cytochromep4503aenzymescatalyzethe 6-demethyla ... · pdf fileanother study demonstrated...
TRANSCRIPT

Cytochrome P450 3A Enzymes Catalyze the O6-Demethyla-tion of Thebaine, a Key Step in Endogenous MammalianMorphine Biosynthesis*
Received for publication, May 14, 2015, and in revised form, June 17, 2015 Published, JBC Papers in Press, July 8, 2015, DOI 10.1074/jbc.M115.665331
Valerie M. Kramlinger, Monica Alvarado Rojas, Tatsuyuki Kanamori, and F. Peter Guengerich1
From the Department of Biochemistry, School of Medicine, Vanderbilt University School of Medicine,Nashville, Tennessee 37232-0146
Background: Mammals synthesize endogenous morphine; the enzyme catalyzing thebaine O6-demethylation, a key latestep, is uncharacterized.Results: Human cytochromes P450 (P450) 3A4 and 3A5 catalyzed thebaine O6-demethylation; the P450 3A-selective drugketoconazole inhibited the reaction in human liver microsomes and rat brain homogenates.Conclusion: P450s 3A4 and 3A5 catalyze thebaine O6-demethylation in humans.Significance: Enzymes catalyzing all oxidations in the latter steps of mammalian morphine biosynthesis have been identified.
Morphine, first characterized in opium from the poppyPapaver somniferum, is one of the strongest known analgesics.Endogenous morphine has been identified in several mamma-lian cells and tissues. The synthetic pathway of morphine in theopium poppy has been elucidated. The presence of commonintermediates in plants and mammals suggests that biosynthesisoccurs through similar pathways (beginning with the amino acidL-tyrosine), and the pathway has been completely delineated inplants. Some of the enzymes in the mammalian pathway havebeen identified and characterized. Two of the latter steps in themorphine biosynthesis pathway are demethylation of thebaineat the O3- and the O6-positions, the latter of which has beendifficult to demonstrate. The plant enzymes responsible forboth the O3-demethylation and the O6-demethylation are mem-bers of the FeII/�-ketoglutarate-dependent dioxygenase family.Previous studies showed that human cytochrome P450 (P450)2D6 can catalyze thebaine O3-demethylation. We report thatdemethylation of thebaine at the O6-position is selectively cata-lyzed by human P450s 3A4 and 3A5, with the latter being moreefficient,andratP4503A2.OurresultsdonotsupportO6-demeth-ylation of thebaine by an FeII/�-ketoglutarate-dependent dioxy-genase. In rat brain microsomes, O6-demethylation was inhib-ited by ketoconazole, but not sulfaphenazole, suggesting thatP450 3A enzymes are responsible for this activity in the brain.An alternate pathway to morphine, oripavine O6-demethyla-tion, was not detected. The major enzymatic steps in mamma-lian morphine synthesis have now been identified.
Morphine is the principal benzylisoquinoline alkaloid pro-duced in the opium poppy, Papaver somniferum. Its mecha-
nism of action is as an agonist for the �-opioid receptors thatare distributed throughout the brain. Activation of these recep-tors is associated with analgesia, sedation, euphoria, physicaldependence, and respiratory depression. The �-binding sitesare discretely distributed in the human brain, with high densi-ties in the posterior amygdala, hypothalamus, thalamus,nucleus caudatus, putamen, and certain cortical areas (1).
A considerable body of evidence exists that morphine is pres-ent in the tissues of various animals that have not been medi-cated with morphine or other related opioids. In mammals,morphine has been detected in skin, lung, spinal cord, and,most notably, in the brain (2– 4). The presence of morphine inbrain is of particular interest due to the presence of the �-opi-oid receptors. In the rat, morphine levels in structures of thebrain have been quantified and range from 26 fmol/g of tissue(found globally in the brain) to 7.2 pmol/g of tissue measured inthe hypothalamus (3, 5–7). One study quantified morphine inthe temporal lobe from one human brain tissue at 3.4 ng/g oftissue (8). The presence of morphine in brain tissue in manymammals and the evidence from a human sample suggest thatendogenous morphine is present in human brain.
Endogenous formation of morphine has been demonstratedin human cells in culture. In a seminal study, human neuroblas-toma cells incubated in the presence of 18O2 produced 18O-labeled morphine (9). Another study demonstrated morphinesynthesis by human white blood cells and polymorphonuclearcells in a precursor-dependent manner (10, 11). Takentogether, these studies suggest that morphine biosynthesis canoccur in the human brain. Humans have also been reported toexcrete both codeine and morphine in urine, in the absence oftreatment with these compounds (12).
The physiological role of endogenous morphine is currentlyunknown. The endogenous levels that have been quantified arebelow the plasma concentrations (�M range) achieved duringtherapeutic use (13). Many hypotheses have been presentedregarding the purpose of endogenous morphine that suggest arole in infection, sepsis, or inflammation, as well as neurologicalpathologies (Parkinson disease and schizophrenia) (14 –17).
* This work was supported in whole or part by National Institute of HealthGrants R37 CA090426 (to F. P. G.) and T32 E5007028 (to V. M. K.). Theauthors declare that they have no conflicts of interest with the contents ofthis article.
1 To whom correspondence should be addressed: Dept. of Biochemistry, Van-derbilt University School of Medicine, 638B Robinson Research Bldg., 2200Pierce Ave., Nashville, TN 37232-0146. Tel.: 615-322-2261; Fax: 615-343-0704; E-mail: [email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 290, NO. 33, pp. 20200 –20210, August 14, 2015© 2015 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
20200 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 33 • AUGUST 14, 2015
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Mammalian morphine biosynthesis appears to be highly sim-ilar to the pathway in the opium poppy, with at least somecommon intermediates (18 –20). The multistep biosyntheticpathway begins with the amino acid L-tyrosine, and the finalsteps in this pathway include the conversion of thebaine tomorphine via two critical demethylation steps at the O3- andO6-positions. Two parallel pathways exist for conversion of the-baine to morphine, in which O3- and O6-demethylation reac-tions occur, potentially in either order (Fig. 1) (21). Both ofthese pathways are plausible, in that thebaine, codeine, andoripavine are present in both mammals as well as the opiumpoppy (5, 22–24).
Although the intermediates are known and there is evidencethat some of the reactions occur in mammals (9, 25, 26), theenzymes that catalyze these demethylation steps have onlyrecently been identified. The enzymes that catalyze the O3- andO6-demethylation steps in the plant pathway are members ofthe FeII/�-ketoglutarate-dependent dioxygenase enzyme fam-ily (27). Specifically, in plants, thebaine O6-demethylase cata-lyzes O6-demethylation of both thebaine and oripavine;codeine O-demethylase catalyzes O3-demethylation of boththebaine and codeine (Fig. 1). Independent of plant studies, thediscovery that O3-demethylation of thebaine and codeine canbe catalyzed by the human enzyme cytochrome P450 (P450)2
2D6 (28) has focused our search for the O6-demethylase toP450s, but this enzyme(s) has remained unknown, and the FeII/�-ketoglutarate dioxygenases can be considered.
Both the P450 and the FeII/�-ketoglutarate-dependent di-oxygenase families are known to catalyze diverse sets of reac-tions and are candidates for mediating the O6-demethylation ofthebaine and oripavine. The P450 superfamily catalyzes theoxidation of most organic substances, generally using NADPHas an electron donor (29). The general reaction catalyzed byP450s is mixed function oxidation, manifestations of whichinclude carbon hydroxylation, dealkylation of heteroatomicsubstrates, heteroatom oxygenation, and the oxidation of
unsaturated compounds to epoxides and phenols (30). FeII/�-ketoglutarate-dependent dioxygenases constitute a large familyof soluble, non-heme iron-containing oxidases that couple thedecarboxylation of �-ketoglutarate to activation of dioxygen tocatalyze a variety of oxidation reactions. This family of enzymesmost frequently catalyzes carbon hydroxylation, but other for-mal two-electron oxidations such as desaturation, cyclization,and halogenation are also known (31, 32). Except for the twodioxygenases involved in the plant morphine biosynthesis(27, 33), O-demethylation by the FeII/�-ketoglutarate-depen-dent dioxygenases has not been previously reported, to ourknowledge.
The present study was designed to identify the enzyme(s)responsible for O6-demethylation of thebaine in mammals,humans in particular. Previous attempts to characterize theenzymes that catalyze O6-demethylation of thebaine andoripavine have been unsuccessful, in large part due to insta-bility of codeinone and morphinone in aqueous solution (34,35). In the present work, we devised and validated a coupledenzyme assay to circumvent this issue. We established thathuman P450s 3A4 and P450 3A5 and rat P450 3A2 catalyzeO6-demethylation of thebaine. Using rat brain microsomes,we also provide evidence that P450 3A enzymes are alsoresponsible for thebaine O6-demethylation in mammalianbrain tissue.
Experimental Procedures
Materials
Thebaine, hydrocodone, and hydromorphone were pur-chased from Sigma-Aldrich. Thebaine was recrystallized fromhot acetone. Oripavine and [N-C2H3]thebaine were providedby the late M. H. Zenk (Donald Danforth Plant Science Center,St. Louis, MO). CYP3cide was a gift from K. D. Hardy (Lip-scomb University, Nashville, TN). All other inhibitors werepurchased from Sigma-Aldrich. Rat brains were a gift fromC. K. Jones (Department of Pharmacology, Vanderbilt Univer-sity). Liver microsomes from human liver tissue (set of 10 indi-vidual donors) were prepared in this laboratory according topublished protocols (36).
2 The abbreviations used are: P450, cytochrome P450; ESI, electrosprayionization; MRM, multiple reaction monitoring; CYP3cide, 1-methyl-3-[1-methyl-5-(4-methylphenyl)-1H-pyrazol-4-yl]-4-[(3S)-3-piperidin-1-ylpyrrolidin-1-yl]-1H-pyrazolo[3,4-d]pyrimidine).
FIGURE 1. Latter steps of morphine biosynthesis (21).
P450 3A and Morphine Synthesis
AUGUST 14, 2015 • VOLUME 290 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 20201
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Enzymes
Human P450 3A4- and 3A5-containing Baculosomes�(microsomal preparation from baculovirus-infected insect cellsco-expressing human NADPH-P450 reductase) were pur-chased from Invitrogen/Life Technologies, and rat P450 3A2-containing Baculosomes� were purchased from BD Biosci-ences. A Pseudomonas putida morphinone reductase cDNAvector was provided by N. C. Bruce (University of York, York,UK), and the enzyme was heterologously expressed and puri-fied according to French and Bruce (37); when subjected toSDS-PAGE on a 10% (w/v) acrylamide gel, it showed a singleband at 41 kDa.
Preparation of Rat Brain Homogenate and Microsomes
Male Sprague-Dawley rats (350 – 450 g, Harlan Laboratories,Indianapolis, IN) used in the present studies were housed undera 12-h light/12-h dark cycle and given ad libitum access to foodand water. All animal experiments were approved by the Van-derbilt University Animal Care and Use Committee, and exper-imental procedures conformed to guidelines established by theNational Research Council Guide for the Care and Use of Lab-oratory Animals. Rats were briefly anesthetized with isofluraneand decapitated. Whole brains were removed and immediatelywashed in ice-cold 150 mM NaCl to remove blood clots. Thebrain tissue was homogenized in four volumes of ice-cold 0.10M potassium phosphate buffer (pH 7.6) containing 0.15 M KClusing 7–10 up-and-down strokes in a manually operated Teflonglass Potter-Elvehjem homogenizer. All steps used to obtainmicrosomal fractions were carried out at 4 °C. Homogenateswere centrifuged at 14,000 � g for 15 min at 4 °C (to removedebris, nuclei, and mitochondria), and the resulting superna-tant was centrifuged at 100,000 � g for 1 h at 4 °C. The micro-somal pellet was resuspended in 10 mM Tris acetate (pH 7.4)buffer containing 1.0 mM EDTA and 20% glycerol (v/v) (micro-somes from 1 g of brain in �0.2 ml of buffer).
Enzymatic Assays
In general, incubations were conducted in a total volume of300 �l in 50 mM potassium phosphate buffer (pH 7.4) contain-ing 10 pmol of P450 enzyme (Baculosomes�, Life Technologiesor BD Biosciences) or 0.5 mg of protein/ml of tissue homoge-nate or microsomal preparation. Unless otherwise noted, neg-ative control samples for each experiment contained the samecomponents but were heated in a boiling water bath (5 min)before initiating the reactions.
Additionally, to trap the primary reaction products, morphi-none reductase (0.5 �M) and NADH (1 mM) were included toconvert codeinone and morphinone to the stable productshydrocodone and hydromorphone, respectively (37) (see Fig.2A). Because the substrates codeinone and morphinone areunstable and we did not have a standard, the conditions werevalidated by two preliminary experiments. A reported substrateof morphinone reductase is 2-cyclohexen-1-one (38). There-fore, to validate the activity of morphinone reductase, werepeated the experiment described by Barna et al. (38) and mea-sured the rate of 2-cyclohexen-1-one reduction. Our resultsgive a rate of 1.3 s�1 under anaerobic conditions, which is sim-ilar to that reported (kcat 0.8 s�1). Anaerobic assays were done
using the basic apparatus design of Burleigh et al. (39) as sub-sequently modified in this laboratory (40, 41). Additionally,morphinone reductase was titrated in an incubation with P4503A5 (described below), and saturation was seen at 0.2 �M mor-phinone reductase (see Fig. 2B).
Following temperature equilibration to 37 °C for 5 min, thereactions were initiated by the addition of an NADPH-regener-ating system (0.5 mM NADP�, 10 mM glucose 6-phosphate, and1 IU ml�1 yeast glucose 6-phosphate dehydrogenase (36)) toanalyze P450-dependent activity (42). Reactions generally pro-ceeded at 37 °C for 20 min and were terminated by the additionof 300 �l of ice-cold CH3OH. Quinidine (as internal standard,0.5 �mol) and sodium borate buffer, pH 9.5 (300 �l of 0.5 M
solution) were added. The samples were mixed using a vortexdevice and extracted with 1.0 ml of CH2Cl2, and the layers wereseparated by centrifugation (103 � g for 10 min). A 0.8-ml ali-quot of the CH2Cl2 layer (lower phase) was transferred to aclean tube. An additional 1.0 ml of CH2Cl2 was added to theresidual layer to extract the products, followed by mixing andanother centrifugation step at 103 � g. The organic layers werecombined, and the solvent was removed under an N2 stream.All samples were analyzed by LC/MS/MS (see below).
Initial Activity Screens
Initial screening for O6-demethylation activity was per-formed as described above using human brain homogenate,human liver microsomes, or no enzyme source. In these exper-iments, P450-dependent activity was initiated by the addition ofthe NADPH-regenerating system, and dioxygenase-dependentactivity was initiated by the addition of �-ketoglutarate (1 mM),Fe(NH4)2(SO4)2 (100 or 20 �M), and L-ascorbate (1.8 mM). In allcases, morphinone reductase and NADH were also present. Con-trol samples contained the same components that were boiledbefore initiating the reactions. Control samples contained humanliver homogenate, but no NADPH-regenerating system or �-ke-toglutarate, Fe(NH4)2(SO4)2, or L-ascorbate.
Steady-state Kinetic Assays
Kinetic assays were performed as described above usingBaculosomes� containing P450 3A4 or 3A5. Additionally, the-baine concentrations of 1.6, 3.1, 6.3, 12.5, 25, 50, and 100 �M
were used. Results were fit to hyperbolic plots in GraphPadPrism (GraphPad Software, La Jolla, CA) to estimate kcat andKm values � S.E.
Morphinone Reductase Titration
To determine a saturating concentration of morphinonereductase, a titration was performed as described above usingBaculosomes� containing P450 3A5; however, the concentra-tion of morphinone reductase varied (0.006, 0.03, 0.06, 0.18, 0.3,0.6, 1.2 �M) (see Fig. 2B).
Oxidation of d3-Thebaine
Assays were performed as described above using pooledhuman liver microsomes or Baculosomes� containing P4503A4 or 3A5. In addition to using thebaine as a substrate, sepa-rate incubations were done with d3-thebaine ([N-2H3]thebaine)(100 �M) as substrate.
P450 3A and Morphine Synthesis
20202 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 33 • AUGUST 14, 2015
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Inhibition Assays
Human Liver Microsomes—Initial P450 inhibition screenswere performed as described above, with inhibitors added.Inhibitor concentrations were as follows: 1 �M �-naphthofla-vone (for P450 1A1 and 1A2), 2 �M quinidine (for P450 2D6),100 �M 4-methylpyrazole (for P450 2E1), 2 �M methoxsalen(for P450 2A6), 50 �M quercetin (for P450 2C8), 5 �M sulfa-phenazole (for P450 2C9), 2 �M ketoconazole (for P450 3A4 and3A5), 10 �M fluconazole (for P450 51A1), or 5 �M ticlopidine(for P450 2B6). The concentrations were chosen based uponvalues from literature that showed efficacy against and selectiv-ity for each P450 (43– 48). The half-maximal inhibitory concen-tration (IC50) values were determined using conditionsdescribed above; however, these incubations contained eitherketoconazole (0, 0.2, 0.4, 2.0, or 10 �M) or CYP3cide (0, 0.4, 2.0,10, or 20 �M). IC50 values were calculated by non-linear regres-sion using the GraphPad Prism equation for inhibitor concen-tration response (Eq. 1)
Y �100
�1 � 10�X � log�IC50���(Eq. 1)
where X is log [inhibitor] and Y is the percentage of the unin-hibited activity. All inhibitors were dissolved in C2H5OH, andthe volume added was 0.1% (v/v). Control incubations con-tained no inhibitor but did include 0.1% C2H5OH (v/v).
P450 3A2 and Rat Brain Microsomes—Ketoconazole or sul-faphenazole (0, 0.2, or 2 �M) was included in incubations con-taining P450 3A2 (4 pmol) or rat brain homogenate (800 �g ofprotein) as described above. Control samples were incubatedwith and without vehicle present (0.5% CH3OH, v/v). Concen-trations used were chosen based on published selectivity ofinhibitors for rat P450s (49).
Iron Titration Incubations
Thebaine (100 �M), potassium phosphate buffer (50 mM, pH7.4), morphinone reductase (0.5 �M), NADH (1 mM), andFe(NH4)2(SO4)2 (0, 20, 100, 225, or 500 �M), in a total volume of300 �l, were incubated at 37 °C for 20 min. Reactions wereterminated, extracted, and analyzed as described under “Enzy-matic Assays.”
LC-MS/MS Analysis
Components of the dried extracts, dissolved in 10 mM
NH4HCO3:CH3CN:HCO2H (94.9:5:0.1, v/v), were separatedon a Waters ACQUITY UPLC system. Separation of (10-�l)samples was achieved using an ACQUITY� UPLC BEH C18octadecylsilane (1.7 �m, 2.1 � 100 mm) column (Waters)equipped with a C18 guard column (Phenomenex, 2 � 4 mm).The mobile phase flow rate was 0.3 ml/min, with binary gradi-ent elution using solvents A (10 mM aqueous NH4HCO3 with0.1% (v/v) HCO2H) and B (CH3CN with 0.1% (v/v) HCO2H).The gradient started with 95% A (v/v) and was increased to 70%B (v/v) over 6 min, and then held at 70% B (v/v) for 2 min.
Compounds were analyzed with a Thermo LTQ mass spec-trometer using the positive-ion electrospray ionization (ESI)mode. ESI conditions were: N2 sheath gas, 40 psi; N2 auxiliarygas, 6 psi; spray voltage, 20 V; capillary temperature, 350 °C;
capillary offset, 35 V; tube lens offset, 80 V. MS-MS conditionswere: normalized collision energy, 35%; activation Q setting,0.250; activation time, 30 ms.
Identification of thebaine product analytes was based onretention times (tR), the ratios of two different MRM transi-tions, and comparison with the expected values for standards.MRM transitions used were thebaine (m/z 3123351, m/z3123381), oripavine (m/z 2983218, m/z 2983249), hydro-codone (m/z 3003199, m/z 3003215), and hydromorphone(m/z 2863201, m/z 2863242). Quantitation was done by con-structing standard curves for each analyte (using the internalstandard quinidine).and integrating the peak areas of one MRMtransition with Xcalibur software (Thermo Scientific). Thelimit of quantitation for hydrocodone was 5 pmol, and the limitof detection was 1 pmol.
Identification of products of d3-thebaine was based on the tRof the non-deuterated standard and the expected m/z. The rel-evant masses of the compounds of interest were: for d3-thebaine,m/z 315; for oripavine, m/z 301; for hydrocodone, m/z 303;and for northebaine, m/z 298. Northebaine was not availableas a standard, and therefore quantitation was based using astandard curve prepared using thebaine. The limit of detectionfor thebaine was 500 fmol, and the limit of quantitation was 2pmol.
Results
Assays for O6-Demethylation—The O6-demethylation prod-ucts morphinone (from oripavine) and neopinone/codeinone(from thebaine) are Michael acceptors and are inherentlyunstable, and preliminary attempts to demonstrate their for-mation in liver homogenates, microsomes, or purified P450susing LC-UV assays were unsuccessful. We also tried LC-MSassays, including ones in which we added NaBH4 or NaBD4 totrap the products, without success, probably because theapproach only stabilized the product in solution at the comple-tion of the incubation.
The use of coupled assays is well established in biochemistry.A bacterial morphinone reductase has been extensively studiedas a model flavoprotein (34, 37, 50) and is known to rapidlyreduce both morphinone and codeinone. The products, hydro-morphone and hydrocodone, are stable and can easily beassayed (e.g. positive-ion ESI LC-MS). In preliminary assays, theconcentration of morphinone reductase we established (0.5�M) was found to be saturating in detecting product formation(see below) (Fig. 2B).
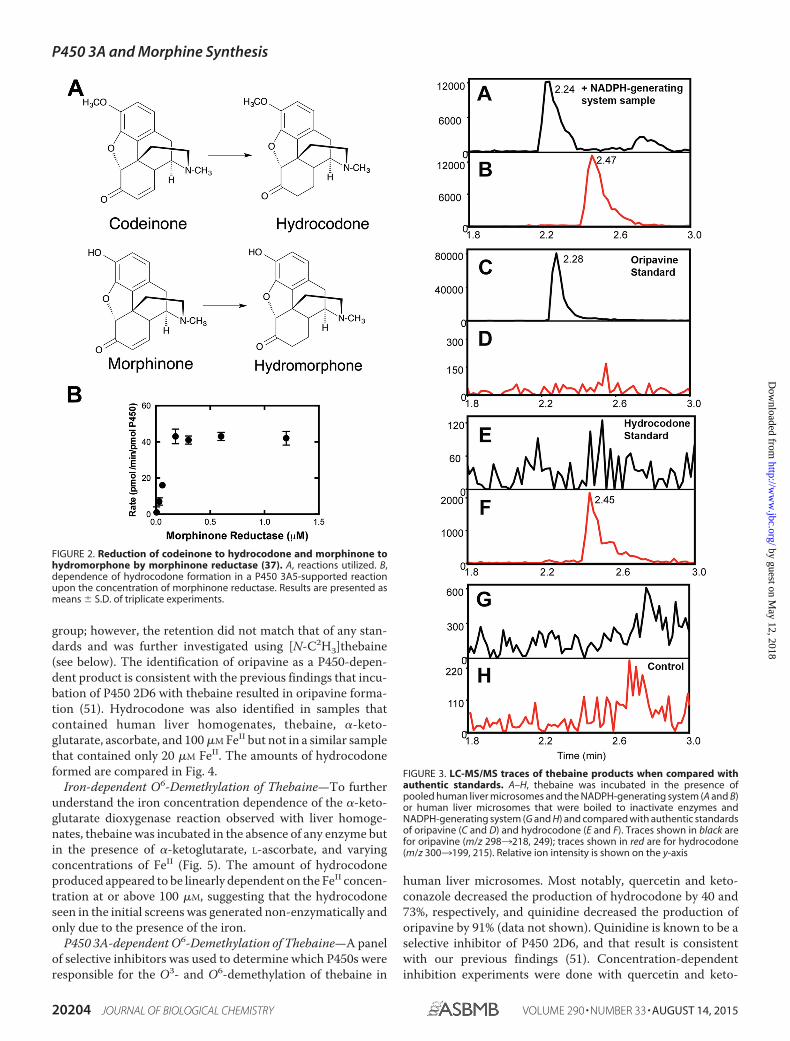
P450-dependent O6-Demethylation of Thebaine—Incuba-tions of thebaine and human liver microsomes in the presenceof an NADPH-regenerating system and the morphinone reduc-tase-NADH trapping system resulted in the formation of threeproducts that were not seen in control reactions with heat-inactivated enzymes. These metabolites were subjected to MSafter separation by UPLC. Comparison of two of the com-pounds with standards of alkaloids that were expected to beformed allowed the identification of the unknown compoundsas oripavine (via O3-demethylation, Fig. 1) (21) and hydro-codone (via O6-demethylation followed by morphinone reduc-tase-catalyzed reduction, Figs. 1 and 2) (37) (Fig. 3). The thirdcompound had an m/z of 298, suggesting the loss of a methyl
P450 3A and Morphine Synthesis
AUGUST 14, 2015 • VOLUME 290 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 20203
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
group; however, the retention did not match that of any stan-dards and was further investigated using [N-C2H3]thebaine(see below). The identification of oripavine as a P450-depen-dent product is consistent with the previous findings that incu-bation of P450 2D6 with thebaine resulted in oripavine forma-tion (51). Hydrocodone was also identified in samples thatcontained human liver homogenates, thebaine, �-keto-glutarate, ascorbate, and 100 �M FeII but not in a similar samplethat contained only 20 �M FeII. The amounts of hydrocodoneformed are compared in Fig. 4.
Iron-dependent O6-Demethylation of Thebaine—To furtherunderstand the iron concentration dependence of the �-keto-glutarate dioxygenase reaction observed with liver homoge-nates, thebaine was incubated in the absence of any enzyme butin the presence of �-ketoglutarate, L-ascorbate, and varyingconcentrations of FeII (Fig. 5). The amount of hydrocodoneproduced appeared to be linearly dependent on the FeII concen-tration at or above 100 �M, suggesting that the hydrocodoneseen in the initial screens was generated non-enzymatically andonly due to the presence of the iron.
P450 3A-dependent O6-Demethylation of Thebaine—A panelof selective inhibitors was used to determine which P450s wereresponsible for the O3- and O6-demethylation of thebaine in
human liver microsomes. Most notably, quercetin and keto-conazole decreased the production of hydrocodone by 40 and73%, respectively, and quinidine decreased the production oforipavine by 91% (data not shown). Quinidine is known to be aselective inhibitor of P450 2D6, and that result is consistentwith our previous findings (51). Concentration-dependentinhibition experiments were done with quercetin and keto-
FIGURE 2. Reduction of codeinone to hydrocodone and morphinone tohydromorphone by morphinone reductase (37). A, reactions utilized. B,dependence of hydrocodone formation in a P450 3A5-supported reactionupon the concentration of morphinone reductase. Results are presented asmeans � S.D. of triplicate experiments.
FIGURE 3. LC-MS/MS traces of thebaine products when compared withauthentic standards. A–H, thebaine was incubated in the presence ofpooled human liver microsomes and the NADPH-generating system (A and B)or human liver microsomes that were boiled to inactivate enzymes andNADPH-generating system (G and H) and compared with authentic standardsof oripavine (C and D) and hydrocodone (E and F). Traces shown in black arefor oripavine (m/z 2983218, 249); traces shown in red are for hydrocodone(m/z 3003199, 215). Relative ion intensity is shown on the y-axis
P450 3A and Morphine Synthesis
20204 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 33 • AUGUST 14, 2015
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

conazole. Quercetin (P450 2C8 inhibitor) did not show concen-tration dependence (results not shown) and therefore was notfurther investigated further.
Ketoconazole showed concentration-dependent inhibition,with an IC50 of 0.16 �M (Fig. 6), typical for P450 Subfamily 3A
reactions (52). Because ketoconazole appears to inhibit allhuman P450 3A enzymes (49, 53), CYP3cide (an inhibitor selec-tive for P450 3A4 (53,54)) was used to estimate the contributionof the human P450 3A enzymes involved in the O6-demethyla-tion of thebaine. CYP3cide also showed concentration depen-dence, with an IC50 of 2.1 �M (Fig. 6), typical for P450 3A4reactions (53, 54).
Thebaine O6-Demethylation Catalyzed by Recombinant P4503A4 and 3A5—Steady-state kinetic parameters for recombi-nant P450 3A4- and P450 3A5-mediated thebaine O6-demeth-ylation were determined based on the resulting hydrocodone.The results displayed Michaelis-Menten kinetics, and the cata-lytic parameters were estimated (Fig. 7). P450 3A4 and 3A5 hadsimilar apparent Km values (22 and 26 �M, respectively); how-ever, the maximum rate (kcat) for the conversion of thebaine tocodeinone by P450 3A5 (52 min�1) was 23-fold higher than thatof P450 3A4 (2.3 min�1) (Fig. 7). For reference, the previouslydetermined parameters for P450 2D6 conversion of thebaine tooripavine were kcat 4.6 min�1 and Km 48 �M (38) (kcat/Km 0.1�M�1 min�1).
N-Demethylation of d3-Thebaine by Human Liver Micro-somes, P450 3A4, and P450 3A5—To confirm the identity of theunidentified analyte with m/z 298 (see above), d3-thebaine wasused as substrate in incubations. An analyte loss of 14 atomicmass units indicates a loss of a methyl group. There are threemethyl groups on thebaine; an O-methyl at both the 3- andthe 6-positions and an N-methyl. The products of O3-demeth-ylation (oripavine) and the O6-demethylation (hydrocodoneafter reduction by coupled enzyme system) were already identi-fied, leaving the N-demethylation product, northebaine, as thelikely third product. By using thebaine and d3-thebaine as sub-strates in parallel experiments and comparing the products, wewere able to determine that northebaine was indeed the thirdanalyte.
When thebaine and d3-thebaine were included as substratesin parallel experiments, with pooled human liver microsomes,three products were observed. The first product (tR 1.9 min),which had an m/z of 298 when starting with thebaine and anm/z of 301 when starting with d3-thebaine, was oripavine. Thesecond (tR 2.05 min) had an m/z of 300 when starting with
FIGURE 4. O6-Demethylation of thebaine in the presence of or absence ofhuman liver microsomes with and without iron. A and B, thebaine wasincubated in the presence of pooled human liver homogenate or microsomes(A) or no enzyme (B) for 20 min. Control samples contained liver homogenatebut did not contain an NADPH-generating system. �NADPH samples con-tained an NADPH-generating system and liver microsomes, and �Fe samplescontained 20 or 100 �M iron, 1 mM �-ketoglutarate, 1.8 mM L-ascorbate, andliver homogenate. Results are presented as means � S.D. of duplicate (n 2)experiments.
FIGURE 5. FeII-dependent O6-demethylation of thebaine. Fe(NH4)2(SO4)2(0, 20, 100, 250, and 500 �M) was incubated with 50 �M thebaine for 20 minunder typical assay conditions (absence of enzymes). Results are presented asmeans � S.D. of triplicate experiments.
FIGURE 6. Inhibition of hydrocodone formation in human liver micro-somes. Thebaine was incubated in the presence of pooled human livermicrosomes and the NADPH-generating system with increasing amountsof ketoconazole (F) or CYP3cide (f). Inhibition constants were derivedfrom non-linear fitting to a single-site inhibition model in Prism software(GraphPad). Results are presented as means � S.D. of triplicateexperiments.
P450 3A and Morphine Synthesis
AUGUST 14, 2015 • VOLUME 290 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 20205
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

thebaine and an m/z of 303 when starting with d3-thebaine,consistent with hydrocodone. The third, tR 2.4 min, had anm/z of 298 when starting with thebaine or with d3-thebaine,consistent with loss of the N-methyl group, giving north-ebaine. The northebaine gave fragment ions distinct fromhydrocodone (m/z of 2983281, 266). Northebaine was alsoidentified as a product when thebaine or d3-thebaine wasincluded as substrates with P450s 3A4 or 3A5. Based on onetime point and a standard curve of thebaine, rate estimateswere calculated for the conversion of thebaine to north-ebaine by P450s 3A4 and 3A5 (88 and 27 pmol/pmol ofP450/min, respectively.)
Thebaine O6-Demethylation by P450 3A2 and Rat BrainMicrosomes—We have been unsuccessful in demonstrating anyP450-dependent activity in frozen brain samples obtained fromcattle or rats. Stored human brain samples (Vanderbilt Univer-sity tissue facility) or those provided by the Cooperative HumanTissue Network (Birmingham, AL; Nashville, TN) were alsonegative for known P450 reactions, e.g. 7-ethoxycoumarinO-deethylation. We were able to detect activities in micro-somes prepared from freshly obtained rat brains, as reportedpreviously (55). Thebaine O6-demethylation was detected withrecombinant rat P450 3A2, a prominent rat Subfamily 3A P450(Fig. 8A), or rat brain microsomes (Fig. 8B). Rates were esti-mated to be 0.75 fmol/min/pmol of P450 and 1.3 fmol/min/mgof total protein for P450 3A2 and rat brain microsomes, respec-tively (based on a single time point, 15 min). Additionally, O6-demethylation of thebaine was inhibited in both P450 3A2 andrat brain microsomes in the presence of either ketoconazole(Subfamily 3A-selective). Sulfaphenazole (P450 Subfamily2C-selective (48)) did not inhibit O6-demethylation by eitherthe P450 3A2 or rat brain microsomes.
Discussion
A wealth of complementary biochemical, molecular, andphysiological studies from independent laboratories have pro-vided evidence supporting the biosynthesis of morphine in avariety of animal cells and organs (see Ref. 21 for comprehen-sive review). These studies have characterized multiple en-zyme-catalyzed reactions and chemically defined intermediateprecursor molecules that share marked similarities to the plantbiosynthetic scheme previously established in the poppyP. somniferum (3–5, 9, 10, 22, 24 –26, 51, 56 –58). The finalsteps of the pathway convert thebaine to morphine, and the-baine must undergo demethylation at both the O3- and O6-po-sitions (Fig. 1). Thus, two parallel pathways are possible for theconversion of thebaine to morphine: one in which O3-demeth-ylation precedes O6-demethylation and the other in which theorder is reversed. In plants, both pathways occur and the O3-andO6-demethylationstepsarecatalyzedbycodeineO-demeth-ylase and thebaine O6-demethylase, respectively, with the bal-ance differing among species and strains (27). Because both ofthese enzymes are in the FeII/�-ketoglutarate-dependent
FIGURE 7. Oxidation of thebaine to codeinone by recombinant P450 3A4and 3A5. A, P450 3A4. kcat 2.3 � 0.3 pmol/min/pmol P450; Km 22 � 7 �M;kcat/Km 0.10 �M
�1 min�1. B, P450 3A5. kcat 52 � 4 pmol/min/pmol P450; Km26 � 6 �M; kcat/Km 2.0 �M
�1 min�1. The results (� S.E.) were calculated usingGraphPad Prism.
FIGURE 8. Inhibition of P450 3A2- or rat brain homogenate-mediatedhydrocodone formation. A and B, thebaine was incubated in the pres-ence of P450 3A2 (A) or rat brain homogenate (B). Control samples con-tained enzyme and the NADPH-generating system. Vehicle Control sam-ples contained 0.05% (v/v) CH3OH. The inhibitor-containing samplescontained 0.2 or 2 �M ketoconazole or sulfaphenazole in the same volumeof CH3OH as the vehicle control. Results are presented as means � S.D. ofduplicate experiments.
P450 3A and Morphine Synthesis
20206 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 33 • AUGUST 14, 2015
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

dioxygenase family (33), our initial hypothesis was that anenzyme from this family would also be responsible for thesedemethylation steps in mammalian systems. None of ourresults support the initial hypothesis; instead, it was shownthat both human and rat P450 3A enzymes catalyze theO6-demethylation of thebaine. Additionally, the same reac-tion is catalyzed by a P450 in rat brain microsomes, appar-ently a member of the P450 3A Subfamily. These findingshighlight a key difference in the biosynthesis of morphine inplants versus mammals. Oripavine and northebaine appearto be final products.
The thebaine and oripavine O6-demethylation reactionsare somewhat unusual in that they both involve cleavage ofan enol ether, as opposed to an aliphatic or a phenolic ether.This cleavage is apparently unprecedented in P450 chemis-try (30) aside from aryl ethers. As mentioned earlier, initialefforts to directly demonstrate this reaction were unsuccess-ful, before implementation of the morphinone reductase-coupled assay.
Efforts were made to determine whether mammalian FeII/�-ketoglutarate-dependent dioxygenases catalyze O3- and O6-demethylation of thebaine, based on the precedence in plants.Our preliminary experiments were designed using typical con-ditions from the literature, which included exogenous iron con-centrations ranging from 50 to 500 �M (33, 59, 60). Althoughthebaine O6-demethylation was observed, we noted that thegeneration of the product was neither time-dependent norenzyme concentration-dependent. Experiments done in theabsence of any enzyme showed that O6-demethylation ofthebaine was dependent on the iron concentration (Fig. 5).At concentrations �20 �M iron, no (non-enzymatic) O6-the-baine demethylation was detected. Although total mamma-lian cellular iron concentrations are estimated between 50and 100 �M, iron homeostasis is highly regulated and freeiron represents only a minor fraction (5%); even 20 �M ironis much higher than the biologically relevant concentrationand would be highly deleterious (61). When experimentswith brain homogenates were repeated with 20 �M iron, nohydrocodone formation was detected. Although no furtherstudies on the mechanism of the iron-catalyzed demethyla-tion were done, it is well known that FeII catalyzes non-enzymatic hydroxyl radical formation from H2O2 via theFenton reaction (62– 64), and thus it is probably not surpris-ing that iron-catalyzed (non-enzymatic) demethylation wasobserved (see also Ref. 65 for iron-catalyzed N-demethyla-tion of oripavine). Because the O-demethylation that wasinitially observed was an artifact resulting from the high ironconcentration, it calls into question the use of high iron con-centrations that are not biologically relevant. We cannotspeculate on the relevance of our results in the general fieldof FeII/�-ketoglutarate-dependent dioxygenases. Manydioxygenases in this field have been extensively character-ized (32, 59, 60, 66 – 68). However, our results do suggestcaution in the assignment of activities to enzymes in thisfamily in the absence of more extensive characterization.
We report here that thebaine O6-demethylation is catalyzedby P450 Subfamily 3A enzymes. The use of P450-selectiveinhibitors, in combination with human liver microsomes,
implicatedhumanP450s3A4and3A5incatalysisofO6-demeth-ylation, confirmed by in vitro experiments with recombinantenzymes. Although both of these enzymes generate the sameproduct, the catalytic efficiency of P450 3A5 was 20-fold higherthan that of P450 3A4. When considering the relevance ofP450s 3A4 and 3A5 in common substrate metabolism, themuch higher liver concentrations and typical superior catalyticefficiency usually focus responsibility toward P450 3A4 (52,69 –71). Here, however, we are considering P450 activity in thebrain, and P450 3A5 is expressed in the brain with mRNA levelscomparable with that of P450 3A4 (72). The higher catalyticefficiency and its measured expression in the brain lead to theproposal that P450 3A5 plays an important role in human mor-phine biosynthesis.
Numerous searches for thebaine O6-demethylation usingmicrosomes derived from human brain tissue wereattempted but were not presented here because no enzy-matic activity was observed for any P450-dependent reac-tion. This was likely due to the difficulty in obtaining humanbrain samples that undergo careful treatment necessary toretain enzymatic activity. Thus, microsomes were preparedfrom freshly acquired rat brains. In the presence of anNADPH-generating system, O6-demethylation was ob-served when thebaine was incubated with these microsomes(Fig. 8B). Additionally, the inhibition of this activity in thepresence of ketoconazole (a P450 3A inhibitor that alsoinhibits rat P450 2C9) and the retention of the activity in thepresence sulfaphenazole (a selective P450 2C inhibitor) (49)indicate that thebaine O6-demethylation in the brain micro-somes is due to one or more of the rat P450 3A enzymes.(P450 activity was also lost in our rat brain tissues uponfrozen storage (see also Ref. 73) consistent with the lack ofactivity in all frozen human brain samples we obtained.)Thus, in addition to showing that the human P450 3Aenzymes can catalyze thebaine O6-demethylation, we reportthat this demethylation reaction occurs in rat brain, cata-lyzed by one or more Subfamily 3A P450 enzymes. Theresults presented here are consistent with a study that mea-sured NADPH-dependent thebaine metabolism in rat brainmicrosomes (26). In that study, codeine was generated at arate of 0.20 � 0.04 pmol/h/mg of protein (as estimated by aradioimmune assay instead of LC-MS), as compared with thepresent study in which hydrocodone was generated at a rateof 0.075 pmol/h/mg of protein in the coupled LC-MS assay.
As described, two parallel pathways lead from thebaine tomorphine (Fig. 1). The first, in which thebaine is demethylatedat the 3-position, yields oripavine, not thebaine, as the substratefor O6-demethylation. When oripavine was incubated witheither human liver microsomes or recombinant P450 3A4 orP450 3A5, no O6-demethylation was observed (data notshown). Thus, the results presented in this study support thesecond pathway, in which thebaine is first demethylated at theO6-position, as the only pathway that contributes to morphinesynthesis, at least in human liver.
No clear function has been attributed to endogenous mor-phine, but suggestions include infection, sepsis, and inflamma-tion, as well as major neurological pathologies (Parkinson dis-ease, schizophrenia) (reviewed in Ref. 21). Our findings indicate
P450 3A and Morphine Synthesis
AUGUST 14, 2015 • VOLUME 290 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 20207
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

that P450 3A5 is most active in the O6-demethylation of the-baine in human biosynthesis of morphine (Fig. 7). P450 3A5 ishighly polymorphic, and the wild-type allele, CYP3A5*1, isexpressed with a frequency of only 5–15% in Caucasian popu-lations, 25– 40% in various Asian ethnic groups, and �40 – 60%in African and African-American populations (69, 74, 75).Thus, individuals with P450 3A5 polymorphisms that affectmorphine biosynthesis might be expected to have differentphysiological outcomes related to this. Mikus et al. (12) mea-sured urinary excretion of codeine and morphine in 40 peoplewho had been phenotyped for P450 2D6 status, utilizing anHPLC separation/radioimmune assay for detection. Theyreported no difference in morphine levels due to P450 2D6status and concluded that other P450s might be involved in thesynthesis of endogenous morphine. The relevance of geneticvariation of P450s 3A4 and 3A5 to the P450 2D6 work isunknown. In the work of Mikus et al. (12), considerable inter-individual variability was reported (103-fold), but 10-fold day-to-day intra-individual variabilities where also seen. Whetherthese variations are real or the result of analytical deficiencies isnot clear.
Our results can be placed in context of the overall schemeof mammalian synthesis of morphine (Fig. 9). Administra-tion of (R,S)-tetrahydropapaveroline to mice yielded reticu-line, salutaridinol, thebaine, and oripavine in urine (24). Allof the oxidation steps following the formation of tetrahydro-papaveroline have now been characterized in humans invitro. P450 2D6 or the two Subfamily 3A P450s (3A4 and3A5) have roles in all oxidation steps examined, includingthe early step of oxidation of tyramine to dopamine (7). Acaveat is that many of the demonstrations of catalytic activ-ity have been done with individual enzymes and the possibil-ity exists that some other P450s may also be found to con-tribute. Although many of the studies involved in elucidatingthe steps in Fig. 9 have been done in liver systems (26, 51),they do not speak to the localization near the neural targets,which may play an important role in endogenous morphinebiosynthesis in the brain. It is possible that an endocrinemechanism is more relevant to the function of endogenousmorphine. In conclusion, our results now implicate severalmajor human P450s in the scheme of Fig. 9, and these arefound in a number of sites in the body (36).
FIGURE 9. Current scheme of enzymatic mammalian morphine biosynthesis. PNMT, phenylethanolamine N-methyltransferase; NMT, N-methyltrans-ferase (7, 58).
P450 3A and Morphine Synthesis
20208 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 33 • AUGUST 14, 2015
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Author Contributions—V. M. K. conducted most of the experi-ments, analyzed the results, and wrote most of the paper. M. A. R.conducted experiments on the roles of P450s 3A4 and 3A5. T. K.conducted experiments searching for dioxygenase function.F. P. G. conceived the idea for the project, conducted the anaero-bic assay of morphinone reductase activity, and wrote the paperwith V. M. K.
Acknowledgments—We thank the late Prof. M. H. Zenk for the gifts ofthe oripavine and d3-thebaine and for encouragement in the earlyphases of this work. We also acknowledge Drs. M. Dostalek and G.Chowdhury for exploratory work on this project and K. Trisler forassistance in the preparation of the manuscript.
References1. Fowler, C. J., and Fraser, G. L. (1994) �-, �-, -Opioid receptors and their
subtypes: a critical review with emphasis on radioligand binding experi-ments. Neurochem. Int. 24, 401– 426
2. Perea-Sasiain, J. (2008) Biosynthesis of morphine: its importance in Par-kinson’s disease. Rev. Fac. Med. Unal. 56, 161–189
3. Lee, C. S., and Spector, S. (1991) Changes of endogenous morphine andcodeine contents in the fasting rat. J. Pharmacol. Exp. Ther. 257, 647– 650
4. Goldstein, A., Barrett, R. W., James, I. F., Lowney, L. I., Weitz, C. J., Knip-meyer, L. L., and Rapoport, H. (1985) Morphine and other opiates frombeef brain and adrenal. Proc. Natl. Acad. Sci. U.S.A. 82, 5203–5207
5. Donnerer, J., Cardinale, G., Coffey, J., Lisek, C. A., Jardine, I., and Spector,S. (1987) Chemical characterization and regulation of endogenous mor-phine and codeine in the rat. J. Pharmacol. Exp. Ther. 242, 583–587
6. Meijerink, W. J. H. J., Molina, P. E., and Abumrad, N. N. (1999) Mamma-lian opiate alkaloid synthesis: lessons derived from plant biochemistry.Shock 12, 165–173
7. Hiroi, T., Imaoka, S., and Funae, Y. (1998) Dopamine formation fromtyramine by CYP2D6. Biochem. Biophys. Res. Commun. 249, 838 – 843
8. Fricchione, G., Zhu, W., Cadet, P., Mantione, K. J., Bromfield, E., Madsen,J., DeGirolami, U., Dworetzky, B., Vaccaro, B., Black, P., and Stefano, G. B.(2008) Identification of endogenous morphine and a �3-like opiate alka-loid receptor in human brain tissue taken from a patient with intractablecomplex partial epilepsy. Med. Sci. Monit. 14, CS45–CS49
9. Poeaknapo, C., Schmidt, J., Brandsch, M., Drager, B., and Zenk, M. H.(2004) Endogenous formation of morphine in human cells. Proc. Natl.Acad. Sci. U.S.A. 101, 14091–14096
10. Boettcher, C., Fellermeier, M., Boettcher, C., Drager, B., and Zenk, M. H.(2005) How human neuroblastoma cells make morphine. Proc. Natl.Acad. Sci. U.S.A. 102, 8495– 8500
11. Zhu, W., Cadet, P., Baggerman, G., Mantione, K. J., and Stefano, G. B.(2005) Human white blood cells synthesize morphine: CYP2D6 modula-tion. J. Immunol. 175, 7357–7362
12. Mikus, G., Bochner, F., Eichelbaum, M., Horak, P., Somogyi, A. A., andSpector, S. (1994) Endogenous codeine and morphine in poor and exten-sive metabolisers of the CYP2D6 (debrisoquine/sparteine) polymor-phism. J. Pharmacol. Exp. Ther. 268, 546 –551
13. Findlay, J. W., Jones, E. C., Butz, R. F., and Welch, R. M. (1978) Plasmacodeine and morphine concentrations after therapeutic oral doses of co-deine-containing analgesics. Clin. Pharmacol. Ther. 24, 60 – 68
14. Laux-Biehlmann, A., Grafe, N., Mouheiche, J., Stuber, D., Welters, I. D.,Delalande, F., Poisbeau, P., Garnero, P., Metz-Boutigue, M. H., Schneider,F., and Goumon, Y. (2012) Comparison of serum and lithium-heparinateplasma for the accurate measurements of endogenous and exogenousmorphine concentrations. Br. J. Clin. Pharmacol. 74, 381–383
15. Charron, G., Doudnikoff, E., Laux, A., Berthet, A., Porras, G., Canron,M. H., Barroso-Chinea, P., Li, Q., Qin, C., Nosten-Bertrand, M., Giros, B.,Delalande, F., Van Dorsselaer, A., Vital, A., Goumon, Y., and Bezard, E.(2011) Endogenous morphine-like compound immunoreactivity in-creases in Parkinsonism. Brain 134, 2321–2338
16. Glattard, E., Welters, I. D., Lavaux, T., Muller, A. H., Laux, A., Zhang, D.,
Schmidt, A. R., Delalande, F., Laventie, B. J., Dirrig-Grosch, S., Colin, D. A.,Van Dorsselaer, A., Aunis, D., Metz-Boutigue, M. H., Schneider, F., andGoumon, Y. (2010) Endogenous morphine levels are increased in sepsis: apartial implication of neutrophils. Plos. One 5, e8791
17. Cousins, D. A., Butts, K., and Young, A. H. (2009) The role of dopamine inbipolar disorder. Bipolar Disord. 11, 787– 806
18. Kirby, G. W. (1967) Biosynthesis of the morphine alkaloids. Science 155,170 –173
19. Kutchan, T. M. (1998) Molecular genetics of plant alkaloid biosynthesis, inThe Alkaloids: Chemistry and Biology (Cordell, G. A., ed), Vol. 50, pp.257–316, Academic Press, San Diego
20. Onoyovwe, A., Hagel, J. M., Chen, X., Khan, M. F., Schriemer, D. C., andFacchini, P. J. (2013) Morphine biosynthesis in opium poppy involves twocell types: sieve elements and laticifers. Plant Cell 25, 4110 – 4122
21. Laux-Biehlmann, A., Mouheiche, J., Veriepe, J., and Goumon, Y. (2013)Endogenous morphine and its metabolites in mammals: history, synthesis,localization and perspectives. Neuroscience 233, 95–117
22. Kodaira, H., Lisek, C. A., Jardine, I., Arimura, A., and Spector, S. (1989)Identification of the convulsant opiate thebaine in mammalian brain. Proc.Natl. Acad. Sci. U.S.A. 86, 716 –719
23. Weitz, C. J., Lowney, L. I., Faull, K. F., Feistner, G., and Goldstein, A. (1986)Morphine and codeine from mammalian brain. Proc. Natl. Acad. Sci.U.S.A. 83, 9784 –9788
24. Grobe, N., Lamshoft, M., Orth, R. G., Drager, B., Kutchan, T. M., Zenk,M. H., and Spiteller, M. (2010) Urinary excretion of morphine and biosyn-thetic precursors in mice. Proc. Natl. Acad. Sci. U.S.A. 107, 8147– 8152
25. Weitz, C. J., Faull, K. F., and Goldstein, A. (1987) Synthesis of the skeletonof the morphine molecule by mammalian liver. Nature 330, 674 – 677
26. Kodaira, H., and Spector, S. (1988) Transformation of thebaine tooripavine, codeine, and morphine by rat liver, kidney, and brain micro-somes. Proc. Natl. Acad. Sci. U.S.A. 85, 1267–1271
27. Hagel, J. M., and Facchini, P. J. (2010) Dioxygenases catalyze the O-demeth-ylation steps of morphine biosynthesis in opium poppy. Nat. Chem. Biol.6, 273–275
28. Dayer, P., Desmeules, J., Leemann, T., and Striberni, R. (1988) Bioactiva-tion of the narcotic drug codeine in human liver is mediated by the poly-morphic monooxygenase catalyzing debrisoquine 4-hydroxylation (cyto-chrome P-450 dbl/bufI). Biochem. Biophys. Res. Commun. 152, 411– 416
29. Ortiz de Montellano, P. R. (2015) Substrate oxidation by cytochrome P450enzymes, in Cytochrome P450: Structure, Mechanism, and Biochemistry(Ortiz de Montellano, P. R., ed), 4th Ed., pp. 111–176, Springer, New York
30. Guengerich, F. P. (2001) Common and uncommon cytochrome P450 re-actions related to metabolism and chemical toxicity. Chem. Res. Toxicol.14, 611– 650
31. Knauer, S. H., Hartl-Spiegelhauer, O., Schwarzinger, S., Hanzelmann, P.,and Dobbek, H. (2012) The Fe(II)/�-ketoglutarate-dependent taurine di-oxygenases from Pseudomonas putida and Escherichia coli are tetramers.FEBS J. 279, 816 – 831
32. Proshlyakov, D. A., Henshaw, T. F., Monterosso, G. R., Ryle, M. J., andHausinger, R. P. (2004) Direct detection of oxygen intermediates in thenon-heme Fe enzyme taurine/�-ketoglutarate dioxygenase. J. Am. Chem.Soc. 126, 1022–1023
33. Farrow, S. C., and Facchini, P. J. (2013) Dioxygenases catalyze O-demeth-ylation and O,O-demethylenation with widespread roles in benzyliso-quinoline alkaloid metabolism in opium poppy. J. Biol. Chem. 288,28997–29012
34. Craig, D. H., Moody, P. C., Bruce, N. C., and Scrutton, N. S. (1998) Reduc-tive and oxidative half-reactions of morphinone reductase from Pseu-domonas putida M10: a kinetic and thermodynamic analysis. Biochemis-try 37, 7598 –7607
35. Lister, D. L., Kanungo, G., Rathbone, D. A., and Bruce, N. C. (1999) Trans-formations of codeine to important semisynthetic opiate derivatives byPseudomonas putida m10. FEMS Microbiol. Lett. 181, 137–144
36. Guengerich, F. P. (2014) Analysis and characterization of enzymes andnucleic acids relevant to toxicology, in Hayes’ Principles and Methods ofToxicology (Hayes, A. W., and Kruger, C. L., eds), 6th Ed., pp. 1905–1964,CRC Press-Taylor & Francis, Boca Raton, FL
37. French, C. E., and Bruce, N. C. (1994) Purification and characterization of
P450 3A and Morphine Synthesis
AUGUST 14, 2015 • VOLUME 290 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 20209
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

morphinone reductase from Pseudomonas putida M10. Biochem. J. 301,97–103
38. Barna, T., Messiha, H. L., Petosa, C., Bruce, N. C., Scrutton, N. S., andMoody, P. C. (2002) Crystal structure of bacterial morphinone reduc-tase and properties of the C191A mutant enzyme. J. Biol. Chem. 277,30976 –30983
39. Burleigh, B. D., Jr., Foust, G. P., and Williams, C. H., Jr. (1969) A methodfor titrating oxygen-sensitive organic redox systems with reducing agentsin solution. Anal. Biochem. 27, 536 –544
40. Guengerich, F. P. (1983) Oxidation-reduction properties of rat liver cyto-chromes P-450 and NADPH-cytochrome P-450 reductase related to ca-talysis in reconstituted systems. Biochemistry 22, 2811–2820
41. Guengerich, F. P., Krauser, J. A., and Johnson, W. W. (2004) Rate-limitingsteps in oxidations catalyzed by rabbit cytochrome P450 1A2. Biochemis-try 43, 10775–10788
42. Sohl, C. D., Cheng, Q., and Guengerich, F. P. (2009) Chromatographicassays of drug oxidation by human cytochrome P450 3A4. Nat. Protoc. 4,1252–1257
43. Turpeinen, M., Uusitalo, J., Jouko, U., Jalonen, J., Jorma, J., Pelkonen, O.,and Olavi, P. (2005) Multiple P450 substrates in a single run: rapid andcomprehensive in vitro interaction assay. Eur. J. Pharm. Sci. 24, 123–132
44. Bu, H. Z., Knuth, K., Magis, L., and Teitelbaum, P. (2001) High-through-put cytochrome P450 (CYP) inhibition screening via a cassette probe-dosing strategy. V. Validation of a direct injection/on-line guard cartridgeextraction: tandem mass spectrometry method for CYP1A2 inhibitionassessment. Eur. J. Pharm. Sci. 12, 447– 452
45. Yin, H., Racha, J., Li, S. Y., Olejnik, N., Satoh, H., and Moore, D. (2000)Automated high throughput human CYP isoform activity assay usingSPE-LC/MS method: application in CYP inhibition evaluation. Xenobi-otica 30, 141–154
46. Zhang, W., Kilicarslan, T., Tyndale, R. F., and Sellers, E. M. (2001) Evalu-ation of methoxsalen, tranylcypromine, and tryptamine as specific andselective CYP2A6 inhibitors in vitro. Drug. Metab. Dispos. 29, 897–902
47. Richter, T., Murdter, T. E., Heinkele, G., Pleiss, J., Tatzel, S., Schwab, M.,Eichelbaum, M., and Zanger, U. M. (2004) Potent mechanism-based inhi-bition of human CYP2B6 by clopidogrel and ticlopidine. J. Pharmacol.Exp. Ther. 308, 189 –197
48. Correia, M. A. (2005) Inhibition of cytochrome P450 enzymes, in Cyto-chrome P450: Structure, Mechanism, and Biochemistry (Ortiz de Mon-tellano, P. R., ed), 3rd Ed., pp. 247–322, Kluwer Academic/Plenum Press,New York
49. Kobayashi, K., Urashima, K., Shimada, N., and Chiba, K. (2003) Selectivi-ties of human cytochrome P450 inhibitors toward rat P450 isoforms: studywith cDNA-expressed systems of the rat. Drug. Metab. Dispos. 31,833– 836
50. Messiha, H. L., Munro, A. W., Bruce, N. C., Barsukov, I., and Scrutton,N. S. (2005) Reaction of morphinone reductase with 2-cyclohexen-1-one and 1-nitrocyclohexene: proton donation, ligand binding, and therole of residues histidine 186 and asparagine 189. J. Biol. Chem. 280,10695–10709
51. Grobe, N., Zhang, B., Fisinger, U., Kutchan, T. M., Zenk, M. H., andGuengerich, F. P. (2009) Mammalian cytochrome P450 enzymes catalyzethe phenol-coupling step in endogenous morphine biosynthesis. J. Biol.Chem. 284, 24425–24431
52. Niwa, T., Murayama, N., Emoto, C., and Yamazaki, H. (2008) Comparisonof kinetic parameters for drug oxidation rates and substrate inhibitionpotential mediated by cytochrome P450 3A4 and 3A5. Curr. Drug. Metab.9, 20 –33
53. Tseng, E., Walsky, R. L., Luzietti, R. A., Jr., Harris, J. J., Kosa, R. E., Goosen,T. C., Zientek, M. A., and Obach, R. S. (2014) Relative contributions ofcytochrome CYP3A4 versus CYP3A5 for CYP3A-cleared drugs assessedin vitro using a CYP3A4-selective inactivator (CYP3cide). Drug. Metab.Dispos. 42, 1163–1173
54. Walsky, R. L., Obach, R. S., Hyland, R., Kang, P., Zhou, S., West, M.,Geoghegan, K. F., Helal, C. J., Walker, G. S., Goosen, T. C., and Zientek,M. A. (2012) Selective mechanism-based inactivation of CYP3A4 by
CYP3cide (PF-04981517) and its utility as an in vitro tool for delineatingthe relative roles of CYP3A4 versus CYP3A5 in the metabolism of drugs.Drug. Metab. Dispos. 40, 1686 –1697
55. Guengerich, F. P., and Mason, P. S. (1979) Immunological comparison ofhepatic and extrahepatic cytochromes P-450. Mol. Pharmacol. 15,154 –164
56. Cardinale, G. J., Donnerer, J., Finck, A. D., Kantrowitz, J. D., Oka, K., andSpector, S. (1987) Morphine and codeine are endogenous components ofhuman cerebrospinal fluid. Life Sci. 40, 301–306
57. Donnerer, J., Oka, K., Brossi, A., Rice, K. C., and Spector, S. (1986) Pres-ence and formation of codeine and morphine in the rat. Proc. Natl. Acad.Sci. U.S.A. 83, 4566 – 4567
58. Grobe, N., Ren, X., Kutchan, T. M., and Zenk, M. H. (2011) An (R)-specificN-methyltransferase involved in human morphine biosynthesis. Arch.Biochem. Biophys. 506, 42– 47
59. Li, D., Fedeles, B. I., Shrivastav, N., Delaney, J. C., Yang, X., Wong, C.,Drennan, C. L., and Essigmann, J. M. (2013) Removal of N-alkyl modifica-tions from N2-alkylguanine and N4-alkylcytosine in DNA by the adaptiveresponse protein AlkB. Chem. Res. Toxicol. 26, 1182–1187
60. Fukumori, F., and Hausinger, R. P. (1993) Alcaligenes eutrophus JMP134“2,4-dichlorophenoxyacetate monooxygenase” is an �-ketoglutarate-de-pendent dioxygenase. J. Bacteriol. 175, 2083–2086
61. Kakhlon, O., and Cabantchik, Z. I. (2002) The labile iron pool: character-ization, measurement, and participation in cellular processes. Free Radic.Biol. Med. 33, 1037–1046
62. Winterbourn, C. C. (1995) Toxicity of iron and hydrogen peroxide:the Fenton reaction. Toxicol. Lett. 82–83, 969–974, 10.1016/0378-4274(95)03532-X
63. Goldstein, S., and Meyerstein, D. (1999) Comments on the mechanism of“Fenton-Like” reaction. Acc. Chem. Res. 32, 547–550
64. MacFaul, P. A., Wayner, D. D. M., and Ingold, K. U. (1998) A radicalaccount of “Oxygenated Fenton Chemistry.” Acc. Chem. Res. 31, 159 –162
65. Kok, G. B., and Scammells, P. J. (2011) Further investigations into theN-demethylation of oripavine using iron and stainless steel. Org. Biomol.Chem. 9, 1008 –1011
66. Fukumori, F., and Hausinger, R. P. (1993) Purification and characteriza-tion of 2,4-dichlorophenoxyacetate/�-ketoglutarate dioxygenase. J. Biol.Chem. 268, 24311–24317
67. Hausinger, R. P. (2004) FeII/�-ketoglutarate-dependent hydroxylases andrelated enzymes. Crit. Rev. Biochem. Mol. Biol. 39, 21– 68
68. Hausinger, R. S., C. (2015) 2-Oxoglutarate-Dependent Oxygenases, 1st Ed.,Royal Society of Chemistry, Abingdon, Oxfordshire, UK
69. Daly, A. K. (2006) Significance of the minor cytochrome P450 3A iso-forms. Clin. Pharmacokinet. 45, 13–31
70. Stevens, J. C. (2006) New perspectives on the impact of cytochrome P4503A expression for pediatric pharmacology. Drug Discov. Today 11,440 – 445
71. Rendic, S. P., and Guengerich, F. P. (2015) Survey of human oxidoreduc-tases and cytochrome P450 enzymes involved in the metabolism of chem-icals. Chem. Res. Toxicol. 28, 39 – 42
72. Agrawal, V., Huang, N., and Miller, W. L. (2008) Pharmacogenetics ofP450 oxidoreductase: effect of sequence variants on activities of CYP1A2and CYP2C19. Pharmacogenet. Genomics 18, 569 –576
73. Tyndale, R. F., Li, Y., Li, N. Y., Messina, E., Miksys, S., and Sellers, E. M.(1999) Characterization of cytochrome P-450 2D1 activity in rat brain:high-affinity kinetics for dextromethorphan. Drug Metab. Dispos. 27,924 –930
74. Kuehl, P., Zhang, J., Lin, Y., Lamba, J., Assem, M., Schuetz, J., Watkins,P. B., Daly, A., Wrighton, S. A., Hall, S. D., Maurel, P., Relling, M., Brimer,C., Yasuda, K., Venkataramanan, R., Strom, S., Thummel, K., Boguski,M. S., and Schuetz, E. (2001) Sequence diversity in CYP3A promoters andcharacterization of the genetic basis of polymorphic CYP3A5 expression.Nat. Genet. 27, 383–391
75. Daly, A. K., King, B. P., and Leathart, J. B. (2006) Genotyping for cyto-chrome P450 polymorphisms. Methods Mol. Biol. 320, 193–207
P450 3A and Morphine Synthesis
20210 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 33 • AUGUST 14, 2015
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

GuengerichValerie M. Kramlinger, Mónica Alvarado Rojas, Tatsuyuki Kanamori and F. Peter
Step in Endogenous Mammalian Morphine Biosynthesis-Demethylation of Thebaine, a Key6OCytochrome P450 3A Enzymes Catalyze the
doi: 10.1074/jbc.M115.665331 originally published online July 8, 20152015, 290:20200-20210.J. Biol. Chem.
10.1074/jbc.M115.665331Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/290/33/20200.full.html#ref-list-1
This article cites 71 references, 28 of which can be accessed free at
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from