightlysealed with butyl rubber stoppers, and H2S gas was injectedinto each flask through the stopper (total H2S amount, about8 ,umol per flask). The following volumes of MT, DMS, orDMDS were then injected to give total amounts of 2, 1, and2 ,umol per flask, respectively: 10 ml of MT gas from acylinder containing 8,200 ,ul of MT liter-' in N2 (Takachiho);0.1 ml of DMS gas from the headspace of a glass vesselcontaining a 98% DMS solution (Kanto Chemical Co. Inc.,Tokyo, Japan) at 25°C; and 2 ml of DMDS gas from theheadspace of a 99% DMDS solution (Tokyo Kasei Co. Inc.,Tokyo, Japan) at 25°C. The fourth flask contained H2S gasonly as a control. During shaking at 120 rpm at 30°C, theheadspace of each flask was sampled, and gas concentra-tions were determined periodically by gas chromatography.H2S removal experiment using a peat biofilter inoculated
with strain DY44. An experiment on the removal of H2S bystrain DY44 inoculated on peat was carried out in a labora-tory-scale peat column, which is referred to as a peatbiofilter (4, 5) (Fig. lb). Forty-five grams of dry fibrous peat,
1184 CHO ET AL.
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
H2S DEGRADATION BY XANTHOMONAS SP. STRAIN DY44 1185
12
10
0 1 2 3 4 5 6
6
5 C
CMo fon E
003 '
2 E
co
- 4cm
0
o
-0. 2i
1
CM n,
10
9
8 ^
I7
6
5
Time (h)
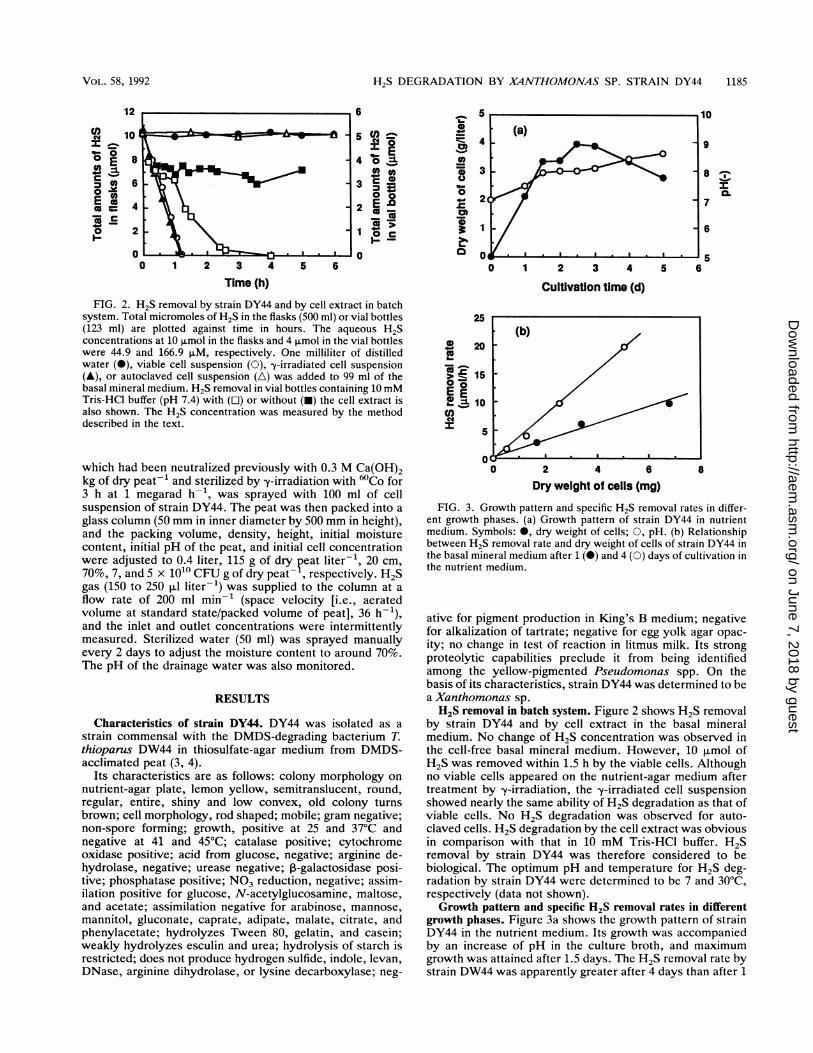
FIG. 2. H2S removal by strain DY44 and by cell extract in batchsystem. Total micromoles of H2S in the flasks (500 ml) or vial bottles(123 ml) are plotted against time in hours. The aqueous H2Sconcentrations at 10 ,umol in the flasks and 4 p.mol in the vial bottleswere 44.9 and 166.9 pLM, respectively. One milliliter of distilledwater (@), viable cell suspension (0), y-irradiated cell suspension(A), or autoclaved cell suspension (A) was added to 99 ml of thebasal mineral medium. H2S removal in vial bottles containing 10 mMTris-HCl buffer (pH 7.4) with ([l) or without (-) the cell extract isalso shown. The H2S concentration was measured by the methoddescribed in the text.
which had been neutralized previously with 0.3 M Ca(OH)2kg of dry peat-' and sterilized by y-irradiation with 60Co for3 h at 1 megarad h-', was sprayed with 100 ml of cellsuspension of strain DY44. The peat was then packed into aglass column (50 mm in inner diameter by 500 mm in height),and the packing volume, density, height, initial moisturecontent, initial pH of the peat, and initial cell concentrationwere adjusted to 0.4 liter, 115 g of dry peat liter-', 20 cm,70%, 7, and 5 x 1010 CFU g of dry peat- , respectively. H2Sgas (150 to 250 RI liter-') was supplied to the column at aflow rate of 200 ml min-' (space velocity [i.e., aeratedvolume at standard state/packed volume of peat], 36 h-1),and the inlet and outlet concentrations were intermittentlymeasured. Sterilized water (50 ml) was sprayed manuallyevery 2 days to adjust the moisture content to around 70%.The pH of the drainage water was also monitored.
RESULTS
Characteristics of strain DY44. DY44 was isolated as astrain commensal with the DMDS-degrading bacterium T.thioparus DW44 in thiosulfate-agar medium from DMDS-acclimated peat (3, 4).
Its characteristics are as follows: colony morphology on
nutrient-agar plate, lemon yellow, semitranslucent, round,regular, entire, shiny and low convex, old colony turnsbrown; cell morphology, rod shaped; mobile; gram negative;non-spore forming; growth, positive at 25 and 37°C andnegative at 41 and 45°C; catalase positive; cytochromeoxidase positive; acid from glucose, negative; arginine de-hydrolase, negative; urease negative; ,3-galactosidase posi-tive; phosphatase positive; NO3 reduction, negative; assim-ilation positive for glucose, N-acetylglucosamine, maltose,and acetate; assimilation negative for arabinose, mannose,mannitol, gluconate, caprate, adipate, malate, citrate, andphenylacetate; hydrolyzes Tween 80, gelatin, and casein;weakly hydrolyzes esculin and urea; hydrolysis of starch isrestricted; does not produce hydrogen sulfide, indole, levan,DNase, arginine dihydrolase, or lysine decarboxylase; neg-
Cultivation time (d)
25
202I-_
:Ir', 15a 0E E0A- =1 10
r
2 4 6
Dry weight of cells (mg)
FIG. 3. Growth pattern and specific H2S removal rates in differ-ent growth phases. (a) Growth pattern of strain DY44 in nutrientmedium. Symbols: *, dry weight of cells; 0, pH. (b) Relationshipbetween H2S removal rate and dry weight of cells of strain DY44 inthe basal mineral medium after 1 (l) and 4 (0) days of cultivation inthe nutrient medium.
ative for pigment production in King's B medium; negativefor alkalization of tartrate; negative for egg yolk agar opac-ity; no change in test of reaction in litmus milk. Its strongproteolytic capabilities preclude it from being identifiedamong the yellow-pigmented Pseudomonas spp. On thebasis of its characteristics, strain DY44 was determined to bea Xanthomonas sp.H2S removal in batch system. Figure 2 shows H2S removal
by strain DY44 and by cell extract in the basal mineralmedium. No change of H2S concentration was observed inthe cell-free basal mineral medium. However, 10 ,umol ofH2S was removed within 1.5 h by the viable cells. Althoughno viable cells appeared on the nutrient-agar medium aftertreatment by y-irradiation, the y-irradiated cell suspensionshowed nearly the same ability of H2S degradation as that ofviable cells. No H2S degradation was observed for auto-claved cells. H2S degradation by the cell extract was obviousin comparison with that in 10 mM Tris-HCl buffer. H2Sremoval by strain DY44 was therefore considered to bebiological. The optimum pH and temperature for H2S deg-radation by strain DY44 were determined to be 7 and 30°C,respectively (data not shown).Growth pattern and specific H2S removal rates in different
growth phases. Figure 3a shows the growth pattern of strainDY44 in the nutrient medium. Its growth was accompaniedby an increase of pH in the culture broth, and maximumgrowth was attained after 1.5 days. The H2S removal rate bystrain DW44 was apparently greater after 4 days than after 1
0I-_- 0
o
o s
asoc
VOL. 58, 1992
5
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
B UL 9
co
=a
500
CM400
0 5300
p-200
C 1000
0
0 5 10 15
Time (d)
FIG. 4. H2S removal by strain DY44 in basal mineral mediumwith a continuous supply of H2S. H2S gas was supplied into thebottle at a flow rate of 100 ml min-1 at 30°C. Symbols: 0, inlet H2Sconcentration; A and A, outlet H2S concentrations in bottlescontaining only the basal mineral medium (A) and inoculated withstrain DY44 in the medium (A); a, cell number; *, pH.
day (Fig. 3b). After 5 days, the rate fell to about one half ofthe 4-day value, mainly because of autolysis of the cells (datanot shown), reflecting the fact that H2S-removing activity isat a maximum during cultivation time. From the slope of Fig.3b, the specific H2S removal rates of cells cultured for 1 and4 days in the nutrient medium were determined as 1.42 and3.92 mmol g of dry cells-' h-1, respectively.Removal of H2S in continuous supply in basal mineral
medium. The removal pattern of H2S which was continu-ously supplied at 100 ml min-' in the basal mineral mediumis shown in Fig. 4b. Xanthomonas sp. strain DY44 removedH2S gas without lag time, indicating that the intracellularenzymes responsible for H2S degradation are constitutive.Viable cell counts gradually decreased, mainly because ofcell autolysis. No change in the pH of the culture broth wasfound (Fig. 4a). The total amount of H2S removed by strainDY44 in the basal mineral medium over 16 days was esti-mated to be 0.1 mol g of dry cells-', which was nearly thesame as that for -y-irradiated cells, 0.11 mol g of dry cells-'.H2S removal in the presence of organic compounds. Xanth-
omonas sp. strain DY44 assimilates glucose, acetate, andmaltose, and its growth in the basal mineral medium supple-mented with these carbon sources (2 g liter-1) was confirmed(data not shown). In the presence of these compounds, H2Sremoval by this strain was studied with a batch system (Fig.
TABLE 1. Changes of cell number in strain DY44 duringcontinuous supply of H2S in the basal mineral medium with or
without 2 g of yeast extract liter-'
No. of cells (CFU ml-')
Medium After supply of H2SInitial
2 days 7 days
Without yeast extract 1.75 x 109 4.61 X 108 2.38 x 107With yeast extract 1.75 x 109 2.45 x 1010 1.11 x 101°
12
"
10
2E 8
:J 60~
0
1-. 2
0
25co
20
0
* 15a -1
.
%3 100 =0C
0 5
0 1 2
Time (h)3
0 2 4 6 8 10 12Time (d)
FIG. 5. H2S removal in the presence of organic compounds bystrain DY44. (a) Removal of batch-supplied H2S by strain DY44 inbasal mineral medium supplemented with 2 g of glucose (0), acetate(A), or maltose (A) liter-'. H2S removal in basal mineral mediumwhen only strain DY44 (0) was monitored as a control is alsoshown. Total micromoles of H2S in the flasks are plotted againsttime in hours. The aqueous H2S concentration at 11 ,umol was 49.3,uM. (b) Removal of continuously supplied H2S in basal mineralmedium supplemented with 2 g of yeast extract liter-'. H2S gas wassupplied into the bottle at a flow rate of 200 ml min-' at 30°C.Symbols: *, inlet H2S concentration; A and A, outlet H2S concen-trations in the bottles containing only the basal mineral medium (A)and strain DW44 in the medium (A); O and *, outlet H2S concen-trations in the bottles containing the basal mineral medium supple-mented with yeast extract ([) and strain DY44 in the medium plusyeast extract (A).
5a). The cell-free basal mineral medium supplemented withthese compounds showed no decline in H2S concentration(data not shown). However, H2S degradation was observedin all of the flasks inoculated with the cells. The presence oforganic compounds had no significantly adverse effects onH2S removal rates, although the rate in the presence ofglucose was relatively slower.
Figure 5b shows the removal of continuously supplied H2Sin the basal mineral medium with or without yeast extract.H2S degradation with only strain DY44 in the basal mineralmedium finally deteriorated in a way similar to that seen inFig. 4. This reflects a decrease in the viable cell number, asshown in Table 1. The addition of yeast extract to the basalmineral medium resulted in longer-lasting and more-efficientH2S degradation. This is mainly because the growth of strainDY44 was significant, as can be seen in Table 1. It is obviousthat cell-free yeast extract in the medium reacted with H2Schemically, and an apparent decrease in H2S concentration
1186 CHO ET AL.
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

H,S DEGRADATION BY XANTHOMONAS SP. STRAIN DY44 1187
(a)I I a I I
300 200Binding energy (eV)
FIG. 6. XPS spectra of the lyophilized culture broth with orwithout supply of H2S in the basal mineral medium. (a) Lyophilizedculture without H2S supply; (b) lyophilized culture with H2S supply;(c) mixture of lyophilized culture with H2S supply and elementalsulfur.
was observed. This fact indicates that the selection of themedium is of primary importance in verifying the purelybiological degradation of H2S.
Identification of product of removed H2S. During continu-ous supply of H2S in the basal mineral medium (Fig. 4),whitish-yellow precipitates appeared in the culture broth.No anionic sulfur compounds were detected in the filtrate ofthe culture broth by ion chromatography. Figure 6 shows thespectra by XPS analysis for lyophilized samples of culturebroth without (Fig. 6a) and with (Fig. 6b) H2S supply. Sulfurpeaks, S2s and S2p, appeared in the H2S-supplied culture at227.4 and 163.1 eV, respectively. The binding energy valueof S2p was very close to that of elemental sulfur, 164.05 eV
8(0
0 6
0. 4I-E |0
0*#- 2
0.0 0.5 1.0
Time (h)1.5 2.0
FIG. 7. H2S removal in the presence of MT (0), DMS (A), orDMDS (A) by strain DY44. H2S removal in a supply of H2S alone(A) served as a control. Total micromoles of H2S in the flasks areplotted against time in hours. The aqueous H2S concentration at 8,umol in the flasks was 36.0 ,uM.
(24), but this difference of 1 eV is significant. Two peaks atS2s and S2p were clearly observed when a mixture oflyophilized culture supplied with H2S and elemental sulfurwas analyzed (Fig. 6c). When the lyophilized sample ofH2S-supplied culture broth was extracted by CS2, elementalsulfur was obtained. On the basis of these results, themetabolite of H2S oxidized by strain DY44 was judged to be
_J polysulfide, which has properties very similar to elementalsulfur.H2S removal in the presence of sulfur-containing gases. The
H2S removal pattern in the presence of the sulfur-containinggases MT, DMS, and DMDS is shown in Fig. 7. Strain DY44removed H2S in the presence of MT, DMS, or DMDS,although the H2S removal rate in the presence of MT was40% of that in the presence either DMS or DMDS. The strainhad the ability to remove MT, but the removal rate was only1/100 that of H2S removal (data not shown). Neither DMSnor DMDS was removed in a single gas or in mixed gaseswith H2S by this strain (data not shown).H2S removal in a peat biofilter. Strain DY44 was inoculated
100 into peat, and its H2S degradation ability on the peat columnshown in Fig. 1 was tested. Figure 8 shows the H2S removalpattern in the peat biofilter. Clearly, strain DY44 exhibited
CMI0c_o2 20~c:
0C.)
300
250
200 1
150
100
50
00 5 10
Time (d)
7
655
15
FIG. 8. H2S removal by the peat biofilter inoculated with strainDY44. H2S gas was supplied into the column at a flow rate of 200 mlmin-' at 23°C. Symbols: *, inlet H2S concentration; 0, outlet H2Sconcentration; *, pH of drainage water.
VOL. 58, 1992
I
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
the ability to degrade H2S on the peat without a lag time. Nodecline of pH in drainage water was detected during theexperimental period.
DISCUSSION
We have demonstrated the characteristics of H2S removalby Xanthomonas sp. strain DY44 isolated from DMDS-acclimated peat. Diverse microorganisms capable of H2Soxidation have previously been reported: colorless sulfurbacteria such as Thiobacillus spp. (4, 5, 12, 18, 21, 23),Thiothrix sp. (1), and Beggiatoa sp. (2); methylotrophs suchas Hyphomicrobium sp. (20); phototrophs such as Chloro-bium spp. (11, 17), Chromatium sp. (7), Rhodobacter sp. (8),and Ectothiorhodospira sp. (22); and cyanobacteria (15).However, there have been few reports on H2S oxidation byaerobic heterotrophic microorganisms (16), and this is thefirst extensive report on H2S removal by a heterotrophicbacterium.H2S removal by Xanthomonas sp. strain DY44 seems to
be specific to this strain because four strains of Xanthomo-nas campestis (03-01420, 03-01424, 03-01426, and 03-01428)supplied by the National Institute of Agrobiological Re-sources (Tsukuba, Japan) were subjected to H2S removaltests, but no H2S removal was detected (data not shown).The specific H2S removal rate by 4-day-old cells in thenutrient medium was maximum at 3.92 mmol g of dry cells-'h-' (6.7 x 10-16 mol cell-1 h-1) (Fig. 3). As strain DY44 wasisolated as a strain commensal with T. thioparus DW44, wespeculate that T. thioparus DW44 and Xanthomonas sp.strain DY44 are major bacteria able to degrade H2S in theneutral pH region and in environmental conditions where theconcentration of organic matter is high. We have alreadyreported other autotrophic bacteria isolated from a peatbiofilter as H2S-oxidizing bacteria, namely Thiobacillus in-tennedius (23) and Thiobacillus sp. strain HA43 (5), in whichoptimal activity was expressed in the acidic pH region.The autoclaved cells (120°C, 20 min) did not degrade H2S,
but the cells treated by y-irradiation showed nearly the sameH2S-degrading ability as that of viable cells, and the cellextract degraded H2S, suggesting the existence of a heat-labile intracellular enzymatic system for H2S oxidation (Fig.2). The fact that strain DY44 cultivated in the nutrientmedium degraded H2S immediately after the supply of H2Sstarted (Fig. 4) suggests that the intracellular enzymaticsystem for H2S degradation formed in the nutrient mediumwas constitutive. After 16 days of H2S degradation, H2Sremoval stopped and the cell number decreased, indicatingthat H2S removal was not a consequence of chemo-lithotrophic activity. Since this strain oxidized neither thio-sulfate nor elemental sulfur in the basal mineral medium, andno growth of the strain was detected (data not shown), it isobvious that it does not grow chemolithotrophically withonly these sulfur compounds. On the other hand, this straindegraded H2S in the presence of organic compounds such asglucose, acetate, maltose, and yeast extract (Fig. 5), and itsgrowth in these compounds was not affected by the presenceof H2S. These results suggest that H2S degradation is notclosely linked to the utilization of organic compounds whichare associated with energy production. The oxidation of H2Sby Xanthomonas sp. strain DY44 is speculated to be phys-iologically an H2S detoxification process.The facts that whitish-yellow precipitates were generated
in the culture broth during the supply of H2S, no decline ofpH was observed (Fig. 4), and no anionic sulfur compoundswere detected in the culture filtrate indicate that the H2S
removal mechanism of this strain is different from that of theautotrophic bacteria reported previously (4, 5, 12, 18, 21,23). Actually, the product of H2S removal by Xanthomonassp. strain DY44 was identified as polysulfide by analysis withXPS (Fig. 6). During H2S oxidation by strain DY44, 02consumption was confirmed with a dissolved-oxygen elec-trode, suggesting that 02 was required for H2S oxidation(data not shown). A detailed biochemical study on themechanism of H2S oxidation in Xanthomonas sp. strainDY44 is in progress.There are several advantages of strain DY44 for practical
application in removing H2S in a peat biofilter. Although thespecific H2S removal rate of strain DY44 is lower than thoseof purified Thiobacillus spp. (4, 5, 23), harvesting of the cellmass for inoculation is easy because of its rapid growth inthe nutrient medium. No pH change was detected in the peatbiofilter inoculated with strain DY44 mainly because the finalproduct was polysulfide (Fig. 8). Sulfate is a final product ofmost autotrophic H2S-oxidizing bacteria, and pH decline issignificant. Polysulfide as a final product is preferable tosulfate, chiefly because a deterioration of microbial activitydue to a decline in pH will not occur. We have also alreadydemonstrated that the removability of MT, DMDS, andDMS, as well as of H2S, was sensitive to a decline in the pHof the peat (3, 10, 25). Considering the result shown in Fig.8b, higher efficiency in H2S removal can be obtained bysupplying organic compounds such as treated wastewater toa peat biofilter inoculated with Xanthomonas sp. strainDY44. The fact that removal of H2S was not influenced bythe coexistence of MT, DMS, or DMDS (Fig. 7) suggeststhat this bacterium can be applied to the treatment ofexhaust gases in wastewater or night soil treatment plantswhere these gases are the main malodorous components.Xanthomonas sp. strain DY44 can be also used to improvethe removability of MT, DMS, and DMDS in mixed cultureswith MT-, DMS-, and DMDS-degrading microorganisms,where the removability of these compounds is inhibited bythe coexistence of H2S.
ACKNOWLEDGMENTSWe thank K. Dohmen and J. Yoshimura, Research Laboratory of
Resource Utilization, Tokyo Institute of Technology, for assistancein measuring XPS and for valuable discussions.
REFERENCES1. Bland, J. A., and J. T. Staley. 1978. Observation on the biology
of Thiothrix. Arch. Microbiol. 117:79-89.2. Cannon, G. C., W. R. Strohl, J. M. Larkin, and J. M. Shively.
1979. Cytochromes in Beggiatoa alba. Curr. Microbiol. 2:263-266.
3. Cho, K. S., M. Hirai, and M. Shoda. 1991. Removal of dimethyldisulfide by the peat biofilter seeded with night soil sludge. J.Ferment. Bioeng. 71:289-291.
4. Cho, K. S., M. Hirai, and M. Shoda. 1991. Degradation charac-teristics of hydrogen sulfide, methanethiol, dimethyl sulfide anddimethyl disulfide by Thiobacillus thioparus DW44 isolatedfrom peat biofilter. J. Ferment. Bioeng. 71:384-389.
5. Cho, K. S., L. Zhang, M. Hirai, and M. Shoda. 1991. Removalcharacteristics of hydrogen sulfide and methanethiol by Thioba-cillus sp. isolated from peat in biological deodorization. J.Ferment. Bioeng. 71:44-49.
6. Cork, D. J., R. Garunas, and A. Sajjad. 1983. Chlorobiumlimicola forma thiosulfatophilum: biocatalyst in the productionof sulfur and organic carbon from a gas stream containing H2Sand CO2. Appl. Environ. Microbiol. 45:913-918.
7. Fukumori, Y., and T. Yamanaka. 1979. Flavocytochrome c ofChromatium vinosum, some enzymatic properties and subunitstructure. J. Biochem. 85:1405-1414.
1188 CHO ET AL.
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

H2S DEGRADATION BY XANTHOMONAS SP. STRAIN DY44 1189
8. Hansen, T. A., and H. van Gemerden. 1972. Sulfide utilizationby purple non-sulfur bacteria. Arch. Microbiol. 86:49-56.
9. Hirai, M., M. Ohtake, and M. Shoda. 1990. Removal kinetics ofhydrogen sulfide, methanethiol and dimethyl sulfide by peatbiofilter. J. Ferment. Bioeng. 70:334-339.
10. Hirai, M., M. Terasawa, I. Inamura, K. Fujie, M. Shoda, and H.Kubota. 1988. Biological removal of organosulfur compoundsusing a peat biofilter. J. Odor Res. Eng. 19:305-312.
11. Kusai, K., and T. Yamanaka. 1973. The oxidation mechanismsof thiosulphate and sulphide in Chlorobium thiosulphatophilum.Biochim. Biophys. Acta 325:304-314.
12. Moriarty, D. J. W., and D. J. D. Nicholas. 1970. Products ofsulphide oxidation in extracts of Thiobacillus concretivorus.Biochim. Biophys. Acta 197:143-151.
13. Murakami, H., T. Okada, T. Niida, and S. Katayama. 1987.Solubility of odor gas in water. J. Chem. Eng. Jpn. 13:43-49.
14. Nishihara Environmental Sanitation Research Corp. October1982. Japanese patent 57,170,181.
15. Oren, A., and E. Padan. 1978. Induction of anaerobic, photoau-totrophic growth in the cyanobacterium Oscillatoria limnetica.J. Bacteriol. 133:558-563.
16. Schook, L. B., and R. S. Berk. 1978. Nutritional studies withPseudomonas aeruginosa grown on inorganic sulfur sources. J.Bacteriol. 133:1377-1382.
17. Steinmetz, M. A., and U. Fisher. 1981. Cytochromes of thenon-thiosulfate utilizing green sulfur bacterium Chlorobiumlimicola. Arch. Microbiol. 130:31-37.
18. Sublette, K. L., and N. D. Sylvester. 1987. Oxidation of hydro-gen sulfide by continuous cultures of Thiobacillus denitrificans.Biotechnol. Bioeng. 29:753-758.
19. Sumitomo Jukikai Envirotech. September 1983. Japanese patent58,152,488.
20. Suylen, G. M. H., G. C. Stefess, and J. G. Kuenen. 1986.Chemolithotrophic potential of a Hyphomicrobium species,capable of growth on methylated sulphur compounds. Arch.Microbiol. 146:192-198.
21. Tanji, Y., T. Kanagawa, and E. Mikami. 1989. Removal ofdimethyl sulfide, methyl mercaptan, and hydrogen sulfide byimmobilized Thiobacillus thioparus TK-m. J. Ferment. Bioeng.67:280-285.
22. Then, J., and H. G. Truper. 1983. Sulfide oxidation in Ec-tothiorhodospira abdelmalekii. Evidence for the catalytic roleof cytochrome c-551. Arch. Microbiol. 135:254-258.
23. Wada, A., M. Shoda, H. Kubota, T. Kobayashi, Y. Katayama-Fujimura, and H. Kuraishi. 1986. Characteristics of H2S oxidiz-ing bacteria inhabiting a peat biofilter. J. Ferment. Technol.64:161-167.
24. Wagner, C. D., W. M. Riggs, L. E. Davis, J. F. Moulder, andG. E. Muilenberg. 1978. Handbook of X-ray photoelectronspectroscopy, p. 56. Perkin-Elmer Corp., Eden Prairie, Minn.
25. Zhang, L., M. Suzuki, M. Terasawa, M. Hirai, and M. Shoda.1990. A long-term experiment of a pilot-scale peat biofilter toremove sulfur-containing gases from a waste water treatmentplant. J. Odor Res. Eng. 21:1-9.
VOL. 58, 1992
on June 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from