the isolated perfused equine distal limb and …€¦ · pharmacology and toxicology, july 8-12,...
TRANSCRIPT

UNIVERSITY OF VETERINARY MEDICINE HANNOVER
DEPARTMENT OF PHARMACOLOGY, TOXICOLOGY AND PHARMACY
THE ISOLATED PERFUSED EQUINE DISTAL LIMB AND
SYNOVIOCYTE CULTURES AS MODELS FOR
PHARMACOLOGICAL STUDIES
THESIS
SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE
DOCTOR OF PHILOSOPHY (PHD)
IN THE FIELD OF PHARMACOLOGY
AWARDED BY THE UNIVERSITY OF VETERINARY MEDICINE HANNOVER
BY
MAREN FRIEBE
(BIELEFELD)
HANNOVER 2013

Supervisor: Prof. Dr. Manfred Kietzmann
Supervision group: Prof. Dr. Manfred Kietzmann
Prof. Dr. Karsten Feige
PD Dr. Fritz Thorey
1st Evaluation: Prof. Dr. Manfred Kietzmann
Department of Pharmacology, Toxicology and Pharmacy
University of Veterinary Medicine Hannover
Prof. Dr. Karsten Feige
Clinic for Horses
University of Veterinary Medicine Hannover
PD Dr. Fritz Thorey
Department of Orthopedic Surgery
Hannover Medical School
2nd Evaluation: Prof. Dr. Anton Fürst
Equine Hospital, Vetsuisse Faculty
University of Zurich
Date of final exam: April 23, 2013
This project was financially supported by the German Equestrian Foundation (FN,
Warendorf, Germany).

Für meine Eltern


This project has been published in parts:
Publications
FRIEBE M., J. STAHL, M. KIETZMANN (2012)
The isolated perfused equine distal limb as an ex vivo model for pharmacokinetic studies
Journal of Veterinary Pharmacology and Therapeutics
Accepted: July 23, 2012. doi: 10.1111/jvp.12001
FRIEBE, M., S. SCHUMACHER, J. STAHL, M. KIETZMANN (2013)
Synovial distribution of “systemically” administered acetylsalicylic acid in the isolated
perfused equine distal limb
BMC Veterinary Research 9, 56
doi: 10.1186/1746-6148-9-56
Poster presentations at scientific meetings
FRIEBE, M., K. BUNTENKÖTTER, S. SCHUMACHER, M. KIETZMANN
Distribution of acetylsalicylic acid into synovia in an ex vivo model of the equine distal limb
Presented at the 12th International Congress of the European Association for Veterinary
Pharmacology and Toxicology,
July 8-12, 2012; Noordwijkerhout, the Netherlands
FRIEBE, M., J. STAHL, S. SCHUMACHER, M. DÜE, M. KIETZMANN
The isolated perfused equine distal limb as a new ex vivo model for pharmacokinetic studies
Presented at the 17th Congress on Alternatives to Animal Testing,
September 5-8, 2012; Linz, Austria
FRIEBE, M., S. SCHUMACHER, K. BUNTENKÖTTER, M. DÜE, M. KIETZMANN
Distribution of acetylsalicylic and salicylic acid into synovia in an ex vivo model of the equine
distal limb
Presented at the 19th International Conference of Racing Analysts and Veterinarians,
September 17-21, 2012; Philadelphia, PA, USA


TABLE OF CONTENT
VII
Table of content
1 INTRODUCTION ..................................................................................................... 13
2 LITERATURE REVIEW ............................................................................................. 15
2.1 Isolated perfused organs in pharmacological research ................................... 15
2.1.1 Perfusion media and systems ............................................................... 15
2.1.2 Viability parameters .............................................................................. 16
2.1.3 Applications........................................................................................... 16
2.2 Substances investigated using the isolated perfused equine distal limb ......... 19
2.2.1 Corticosteroids ...................................................................................... 19
2.2.1.1 General pharmacology ................................................................... 19
2.2.1.2 Duration of action ........................................................................... 20
2.2.1.3 Betamethasone .............................................................................. 21
2.2.1.4 Mechanism of action – genomic effects ......................................... 22
2.2.1.5 Intra-articular use of glucocorticoids ............................................... 22
2.2.1.6 Pharmacokinetics after intra-articular application ........................... 23
2.2.2 Nonsteroidal anti-inflammatory drugs (NSAIDs) ................................... 25
2.2.2.1 Role of prostaglandins in inflammation ........................................... 26
2.2.2.2 Isoforms of cyclooxygenase ........................................................... 26
2.2.2.3 NSAIDs for treating joint disease .................................................... 27
2.2.2.4 Acetylsalicylic and salicylic acid ..................................................... 30
2.2.2.5 Salicylic acid as a threshold substance .......................................... 31
2.3 Determination of an effective concentration in the joint ................................... 32
2.3.1 Cultured equine synovioctes as a model of inflammation ..................... 32
2.3.2 Role of prostaglandin E2 in joint disease ............................................... 33

TABLE OF CONTENT
VIII
3 THE ISOLATED PERFUSED EQUINE DISTAL LIMB AS AN EX VIVO MODEL FOR
PHARMACOKINETIC STUDIES .................................................................................. 34
3.1 Abstract ........................................................................................................... 35
4 SYNOVIAL DISTRIBUTION OF “SYSTEMICALLY” ADMINISTERED ACETYLSALICYLIC ACID IN
THE ISOLATED PERFUSED EQUINE DISTAL LIMB ........................................................ 36
4.1 Abstract ........................................................................................................... 37
5 PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION ............................... 39
5.1 Abstract ........................................................................................................... 39
5.2 Materials and Methods .................................................................................... 39
5.2.1 Cell culture ............................................................................................ 39
5.2.2 Verification of equine synoviocytes ....................................................... 45
5.2.3 In vitro inflammation model with equine synoviocytes ........................... 47
5.2.4 Statistical analysis ................................................................................. 49
5.3 Results ............................................................................................................ 49
5.3.1 Isolation and culture of synoviocytes .................................................... 49
5.3.2 Verification of synoviocytes with immunocytochemistry ........................ 50
5.3.3 In vitro inflammation model ................................................................... 51
6 GENERAL DISCUSSION .......................................................................................... 55
6.1 Model characteristics ...................................................................................... 55
6.2 Betamethasone ............................................................................................... 57
6.3 Acetylsalicylic and salicylic acid ...................................................................... 58
6.4 Outlook ............................................................................................................ 60
7 SUMMARY ............................................................................................................ 63
8 ZUSAMMENFASSUNG ............................................................................................ 65
9 REFERENCES ....................................................................................................... 67

TABLE OF CONTENT
IX
10 APPENDIX ............................................................................................................ 83
10.1 Cell culture experiments ................................................................................ 83
10.1.1 Cell viability ........................................................................................ 83
10.1.2 PGE2 concentrations ......................................................................... 84
10.2 Maren Friebe’s contribution to the manuscripts ............................................ 85
ABBREVIATIONS
X
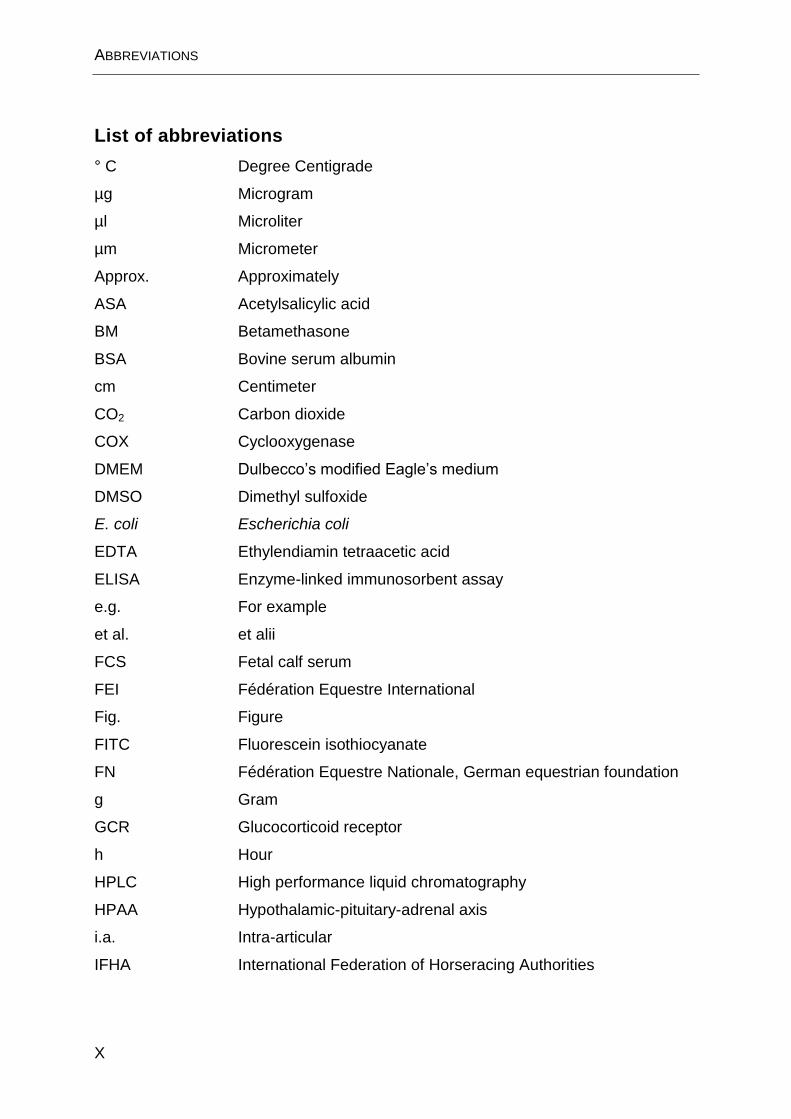
List of abbreviations
° C Degree Centigrade
µg Microgram
µl Microliter
µm Micrometer
Approx. Approximately
ASA Acetylsalicylic acid
BM Betamethasone
BSA Bovine serum albumin
cm Centimeter
CO2 Carbon dioxide
COX Cyclooxygenase
DMEM Dulbecco’s modified Eagle’s medium
DMSO Dimethyl sulfoxide
E. coli Escherichia coli
EDTA Ethylendiamin tetraacetic acid
ELISA Enzyme-linked immunosorbent assay
e.g. For example
et al. et alii
FCS Fetal calf serum
FEI Fédération Equestre International
Fig. Figure
FITC Fluorescein isothiocyanate
FN Fédération Equestre Nationale, German equestrian foundation
g Gram
GCR Glucocorticoid receptor
h Hour
HPLC High performance liquid chromatography
HPAA Hypothalamic-pituitary-adrenal axis
i.a. Intra-articular
IFHA International Federation of Horseracing Authorities

ABBREVIATIONS
XI
i.m. Intramuscular
i.e. That is
i.v. Intravenous
kg Kilogram
L Liter
LDH Lactate-dehydrogenase
LOD Limit of detection
LOQ Limit of quantification
LPS Lipopolysaccharide
mg Milligram
min Minute
mL Milliliter
mm Millimeter
MP Methylprednisolone
n Number of experiments
ng Nanogram
NSAID Nonsteroidal anti-inflammatory drug
PBS Phosphate buffered saline
PG Prostaglandin
pg Picogram
PLA2 Phospholipase A2
p.o. Per os
RMTC Racing Medication and Testing Consortium
s Second
SA Salicylic acid
s.c. Subcutaneous
SD Standard deviation
SEM Standard error of the mean
Tab. Table
TM Trademark
UV Ultraviolet

FIGURES & TABLES
XII
List of figures
FIGURE 1: Generalized ring structure and numbering of corticosteroids .................... 20
FIGURE 2: The arachidonic acid cascade. ................................................................. 25
FIGURE 3: Chemical structures of acetylsalicylic and salicylic acid. ........................... 30
FIGURE 4: Verification of equine synoviocytes Type B. ............................................. 50
FIGURE 5: Viability testing using the MTS-test ........................................................... 52
FIGURE 6: PGE2 concentration in the culture medium after 24 h of stimulation ......... 54
List of tables
TABLE 1: Viability parameters determined in isolated perfused organs. .................... 18
TABLE 2: Different glucocorticoid esters and their duration of release. ...................... 21
TABLE 3: Studies investigating pharmacokinetic properties of glucocorticoids .......... 24
TABLE 4: NSAIDs currently available in Germany ..................................................... 29
TABLE 5: Ingredients of the DMEM basic medium ..................................................... 40
TABLE 6: Protocol for immunostaining of PGP 9.5..................................................... 46
TABLE 7: Protocol for stimulation and treatment of equine synoviocytes. .................. 48
TABLE 8: Results of the modified MTS-test ............................................................... 83
TABLE 9: PGE2 concentration (ng/mL) in the culture supernatant ............................. 84

INTRODUCTION
13
1 Introduction
Lameness caused by inflammatory or degenerative joint conditions is the most
important source of reduced performance in equine athletes (CARON 2005;
GOODRICH and NIXON 2006). Treatment options include administration of
substances directly into the joint or systemically via the blood circulation
(GOODRICH and NIXON 2006).
In order to investigate how long a drug remains in the joint as well as its duration of
action within the joint, pharmacokinetic and pharmacodynamic studies are needed.
In vivo studies that involve living horses, however, are expensive, elaborate and
ethically questionable. An alternative might be ex vivo models of the organs and
tissues of interest, as they have already been successfully established in other fields
of pharmacological research.
It was the aim of this PhD-project to set up appropriate ex vivo alternatives for
pharmacokinetic as well as pharmacodynamic investigations concerning treatment of
the equine joint. The first two studies in this thesis deal with the ex vivo model of the
isolated perfused equine distal limb that was established to investigate the
distribution of substances administered to a horse’s joint either locally (i.e. via intra-
articular application) or systemically. In the third study, a cell culture model of primary
equine synoviocytes was established as a tool for assessing anti-inflammatory
potency of drugs within a joint. As a first application, those concentrations measured
in the isolated perfused equine distal limb were applied to the cells and examined for
their anti-inflammatory properties.


LITERATURE REVIEW
15
2 Literature review
2.1 Isolated perfused organs in pharmacological research
Ever since RUSSELL and BURCH (1959) introduced their concept of the 3Rs -
replacement, reduction, and refinement - for the use of animals in science, various
kinds of ex vivo or in vitro models have been established in many areas of
pharmacological research. Isolated perfused organs have been introduced because
they hold the advantage of resembling the in vivo situation as closely as possible,
including the preservation of anatomic structures (GRONEBERG et al. 2002). While
isolated perfused organs of laboratory animals have been employed in
pharmacological research for many decades (LANGENDORFF 1895; MILLER et al.
1951; WEISS et al. 1959), organs of larger animals have been introduced in the last
30 years as a useful tool mainly in toxicity testing and absorption studies (RIVIERE
and MONTEIRO-RIVIERE 1991). Since they mostly employ organs from animals
killed for unrelated purposes, a reduction of the number of animals used for research
could be achieved (KIETZMANN et al. 1993; VON BAEYER et al. 1997; BÄUMER et
al. 2002; GROSSE-SIESTRUP et al. 2002). Examples include the isolated perfused
porcine skin flap (RIVIERE et al. 1986), equine skin flap (BRISTOL et al. 1991),
bovine udder (KIETZMANN et al. 1993), bovine uterus (BÄUMER et al. 2002), and
porcine forelimb (WAGNER et al. 2003). PATAN et al. (2009) have investigated
effects of long-term extracorporeal blood perfusion on an isolated equine forelimb
with regard to changes in the laminar tissue of the hoof and used this setup as a
model for studying laminitis induction with endotoxin (PATAN-ZUGAJ et al., 2012).
Apart from ethical reasons for the replacement of animal experiments, it also entails
reduction of costs and labor associated with keeping large animals such as horses.
2.1.1 Perfusion media and systems
Generally, perfusion media can be categorized in two major groups: (diluted) blood
and acellular fluids such as Tyrode or Krebs-Ringer solution. The latter group is
easier to supply but frequently needs substitution with components to increase the

LITERATURE REVIEW
16
colloid osmotic pressure within the vessels, e.g. albumin (RIVIERE et al. 1986),
bovine serum albumin (BRISTOL et al. 1991) or dextran (KIETZMANN et al. 1993).
Blood is diluted and needs anticoagulation in order to be usable (BÄUMER et al.
2002; WAGNER et al. 2003; PATAN et al. 2009). It holds the advantage of
resembling the in vivo situation even more closely but is often hard to obtain,
especially if autologous blood is desired. Furthermore, it is more demanding to
handle with regard to oxygen supply and because of blood clotting and hemolysis.
Perfusion systems are either closed, with the venous perfusate being recirculated, or
open, meaning that the venous perfusate is disposed of. The former type consumes
less perfusion medium while it is more challenging to restore the perfusion fluid for
recirculation.
2.1.2 Viability parameters
The monitoring of vitality is essential when working with isolated perfused organs
(WARD and BUTTERY 1979) to ensure sufficient tissue viability and to determine the
maximum timespan for which the organ can be used. To do so, various biochemical,
physiological and morphological endpoints are usually looked at (RIVIERE and
MONTEIRO-RIVIERE 1991). TABLE 1 summarizes viability parameters analyzed in
several studies implementing isolated perfused organs.
2.1.3 Applications
As indicated above, isolated perfused organs have mainly been used for toxicity
testing and absorption studies. Examples include percutaneous absorption studies of
organophosphates, steroids, benzoic acid, caffeine, dexamethasone, benzoyl
peroxide, etofenamate, nitroglycerin, and estradiol as well as mucous membrane
irritation and - inflammation studies (CARVER et al. 1989; WILLIAMS et al. 1990;
KIETZMANN et al. 1993; BÄUMER et al. 2002; WAGNER et al. 2003). It has to be
emphasized that these ex vivo models were not designed to completely replace
animal experiments but to reduce them.

LITERATURE REVIEW
17
Likewise, pathophysiological mechanisms can be studied using isolated perfused
organs: PATAN-ZUGAJ et al. (2012) demonstrated that an exposure to LPS led to
significant laminitis-like changes in the laminar tissue as well as to metabolic
changes in their model of the isolated perfused equine distal limb.

LITERATURE REVIEW
18
TABLE 1: Viability parameters determined in isolated perfused organs.
Viability parameter Indicative of Author
Glucose consumption Glucose metabolism (RIVIERE et al. 1986; BRISTOL et al.
1991; KIETZMANN et al. 1993;
BÄUMER et al. 2002; WAGNER et al.
2003; PATAN et al. 2009)
Lactate production Anaerobic conditions (RIVIERE et al. 1986; BRISTOL et al.
1991; KIETZMANN et al. 1993;
BÄUMER et al. 2002; WAGNER et al.
2003; PATAN et al. 2009)
LDH activity Cell destruction (RIVIERE et al. 1986; KIETZMANN et
al. 1993; BÄUMER et al. 2002;
PATAN et al. 2009)
Blood gas analysis Oxygen supply (PATAN et al. 2009)
Free hemoglobin Hemolysis (WAGNER et al. 2003; PATAN et al.
2009)
Vascular resistance Functionality (RIVIERE et al. 1986; BRISTOL et al.
1991; WAGNER et al. 2003)
Venous perfusate pH Anaerobic conditions (RIVIERE et al. 1986; KIETZMANN et
al. 1993; PATAN et al. 2009)
Skin surface temperature Peripheral blood
supply
(KIETZMANN et al. 1993)
Weight increase Edema formation (BRISTOL et al. 1991; KIETZMANN et
al. 1993; BÄUMER et al. 2002;
PATAN et al. 2009)
Skin fold thickness Edema formation (KIETZMANN et al. 1993)
Histological examination
Morphological
alterations
(RIVIERE et al. 1986; BRISTOL et al.
1991; KIETZMANN et al. 1993;
WAGNER et al. 2003; PATAN et al.
2009)
LDH = Lactate dehydrogenase, Italics: Hemoperfused organs

LITERATURE REVIEW
19
2.2 Substances investigated using the isolated perfused equine distal limb
In the study presented, we chose to investigate the distribution of two representatives
of those substance classes that are commonly used for the treatment of inflammatory
joint disease in horses, namely glucocorticoids and nonsteroidal anti-inflammatory
drugs (LEES 2003). These representatives were betamethasone and (acetyl-)
salicylic acid.
2.2.1 Corticosteroids
2.2.1.1 General pharmacology
Corticosteroids are 21-carbon steroid hormones composed of four rings that are
synthesized in the adrenal cortex from cholesterol. They can be further divided into
two groups: mineralocorticoids, mainly responsible for regulating water and
electrolyte balance, and glucocorticoids, which have an effect on various endocrine
systems in the body.
Glucocorticoids influence the intermediary metabolism by increasing liver glycogen
synthesis and storage, gluconeogenesis, lipolysis and redistribution of lipids. They
may also stimulate the central nervous system and have various effects on the
cardiovascular system, such as increasing myocardial contractions and the
expression of α-adrenergic receptors in the vascular smooth muscle and β-
adrenergic receptors in the myocardium. Glucocorticoids also decrease capillary
permeability and affect the respiratory system by increasing expression of β2–
receptors, which in turn leads to bronchodilatation. Effects on blood cells and
lymphoid tissues include an increase in the number of circulatory erythrocytes,
neutrophils, monocytes and platelets whereas the number of circulatory lymphocytes,
eosinophils and basophils is decreased (HSU 2008).
In therapeutic doses, glucocorticoids exhibit anti-inflammatory and anti-allergic
properties which are linked to the suppression of immune responses: leukocyte
migration and function are suppressed, and plasma and lysosomal membranes are
stabilized, attributable to a decrease in phospholipase A2 (PLA2) activity. As depicted
in FIGURE 2, PLA2 converts phospholipids into arachidonic acid, a precursor of

LITERATURE REVIEW
20
eicosanoids and therefore, synthesis of proinflammatory derivatives such as
prostaglandins, bradykinins, histamines and leukotrienes is suppressed (CREAMER
1999). Since bradykinin and histamine can cause pain by directly stimulating primary
afferent nociceptive fibers, and prostaglandins and leukotrienes have been shown to
sensitize nociceptors, reduction of these mediators also explains the analgesic
effects of glucocorticoids (HAMEED and IHM 2012).
2.2.1.2 Duration of action
Glucocorticoids are traditionally categorized according to the duration of their
hypothalamic-pituitary-adrenal axis - suppression. Hydrocortisone, cortisone, and
prednisolone are representatives of the short-acting (< 24 h) glucocorticoids,
triamcinolone belongs to the intermediate-acting (24 - 48 h) group, and
flumethasone, dexamethasone as well as BM are examples of long-acting (> 48 h)
glucocorticoids (UNGEMACH 2006). However, some studies suggest that the HPAA-
suppression should not be taken as an indicator for duration of pharmacological
effect (KLAUS and HAPKE 1996). Apart from the chemical structure, the formulation
FIGURE 1: Generalized ring structure and numbering of corticosteroids (left). Structure
of cortisol (top right) and betamethasone (bottom right).

LITERATURE REVIEW
21
influences the duration of action: steroid esters are considered prodrugs because the
active moiety is the free alcohol or steroid base resulting from hydrolysis
(AUTEFAGE et al. 1986; WRIGHT et al. 1986). Esterification of the alcohol at C21
significantly determines the drug’s characteristics such as the water/lipid solubility
ratio and the duration of action since tissue esterases cleave the ester to release the
free and therapeutically active base (FERGUSON et al. 2009). Examples for duration
of release are given in TABLE 2.
TABLE 2: Different glucocorticoid esters and their duration of release. Modified from
UNGEMACH (2006) .
Prodrug / Ester Duration of release
Phosphate Rapid
Hemisuccinate
Acetate 2-14 days
Diacetate
Pivalate Weeks to months
2.2.1.3 Betamethasone
The glucocorticoid betamethasone was synthesized through selective modification of
the endogenous glucocorticoid cortisol: an extra double bond was included between
C1 and C2, a methyl group was added at position C16, and a 9-α-fluoro group was
added to C9 (see FIGURE 1). These changes have increased the glucocorticoid
potency in comparison with hydrocortisone 30-fold whereas the mineralocorticoid
potency was reduced to virtually zero (FERGUSON et al. 2009).
BM belongs to the group of long-acting glucocorticoids, with a HPAA - suppression
between 36 – 72 h (UNGEMACH 2006). In Germany, BM is currently available as
Celestovet® (12.0 mg BM-21-acetate and 3.9 mg BM-21-disodiumphosphate) for
therapeutic use in horses and dogs (VETIDATA 2012). However, the preparation
used in this study was Celestan® solubile (5.3 mg BM-21-disodiumphosphate)

LITERATURE REVIEW
22
because it is known that its active agent BM-21-disodiumphosphate is used as a
rapid-acting glucocorticoid in sport horses.
2.2.1.4 Mechanism of action – genomic effects
Like other steroid hormones, glucocorticoids act by altering mRNA synthesis. After
diffusion through the cell membrane, they bind to cell receptors (GCRs), which enter
the cell nucleus and modulate the expression of target genes. One important
mechanism for anti-inflammatory and immunosuppressive effects is the
aforementioned inhibition of PLA2 via the steroid-inducible group of proteins called
lipocortin. PLA2 is the enzyme responsible for the release of arachidonic acid from
cell membranes before its further metabolization by the cyclooxygenase (COX) and
lipoxygenase (LOX) pathways (FERGUSON et al. 2009). Together with their potent
inhibition of the protein NF-κB, which is essential for enhancing inflammatory
cytokine production (BOUMPAS and WILDER 2001), the formation of prostaglandins,
leukotrienes, and platelet activating factor is inhibited. Genomic effects are generally
observed after at least 30 min because of the time needed for GCR activation,
transcription, and translation (FERGUSON et al. 2009). GCRs are present in
neutrophils, lymphocytes, monocytes, and eosinophils (AXELROD 1993).
2.2.1.5 Intra-articular use of glucocorticoids
Glucocorticoids are commonly injected intra-articularly in horses to minimize pain and
inflammation with joint disease (TROTTER et al. 1991). The use of hydrocortisone for
treatment of musculoskeletal conditions in cattle and horses was first described by
WHEAT (1955). Local, i.e. intra-articular, glucocorticoid therapy has the advantage of
directly targeting the inflamed area even with doses much lower compared with
systemic administration (DERENDORF et al. 1986; HARKINS et al. 1993;
FERGUSON et al. 2009). Glucocorticoids frequently used for intra-articular treatment
include different esters of BM, dexamethasone, methylprednisolone, and
triamcinolone (YARBROUGH 2004). Glucocorticoid activity is dependent upon the
presence of a hydroxyl group at C11, therefore, cortisone and prednisone have to be

LITERATURE REVIEW
23
converted to cortisol and prednisolone in the liver in order to be biologically active
(TROTTER 1996). Hence, glucocorticoid preparations for intra-articular use are 11-β-
hydroxyl compounds that do not require biotransformation. Traditionally, the duration
of action after intra-articular injection has been assumed to be inversely correlated
with the glucocorticoid preparation’s solubility in water: water-soluble preparations
such as phosphate and succinate esters are thought to be short-acting, whereas
more lipid-soluble esters such as acetate and acetonide have a longer duration of
action because they are absorbed with a delay (GRAY and GOTTLIEB 1983;
AUTEFAGE et al. 1986). Other determinants for the duration of action include the
rate of hydrolysis of the drug by synovial tissue esterases and the binding affinity of
the glucocorticoid to the steroid receptor in the cytoplasm of target cells (WRIGHT et
al. 1986).
2.2.1.6 Pharmacokinetics after intra-articular application
Results of studies that have investigated the distribution of glucocorticoids after intra-
articular injection to horses are listed in TABLE 3. Some preparations used consist of a
mixture of a rapid-acting solution and a more long acting suspension. However,
because of the high solubility associated with some of the short-acting solutions, it
has been suggested that they might be cleared from the joint very quickly and may
therefore be an unnecessary component (AUTEFAGE et al. 1986; WRIGHT et al.
1986).

LIT
ER
AT
UR
E R
EV
IEW
TABLE 3: Studies investigating pharmacokinetic properties of glucocorticoids after intra-articular administration to horses.
Authors Number
of horses
Joint
injected
Glucocorticoid
and dose
administered
Ester used Detection
time in
plasma
Duration of
cortisol
suppression [h]
Detection
time in urine
(AUTEFAGE
et al. 1986)
5 Tibiotarsal
joint
100 mg MP MP acetate 24 h 3 d n.i.
(LILLICH et al.
1996)
4
4
Tarsocrural
joint
100 mg
100 mg
MP acetate
Isoflupredone acetate
12 h
12 h
n.i. 72 h
22 h
(POPOT et al.
2002)
2 8 mg BM BM phosphate n.i. 36 48 h
(VINE 2006) 2 Carpal joint 11.4 mg BM BM phosphate + acetate 24 h 72 72 h
(SOMA et al.
2006)
6 Carpal joint 200 mg MP acetate > 144 h 144 n.i.
(MACHNIK et
al. 2007)
10 27 - 70 mg BM BM phosphate
BM acetate
n.i. n.i. 35 - 45 d
(SOMA et al.
2011)
6 Carpal joint 0.04 mg/kg Triamcinolone acetonide > 96 h > 96 n.i.
(LÖVENICH
2012)
6
4
Fetlock joint 4 mg BM
34.6 mg BM
BM phosphate
BM phosphate + acetate
48 - 168 h
4 d
48
48 - 384
71 - 145 h
10 d
24
BM Betamethasone, MP Methylprednisolone, n.i. not investigated
LITERATURE REVIEW
25
GLUCOCORTICOID
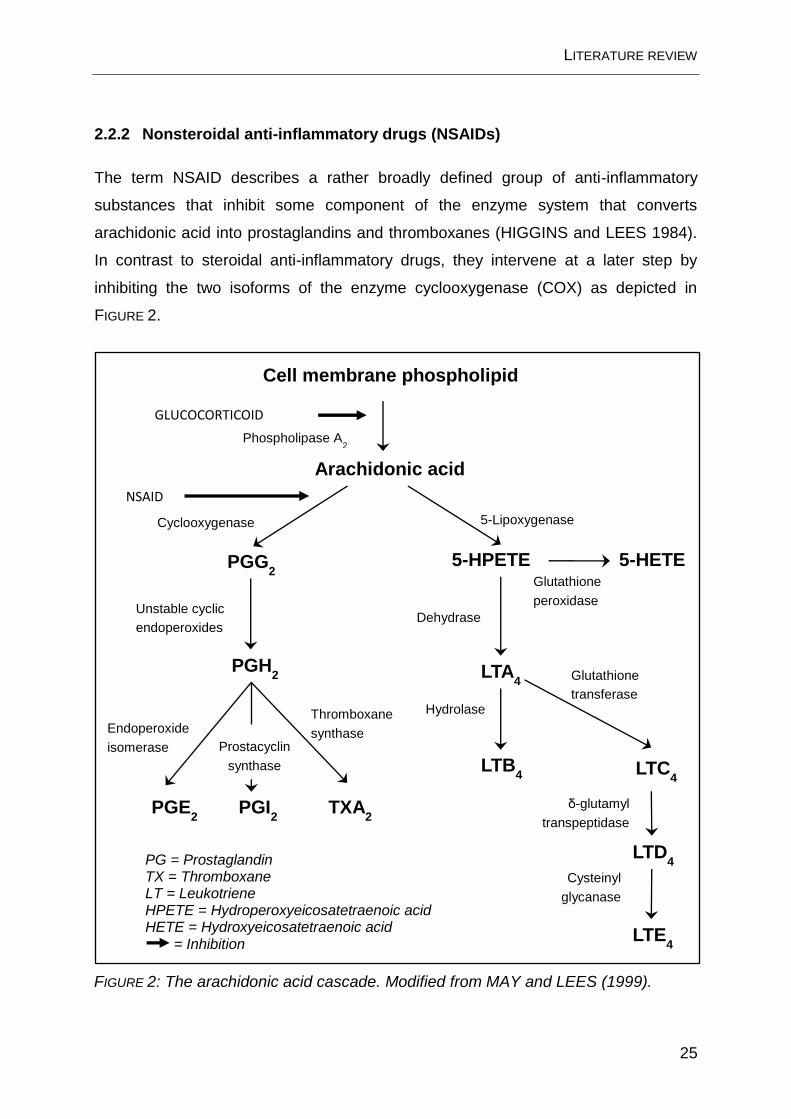
2.2.2 Nonsteroidal anti-inflammatory drugs (NSAIDs)
The term NSAID describes a rather broadly defined group of anti-inflammatory
substances that inhibit some component of the enzyme system that converts
arachidonic acid into prostaglandins and thromboxanes (HIGGINS and LEES 1984).
In contrast to steroidal anti-inflammatory drugs, they intervene at a later step by
inhibiting the two isoforms of the enzyme cyclooxygenase (COX) as depicted in
FIGURE 2.
PGG2
Unstable cyclic
endoperoxides
Prostacyclin
synthase LTB4 LTC
4
LTD4
LTE4
PG = Prostaglandin TX = Thromboxane LT = Leukotriene HPETE = Hydroperoxyeicosatetraenoic acid HETE = Hydroxyeicosatetraenoic acid = Inhibition
Cysteinyl
glycanase
δ-glutamyl
transpeptidase
LTA4
Hydrolase
Dehydrase
Glutathione
transferase
5-HETE 5-HPETE
Glutathione
peroxidase
PGH2
PGE2 PGI
2 TXA
2
Endoperoxide
isomerase
Thromboxane
synthase
Cell membrane phospholipid
Arachidonic acid
Phospholipase A2
Cyclooxygenase 5-Lipoxygenase
FIGURE 2: The arachidonic acid cascade. Modified from MAY and LEES (1999).
NSAID

LITERATURE REVIEW
26
2.2.2.1 Role of prostaglandins in inflammation
Arachidonic acid plays a pivotal role in inflammation as the precursor of the
eicosanoid group of mediators (see FIGURE 2). It is a component of cell membrane
phospholipid, which is released in tissue damage following activation of
phospholipase A2. After its release from the cell membrane phospholipid, arachidonic
acid serves as a substrate not only for cyclooxygenase (COX), of which there are the
two isoforms COX-1 and COX-2, but also for several lipoxygenases. Each enzyme is
part of a cascade, in which the action of further enzymes leads to the formation of
many inflammatory mediators of the eicosanoid family. The mediators are short-lived,
so that continued effect depends on maintained synthesis and release. COX
catalyzes both the formation of PGG2 and then PGG2 conversion to PGH2 via a
peroxidase function. Prostaglandins play an important role as inflammatory mediators
but also participate in synergisms: primary inflammatory mediators such as histamine
and bradykinine stimulate nociceptors to increase the discharge in afferent nerves so
that pain is sensed in spinal and brain centers (LEES 2009). Prostaglandins, e.g.
PGE2, increase intensity and duration of this afferent discharge, causing hyperalgesia
and allodynia (HSU and KANTHASAMY 2008; LEES 2009). PGE2 is an endogenous
pyrogen that leads to upward resetting of the temperature regulating center in the
anterior hypothalamus. Furthermore, prostaglandins enhance edema formation
mainly caused by primary mediators such as histamine and bradykinin by dilating
small arterioles (LEES 2009).
2.2.2.2 Isoforms of cyclooxygenase
COX-1 is a membrane-bound enzyme. It first cyclizes arachidonic acid to form PGG2
and adds a 15-hydroperoxy group to convert PGG2 to PGH2. COX-1 is expressed
constitutively in many tissues and in blood platelets. It is involved in “housekeeping”
functions such as blood clotting, regulation of vascular homeostasis, renoprotection,
gastroprotection, and coordination of the actions of circulating hormones (LEES
2009). COX-2 is both an inducible and a constitutive isoform that is encoded by a
different gene than COX-1, but it has a 60 % structure homology with COX-1 at the

LITERATURE REVIEW
27
amino acid level (HSU and KANTHASAMY 2008). Induction of COX-2 synthesis is
stimulated by proinflammatory cytokines, growth factors, and lipopolysaccharide as
well as mitogens, but it is also constitutively expressed in monocytes, macrophages,
endothelial cells, brain, spinal cord, ovary, uterus, and ciliary body in the eye (LEES
2009). COX-2 is a better competitor than COX-1 for arachidonic acid released within
the cell and produces both pro- and anti-inflammatory prostaglandins at sites of
inflammation (GILROY et al. 1999; LEES 2009). While most data support COX-2 as
the isoform which generates prostaglandins at sites of inflammation, some findings
also indicate a role for COX-1 (SMITH et al. 1998). Many NSAIDs, including
acetylsalicylic acid and indomethacin, are more effective at inhibiting COX-1 than
COX-2 while others, such as ibuprofen and meclofenamic acid, are equipotent at
inhibiting both isoforms (MEADE et al. 1993; MITCHELL et al. 1993). It has been
proposed that this inhibition of the constitutively produced COX-1 is responsible for
many adverse effects of NSAIDs, including gastric ulceration, renal function
impairment and platelet dysfunction (VANE and BOTTING 1995). In an attempt to
decrease these side-effects, new compounds that more selectively inhibit the COX-2
isoform have been developed (LEES 2009). Although numerous COX-2 selective
products are available for use in humans, few of these compounds have been
studied in veterinary species. Further complicating the issue, it appears that enzyme
selectivity is very species dependent (KOLLIAS-BAKER and COX 2004). For
example, carprofen has been reported to be a selective COX-2 inhibitor in the dog
but this selectivity has not been confirmed in the horse (VANE and BOTTING 1995;
RICKETTS et al. 1998).
2.2.2.3 NSAIDs for treating joint disease
The administration of NSAIDs began over 100 years ago with the introduction of
salicylic acid for the treatment of rheumatic disease (BRUNE and BECK 1991).
Nowadays, they are the most commonly prescribed agents for use in horses, with
musculoskeletal pain and inflammation being the most widespread indications for
chronic administration (KOLLIAS-BAKER and COX 2004). Their ability to inhibit the
arachidonic acid cascade has been a mainstay for the treatment of joint disease for

LITERATURE REVIEW
28
many years (FRISBIE 2011). Analgesic and antiphlogistic properties are mediated
through the inhibition of prostaglandin and thromboxane production, which are
intimately involved in pain, altered cartilage metabolism and ongoing inflammation in
damaged joints. It has also been suggested that NSAIDs may play a role in
mediating joint pain at the level of the spinal cord (MALMBERG and YAKSH 1992),
even though this has not yet been investigated in the horse.
Clinically recommended routes of administration include the oral, intravenous,
intramuscular, and subcutaneous injection. Substances used for the treatment of
equine joint disease are listed in TABLE 4.

LITERATURE REVIEW
29
TABLE 4: NSAIDs currently available in Germany for treating musculoskeletal pain in
horses. Note that drugs in italics are not available for food-producing horses.
Drug Route of
administration
Recommended dosage
Firocoxib i.v.
p.o.
0.09 mg/kg as initial treatment
0.1 mg/kg s.i.d. up to 14 d (PLUMB 2008)
Flunixin
meglumine
i.m., i.v.
p.o.
1.1 mg/kg (SOMA et al. 1988; LANDONI and LEES 1995);
0.5 - 2.0 mg/kg (TOUTAIN et al. 1994)
1.1 mg/kg (LANDONI and LEES 1995)
Ketoprofen i.m., i.v. 2.2 mg/kg s.i.d. for 5 days (OWENS et al. 1995)
Meloxicam i.m., i.v., s.c.
p.o.
0.6 mg/kg as initial treatment
0.6 mg/kg s.i.d. up to 14 d (LÖSCHER et al. 2006)
Metamizol i.m., i.v. 5.0 - 22.0 mg/kg (J.J. BERTONE and HORSPOOL 2004)
Phenylbutazone i.v.
p.o.
4.4 mg/kg (LEES et al. 1987b; TOUTAIN et al. 1994; OWENS et
al. 1996); 4.4 mg/kg then 2.2 mg/kg (RAEKALLIO et al. 1997)
4.4 mg/kg b.i.d. for 1 day, 2.2 mg/kg b.i.d. for 4 days; 2.2 mg/kg
s.i.d. for 7 days (TAYLOR et al. 1983; LEES et al. 1986)
Suxibuzone p.o. 6.25 mg/kg b.i.d. for 2 d, then 3.1 mg/kg b.i.d. for 3 d, then 3.1
mg/kg q48h (BISHOP 2005)
Vedaprofen p.o. 2 mg/kg initially, then 1 mg/kg b.i.d. for up to 14 d
(J.J. BERTONE and HORSPOOL 2004)
i.m. intramuscular, i.v. intravenous, s.c. subcutaneous, p.o. orally, s.i.d. once daily, b.i.d.
twice daily, q48h every 48 h

LITERATURE REVIEW
30
2.2.2.4 Acetylsalicylic and salicylic acid
Acetylsalicylic acid is one of the oldest known NSAIDs and the prototype of the
salicylate drugs (COLLIER 1971). It elicits its effect via acetylation and irreversible
inhibition of COX-1, resulting in decreased prostaglandin synthesis and anticoagulant
effects due to the blockade of thromboxane A2 production (HSU and KANTHASAMY
2008). In higher concentrations, it also irreversibly inhibits COX-2 (LEES 2009). ASA
is rapidly deacetylated to SA in virtually all body tissues and blood by endogenous
esterases (KIETZMANN et al. 2002; BROOME et al. 2003).
In horses, the half-life of ASA after intravenous administration is reported to be as
short as 5-10 min (LEES et al. 1987a). Despite this rapid clearance, ASA is very
effective as an antithrombotic agent: relatively small doses can significantly prolong
bleeding times. This prolongation is due to the irreversible inhibition thromboxane A2
in platelets. Since platelets do not contain nuclei, this anticoagulant effect cannot be
reversed until new platelets are formed (KIETZMANN et al. 2002; LEES 2009). As an
analgesic or anti-inflammatory agent, large amounts of ASA have to be administered
because of its very short half-life, rendering its oral application rather impractical
(MAY and LEES 1996). Therefore, ASA is almost exclusively used for the treatment
and prevention of diseases that have arterial thrombosis as a part of their
pathogenesis, e.g. thromboembolic colic or disseminated intravascular coagulation
(BROOME et al. 2003).
FIGURE 3: Chemical structures of acetylsalicylic and salicylic acid.

LITERATURE REVIEW
31
2.2.2.5 Salicylic acid as a threshold substance
Formulating threshold levels is necessary for those substances that a horse is
frequently, but inadvertently exposed to and that may e.g. be a natural part of the
horse’s diet. Without a threshold level, any detection of the substance would have to
be reported as a violation of medication rules. On the other hand, there are also
threshold levels for endogenous substances, an example would be testosterone, for
which different threshold levels exist for geldings and mares (TOUTAIN 2010b). It is
important that the concentration of the substance below the threshold level is not a
threat to the horse’s welfare, does not alter its performance nor compromise the
integrity of the sport (TOUTAIN 2010a).
There are differences between the equestrian federations concerning not only the
concentrations at which threshold levels are fixed but also the threshold substances
as such. An example is dimethyl sulfoxide (DMSO), for which the international
federation for equestrian sports (FEI) and the international federation of horseracing
authorities (IFHA) have established a threshold, whereas this is not the case in the
regulations of the racing medication and testing consortium (RMTC), the national
horseracing authority of the United States (TOUTAIN 2010b).
Since SA naturally occurs in certain plants used as horse feed (such as alfalfa hay), it
may be a component of horse urine (BEAUMIER et al. 1983). Therefore, the rule in
equine sports, according to which neither a drug nor its metabolite may be detectable
in blood or urine during competition, cannot be applied (ANONYMUS 2012). In order
to investigate the normal concentration of SA in horse urine and to formulate
threshold levels, two research groups independently conducted feeding experiments
and analyzed urine samples (BEAUMIER et al. 1983; LAKHANI et al. 2004).
BEAUMIER et al. (1983) postulated a urine threshold concentration of 750 µg/mL,
whereas LAKHANI et al. (2004) suggested the limit of 625 µg/mL. The latter
concentration was adapted by the FEI and the German equestrian federation (FN).
Later, in an attempt to harmonize SA threshold levels between different horse sport
organizations, the FEI has raised the original threshold levels of 625 µg/mL (urine)
and 5.4 µg/mL (plasma) to 750 µg/mL and 6.5 µg/mL, respectively (ANONYMUS
2010). Threshold levels are now aligned with those of the IFHA, whereas the FN has

LITERATURE REVIEW
32
maintained its original concentrations of 625 µg/mL (urine) and 5.4 µg/mL (plasma)
(ANONYMUS 2011).
2.3 Determination of an effective concentration in the joint
Drug effects can be readily assessed using in vitro or in vivo pharmacodynamic
models (KAMERLING and OWENS 1994). Classically, the effective dose of an
antiphlogistic drug is determined from a dose-titration study by use of an appropriate
experimental method of inducing inflammation.
2.3.1 Cultured equine synovioctes as a model of inflammation
In living horses, intra-articular lipopolysaccharide (LPS) injection is an established
model for induction of transient sterile inflammation. It has been used to study e.g.
clinical symptoms, drug pharmacokinetics and the effect of therapeutic intervention
(PALMER and BERTONE 1994a; MORTON et al. 2005; DE GRAUW et al. 2009;
MEULYZER et al. 2009). Also in vitro, cultured equine synoviocytes synthesize large
amounts of PGE2 after stimulation with LPS (LANDONI et al. 1996; MOSES et al.
2001; BRISTON et al. 2009), which is regarded as a surrogate effect of inflammation
(TOUTAIN and CESTER 2004). Because of their importance in the pathogenesis of
osteoarthritis, synoviocytes are considered to be a target for therapeutic activity in
horses (BYRON et al. 2008). This has led to numerous studies investigating anti-
inflammatory properties of substances in the joint, using cultured synoviocytes as an
in vitro model for articular inflammation (LANDONI et al. 1996; FREAN and LEES
2000; BYRON et al. 2008).
Synoviocytes can be divided into two groups: type A cells, or macrophagic cells, and
type B cells, or fibroblast-like cells. The former cells are non-fixed and can
phagocytose cell debris and wastes in the joint cavity; furthermore, they possess the
ability to present antigen. Type B synoviocytes are regarded as proper synoviocytes;
their dendritic processes form a regular network on the luminal surface of the
synovial membrane (IWANAGA et al., 2000). They are responsible for production of

LITERATURE REVIEW
33
both synovial fluid and the extracellular matrix in the synovial intima (KITAMURA et
al. 1999).
2.3.2 Role of prostaglandin E2 in joint disease
Joint disease in the horse is commonly characterized by inflammation of the
synovium (OWENS et al. 1996), involving synoviocytes producing pro-inflammatory
cytokines (DINGLE et al. 1979; SAKLATVALA and DINGLE 1980),
metalloproteinases (OKADA et al. 1992), and eicosanoids (MAY et al. 1989;
LINDSLEY and SMITH 1990; HULKOWER et al. 1993). Of the latter group,
prostaglandin E2 (PGE2) has been identified as an important mediator of
inflammation and hyperalgesia through its enhancement of vascular permeability,
vasodilatory properties, and sensitization of joint nociceptors (HIGGINS et al. 1987;
PALMER and BERTONE 1994b; MAY and LEES 1996). In the joint, synovial tissue
is believed to be the major source of PGE2 in synovial fluid (WITTENBERG et al.
1993). In horses, increased PGE2 concentrations are present in synovial fluid with a
range of joint pathologies (MAY et al. 1994; KIRKER-HEAD et al. 2000). BERTONE
et al. (2001) reported PGE2 to be a good to excellent marker of acute and chronic
joint disease in horses.

THE ISOLATED PERFUSED EQUINE DISTAL LIMB
34
3 The isolated perfused equine distal limb as an ex vivo model for
pharmacokinetic studies
Maren Friebe, Jessica Stahl, Manfred Kietzmann
Department of Pharmacology, Toxicology and Pharmacy; University of Veterinary
Medicine Hannover, Foundation, Hannover, Germany
Accepted for publication in: Journal of Veterinary Pharmacology and Therapeutics
Accepted: July 23, 2012. doi: 10.1111/jvp.12001
Published online: August 23, 2012

THE ISOLATED PERFUSED EQUINE DISTAL LIMB
35
3.1 Abstract
Even though intra-articular treatment plays an important role for the treatment of
joint-related lameness in horses, little is known about pharmacokinetic properties of
substances used. Therefore, an ex vivo model for pharmacokinetic studies was
developed using distal forelimbs of slaughtered horses. The extremity was perfused
with gassed tyrode solution for up to 8 h; tissue viability was confirmed by
measurements of glucose consumption, lactate production and lactate
dehydrogenase activity in the perfusate. Standard criteria for tissue viability had been
determined in preliminary experiments (n=11) which also included histological
examinations of the joint capsule. As the model’s first implementation, the articular
efflux rate of betamethasone (BM), administered as betamethasone disodium
phosphate intra-articularly to the fetlock joint (4 mg BM / joint), was investigated. The
concentration of BM in the venous perfusate of the radial vein was measured by
means of high performance liquid chromatography. The average BM efflux rate per
minute was calculated to be 5.1 µg/min with values ranging from 9 µg/min to
2.9 µg/min. 7.5 h after i.a.-application, 2.3 mg BM had left the joint via the radial vein.
Using this inexpensive setup, the model presented allows studying a variety of
pharmacological topics without the ethical limitations of animal studies.
KEYWORDS: Isolated perfused equine distal limb, betamethasone, intra-articular
injection, articular efflux

SYNOVIAL DISTRIBUTION OF “SYSTEMICALLY” ADMINISTERED ACETYLSALICYLIC ACID
36
4 Synovial distribution of “systemically” administered
acetylsalicylic acid in the isolated perfused equine distal limb
Maren Friebe, Stephan Schumacher , Jessica Stahl, Manfred Kietzmann (2013)
Department of Pharmacology, Toxicology and Pharmacy; University of Veterinary
Medicine Hannover, Foundation, Bünteweg 17, 30559 Hannover, Germany
BMC Veterinary Research 9, 56
doi: 10.1186/1746-6148-9-56

SYNOVIAL DISTRIBUTION OF “SYSTEMICALLY” ADMINISTERED ACETYLSALICYLIC ACID
37
4.1 Abstract
Background
This study investigated synovial concentrations of acetylsalicylic acid (ASA) and its
metabolite salicylic acid (SA) in the equine fetlock joint following systemic
administration of ASA. Salicylates were chosen because SA is the only nonsteroidal
anti-inflammatory drug for which threshold levels exist for plasma and urine in equine
sports. To avoid animal experiments, the study was conducted using an ex vivo
model of the isolated perfused equine distal limb in combination with plasma
concentrations obtained from literature. Salicylate concentrations in the joint were
determined using microdialysis and high performance liquid chromatography (HPLC).
Any anti-inflammatory effect of synovial ASA concentrations was assessed using an
ASA EC50 (half maximal effective concentration) determined in equine whole blood.
Results
The ASA concentration in the synovial fluid (n=6) reached a maximum of 4 µg/mL,
the mean concentration over the entire perfusion period was 2 µg/mL. Maximum SA
concentration was 17 µg/mL, the average was 14 µg/mL. ASA and SA concentration
in the synovial fluid exceeded systemic concentrations 2 h and 3.5 h after “systemic”
administration, respectively.
Conclusions
ASA and SA accumulated in the in the synovial fluid of the ex vivo model despite
decreasing systemic concentrations. This suggests a prolonged anti-inflammatory
effect within the joint that remains to be further elucidated.
Keywords
Acetylsalicylic acid, salicylic acid, isolated perfused equine distal limb, synovial fluid,
horse, microdialysis


PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
39
5 Primary equine synoviocytes in a model of inflammation
5.1 Abstract
In order to test anti-inflammatory effects of substances, cultured synoviocytes in vitro
are a common approach (LANDONI et al. 1996; ARMSTRONG and LEES 2002).
Prostaglandin E2 (PGE2) has been reported to be an important mediator of
inflammation and hyperalgesia (MAY et al. 1994; PALMER and BERTONE 1994b).
Also, it has been demonstrated that cultured equine synoviocytes synthesize large
amounts of PGE2 when stimulated with LPS in vitro (LANDONI et al. 1996). To
assess the anti-inflammatory potency of substances, cells were stimulated with LPS
and either treated with decreasing concentrations of betamethasone, acetylsalicylic
acid, or salicylic acid. The resulting PGE2 production was quantified and compared
with the untreated vehicle group. The lowest concentrations capable of significantly
suppressing PGE2 production were 2.55x10-9 mol/L for BM and 7.77x10-6 for ASA.
For SA, a decrease in PGE2 production was present with increasing SA
concentration, but this decrease was not statistically significant. Results of this study
help to range in the anti-inflammatory effects of intra-articular drug concentrations
measured in the ex vivo model of the isolated perfused equine distal limb.
5.2 Materials and Methods
5.2.1 Cell culture
Culture media
The basic culture medium used for the primary equine synoviocytes was Dulbecco’s
modified Eagle’s medium (DMEM; PAA, Pasching, Germany). All ingredients of this
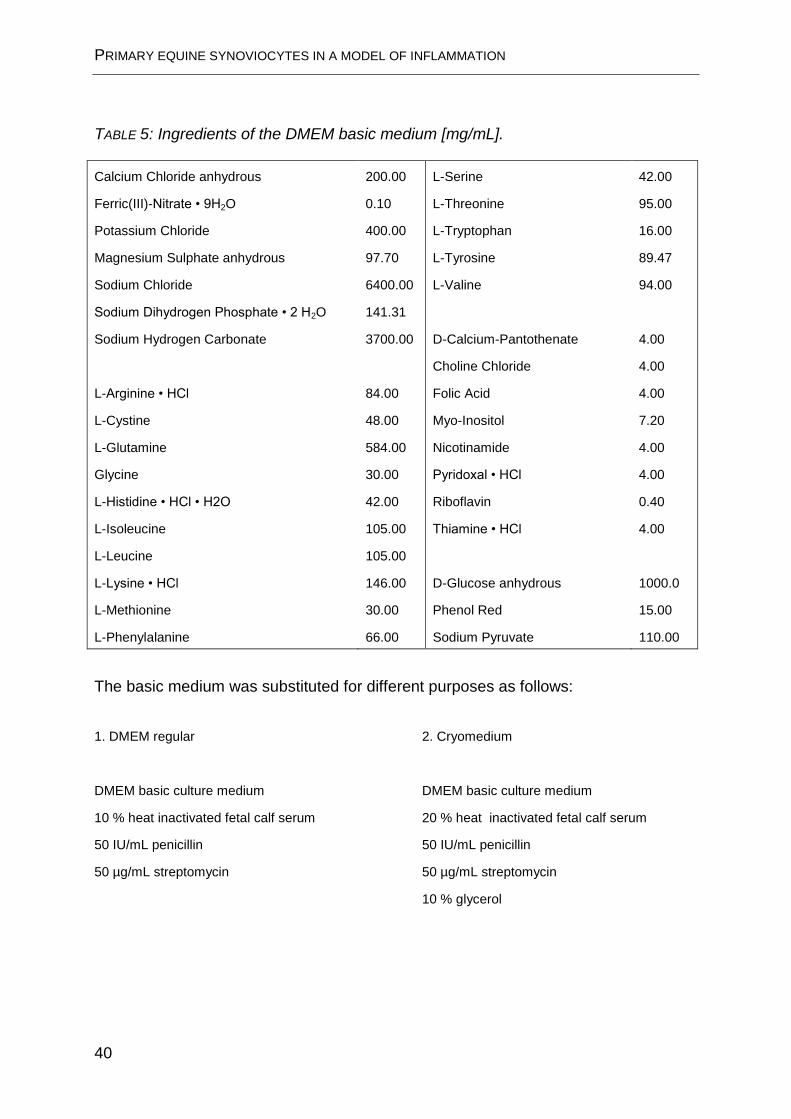
basic medium are listed in TABLE 5.
PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
40
TABLE 5: Ingredients of the DMEM basic medium [mg/mL].
Calcium Chloride anhydrous
Ferric(III)-Nitrate • 9H2O
Potassium Chloride
Magnesium Sulphate anhydrous
Sodium Chloride
Sodium Dihydrogen Phosphate • 2 H2O
Sodium Hydrogen Carbonate
L-Arginine • HCl
L-Cystine
L-Glutamine
Glycine
L-Histidine • HCl • H2O
L-Isoleucine
L-Leucine
L-Lysine • HCl
L-Methionine
L-Phenylalanine
200.00
0.10
400.00
97.70
6400.00
141.31
3700.00
84.00
48.00
584.00
30.00
42.00
105.00
105.00
146.00
30.00
66.00
L-Serine
L-Threonine
L-Tryptophan
L-Tyrosine
L-Valine
D-Calcium-Pantothenate
Choline Chloride
Folic Acid
Myo-Inositol
Nicotinamide
Pyridoxal • HCl
Riboflavin
Thiamine • HCl
D-Glucose anhydrous
Phenol Red
Sodium Pyruvate
42.00
95.00
16.00
89.47
94.00
4.00
4.00
4.00
7.20
4.00
4.00
0.40
4.00
1000.0
15.00
110.00
The basic medium was substituted for different purposes as follows:
1. DMEM regular
DMEM basic culture medium
10 % heat inactivated fetal calf serum
50 IU/mL penicillin
50 µg/mL streptomycin
2. Cryomedium
DMEM basic culture medium
20 % heat inactivated fetal calf serum
50 IU/mL penicillin
50 µg/mL streptomycin
10 % glycerol

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
41
Substances
Heat inactivated fetal calf serum
EDTA (Versen)
Trypsin
Penicillin
Streptomycin
Glycerol
DMSO
Betamethasone
LPS (E.coli, O111:B4)
Collagenase, type VIII from
Clostridium histolyticum
Trypan blue
PAA Laboratories, Pasching, Germany
Biochrom AG, Berlin, Germany
Biochrom AG, Berlin, Germany
PAA Laboratories, Pasching, Germany
PAA Laboratories, Pasching, Germany
Merck, Darmstadt, Germany
Sigma-Aldrich, Steinheim, Germany
Sigma-Aldrich, Steinheim, Germany
Sigma-Aldrich, Steinheim, Germany
Sigma-Aldrich, Steinheim, Germany
Sigma-Aldrich, Steinheim, Germany
Disposable materials
6-mm biopsy punch
96-well tissue culture plate
6-well tissue culture plate
25 cm2 cell culture flask, 50 mL
Sterile cell scraper
Scalpel blade
Minisart® filter unit
Cryovials, 1 mL
kai Europe, Solingen, Germany
Greiner BIO-ONE, Frickenhausen, Germany
Greiner BIO-ONE, Frickenhausen, Germany
Greiner BIO-ONE, Frickenhausen, Germany
TPP, Omnilab, Mettmenstetten, Germany
Bayha, Tuttlingen, Germany
Millipore, Carrigtwohill, Ireland
Greiner BIO-ONE, Frickenhausen, Germany
Cell culture equipment
Incubator CO2 -auto-zero
Centrifuge
Sterile work bench
Phase contrast microscope
Freezing container
Neubauer counting chamber
Heraeus-Kulzer, Hanau, Germany
5804 R, Eppendorf, Hamburg, Germany
LaminAir HLP 2472, Heraeus-Kulzer, Hanau, Germany
Axiovert 25, Zeiss, Oberkochen, Germany
"Mr. Frosty", Sigma-Aldrich, Steinheim, Germany
VWR International, Darmstadt, Germany

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
42
Prostaglandin measurement
Prostaglandin E2 Express EIA Kit -
Monoclonal
Cayman Chemical, Ann Arbor, MI, USA
Substances and materials for immunocytochemistry
Methanol
Acetone
Bisbenzimide
Primary antibody
Secondary antibody
Microscope slides
Mounting medium
Cover slips
Fluorescence microscope
AppliChem, Darmstadt, Germany
Sigma-Aldrich, Steinheim, Germany
Sigma-Aldrich, Steinheim, Germany
Mouse-anti-human protein gene product 9.5 IgG;
Biozol, Eching, Germany
F(ab)2 Goat anti-mouse IgG:FITC;
Serotec, Düsseldorf, Germany
Roth, Karlsruhe, Germany
Fluoromount, Sigma-Aldrich, Steinheim, Germany
VWR International, Darmstadt, Germany
Leica DM LB, Wetzlar, Germany
Equipment
Microplate Reader
Orbital shaker
Precision scale
MRX , Dynatech Laboratories, Denkendorf, Germany
Reax 2000, Heidolph, Kehlheim, Germany
AKS 120-4, Kern & Sohn, Balingen, Germany
Solutions
PBS (0.01 mol/L; pH 7.4)
NaCl
KCl
Na2HPO4 2 • H2O
KH2PO4
Blocking buffer
Triton X-100
BSA
in 1 mL PBS
137 mmol/L
2.7 mmol/L
6.5 mmol/L
1.5 mmol/L
2.5 µL
10 mg
Merck, Darmstadt, Germany
Merck, Darmstadt, Germany
Merck, Darmstadt, Germany
Merck, Darmstadt, Germany
Sigma-Aldrich, Steinheim, Germany
Sigma-Aldrich, Steinheim, Germany

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
43
Animals
All equine distal limbs used for the isolation of synoviocytes were obtained from
warmblood horses of various sex and age slaughtered for meat production at a local
abattoir. Cells were isolated within 3 h after death of the respective horse. Only
synoviocytes from macroscopically healthy joints were included in this study, criteria
for selection were as follows: synovial membrane did not show signs of acute or
chronic inflammation; articular cartilage was free of erosion or wear lines,
periarticular osteophytes were not observed; synovial fluid appeared normal in
quantity, viscosity, and color (FREAN and LEES, 2000).
Isolation of equine synoviocytes
Synoviocytes were isolated based on a protocol by FREAN and LEES (2000).
Synovial membrane was harvested aseptically from the cranial joint recess of the
metacarpophalangeal (fetlock) joint using 6-mm-biopsy punches. The synovial
membrane was dissected from the underlying adipose subintima. Tissue pieces were
collected in 10 mL PBS containing 50 IU/mL penicillin and 50 µg/mL streptomycin
and transferred to a laminar flow hood for further processing. The intimal side of the
membrane section was subjected to enzymatic digestion in type VIII collagenase
from Clostridium histolyticum (312.5 collagen digestion units/mL) in a 6-well-plate for
3 h at 37 °C and 5 % CO2 in a humidified atmosphere. Subsequently, the
supernatant was removed and centrifuged at 4 °C at 350 x g for 10 min. Pooled from
both joints of the same horse, cells were resuspended in 5 mL DMEM regular and
seeded in 25 cm2 flasks so that synoviocytes of six different horses were available.
Culture of equine synoviocytes and cryoconservation
Cells were cultured under standard conditions (37 °C, 5 % CO2 in a humidified
atmosphere). Culture medium was exchanged every two to three days. Synoviocytes
were passaged after six to eight days. Before detachment, all medium was removed
and the cells were washed with 2 mL sterile PBS per flask. Cells were then detached
by adding 2 mL EDTA solution for 5 min at room temperature. The EDTA was

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
44
removed and 1.5 mL trypsin solution was added for 10 min at 37 °C, 5 % CO2 until
the cells were detached and separated. Regular DMEM medium containing 10 %
heat inactivated fetal calf serum, 50 IU/mL penicillin and 50 µg/mL streptomycin was
added to inactivate trypsin before centrifugation of the cells at 4 °C at 350 x g for
10 min. The supernatant was decanted and the cells were seeded in fresh medium at
a density of 2,000 cells/cm2 into 25 cm2 cell culture flasks.
Equine synoviocytes were cryoconserved and stored in liquid nitrogen at -196 °C in
order to have cells available at any time. The cells were detached as described
earlier. After centrifugation, the cells were resuspended in 1 mL of cryomedium and
transferred to cryovials at a density of 1,000,000 – 4,000,000 cells/mL. The vials
were stored at – 80 °C in a freezing container for 4 h and then transferred to liquid
nitrogen. For thawing, cryovials were put in a water bath at 37 °C until only a frozen
core of approx. 2 mm diameter remained. Cryovials were then placed under the
sterile bench where the cells were carefully resuspended in pre-warmed regular
DMEM, transferred to culture flasks and placed in the incubator. Medium was
exchanged 4 h after seeding and from then on every two to three days.
Cell count was routinely determined during passaging: 50 µL of a cell suspension
were diluted with 100 µL trypan blue suspension (40 mg trypan blue in 10 mL aq.
dest.) and 10 µL of this suspension placed in a Neubauer counting chamber. The
cells were counted in the four quadrants of the chamber using a 100-fold
magnification under the microscope. Since all cells incorporate the blue dye but only
viable cells are able to eliminate it, the trypan blue staining allows for a differentiation
between viable and dead cells. Under the microscope, dead cells display a blue
cytoplasm and nucleus, whereas viable cells appear unstained. The total cell count
was calculated using the following equation:
Cells counted = Number of cells in 4 counting chamber quadrants
Cells total = Cell count in 1mL of culture medium

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
45
Cell viability after stimulation and/or treatment was evaluated using a colorimetric test
in which the yellow MTS tetrazolium compound 3-(4,5-dimethylthiazol-2-yl)-5-(3-
carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium is reduced to a brown
formazan compound by enzymatic activity of viable cells. The resulting intracellular
formazan can be quantified by spectrophotometric means.
5.2.2 Verification of equine synoviocytes
Protein Gene Product 9.5, originally isolated from human brain as a brain-specific
protein, has also been described as a specific cytochemical marker for the
identification of equine type B synoviocytes (KITAMURA et al. 1999). In equine type
B synoviocytes, immunoreactivity for PGP 9.5 is diffusely distributed throughout the
cytoplasm. Synoviocytes of each horse were analyzed by immunocytochemical
staining for PGP 9.5 and only the positive cultures were employed in the
inflammation experiments.
Immunocytochemistry
For immunocytochemistry staining, cells were first seeded on 12-mm-cover glasses
and grown to confluence. To do so, sterilized cover glasses were placed in a 24-well-
plate and covered with 10,000 cells in 500 µL regular DMEM per well. As soon as the
cells were confluent, all medium was removed and cells were fixed with cold
methanol and acetone (1:1) for 5 min. Slides were either stained immediately
afterwards or stored in sterile PBS at 4 °C.
During the entire staining protocol, care was taken to avoid drying of the slides. The
detailed staining protocol is given in TABLE 6. Bisbenzimide binds to DNA and is a
nuclear counterstain. After staining, slides were stored at 4 °C and protected from
light for 24 h before visualizing the results using a fluorescence microscope and the
software KS 400.

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
46
TABLE 6: Protocol for immunostaining of PGP 9.5.
Washing of cover glasses containing the fixed cells twice with 300 µL of PBS.
Blocking of non-specific staining by adding 300 µL of blocking buffer and incubation for
30 min at room temperature.
Removal of blocking buffer without rinsing.
Dilution of the primary antibody in blocking buffer (1:500). Addition of 150 µL per cover
glass and incubation at 37 °C for 1.5 h. For the negative control, addition of blocking
buffer instead of primary antibody.
Removal of primary antibody, washing twice with 300 µL of blocking buffer.
Dilution of the secondary antibody in blocking buffer (1:200) and addition of 150 µL to
each well. Incubation at 37 °C for 30 min in the dark.
- From this step forward, protection of samples from light -
Removal of secondary antibody and rinsing twice with 300 µL of PBS.
Addition of 150 µL of the diluted bisbenzimide solution (1 µg/mL) to each well,
incubation for 10 min at room temperature.
Rinsing once with 300 µL PBS, then careful removal of the cover glasses from the wells
and blotting dry to remove any excess water.
Addition of one drop of mounting medium per cover glass onto the microscope slide.
Mounting of the coverslip with the cells facing towards the microscope slide.

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
47
5.2.3 In vitro inflammation model with equine synoviocytes
Pooled synoviocytes from both fetlock joints of six horses were seeded in 96-well-
plates at a density of 10,000 cells per well and grown to confluence in 1-2 d. All anti-
inflammatory substances except for BM were added simultaneously with LPS in
100 µL of regular DMEM; BM was added 4 h prior to stimulation with LPS because of
the time needed for GCR activation, transcription, and translation. Cells were
incubated at 37 °C and 5 % CO2 in a humidified atmosphere. After 24 h,
supernatants were taken and centrifuged for 5 min at 4 °C and 3000 x g. Four
hundred µL of the resulting supernatant were removed and stored at - 80 °C until
analysis.
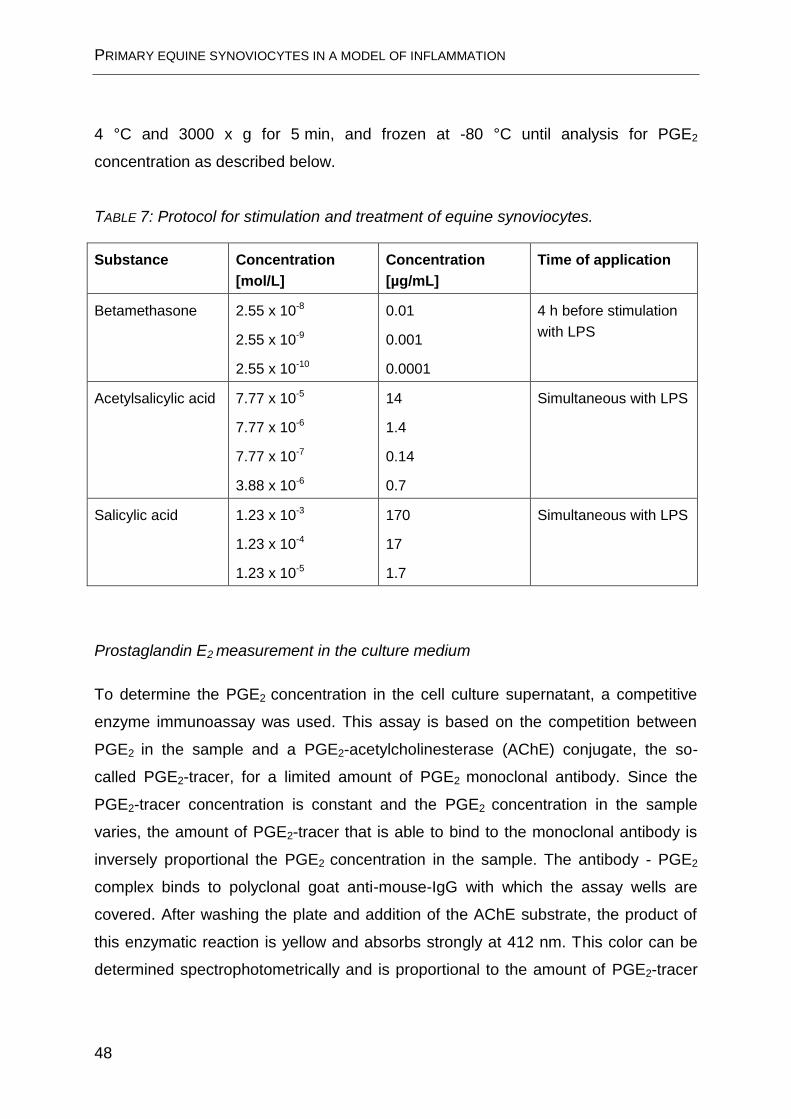
Stimulation with LPS and treatment
Synoviocytes were stimulated with LPS (E.coli O111:B4) to trigger an inflammation-
like reaction. In preliminary experiments, different concentrations of LPS were tested
for their capability to stimulate PGE2 production in the cells. They were stimulated
with 10 µg/mL or 100 µg/mL of LPS for 24 h. The optimal concentration with regard to
cell viability and PGE2 production was 10 µg/mL LPS (data not shown). This
concentration was applied in the main experiments, where LPS was diluted in regular
DMEM.
Those three anti-inflammatory agents that had been used in experiments with the
isolated perfused equine distal limb (BM, ASA and SA) were added to the LPS-
stimulated synoviocytes. BM concentrations had been determined in preliminary
experiments (data not shown). ASA and SA concentrations were those measured in
synovial fluid at the end of the ex vivo experiments plus one order of magnitude
greater and lesser, as indicated in TABLE 7. Immediately before each experiment, all
anti-inflammatory substances were dissolved in DMSO and diluted in regular DMEM
to a final DMSO concentration of 1 ‰. Four wells received the same treatment with
LPS stimulation. The control group was stimulated with LPS and treated with vehicle
only (medium with 1 ‰ DMSO). After 24 h of incubation at 37 °C, 5 % CO2 in a
humidified atmosphere, supernatants were taken, pooled, immediately centrifuged at
PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
48
4 °C and 3000 x g for 5 min, and frozen at -80 °C until analysis for PGE2
concentration as described below.
TABLE 7: Protocol for stimulation and treatment of equine synoviocytes.
Substance Concentration
[mol/L]
Concentration
[µg/mL]
Time of application
Betamethasone 2.55 x 10-8
2.55 x 10-9
2.55 x 10-10
0.01
0.001
0.0001
4 h before stimulation
with LPS
Acetylsalicylic acid 7.77 x 10-5
7.77 x 10-6
7.77 x 10-7
3.88 x 10-6
14
1.4
0.14
0.7
Simultaneous with LPS
Salicylic acid 1.23 x 10-3
1.23 x 10-4
1.23 x 10-5
170
17
1.7
Simultaneous with LPS
Prostaglandin E2 measurement in the culture medium
To determine the PGE2 concentration in the cell culture supernatant, a competitive
enzyme immunoassay was used. This assay is based on the competition between
PGE2 in the sample and a PGE2-acetylcholinesterase (AChE) conjugate, the so-
called PGE2-tracer, for a limited amount of PGE2 monoclonal antibody. Since the
PGE2-tracer concentration is constant and the PGE2 concentration in the sample
varies, the amount of PGE2-tracer that is able to bind to the monoclonal antibody is
inversely proportional the PGE2 concentration in the sample. The antibody - PGE2
complex binds to polyclonal goat anti-mouse-IgG with which the assay wells are
covered. After washing the plate and addition of the AChE substrate, the product of
this enzymatic reaction is yellow and absorbs strongly at 412 nm. This color can be
determined spectrophotometrically and is proportional to the amount of PGE2-tracer

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
49
bound to the well, which is inversely proportional to the amount of free PGE2 present
in the sample. A standard curve was performed with each assay and the percentage
of binding used to calculate PGE2 concentrations. Each assay included the following
controls: blank, total activity, non-specific binding, and maximum binding.
Viability testing
After removal of the supernatants, a colorimetric test was performed as described
above to assess cell viability after stimulation and treatment, and to ensure that the
PGE2 concentration in the culture medium was not a result of cell decay.
5.2.4 Statistical analysis
The stimulation / treatment experiments were conducted six times with synoviocytes
from six different horses. Each concentration of an anti-inflammatory drug was
applied in quadruplicate. To compare PGE2 concentrations of LPS-stimulated cells
after treatment with those of that only received vehicle treatment, PGE2
concentrations were analyzed by means of a one-way analysis of variance followed
by a Dunnett’s multiple comparison test. For viability testing, metabolic activity of the
positive control group was normalized to 100 % and results of the treated cells
expressed in relation to that. They were analyzed with a one-way analysis of
variance followed by a Kruskal-Wallis test. In both cases, a p-value < 0.05 was
considered significant. Statistical calculations were performed with GraphPad Prism
5.03 software (GraphPad Software, La Jolla, CA, USA).
5.3 Results
5.3.1 Isolation and culture of synoviocytes
Equine synoviocytes were enzymatically and mechanically isolated, and taken into
culture. Cells exhibited the typical spindle shaped morphology under the phase
contrast microscope (FIGURE 4).

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
50
5.3.2 Verification of synoviocytes with immunocytochemistry
Primary equine synoviocyte cultures were analyzed by staining with a primary
antibody against PGP 9.5. None of the negative controls showed any staining
whereas synoviocyte cultures from all horses exhibited a diffuse staining throughout
their cytoplasm as depicted in FIGURE 4.
FIGURE 4: Verification of equine synoviocytes Type B. 1: Spindle – like morphology
(Phase contrast microscope). 2: Positive staining for protein gene product 9.5
(Fluorescence microscope).
1 100 µm
50 µm
2

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
51
5.3.3 In vitro inflammation model
Viability testing after LPS-stimulation
To assess the influence of anti-inflammatory treatment on the viability of cells and to
make sure that PGE2 production was not due to cell death, a colorimetric test was
performed with all 96-well-plates used as described earlier. Results are shown in
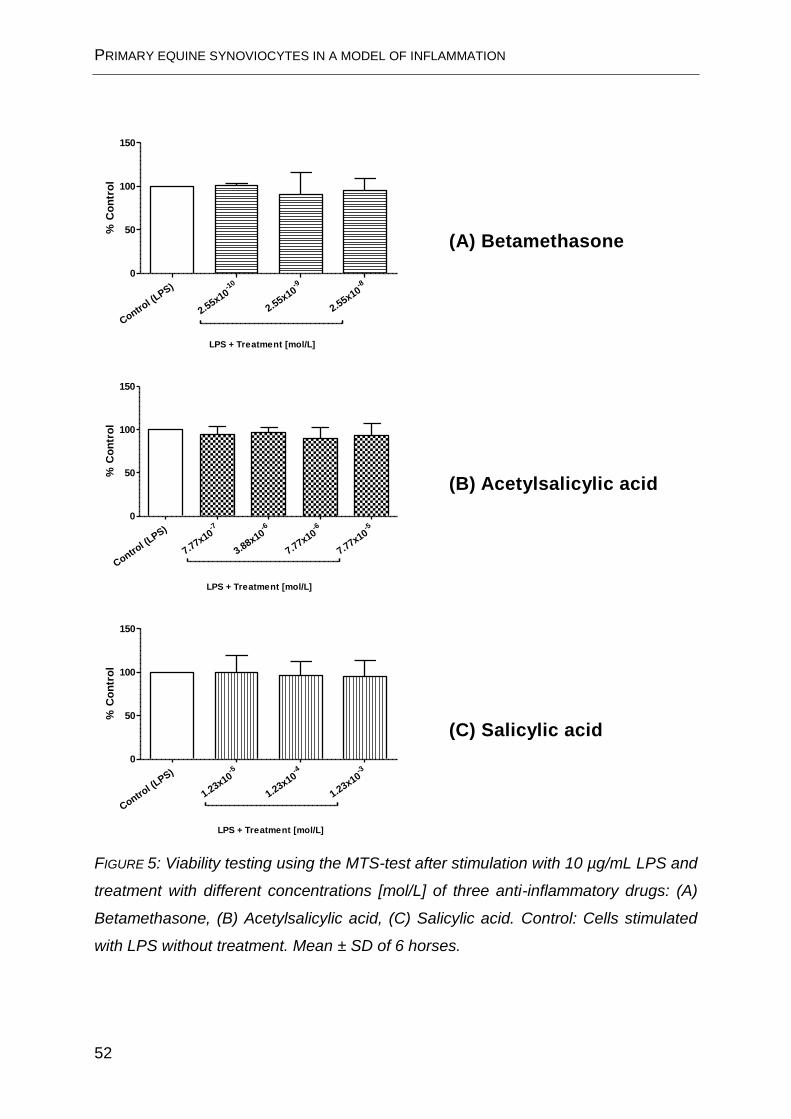
FIGURE 5. No significant deterioration in cell viability can be seen as a result of
treatment.
PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
52
Control (
LPS) -10
2.55x10
-9
2.55x10
-8
2.55x10
0
50
100
150
LPS + Treatment [mol/L]
% C
on
tro
l
Control (
LPS) -7
7.77x10
-6
3.88x10
-6
7.77x10
-5
7.77x10
0
50
100
150
LPS + Treatment [mol/L]
% C
on
tro
l
Control (
LPS) -5
1.23x10
-4
1.23x10
-3
1.23x10
0
50
100
150
LPS + Treatment [mol/L]
% C
on
tro
l
FIGURE 5: Viability testing using the MTS-test after stimulation with 10 µg/mL LPS and
treatment with different concentrations [mol/L] of three anti-inflammatory drugs: (A)
Betamethasone, (B) Acetylsalicylic acid, (C) Salicylic acid. Control: Cells stimulated
with LPS without treatment. Mean ± SD of 6 horses.
(A) Betamethasone
(B) Acetylsalicylic acid
(C) Salicylic acid

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
53
LPS stimulation and treatment
Results for the stimulation of equine synoviocytes with LPS (10 µg/µL) with or without
anti-inflammatory treatment are illustrated in FIGURE 6.
For BM and ASA, the PGE2 inhibition was dose-dependent: for cells treated with BM
(A), the increase in PGE2 production was significantly inhibited at 2.55x10-8 mol/L
(0.01 µg/mL) and 2.55x10-9 mol/L (0.001 µg/mL) but not at 2.55x10-10 mol/L
(0.0001 µg/mL). ASA (B) significantly reduced PGE2 production at 7.77x10-5 mol/L
(14 µg/mL) and 7.77x10-6 mol/L (1.4 µg/mL), but not at 3.88x10-6 mol/L (0.7 µg/mL)
and 7.77x10-7 mol/L (0.14 µg/mL). None of the three SA (C) concentrations tested
were able to significantly inhibit PGE2 production: 1.23x10-3 mol/L (170 µg/mL),
1.23x10-4 mol/L (17.0 µg/mL), 1.23x10-5 mol/L (1.7 µg/mL).

PRIMARY EQUINE SYNOVIOCYTES IN A MODEL OF INFLAMMATION
54
+LPS -10
2.55x10
-9
2.55x10
-8
2.55x10
0
50
100
**
*
LPS + Treatment [mol/L]
PG
E2 (
ng
/mL
)
+LPS -7
7.77x10
-6
3.88x10
-6
7.77x10
-5
7.77x10
0
50
100
*
***
LPS + Treatment [mol/L]
PG
E2 (
ng
/mL
)
+LPS -5
1.23x10
-4
1.23x10
-3
1.23x10
0
50
100
LPS + Treatment [mol/L]
PG
E2 (
ng
/mL
)
FIGURE 6: PGE2 concentration in the culture medium after 24 h of stimulation with
10 µg/mL LPS and treatment with (A) Betamethasone, (B) Acetylsalicylic acid or
(C) Salicylic acid. Data are given as mean ± SD of 6 horses (Analysis of variance and
Dunett’s multiple comparison test, p < 0.05).
(A) Betamethasone
(B) Acetylsalicylic acid
(C) Salicylic acid

GENERAL DISCUSSION
55
6 General discussion
It was the aim of the study presented to establish an ex vivo model of the isolated
perfused equine distal limb in order to facilitate pharmacokinetic studies concerning
the equine foot. Pharmacokinetic data are urgently needed by veterinarians who treat
equine athletes to ensure that recovered horses do not return to competition until
they are clear of all medication. The established ex vivo model was designed to
reduce the number of living horses used, which renders these studies less labor-
intensive and less expensive. Perfusion medium and -conditions were optimized with
regard to the following viability parameters: Glucose consumption, lactate production,
LDH liberation, weight increase, skin surface temperature, and histologic changes in
the joint capsule.
Two substances were administered either locally, i.e. intra-articularly, or systemically,
i.e. via the systemic circulation, as it is done to target equine joint disease in vivo.
The ex vivo model was subsequently used to quantify the amount of drug in the joint
over the perfusion period of 8 h. The remaining anti-inflammatory effect of the
respective intra-articular concentration was assessed by examining the ability to
inhibit PGE2 production following stimulation with LPS in an in vitro inflammation
model of cultured equine synoviocytes.
6.1 Model characteristics
For the ex vivo model of the isolated perfused equine distal limb, criteria have been
formulated in order to set minimum standards for tissue viability. However, one
disadvantage of the model is the relative heterogeneity of animals at the abattoir.
Even though only warmblood horses showing no signs of lameness were included in
this study, some variables such as different age, sex, and history of joint disease
remain. It has to be considered, though, that also in vivo studies recruit horses of
different age, sex, and often breed. These horses are not as uniform as laboratory
animals such as mice or rats, which are not only genetically identical due to
inbreeding but are also kept under standardized conditions.

GENERAL DISCUSSION
56
The other limitation of all isolated perfused organs, the restricted time span in which
they are viable, could be overcome by extrapolation of data using pharmacokinetic
parameters determined elsewhere, as it was done for BM.
The synovial clearance of a substance is among others dependent on the blood flow,
whose increase e.g. with exercise also increases the transport of solutes from the
joint to the circulation (DULIN et al. 2012). These physiological changes in blood flow
were not mirrored in the ex vivo model; the perfusion rate was constantly set at the
lower limit of flow rates reported in literature as a compromise between sufficient
tissue supply and prevention of edema formation (ROBINSON et al. 1975;
WONGAUMNUAYKUL et al. 2006).
The model’s most evident advantage is the possibility to investigate the distribution of
drugs administered to the equine foot without the necessity to use living horses.
DERENDORF et al. (1986) have described the so-called “flip-flop case”, according to
which the terminal phase of the concentration-time-curve of intra-articularly
administered triamcinolone acetonide and BM is represented rather by the absorption
rate from the joint than by the elimination phase. The absorption rates can be
investigated in the model since they do not require the animal’s systemic circulation
or excretory organs. It should also be possible to compare joint clearance rates of
different substance formulations. Furthermore, the model offers sampling sites that
are not accessible in the living animal such as efferent veins, circumventing the
substance’s dilution in the entire blood volume as it would be case in vivo.
In order to get some orientation concerning the influence of plasma protein binding
on synovial distribution of systemically administered substances, two extremities
were perfused with a blood-tyrode mixture (4:1) instead of tyrode solution. The
resulting synovia concentrations of ASA and SA were 49.5 % and 47.8 %,
respectively, in comparison with synovial concentrations after perfusion with cell-free
tyrode solution. These results reflect the plasma protein binding of around 50 % for
salicylates (FRASER et al. 1986). It can therefore be speculated that synovial fluid
concentrations of substances in the tyrode-perfused model represent the
concentration without plasma protein binding. Since this characteristic is known for
most drugs, synovial concentrations in tyrode-perfused limbs would simply have to

GENERAL DISCUSSION
57
be adjusted by the plasma protein binding in order to approximate the in vivo
situation.
The model has been established to mirror conditions of the healthy limb and can
therefore not reflect possible alterations due to pathologic processes. Even though
these processes may have an influence on drug distribution, the investigation under
non-diseased conditions was priority.
6.2 Betamethasone
Glucocorticoids are one of the most commonly administered classes of intra-articular
medication in equine medicine (LEES 2003; YARBROUGH 2004). In the study
presented BM, a typical representative of this drug class, was chosen as model
substance.
In order to assess the value of data obtained from the ex vivo model of the isolated
perfused equine distal limb, an extrapolation was made to estimate the time until BM
would no longer be detectable in synovial fluid of the fetlock joint. This time span was
then compared with plasma concentrations from an in vivo study, in which an intra-
articular injection of the identical dose of BM had been performed (LÖVENICH 2012).
Pharmacokinetic parameters that could not be obtained from the ex vivo model were
adapted from a study that had investigated them in vivo (DIKKER 2009).
For calculation, a three-compartment-model was used (joint, blood, peripheral
compartment). A horse of 500 kg bodyweight and a blood volume of 35 L was
assumed. In the aforementioned in vivo study (n=6 horses), LÖVENICH (2012) found
that BM was detectable in plasma for up to 168 h (LOD 4 pg/mL). With data from the
isolated perfused equine distal limb, it was extrapolated that BM concentration in the
joint would decrease to 36 ng/mL after 168 h, possibly indicating an overestimation of
the real concentration. However, it has also been reported that after intra-articular
injection, a glucocorticoid is longer detectable in synovial fluid than in plasma (CHEN
et al. 1992). Also, the return to normal concentrations is described to be slightly
delayed in synovial fluid in comparison with plasma (VINE 2006). As indicated
before, BM phosphate is known to be a more rapid-acting ester (FERGUSON et al.
2009). Mean BM concentration in the joint 7.5 h after intra-articular injection of 4 mg

GENERAL DISCUSSION
58
BM per joint (as 5.3 mg BM disodium phosphate) was calculated to be 1.7 mg. This
finding is supported by other studies in which hydrosoluble esters such as
phosphates are cleared from the joint rather quickly (AUTEFAGE et al. 1986;
DERENDORF et al. 1986). Less soluble glucocorticoid esters such as
methylprednisolone acetate, triamcinolone acetonide or BM acetate are detectable
for longer periods of time (SOMA et al. 2006; LÖVENICH 2012).
In this context, it would be interesting to know which concentration still leads to a
pharmacologic, i.e. anti-inflammatory, effect. Results of the in vitro experiments with
cultured equine synoviocytes described in this thesis have shown that a BM
concentration of as low as 1 ng/mL (2.55x10-9 mol/L) significantly suppresses PGE2
production after LPS stimulation of the cells. A study investigating PGE2 inhibition in
human rheumatoid synovia reports dexamethasone (whose structure is virtually
identical with that of BM) concentrations of 10-8 mol/L (3.93 ng/mL) to 10-6 mol/L
(393 ng/mL) to be effective (KANTROWITZ et al. 1975).
These findings indicate that rather small BM concentrations still exert an anti-
inflammatory effect, even though its presence in the joint after intra-articular
application might not be detectable in plasma samples.
6.3 Acetylsalicylic and salicylic acid
Although not frequently used for treatment of joint disease in horses, synovial
concentrations of ASA and its metabolite SA are of special interest because SA is a
threshold substance. This means that a horse may feature SA concentrations of up
to 625 µg/mL (urine) and 5.4 µg/mL (plasma) in national events in Germany
(ANONYMUS 2011) and up to 750 µg/mL (urine) and 6.5 µg/mL (plasma)
internationally (ANONYMUS 2010). Furthermore, ASA is known to have a very short
half-life in horses and might therefore be chosen if treatment with an NSAID should
appear necessary shortly before a competition.
Synovial fluid concentrations after simulated systemic administration of ASA were
obtained from the ex vivo model of the isolated perfused equine distal limb. Systemic
plasma concentrations of ASA and SA were taken over from a study that had
investigated their pharmacokinetic properties after systemic administration of ASA

GENERAL DISCUSSION
59
(20 mg/kg bodyweight) to horses (BROOME et al. 2003). ASA and SA were detected
by means of high performance liquid chromatography and UV detection with limits of
detection (LOD) at 53 ng/mL and 84 ng/mL and limits of quantification (LOQ) at
106 ng/mL and 169 ng/mL, respectively.
None of the synovial blank samples analyzed before the onset of systemic ASA
application contained any measurable concentrations of ASA or SA. After systemic
administration, the decrease of ASA and SA concentrations in the joint did not
parallel the decrease in the perfusion fluid. This accumulation in synovial fluid is in
accordance with findings in humans (SHOLKOFF et al. 1967; SOREN 1977).
Although the exact reasons for this phenomenon have never been elucidated, it can
be speculated that the accumulation is due to at least two factors: firstly, the
continuous deacetylation of ASA to SA in plasma leads to a constant supply of ASA
and SA to the joint and secondly, the lower esterase activity in the synovial fluid
results in less ASA being converted to SA so that ASA concentrations remain
relatively constant once ASA has diffused into the synovial cavity.
Particularly those synovial fluid concentrations are of clinical relevance that were
measured when the SA plasma concentration had fallen below the FEI threshold
level. In the systemic plasma concentrations adapted from BROOME et al. (2003),
this was the case after 3.5 h. Corresponding synovial fluid concentrations of ASA and
SA until the end of the experiment were 1x10-5 to 8x10-6 mol/L and 1.15x10-4 to
1.24x10-4 mol/L, respectively. A potential anti-inflammatory effect of these
concentrations would be of special importance because at the time these
concentrations were measured within the joint, the horse’s plasma concentration
would have been below the permitted threshold.
In vitro experiments within the framework of this thesis have revealed a dose-
dependent inhibition of PGE2 production after LPS stimulation of primary equine
synoviocytes for ASA, the lowest ASA concentrations capable of significantly
inhibiting PGE2 production being 7.77x10-6 mol/L. For SA, a decrease in PGE2
production could be noted with increasing SA concentration, but this decrease was
not statistically significant. These results are in accordance with data from
BUNTENKÖTTER (2012), who has calculated the following EC50 values (which

GENERAL DISCUSSION
60
describe those plasma concentrations at which half of the maximum possible effect is
provided (TOUTAIN 2009)) for horses: for ASA, the EC50 values were 1.17x10-6 mol/L
for COX I and 1.23x10-5 mol/L for COX II. For SA, an EC50 value could not be
calculated because the inhibition at the concentrations tested was too weak. Four
other studies have determined EC50 values for COX I inhibition by ASA between
1.67x10-6 mol/L and 4.45x10-6 mol/L and for COX II inhibition between 5.35x10-6
mol/L and 1.59x10-5 mol/L. However, these values were not determined specifically
for horses (CROMLISH and KENNEDY 1996; CRYER and FELDMAN 1998;
GIULIANO and WARNER 1999; HINZ et al. 2000).
With regard to SA concentrations in urine samples, BUNTENKÖTTER (2012)
observed that after oral administration of 12.5 µg/kg, 25 µg/kg, and 50 µg/kg ASA,
urine concentrations decreased rather quickly: values fell below the FEI threshold
level of 750 µg/mL after 12 h (12.5 µg/kg) or 24 h (25 µg/kg and 50 µg/kg). However,
concentrations were broadly distributed so that three out of eight horses showed a
urine concentration below the threshold as early as 7 h after administration
(12.5 µg/kg), two out of eight 12 h after administration (25 µg/kg), and two out of
sixteen horses did not show a SA urine concentration above the threshold level at all
(one each for 12.5 µg/kg and 25 µg/kg).
Together, these data give a strong hint that the observed accumulation of ASA in
synovial fluid might lead to an anti-inflammatory effect despite plasma concentrations
below the threshold level. Likewise, it has been reported that NSAIDs accumulate in
and are cleared slowly from inflammatory exudate (LANDONI et al. 1995; LANDONI
and LEES 1995). Hence, the level of SA threshold levels might have to be
reassessed in order to avert the option of treating inflammatory joint conditions in
horses with ASA by taking advantage of its short half-life in plasma and urine coupled
with an accumulation in synovial fluid.
6.4 Outlook
The ex vivo model of the isolated perfused equine distal limb appears to be a
promising tool to investigate the distribution of substances targeting the equine joint,
either administered intra-articularly or systemically, as demonstrated with the model

GENERAL DISCUSSION
61
substances BM and ASA / SA. Further applications might include the comparison of
different formulations of e.g. glucocorticoids or NSAIDs. Also, it would be interesting
to compare synovial fluid concentrations of other therapeutic agents such as
antibiotics after intra-articular with those after systemic administration.
The in vitro equine synoviocyte model of inflammation has proven to be useful for
determining effective concentrations of anti-inflammatory substances. Measurement
of the cells’ inflammatory response might be extended to other parameters of
inflammation.


SUMMARY
63
7 Summary
Maren Friebe (2013)
The isolated perfused equine distal limb and synoviocyte cultures as models
for pharmacological studies
It was the aim of this PhD-project to establish suitable models for investigating
pharmacokinetic and pharmacodynamic properties of substances administered to the
equine joint.
Firstly, in order to assess the distribution of drugs either after intra-articular or
systemic application, the ex vivo model of the isolated perfused equine distal limb
was established. Distal forelimbs of horses obtained from a slaughterhouse were
exarticulated in the carpal joint. Within 1 h after slaughter, the median artery was
canulated and extremities were perfused with oxygenated tyrode solution containing
sodium carboxymethyl cellulose at a flow rate of approx. 65 mL/min for up to 8 h.
Tissue viability during the perfusion period was verified by determination of glucose
consumption, lactate production and LDH liberation in the venous perfusate sampled
from the radial vein. Furthermore, skin surface temperature and weight increase were
monitored as indicators of peripheral blood supply and edema formation. The
integrity of the joint capsule was assessed by histologic examination. Minimum
requirements for parameters were formulated to ensure sufficient viability of the distal
limb.
The articular efflux rate of BM in the venous perfusate of the radial vein after intra-
articular injection of BM disodium phosphate (4 mg BM) into the fetlock joint was
measured with HPLC and UV detection (n=6). The average BM efflux rate per minute
was calculated to be 5.1 µg/min with values ranging from 9 µg/min to 2.9 µg/min. 7.5
h after i.a.-application, 2.3 mg BM had left the joint.

SUMMARY
64
Also, the distribution of ASA and its metabolite SA into the fetlock joint after systemic
administration of ASA was determined in the isolated perfused equine distal limb
(n=8). Plasma concentrations of ASA and SA were adapted from an in vivo study and
added to the perfusate (BROOME et al. 2003). ASA and SA concentrations in the
synovial fluid were determined using calibrated microdialysis probes and HPLC-UV.
Two distal limbs were perfused with diluted autologous blood in order to give some
orientation concerning the influence of plasma protein binding on the synovial
distribution. For the tyrode-perfused limbs (n=6), ASA concentrations in the synovial
fluid reached a maximum of 2.1x10-5 mol/L, maximum SA concentrations were
1.27x10-4 mol/L. Synovial salicylate concentrations in the hemoperfused limbs were
about 50 % lower, presumably owing to the salicylate plasma protein binding of
50 %. The decrease of ASA and SA in synovial fluid did not parallel the decrease in
plasma concentrations, indicating an accumulation in the joint.
Secondly, in order to assess the anti-inflammatory potency of concentrations
measured in the isolated perfused equine distal limb, a cell culture model of primary
equine synoviocytes was established. Cells isolated from macroscopically healthy
fetlock joints (n=6) were stimulated with LPS and treated with decreasing doses of
BM, ASA and SA. The inhibition of prostaglandin E2 production in comparison with an
untreated control was determined as an indicator of an anti-inflammatory effect.
The lowest concentrations causing a significant inhibition of PGE2 production were
2.55x10-9 mol/L and 7.77x10-6 mol/L for BM and ASA, respectively. SA
concentrations employed did not cause a significant PGE2 inhibition. These results
may be a hint for a prolonged anti-inflammatory effect in the joint in comparison with
plasma concentrations.
In summary, the isolated perfused equine distal limb and primary equine synoviocyte
cultures were successfully established as useful tools for the investigation of
pharmacokinetic and pharmacodynamic properties of substances administered to the
equine joint without the necessity to use living horses.

ZUSAMMENFASSUNG
65
8 Zusammenfassung
Maren Friebe (2013)
Das isoliert perfundierte Pferdebein und kultivierte Synoviozyten als Modelle
für pharmakologische Studien
Ziel der vorliegenden PhD-Arbeit war es, geeignete Modelle zur Untersuchung
pharmakokinetischer und pharmakodynamischer Daten von Substanzen zu
etablieren, mit denen beim Pferd Gelenksbehandlungen durchgeführt werden.
Um zunächst die Verteilung von Stoffen nach intra-artikulärer oder systemischer
Applikation zu untersuchen, wurde das ex vivo Modell des isoliert perfundierten
distalen Pferdebeins etabliert. Vordergliedmaßen von Pferden wurden an einem
Schlachthof im Karpalgelenk exartikuliert und innerhalb von 1 h nach der
Schlachtung über die A. mediana mit oxygenierter Tyrodelösung, die Natrium-
Carboxymethylcellulose enthielt, bei einer Flussrate von ca. 65 ml/min für 8 Stunden
perfundiert. Die Gewebevitalität während der Perfusion wurde durch die Bestimmung
von Glukoseverbrauch, Laktatproduktion und LDH-Freisetzung im venösen Perfusat
der V. radialis überprüft. Des Weiteren wurden Hautoberflächentemperatur und
Gewichtszunahme als Indikatoren für periphere Blutversorgung und Ödembildung
kontrolliert. Die Unversehrtheit der Gelenkkapsel wurde mittels histologischer
Untersuchung beurteilt. Mindestanforderungen für eine ausreichende Vitalität der
distalen Gliedmaße wurden formuliert.
Die Abflutung von Betamethason über die V. radialis nach intra-artikulärer Injektion
von BMdihydrogenphosphat-Dinatrium (4 mg BM) ins Fesselgelenk wurde mittels
HPLC und UV-Detektion gemessen (n=6). Die durchschnittliche BM - Abflutung pro
Minute betrug 5.1 µg/min, mit Werten zwischen 9 µg/min und 2.9 µg/min. 7.5
Stunden nach i.a.-Applikation waren 2.3 mg BM aus dem Gelenk abgeflutet.
Die Distribution von Acetylsalicylsäure und ihres Metaboliten Salicylsäure in das
Fesselgelenk nach systemischer Applikation von ASA wurde ebenfalls am isoliert
perfundierten Pferdebein untersucht (n=8). Plasmakonzentrationen von ASA und SA

ZUSAMMENFASSUNG
66
wurden aus einer entsprechenden in vivo Studie übernommen und dem Perfusat
zugesetzt (BROOME et al. 2003). Ihre Konzentrationen in der Synovialflüssigkeit
wurden mit einer kalibrierten Mikrodialysesonde und HPLC-UV bestimmt. Zwei Beine
wurden mit verdünntem autologem Blut perfundiert, um eine Orientierung hinsichtlich
des Einflusses der Plasmaproteinbindung zu erhalten. In den Tyrode-perfundierten
Gliedmaßen (n=6) wurden maximale synoviale Konzentrationen von 2.1x10-5 mol/l
(ASA) und 1.27x10-4 mol/l (SA) erreicht. Die Salicylatkonzentration in der
Synovialflüssigkeit der hämoperfundierten Beine war ca. 50 % niedriger,
wahrscheinlich aufgrund der Plasmaproteinbindung der Salicylate von 50 %. Die
Abnahme von ASA und SA in der Synovia verlief bedeutend langsamer als im
Plasma, was für eine Akkumulation im Gelenk spricht.
Um im Anschluss die antiphlogistische Wirkung der im isoliert perfundierten
Pferdebein gemessenen Synovialkonzentrationen einzuordnen, wurde ein primäres
Zellkulturmodell aus equinen Synoviozyten etabliert. Die aus makroskopisch
gesunden Fesselgelenken (n=6) isolierten Zellen wurden mit LPS stimuliert und mit
abnehmenden Konzentrationen von BM, ASA und SA behandelt. Die Inhibition der
Prostaglandin E2-Produktion im Vergleich mit einer unbehandelten Kontrolle wurde
als Indikator für einen entzündungshemmenden Effekt verwendet. Die niedrigsten
Konzentrationen, die zu einer signifikanten Hemmung der PGE2-Produktion führten,
betrugen 2.55x10-9 mol/l für BM und 7.77x10-6 mol/l für ASA. Die eingesetzten SA-
Konzentrationen führten nicht zu einer signifikanten PGE2-Hemmung. Die Ergebnisse
bedeuten einen Hinweis darauf, dass die entzündungshemmende Wirkung im
Gelenk im Vergleich zum Plasma länger andauert.
Zusammenfassend lässt sich sagen, dass das isoliert perfundierte Pferdebein und
die Synoviozytenkultur erfolgreich etabliert werden konnten. Ohne dass
Versuchspferde eingesetzt werden, ermöglichen sie die Untersuchung
pharmakokinetischer und pharmakodynamischer Charakteristika von Substanzen,
die für die Gelenksbehandlung beim Pferd eingesetzt werden.

REFERENCES
67
9 References
ARMSTRONG, S. and P. LEES (2002):
Effects of carprofen (R and S enantiomers and racemate) on the production of IL-1, IL-6 and TNF-
alpha by equine chondrocytes and synoviocytes.
Journal of veterinary pharmacology and therapeutics 25, 145-153
AUTEFAGE, A., M. ALVINERIE and P. L. TOUTAIN (1986):
Synovial fluid and plasma kinetics of methylprednisolone and methylprednisolone acetate in horses
following intra-articular administration of methylprednisolone acetate.
Equine veterinary journal 18, 193-198
AXELROD, L. (1993):
Glucocorticoids.
In: Textbook of Rheumatology
Saunders, Philadelphia, pp. 779-796
BÄUMER, W., A. MERTENS, M. BRAUN and M. KIETZMANN (2002):
The isolated perfused bovine uterus as a model for mucous membrane irritation and inflammation.
Altex 19, 57-63
BEAUMIER, P., J. FENWICK, L. YOUNG, S. HAZEN, K. HAMA, B. LAVIOLETTE, Z. DEOCAMPO, H.
PARK, N. SIO, S. TIMMINGS and F. TODI (1983):
Salicylic Acid in the Horse: Studies Conducted for Agriculture Canada.
In: Proceedings of the 5th International Conference on the Control of the Use of Drugs in Racehorses.
Toronto, pp. 12-20
BERTONE, A. L., J. L. PALMER and J. JONES (2001):
Synovial fluid cytokines and eicosanoids as markers of joint disease in horses.
Veterinary surgery 30, 528-538
BERTONE, J. J. and L. J. HORSPOOL (2004):
Appendix.
In: Equine Clinical Pharmacology
Saunders, pp. 367 - 380

REFERENCES
68
BISHOP, Y. (2005):
In: The Veterinary Formulary.
Pharmaceutical Press, London
BOUMPAS, D. T. and R. L. WILDER (2001):
Corticosteroids.
In: Arthritis and Allied Conditions
Williams and Wilkins, Philadelphia, S. 827-847
BRISTOL, D. G., J. E. RIVIERE, N. A. MONTEIRO-RIVIERE, K. F. BOWMAN and R. A. ROGERS
(1991):
The isolated perfused equine skin flap. Preparation and metabolic parameters.
Veterinary surgery 20, 424-433
BRISTON, L., J. DUDHIA and P. LEES (2009):
Age-related differences in prostaglandin E2 synthesis by equine cartilage explants and synoviocytes.
Journal of veterinary pharmacology and therapeutics 33, 268-276
BROOME, T. A., M. P. BROWN, R. R. GRONWALL, M. F. CASEY and K. A. MERITT (2003):
Pharmacokinetics and plasma concentrations of acetylsalicylic acid after intravenous, rectal, and
intragastric administration to horses.
Canadian journal of veterinary research 67, 297-302
BRUNE, K. and W. S. BECK (1991):
Towards safer nonsteroidal anti-inflammatory drugs.
Agents and actions 32, 13-25
BUNTENKÖTTER, K. (2012):
Untersuchung zur Pharmakokinetik von Salicylsäure nach oraler Applikation von Acetylsalicylsäure
beim Pferd.
Thesis, University of Veterinary Medicine Hannover, Department of Pharmacology, Toxicology and
Pharmacy
BYRON, C. R., M. C. STEWART, A. A. STEWART and H. C. PONDENIS (2008):
Effects of clinically relevant concentrations of glucosamine on equine chondrocytes and synoviocytes
in vitro.
American journal of veterinary research 69, 1129-1134

REFERENCES
69
CARON, J. P. (2005):
Intra-articular injections for joint disease in horses.
The Veterinary clinics of North America 21, 559-573
CARVER, M. P., P. L. WILLIAMS and J. E. RIVIERE (1989):
The isolated perfused porcine skin flap. III. Percutaneous absorption pharmacokinetics of
organophosphates, steroids, benzoic acid, and caffeine.
Toxicology and applied pharmacology 97, 324-337
CHEN, C. L., J. A. SAILOR, J. COLLIER and J. WIEGAND (1992):
Synovial and serum levels of triamcinolone following intra-articular administration of triamcinolone
acetonide in the horse.
Journal of veterinary pharmacology and therapeutics 15, 240-246
COLLIER, H. O. (1971):
Prostaglandins and aspirin.
Nature 232, 17-19
CREAMER, P. (1999):
Intra-articular corticosteroid treatment in osteoarthritis.
Current opinion in rheumatology 11, 417-421
CROMLISH, W. A. and B. P. KENNEDY (1996):
Selective inhibition of cyclooxygenase-1 and -2 using intact insect cell assays.
Biochemical pharmacology 52, 1777-1785
CRYER, B. and M. FELDMAN (1998):
Cyclooxygenase-1 and cyclooxygenase-2 selectivity of widely used nonsteroidal anti-inflammatory
drugs.
The American journal of medicine 104, 413-421
DE GRAUW, J. C., C. H. VAN DE LEST and P. R. VAN WEEREN (2009):
Inflammatory mediators and cartilage biomarkers in synovial fluid after a single inflammatory insult: a
longitudinal experimental study.
Arthritis research & therapy 11, R 35

REFERENCES
70
DERENDORF, H., H. MOLLMANN, A. GRUNER, D. HAACK and G. GYSELBY (1986):
Pharmacokinetics and pharmacodynamics of glucocorticoid suspensions after intra-articular
administration.
Clinical pharmacology and therapeutics 39, 313-317
DIKKER, L. (2009)
Untersuchung zur Pharmakokinetik von Betamethason nach intravenöser und dermaler Applikation
hinsichtlich der Dopingrelevanz beim Pferd.
Thesis, University of Veterinary Medicine Hannover, Department of Pharmacology, Toxicology and
Pharmacy
DINGLE, J. T., J. SAKLATVALA, R. HEMBRY, J. TYLER, H. B. FELL and R. JUBB (1979):
A cartilage catabolic factor from synovium.
The Biochemical journal 184, 177-180
DULIN, J. A., W. T. DROST, M. A. PHELPS, E. M. SANTSCHI, M. I. MENENDEZ and A. L.
BERTONE (2012):
Influence of exercise on the distribution of technetium Tc 99m medronate following intra-articular
injection in horses.
American journal of veterinary research 73, 418-425
FERGUSON, D. C., L. DIRIKOLU and M. HOENIG (2009):
Glucocorticoids, mineralocorticoids and adrenolytic drugs.
In: Veterinary Pharmacology and Therapeutics
Wiley-Blackwell, pp. 771 - 793
FRASER, C. M., A. MAYS, H. E. AMSTUTZ, J. ARCHIBALD and J. ARMOUR (1986):
In: The Merck Veterinary Manual: a Handbook of Diagnosis, Therapy and Disease Prevention and
Control for the Veterinarian.
Merck & Co Inc, Rahway (USA), p. 1677
FREAN, S. P. and P. LEES (2000):
Effects of polysulfated glycosaminoglycan and hyaluronan on prostaglandin E2 production by cultured
equine synoviocytes.
American journal of veterinary research 61, 499-505

REFERENCES
71
FRISBIE, D. D. (2011):
Medical Treatment of Joint Disease In: Equine Surgery
Saunders Elsevier, pp. 1114-1117
GILROY, D. W., P. R. COLVILLE-NASH, D. WILLIS, J. CHIVERS, M. J. PAUL-CLARK and D. A.
WILLOUGHBY (1999):
Inducible cyclooxygenase may have anti-inflammatory properties.
Nature medicine 5, 698-701
GIULIANO, F. and T. D. WARNER (1999):
Ex vivo assay to determine the cyclooxygenase selectivity of non-steroidal anti-inflammatory drugs.
British journal of pharmacology 126, 1824-1830
GOODRICH, L. R. and A. J. NIXON (2006):
Medical treatment of osteoarthritis in the horse - a review.
Veterinary journal 171, 51-69
GRAY, R. G. and N. L. GOTTLIEB (1983):
Intra-articular corticosteroids. An updated assessment.
Clinical orthopaedics and related research 177, 235-263
GRONEBERG, D. A., C. GROSSE-SIESTRUP and A. FISCHER (2002):
In vitro models to study hepatotoxicity.
Toxicologic pathology 30, 394-399
GROSSE-SIESTRUP, C., C. FEHRENBERG, H. VON BAEYER and D. A. GRONEBERG (2002):
Multiple-organ harvesting for models of isolated hemoperfused organs of slaughtered pigs.
Altex 19, 9-13
HAMEED, F. and J. IHM (2012):
Injectable medications for osteoarthritis.
Physical medicine and rehabilitation 4, 75-81
HARKINS, J. D., J. M. CARNEY and T. TOBIN (1993):
Clinical use and characteristics of the corticosteroids.
The Veterinary clinics of North America 9, 543-562

REFERENCES
72
HIGGINS, A. J. and P. LEES (1984):
The acute inflammatory process, arachidonic acid metabolism and the mode of action of anti-
inflammatory drugs.
Equine veterinary journal 16, 163-175
HIGGINS, A. J., P. LEES and A. D. SEDGWICK (1987):
Development of equine models of inflammation. The Ciba-Geigy Prize for Research in Animal Health.
The Veterinary record 120, 517-522
HINZ, B., V. KRAUS, A. PAHL and K. BRUNE (2000):
Salicylate metabolites inhibit cyclooxygenase-2-dependent prostaglandin E2 synthesis in murine
macrophages.
Biochemical and biophysical research communications 274, 197-202
HSU, W. H. (2008):
Endocrine Pharmacology In: Handbook of Veterinary Pharmacology
Wiley-Blackwell, pp. 264-267
HSU, W. H. and A. KANTHASAMY (2008):
Nonsteroidal Anti-inflammatory Drugs.
In: Handbook of Veterinary Pharmacology
Wiley-Blackwell, pp. 153 - 168
HULKOWER, K. I., J. W. COFFEY, W. LEVIN, C. M. ANDERSON, T. CHEN, W. C. HOPE, D. R.
BOLIN and D. W. MORGAN (1993):
Interleukin-1 beta induces cytosolic PLA2 in parallel with prostaglandin E2 in rheumatoid synovial
fibroblasts.
Agents Actions 39 Spec No, C5-7
IWANAGA, T., M. SHIKICHI, H. KITAMURA, H. YANASE and K. NOZAWA-INOUE (2000):
Morphology and functional roles of synoviocytes in the joint.
Archives of Histology and Cytology 63 (1), 17-31.
KAMERLING, S. G. and J. G. OWENS (1994):
Models for assessing the relationship between drug concentration and drug effect in performance
horses.
The British veterinary journal 150, 507-525

REFERENCES
73
KANTROWITZ, F., D. R. ROBINSON, M. B. MCGUIRE and L. LEVINE (1975):
Corticosteroids inhibit prostaglandin production by rheumatiod synovia.
Nature 258, 737-739
KIETZMANN, M., W. LÖSCHER, D. ARENS, P. MAASS and D. LUBACH (1993):
The isolated perfused bovine udder as an in vitro model of percutaneous drug absorption. Skin
viability and percutaneous absorption of dexamethasone, benzoyl peroxide, and etofenamate.
Journal of pharmacological and toxicological methods 30, 75-84
KIETZMANN, M., R. SCHERKL and R. SCHULZ (2002):
Pharmakologie der Entzündung und der Allergie.
In: Lehrbuch der Pharmakologie und Toxikologie für die Veterinärmedizin
Enke, Stuttgart, pp. 318 - 328
KIRKER-HEAD, C. A., V. K. CHANDNA, R. K. AGARWAL, E. A. MORRIS, A. TIDWELL, M. W.
O'CALLAGHAN, W. RAND and M. S. KUMAR (2000):
Concentrations of substance P and prostaglandin E2 in synovial fluid of normal and abnormal joints of
horses.
American journal of veterinary research 61, 714-718
KITAMURA, H. P., H. YANASE, H. KITAMURA and T. IWANAGA (1999):
Unique localization of protein gene product 9.5 in type B synoviocytes in the joints of the horse.
Journal of histochemistry and cytochemistry 47, 343-352
KLAUS, A. M. and H. J. HAPKE (1996):
[Natural and synthetic glucocorticoids in the racing horse: a review of the literature].
Deutsche tierärztliche Wochenschrift 103, 494-500
KOLLIAS-BAKER, C. and K. COX (2004):
Non-steroidal anti-inflammatory drugs.
In: Equine Clinical Pharmacology
Saunders, pp. 247-266
LAKHANI, K. H., M. LAMBERT, F. SLUYTER, R. DEVOLZ, G. MAYLIN and A. J. HIGGINS (2004):
Estimation of the critical threshold value for presence of salicylic acid in the urine of thoroughbred
horses.
In: Proceedings of the 15th International Conference of Racing Analysts and Veterinarians, Dubai, 67-
77

REFERENCES
74
LANDONI, M. F., F. M. CUNNINGHAM and P. LEES (1995):
Comparative pharmacodynamics of flunixin, ketoprofen and tolfenamic acid in calves.
The Veterinary record 137, 428-431
LANDONI, M. F., R. FOOT, S. FREAN and P. LEES (1996):
Effects of flunixin, tolfenamic acid, R(-) and S(+) ketoprofen on the response of equine synoviocytes to
lipopolysaccharide stimulation.
Equine veterinary journal 28, 468-475
LANDONI, M. F. and P. LEES (1995):
Comparison of the anti-inflammatory actions of flunixin and ketoprofen in horses applying PK/PD
modelling.
Equine veterinary journal 27, 247-256
LANGENDORFF, O. (1895):
Untersuchungen am überlebenden Säugethierherzen.
Arch Ges Physiol 61, 291-332
LEES, P. (2003):
Pharmacology of drugs used to treat osteoarthritis in veterinary practice.
Inflammopharmacology 11, 385-399
LEES, P. (2009):
Analgesic, antiinflammatory, antipyretic drugs.
In: Veterinary Pharmacology and Therapeutics
Wiley-Blackwell, pp. 457 - 486
LEES, P., C. P. EWINS, J. B. TAYLOR and A. D. SEDGWICK (1987a):
Serum thromboxane in the horse and its inhibition by aspirin, phenylbutazone and flunixin.
The British veterinary journal 143, 462-476
LEES, P., J. B. TAYLOR, A. J. HIGGINS and S. C. SHARMA (1986):
Phenylbutazone and oxyphenbutazone distribution into tissue fluids in the horse.
Journal of veterinary pharmacology and therapeutics 9, 204-212
LEES, P., J. B. TAYLOR, T. E. MAITHO, J. D. MILLAR and A. J. HIGGINS (1987b):
Metabolism, excretion, pharmacokinetics and tissue residues of phenylbutazone in the horse.
The Cornell veterinarian 77, 192-211

REFERENCES
75
LILLICH, J. D., A. L. BERTONE, L. M. SCHMALL, A. J. RUGGLES and R. A. SAMS (1996):
Plasma, urine, and synovial fluid disposition of methylprednisolone acetate and isoflupredone acetate
after intra-articular administration in horses.
American journal of veterinary research 57, 187-192
LINDSLEY, H. B. and D. D. SMITH (1990):
Enhanced prostaglandin E2 secretion by cytokine-stimulated human synoviocytes in the presence of
subtherapeutic concentrations of nonsteroidal antiinflammatory drugs.
Arthritis and rheumatism 33, 1162-1169
LÖSCHER, W., F. R. UNGEMACH and R. KROKER (2006):
Pharmakotherapie bei Haus- und Nutztieren.
Enke Verlag, Stuttgart
LÖVENICH, B. (2012)
Pharmacokinetics of betamethasone and triamcinolone acetonide after intra-articular injection
regarding doping relevance in the horse.
Thesis, University of Veterinary Medicine Hannover, Department of Pharmacology, Toxicology and
Pharmacy
MACHNIK, M., W. LÖHLEIN, Y. SCHRADER, K. KÖHLER, J. FREYMARK and W. SCHÄNZER
(2007):
Behandlung von Sportpferden mit Corticosteroiden: Nachweiszeiten von Flumethason und
Betamethason beim Pferd nach i.v. Applikation von Acutol und i.a. Applikation von Celestovet.
Pferdeheilkunde 5, 476 - 480
MALMBERG, A. B. and T. L. YAKSH (1992):
Hyperalgesia mediated by spinal glutamate or substance P receptor blocked by spinal
cyclooxygenase inhibition.
Science 257, 1276-1279
MAY, S. A., R. E. HOOKE and P. LEES (1989):
Identity of the E-series prostaglandin produced by equine chondrocytes and synovial cells in response
to a variety of stimuli.
Research in veterinary science 46, 54-57

REFERENCES
76
MAY, S. A., R. E. HOOKE, K. Y. PEREMANS, F. VERSHOOTEN and P. LEES (1994):
Prostaglandin E2 in equine joint disease.
Vlaams Diergeneeskd Tijdschr 63, 187-191
MAY, S. A. and P. LEES (1996):
Nonsteroidal anti-inflammatory drugs.
In: Joint disease in the horse
Saunders, Philadelphia, pp. 223 - 237
MEADE, E. A., W. L. SMITH and D. L. DEWITT (1993):
Differential inhibition of prostaglandin endoperoxide synthase (cyclooxygenase) isozymes by aspirin
and other non-steroidal anti-inflammatory drugs.
The Journal of biological chemistry 268, 6610-6614
MEULYZER, M., P. VACHON, F. BEAUDRY, T. VINARDELL, H. RICHARD, G. BEAUCHAMP and S.
LAVERTY (2009):
Joint inflammation increases glucosamine levels attained in synovial fluid following oral administration
of glucosamine hydrochloride.
Osteoarthritis and cartilage / OARS, Osteoarthritis Research Society 17, 228-234
MILLER, L. L., C. G. BLY, M. L. WATSON and W. F. BALE (1951):
The dominant role of the liver in plasma protein synthesis; a direct study of the isolated perfused rat
liver with the aid of lysine-epsilon-C14.
The Journal of experimental medicine 94, 431-453
MITCHELL, J. A., P. AKARASEREENONT, C. THIEMERMANN, R. J. FLOWER and J. R. VANE
(1993):
Selectivity of nonsteroidal antiinflammatory drugs as inhibitors of constitutive and inducible
cyclooxygenase.
Proceedings of the National Academy of Sciences of the United States of America 90, 11693-11697
MORTON, A. J., N. B. CAMPBELL, J. M. GAYLE, W. R. REDDING and A. T. BLIKSLAGER (2005):
Preferential and non-selective cyclooxygenase inhibitors reduce inflammation during
lipopolysaccharide-induced synovitis.
Research in veterinary science 78, 189-192

REFERENCES
77
MOSES, V. S., J. HARDY, A. L. BERTONE and S. E. WEISBRODE (2001):
Effects of anti-inflammatory drugs on lipopolysaccharide-challenged and -unchallenged equine
synovial explants.
American journal of veterinary research 62, 54-60
OKADA, Y., M. SHINMEI, O. TANAKA, K. NAKA, A. KIMURA, I. NAKANISHI, M. T. BAYLISS, K.
IWATA and H. NAGASE (1992):
Localization of matrix metalloproteinase 3 (stromelysin) in osteoarthritic cartilage and synovium.
Laboratory investigation 66, 680-690
OWENS, J. G., S. G. KAMERLING and S. A. BARKER (1995):
Pharmacokinetics of ketoprofen in healthy horses and horses with acute synovitis.
Journal of veterinary pharmacology and therapeutics 18, 187-195
OWENS, J. G., S. G. KAMERLING, S. R. STANTON, M. L. KEOWEN and J. S. PRESCOTT-
MATHEWS (1996):
Effects of pretreatment with ketoprofen and phenylbutazone on experimentally induced synovitis in
horses.
American journal of veterinary research 57, 866-874
PALMER, J. L. and A. L. BERTONE (1994a):
Experimentally-induced synovitis as a model for acute synovitis in the horse.
Equine veterinary journal 26, 492-495
PALMER, J. L. and A. L. BERTONE (1994b):
Joint structure, biochemistry and biochemical disequilibrium in synovitis and equine joint disease.
Equine veterinary journal 26, 263-277
PATAN-ZUGAJ, B., F. C. GAUFF and T. F. LICKA (2012):
Effects of the addition of endotoxin during perfusion of isolated forelimbs of equine cadavers.
American journal of veterinary research 73, 1462-1468
PATAN, B., K. D. BUDRAS and T. F. LICKA (2009):
Effects of long-term extracorporeal blood perfusion of the distal portion of isolated equine forelimbs on
metabolic variables and morphology of laminar tissue.
American journal of veterinary research 70, 669-677

REFERENCES
78
PLUMB, D. C. (2008):
In: Veterinary Drug Handbook.
Wiley-Blackwell
POPOT, M. A., P. GARCIA, Y. MOULARD and Y. BONNAIRE (2002):
Detection of triamcinolone, betamethasone and methylprednisolone in the horse: Effect on urine
cortisol and cortisol metabolite concentrations.
In: Proceedings of the 14th international conference of racing analysts and veterinarians, Florida, USA
pp. 408 - 412
RAEKALLIO, M., P. M. TAYLOR and R. C. BENNETT (1997):
Preliminary investigations of pain and analgesia assessment in horses administered phenylbutazone
or placebo after arthroscopic surgery.
Veterinary surgery 26, 150-155
RICKETTS, A. P., K. M. LUNDY and S. B. SEIBEL (1998):
Evaluation of selective inhibition of canine cyclooxygenase 1 and 2 by carprofen and other
nonsteroidal anti-inflammatory drugs.
American journal of veterinary research 59, 1441-1446
RIVIERE, J. E., K. F. BOWMAN, N. A. MONTEIRO-RIVIERE, L. P. DIX and M. P. CARVER (1986):
The isolated perfused porcine skin flap (IPPSF). I. A novel in vitro model for percutaneous absorption
and cutaneous toxicology studies.
Fundamental and applied toxicology 7, 444-453
RIVIERE, J. E. and N. A. MONTEIRO-RIVIERE (1991):
The isolated perfused porcine skin flap as an in vitro model for percutaneous absorption and
cutaneous toxicology.
Critical reviews in toxicology 21, 329-344
ROBINSON, N. E., J. M. DABNEY, W. J. WEIDNER, G. A. JONES and J. B. SCOTT (1975):
Vascular responses in the equine digit.
American journal of veterinary research 36, 1249-1253
RUSSELL, W. M. S. and R. L. BURCH (1959):
The Principles of Humane Experimental Technique.
Methuen, London, UK.

REFERENCES
79
SAKLATVALA, J. and J. T. DINGLE (1980):
Identification of catabolin, a protein fro synovium which induces degradation of cartilage in organ
culture.
Biochemical and biophysical research communications 96, 1225-1231
SHOLKOFF, S. D., E. J. EYRING, M. ROWLAND and S. RIEGELMAN (1967):
Plasma and synovial fluid concentrations of acetylsalicylic acid in patients with rheumatoid arthritis.
Arthritis and rheumatism 10, 348-351
SMITH, C. J., Y. ZHANG, C. M. KOBOLDT, J. MUHAMMAD, B. S. ZWEIFEL, A. SHAFFER, J. J.
TALLEY, J. L. MASFERRER, K. SEIBERT and P. C. ISAKSON (1998):
Pharmacological analysis of cyclooxygenase-1 in inflammation.
Proceedings of the National Academy of Sciences of the United States of America 95, 13313-13318
SOMA, L. R., E. BEHREND, J. RUDY and R. W. SWEENEY (1988):
Disposition and excretion of flunixin meglumine in horses.
American journal of veterinary research 49, 1894-1898
SOMA, L. R., C. E. UBOH, Y. LUO, F. GUAN, P. J. MOATE and R. C. BOSTON (2006):
Pharmacokinetics of methylprednisolone acetate after intra-articular administration and its effect on
endogenous hydrocortisone and cortisone secretion in horses.
American journal of veterinary research 67, 654-662
SOMA, L. R., C. E. UBOH, Y. YOU, F. GUAN and R. C. BOSTON (2011):
Pharmacokinetics of intra-articular, intravenous, and intramuscular administration of triamcinolone
acetonide and its effect on endogenous plasma hydrocortisone and cortisone concentrations in
horses.
American journal of veterinary research 72, 1234-1242
SOREN, A. (1977):
Dissociation of acetylsalicylic acid in blood and joint fluid.
Scandinavian journal of rheumatology 6, 17-22
TAYLOR, J. B., A. WALLAND, P. LEES, E. L. GERRING, T. E. MAITHO and J. D. MILLAR (1983):
Biochemical and haematological effects of a revised dosage schedule of phenylbutazone in horses.
The Veterinary record 112, 599-602

REFERENCES
80
TOUTAIN, P. L. (2009):
Mechanisms of drug action and pharmacokinetics/pharmacodynamics integration in dosage regimen
optimization for veterinary medicine.
In: Veterinary Pharmacology and Therapeutics
Wiley-Blackwell, pp. 75-97
TOUTAIN, P. L. (2010a):
How to extrapolate a withdrawal time from an EHSLC published detection time: a Monte Carlo
simulation appraisal.
Equine veterinary journal 42, 248-254
TOUTAIN, P. L. (2010b):
Veterinary medicines and competition animals: the question of medication versus doping control.
Handbook of experimental pharmacology 199, 315-339
TOUTAIN, P. L., A. AUTEFAGE, C. LEGRAND and M. ALVINERIE (1994):
Plasma concentrations and therapeutic efficacy of phenylbutazone and flunixin meglumine in the
horse: pharmacokinetic/pharmacodynamic modelling.
Journal of veterinary pharmacology and therapeutics 17, 459-469
TOUTAIN, P. L. and C. C. CESTER (2004):
Pharmacokinetic-pharmacodynamic relationships and dose response to meloxicam in horses with
induced arthritis in the right carpal joint.
American journal of veterinary research 65, 1533-1541
TROTTER, G. W. (1996):
Intra-articular corticosteroids.
In: Joint disease in the horse
Saunders, Philadelphia, pp. 237 - 256
TROTTER, G. W., C. W. MCILWRAITH, J. V. YOVICH, R. W. NORRDIN, R. H. WRIGLEY and C. H.
LAMAR (1991):
Effects of intra-articular administration of methylprednisolone acetate on normal equine articular
cartilage.
American journal of veterinary research 52, 83-87

REFERENCES
81
UNGEMACH, F. R. (2006):
Pharmaka zur Beeinflussung von Entzündungen.
In: Pharmakotherpapie bei Haus- und Nutztieren
Parey, Stuttgart, pp. 364-403
VANE, J. R. and R. M. BOTTING (1995):
New insights into the mode of action of anti-inflammatory drugs.
Inflammation research 44, 1-10
VETIDATA (2012)
Glukokortikoide
www.vetidata.de, 21.12.2012
VINE, J. S., N; GLOWACKI, L; KRSTEVSKA, D; ZAHRA, P; O'CALLAGHAN, P (2006):
The metabolism and excretion of betamethasone sodium phosphate and betamethasone acetate in
the horse after intra-articular injection.
In: Proceedings of the 16th International Conference of Racing Analysts and Veterinarians
Tokyo, Japan, pp. 167-171
VON BAEYER, H., K. STAHL, M. HAUSLER, M. MEISSLER, V. UNGER, J. FRANK, C. GROSSE-
SIESTRUP, G. KACZMARCZYK, K. AFFELD, H. J. FLAIG and B. STEINBACH (1997):
[A new method of ex vivo whole blood perfusion of isolated mammalian organs, exemplified by the
kidney of swine].
Biomedizinische Technik 42, 61-68
WAGNER, S. M., A. C. NOGUEIRA, M. PAUL, D. HEYDECK, S. KLUG and B. CHRIST (2003):
The isolated normothermic hemoperfused porcine forelimb as a test system for transdermal
absorption studies.
The international journal of artificial organs 6, 183-191
WARD, L. C. and P. J. BUTTERY (1979):
The patho-physiological basis for tests of viability in isolated perfused organs.
Biomedicine 30, 181-186
WEISS, C., H. PASSOW and A. ROTHSTEIN (1959):
Autoregulation of flow in isolated rat kidney in the absence of red cells.
The American journal of physiology 196, 1115-1118

REFERENCES
82
WHEAT, J. D. (1955):
The use of hydrocortisone in the treatment of joint and tendon disorders in large animals.
Journal of the American Veterinary Medical Association 127, 64-67
WILLIAMS, P. L., M. P. CARVER and J. E. RIVIERE (1990):
A physiologically relevant pharmacokinetic model of xenobiotic percutaneous absorption utilizing the
isolated perfused porcine skin flap.
Journal of pharmaceutical sciences 79, 305-311
WITTENBERG, R. H., R. E. WILLBURGER, K. S. KLEEMEYER and B. A. PESKAR (1993):
In vitro release of prostaglandins and leukotrienes from synovial tissue, cartilage, and bone in
degenerative joint diseases.
Arthritis and rheumatism 36, 1444-1450
WONGAUMNUAYKUL, S., C. SIEDLER, H. SCHOBESBERGER and C. STANEK (2006):
Doppler sonographic evaluation of the digital blood flow in horses with laminitis or septic
pododermatitis.
Veterinary radiology & ultrasound 47, 199-205
WRIGHT, J. M., C. G. KNIGHT and I. M. HUNNEYBALL (1986):
The effect of side chain structure on the biochemical and therapeutic properties of intra-articular
dexamethasone 21-esters.
Clinical and experimental rheumatology 4, 331-339
YARBROUGH, T. (2004):
Intraarticular medication.
In: Equine Clinical Pharmacology
Saunders, pp. 121-134
APPENDIX
83
10 Appendix
10.1 Cell culture experiments
10.1.1 Cell viability
TABLE 8: Results of the modified MTS-test following synoviocyte stimulation with LPS and treatment
with BM, ASA or SA, measured as spectrophotometrical extinction. Control: Stimulation with LPS and
vehicle treatment. Each value is a mean of 4 wells. SD = Standard deviation.
Horse # 1 2 3 4 5 6 Mean SD
Control 1.065 1.024 0.726 0.858 1.156 0.782 0.935 0.171
BM (mol/L)
2.55x10-8
0.750 1.0785 0.751 0.909 1.078 0.732 0.883 0.164
2.55x10-9
0.744 0.494 0.744 0.974 1.046 0.749 0.792 0.196
2.55x10-10
0.756 0.678 0.743 0.840 1.162 0.811 0.832 0.171
ASA (mol/L)
7.77x10-5
0.710 0.941 0.771 0.866 1.126 0.761 0.863 0.153
7.77x10-6
0.692 0.939 0.707 0.831 1.054 0.762 0.831 0.142
7.77x10-7
0.800 0.991 0.718 0.893 1.060 0.756 0.870 0.136
3.88x10-6
0.730 0.926 0.756 0.917 1.058 0.757 0.857 0.130
SA (mol/L)
1.23x10-3
0.668 1.056 0.731 0.875 1.019 0.745 0.849 0.161
1.23x10-4
0.729 0.895 0.808 0.965 1.152 0.767 0.886 0.156
1.23x10-5
0.719 0.970 0.883 0.711 1.182 0.804 0.878 0.179

APPENDIX
84
10.1.2 PGE2 concentrations
TABLE 9: PGE2 concentration (ng/mL) in the culture supernatant 24 h after stimulation with LPS and
treatment with BM, ASA or SA. Control: Stimulation with LPS and vehicle treatment. Each
concentration is a mean of 4 wells. SD = Standard deviation.
Horse # 1 2 3 4 5 6 Mean SD
Control 58 82 112 68 43 73 73 23
BM (mol/L)
2.55x10-8
13 10 52 27 15 40 26 17
2.55x10-9
39 12 68 36 23 55 39 23
2.55x10-10
63 83 103 65 43 79 73 25
ASA (mol/L)
7.77x10-5
2 5 9 4 2 4 4 3
7.77x10-6
28 48 62 29 23 47 40 15
7.77x10-7
41 68 103 60 30 61 61 25
3.88x10-6
53 56 75 61 28 47 53 17
SA (mol/L)
1.23x10-3
33 60 73 60 25 55 51 18
1.23x10-4
45 80 86 65 35 60 62 20
1.23x10-5
37 90 92 68 46 61 66 22

APPENDIX
85
10.2 Maren Friebe’s contribution to the manuscripts
1. The isolated perfused equine distal limb as an ex vivo model for pharmacokinetic studies:
Participation in the study design
Performance of the experiments
Writing of the manuscript.
2. Synovial distribution of “systemically” administered acetylsalicylic acid in the isolated perfused
equine distal limb
Participation in the study design
Performance of the experiments
Writing of the manuscript

THANK YOU
Danksagung
Herrn Prof. Dr. M. Kietzmann danke ich herzlich für die Überlassung des interessanten
Themas, die gute und stets freundliche Betreuung, die fachlichen Denkanstöße und
schließlich die konstruktive Kritik bei der Anfertigung dieser Arbeit.
Herrn Prof. Dr. K. Feige und PD Dr. F. Thorey danke ich für die Ratschläge und
Anregungen im Rahmen der Betreuergruppe.
Der Deutschen Reiterlichen Vereinigung (FN), insbesondere Herrn Dr. Michael Düe,
danke ich für die finanzielle Unterstützung der Studie.
Allen Mitarbeitern der Arbeitsgruppe Toxikologie danke ich für die abwechslungsreichen
drei Jahre. Speziell möchte ich Frau Dr. Jessica Stahl für den Beistand rund um die
HPLC sowie die jederzeit hilfsbereite Art und Herrn Stephan Schumacher, PhD, für die
umfangreiche Einführung in Mikrodialyse und Statistik danken. Viktoria Garder und
Caroline Schulz danke ich für die souveräne Unterstützung in allen größeren und
kleineren Fragen und die unterhaltsame Zeit. Danken für die Hilfe im Labor möchte ich
außerdem Hans-Herbert Bohr, Theiss Wystemp und Alina Narten.
Meinen Mitdoktoranden im Laufe der Jahre (auch denen aus der 1. Etage), ganz
besonders der aktuellen Besetzung des „kleinen“ Doktorandenzimmers Isabelle Roth und
Stefanie Mielke, danke ich für die fachlichen und nicht fachlichen Gespräche und
Ratschläge, den Spaß und das Gefühl, nicht alleine zu sein.
Sonja, Maike, Hellen, Kathrin, Katha & Anne-Sophie danke ich für ihre Freundschaft seit
dem Studium und viele schöne Erinnerungen an unsere gemeinsame Zeit in Hannover.
Meinen Eltern und meinem Bruder Thomas danke ich von Herzen für die Unterstützung
während meines Studiums und meiner Doktorarbeit. Ohne Euch wäre das alles nicht
möglich gewesen.
Meinem lieben Jan danke ich für die Zuneigung und die Ablenkung und dafür, dass er
immer an mich glaubt. Ich freue mich auf die Zukunft mit Dir!