coral disease diagnostics whataem.asm.org/content/early/2006/12/08/aem.02172-06.full.pdf3 48...
TRANSCRIPT

1
CORAL DISEASE DIAGNOSTICS: WHAT’S BETWEEN A PLAGUE AND A BAND? 1
2
Ainsworth TD1, Kramasky–Winter E
2, Loya Y
2, Hoegh-Guldberg O
1, Fine M
3 3
4
5
6
7
1Centre for Marine Studies, and The ARC Centre of Excellence for Coral Reef Studies 8
The University of Queensland, St. Lucia, QLD, 4072 Australia, 2Tel Aviv University, Tel 9
Aviv, Israel, and Faculty of Life Sciences, Bar-Ilan University, The Interuniversity 10
Institute for Marine Science, Eilat 88103, Israel. 11
12
Corresponding Author: 13
Tracy Ainsworth 14
Centre for Marine Studies 15
University of Queensland 16
Brisbane, 4072. Australia 17
Email: [email protected] 18
Phone: +61 7 3365 3548 19
Fax: +61 7 3365 4755 20
21
Keywords: Coral disease, histopathology, FISH, The Red Sea 22
Running Title: Coral diseases of Eilat, The Red Sea. 23
24
ACCEPTED
Copyright © 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.02172-06 AEM Accepts, published online ahead of print on 8 December 2006
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2
Abstract 25
Recently reports of coral disease have increased significantly across the world’s tropical 26
oceans. Despite increasing efforts to understand the changing incidence of coral disease 27
very few primary pathogens have been identified and most studies remain dependent on 28
the external appearance of corals for diagnosis. As such our current understanding of 29
coral disease, disease progression and the underlying cause of disease is very limited. In 30
the present study we use structural and microbial studies to differentiate different forms 31
of black band disease; atypical black band disease and typical black band disease. 32
Atypical black band diseased corals were infected with the black band disease microbial 33
consortium yet did not show any of the typical external signs of black band disease based 34
on macroscopic observations. In previous studies, these examples, here referred to as 35
atypical black band disease, would have not been correctly diagnosed. We also 36
differentiate white syndrome from white disease on the basis of tissue structure and 37
presence/absence of microbial associates. White diseases are those with dense bacterial 38
communities associated with lesions of symbiont loss and/or extensive necrosis of 39
tissues, while white syndromes are characteristically bacterial free with evidence for 40
extensive programmed cell death/apoptosis associated with the lesion and the adjacent 41
tissues. The pathology of coral disease as a whole requires further investigation. This 42
study emphasizes the importance of going beyond the external macroscopic signs of coral 43
disease for accurate disease diagnosis. 44
45
46
47
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3
Introduction 48
Coral disease is considered an important factor in the recent decline of coral reefs 49
worldwide (29, 30, 38, 64). Reports of disease and disease-like syndromes in reef-50
building corals have increased substantially since first being reported in 1973 (10). This 51
increase in the incidence of disease is due in part to a better awareness of coral health, but 52
is also linked to the increased environmental stresses affecting coral reefs (30, 31, 35, 53
44). Between 18 to 30 diverse coral diseases and syndromes are described worldwide 54
(29, 64, 65) on the basis of macroscopic features. Coral disease diagnosis is primarily 55
macroscopic, taking into account characteristics such as the extent of tissue loss, tissue 56
colour and exposure of coral skeleton. These macroscopic disease signs then become the 57
basis for nomenclature and diagnosis. While these characteristics allow broad 58
descriptions of change on reefs, they are unreliable for accurate disease diagnosis and do 59
not increase our understanding of the causes of coral disease and disease progression. 60
61
The reefs of Eilat have endured several decades of high levels of anthropogenic impact 62
(38, 39, 66). As a result coral disease and coral mortality have increased (39, 67) 63
resulting in decreases in coral abundance and diversity over the past 3 decades. 64
Increasing stress from global warming and ocean acidification has also been associated 65
with the overall declining health of Red Sea corals (40). Al- Moghrabi (5) have reported 66
outbreaks of black band disease in the northern Red Sea, and Antonius and Riegl (13) 67
have reported white syndrome on the reefs of the Sinai Peninsula with the most abundant 68
reef building species (Acropora hemprichii) suffering heaviest losses. Ben-Haim et al 69
(17) also reported a new disease of Pocillopora damicornis from the Gulf of Aqaba. In 70
addition anecdotal reports of black band disease became common in the early 1990s in 71
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4
the Eilat side of the Gulf of Aqaba although no study was conducted (Loya pers comm.). 72
Recently Barash et al (17) reported that many of the massive corals of Eilat were affected 73
by an infectious white plague-like disease. Despite reports and field observations of these 74
diseases and syndromes within the region, relatively little is known about the pathology, 75
cytology, microbial ecology and disease processes of corals in the Gulf. 76
77
While there have been numerous ecological and field based studies of corals exhibiting 78
disease (49, 50, 51), few primary pathogens have been identified to date (11, 16, 18, 24, 79
31, 32, 34, 36, 48, 53). Also few studies have examined the histopathological and 80
microbial characteristics of diseased corals. Black band disease (BBD) is considered one 81
of the major diseases impacting coral reefs worldwide. The first report of BBD was by 82
Antonius in 1973 (10). Since that time BBD has been observed to affect corals 83
worldwide, especially in polluted environments (5, 13). The black band that is typical of 84
this disease is composed of a mixed microbial mat that is dominated by cyanobacteria 85
and comprises sulphur reducing and sulphur oxidising bacteria, as well as a number of 86
other micro-organisms. The mat overgrows coral tissues, creating a toxic environment 87
and tissue loss is attributed to the presence of high sulphide levels (0.8 mM) in the tissues 88
adjacent the black band (22, 54). The distinct macroscopic signs of this disease and the 89
growth pattern of the mat in a top-down manner across polyps and tissue structures (10, 90
27, 54) are used as the primary method of the disease identification. BBD has been 91
described as one of the main diseases in the Caribbean and Florida Keys where it has 92
been responsible for reef decline (37). Most active during the summer months, BBD is 93
one of the most widespread and destructive coral diseases due to its high impact on 94
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

5
massive and framework building corals with rates of tissue loss of up to 2 cm per day (27, 95
54). 96
97
White band and white plague diseases have also been described in many regions 98
worldwide, including the Red Sea and Gulf of Aqaba (5, 12, 13, 14, 17, 59). Field based 99
surveys and visual descriptions of these diseases have described them as typically having 100
a lesion of white recently exposed coral skeleton. It is believed that the white band 101
diseases have had a major role in the community structure shift occurring in the 102
Caribbean (15). Reefs off the coast of Florida have experienced increasing occurrence of 103
white diseases with patterns of disease spread suggesting a highly infectious nature (49). 104
While pathogens have been identified with some of the white diseases (24, 46, 48, 51), 105
others have been described as having no observable microbial community (13, 20, 21, 106
47). As such the majority of casual factors underpinning the rise in coral disease have 107
remained elusive particularly for many of the white diseases and white syndromes (21, 108
64). Richardson et al (52) has rightly called for an integrated approach to disease 109
diagnosis that incorporates field and laboratory studies. 110
111
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

6
In the present study we demonstrate that integrating observations on microbial diversity 112
characteristics with specific cytological observations may be a useful tool in 113
understanding the disease process of corals and improving the basis on which disease is 114
diagnosed. This study has also pointed to some significant problems associated with 115
the simplistic use of macroscopic signs as the only means of identifying coral disease as 116
observed on the Reef of Eilat, Gulf of Aqaba in the summer of 2005. 117
118
Materials and Methods 119
Samples collection and disease identification 120
Corals were surveyed visually for signs of disease in June (early summer) 2005 at depths 121
of 1 to 18 m on coral reefs near the Marine Biological Laboratory at Eilat in the Gulf of 122
Aqaba. Massive and branching corals were assessed for the typical macroscopic signs of 123
disease including: tissue loss and apparent rapid exposure of coral skeleton which is 124
indicative of white diseases and white syndromes, as well as general paling from 125
bleaching and the presence/absence of black bands. All colonies exhibiting signs of 126
disease were photographed using a Nikon CoolPix 5000 (Nikon, Inc) digital camera 127
inside a Subal (Steyr, Austria) underwater housing prior to sampling. Replicate tissue 128
and skeleton samples (n,3) were collected from corals displaying each the observed 129
diseases and syndromes to investigate microbial populations and histopathological 130
features of these diseases. Of the diseases observed in massive (Favid) coral colonies, 6 131
classified as having black band disease (BBD) and 6 classified as having white band 132
disease were sampled in both early and late June 2005. Of the white diseases and white 133
syndromes of other massive and of branching corals replicate samples of each type (n, 3) 134
were collected. 135
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

7
Sample preservation and tissue processing 136
Coral samples were fixed in 4 % (w/v) paraformaldehyde in sterile phosphate buffered 137
saline (PBS, pH 7.4)(20) for 12 hr, prior to decalcification with 20 % (w/v) EDTA (pH 8) 138
(63) and standard processing for paraffin embedding. Tissues were processed through 139
washes of 70 %, 80, two of 95 % and three of 100 % ethanol for 40 min each, three 140
xylene washes for 40 min and then 3 paraffin washes under vacuum for 40 min each prior 141
to embedding in paraffin. Serial tissue sections (4 µm) were collected onto Superfrost 142
Plus slides (Menzel, Brauschweig, Germany) for use in fluorescence in situ hybridisation 143
(FISH) and histopathology. 144
145
Fluorescence in situ hybridisation 146
Oligonucleotides were Cy3 labeled by Thermo Electron Corporation Pty Ltd, and used in 147
a conventional FISH protocol (6, 7, 8, 9, 41, 42, 43, 58). Paraffin sections were dewaxed 148
through 3 x 5 min washes in xylene and 4 x 5 min washes in fresh 100% ethanol. The 149
hybridisation was conducted in hybridisation buffer (0.9 M NaCl, 0.01% SDS, 0.01 M 150
Tris/HCl pH 7.2) for 1.5 to 2 hr at 46°C, followed by a 10 to 20 min wash in pre-warmed 151
(48°C ) wash buffer (0.08 M NaCl, 0.01 % SDS, 0.01 M Tris/HCl, 0.05 M EDTA) (3). A 152
Zeiss Meta 510 confocal scanning laser microscope (Zeiss, Germany) and spectral profile 153
imaging via the Zeiss Image Browser software were used to visualize tissue 154
autofluorescence and probe conferred fluorescence. Coral tissue sections treated with the 155
FISH protocol without the application of probe were used for spectral profiling (3). This 156
was repeated on several tissue sections for each coral species to determine variability in 157
fluorescence profiles. FISH probes used within this study include a universal bacterial 158
probe mix (EUBmix) and specific group probes for δ-proteobacteria (GAM42A), 159
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

8
Cytophaga-Flavobacterium (CF319) and Vibrio sp. (MV) (Table 1). These probes were 160
selected as these represent major bacterial groups within oceanic communities and as 161
many Vibrio sp are pathogenic. 162
163
Histopathology 164
The general tissue condition associated with the lesion and the adjacent tissues of each of 165
the diseases was ascertained following staining using Harris’s haematoxylin and eosin 166
(with phyloxine B) (Sigma-Aldrich Pty Ldt, # HHS32 and HT110-1-32). The extent of 167
mass tissue necrosis (swelling and lysis of cells, disruption of cell structure) was 168
recorded. In situ labelling of 3’ end of DNA fragments was used to investigate the 169
presence and extent of programmed cell death (1, 4, 25, 26) using the ApopTag in situ 170
apoptosis detection kit as per manufacturers recommendations (S7101, Chemicon 171
International, Inc. USA). This has been shown to distinguish apoptosis from necrosis by 172
specifically detecting DNA cleavage and chromatin condensation associated with 173
apoptosis and confirmed by lack of necrotic morphology. Cells were defined as apoptotic 174
if the nuclear area of the cell was positively labeled as indicated by red stain as opposed 175
to the blue haematoxylin counter-stained non-apoptotic nuclei (1). 176
177
Results 178
Black band disease. 179
A simultaneous outbreak of black band disease and white band disease in massive 180
corals (from the genus Favia) on reefs of Eilat (Red Sea) was observed during June 181
2005. Black band diseased corals showed typical signs of the disease characterized by 182
a thick black microbial mat that appeared to grow over and into underlying coral tissues 183
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

9
(Figure 1 A1, a1). The typical macro-scale signs of white band / plague disease in 184
Favia included an apparently clear lesion border between recently exposed bare white 185
skeleton and tissues. The lesion lacked any observable black, mixed microbial band on 186
the coral surface (Figure 1 B1, b1). Newly exposed clean white coral skeleton was 187
visible at the lesion interface and away from the disease lesion (Figure 1 b1). Based on 188
macro-scale signs these corals would be classified/diagnosed as suffering a white 189
disease or white plague. 190
191
Microbial and histopathological investigation of both black band (BBD) and white 192
band/white plague corals identified a similar distinctive cyanobacteria dominated 193
microbial mat. In the BBD the black encircling mat was easily visible on the colony 194
surface. In contrast the white disease/plague of Favia a similar black mat was found 195
deep within the polyp structure and underneath the disease lesion interface. This same 196
mat was identified in all white band sampled colonies in early and late June 2005 (n, 197
12). Given the similarity in cytological response and microbial community we refer to 198
this type of white band as atypical BBD (aBBD). Similar distinct cyanobacterial 199
dominated microbial communities was observed in both BBD and aBBD and appeared 200
to cause tissue lysis and necrosis (compare Figure 1 A2, a2 with Figure 1 B2, b2). 201
Imaging of the microbial communities identified similar morpho-type cyanobacteria 202
dominating the microbial community in both the typical (Figure 1A3,a3) and atypical 203
(Figure 1 B3, b3) black band diseased colonies. 204
205
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

10
White disease and white syndrome. 206
Both massive and branching corals on the Eilat reefs appeared to be suffering from white 207
disease, typified by abrupt lesions and areas of exposed skeleton adjacent to otherwise 208
normally pigmented tissues. White disease was identified in the massive colonies of 209
Hydnophora sp. (Figure 2 A1, a1) and Porites sp. as well as the branching coral S. 210
pistillata (Figure 4 A1, a1). White syndrome was identified in coral colonies of the 211
plating coral Acropora sp. (Figure 5 A1, a1) in the region. 212
213
There were distinct cytological and microbial differences between these macroscopically 214
similar syndromes. The white disease of Hydnophora sp. was characterized by extensive 215
tissue breakdown and mass tissue necrosis within the lesion areas (Figure 2 A2, a2). The 216
lesions were also extensively populated by bacteria, which were found to penetrate all 217
tissue layers and infiltrating adjacent regions of tissue (Figure 2 A3, a3). Investigation of 218
several tissue sections showed the bacteria were from both δ-proteobacterial (Figure 3 b) 219
and Cytophaga-flavobacterium (Figure 3c) groups, and communities of Vibrio sp. were 220
also found within the lesion (Figure 3d). 221
222
In contrast the white disease observed in the branching coral S. pistillata (Figure 4 A1) 223
exhibited patches of bleaching close to the lesion border and regions of tissue loss (Figure 224
4 a1). Within this white disease or form of white disease, extensive bacterial communities 225
were found in tissue layers associated with the lesion (Figure 3 A3, a3) where a range of 226
bacterial groups were evident, with no single bacterial group being dominant. There was a 227
lack of evidence for extensive tissue breakdown and necrosis (Figure 3 A2, a2), with only 228
small regions of necrosis in the lesion of white disease of S. pistillata, this necrosis was 229
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

11
limited only to regions of the gastroderm (Figure 4 aa2) presumably associated with 230
macroscopic areas of bleaching. 231
232
Disease signs consistent with descriptions of white syndrome were also observed in the 233
plating Acropora sp. with clear lesions evident between apparently healthy tissues and 234
recently exposed skeleton. The white syndrome identified on Acropora sp. (Figure 5 235
A1) had a distinctly clear lesion border that appeared to be moving quickly across 236
affected corals, as determined by the lack of macro-algal overgrowth over the large 237
exposed areas of skeleton. The exposed skeleton at the lesion was free of any evidence 238
for macro-algal overgrowth (Figure 5 a1) and tissues at the lesion border appeared 239
healthy despite disease progression (Figure 5 B1,b1). Microscopic analysis of the 240
lesion border showed it to be distinctly devoid of any significant bacterial populations 241
with tissue adjacent to the lesion lacking any evidence of tissue breakdown or necrosis 242
associated with the disease (Figure 5 A3, a3). This provides clear evidence of distinct 243
microbial community differences and morphological differences between white 244
syndrome and white diseases (Table 2), which requires further investigation. 245
246
Evidence of necrosis and apoptosis in disease. 247
Haematoxylin and eosin staining of tissue sections revealed extensive evidence for 248
necrosis associated with the tissue loss in Favid corals with black band disease. Tissue 249
breakdown consistent with necrosis was also evident in white disease in Hydnophora 250
sp. and Porites sp. Comparatively little/or no sign of necrosis or a mass loss of tissue 251
integrity were evident in the white disease of S. pistillata or Acropora sp.. In situ end 252
labeling of fragmented DNA used as a marker of programmed cell death, showed no 253
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

12
evident staining in tissues of the black band diseased massive corals, either typical 254
(BBD, Figure 6 a) or atypical black band (aBBD, Figure 6b,c), or in any of the tissues 255
associated with the necrotic white disease of Hydrophora sp. (Figure 6e,f, g). However 256
positive staining was evident in large regions of tissues associated with white syndrome 257
of Acropora sp. within both epithelial (Figure 6h) and gastrodermal tissue layers 258
(Figure 6i) while no staining was evident in healthy tissue in the same colonies (Figure 259
6j). Positive staining was also evident with the tissue layers of white disease of S. 260
pistillata, (Figure 6k, l) however this was limited only to cells directly adjacent to the 261
tissue lesions (Figure 6m). Cells showing morphology associated with necrotic cell 262
death showed no staining using in situ end labeling for fragmented DNA (Figure 6a-f). 263
Cell death markers consistent with programmed cell death were therefore only 264
identified in the apparent bacteria free lesion of Acropora sp. white syndrome and in 265
the lesion edge of the white disease of S. pistillata and not associated with BBD, or 266
where mass tissue necrosis was evident (Table 2) again demonstrating that white 267
syndrome is distinctly different from white diseases, here based on patterns of cell 268
death associated with the necrotic diseases (Table 2). 269
270
Discussion 271
While coral disease has been reported on the reefs of the northern Red Sea (5, 13, 14, 17, 272
39, 40, 59) this is the first study to explore histopathological changes and in situ 273
microbiology associated with these diseases and syndromes. This approach to the study 274
of coral disease has revealed that dependence on visual macroscopic characteristics alone 275
is unreliable for accurate diagnosis and overlooks critical information on the cellular and 276
microbial processes associated with disease-like states. The incorporation of 277
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

13
physiological assays, for example descriptions of photosynthetic assimilate translocation 278
(28, 55) which demonstrate colonial activity and integration, and assays addressing host 279
mechanisms (33) are also useful in understanding the coral colony responses to different 280
diseases and may provide important information for disease diagnosis. We suggest that 281
studies going beyond macroscopic disease signs by incorporating cytological, microbial 282
and physiological assays provide a greater detail to truly understand the processes of 283
disease in corals and will provide a better basis on which to make accurate disease 284
diagnosis (Figure 7). Accurate diagnosis of coral disease can direct research and 285
management practices towards addressing and managing the true underlying cause of 286
disease on reefs. 287
288
Atypical Black Band Disease 289
Rosenberg et al (59) and Barash et al (17) recently reported an anomalous temperature 290
spike in the Red Sea and a simultaneous outbreak of black band disease and white plague 291
in the summer months from 2001 to 2004. Attempts by these authors to determine 292
causative agents of the apparent white plague experienced difficulties in pathogen culture 293
and transmission of a suspected pathogen. In the present study histopathological 294
investigation revealed the presence of cyanobacterial dominated microbial mats deep 295
within the coral polyp structure of Favia exhibiting characteristics of white 296
disease/plague. These microbial populations and the pattern of coral tissue disruption 297
were consistent with those observed in the typical black band disease where 298
cyanobacterial filaments were found penetrating tissues deep within the polyp structure. 299
Based on the fact that the apparent white plague corals all contained cyanobacterial mats 300
deep in the tissues, this disease state appears to be an atypical form of black band disease, 301
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

14
also it appears the black band consortium can cause an apparent white disease or there 302
can be a progression from a white disease into black band disease. We suggest the use of 303
the term atypical black band disease (aBBD) to describe this different form of black band 304
disease. This conclusion is supported by the results of Bythell et al (21) who previously 305
observed an apparent progression from white plague to black band disease in the 306
Caribbean. The difficulties in pathogen identification and infection studies experienced 307
by Barash et al (17) may be due to the fact that some of these diseased corals were 308
actually infected by atypical BBD, illustrating the confusion that relying on macroscopic 309
characteristics alone may incur. We also suggest that the incidence of BBD on reefs 310
maybe estimated if this type of atypical form is common. Studies investigating BBD 311
should also attempt to determine the extent and impact of atypical forms of this disease in 312
other regions. 313
314
White Disease and White Syndrome. 315
White diseases and white syndrome of corals showed marked differences, evident both 316
from microbiological and cytological investigation; here we define ‘white diseases’ and 317
“white syndrome” of corals based on specific microbial and cytological parameters. 318
White diseases characteristically show extensive bacterial infiltration of the lesion tissue 319
layers and tissues adjacent to the disease lesion, as well as extensive necrosis, loss of 320
tissue structure and/or symbiont loss. White diseases also characteristically had little or 321
no evidence for programmed cell death associated with the disease lesions. The term 322
‘white disease’ may encompass a range of diseases or states of the disease, as seen in the 323
example of white disease of S. pistillata in this study. A similar macroscopic pattern of 324
tissue loss was apparent in the white disease of S. pistillata and the white disease of 325
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

15
Hydnophora sp., yet no mass loss of tissue structure and only small regions of necrosis of 326
the tissue layers were apparent in the white disease of S. pistillata. However dense and 327
mixed microbial communities were evident within the tissue layers characterising both 328
the white disease of massive and branching colonies, bacteria of Cytophaga-329
Flavobacterium and δ-Proteobacterial groups were present in both, as well as 330
communities of Vibrio sp., yet no single bacterial group appeared dominant. The potential 331
of secondary colonizers involved in white diseases and the role of opportunistic 332
pathogens in white disease progression maybe very important as evident by the diverse 333
microbial community of these diseases and also considering the source of highly diverse 334
microbial environments of the coral holobiont (19, 27, 45, 60, 61) and the coral reef. 335
336
In contrast there was a distinct lack of microbial community interaction associated with 337
the disease lesion of white syndrome. The disease lesion of white syndrome is 338
characteristically free of any evidence of mass tissue breakdown, with no loss of 339
structural integrity or necrosis, apparently healthy tissues bordered exposed coral 340
skeleton at the disease lesion. Extensive in situ end labeling of fragmented DNA, 341
suggestive of programmed cell death or apoptosis, was associated not only with the 342
tissue layers of the white syndrome lesion but also the adjacent tissues not directly 343
associated with the disease lesion. This is consistent with previous observations of 344
white syndrome of tabular Acropora sp. on the Great Barrier Reef (4, 56, 57). 345
Observations of extensive programmed cell death associated with white syndrome and 346
a lack of resource translocation to the white syndrome lesion site suggested the disease 347
to be a reaction of the host, which is progressing independently of an apparent pathogen 348
(4, 57). We suggest that white syndrome of corals in Eilat may be typical of a similar 349
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16
host reaction, or white syndrome, and that the syndrome is a divergent process of 350
disease progression to that seen in necrotic and bacterial white diseases. We therefore 351
define white syndrome of coral as characterized by a disease state that progresses 352
independently of an apparent pathogen or microbial colonization, with little or no 353
evidence for mass tissue necrosis and with extensive programmed cell death associated 354
with the lesion. 355
356
In this study we differentiate white syndrome from white diseases based on specific 357
cytological/morphological and microbial differences. Previous studies of coral disease 358
have also suggested white syndrome as being a distinct coral disease apparently linked 359
to a lack of observable microbial population associated with the disease progression 360
(13, 21). Weil (65) has defined a syndrome as a disease that has yet to have a causative 361
agent identified. However we suggest adhering to a medical definition of a “syndrome” 362
as one fulfilling a collection of specific signs that occur together to characterize the 363
particular disease or abnormality. We suggest that microbial, cytological and 364
physiological characteristics are useful criteria for differentiating coral diseases (Figure 365
7) and for determining the specific signs suitable for accurate disease diagnosis. This 366
also allows for the addition of other criteria, as shown in Figure 7, that may further 367
differentiate these diseases. Furthermore we postulate that the mechanisms of tissue 368
loss and disease progression in white diseases and white syndrome are divergent and 369
indicate the importance of understanding the physiological mechanisms that are 370
associated with these diseases to truly identify the underlying cause of their increases 371
worldwide. 372
373
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17
Primary versus opportunistic pathogens 374
Opportunistic pathogens are defined as those which infect compromised or previously 375
stressed individuals, whereas primary pathogens are those that cause disease in an 376
uncompromised host (62). The involvement of opportunistic, rather than primary 377
pathogens has been overlooked in coral disease research and needs to be considered 378
when attempting to understand disease and determine disease causation, disease 379
progression and colony mortality. This is further evident when considering the rapidly 380
changing and highly impacted environment of coral reefs. Increased environmental 381
stress and pollution may de-stabilize the coral holobiont creating increased potential for 382
possible opportunistic pathogens to affect the stressed corals. The question of 383
differentiating microbial pathogens involved in disease initiation and those involved in 384
disease progression or sources of secondary infections is of great importance. Future 385
work addressing disease causation should consider the definitions of various pathogens: 386
a primary pathogen, being the first infection by a pathogen in an uncompromised host 387
(62) and a secondary pathogen being one causing a secondary or subsequent infection 388
by multiplying within already diseased tissue but is not the primary pathogen (2). 389
Finally an opportunistic pathogen being a pathogenic organism that is normally 390
commensal but which gives rise to infection in compromised hosts (62). The use of a 391
range of diagnostic techniques will allow us to better understand the processes of 392
disease and address questions of underling causes of disease progression in dense and 393
diverse microbial ecosystems. 394
395
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18
Conclusions 396
This study has demonstrated the importance of cytological and in situ microbiological 397
studies in the investigation of coral disease. Comparative analysis of disease states 398
indicates that the current use of macroscopic disease signs is insufficient for 399
characterizing and understanding coral disease. A particularly good example is that of 400
black band disease within this study. The extent and impact of black band disease on 401
reefs worldwide may be underestimated if the lack of macroscopic disease signs, as 402
observed for Favia in Eilat, is common. Black band disease etiology and epidemiology 403
requires further analysis especially to determine if there is the potential of other diseases 404
to progress into this disease during summer months. Our study demonstrates that research 405
depending solely on macroscopic signs of disease run the risk of misdiagnosing diseases 406
among corals. 407
408
We strongly suggest that studies incorporating cytological, microbiological and 409
physiological investigations provide critical insights into the inception, progression and 410
causal factors underpinning the current global increase in coral disease. Accurate 411
diagnosis of coral disease is vital in providing researchers and managers with a better 412
basis for understanding disease causation on reefs. We therefore conclude that future 413
studies of coral disease, whether they be field or laboratory based must examine 414
fundamental cellular characteristics and physiological processes underlying disease 415
progression. 416
417
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

19
Acknolwedgements 418
The authors would like to thank the Eilat Coral Beach Nature Reserve and Dr David 419
Zakai for assistance and expertise in sampling and surveying corals of Eilat Reefs. The 420
authors are grateful for support provided by the GEF Coral Reef Targeted Research 421
Program (www.gefcoral.org) and the ARC Centre of Excellence for Coral Reef Studies 422
(www.coralcoe.org.au). The authors would like to thank the Centre for Advanced Light 423
Microscopy at the University of Queensland for assistance with confocal microscopy, and 424
also Professor Oded Yarden and Dr Bill Leggat for conceptual, logistical and editorial 425
assistance. 426
427
References 428
1. Adle-Biassette H, Bell JE, Creange A, Sazdovitch V, Authier FJ, Gray F, Hauw J-J, 429
and Gherardi R. DNA breaks detected by in situ end labelling in dorsal root ganglia of 430
patients with AIDS. Neuropathology and Applied Neurobiology (1998) 24:373-380. 431
2. Agrios G.N 1997. Plant Pathology 4th
Edition, Academic Press. 432
3. Ainsworth TD, Fine M, Blackall LL and Hoegh-Guldberg O (2006). Fluorescence in 433
situ hybridisation and spectral imaging of coral associated bacterial communities. Appl. 434
Environ. Micro. 72(4): 3016-3020 435
4. Ainsworth TD, Kvennefors EC, Blackall LL, Fine M. and Hoegh-Guldberg O. (In 436
Press). Disease and cell death in white syndrome of Acroporid corals on the Great Barrier 437
Reef. Mar Biol (In Press). 438
5. Al-Moghrabi, S. M., 2001: Unusual black band disease (BBD) outbreak in the 439
northern tip of the Gulf of Aqaba (Jordan). Coral Reefs, 19(4): 330–331. 440
6. Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, and Stahl DA (1990). 441
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

20
Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for 442
analysing mixed microbial populations. Appl. Environ. Micro. 56:1919–1925 443
7. Amann RI, Ludwig W, and Schleifer KH (1995). Phylogenetic identification and in 444
situ detection of individual microbial cells without cultivation. Microbiol. Rev. 59:143-445
169 446
8. Amman R, Snaidr J, Wagner M, Ludwig W, and Scleifer KH (1996). In situ 447
visualisation of high genetic diversity in a natural microbial community. J. Bacteriol. 178: 448
3496 – 3500. 449
9. Amann R, Fuchs BM, and Behrens S (2001). The identification of micro-organisms by 450
fluorescence in situ hybridisation. Curr. Opin. Biotech. 12: 231- 236. 451
10. Antonius A (1973). New observations on coral destruction in reefs. Abs. Assoc. Isl. 452
Mar. Lab. Caribb.10:3. 453
11. Antonius A (1977). Coral mortality in reefs: a problem for science and management. 454
Proc 3rd
Int Coral Reef Symp. Miami 2: 617–623. 455
12. Antonius A (1981). The ‘band’ diseases in coral reefs. Proc 4th
Int Coral Reef Symp. 456
Philippines 2: 7–14. 457
13. Antonius A and Riegl B (1997). A possible link between coral diseases and a 458
corallivorous snail (Drupella cornus) outbreak in the Red Sea. Atoll. Res. Bull. 447: 1–9. 459
14. Antonius A and Riegl B (1998). Coral diseases and Drupella cornus invasion in the 460
Red Sea. Coral Reefs 17: 48. 461
15. Aronson RB and Precht WF (2001). White-band diseases and the changing face of 462
Caribbean coral reefs. Hydrobiol. 460:25–38. 463
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

21
16. Banin E, Israely T, Fine M, Loya Y, and Rosenberg E (2001). Role of endosymbiotic 464
zooxanthellae and coral mucus in the adhesion of the coral-bleaching pathogen Vibrio 465
shiloi to its host. FEMS Micro. Lett. 199:33–37 466
17. Barash Y, Sulam R, Loya Y, and Rosenberg E (2005). Bacterial strain BA-3 and a 467
filterable factor cause a white plague-like disease in corals from the Eilat coral reef. 468
Aquat. Micro. Ecol. 40:183-189 469
18. Ben-Haim Y, and Rosenberg E (2002). A novel Vibrio sp pathogen of the coral 470
Pocillopora damicornis. Mar. Biol. 141:47–55 471
19. Breitbart M, Bhagooli R, Griffin S, Johnston I, and Rowher F (2005). Microbial 472
communities associated with skeletal tumors on Porites compressa. FEMS Micro. lett. 473
Online release. 474
20. Bythell JC, Barer MR, Cooney RP, Guest JR, O’Donnell AG, Pantos O, Le Tissier 475
MDA (2002). Histopathological methods for the investigation of microbial communities 476
associated with disease lesions in reef corals. Lett. Appl. Micro. 34: 359–364. 477
21. Bythell JC, Pantos O, and Richardson L, (2004). White plague, White Band and 478
other “White” Diseases. In Coral health and Disease. (ed. E. Rosenberg and Y. Loya) 479
Springer-Verlag, Germany. 480
22. Carlton R, and Richardson LL (1995). Oxygen and sulphide dynamics in a 481
horizontally migrating cyanobacterial mat: black band disease of corals. FEMS Micro. 482
Ecol. 18:155-162 483
23. Daims H, Brühl A, Amann R, Schleifer KH, and Wagner M (1999). The domain-484
specific probe EUB338 is insufficient for the detection of all Bacteria: development and 485
evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 22:434-444. 486
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

22
24. Denner EBM, Smith G, Busse HJ, Schumann P, Narzt T, Polson SW, Lubitz W, and 487
Richardson LL (2003). Aurantimonas coralicida gen. nov., sp. nov., the causative agent 488
of white plague type II on Caribbean scleractinian corals. Int. J .Syst. Evol. Microbiol. 489
53:1115–1122. 490
25. Dunn SR, Bythell JC, Le Tissier MDA, Burnett WJ, and Thomason JC (2002). 491
Programmed cell death and cell necrosis activity during hyperthermic stress induced 492
bleaching of the symbiotic sea anemone Aiptasia sp. J. Exp. Mar. Biol. Ecol. 272: 29–53. 493
26. Dunn SR, Thomason JC, Le Tissier MDA, and Bythell JC (2004). Heat stress 494
induces different forms of cell death in sea anemones and their endosymbiotic algae 495
depending on temperature and duration. Cell death and Diff. (2004): 1-10. 496
27. Frias-lopez J, Zerkle AL, Bonheyo GT, and Fouke BW (2002). Partitioning of 497
bacterial communities between seawater and healthy, black band diseased, and dead coral 498
surface. Appl. Env. Micro. 68(5): 2214 – 2228. 499
28. Fine M, Oren U, and Loya Y (2002). Bleaching effect on regeneration and resource 500
translocation in the coral Oculina patagonica. MEPS 234:119 -125 501
29. Green EP, and Bruckner AW (2000). The significance of coral disease epizootiology 502
for coral reef conservation. Biol. Conserv. 96: 347–361. 503
30. Harvell CD, Kim K, Burkholder JM, Colwell RR, Epstein PR, Grimes EE, et al. 504
(1999) Emerging marine diseases-climate links and anthropogenic factors. Science 285: 1 505
505–1510. 506
31. Harvell CD, Mitchell CE, Ward JR, Altizer S, Dobson AP, Ostfeld RS, and Samuel 507
MD (2002). Climate Warming and disease risks for terrestrial and marine biota. Science 508
296: 2158 – 2162. 509
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
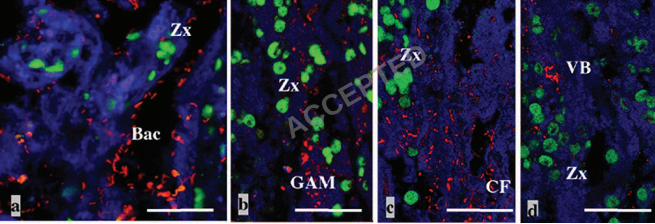
Dow
nloaded from

23
32. Kim K, Harvell CD, Kim PD, Smith GW, and Merkel SM (2000a). Fungal disease 510
resistance of Caribbean sea fan corals (Gorgonia spp). Mar. Biol. 36: 259–267. 511
33. Kim K, Kim PD, Alker AP, and Harvell CD (2000b). Chemical resistance of 512
gorgonian corals against fungal infections. Mar. Biol. 137: 393–401. 513
34. Kushmaro A, Loya Y, Fine M, and Rosenberg E (1996). Bacterial infection and 514
coral bleaching. Nature 380: 396 515
35. Kushmaro A, Rosenberg E, Fine M, Ben-Haim Y, and Loya Y (1998). Effect of 516
temperature on bleaching of the coral Oculina patagonica by Vibrio shiloi AK-1. MEPS 517
171: 131–137. 518
36. Kushmaro A, Banin E, Loya Y, Stackebrandt E, and Rosenberg E (2001). Vibrio 519
shiloi sp nov the causative agent of bleaching of the coral Oculina patagonica. Int. J. 520
Syst. Evol. Micro. 51: 1383–1388. 521
37. Kuta KG, and Richardson LL (1996). Abundance and distribution of black band 522
disease of corals in the northern Florida Keys. Proc 8th
Int Coral Reef Symp. 15:219 – 523
223. 524
38. Lesser MP (2004). Experimental biology of coral reef ecosystems. J. Exp. Mar. Bio. 525
Ecol. 300: 217 – 252. 526
39. Loya Y, and Kramarsky-Winter E (2003). In situ eutrophication caused by fish farms 527
in the northern Gulf of Eilat (Aqaba) is beneficial for its reefs: a critique. MEPS 261:299 528
- 303 529
40. Loya Y. (2004). The Coral Reefs of Eilat – Past, Present and Future: Three Decades 530
of Coral Community Structure Studies. In Coral health and Disease. (ed. E. Rosenberg 531
and Y. Loya) Springer-Verlag, Germany. 532
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

24
41. Manz W, Amann R, Ludwig W, Wagner M, and Schleifer KH (1992). Phylogenetic 533
oligonucleotide probes for the major subclasses of proteobacteria: problems and 534
solutions. Syst. Appl. Micro. 15: 593-600 535
42. Manz W, Arp G, Schumann-Kindel G, Szemzyke U, and Reitner J (2000). 536
Widefield deconvolution epifluorescence microscopy combined with fluorescence in situ 537
hybridisation reveals the spatial arrangement of bacteria in sponge tissue. J. Micro. Meth. 538
40:125-134. 539
43. Moreno Y, Arias CR, Meier H, Garay E, and Aznar R (1999). In situ analysis of the 540
bacterial communities associated to farmed eel by whole-cell hydridisation. Lett Appl 541
Micro 29:160-165. 542
44. Mullen KM, Peters EC, and Harvell CD (2004). Coral Resistance to disease. In Coral 543
health and Disease. (ed. E. Rosenberg and Y. Loya) Springer-Verlag, Germany. 544
45. Pantos O, and Bythell JC (2006). Bacterial community structure associated with 545
white band disease in the elkhorn coral Acropora palmata determined using culture-546
independent 16S rRNA techniques. Dis. Aquat. Org. 69:79 – 88 547
46. Patterson KL, Porter JW, Ritchie KB, Polson SW, Mueller E, Peters EC, Santavy 548
DL, Smith GW(2002). The eitiology of white pox a lethal disease of the Caribbean 549
Elkhorn coral Acropora palmata. Proc Natl Acad Sci USA 99:8725-8730. 550
47. Peters EC (1984). A survey of cellular reactions to environmental stress and disease 551
in Caribbean scleractinian corals. Helgol Meeresunters 37: 113–137. 552
48. Peters EC, Oprandy JJ, and Yevich PP (1983). Possible causal agent of “white band 553
disease” in Caribbean acroporids corals. J. Invert. Pathol. 41: 394-396. 554
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

25
49. Porter JW, Dunstan P, Jaap WC, Patterson KL, Kosmynin Y, Meir OW, Patterson 555
ME, Parsons M (2001). Patterns of spread of coral disease in the Florida Keys. 556
Hydrobiologia 460: 1 – 24. 557
50. Richardson LL, Goldberg WM, Carlton RG, and Halas JC (1998a). Coral disease 558
outbreak in the Florida Keys: plague type II. Rev. Biol. Trop. 46:187–198. 559
51. Richardson LL, Goldberg WM, Kuta KG, Aronson RB, Smith GW, Ritchie KB, 560
Halas JC et al. (1998b). Florida’s mystery coral killer identified. Nature 392: 557–558. 561
52. Richardson LL, Smith GW, Ritchie KB, and Carlton RG (2001). Integrating 562
microbiological microsensor molecular and physiologic techniques in the study of coral 563
disease pathogenesis. Hydrobiol. 460: 71–89. 564
53. Richardson LL, and Kuta KG (2003). Ecological physiology of the black band 565
disease cyanobacterium Phormidium corallyticum. FEMS Micro. Ecol, 43: 287 – 298 566
54. Richardson LL (2004). Black Band Disease. In Coral health and Disease. (ed. E. 567
Rosenberg and Y. Loya) Springer-Verlag, Germany. 568
55. Ritchie KB, and Smith GW (1997). Physiological comparison of Bacterial 569
communities from various species of scleractinian corals. Proceeding from 8th
570
International Coral Reef Symp. 1: 521-526. 571
56. Roff J (2004). Tabular Acropora syndrome on the Great Barrier Reef. Honours 572
Thesis University of Queensland, Australia. 573
57. Roff G, Hoegh-Guldberg O, and Fine M (2006). Intra-colonial response to Acroporid 574
“white syndrome” lesions in tabular Acropora app. (Scleractinia). Coral Reefs: Online 575
First 576
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

26
58. Roller C, Wagner M, Amann R, Ludwig W, and Schleifer KH (1994). In situ probing 577
of gram positive bacteria with hight G+C content using 23S rRNA-targeted 578
oligonucleotides. Micro. 140; 2849 – 2858. 579
59. Rosenberg E, and Ben-Haim Y (2002). Microbial diseases of corals and global 580
warming. Environ. Micro. 4(6): 318 – 326. 581
60. Rowher F, Breitbart M, Jara J, Azam F, and Knowlton N (2001). Diversity of 582
bacteria associated with the Caribbean coral Montastrea franksi. Coral Reefs 20: 85-95. 583
61. Rowher F, Seguritan V, Azam F, and Knowlton N (2002). Diversity and distribution 584
of coral-associated bacteria. MEPS. 243:1-10 585
62. Stanier RY, Doudoroff M, and Adelberg EA (1987). General Microbiology. 586
Macmillian, London (1971). 587
63. St John JA, Tisay KT, Caras IW, and Key B (2000). Expression of EphA5 during 588
development of the olfactory nerve pathway in rat. J. Comp. Neuor. 416:540-550. 589
64. Sutherland KP, Porter JW, and Torres C (2004). Disease and immunity in Caribbean 590
and Indo-pacific zooxanthellate corals. MEPS 266: 273 – 302. 591
65. Weil E., Smith G., and Gil-Agudelo DL (2006). Status and progress in coral reef 592
disease research. Dis. Aquat. Org. 69: 1-7 593
66. Wielgus J. (2003) The coral reef of Eilat (northern Red Sea) requires immediate 594
protection. MEPS 263:307. 595
67. Winkler R, Antonius A, and Renegar DA (2004). The skeleton Eroding Band 596
Disease on Coral Reefs of Aqaba, Red Sea. Mar. Ecol. 25(2): 129 –144. 597
598
599
600
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

27
Figure legends 601
602
Figure 1. Typical (A,a) and atypical black band disease (B,b) of Eilat, The Red Sea. 603
Underwater photographs of infected black band diseased colonies displaying typical 604
(A1,a1) and atypical (B1,b1) signs of disease. Tissue necrosis and microbial penetration 605
and disruption of tissue layers in black band diseased colonies evident by histopathology, 606
using Haematoxylin and Eosin staining of typical (A2,a2) and atypical (B2,b2) diseased 607
colonies, and FISH of typical (A3,a3) and atypical (B3,b3) diseased colonies. Scale 608
50um. Polyp (Po), Epithelium (Ep), Gastroderm (Ga), necrosis (Ne), Bacteria (bac). FISH 609
and spectral codes: Coral tissue (blue), Symbiodinium sp. (green), bacteria (red). 610
Uppercase labels demonstrates lower magnification images, lowercase denotes higher 611
magnification. 612
613
614
615
616
617
618
619
620
621
622
623
624
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

28
625
Figure 2. Diseased Hydnophora sp. (a) with evident rapid tissue loss and clear lesion 626
borders between tissue and recently exposed skeleton (b), histopathology showed extensive 627
tissue necrosis (Ne) of cell layers (c) including epithelium, gastroderm (d), FISH revealed 628
extensive microbial population using the EUB universal bacterial probe(e, f, g) associated 629
with tissue necrosis and adjacent tissues. Scale 50 µm. Epithelium (Ep), Gastroderm (Ga), 630
Symbiodinium sp. (Zx), tissue necrosis (Ne), Bacteria (Bac). FISH and spectral color 631
codes: coral tissue (Blue), Symbiodinium sp. (Green), Bacterial (Red). Uppercase label 632
demonstrates lower magnification images, lowercase denotes higher magnification. 633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

29
Figure 3. FISH identified bacterial communities associated with diseased Hydnophora sp. 649
identified with EUBmix (a), as belonging to δ-proteobacteria (b) and a filamentous 650
Cytophaga-Flavobacterium (c), populations of a Vibrio species were also identified (d). 651
Scale 50 µm. Epithelium (Ep), Gastroderm (Ga), Symbiodinium sp. (Zx), tissue necrosis 652
(Ne), Bacteria (Bac), δ-proteobacteria (GAM), Cytophaga-Flavobacterium (CF) and Vibrio 653
sp (VB). FISH and spectral color codes: coral tissue (Blue), Symbiodinium sp. (Green), 654
Bacterial (Red). Uppercase label demonstrates lower magnification images, lowercase 655
denotes higher magnification. 656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
j ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

30
Figure 4. White disease of Stylophora pistillata displaying tissue loss and partial bleaching 673
in pattern starting at the branch base and progressing rapidly up the branches and through 674
the colony (A1, a1), with 50% mortality of the colony is evident (A). Haematoxylin and 675
eosin staining shows tissue structures remain intact at the lesion border (A2,a2) with some 676
regions of structural loss of the gastroderm adjacent to the lesions (aa2) and FISH using the 677
EUB general bacterial probe identified large populations of bacteria associated with tissues 678
adjacent to the lesion border (A3), a mixed bacterial population dominated by δ-679
proteobacterial taxa (a3). Scale 50um. Epithelium (Ep), Gastroderm (Ga), Symbiodinium 680
sp. (Zx), tissue necrosis (Ne). FISH and spectral color codes: coral tissue (Blue), 681
Symbiodinium sp. (Green), Bacterial (Red). Uppercase label demonstrates lower 682
magnification images, lowercase denotes higher magnification. 683
684
685
686
687
688
689
690
691
692
693
694
695
e
j ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

31
Figure 5. White syndrome of Acropora sp. (a), evidence of clear lesion boundary between 696
healthy tissue and recently exposed skeleton (b), investigation of tissue structure using 697
haematoxylin and Eosin appear (c,d), a lack of microbial communities associated with the 698
tissues other than dense bacterial aggregates within gastrodermal tissue layers (e,f). Scale 699
50 µm. (Ep), Gastroderm (Ga), Symbiodinium sp. (Zx), tissue necrosis (Ne), Bacteria 700
(Bac),). FISH and spectral color codes: coral tissue (Blue), Symbiodinium sp. (Green), 701
Bacteria (Red). Uppercase label demonstrates lower magnification images, lowercase 702
denotes higher magnification. 703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
j ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

32
Figure 6. In situ end labelling of fragmented DNA evident of apoptotic cell death is not 719
detected in Black band disease (atypical) (a), typical (b,c), white diseases of massive’s 720
(d,e,f), positive staining was evident at the lesion border in the white disease of 721
Stylophora pistillata (g,h), but not of tissues away from border (i) and a high density of 722
apoptotic cells associated with white syndrome (k,l), but not away from lesion border 723
(m). Uppercase label demonstrates lower magnification images, lowercase denotes higher 724
magnification. 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

33
Figure 7. Profiling the interaction of holobiont changes and multiple levels of diseaase 743
signs can be used as a basis for identifying specific criteria for disease diagnosis and to 744
improve understanding of divergent disease processes underlieng coral disease 745
progession. 746
747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
763
764
765
766
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

34
Tables 767
Table 1. Bacterial Probes used in FISH. 768
Probe Target group Sequence (5’ – 3’) formamide From
EUBmix Universal
bacterial probe
GCTGCCTCCCGTAGGAGT
GCAGCCACCCGTAGGTGT
GCTGCCACCCGTAGGTGT
35% 6
23
BET42A β-proteobacteria GCCTTCCCACTTAGTTT 35% 41
GAM42A γ-proteobacteria GCCTTCCCACATCGTTT 35% 41
CF319 Cytophaga-
Flavobacterium
TGGTCCGTGTCTCAGTAC 35% 42
MV Vibrio spp. ACAGTACTCTAGTCTCGCCAG 35% 40
769
770
771
772
773
774
775
776
777
778
779
780
781
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

35
782
Table 2. Summary of microbial and disease characteristics of coral diseases in Eilat*. 783
Black band disease
typical / atypical
White disease
(massive)
White disease
(branching)
White syndrome
(branching)
Bacterial
infiltration of
tissue layers
+ ve / +ve + ve + ve - ve
Mass tissue
necrosis
+ ve / +ve + ve - ve
(small region)
- ve
Programmed
cell death
- ve / -ve - ve + ve
(only adjacent
lesion border)
+ ve
*Presence of the characteristic indicated by +ve, a lack of the characteristic indicated by –784
ve. 785
786
787
788
789
790
791
792
793
794
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from






Example 3:
Black band disease
1. Black microbial
growth at lesion border
(typical or atypical)
2. Black microbial
penetration of tissue
layers
3. Mass tissue necrosis
Example 2:
White disease 1. Clear lesion boundary
2. Exposed skeleton
3. Extensive microbial
involvement
4. No evidence for black
band consortium
5. Mass tissue necrosis
5.
Example 1:
White syndrome
1. Clear lesion boundary
2. Exposed skeleton
3. Lack of microbial
involvement in disease
progression
5. Evidence for extensive
apoptosis
Physiological changes
Scale
Microbial isolation and in situ detection/identification
• Presence /absence of microbial interaction & and extent of
microbial colonization?
Cytological disease indications?
• Mass necrosis / loss of structure or programmed cell death?
• Changes in respiration /photosynthesis / translocation?
• Microbial communities & community shifts
• Ability to determine pathogenic capacity?
Reefal observations & macroscopic indications of disease
• Ecological and colonial patterns?
• Tissue loss / exposed skeleton or bleaching /symbiont loss?
ACCEPTED
on June 15, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from