cytotoxicity of bile salts against biliary epithelium: a study in isolated bile ductule fragments...
TRANSCRIPT

Cytotoxicity of Bile Salts Against Biliary Epithelium: A Study inIsolated Bile Ductule Fragments and Isolated Perfused
Rat Liver
ANTONIO BENEDETTI,1 DOMENICO ALVARO,3 CRISTINA BASSOTTI,1 ALESSANDRO GIGLIOZZI,3 GIANNA FERRETTI,4
TIZIANA LA ROSA,3 ANTONIO DI SARIO,1 LEONARDO BAIOCCHI,3 AND ANNE MARIE JEZEQUEL2
hepatocytes showed prominent subcellular damage with CAWe evaluated cytotoxic effects of different unconjugatedand CDCA, and profound alterations of the canalicular mem-and glycine- and taurine-conjugated bile salts (BS) against bilebrane with LCA and tauro-LCA. We have shown that, in vitro,duct epithelial cells in isolated bile ductule fragments andBDE cells are not damaged by taurine- or glycine-conjugatedisolated perfused rat liver. Ultrastructural morphometric stud-BS, but they are very sensitive to cytotoxicity of hydrophobicies were performed in polarized rat bile ductule fragmentsunconjugated BS. Such sensitivity is not present in the wholeexposed in vitro to increasing concentrations (10-100 mmol/liver, probably because of the specificity of BS transport pro-L) of lithocholate (LCA), deoxycholate (DCA), chenodeoxy-cesses, the microvascular architecture of the bile ductal sys-cholate (CDCA), cholate (CA), ursodeoxycholate (UDCA),tem, and the presence in bile of a physiological surfactant,their taurine-conjugates, and glycoconjugates of cholic (GCA)such as phospholipids. (HEPATOLOGY 1997;26:9-21.)or chenodeoxycholic acid (GCDCA) for 20, 30, or 75 minutes.
To evaluate the cytotoxicity of unconjugated hydrophobic bilesalts against biliary epithelium (BDE) in the whole liver, livers
The cytotoxicity of bile salts (BS) against different cell types,were isolated from rats with impaired taurine-conjugation ca-including hepatocytes,1-4 HepG2 cell,5 erythrocytes,2 mastpacity (b-alanine treatment) and perfused for 70 minutes withcells,6 and gastric and intestinal cells,7 has been extensively2 mmol/min LCA (n Å 6), CDCA (n Å 6), CA (n Å 6), orinvestigated in vitro. In general, the cytotoxicity increases with0.5 mmol/min tauro-LCA (n Å 4). In isolated bile ductuleincreasing BS hydrophobicity (unconjugated ú glycoconju-fragments, hydrophobic unconjugated bile salts (LCA, CDCA,gated ú taurine-conjugated; monohydroxy- ú dihydroxy- úDCA) induced a marked damage of intracellular organelles,trihydroxy-BS). The position of hydroxy groups is also verymainly mitochondria. The damage started at a concentrationimportant, because the 7b-dihydroxy BS, ursodeoxycholateof 10 mmol/L and became prominent at concentrations higher(UDCA), shows no cytotoxicity, and is instead protective.8than 50 mmol/L. No damage of the apical and basolateralStudies in vivo have also shown that the capability of inducingmembrane was seen and tight junctions appeared intact.hepatocellular damage correlates with the hydrophobicity ofUDCA, taurine and glycoconjugated bile salts failed to induceadministered BS.9-13 In light of these findings, current opinionany evident ultrastructural alteration. In taurine-depleted iso-concurs that in the course of cholestatic disorders, retentionlated livers, perfused with LCA, CDCA, or CA, bile ductof hydrophobic BS could have a role in the development ofepithelial cells showed no evidence of intracellular damage,hepatocellular damage. Although speculative, a direct, patho-despite the increased biliary excretion of unconjugated BS.genetic role of hydrophobic BS in human chronic cholestaticMarked alterations of the apical cell membrane were seenliver diseases has been proposed.2,9-12
only in livers perfused with LCA and in isolated segments ofBile duct epithelial (BDE) cells lining the biliary tree arethe biliary epithelium. In contrast with biliary epithelium,
continuously exposed, at their apical site, to millimolar con-centrations of BS.9-12 Presumably, damage to the biliary treeduring bile flow from the site of formation to the duodenum isAbbreviations: BS, bile salt; UDCA, ursodeoxycholate; BDE, bile duct epithelial;avoided by endogenous protective factors or by a constitutiveIPRL, isolated perfused rat liver; TCA, taurocholate; TDCA, taurodeoxycholate;
TCDCA, taurochenodeoxycholate; TLCA, taurolithocholate; CA, cholate; DCA, deoxy- resistance of BDE cells to detergents. One mechanism of pro-cholate; CDCA, chenodeoxycholate; LCA, lithocholate; GCA, glycocholate; GCDCA, tection is the conjugation of BS with taurine or glycine beforeglycochenodeoxycholate; TLC, thin-layer chromatography; HPLC, high-pressure liquid bile excretion, because conjugated BS, especially taurine con-chromatography.
jugates are significantly less cytotoxic than parent unconju-From the 1Department of Gastroenterology, University of Ancona, Ancona, Italy;2Institute of Experimental Pathology, University of Ancona, Ancona, Italy; 3II Depart- gates.1,3 Biliary phospholipids represent an additional physio-ment of Gastroenterology, University ‘‘La Sapienza’’ of Rome, Rome, Italy; and 4Institute logical protective factor against BS cytotoxicity, as recentlyof Biochemistry, University of Ancona, Ancona, Italy. emphasized by observations that in mdr2-mice the lack of
Received March 10, 1996; accepted February 6, 1997.phospholipids in bile causes a destructive damage of bileA. Benedetti and D. Alvaro are both first coauthors.ducts.14 However, although BDE cells are in a condition suchSupported by Consiglio Nazionale delle Ricerche (Progetto Finalizzato FATMA,
94.00527PF41) and by Ministero dell’Universita e della Ricerca Scientifica e Tecnolo- that they are continuously exposed to very high concentra-gica (Progetto Nazionale Cirrosi). tions of BS, studies exploring their sensitivity to the cytotoxic
Address reprint requests to: A. Benedetti, M.D., Department of Gastroenterology, effects of different BS are lacking. This is further emphasizedUniversity of Ancona CP 68, 60020 Torrette, Ancona, Italy. Fax: 39-71-2206044.
by the fact that BDE cells are the primary target in the groupCopyright q 1997 by the American Association for the Study of Liver Diseases.0270-9139/97/2601-0003$3.00/0 of chronic cholestatic liver diseases recently classified as the
9
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

10 BENEDETTI ET AL. HEPATOLOGY July 1997
vanishing bile duct syndrome.15 The recent development ofinnovative techniques to isolate sufficiently pure BDE cellsor polarized bile duct or ductule fragments,16-21 has led toconsiderable progress in the biology of the biliary epithelium.
This study aimed to investigate, at the ultrastructural level,the cytotoxicity of BS with different hydrophobicity, againstBDE cells. The study was performed using an in vitro modelof polarized bile ductule fragments exposed to different un-conjugated, taurine- and glycine-conjugated BS. Becausethese experiments showed cytotoxicity only for unconju-gated BS, we next evaluated whether unconjugated BS arecytotoxic against biliary epithelium also in the whole liver.For this purpose, we performed experiments where isolatedperfused rat livers (IPRL) obtained from rats with impairedhepatic tauroconjugation capacity (taurine-depleted rats)
FIG. 1. Low-power micrographs showing specimens of isolated bilewere perfused with different unconjugated BS.ductules after staining for g-glutamyl transpeptidase. Bar Å 10 mm.
MATERIALS AND METHODS
and 19,16 4-mm cryostat sections were cut and fixed in absoluteAnimals. Sprague-Dawley rats, CD strain (Charles River, Como, acetone for 10 minutes.
Italy) weighing 250 to 400 g, were maintained on Good Laboratory Biliary fragments were then gently and finely minced with scissorsPractice diet (Nossan, Corezzana, MI, Italy) in pellets and given under a light microscope, plated on glass coverslips, precoated withwater ad libitum. Rats were housed in plastic cages with a wire the extracellular biomatrix Matrigel (Collaborative Research, Lex-bottom mesh providing isolation from cage bedding and were ex- ington, MA) diluted 1:1 with L-15 HEPES medium, and incubatedposed to a 12-hour light/dark cycle. Study protocols respected insti- in bicarbonate containing medium (alpha-MEM medium, GIBCO,tutional guidelines. Grand Island, NY) up to 36 to 48 hours.
Chemicals. Bovine serum albumin (fraction V), phosphatidylcho- For light microscopy, fragments of cultured bile ductules wereline, cholesterol, sodium salts of taurocholate (TCA), taurodeoxycho- fixed with 4% buffered formaldehyde followed by inclusion in paraf-late (TDCA), taurochenodeoxycholate (TCDCA), taurolithocholate fin. Four-mm sections stained with hematoxylin-eosin were exam-(TLCA), cholate (CA), deoxycholate (DCA), chenodeoxycholate ined under an Olympus-light microscope equipped with an auto-(CDCA), lithocholate (LCA), glycocholate (GCA), glycochenodeoxy- matic image analyzer system (Olympus Cue-3 VS [Olympus Opticalcholate (GCDCA), paraformaldehyde, b-alanine, and collagenase A Co., Ltd., Tokyo, Japan]).were obtained from Sigma Chemical (St. Louis, MO). Standards of The mean ductular diameter of isolated segments of bile ductulesb-muricholic acid and tauro-b-muricholic acid were kindly provided was 19 { 5 mm (range 10-28 mm) and the epithelium consisted ofby Dr. Alan Hofmann, San Diego, CA. Standard of glyco-b-muricholic 2 to 10 lining cells.acid was kindly provided by Dr. A. Cantafora, Rome, Italy. Glutaralde- Most of the cells of the isolated intrahepatic bile ductule fractionhyde was obtained from Fluka, Chemica-Biochemika (Buchs, Switzer- stained positively for GGT (81 { 1.9%, range 76-87%) (Fig. 1) andland), sodium cacodylate from Farmitalia Carlo Erba (Milan, Italy), for cytokeratins 7 and 19 (81 { 2.0%, range 76%-88%), establishedand sucrose from Merck (Darmstadt, Germany). All other chemicals markers for biliary epithelial cells.were the purest available commercially. For electron microscopy, fragments were fixed with 1% osmium
Isolation and Characterization of Intrahepatic Bile Ductules. Intrahepa- tetroxide in 0.1 mol/L phosphate buffer, pH 7.4, followed by dehy-tic bile ductules were isolated by collagenase digestion and mechani- dration and embedding in Epon/Araldite (Epon, Electron Micros-cal separation of the biliary tree from parenchymal tissue, as pre- copy Sciences, Fort Washington, PA; Araldite, Fluka Chemika-Bio-viously described.16,17 Briefly, rats were anesthetized with chemika, Buchs, Switzerland). Ultrathin 60-nm sections wereintraperitoneal sodium thiopental (Farmotal, 100 mg/Kg body examined under the electron microscope, after staining with leadweight). The portal vein was cannulated with a 16-gauge cannula citrate.and perfused in situ for 10 minutes with a Ca//, Mg//-free Hank’s Morphological Studies on Biliary Fragments. The cultured biliarysolution supplemented with bovine serum albumin ( 1 g/L, Sigma fragments were incubated for 20, 30, or 40 minutes at 377C in L-Chemical Co.) and heparin (2,000 U/L, Heparin Vister, Parke-davis, 15 medium containing 10 (n Å 5), 50 (n Å 5), or 100 mmol/L (nMilan, Italy) (pH 7.2) at 377C. The liver was then perfused in a Å 5) of the following BS: CDCA (n Å 5), TCDCA (n Å 5), DCArecirculating mode at 377C for 20 minutes with Ca//, Mg//-con- (n Å 5), TDCA (n Å 5), LCA (n Å 5), TLCA (n Å 5), CA (n Åtaining L-15 medium (GIBCO, Grand Island, NY) to which colla- 5), TCA (n Å 5), UDCA (n Å 5), TUDCA (n Å 5), GCA (n Å 5),genase A (360 U/L, Sigma Chemical Co.), HEPES (25 mmol/L, and GCDCA (n Å 5). Different BS were diluted in L-15 mediumSigma Chemical Co.), and bovine serum albumin (1 g/L) (pH 7.4) or in dimethyl sulfoxide (10 mL) and then in L-15 medium, ashad been added. When the capsula began to detach, the perfusion appropriate. Paired controls were exposed to the dilution mediumwas stopped and the liver was immersed in ice-cold modified L-15 without BS.medium. Portal tissue containing the intrahepatic biliary tree was Similar experiments were also performed (n Å 4 for each BS) inseparated mechanically from the parenchymal tissue and from other the presence of liposomes made of phosphatidylcholine and choles-cell types by shaking in cold L-15 medium and gentle combing with terol. Small fragments were fixed at the similar time intervals anda fine-tooth comb. The biliary tree was then placed in a beaker prepaqred for electron microscopy. After staining with lead citrate,containing L-15 medium to which collagenase, pronase and DNAase 60-nm thick sections were examined under the electron microscope.had been added; the beaker was shaken twice at 377C for 30 min- Liposome Preparation. Aliquots of phosphatidylcholine (dipalmi-utes. toyl) and cholesterol in chloroform-methanol were placed in a vial.
Part of the material was embedded in Tissue Tek (Triangle Bio- Organic solvents were evaporated under a stream of nitrogen. Themedical Sciences, Durham, NC), snap-frozen in isopentane pre- residue was resolubilized in chloroform-methanol (2:1, vol/vol).cooled in liquid nitrogen, and stored at 0707C for histochemical The same procedure was repeated and the mixture was lyophilized
to complete solvent removal. Multilamellar liposomes were formedstudies. To show g-glutamyl transpeptidase22,23 and cytokeratin 7
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

HEPATOLOGY Vol. 26, No. 1, 1997 BENEDETTI ET AL. 11
by resuspending lipids, at a higher temperature than the main phase were stained with lead citrate and examined with Philips EM 201electron microscope (Philips, Eindoven, the Netherlands). Observa-transition (427C for dipalmitoyl-phosphatidylcholine) in 0.1 mol/L
NaCl, Tris-HCl 20 mmol/L, pH 7.4. Total liposomal phospholipid tions were made by two expert morphologists who were unawareof the treatment.and cholesterol concentrations in the supernatant was assayed by
phosphate determination after perchloric acid extraction. The cho- Analytical Methods. Bile samples were collected in preweighedtubes and the amount evaluated gravimetrically. Total biliary BSlesterol/phosphatidylcholine molar ratio of multilamellar liposomes
was 0.424 and the final concentrations of phosphatidylcholine and in bile were measured by the 3a-hydroxysteroid dehydrogenasemethod.31 Total phosphatidylcholine was measured by choline oxi-cholesterol were 1 mmol/L and 0.4 mmol/L, respectively.
Quantitative Morphometric Analysis. Morphometric studies were dase method.32
BS composition of bile was analyzed, in 10- to 70-minute-pooledperformed in a coded fashion on ultrathin sections by two indepen-dent observers. The reliability of the morphometric analysis was bile samples, by thin-layer chromatography (TLC) and high-pres-
sure liquid chromatography (HPLC). For TLC analysis, 50 mL ofevaluated as ‘‘overall agreement,’’ i.e., the proportion of pictures forwhich observers agreed on quantitative data. Three different blocks bile was deposited on two distinct silica gel 60 F254 plates (0.25
mm thickness; 10 1 20 cm; Merck, Darmstadt, Germany) that werefrom three different samples were examined for each animal. Fivepictures of adjacent nonoverlapping fields of each sample were run in parallel with the solvent system isoamyl acetate/propionic
acid/n-propanol/water, (4:3:2:1; vol/vol/vol/vol).33 TCA, TCDCA,taken at an initial magnification of 17,000 and printed at a finalmagnification of121,000. On each print, a 1-mm-wide area beneath TLCA, T-b-muricholic acid, TLCA-sulfate, GCA, GCDCA, GLCA,
glyco-b-muricholic acid, glycolithocholate-sulfate, CA, CDCA,the basolateral or the apical membranes was delimited, leaving anintermediate (juxtanuclear) region that included all the remaining LCA, LCA-sulfate, and b-muricholic acid were used as standards.
Spots were visualized on one plate by spraying with sulfuric acid.cytoplasm.The surface density (SD Å mm2/mm3 of cytoplasm) of plasma Corresponding spots were scraped off the second plate run in paral-
lel, eluted with methanol, and quantified by 3a-hydroxysteroid de-membrane and microvilli (apical membrane) was evaluated by lineintersection counting. The volume density (VD Å % of cytoplasm hydrogenase. The recovery of BS was calculated by comparison with
total BS concentration measured before TLC. By using the samevolume) of mitochondria and of the Golgi network was measuredby point-counting technique.25 procedure, more than 95% of authentic standards were recovered.
To identify the presence of BS glucuronides, another TLC system,34All morphometric data were analyzed statistically using Student’spaired or unpaired t test, as appropriate. A value of P õ .05 was which permits the separation of glucuronides from other conju-
gates, was used (ethanol/ethyl acetate/ammonia, 3:3:1, vol/vol/vol).considered statistically significant.Taurine Depletion. Depletion of taurine stores was obtained, as pre- The plates were then sprayed with naphthoresorcinol (0.2% in etha-
nol/phosphoric acid 9:1, vol/vol), a specific reagent for glucuro-viously described,26-28 by administration of b-alanine (20 mmol/d,orally) in distilled water for 2 consecutive days. On the third consec- nide.35 Taurine and glycine conjugates of BS were also identified
and quantified by reversed-phase HPLC.36utive day, liver was isolated and perfused as follows.IPRL System. Animal surgery, liver isolation and perfusion tech- Lactic dehydrogenase and aspartate transaminase in the perfusate
were determined using standard kit methods.nique, and solution preparation were performed as previouslydescribed.29,30 Briefly, under sodium pentobarbital anesthesia (50 Oxygen liver consumption was calculated as the difference be-
tween oxygen concentration measured (Schott-Seraete, Italscienti-mg/kg/body wt, intraperitoneally), the bile duct was cannulatedwith a PE-10 tubing (Clay Adams, Parsippany, NJ). The pancre- fica, Genova, Italy) in influent and effluent perfusate.
Bicarbonate biliary concentration was analyzed immediately afteratic-duodenal branch of the portal vein was ligated and the portalvein cannulated with a 14-gauge Teflon intravenous catheter. The collection in 25 to 30 mL bile samples by using a microgasameter
(AVL, Medical Instruments, Schaffahusen, Switzerland).liver was immediately perfused at a constant flow rate of 25 mL/Statistical Analysis. Data are presented as means { SE. Statisticalmin (377C) with CO2 /O2 (5%/95%) oxygenated Krebs-Ringer bi-
comparison was made using repeated ANOVA.carbonate buffer containing 5.5 mmol/L glucose and 200 U hepa-rin per 100 mL. The diaphragm was opened, the suprahepatic
RESULTSvena cava cannulated, and the liver transferred into a heated per-fusion chamber and then perfused in a single-pass system (nonre- Morphological Studies of Isolated Bile Ductules. Isolated bilecirculating) with CO2 /O2 (5%/95%) oxygenated Krebs-Ringer bi- ductules incubated with hydrophobic unconjugated BScarbonate buffer (pH 7.4), containing 5.5 mmol/L glucose, at a (CDCA, DCA, LCA) presented a good preservation of theflow rate of 3.5 mL/min/g liver. The perfusion buffer was continu- general architecture with intact junctional complexes andously gassed using an artificial lung, warmed at 377C and continu- numerous microvilli surrounding the lumen. The cyto-ously filtered through a 2-mm filter placed in front of the portal plasmic organelles, however, were altered, especially the mi-cannula. Portal pressure, which was continuously monitored dur-
tochondria, which appeared swollen with rarefaction of theing the experiments, averaged 10 cm H2O.matrix, globular shapes, and loss or shortening of cristae inExperimental Design. IPRL were perfused from 0 to 70 minutesall the cells (Fig. 2A and 2B). At times, the Golgi apparatuswith 2 mmol/min of CA (n Å 6), CDCA (n Å 6), LCA (n Å 6), orwas also prominent, with evidence of dilatation of the cister-with 0.5 mmol/min TLCA (n Å 4), dissolved in the perfusate at anae (Fig. 3A). The nuclei were normal in appearance (Fig.final concentration of 50 mmol/L for CA, CDCA, or LCA and 12.5
mmol/L for TLCA. Control experiments in taurine-depleted IPRL 3A).perfused without BS in the perfusate were also performed. The treatment of bile ductule samples either with taurine-
Morphological Studies on IPRL. Fragments of liver tissue were taken or glycine-conjugated hydrophobic BS or with hydrophilicat the end of the experiment. For light microscopy, the fragments unconjugated BS (CA, UDCA) did not show significant modi-were fixed in 4% formaldehyde and embedded in paraffin. Sections fications of the general architecture of the biliary epitheliumwere stained with hematoxylin-eosin for routine examination. For with respect to controls (Fig. 3C).ultrastructural studies, small fragments were fixed for 1 hour in
Finally, the incubation of bile ductule samples with mixed0.8% paraformaldehyde/2.5% glutaraldehyde in a 0.1 mol/L cacodyl-liposomes did not prevent morphological damage. Indeed,ate buffer pH 7.4, transferred overnight in a 0.1 mol/L cacodylatethe mitochondrial alterations induced by DCA or LCA (50buffer pH 7.4 containing 0.25 mol/L sucrose. The samples weremmol/L) in the presence of liposomes were similar with re-then post-fixed for 1 hour in 1% osmium tetroxide in a 0.1 mol/spect to paired experiments performed in the absence ofL cacodylate buffer pH 7.6 and embedded in Epon/Araldite after
dehydration with ethanol and propylene oxide. Ultrathin sections liposomes (Fig. 3B).
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

12 BENEDETTI ET AL. HEPATOLOGY July 1997
FIG. 2. (A) Isolated bile ductules exposed to DCA (50 mmol/L) in L-15 medium at 377C for 30 minutes. The micrograph shows the good preserva-tion of the general architecture with intact junctional complexes (arrows). There is, however, evidence of swelling of mitochondria [M] with electronlucent matrix in all the cells present in the field. The nuclei [N] appear normal. L, lumen; Bar Å 1 mm. (B) Isolated bile ductules exposed to CDCA(50 mmol/L) in L-15 medium at 377C for 30 minutes. The general architecture is well preserved with intact junctional complexes (arrows) but allmitochondria [M] are deeply altered, with globular shapes, rarefaction of the matrix, and loss of cristae. Bar Å 1 mm.
Ultrastructural Morphometric Analysis. Table 1 shows the Viability and Bile Secretory Parameters in IPRL From Taurine-Depleted Rats. The parameters of viability of IPRLs from tau-morphometric analysis of cellular structures after incubation
with different BS. As mentioned above, after treatment with rine-depleted rats are reported in Table 2. Oxygen liver con-sumption remained stable in IPRLs perfused without BSs ashydrophobic unconjugated BS, the volume density of mito-
chondria was significantly increased with respect to controls. well as in CA- and LCA-perfused livers. A significant decreasein oxygen liver consumption occurred after 65 minutes perfu-These modifications appeared to be dose- and time-depen-
dent and were observed in a high percentage (92 { 0.8%, sion with CDCA, compared with values measured after 20minutes (Põ .05). On the contrary, oxygen liver consumptionrange 87-98) of mitochondria (Table 1). The minimum con-
centration at which morphological damage was evident was significantly increased after 65 minutes perfusion with TLCA,compared with both values measured after 20 minutes (P õ10 mmol/L.
A dilatation of the cisternae of the Golgi apparatus was .02) and with IPRLs perfused without BS or with CA, CDCA,and LCA (P õ .02). LDH release in the perfusate remainedsometimes evident, but the morphometric data were not sig-
nificantly different from that obtained in controls (Fig. 2A stable in IPRLs perfused without BS. In comparison, LDHrelease in the perfusate significantly increased after 65 minutesand Table 1).
Finally, in bile duct fragments treated with either 50 or perfusion with CA (P õ .05) and more markedly (P õ .02)after perfusion with CDCA, LCA, and TLCA. AST release in100 mmol/L CDCA, DCA, or LCA, the SD of plasma mem-
brane and of microvilli did not appear significantly modified the perfusate remained stable in BS-free and CA-perfusedIPRLs, while it was significantly (P õ .04) increased after 65with respect to controls (Table 1).
The coincubation with hydrophobic BS (DCA or LCA, 50 minutes exposure to CDA, LCA, and TLCA. Portal pressureremained stable during experiments, without significant differ-mmol/L), together with cholesterol/phosphatidylcholine (molar
ratio 0.4) mixed liposomes, did not prevent the ultrastructural ences between the various perfused BS (data not shown).Figure 4 shows bile flow and BS biliary secretion in IPRLsmodifications induced by DCA or LCA alone (Fig. 3B).
On the other hand, morphometric analysis did not show perfused with various BS. Administration of CA and CDCA,in a similar fashion, significantly increased bile flow in com-any dose- or time-dependent structural modification in bile
duct fragments treated with taurine- or glycine-conjugated parison with IPRLs perfused without BS (Põ .01). BS biliarysecretion was significantly (Põ .04) higher during perfusionhydrophobic BS or with the hydrophilic BS (unconjugated
and taurine- or glycine-conjugated) compared with controls with CA than with CDCA. Administration of LCA did notchange bile flow with respect to IPRLs perfused without BS(Table 1 and Fig. 3C).
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

HEPATOLOGY Vol. 26, No. 1, 1997 BENEDETTI ET AL. 13
FIG. 3. (A) Isolated bile ductules exposed to LCA (50 mmol/L) in L-15medium at 377C for 30 minutes. The general architecture of the duct is wellpreserved, with intact tight junctions (arrows) and numerous microvillisurrounding the lumen [L]. The cytoplasmic organelles however are altered,especially the mitochondria [M], which appear swollen with an electronlucent matrix and short cristae in all the cells. The Golgi apparatus [G] isalso prominent, at times, with evidence of dilatation of the cisternae. Thenuclei [N] are normal in appearance. Bar Å 1 mm. (B) Fragment of isolatedbile duct exposed for 30 minutes to LCA (50 mmol/L) in the presence ofphosphatidylcholine/cholesterol liposomes (molar ratio, 0.4). There is agood preservation of the general architecture with absence of cytoplasmicblebs on the basolateral membrane, intact microvilli, and junctional com-plexes (arrows). A diffuse mitochondrial damage is evident: the organelles[M] tend to assume a globular shape with electron lucent matrix and short,peripheral cristae. The aspect is similar to that observed after incubationwith LCA alone (compare with Fig. 3A). N, nuclei. BarÅ 1 mm. (C) Isolatedbile ductules exposed to TLCA (50 mmol/L) in L-15 medium at 377C for30 minutes. The architecture of the ductule is well preserved as shown bythe intact junctional complexes (arrows) surrounding the lumen [L]. Thecytoplasmic organelles such as mitochondria [M] appear normal. N, nucleus.Bar Å 1 mm.
until after 50 minutes of perfusion. Thereafter, bile flow was pared with IPRLs perfused without BS (at least until 50 min-utes perfusion). Administration of TLCA induced an immedi-inhibited by approximately 45% by LCA in comparison with
IPRLs perfused without BS (P õ .05). During LCA adminis- ate (within 15 minutes) marked inhibition of bile flow andBS biliary secretion.tration, BS biliary secretion was lower (P õ .01) than during
perfusion with CA and CDCA and higher (P õ .05) com- The biliary BS composition analyzed by TLC and reversed-
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

14 BENEDETTI ET AL. HEPATOLOGY July 1997
TABLE 1. Morphometric Analysis of Ultrastructural Modifications in Isolated Intrahepatic Bile Ductules Incubated in L-15 Medium / BA or TheirTauro- or Glycoconjugated Derivatives (50 mmol/L) at 377C for 30 Minutes
Plasma MembraneSurface Density Microvilli Surface
Mitochondria Volume Density Golgi Volume Density mm2/mm3 Density mm2/mm3
Bile Acid % of Cytoplasm % of Cytoplasm Cytoplasm Cytoplasm
Controls 8.88 { 0.62 0.14 { 0.007 16.08 { 1.35 0.12 { 0.02LCA 14.13 { 0.45*† 0.14 { 0.005 16.04 { 1.42 0.15 { 0.01TLCA 10.26 { 1.02 0.14 { 0.005 15.99 { 1.14 0.13 { 0.03DCA 14.96 { 1.00*‡ 0.14 { 0.004 14.98 { 1.45 0.15 { 0.01TDCA 9.82 { 0.30 0.14 { 0.007 15.03 { 1.40 0.13 { 0.01CDCA 18.56 { 0.63*† 0.14 { 0.010 16.15 { 1.00 0.17 { 0.01TCDCA 11.01 { 0.68 0.14 { 0.005 16.30 { 0.81 0.13 { 0.01GCDCA 10.98 { 0.71 0.14 { 0.006 16.26 { 0.87 0.14 { 0.03CA 10.23 { 1.00 0.14 { 0.004 16.01 { 1.40 0.15 { 0.02TCA 9.46 { 0.32 0.14 { 0.004 16.28 { 0.95 0.13 { 0.01GCA 10.01 { 0.78 0.14 { 0.003 16.32 { 0.83 0.13 { 0.02UDCA 8.57 { 0.36 0.14 { 0.007 15.89 { 1.15 0.12 { 0.01TUDCA 8.61 { 0.25 0.14 { 0.005 16.09 { 0.55 0.12 { 0.01
* P õ .001 vs. controls (biliary ductules incubated in the culture medium without BA).† P õ .001 vs. the corresponding tauro- or glyco-conjugated BA.‡ P õ .01 vs. the corresponding tauro- or glyco-conjugated BA.
phase HPLC, after pooling 10- to 70-minute bile samples, is reaches a 25.8{ 2.6% of the total BS. No spots correspondingto sulfate derivatives were seen during administration of LCAshown in Table 3. In IPRL perfused without BS, unconjugated
tri-hydroxylated BS account for only 2.7 { 1.1 of total biliary or TLCA. The occurrence of BS glucuronides were evaluatedby using TLC and a solvent system that permits their resolu-BS. In IPRLs perfused with CA, this BS was significantly
secreted as unconjugated, representing 42.5 { 3.8% of the tion with respect to other conjugates. After spraying TLCplates with naphthoresorcinol, a specific stain for glucuro-total BS (average concentration Å 29 mmol/L). On the con-
trary, during perfusion with CDCA and LCA, only 12.5 { nides, we failed to evidence specific spots.Biliary phospholipid concentration was significantly higher1.1% (average concentration Å 4.2 mmol/L) and 5.9 { 0.5%
(average concentration Å 0.97 mmol/L), respectively, of (P õ .02) during perfusion with CA and CDCA than duringperfusion with LCA or no BS (Table 3).these BS were excreted without prior conjugation. CDCA
was mainly secreted in the form of tauro- and glyco-CDCA. Biliary bicarbonate concentration was significantly higherduring perfusion with CDCA (P õ .01) than with the otherHowever, in bile of CDCA-perfused IPRL, two unknownBS or IPRL perfused without BS (Table 3). CA administrationpeaks, with an HPLC retention time comprised between TCAslightly (Põ .05) increased bicarbonate concentration in bileand TCDA, were evidenced (approximately 10% of total bili-compared with IPRL perfused without BS (Table 3).ary BS), indicating partial metabolism of CDCA to more polar
Morphological Studies in IPRL From Taurine-Depleted Rats. Nometabolites. LCA was markedly hydroxylated to b-muri-morphological alterations were seen by light or electron mi-cholic acid that was secreted as taurine- or glycine-conju-croscopy in the liver tissue of taurine-depleted IPRL perfusedgated. Only 19% of LCA appears in bile as taurine- or glycine-
conjugated. When IPRL was perfused with TLCA, this BS without BS.
TABLE 2. Viability Parameters of Taurine-Depleted IPRL Perfused With Different Bile Salts
NO BS CA CDCA LCA TLCA
O2 Liver Consumption (mmol/min/g liver)20 min 1.52 { 0.15 1.59 { 0.08 1.90 { 0.06 1.58 { 0.13 1.90 { 0.1045 min 1.74 { 0.16 1.59 { 0.08 1.49 { 0.11* 1.60 { 0.10 2.56 { 0.11†65 min 1.66 { 0.10 1.61 { 0.09 1.55 { 0.12* 1.63 { 0.11 2.45 { 0.11†
LDH Release (U/L perfusate)20 min 20.7 { 1.4 23.3 { 3.7 33.2 { 4.8 19.9 { 4.8 27.5 { 3.145 min 23.1 { 3.1 34.4 { 4.2* 45.4 { 8.2 68.9 { 4.7† 32.6 { 4.765 min 21.2 { 4.8 32.6 { 4.1* 61.6 { 4.2† 51.5 { 2.6† 43.5 { 5.8†
AST Release (U/L perfusate)20 min 8.3 { 0.6 8.8 { 0.4 9.52 { 0.9 6.2 { 0.5 7.2 { 0.945 min 9.1 { 0.7 8.9 { 0.5 9.95 { 2.2 10.4 { 1.5 9.7 { 0.965 min 9.0 { 0.7 10.3 { 2.8 14.27 { 3.4* 14.9 { 3.4* 12.6 { 1.6*
NOTE. IPRLs were isolated from taurine-depleted rats and perfused with 2 mmol/min CA (n Å 6), CDCA (n Å 6), LCA or with 0.5 mmol/min TLCA(n Å 4) from 0 to 70 minutes or without bile salts (no BS; n Å 6). Data are means { SE.
Abbreviations: CA, cholic acid; CDCA, chenodeoxycholic acid; LCA, lithocholic acid; TLCA, taurolithocholic acid.* P õ .05 vs. 20-minute values.† P õ .02 vs. 20-minute values.
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

HEPATOLOGY Vol. 26, No. 1, 1997 BENEDETTI ET AL. 15
completely occluding the lumen (Fig. 6). The extent anddistribution of alterations varied accordingly to the natureof the various BS and the lobular topography. Maximal dam-age was observed after perfusion with LCA or TLCA. Inaddition, although nonspecific for each BS, the alterationsshowed a preferential pattern, so that the main lesion con-sisted of canalicular damage after LCA and TLCA, whilecytoplasmic vacuoles were more frequently observed afterCDCA or CA infusion. The structural damage affected almostall hepatocytes in the sections including portal tract compo-nents. This was less prominent or even absent in other sam-ples, suggesting a preferential localization in acinar zone 1.Despite the cytoplasmic and canalicular damage, there wasno morphological evidence of reduced junctional complexintegrity in any group. Occasional necrotic hepatocytes wereobserved in all groups, although more easily after CDCA.They showed swollen mitochondria and endoplasmic reticu-lum and appeared isolated or in groups of no more thanthree cells, mostly in juxtaportal areas.
The bile ductules showed a gross preservation of the archi-tecture. However, in isolated segments of the biliary epithe-lium, in liver perfused with LCA, the ductular lumen wasfilled with complex membranous formations, at times nar-rowly apposed to the epithelial cells. At this level profoundalterations of the apical cell membrane were evident, withdisappearance of microvilli and lysis of the apical cytoplasm(Fig. 7). The basolateral membrane appeared structurallypreserved. The subcellular configuration of the neighboringbiliary cells was normal, with integrity of the cytoplasmicorganelles and of the tight junctions.
FIG. 4. Bile flow (A) and bile salt biliary secretion (B) in IPRL isolatedDISCUSSIONfrom taurine-depleted rats and perfused with different bile salts from 0 to
70 minutes. Bile flow was significantly higher (P õ .01) during perfusion In Vitro Studies on Bile Ductule Fragments. We investigatedwith CA (m; nÅ 6, 50 mmol/L) and CDCA (l; nÅ 6, 50 mmol/L), compared
the cytotoxicity of BS with different hydrophobicity againstwith LCA (j; n Å 6, 50 mmol/L), NO BS (l; n Å 6), or TLCA (h; n ÅBDE cells using a recently developed in vitro model of polar-4, 12.5 mmol/L). BS biliary secretion was significantly (P õ .04) higher
during perfusion with CA than with CDCA. Administration of LCA did not ized bile ductule fragments.16,17 These fragments maintainchange bile flow with respect to IPRLs perfused without BS until after 50 their anatomical polarity with a good preservation of theminutes of perfusion. Thereafter, bile flow was inhibited by approximately
basement membrane, an apical pole with numerous micro-45% by LCA in comparison with IPRLs perfused without BS (P õ .05).villi surrounding a ductular lumen and intact junctional com-During LCA administration, BS biliary secretion was lower (P õ .01) than
during perfusion with CA and CDCA and higher (P õ .05) as compared plexes. In addition, these fragments maintain in culture theirwith IPRLs perfused without BS (at least until 50 minutes perfusion). Ad- functional properties because they secrete alkaline fluid inministration of tauro-LCA induced an immediate (within 15 minutes)
the lumen in basal conditions as well as in response tomarked inhibition of bile flow and BS biliary secretion. Data are mean {hormonal stimulation.17 We exposed these fragments to in-SE. For some time points SE are within symbols.creased concentrations of unconjugated, taurine- or glycine-conjugated BS. Our morphological study indicated that cyto-toxicity occurs only with unconjugated hydrophobic BSIn the liver of the animals perfused with BS, the general
architecture appeared well preserved under light microscopy. (CDCA, DCA, LCA). This occurs at the intracellular level,because no evidence of damage to basolateral or apical mem-Occasionally, small necrotic foci were seen in the paren-
chyma with swollen and poorly stained hepatocytes, evidence branes of BDE cells were evident. This could indicate thathydrophobic BS exert their toxicity in relation to their trans-of karyolysis or karyorrhexis, but without associated inflam-
mation or apoptosis. These necrotic foci were more fre- port inside the cell and partitioning or accumulation in theintracellular organelles. This also excludes a direct ‘‘deter-quently observed after infusion with CDCA and mostly local-
ized in lobular zone 1, sometimes in zone 2. The portal areas gent’’ effect of BS bathing basolateral membranes as well ascanalicular membranes after excretion in the ductular lumen.appeared grossly normal.
Under electron microscopy, numerous hepatocytes showed On the other hand, the concentrations of BS used were farfrom their critical micellar concentration values, at which BSsubcellular damage after perfusion with BS. Profound alter-
ations of the canalicular wall were evident, together with the are known to solubilize biological membranes 37,38 and mixedliposomes showed no protection. The damaging effect of un-presence of intracytoplasmic vacuoles. The latter showed an
electrolucent matrix often containing remnants of altered conjugated hydrophobic BS features as a prominent mito-chondrial damage that begins to be evident at a concentrationmitochondria and membrane debris arranged as dense whorls
(Fig. 5). At the apical pole of hepatocytes, the microvilli were of 10 mmol and becomes significant at a concentration of50 mmol/L. In hepatocytes, mitochondrial damage has beenreplaced by irregular, often lamellar projections, at times
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

16 BENEDETTI ET AL. HEPATOLOGY July 1997
TABLE 3. Biliary Bile Salt Composition, Phospholipid, and Bicarbonate Biliary Concentration in Taurine-Depleted IPRLPerfused With Different Bile Salts
NO BS CA CDCA LCA TLCA
BS Concentration (10-70 min. mmol/L) 5.57 { 0.47 69.26 { 2.42 33.7 { 2.76 16.6 { 1.7 6.5 { 0.5BS Composition (10-70 min. molar %)Unconjugated
Tri-hydroxy 2.7 { 1.1 42.5 { 3.8 — 1.2 { 0.2 1.8 { 0.3Di-hydroxy — — 12.5 { 1.1 — —Mono-hydroxy — — — 5.9 { 0.5 —
Taurine-ConjugatedTCA 27.6 { 3.7 26.4 { 3.5 7.2 { 0.3 13.5 { 1.3 20.6 { 2.8T-b-muricholic 34.7 { 4.2 2.8 { 0.6 5.7 { 0.6 41.9 { 5.7 38.6 { 4.3TCDCA — — 45.5 { 5.3 — —TLCA — — — 13.7 { 0.9 25.8 { 2.6
Glycine-conjugatedGCA 13.5 { 3.7 24.5 { 3.2 — 3.0 { 0.4 5.6 { 0.3G-b-muricholic 15.5 { 3.2 1.3 { 0.2 — 12.1 { 2.1 6.3 { 0.4GCDCA — — 16.5 { 2.7 — —GLCA — — — 5.4 { 0.6 —
Phospholipids (mmol/L)20 min. 1.1 { 0.11 2.8 { 0.1† 2.6 { 0.1† 0.7 { 0.3 ND40 min. 0.9 { 0.20 2.7 { 0.2† 2.3 { 0.3† 0.9 { 0.3 ND60 min. 0.6 { 0.13 2.4 { 0.2† 2.2 { 0.3† 1.1 { 0.2* ND
[HCO3-] (mmol/L)20 min. 23.9 { 0.8 27.5 { 0.7* 35.8 { 1.1‡ 24.1 { 0.8 ND40 min. 24.7 { 1.1 27.1 { 0.5* 37.9 { 1.1‡ 24.7 { 0.6 ND60 min. 24.7 { 1.1 28.7 { 0.6* 36.5 { 0.7‡ 24.8 { 0.9 ND
NOTE. IPRLs were isolated from taurine-depleted rats and perfused with 2 mmol/min CA (n Å 6), CDCA (n Å 6), LCA or with 0.5 mmol/min T-LCA(nÅ 4) for 0 to 70 minutes or without bile salts (no BS; n Å 6). Biliary BS composition was analyzed in 10- to 70-minute pooled bile samples. UnconjugatedBS were fractionated by TLC, quantified by 3a-hydroxysteroid dehydrogenase method, and expressed as molar percentage of total BS. Bands correspondingto authentic standards of CA and b-muricholic were considered as tri-hydroxy, to CDCA as di-hydroxy, and to LCA as mono-hydroxy bile salts. Taurine-and glycine-conjugated bile salts were analyzed by reversed phase HPLC. In CDCA-perfused IPRL two unknown peaks with a HPLC retention timecomprised between TCA and TCDA were evidenced (approximately 10% of total biliary BS), indicating partial metabolism of CDCA to more polarmetabolites. Data are mean { SE.
Abbreviations: CA, cholic acid; CDCA, chenodeoxycholic acid; LCA, lithocholic acid; T, taurine-conjugates; G, glycine-conjugates; ND, not determined.* P õ .05 vs. NO BS.† P õ .02 vs. NO BS or LCA.‡ P õ .01 vs. NO BS, CA or LCA.
recently emphasized, as a preferential feature of intracellular on other cell types, including hepatocytes, with regard to thetype of damage (mitochondrial), the relationship with thecytotoxicity of lipophilic BS.4,39,40 It has been described in
isolated human hepatocytes3 incubated with hydrophobic BS, hydrophobicity of unconjugated BS, and finally the lack ofdamage by taurine-conjugated BS.1-3,7,13 In comparison withsuch as CDCA, and also in vivo in the rat.9 In addition,
hydrophobic BS exert toxic effects against isolated mitochon- hepatocytes,1-4 erythrocytes2 or HepG2 cells,5 BDE cells seemto be much more susceptible to the cytotoxic effect of uncon-dria even at a concentration as low as 10 mmol/L.39 However,
while in our bile ductule fragments, evidence of cytotoxicity jugated BS. The sensitivity of BDE cells to BS cytotoxicity inour in vitro model, could be explained by an intracellularstarted at 10 mmol/L, in many different cell types including
hepatocytes, the cytotoxicity of unconjugated hydrophobic accumulation of BS in BDE cells. Two different BS transport-ers have recently been described in cholangiocytes. An active,BS occurs at a concentration higher than 100 mmol/L.1-5,7 In
agreement with many other cell types and with in vivo stud- Na/-independent transport system with a broad specificityfor different unconjugated and amidated BS and that acts inies, conjugation with taurine appears to completely prevent,
in BDE cells, the intracellular cytotoxic effects of hydropho- both directions, has been found at the basolateral pole.42
In the apical pole of cholangiocytes, genetic and functionalbic unconjugated BS.1-3,7,13 Glycine conjugation has also beenreported1,3 to decrease cytotoxicity of hydrophobic BS includ- expression of the ileal Na/-dependent BS transporter has
recently been described.43,44 The kinetic and dynamic param-ing CDCA.1,3 In our study, both GCA and GCDCA failed toinduce morphological damage up to a concentration of 100 eters of these two different transporters are yet to be defined.
In light of all these considerations, our study suggests thatmmol and with a period of exposure up to 40 minutes. Withregard to GCDCA, however, cytotoxic effects including BS accumulated in BDE cells intracellularly, where, as shown
in other cell types, their toxicity is related to hydrophobicityapoptosis,1,3,4,41 have been reported in isolated hepatocyteseven at a concentration of 50 mmol, but with a longer period and as a consequence is higher for unconjugated hydropho-
bic BS. Unconjugated BS are always present in the systemicof exposure than that we used with bile ductule frag-ments.1,3,4 Apart from these considerations concerning circulation because they are continually reabsorbed from the
intestine where they represent the amount of BS undergoingGCDCA, our findings are in keeping with previous studies
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

HEPATOLOGY Vol. 26, No. 1, 1997 BENEDETTI ET AL. 17
FIG. 5. Part of a hepatocyte inliver perfused with CA (2 mmol/min). The micrograph shows the ap-pearance of cytoplasmic vacuolescontaining complex lamellar forma-tions (arrows). The boundaries ofthe vacuole are not clearly outlinedbut rather appear as a gradual struc-tural modification of the cyto-plasmic content. M, mitochondria.Bar Å 1 mm.
deconjugation by the intestinal bacteria.45,46 A concentration tion of unconjugated CDCA in portal blood has been esti-mated to be 18 mmol.45-47 Our finding of evidence of cytotox-of unconjugated cholic acid of 0.88 to 0.47 mmol has been
reported in peripheral plasma of fasting healthy subjects,47 a icity against bile duct fragments for CDCA and LCA, startingat a concentration as low as 10 mmol, provided us with theconcentration which should be 4- to 20-fold higher in the
portal venous blood.45-47 Given the number of anastomoses incentive to verify whether this sensitivity is present only invitro or also in the whole liver. For this reason, we appliedbetween the portal and hepatic arterial vascular system, cho-
langiocytes should also be exposed physiologically to mmol an experimental model, that is the IPRL from taurine-de-pleted rats, where cholangiocytes were exposed continuouslyconcentration of unconjugated BS. In addition, serum con-
centration of unconjugated BS increases in conditions of to unconjugated BS both at their basolateral side and apicalpole. This model provides the absolute certainty that cholan-small bowel bacterial overgrowth, in liver cirrhosis, during
cholestasis or during oral administration of CDCA or UDCA giocytes are exposed to unconjugated BS along the entirelength of the biliary tree.as litholitic agents.48,49 During CDCA therapy, the concentra-
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology
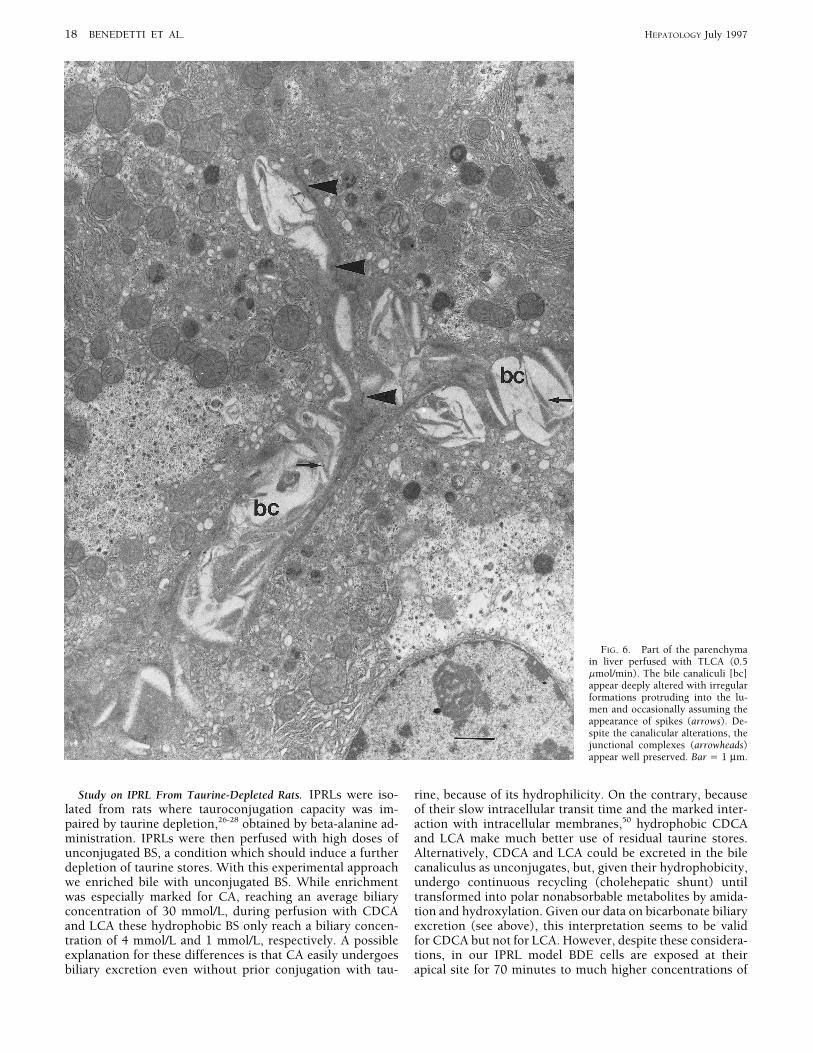
18 BENEDETTI ET AL. HEPATOLOGY July 1997
FIG. 6. Part of the parenchymain liver perfused with TLCA (0.5mmol/min). The bile canaliculi [bc]appear deeply altered with irregularformations protruding into the lu-men and occasionally assuming theappearance of spikes (arrows). De-spite the canalicular alterations, thejunctional complexes (arrowheads)appear well preserved. Bar Å 1 mm.
Study on IPRL From Taurine-Depleted Rats. IPRLs were iso- rine, because of its hydrophilicity. On the contrary, becauseof their slow intracellular transit time and the marked inter-lated from rats where tauroconjugation capacity was im-
paired by taurine depletion,26-28 obtained by beta-alanine ad- action with intracellular membranes,50 hydrophobic CDCAand LCA make much better use of residual taurine stores.ministration. IPRLs were then perfused with high doses of
unconjugated BS, a condition which should induce a further Alternatively, CDCA and LCA could be excreted in the bilecanaliculus as unconjugates, but, given their hydrophobicity,depletion of taurine stores. With this experimental approach
we enriched bile with unconjugated BS. While enrichment undergo continuous recycling (cholehepatic shunt) untiltransformed into polar nonabsorbable metabolites by amida-was especially marked for CA, reaching an average biliary
concentration of 30 mmol/L, during perfusion with CDCA tion and hydroxylation. Given our data on bicarbonate biliaryexcretion (see above), this interpretation seems to be validand LCA these hydrophobic BS only reach a biliary concen-
tration of 4 mmol/L and 1 mmol/L, respectively. A possible for CDCA but not for LCA. However, despite these considera-tions, in our IPRL model BDE cells are exposed at theirexplanation for these differences is that CA easily undergoes
biliary excretion even without prior conjugation with tau- apical site for 70 minutes to much higher concentrations of
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

HEPATOLOGY Vol. 26, No. 1, 1997 BENEDETTI ET AL. 19
FIG. 7. Part of a bile duct in liverperfused with LCA (2 mmol/min).Various alterations of the epitheliallining are evident and show as blebsof the apical pole (large arrows), orcompression of the cells (arrow-heads) by the lamellar material fill-ing the lumen. In some areas, theepithelial lining appears only as athin cytoplasmic sheet (smallarrows), but the basement mem-brane appears intact at this level.Bar Å 1 mm.
unconjugated BS (CA, CDCA, LCA) than those tested in and, predominantly, the canalicular pole after LCA andtauro-LCA administration. In these latter experiments, asvitro. In addition, given the numerous anastomoses between
portal microvasculature and peribiliary plexus,51 the bile duc- previously described,52 the canalicular wall showed markedalterations, with microvilli replaced by irregular, often lamel-tular system should also be exposed at the basolateral pole
to the unconjugated BS perfused via portal vein. Although lar projections, occluding the lumen. There are marked simi-larities between the damage to the apical membrane of hepa-exposed to high concentrations of unconjugated BS, the bili-
ary epithelium showed no evidence of intracellular damage tocytes and biliary cells in LCA-perfused livers, suggesting a‘‘detergent’’ effect exerted by this highly hydrophobic BS inwith any of the perfused BS. Only in LCA-perfused livers
and in isolated segments of the biliary epithelium, there was bile.Our functional study on taurine-depleted IPRL deservesevidence of apical cell membrane damage. Compared with
biliary epithelium, hepatocytes were noticeably damaged further comment. Despite the very high concentrations ofCDCA and LCA administered, no marked cholestasis waseven with the hydrophilic CA. The damage mainly affected
intracellular organelles in CDCA- and CA-exposed IPRLs observed. Rather, CDCA induced the same choleretic effect
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

20 BENEDETTI ET AL. HEPATOLOGY July 1997
as CA. Both CDCA and LA are known to be highly cholestatic Acknowledgment: We would like to express our most sin-cere thanks to Prof. Alan F. Hofmann for helpful discussionswhen administered at doses comparable with those used inand to Luciano Trozzi and Antonella Fava for their excellentthis study, in IPRL or bile-fistula rats with a normal taurocon-technical assistance.jugation capacity.11,35,52 Our findings may certainly be traced
to impaired tauroconjugation. Indeed, when tauro-LCA wasREFERENCESadministered, it induced an almost complete inhibition of
bile flow at a concentration four times lower than that of1. Scholmerich J, Becher MS, Schmidt K, Schubert R, Kremer B, Feldhaus
parent unconjugates. However, to explain the lack of chole- S, Gerok W. Influence of hydroxylation and conjugation of bile saltsstatic effect for LCA and CDCA different reasons could be on their membrane-damaging properties. Studies on isolated hepato-
cytes and lipid membrane vesicles. HEPATOLOGY 1984;4:661-666.considered. As far as CDCA is concerned, we have shown2. Heuman DM, Pandak WM, Hylemon PB, Vlahcevic ZR. Conjugates ofthat HCO0
3 concentration in bile was significantly increased,ursodeoxycholic acid protect against cytotoxicity of more hydrophobic
a finding consistent with the hypothesis that unconjugated bile salts: in vitro studies in rat hepatocytes and human erythrocytes.CDCA, in taurine-depleted IPRL, undergo a cholehepatic HEPATOLOGY 1991;14:920-926.
3. Miyazaki K, Nakayama F, Koga A. Effect of chenodeoxycholic and urso-shunting and thus induce a bicarbonate rich hypercholer-deoxycholic acids on isolated adult human hepatocytes. Dig Dis Sciesis.53-55 The cholehepatic shunting requires, as anatomic1984;29:1123-1130.
substrate, the peribiliary plexus that drains the absorbed BS 4. Spivey JR, Bronk SF, Gores G. Glycochenodeoxycholate-induced lethalback to hepatocytes. This vascular architecture is also effi- hepatocellular injury in rat hepatocytes. J Clin Invest 1993;92:17-24.
5. Concari M, Guicciardi ME, Carubbi F, Bozzoli M, Tagliazucchi M,cient in the IPRL system because of the numerous anastomo-Crema G, Loria P, et al. Effect of S-adenosyl-L-methionine (SAMe)ses between the peribiliary plexus and portal microvascula-reduced hepatotoxicity induced by hydrophobic bile salts (BS) in HepG2
ture.51 Thus, in our system unconjugated CDCA should cell line [Abstract]. J Hepatol 1994;21:S77A.undergo continuous recycling until the molecule is excreted 6. Quist RG, Huong-Thu Ton-Nu, Lillienau J, Hofmann AF, KE Barrett.
Activation of mast cells by bile acids. Gastroenterology 1991;101:446-or transformed into polar, nonabsorbable conjugates. On the456.basis of this interpretation, BDE cells are continuously ex-
7. Ota S, Tsukahara H, Terano A, Hata Y, Hiraishi H, Mutoh H, Sugimotoposed to recycling CDCA, however no ultrastructural fea- T. Protective effect of tauroursodeoxycholate against chenodeoxycho-
late-induced damage to cultured rabbit gastric cells. Dig Dis Sci 1991;tures of damage were seen. As far as LCA is concerned,30:409-416.our findings indicate, in keeping with previous reports,35,52
8. Heuman DM. Hepatoprotective properties of ursodeoxycholic acid [Edi-torial]. Gastroenterology 1993;104:1865-1870.a significant metabolization to trihydroxy polar metabolites,
9. Schmucker DL, Ohta M, Kanai S, Sato Y, Kitani K. Hepatic injurya metabolization that could be enhanced in taurine-depletedinduced by bile salts: correlation between biochemical and morphologi-
liver and perhaps explain the decreased cholestatic effect of cal events. HEPATOLOGY 1990;12:1216-1221.10. Heuman DM, Scott Mills A, McCall J, Hylemon PB, Pandak WM, Vlah-LCA.
cevic R. Conjugates of ursodeoxycholate protect against cholestasis andThus, our study shows that the biliary epithelium is very hepatocellular necrosis caused by more hydrophobic bile salts. In vivostudies in the rat. Gastroenterology 1991;100:203-211.sensitive in vitro but not in the whole liver to cytotoxic effects
11. Drew R, Priestly BG. Choleretic and cholestatic effects of infused bileof hydrophobic unconjugated BS. This indicates that the bili-salts in the rat. Experientia 1978;35:809-811.
ary epithelium is not characterized by an intrinsic resistance 12. Miyai K, Price VM, Fisher MM. Bile acid metabolism in mammals.Ultrastructural studies on the intrahepatic cholestasis induced by litho-to BS cytotoxicity because of a particular composition of cellcholic and chenodeoxycholic acids in the rat. Lab Invest 1971;24:292-membranes. Rather our study suggests the existence in vivo 302.
13. Kitani K, Ohta M, Kanai S. Tauroursodeoxycholate prevents biliaryof mechanisms that avoid intracellular accumulation of hy-protein excretion induced by other bile salts in the rat. Am J Physioldrophobic BS in BDE cells as well as counteracting the deter-1985;248:G407-G417.
gent effect of biliary BS. One of these peculiar mechanisms 14. Smit JJ, Schinkel AH, Oude Elferink RPJ, Groen AK, Wagenaar E, VanDeemter L, Ottenhoff R, et al. Homozygous disruption of the murinecould be the transport processes of BS in BDE cells which, inmdr2 P-glycoprotein gene leads to a complete absence of phospholipidassociation with the peribiliary plexus, ensure a very efficient from bile and to a liver disease. Cell 1993;75:451-462.
15. Sherlock S. Cholestasis: definition and classification of the major clinicalclearance of reabsorbed unconjugated BS. This could haveforms. In: Gentilini P, Arias IM, McIntyre N, Rodes J, eds. Cholestasis.pathogenetic implications in the development of bile ductAmsterdam: Elseevier Science B.V., 1994:3-18.
injury during human cholangiopathies, where injury of the 16. Benedetti A, Marucci L, Bassotti C, Mancini R, Contucci S, JezequelAM, Orlandi F. Tubulo-vesicular transcytotic pathway in rat biliaryperibiliary plexus has been shown to induce damage andepithelium: a study in perfused liver and in isolated intrahepatic bilenecrosis of cholangiocytes.56 In such conditions, the clear- duct. HEPATOLOGY 1993;18:422-432.
17. Mennone A, Alvaro D, Cho W, Boyer JL. Isolation of small polarizedance of toxic reabsorbed BS, even if at very low concentra-bile duct unit. Proc Natl Acad Sci U S A 1995;92:6527-6531.tions, could be impaired and could thus play a role in cholan-
18. Alpini G, Lenzi R, Zhai WR, Liu MH, Slott PA, Paronetto F, Tavolonigiocyte injury. Another important mechanism of defense is N. Isolation of a nonparenchymal liver cell fraction enriched in cells
with biliary epithelial phenotypes. Gastroenterology. 1989;97:1248-represented by biliary phospholipids which, by decreasing1260.CMC and monomeric concentrations of BS,57,58 reduce the 19. Ishii M, Vroman BT, LaRusso NF. Isolation and morphological charac-terization of bile duct epithelial cells from normal rat liver. Gastroenter-surface activity of BS bathing the apical membrane of BDEology 1989;97:1236-1247.cells. To this regard, it is noteworthy that in mdr2-mice, the
20. Roberts SK, Kuntz SM, Gores GJ, La Russo NF. Regulation of bicarbon-lack of phospholipids in bile causes a destructive damage of ate-dependent ductular secretion assessed by lumenal micropuncture
of isolated rodent intrahepatic intact bile ducts. Proc Natl Acad Sci Ubile ducts, 14 which is significantly influenced by the hydro-S A 1995;90:9080-9084.phobicity of excreted BS.59
21. Alpini G, Roberts SK, Kuntz SM, Ueno Y, Gubba S, Podila P, LeSageG, et al. Morphological, molecular and functional heterogeneity of cho-In conclusion, we have shown that BDE cells are verylangiocytes from normal rat liver. Gastroenterology 1996;110:1636-sensitive, in vitro, to cytotoxicity of hydrophobic unconju-1643.
gated BS. Such sensitivity is not present in the whole liver 22. Rutemberg AM, Kim H, Fischbein JW, Hanker JS, Wassenkrug HL,Selingman AM. Histochemical and ultrastructural demonstration ofprobably because of BS transport processes, the microvascu-gamma-glutamyl-transpeptidase activity. J Histochem Cytochem 1969;lar architecture of the bile ductal system, and the presence 17:517-526.
23. Jacobs JMJ, Pretlow TP, Fausto N, Pitts A, Pretlow TG. Separationin bile of a physiological surfactant, such as phospholipids.
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology

HEPATOLOGY Vol. 26, No. 1, 1997 BENEDETTI ET AL. 21
of two populations of cells with gamma-glutamyl-transpeptidase from Ton-Nu H-T, Hofmann AF. Serial quantitative image analysis of uptakeand transport of fluorescent bile acids in polarized biliary epithelialcarcinogen-treated rat liver. J Natl Cancer Inst 1981;66:967-973.
24. Muraca M, Vilei MT, Micromi L, Petrini P. A simple method for the cells. In: Hofmann AF, Paumgartner G, Stiehl A, eds. Bile Acids inGastroenterology. Basic and Clinical Advances. London: Kluwer Aca-determination of lipid composition of human bile. J Lipid Res 1991;
32:371-379. demic Publishers, 1995:191-194.43. Alpini G, Glaser S, Robertson WE, Phinizy IL, Rodgers R, Le Sage GD.25. Weibel ER, Staubli W, Gnagi R, Hess FA. Correlated morphometric and
biochemical studies on the liver cell. J Cell Biol 1969;42:68-91. Genetic and functional expression of the ileal Na-dependent bile acidtransporter and 14 kD cytosolic bile acid binding protein in large but26. Kitani, K, Kanai S. Effect of ursodeoxycholate-induced choleresis in
taurine-deprived and taurine-supplemented rats. Jpn J Physiol 1985;35: not small cholangiocytes from normal rat liver [Abstract]. HEPATOLOGY1996;24:255A.443-462.
27. Shaffer JE, Kocsis JJ. Taurine mobilizing effects of beta alanine and 44. Larazidis K, Pham L, deGroen P, Dawson P, La Russo N. Rat cholangio-cytes express the ileal Na/-dependent taurocholate cotransporter [Ab-other inhibitors of taurine transport. Life Sci 1981;28:2727-2736.
28. Strazzabosco M, Sakisaka S, Hayakawa T, Boyer JL. Effect of UDCA on stract]. HEPATOLOGY 1996;24:351A.45. Hofmann AF. Bile acids. In: Arias IM, Boyer JL, Fausto N, Jakoby WB,intracellular and biliary pH in isolated rat hepatocyte couplets and
perfused livers. Am J Physiol 1991;260:G58-G69. Dchachter D, Shafritz DA, eds. The Liver. Biology and Pathobiology.Third Edition. New York: Raven, 1994:677-718.29. Alvaro D, Benedetti A, Gigliozzi A, Bini A, Della Guardia P, La Rosa T,
Jezequel AM, et al. Functional and ultrastructural features of ethanol/ 46. Carey MC, Duane WC. Enterohepatic circulation. In: Arias IM, BoyerJL, Fausto N, Jakoby WB, Dchachter D, Shafritz DA, eds. The Liver.bile salts interaction in the isolated perfused rat liver. HEPATOLOGY
1995;21:1120-1129. Biology and Pathobiology. Third Edition. New York: Raven, 1994:719-768.30. Alvaro D, Benedetti A, Gigliozzi A, Bini A, Furfaro S, Bassotti C, La
Rosa T, Jezequel AM, Capocaccia L. Effect of brefeldin A on transcytotic 47. Matern S, Jerok W. Pathophysiology of the enterohepatic circulation ofbile acids. Rev Physiol Pharmacol 1979;85:126-204.vesicular pathway and bile secretion: A study on the isolated perfused
rat liver and isolated rat hepatocyte couplets. HEPATOLOGY 1995;21: 48. Tabaqchali S, Booth CC. Bacteria and the small intestine. In: Card WI,Creamer G, eds. Modern Trend in Gastroenterology. London: Butter-450-459.
31. Talalay P. Enzymatic analysis of steroid hormones. Methods Biochem worth, 1970:143-179.49. Angelin B, Ahlberg J, Bjorkhem I, Einarrson K, Ewerth S. Serum levelsAnal 1960;8:119-149.
32. Takayama M, Itoh S, Nagasaki T, Tanimizu I. A new enzymatic method of individual bile acids in portal venous and systemic circulation duringtreatment with chenodeoxycholic acid and cholic acid. In: Paumgartnerfor determination of serum choline-containing phospholipids. Clin
Chim Acta 1977;79:93-98. G, Stiehl A, Gerok W, eds. Biological Effects of Bile Acids. Lancaster:MTP Press, 1979:107-108.33. Hofmann A.F. Thin-layer adsorption chromatography of free and conju-
gated bile acids on silicic acid. J Lipid Res 1962;2:127-128. 50. LeSage GD, Schteingart CD, Hofmann AF. Effect of bile acid hydropho-bicity on biliary transit time and intracellular mobility: a comparison34. Radominska-Pyrek A, Ziminiak R, Chari M, Golunski E, Lester R, Pyrek
St. Glucuronides of monohydroxylated bile acids: specificity of micro- of four fluorescent bile acid analogues [Abstract]. Gastroenterology1994;106:A929.somal glucuronyl-transferase for the glucuronidation site, c-3 configura-
tion, and side chain length. J Lipid Res 1986;27:89-101. 51. Millward GH, Jezequel AM. Normal histology and ultrastrucure. In:Millward-Saddler GH, Wright R, Arthur MJP, eds. Liver and Biliary35. Takikawa H, Ohki H, Sano N, Kasama T, Yamanaka M. Cholestasis
induced by lithocholate and its glucuronide: their biliary excretion and Diseases. Third edition. Philadelphia: Saunders, 1992:12-43.52. Kakis G, Yousef IM. Pathogenesis of lithocholate- and taurolithocholate-metabolism. Biochim Biophys Acta 1991;1081:39-44.
36. Cantafora A, Di Biase A, Alvaro D, Angelico M. Improved method for induced intrahepatic cholestasis in rats. Gastroenterology 1978;75:595-607.measuring the glycine and taurine conjugates of bile salts by high-
performance liquid chromatography with tauro 7a-12a-dihydroxy-5b- 53. Gurantz D, Schteingart CD, Hagey LR, Steinbach JH, Grotmol T, Hof-mann AF. Hypercholeresis induced by unconjugated bile acid infusioncholanic acid as internal standard. J Chromatogr 1987;386:367-370.
37. Lichtenberg D, Robson RJ, Dennis EA. Solubilization of phospholipids correlates with recovery in bile of unconjugated bile acids. HEPATOLOGY1991;13:540-550.by detergents. Biochim Biophys Acta 1983;737:285-294.
38. Armstrong MJ, Carey MC. The hydrophobic-hydrophilic balance of bile 54. Dumont M, Erlinger S, Uchman S. Hypercholeresis induced by urso-deoxycholic acid and 7-Ketolithocholic acid in the rat: possible role ofsalts. Inverse correlation between reverse-phase high performance liquid
chromatographic mobilities and micellar cholesterol-solubilizing bicarbonate transport. Gastroenterology 1980;79:82-89.55. Erlinger S. Hypercholeretic bile acids: A clue to the mechanism? HEPA-capacities. J Lipid Res 1982;23:70-80.
39. Krahenbuhl S, Talos C, Fischer S, Reichen J. Toxicity of bile acids on the TOLOGY 1990;11:888-890.56. Woolf GM, Vierling JM, Disappearing intrahepatic bile ducts. The syn-electron transport chain of isolated rat liver mitochondria. HEPATOLOGY
1994;19:471-479. dromes and their mechanisms. Semin Liver Dis 1993;13:261-275.57. Carey MC. Aqueous bile salt -lecithin cholesterol systems: equilibrium40. Guldutuna S, Elze A, Bhatti S, Imhof M, You T, Zimmer G, Leuschner U.
Ursodeoxycholate (UDC) prevents glutamate dehydrogenase (GLDH) aspects. HEPATOLOGY 1984;4:151S-154S.58. Lindheimer M, Montet JC, Bontemps R, Rouviere J, Brun B. Self- diffu-release of isolated liver mitochondria by chenodeoxycholate
(CDCA)[Abstract]. Gastroenterology 1993;104:A361. sion study of bile salt-monoolein micelles. Determination of the intermi-cellar bile salt concentration. J Chim Phys 1983;80:315-323.41. Roberts LR, Patel T, Bronk SF, Gores GJ. Translocation of cathepsin B
from the cytoplasm to the nucleus contributes to bile salt-induced 59. Oude Elferink RPJ, Groen AH. The role of mdr2 P-glycoprotein inbiliary lipid secretion. Cross-talk between cancer research and biliaryapoptosis [Abstract]. HEPATOLOGY 1996;24:508A.
42. Benedetti A, Marucci L, Di Sario A, Svegliati Baroni G, Schteingart CD, physiology. J Hepatol 1995;23:617-625.
AID Hepa 0073 / 5p22$$1441 06-12-97 15:53:33 hepa WBS: Hepatology