corticosteroids and ascorbicacid transport in … · ascorbic acid in guinea-pig adrenal-cortex...
TRANSCRIPT

564 S. E. DREWES AND D. G. ROUX 1964Bognar, R., Rakosi, M. & Wheeler, T. S. (1962). Tetrahedron,
18, 135.Bokadia, M. M., Brown, B. R. & Cummings, W. (1960). J.
chem. Soc. p. 3308.Chan, W. R., Forsyth, W. G. C. & Hassall, C. H. (1958).
J. chem. Soc. p. 3174.Drewes, S. E. & Roux, D. G. (1964a). Biochem. J. 90,
343.Drewes, S. E. & Roux, D. G. (1964 b). Biochem. J. 92, 555.Freudenberg, K. & Maitland, P. (1934). Liebigs Ann. 510,
193.Geissman, T. A. & Clinton, R. 0. (1946a). J. Amer. chem.
Soc. 68, 697.Geissman, T. A. & Clinton, R. 0. (1946b). J. Amer. chem.
Soc. 68, 706.Goel, R. N., Jain, A. C. & Seshadri, T. R. (1958). Proc.Indian Acad. Sci. 48, 180.
Hattori, S. (1925). Acta phytochim. 2, 99.Hill, D. W. (1936). Chem. Rev. 19, 23.
Jurd, L. & Geissman, T. (1963). J. Amer. chem. Soc. 28,2394.
Lillya, C. P., Drewes, S. E. & Roux, D. G. (1963). Chem. &Ind. p. 783.
Nadkarni, D. R. & Wheeler, T. S. (1938). J. chem. Soc.p. 1320.
Roux, D. G. (1957). Nature, Lond., 179, 305.Roux, D. G. (1963 a). Biochem. J. 87, 435.Roux, D. G. (1963 b). Leder, 14, 179.Roux, D. G. (1963c). J. Chromat. 10, 473.Roux, D. G. & de Bruyn, G. C. (1963). Biochem. J. 87, 439.Roux, D. G. & Freudenberg, K. (1958). Liebigs Ann. 613,
56.Roux, D. G., Maihs, A. E. & Paulus, E. (1961). J. Chromat.
5,9.Roux, D. G. & Paulus, E. (1961). Biochem. J. 80, 476.Roux, D. G. & Paulus, E. (1962). Biochem. J. 84, 416.Saiyad, I. Z., Nadkarni, D. R. & Wheeler, T. S. (1937).
J. chem. Soc. p. 1737.
Biochem. J. (1964), 92, 564
Corticosteroids and Ascorbic Acid Transport in Adrenal Cortex in vitro
BY SHAIL K. SHARMA, ROSE M. JOHNSTONE AND J. H. QUASTELMcGill-Montreal General Ho8pital Re8earch Institute, 3619 University Street, Montreal, P.Q., Canada
(Received 30 December 1963)
We have previously reported (Sharma, John-stone & Quastel, 1963) that the uptake of [1-14C]-ascorbic acid in guinea-pig adrenal-cortex slices is aprocess dependent on metabolic energy and inhibit-ed by dinitrophenol and anaerobiosis. The con-centration of [1_-4C]ascorbic acid in the tissue mayrise to five times that of the medium. Extraction ofthe tissue with 85 % (v/v) ethanol, followed bychromatography and radioautography, showed thepresence of a single component whose RF corre-sponded to that of ascorbic acid. ACTH,* whichdepletes the adrenal gland of ascorbic acid in vivo(Sayers, Sayers, Liang & Long, 1945, 1946; Long &Fry, 1945), inhibits the active uptake of [1-14C]-ascorbic acid in guinea-pig adrenal-cortex slicesin vitro. Moreover, the inhibition was obtainedonly under conditions leading to adrenal-steroidsynthesis. In the absence of glucose, ACTH had noeffect on ascorbic acid uptake. It is already wellestablished that added glucose is required to obtainenhanced steroid synthesis by adrenal-corticaltissue in vitro in the presence ofACTH (Schonbaum,Birmingham & Saffran, 1956). Adenosine 3',5'-monophosphate, which stimulates adrenal steroidsynthesis in vitro (Haynes, Koritz & Peron, 1959;Birmingham, Kurlents, Lane, Muhlstock & Trai-kov, 1960), also inhibited the uptake of [1-14C]-
ascorbic acid (Sharma et al. 1963). As corticosteroidsinhibit ascorbic acid uptake (Sharma et al. 1963), weinvestigated further the nature and the specificity ofthe effects of steroids on ascorbic acid uptake.
MATERIALS AND METHODS
Adrenal-cortex slices. The preparation of guinea-pigadrenal-cortex slices and the incubation procedure that wehave used are fully described by Sharma et al. (1963). Theincubations were carried out in Krebs-Ringer phosphatecontaining NaCl (134 mM), KCI (5-2 mM), KH2P04 (1-3 mM),MgSO4 (1 3 mM), CaCl2 (2-8 mM) and sodium phosphatebuffer, pH 7-4 (10 mM). Glucose (10 mm) and thiourea(1 mM) were also added. All experiments were run induplicate.
Adrenal-cortex homogenate. Adrenal-cortex slices werehomogenized in ice-cold Krebs-Ringer phosphate solution(400 mg. wet wt. of tissue in 20 ml. of Krebs-Ringer phos-phate) with a Teflon-pestle homogenizer. A 0-5 ml. sampleof the homogenate was added to each Warburg manometricvessel. The total volume used was 4 0 ml.
Influx experiments. The influx of [1-_4C]ascorbic acid wasmeasured as described by Sharma et al. (1963). When mea-suring uptake of [1-_4C]ascorbic acid it is necessary to addthiourea (1 mM) to the incubation medium to preventoxidation of ascorbic acid in the adrenal tissue. This is notnecessary for brain slices.
Efflux experiments. The adrenal-cortex slices, after pre-incubation as described by Sharma et al. (1963), were* Abbreviation: ACTH, adrenocorticotrophic hormone.

CORTICOSTEROIDS AND ASCORBIC ACID TRANSPORTincubated aerobically (in 02) at 370 in the presence of[1-_4C]ascorbic acid for 60 min. The vessels were thenplaced in crushed ice.The slices were rinsed in ice-cold Krebs-Ringer medium
and transferred to another set of vessels containing a freshportion of 3 ml. of Krebs-Ringer phosphate medium. Thisnew medium is the same as that above but no [14C]ascorbicacid was added. The vessels were regassed and reincubatedat 37°. After given intervals of time, the vessels wereremoved from the bath and a sample (0-2 ml.) of the fluidwas plated quickly on an aluminium planchet. The vesselswere regassed after each sample had been taken out and theincubation was continued. Each sampling time took 5-6 min. Finally the vessels were placed in crushed ice, andthe slices were rinsed, homogenized and extracted with 80%(v/v) ethanol as described by Sharma et al. (1963).
Dialysis. After incubation at 370 the homogenates weredialysed in a cellophan dialysing bag (i in. diam.) containingfour to six glass beads. The bag was placed in a conical flaskcontaining 600 ml. of Krebs-Ringer phosphate solutionwithout Ca2+ ions at pH 7-4. To agitate the dialysing solu-tion, a magnetic stirrer was used. The dialysing sac wasrotated approx. 12 times/min. at 40 for 24 hr. The dialysingsolution in the conical flask was changed five times duringthe dialysis.The dialysis residue was plated on weighed planchets,
dried, weighed and its radioactivity measured. A controlexperiment with the radioactive substance without tissuewas also carried out.
Steroid estimation. The steroids were estimated by a pro-cedure based on methods used by Saffran & Schally (1955)and Elliott, Birmingham, Schally & Schonbaum (1954).After incubation, the medium was divided into two equalportions: one portion (A) was used to estimate the steroidconcentration and the other (B) was used as a source ofsteroid synthesized in situ for further experiments.Medium incubated without tissue served as a control andwas treated in the same way as that for the medium inwhich tissue had been incubated. A 0-4 ml. sample of themedium was transferred into each of two glass-stopperedcentrifuge tubes of 2-5 ml. capacity. The contents of eachtube were extracted with methylene dichloride (3 x 1 ml.)by shaking and centrifuging. The combined methylenedichloride extracts were evaporated at room temperature ina current of pure N2.
When the tubes were completely dry, 1-0 ml. of methanolwas added to tube A. The tube was stoppered, shaken welland kept for 30 min. at room temperature. Part of thesolution was transferred to a quartz microcuvette andreadings were taken at 225, 240, 255 and 260 mu in a Beck-mann model DU spectrophotometer. The steroids have amaximum absorption peak at 240 m,u. When no peak atabout 240 m,u was obtained, interfering substances wereprobably present, and the readings were discarded. Thiswas a rare occurrence. The formula suggested by McKerns& Nordstrand (1955) has been used for the calculation ofthe amount of steroid present.
Resuspension of the steroid in tube B was made in 50%(v/v) ethanol. A 0-02 ml. sample of this solution was usedin the incubation medium. In the experiments in whichsteroids dissolved in ethanol were used, the effect of 0-02 ml.of50% ethanol was examined separately. All steroids used,including [4-14C]cortisone, were made up in 50% ethanoland the final ethanol concentration in the incubationmedium was 1% (v/v). [1-_4C]Ascorbic acid was freshlyprepared for each experiment and dissolved in 10 mM-thiourea (the final concentration of thiourea in the incuba-tion medium was 1 mM).
RESULTS
Effects of sodium ions and of ouabain on ascorbicacid uptake. It has become increasingly apparentthat Na+ ions are required for the active transportof a number of amino acids and carbohydrates in avariety of cell types (Ricklis & Quastel, 1958; Csaky& Thale, 1960; Bihler & Crane, 1962; Takagaki,Hirano & Nagata, 1959; Abadom & Scholefield,1962). The active transport of [1-14C]ascorbic acidin guinea-pig adrenal-cortex slices is also dependenton Na+ ions, 50 % inhibition of transport beingobserved when the sodium chloride is replaced bycholine chloride (Table 1). When 50% ofthe sodiumchloride is replaced by choline chloride, inhibitionof ascorbic acid uptake is not as extensive but it isstill significant. In the absence of Na+ ions, ascor-bate transport is as effectively inhibited as in the
Table 1. Sodium ion requirement for the uptake of [1-14C]ascorbic acid by guinea-pig adrenal-cortex slicesGuinea-pig adrenal-cortex slices were preincubated in Krebs-Ringer phosphate medium, pH 7 4, containing
glucose (10 mM) at 370 for 45 min., and then incubated in a fresh medium, pH 7 4, containing glucose (10 mM) and[1-14C]ascorbic acid (0-1 mM; 50 000 counts/min./ml. of incubation medium) at 37° for 60 min. Choline chloridereplaced NaCl when this was omitted from the medium. Substances also present during this incubation were:KCI (5.2 mM), CaCl2 (2-8 mm), KH2PO4 (1.3 mM), MgSO4 (1-3 mm) and thiourea (1 mm). The final volume was1 ml. The gas phase was 100% 02. Each value reported in this and subsequent Tables is the mean ±S.D. of atleast four separate determinations.
Concn. of additions to finalincubation medium (mM)
Cholinechloride NaCl Ouabain
0 134 00 134 0.01
67 67 0134 0 0
Uptake of [1-14C]ascorbic acid
Tissue [1-14C]ascorbic acidCounts/min./100 mg. medium [1-14C]ascorbic acidwet wt. of tissue concentration ratio18500+1500 509 600+800 2-513 600+1 100 3-69 900+800 2'6
Qo23-43-13-03-2
Vol. 92 565
S. K. SHARMA, R. M. JOHNSTONE AND J. H. QUASTEL
Table 2. Uptake of [1-14C]ascorbic acid by guinea-pig adrenal-cortex slices in the presenceof steroid hormones
The conditions were as given in Table 1. All flasks contained 134 mM-NaCl. Steroids were dissolved in 50 %ethanol to give a final concn. of 0.1 mm, and 0-02 ml. of the ethanolic solution was added to each vessel. The totalvolume was 1-0 ml. A control with the same concentration of ethanol was carried out with each experiment.
Uptake of [1-14C]ascorbic acid
Steroid (0-1 mM)NoneHydrocortisoneOestradiolTestosteroneProgesteronePregnenaloneDehydroisoandrosteroneAldosterone
Tissue [1-14C]ascorbic acidCounts/min./100 mg. medium [1-14C]ascorbic acidwet wt. of tissue concentration ratio19 000±1 50013 000±1 10019 000±1 50019000±150017 000±1 40019 000±1 50017 000±1 40013 000±1 100
5-13.55-15-14-55.14-53.4
absence of metabolic energy (Sharma et al. 1963).There is an inhibitory effect of ouabain on ascorbicacid uptake (Table 1) which no doubt is related toits known effect on Na+ ion transport at cellmembranes.
Specificity of steroids inhibiting uptake of [1-14C]-ascorbic acid. Of the steroids normally producedby the adrenal cortex that were examined, onlythose that possess a ketol (HO- CH2 *CO-) sidechain at C-17 of the steroid nucleus inhibit theuptake of [1-14C]ascorbic acid (Table 2). We haveshown (Sharma et al. 1963) that cortisone, cortico-sterone and deoxycorticosterone are as effective as
hydrocortisone in inhibiting the uptake of [1-14C]-ascorbic acid. Progesterone, pregnenalone anddehydroisoandrosterone are all produced by theadrenal cortex but do not inhibit ascorbate uptakein guinea-pig-adrenal tissue.
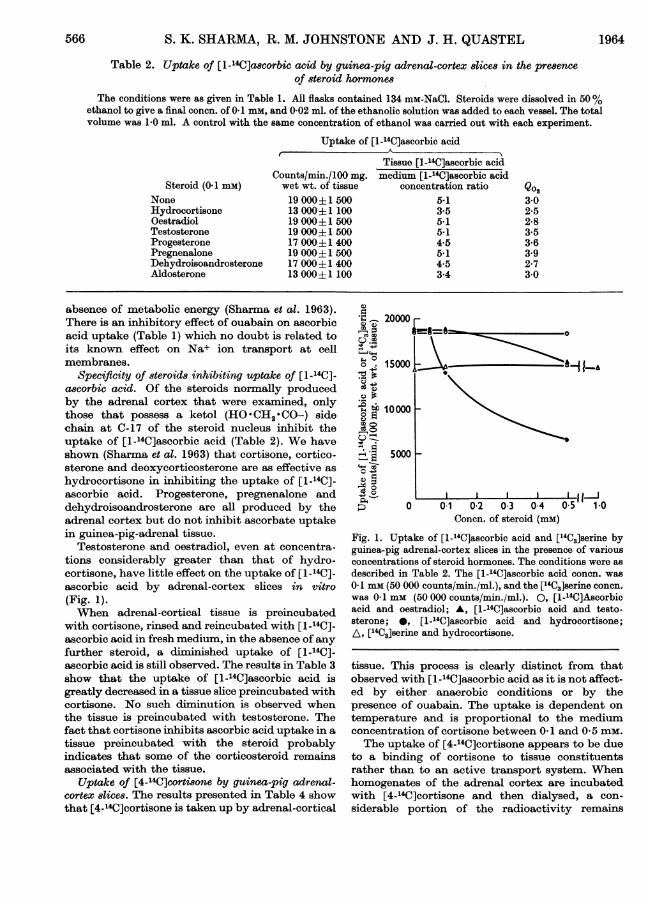
Testosterone and oestradiol, even at concentra-tions considerably greater than that of hydro-cortisone, have little effect on the uptake of [1-14C]-ascorbic acid by adrenal-cortex slices in vitro(Fig. 1).When adrenal-cortical tissue is preincubated
with cortisone, rinsed and reincubated with [1-14C]-ascorbic acid in fresh medium, in the absence of anyfurther steroid, a diminished uptake of [1-14C]-ascorbic acid is still observed. The results in Table 3show that the uptake of [1-14C]ascorbic acid isgreatly decreased in a tissue slice preincubated withcortisone. No such diminution is observed whenthe tissue is preincubated with testosterone. Thefact that cortisone inhibits ascorbic acid uptake in atissue preincubated with the steroid probablyindicates that some of the corticosteroid remainsassociated with the tissue.
Uptake of [4-_4C]cortisone by guinea-pig adrenal-corteX slices. The results presented in Table 4 showthat [4-'4C]cortisone is taken up by adrenal-cortical
5---_ 20000r
0o 15000
0
B
_>_ 5000
p4J'-Cao0
Is
i,,01 0-2 03 0-4 05 1.0
Concn. of steroid (mM)
Fig. 1. Uptake of [1-14C]ascorbic acid and [j4C3]serine byguinea-pig adrenal-cortex slices in the presence of variousconcentrations of steroid hormones. The conditions were as
described in Table 2. The [1-14C]ascorbic acid concn. was
0-1 mm (50 000 counts/min./ml.), and the ["4C3]serine concn.
was 0-1 mM (50 000 counts/min./ml.). 0, [1-14C]Ascorbicacid and oestradiol; A, [1-14C]ascorbic acid and testo-sterone; 0, [1-l4C]ascorbic acid and hydrocortisone;A, ['4C3]serine and hydrocortisone.
tissue. This process is clearly distinct from thatobserved with [1-14C]ascorbic acid as it is not affect-ed by either anaerobic conditions or by thepresence of ouabain. The uptake is dependent on
temperature and is proportional to the mediumconcentration of cortisone between 0.1 and 0 5 mM.The uptake of [4-14C]cortisone appears to be due
to a binding of cortisone to tissue constituentsrather than to an active transport system. Whenhomogenates of the adrenal cortex are incubatedwith [4-14C]cortisone and then dialysed, a con-
siderable portion of the radioactivity remains
Q023-02-52-83-53-63.92-73*0
1=6 _ a
566 1964
18s
L-

CORTICOSTEROIDS AND ASCORBIC ACID TRANSPORT
associated with the dialysis residue. [4-14C]-Cortisone, in the absence of adrenal-cortical tissue,is not retained by the dialysis sac. It is well knownthat corticosteroids are bound byproteins(Bellamy,Phillips, Jones & Leonard, 1962).
Transport of amino acid8 in guinea-pig adrenal-cortex slices in the presence of corticosteroids andadrenocorticotrophic hormone. The results in Table 5show that the uptake of [14C,]serine or ac-amino-[1-14C]isobutyrate is not affected by either ACTHor cortisone under conditions that cause a markedinhibition of ascorbic acid uptake.
Concentrations of hydrocortisone up to 1 mmscarcely affect [14C3]serine uptake, in contrast with
the marked inhibition of [1-14C]ascorbic acid uptake(Fig. 1). [14C3]Serine (0-1 mm) is concentrated byadrenal-cortex slices, a 14C ratio (cell water/medium) of 3-0 being obtained after 60 min. ofincubation. With ax-aminoisobutyrate (0.1 mm) theconcentration ratio is 2-0. A number of amino acids(all at 0- 1 mM), namely proline, glycine, glutamate,aspartate, methionine, phenylalanine, valine, leu-eine, y-aminobutyrate and 5-hydroxytryptophan,attained a concentration ratio of about 1.The uptake of [14C,]serine is inhibited by ouabain
and by anaerobiosis, but oc-amino[1-14C]isobutyrateuptake is apparently not affected. These resultssuggest that, of the amino acids examined, only the
Table 3. Uptake of [1-14C]ascorbic acid in adrenal-cortex slices preincubated with steroid hormones
Tissue slices were preincubated for 45 min. at 370 in Krebs-Ringer phosphate medium, pH 7-4, containingglucose (10 mM) and the steroids. The slices were rinsed thoroughly in cold Krebs-Ringer phosphate medium andreincubated in a fresh medium without any further addition of steroid and containing [1-14C]ascorbic acid (0.1 mM;50 000 counts/min./flask). The gas phase was 02 and the incubation time was 60 min. at 37°. All other conditionswere as given in Table 2.
Uptake of [1-14C]ascorbic acid
Addition topreincubation mediumNoneCortisone (1-0 mM)Testosterone (1.0 mm)
Counts/min./100 mg.wet wt. of tissue17 200±1 4009 000±700
17 000±1 400
Tissue [1-14C]ascorbic acidmedium [1-14C]ascorbic acid
concentration ratio4-82-54-7
Table 4. Uptake of [4-14C]cortisone by guinea-pig adrenal-cortex slices
The preincubation time was 45 min., and the final incubation was 60 min. at 37°. The conditions were as givenin Table 2.
AdditionNoneNoneNoneNoneOuabain (0-1 mM)Ascorbic acid (10 mM)
Temp.00
3737373737
Gasphase020202N20202
Concn. of[4-14C]cortisone
added(mM)0-10-10-50-10-10-1
Uptake of[4-14C]cortisone
(,umoles/g. wet wt.of tissue)
0-060-281-300-290-270-19
Table 5. Amino acid uptake by guinea-pig adrenal-cortex slices in the presence of adrenocorticotrophihormone and corticosteroids
The preincubation time was 45 min. and the final incubation was 60 min. at 37°. The conditions were as givenin Table 2. The ACTH conen. was 0-2 unit/ml., the cortisone conen. was 0-1 mm, and the test substance concn.was 0-1 mM (radioactivity 50000 counts/min./ml.).
Test substanceL-[14C3]SerineL-[14C,]SerineL-['4C3]Serineoc-Amino[1-14C]isobutyrateax-Amino[1-14C]isobutyrateac-Amino[l-14C]isobutyrate[1-14C]Ascorbic acid[1-14C]Ascorbic acid
Uptake of test substance(counts/min.1100 mg.wet wt. of tissue)13 700±1 10013700±1 10013600±11008 000±6008 000±6007 800±60019 000±1 50013600±1 100
Q023-02-52-4
AdditionNoneACTHCortisoneNoneACTHCortisoneNoneCortisone
Vol. 92 567

S. K. SHARMA, R. M. JOHNSTONE AND J. H. QUASTELuptake of [14C3]serine is an energy-dependentprocess in the adrenal cortex in vitro.The fact that ACTH and cortisone do not inhibit
the uptake of [14C3]serine shows that their effects onascorbic acid uptake are due to their associationwith a specific transport system rather than with ageneral transport phenomenon.
Uptake of [14C6]glucose and [2,3-'4C2]succinate inthe presence of adrenocorticotrophic hormone. With[14C6]glucose or [2,3-14C2]succinate as substrate, theamount of radioactivity found in the ethanol-soluble fraction of the tissue is unaltered inpresence of ACTH (Table 6). Moreover, the pro-duction of 14CO2 from [1-14C]glucose and [6-14C]-glucose is not affected by cortisone.
Steroids and ascorbic acid uptake in other tisses.Rat-brain-cortex slices, as well as guinea-pigadrenal-cortex slices, bring about the activetransport of [1-14C]ascorbic acid (Sharma et al.1963). The present results show that guinea-pig-ovary slices are also able to transport [1_14C]_ascorbic acid against a concentration gradient.Under the present experimental conditions a con-
Table 6. Uptake of radioactivity from [14C]glucoseand [14C]succinate by adrenal-cortex slices
The conditions were as given in Table 2, except thatascorbic acid was omitted. The radioactivity added in allcases was 50 000 counts/min./ml. of incubation medium.
Addition to finalincubation medium
[14C6]Glucose (2-0 mm)[14C65]Glucose (0-1 mM)[140C]Glucose (2-0 mM)[2,3-14C2]Succinate (2-0 mM)[2,3-14C2]Succinate (0-1 mM)[2,3-14C2]Succinate (2-0 mM)
Concn.(uniincmE
. ofACTH Uptake of 14Cit/ml. of (counts/min./ubation 100 mg. wetedium) wt. of tissue)0 24000 33000-2 24000 23000 37000-2 2200
centration ratio of 3 for the cell/medium distribu-tion of [1-_4C]ascorbic acid was obtained after1 hr. incubation with 041 mM-[1-14C]ascorbic acid(Table 7). As with brain and adrenal-cortex slices,the uptake of [1-14C]ascorbic acid in guinea-pig-ovary slices is decreased under anaerobic condi-tions.The effect of steroids on [1-14C]ascorbic acid up-
take in brain and ovary slices differs from thatobtained with adrenal-cortex slices. Oestradiol,testosterone and progesterone, as well as theadrenal-cortical steroids, inhibit the uptake ofascorbic acid in rat-brain-cortex slices. Most ofthese steroids also inhibit the uptake of [1-14C]-ascorbic acid in ovary slices (Table 7). ACTH,however, has no effect on ascorbic acid uptake inbrain or ovary preparations. The steroids thatinhibit ascorbic acid uptake at a concentration of0 1 mM have no effect on the respiratory activity ofthe rat-brain slices or the guinea-pig-ovary slices.Deoxycorticosterone and corticosterone, but nothydrocortisone or cortisone, inhibit the respira-tion of rat-brain slices at concentrations of 0 5 mm.In adrenal-cortex preparations, a concentration of0-1 mm-corticosteroid, which inhibits ascorbic aciduptake, has no effect on the respiratory activity.The uptake of [1-_4C]ascorbic acid in brain and
ovary slices, therefore, is not specifically inhibitedby adrenal-cortical steroids with a ketol side chainat C-17. All the steroids examined, whether ofovarian, testicular or adrenal origin, have similarinhibitory effects at equivalent concentrations onascorbic acid uptake in brain and ovary.Action of corticosteroids synthesized in situ on
ascorbic acid uptake. It has been shown that theuptake of [1-14C]ascorbic acid is inhibited by ACTHunder conditions that promote steroid synthesisand also by certain known corticosteroids. If thesteroids produced in response to ACTH are the
Table 7. Uptake of [1-14C]ascorbic acid by rat-brain-cortex and guinea-pig-ovary slicesThe [1-14C]ascorbic acid conen. was 0 1 mM, and the steroid concn. was 0 1 mm. The conditions of the experi-
ment were as given in Table 2. There was no preincubation period with rat-brain-cortex slices.
Uptake of [1-14C]ascorbic acid
By rat-brain-cortex slices By guinea-pig-ovary slices
Counts/min./100 mg.wet wt. of tissue12 700+1 0007 100+6006 500+500
6 400±5006 900+6007 600+6008 000+60010 000±800
Tissue [1-14C]ascorbic acidmedium [1-_4C]ascorbic acid
concentration ratio3-81.91-7
1-71-82-12-32-7
Counts/min./100 mg.wet wt. of tissue
11 500+9007 600±600
7 400+6006 800+600
Tissue [1-14C]ascorbic acidmedium [1-14C]ascorbic acid
concentration ratio3-01-8
1-81-7
SteroidNoneHydrocortisoneDeoxycortico-sterone
CortisoneCorticosteroneOestradiolTestosteroneProgesterone
568 1964

CORTICOSTEROIDS AND ASCORBIC ACID TRANSPORT
inhibitors of ascorbic acid uptake, it should bepossible to demonstrate that the steroids producedin 8itU are capable of inhibiting the uptake ofascorbic acid by adrenal-cortex slices.The results in Table 8 demonstrate the well-
known fact that ACTH stimulates the production ofsteroids in guinea-pig adrenal cortex. A two- tothree-fold increase in corticosteroids is usuallyobtained after 2 hr. of incubation with 0-2 unit ofACTH/ml.To estimate steroid synthesis under conditions
identical with those used for measurements ofascorbic acid uptake, we have examined the effectsof thiourea on steroid production. The results inTable 8 clearly show that 1 mM-thiourea has littleeffect on corticosteroid production in the presenceor absence of ACTH.
Steroids formed in situ in response to ACTH areconsiderably more active than the individualsynthetic corticosteroids tested in bringing aboutinhibitions of ascorbic acid uptake (Table 9).
Table 8. Steroid production in guinea-pig adrenal-cortex slices in the presence and absence of adreno-corticotrophic hormone and thiourea
TheACTH concn. was 0-2 unit/ml. ofincubation medium,and the thiourea conen. was 1-0 mm. Other conditions ofincubation were as given in Table 2. Adrenal-cortex sliceswere preincubated for 45 min. and then incubated for 2 hr.with 10 mM-glucose at pH 7-4 and 37°. The steroids in themedium were extracted with methylene dichloride andmeasured by the ultraviolet-absorption method.
AdditionsNoneThioureaACTHThiourea +ACTH
Steroid production(,ug./100 mg. wetwt. of tissue)
3-84-09-28-2
To obtain and test the steroids synthesized in 8ituthe procedure adopted was as follows. Equalweights of guinea-pig adrenal-cortex slices wereincubated for 2 hr. with and without ACTH in anormal Krebs-Ringer medium containing glucose(10 mM). The tissue was discarded and the mediumwas extracted with methylene dichloride (see theMaterials and Methods section). Methylene di-chloride was evaporated under a stream of nitrogenand the residue redissolved in 50% ethanol. Freshadrenal-cortex slices were prepared in a freshincubation medium containing [1-14C]ascorbic acidand appropriate samples of the ethanol extractwere added to give the desired concentration ofsteroid (see the Materials and Methods section). Theethanol extract from tissue, incubated withoutACTH, served as a control, and the volumes ofethanol extract used from this preparation cor-responded to those used from the ACTH-containingpreparation. Uptake of ascorbic acid was thenmeasured in the usual way. The results in Table 9show that the steroids produced in 8itu are muchmore effective than an equivalent concentration ofcortisone in inhibiting the uptake of [1-14C]ascorbicacid. Even at a concentration of 5jug./rnl. (equiva-lent to 0-017 mM-cortisone) there is an appreciabledecrease in uptake, corresponding to that obtainedwith 30 ag. of cortisone. At concentrations below30,ug./ml. (i.e. 0- 1 mM) none of the conunerciallyavailable corticosteroids produces a significantinhibition of uptake of [1-14C]ascorbic acid.An ethanol extract ofthemedium incubated with-
out ACTH has no significant effect on the uptake of[1-14C]ascorbic acid. These results therefore supportthe hypothesis that the substances, presumablycorticosteroids, produced in response to ACTH arethe most effective inhibitors of ascorbic acid uptake.
All the commercial corticosteroids we have testedwith a ketol side chain at C-17 were found to give
Table 9. Effect of steroids formed in situ on [1-14C]ascorbic acid uptake by guinea-pig adrenal-cortex slicesThe preincubation time was 45 min., and the final incubation was 60 min. at 37°. The conditions were as given
in Table 2.Uptake of [1-14C]ascorbic acid
A
Source of steroidNoneFormed in situ*
Cortisone
Control medium
Concn. ofsteroid Counts/min./100 mg.(pg./m1.) wet wt. of tissue
0 19400+150030 10 000±80015 14000±11005 15000±120030 14000±110015 18700±15005 19 000±1 5000 18400+1 5000 22400±1 5000 20000±1 500
Tissue [1-14C]ascorbic acidmedium [1-14C]ascorbic acid
concentration ratio5.32-53-63-83-75-05-25-05-75-3
* The quantities of steroid formed in situ are given in equivalents of cortisone (see the Materials and Methods section).
569Vol. 92

S. K. SHARMA, R. M. JOHNSTONE AND J. H. QUASTELalmost the same amount of inhibition of ascorbicacid uptake at the concentrations used. The factthat the steroids synthesized in BitU are appreciablymore effective than any of the connnercially avail-able products at equivalent concentrations indi-cates that either (a) a mixture of steroids is moreeffective than a single compound, or (b) a particularsteroid synthesized in 8itu is a far more effectiveinhibitor than any of the compounds examined. Totest the first possibility a mixture of four steroids,namely cortisone, hydrocortisone, corticosteroneand deoxycorticosterone, was used at a final totalconcentration of 0-2 mm, each steroid concentrationin the mixture being 0 05 mm, and its effects werecompared with that of each of the four at 02 mm.The results in Table 10 show that a mixture ofsteroids is more effective in bringing about inhibi-tions of uptake of [1-14C]ascorbic acid than is eachsteroid examined separately. However, even amixture of the steroids is considerably less effectivethan equivalent concentrations of the steroids syn-thesized in 8itu. These results suggest that a particu-lar corticosteroid, which has so far not beenexamined in the present work, is more effectivethan other corticosteroids in blocking the uptake of[1-14C]ascorbic acid in adrenal-cortex slices.
Efflux of ascorbic acid. ACTH is known to depletethe adrenal gland of ascorbic acid in vivo. Theadrenal gland is not known to synthesize ascorbicacid. Presumably, under normal conditions, theascorbic acid in the tissue is at a steady state, theuptake from the blood stream being balanced by acorresponding loss from the tissue. It is known thatin certain transport systems inhibition of the up-take will result in an efflux of material from the cell(Johnstone & Scholefield, 1959; Gonda & Quastel,1962).We have now been able to demonstrate that con-
ditions that bring about a decreased uptake of[14C]ascorbate also bring about a depletion of tissue
[14C]ascorbate. Thus the addition of ACTH orcorticosteroids to adrenal tissue preincubated with[1 -14C]ascorbic acid increases the loss of [14C].ascorbic acid from the tissue.
In these experiments the tissue slices were pre-incubated with 0.1 mM-[1-14C]ascorbic acid, thenrinsed and placed in fresh medium without furtherascorbic acid but containing, where indicated,ACTH or corticosteroids.
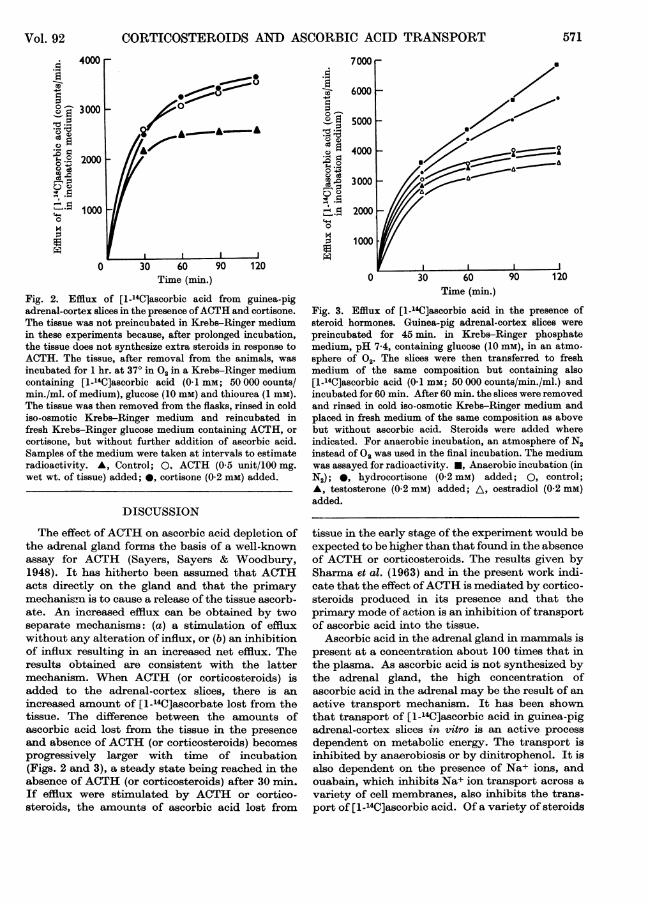
It is to be expected that some [1-14C]ascorbic acidwill be lost from all the tissues on reincubation infresh medium containing no ascorbic acid. Theefflux ceases when the rate of loss from the tissueand the rate of transport back into the tissuebecome balanced, i.e. when a new steady state isreached. If, however, the transport back into thetissue is blocked by ACTH or corticosteroids, theefflux of radioactivity will continue for a longer time.The results presented in Fig. 2 show the effects of
ACTH and cortisone on the rate of efflux of [1-_4C]-ascorbic acid from adrenal-cortex slices. It isapparent that initially (during the first 30 min.) theamount of radioactivity lost is approximately thesame under all conditions. In the presence ofACTH or cortisone the efflux continues, whereas intheir absence a steady state is attained. Similarlyhydrocortisone increases the loss of [1_-4C]ascorbicacid from the tissue, but oestradiol and testosteronehave no effect (Fig. 3).
Anaerobiosis, which also inhibits transport of[1-14C]ascorbic acid in adrenal-cortex slices, en-hances the loss of [14C]ascorbate from the tissue(Fig. 3). Clearly, therefore, under conditions whichinhibit the transport of ascorbic acid, an increasedloss of [14C]ascorbate from tissue preincubated with[1-14C]ascorbic acid can be attained. These condi-tions are analogous to conditions in vivo and indi-cate that ACTH and corticosteroids 'deplete' theadrenal gland of ascorbic acid in vitro as well asin vivo.
Table 10. Inhibition of [1-_4C]ascorbic acid uptake in guinea-pig adrenal-cortex slice8 by a mixtureof 8teroids
The [1-14C]ascorbic acid conen. was 0-1 mm. The preincubation time was 45 min., and the final incubation was60 min. at 37°. The commercial mixture contained hydrocortisone, cortisone, corticosterone and deoxycortico-sterone, each at 0 05 mM. The conditions were as given in Table 2.
Uptake of [1-14C]ascorbic acid
AdditionNoneFormed steroidsCommercial mixtureHydrocortisoneCortisoneCorticosteroneDeoxycorticosterone
Conen. ofsteroid(mM)
0
0-080-20-20-20-20-2
Counts/min./100 mg.wet wt. of tissue19 200+1 50010 000+80010 000+80014 400+1 10013000±1 10014400+1 10012 800+1 000
Tissue [1-14C]ascorbic acidmedium [1-14C]ascorbic acid
concentration ratio5.32-52-53.73.43-73-4
570 1964
CORTICOSTEROIDS AND ASCORBIC ACID TRANSPORT
* 4UU _
2000
, 1000 /0
0 30 60 90 120Time (min.)
Fig. 2. Efflux: of [1_14Clascorbic acid from guinea-pigadrenal-cortexr slices in the presence ofAC:TH and cortisone.The tissue was not preincubated in Krebs-Ringer mediumin these exrperiments because, after prolonged incubation,the tissue does not synthesize extra steroids in response toACTH. The tissue, after removal from the animals, wasincubated for 1 hr. at 37° in°2 in a Krebs-Ringer mediumcontaining [1_14C]ascorbic acid (0- 1 mm; 50 000 counts/min./ml. of medium), glucose (10 mm) and thiourea (1 mm).The tissue was then removed from the flasks, rinsed in coldiso-osmotic Krebs-Ringer medium and reincubated infresh Krebs-Ringer glucose medium containing ACTH, orcortisone, but without further addition of ascorbic acid.Samples of the medium were taken at intervals to estimateradioactivity. A, Control; O, ACTH (0-5 unit/100 mg.wet wt. of tissue) added; *, cortisone (0-2mm) added.
DISCUSSION
The effect of ACTH on ascorbic acid depletion ofthe adrenal gland forms the basis of a well-knownassay for ACTH (Sayers, Sayers & Woodbury,1948). It has hitherto been assumed that ACTHacts directly on the gland and that the primarymechanism is to cause a release of the tissue ascorb-ate. An increased efflux can be obtained by twoseparate mechanisms: (a) a stimulation of effluxwithout any alteration of influx, or (b) an inhibitionof influx resulting in an increased net efflux. Theresults obtained are consistent with the lattermechanism. When ACTH (or corticosteroids) isadded to the adrenal-cortex slices, there is anincreased amount of [1-14C]ascorbate lost from thetissue. The difference between the amounts ofascorbic acid lost from the tissue in the presenceand absence of ACTH (or corticosteroids) becomesprogressively larger with time of incubation(Figs. 2 and 3), a steady state being reached in theabsence of ACTH (or corticosteroids) after 30 min.If efflux were stimulated by ACTH or cortico-steroids, the amounts of ascorbic acid lost from
7000f
- 6000
-~~50000
o 4000 ;
o 4 *.M0
4 3000
0 0 30 6 9 2
2000-~4-0
0 1000
0 30 60 90 120Time (min.)
Fig. 3. Efflux of [1-L4C]ascorbic acid in the presence ofsteroid hormones. Guinea-pig adrenal-cortex slices werepreincubated for 45 min. in Krebs-Ringer phosphatemedium, pH 7-4, containing glucose (10 mM), in an atmo-sphere Of 02. The slices were then transferred to freshmedium of the same composition but containing also[1-_4C]ascorbic acid (0-1 mM; 50 000 counts/min./ml.) andincubated for 60 min. After 60 min. the slices were removedand rinsed in cold iso-osmotic Krebs-Ringer medium andplaced in fresh medium of the same composition as abovebut without ascorbic acid. Steroids were added whereindicated. For anaerobic incubation, an atmosphere of N2instead of 02 was used in the final incubation. The mediumwas assayed for radioactivity. *, Anaerobic incubation (inN2); 0, hydrocortisone (0-2 mM) added; 0, control;A, testosterone (0-2 mM) added; A, oestradiol (0-2 mM)added.
tissue in the early stage of the experiment would beexpected to be higher than that found in the absenceof ACTH or corticosteroids. The results given bySharma et al. (1963) and in the present work indi-cate that the effect ofACTH is mediated by cortico-steroids produced in its presence and that theprimary mode of action is an inhibition of transportof ascorbic acid into the tissue.
Ascorbic acid in the adrenal gland in mammals ispresent at a concentration about 100 times that inthe plasma. As ascorbic acid is not synthesized bythe adrenal gland, the high concentration ofascorbic acid in the adrenal may be the result of anactive transport mechanism. It has been shownthat transport of [1-_4C]ascorbic acid in guinea-pigadrenal-cortex slices in vitro is an active processdependent on metabolic energy. The transport isinhibited by anaerobiosis or by dinitrophenol. It isalso dependent on the presence of Na+ ions, andouabain, which inhibits Na+ ion transport across avariety of cell membranes, also inhibits the trans-port of [1-14C]ascorbic acid. Of a variety of steroids
571Vol. 92

S. K. SHARMA, R. M. JOHNSTONE AND J. H. QUASTELexamined, only adrenal-cortical steroids with aketol group at C-17 inhibit the uptake of [1-14C]-ascorbic acid in adrenal-cortex slices. Rat-brain-cortex slices and ovary slices, which also activelytransport [1 -_4C]ascorbic acid, do not show anyspecificity for the ketol-containing corticosteroids,and are not affected by ACTH. Transport of[1-4C]ascorbic acid in brain and ovary preparationsis inhibited by oestradiol, testosterone and pro-gesterone as well as corticosteroids. In other tissuesexamined, including liver, pancreas, spleen, dia-phragm and testes, active transport of [1 -14C]-ascorbic acid was not observed.To determine whether the corticosteroids specifi-
cally inhibit ascorbic acid transport, the effects ofthe corticosteroids on the transport of other sub-stances were also studied. The uptake of ten aminoacids by adrenal-cortex slices was examined. Onlyserine was found to be transported by an energy-dependent ouabain-sensitive mechanism. ACTHand corticosteroids had no effect on the transport ofserine under conditions that brought about a con-siderable decrease of ascorbic acid transport.The uptake of radioactivity from [14C]glucose and
[2,3-_4C2]succinate was not affected by ACTH orcorticosteroids. 14CO2 production from [1 -14C]-glucose and [6-14C]glucose was not affected bycortisone. Neither ACTH nor corticosteroidsinhibited the respiratory activity of the tissuesexamined at the concentrations tested.
Clearly the corticosteroids specifically inhibit thetransport of ascorbic acid in adrenal-cortex slices.We have previously observed that the uptake of[1-14C]ascorbic acid is inhibited by a medium thathad been exposed to ACTH-stimulated slices.Steroids synthesized in situ in response to ACTHhave now been extracted and their effects onascorbic acid uptake measured. These steroids are atleast three times more effective inhibitors of ascorb-ate uptake than any of the synthetic corticosteroidsso far examined, significant inhibitions beingobtained at a concentration of 0-016 mM-steroid.Since the material synthesized in 8itU is not anindividual component but presumably a mixture ofsteroids, a mixture of commercial corticosteroidswas prepared and tested. This mixture was not aspotent as the mixture of steroids synthesized insitu. The high inhibitory activity of the steroidssynthesized in situ suggests that a particularcorticosteroid formed in the presence of ACTH ismore effective than others in blocking ascorbatetransport in the adrenal cortex. It seems possiblethat this hypothetical steroid might control ascorb-ate transport in a manner analogous to the actionof aldosterone on ion movements.
It has also been demonstrated that pre-exposureof the adrenal tissue to cortisone decreases thetransport of [1-14C]ascorbic acid. This effect is also
characteristic of the corticosteroids since pre-exposure to testosterone does not alter the trans-port. [4-_4C]Cortisone appears to be bound to thistissue by a temperature-dependent but energy-independent process.
It is suggested that the corticosteroids inhibittransport of ascorbic acid in adrenal-cortical tissueby their combination with the specific transport sitefor ascorbic acid. As, of the steroids examined,only corticosteroids with a ketol group at C-17 areeffective inhibitors, it is suggested that the ketolgroup of the steroid combines with this site. Themechanism of depletion of adrenal ascorbic acid byACTH seems to be due to an inhibition of the activetransport of ascorbic acid brought about by thecorticosteroids produced in response to ACTH.
SUMMARY
1. The transport of [1_14C]ascorbic acid in guinea-pig adrenal cortex in vitro is a Na+ ion-dependentouabain-sensitive process.
2. Of the steroids examined, only corticosteroidswith a ketol group at C-17 inhibit the transport of[1-14C]ascorbic acid in guinea-pig adrenal cortex.A number of other steroids of adrenal, ovarian andtesticular origin have no effect.
3. The transport of serine, an energy-dependentand Na+ ion-dependent process, in guinea-pigadrenal-cortex tissue is not inhibited by cortico-steroids. The uptake of a number of other aminoacids, glucose and succinic acid is not affected bycorticosteroids.
4. The pre-exposure of adrenal-cortex slices tocortisone, but not to testosterone, diminishes[1-14C]ascorbic acid uptake.
5. [4-14C]Cortisone is bound by adrenal cortex bya temperature-dependent but energy-independentprocess.
6. Guinea-pig-ovary slices, as well as rat-brain-cortex slices, transport [1_-4C]ascorbic acid by anenergy-dependent process. The uptake of [1-14C]-ascorbic acid in a number of other tissues, includingliver, pancreas and spleen, does not appear to beenergy-dependent.
7. The transport of [1-14C]ascorbic acid in rat-brain-cortex slices and guinea-pig-ovary slices isinhibited by corticosteroids as well as by oestradioland testosterone.
8. Steroids formed in 8itU in response to adreno-corticotrophic hormone inhibit the transport of[1-14C]ascorbic acid to a greater extent than doequivalent concentrations of commercially avail-able individual steroids or mixtures of the latter.
9. In adrenal-cortex slices preincubated with[1-14C]ascorbic acid an increased loss of the label isobtained under conditions that have been shown toinhibit transport of ascorbic acid. It is evident that
572 1964

Vol.92 CORTICOSTEROIDS AND ASCORBIC ACID TRANSPORT 573adrenocorticotrophic hormone and corticosteroidscause an efflux of ascorbic acid from adrenal-cortexslices in vitro, this being due to inhibition of activetransport into the tissue.We thank the National Cancer Institute of Canada and
the Medical Research Council of Canada for grants tosupport this work. We also gratefully acknowledge gifts ofpure ACTH from Canada Packers Ltd., Toronto.
REFERENCES
Abadom, P. N. & Scholefield, P. G. (1962). Canad. J.Biochem. Physiol. 40, 1603.
Bellamy, D., Phillips, J. G., Jones, C. & Leonard, R. A.(1962). Biochem. J. 85, 537.
Bihler, I. & Crane, R. K. (1962). Biochim. biophys. Acta, 59,78.
Birmingham, M. K., Kurlents, E., Lane, R., Muhlstock, B.& Traikov, H. (1960). Canad. J. Biochem. Physiol. 38,1077.
Csaky, T. Z. & Thale, M. (1960). J. Physiol. 151, 59.Elliott, F. H., Birmingham, M. K., Schally, A. V. &Schonbaum, E. (1954). Endocrinology, 55, 721.
Gonda, 0. & Quastel, J. H. (1962). Biochem. J. 84, 394.Haynes, R. C., Koritz, S. B. & Peron, F. G. (1959). J. biol.
Chem. 234, 1421.Johnstone, R. M. & Scholefield, P. G. (1959). Cancer Res.
19, 1140.Long, C. N. H. & Fry, E. G. (1945). Proc. Soc. exp. Biol.,N. Y., 59, 67.
McKerns, K. W. & Norstrand, E. (1955). Canad. J.Biochem. Phy8iol. 33, 209.
Ricklis, E. & Quastel, J. H. (1958). Canad. J. Biochem.Physiol. 36, 363.
Saffran, M. & Schally, A. V. (1955). Endocrinology, 56, 523.Sayers, G., Sayers, M. A., Liang, T. Y. & Long, C. N. H.
(1945). Endocrinology, 37, 96.Sayers, G., Sayers, M. A., Liang, T. Y. & Long, C. N. H.
(1946). Endocrinology, 38, 1.Sayers, M. A., Sayers, G. & Woodbury, A. (1948). Endo-
crinology, 42, 379.Schonbaum, E., Birmingham, M. R. & Saffran, M. (1956).
Canad. J. Biochem. Phy8iol. 34, 527.Sharma, S. K., Johnstone, R. M. & Quastel, J. H. (1963).
Canad. J. Biochem. Phy8iol. 41, 597.Takagaki, G., Hirano, S. & Nagata, Y. (1959). J. Neuro-
chem. 4, 124.
Biochem. J. (1964), 92, 573
Metabolism of Palmitate in Sheep
BY C. E. WEST AND E. F. ANNISONDepartment of Biochemistry and Nutrition, University of New England, Armidale, N.S.W., Australia
(Received 6 January 1964)
The metabolic importance of the plasma freefatty acid fraction of circulating lipids, first sug-gested by the observations of Dole (1956) andGordon & Cherkes (1956), has been confirmed bymeasurements of turnover rate based on isotopedilution in man (Laurell, 1957; Frederickson &Gordon, 1958; Havel, 1961), dogs (Bierman,Schwartz & Dole, 1957; Armstrong et al. 1961) andrats (Olivecrona, 1962). Single-injection studieswith [14C]palmitate or [14C]oleate have formed thebasis of most investigations, but Armstrong et al.(1961) measured turnover of free fatty acid in dogsby a continuous-infusion technique. In this investi-gation, as in most others, plasma free fatty acid wastreated as a single metabolic fraction and theassumption made that the metabolism of theinjected fatty acid (palmitate) was representativeof that of the whole plasma free fatty acid fraction.We have measured entry rates of palmitate in sheepby isotope-dilution procedures based on the con-tinuous infusion of [14C]palmitate. This acid wasisolated from the other components of plasma freefatty acid by chromatography before measurement
of specific radioactivity. Comparison of the specificradioactivities of plasma palmitate and bloodcarbon dioxide allowed the contribution ofpalnitate to total carbon dioxide production to beestimated roughly. The transfer of radioactivityfrom palmitate into other plasma lipids and certainblood constituents was also followed.
MATERIALS AND METHODS
Animals. Merino ewes and wethers (age 2-3 years) werehoused indoors and fed with lucerne chaff (800 g./day). Theanimals were trained to stand quietly in stocks, sheep fail-ing to respond to training (about 50%) being rejected.
Measurement of rate of entry of palmitate. The generalprocedure described by Annison & White (1961) for themeasurement of glucose entry rates was used. Labelledpalmitate in albumin (0 75%) was infused at 100c,0-05 jLmole/min., without using a priming dose (Armstronget al. 1961). Constancy of plasma FFA* concentration is aprerequisite for entry rate measurements, but FFA con-centrations in sheep rise sharply in response even to mildexcitement. Disturbance of the animal due to handling or
* Abbreviation: FFA, free fatty acid.