control of gluconeogenesis in liver · the journal of bmm~ca~ chemistry vol. 243, no. 16, issue of...
TRANSCRIPT

THE JOURNAL OF Bmm~ca~ CHEMISTRY Vol. 243, No. 16, Issue of August 25, pp. 4189-4196, 1968
P&ted in U.S.A.
Control of Gluconeogenesis in Liver
II. EFFECTS OF GLUCAGON, CATECHOLAMINES, AND ADENOSINE 3’) 5’-MONOPHOSPHATE ON GLUCONEOGENESIS IN THE PERFUSED RAT LIVER*
(Received for publication, April 1, 1968)
J. H. EXTON~ AND C. R. PARK
From the Department of Physiology, Vanderbilt University, Nashville, Tennessee 37203
SUMMARY
Glucagon, epinephrine, norepinephrine, and cyclic AMP stimulated gluconeogenesis from lactate in perfused livers from fasted and fed rats. The stimulation was most marked when these agents were added to livers from fasted rats in which gluconeogenesis had become partially inactive during a preliminary perfusion without substrate. It is suggested that inactivation is consequent to the degradation of endog- enous glucagon and catecholamines. Inactivation could be prevented by high levels of lactate.
On a molar basis, glucagon was about lOO-fold more effective in stimulating gluconeogenesis than the catechol- amines, and 100,000 times more effective than exogenous cyclic AMP. Gluconeogenesis was as sensitive as glyco- genolysis to stimulation by any of the agents and was as rapidly activated. It is proposed that cyclic AMP mediates the action of glucagon and catecholamines on gluconeo- genesis.
Glucagon stimulated gluconeogenesis from lactate and pyruvate but not from dihydroxyacetone or fructose, sug- gesting a point of action in the gluconeogenic pathway be- tween pyruvate and triose phosphate. The maximum rate of gluconeogenesis from oxalacetate was about the same as that from pyruvate. Because lactate production from oxal- acetate was substantial and the same tissue level of malate was found with pyruvate or oxalacetate, it appeared that oxalacetate was not metabolized as such by the liver but only after breakdown to pyruvate. Ethanol stimulated gluconeogenesis and lactate formation from pyruvate. The effects of ethanol and glucagon were additive indicating that glucagon does not act by increasing NADH supply. The gluconeogenic effect of glucagon was observed with con- centrations of lactate in the physiological range.
It is proposed that glucagon plays a physiological role in the maintenance of the blood glucose through control of
* This investigation was supported by program project Grant AM-07462 from the National Institutes of Health. United States Public Health Service. Preliminary reports and’an abstract of part of this work have been published-(References 1 and 2 and EXTON, J. H., AND PARK, C. R., Fed. Proc., 24, 537 (1965)).
$ Investigator of the Howard Hughes Medical Institute.
gluconeogenesis as well as glycogenolysis in the liver. Sym- pathetic nervous activity, leading to the lntrahepatic release of catecholamine, may also play an important role in in- creasing glucose output and lactate uptake by the liver, particularly during exercise or hypoglycemia.
A number of observations in the intact animal have suggested that glucagon stimulates gluconeogenesis. It increases urinary nitrogen excretion and urea production in rats (3-7) and stimu- lates hepatic uptake of amino acids in dogs (8). It increases the incorporation of 1°C from labeled glycine into liver glycogen after an initial period of glycogen depletion (9) and lowers the con- centration of amino acids in the blood (6, 7, 10).
More direct evidence has been obtained in studies with the perfused liver. Miller (11, 12) and others (13-15) have found that glucagon increases hepatic urea production. Schimassek and Mitekat (16) have shown that it stimulates lactate utiliaa- tion and, concomitantly, the formation of glucose. Garcia, Williamson, and Cahill (17) and Exton et al. (1, 2) have found an increased incorporation of isotope from 14C-labeled lactate, pyruvate, alanine, or bicarbonate into glucose. Struck, Ash- more, and Wieland (18) have reported an accelerated conversion of lactate to glucose and have suggested that the hormone effect on gluconeogenesis is the result of lipase activation. William- son et al. (19-21) have presented data in support of this hypothe- SiS.
The action of catecholamines on gluconeogenesis has not been extensively studied. Experiments in whole animals have yielded conflicting results (5, 7). Miller (22) has reported that doses of epinephrine sufficient to produce maximum glycogenolysis did not affect urea production in the perfused liver. On the other hand, Schimassek and Mitzkat mentioned that epinephrine increased gluconeogenesis from lactate in the perfused liver (16). Exton et al. (1, 2) verified this and presented evidence that the effects of catecholamines and glucagon were mediated by adeno- sine 3’ , 5’-monophosphate (cyclic AMP).
In this study, further investigations have been undertaken of
4189
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4190
400 -
2 3
z 0
In 300- w- 0” - s Y z a 200-
Control of Gluconeogenesis in Liver. II
T 4oor
A T = I S.E.M.
/
LACTATE T FROM OMIN
LACTATE t
T = I S.E.M.
Vol. 243, No. 16
B
LACTATE t CYCLIC AMP
J
AT 60 MIN.
LACTATE t EPINEPHRINE AT 60 MIN. ! ,j+; ! o~tg
0 20 40 60 80 100 120 0 20 40 60 80 100 120
MIN. OF PERFUSION MIN. OF PERFUSION
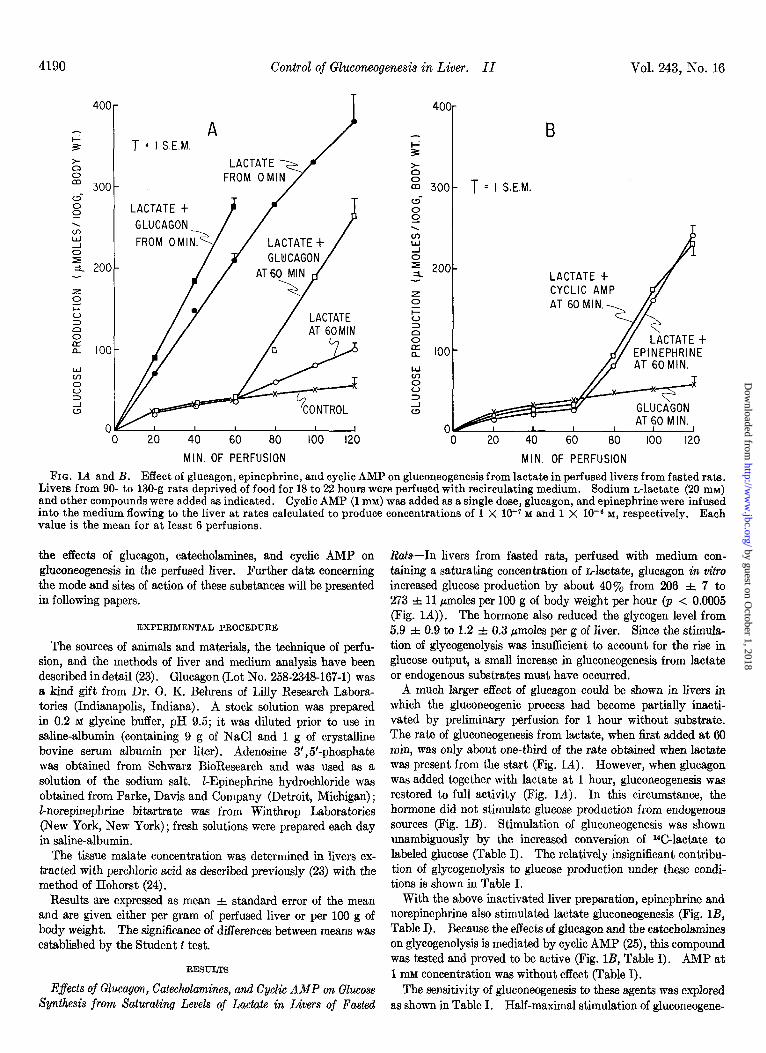
FIG. 1A and B. Effect of glucagon, epinephrine, and cyclic AMP on gluconeogenesis from lactate in perfused livers from fasted rats. Livers from 90- to 130-g rats deprived of food for 18 to 22 hours were perfused with recirculating medium. Sodium n-lactate (20 mru) and other compounds were added as indicated. Cyclic AMP (1 mM) was added as a single dose, glucagon, and epinephrine were infused into the medium flowing to the liver at rates calculated to produce concentrations of 1 X 10-r M and 1 X 10e6 M, respectively. Each value is the mean for at least 6 perfusions.
the effects of glucagon, catecholamines, and cyclic AMP on R&--In livers from fasted rats, perfused with medium con- gluconeogenesis in the perfused liver. Further data concerning taining a saturating concentration of nlactate, glucagon in vitro the mode and sites of action of these substances will be presented increased glucose production by about 40% from 206 =t 7 to in following papers. 273 f 11 pmoles per 100 g of body weight per hour (p < 0.0005
(Fig. 1A)). The hormone also reduced the glycogen level from EXPERTMENTAL PROCEDURE 5.9 f 0.9 to 1.2 f 0.3 pmoles per g of liver. Since the stimula-
The sources of animals and materials, the technique of perfu- tion of glycogenolysis was insufhcient to account for the rise in sion, and the methods of liver and medium analysis have been glucose output, a small increase in gluconeogenesis from lactate
described in detail (23). Glucagon (Lot No. 258-2348-167-1) was or endogenous substrates must have occurred. a kind gift from Dr. 0. K. Behrens of Lilly Research Labora- A much larger effect of glucagon could be shown in livers in tories (Indianapolis, Indiana). A stock solution was prepared which the gluconeogenic process had become partially inacti-
in 0.2 M glycine buffer, pH 9.5; it was diluted prior to use in vated by preliminary perfusion for 1 hour without substrate. saline-albumin (containing 9 g of NaCl and 1 g of crystalline The rate of gluconeogenesis from lactate, when 6rst added at 60 bovine serum albumin per liter). Adenosine 3’,5’-phosphate min, was only about one-third of the rate obtained when lactate
was obtained from Schwarz BioResearch and was used as a was present from the start (Fig. IA). However, when glucagon solution of the sodium salt. Z-Epinephrine hydrochloride was was added together with lactate at 1 hour, gluconeogenesis was
obtained from Parke, Davis and Company (Detroit, Michigan) ; restored to full activity (Fig. 1A). In this circumstance, the
E-norepinephrine bitartrate was from Winthrop Laboratories hormone did not stimulate glucose production from endogenous (New York, New York) ; fresh solutions were prepared each day sources (Fig. 1B). Stimulation of gluconeogenesis was shown in saline-albumin. unambiguously by the increased conversion of “C-lactate to
The tissue malate concentration was determined in livers ex- labeled glucose (Table I). The relatively insignificant contribu-
tracted with perchloric acid as described previously (23) with the tion of glycogenolysis to glucose production under these condi-
method of Hohorst (24). tions is shown in Table I. Results are expressed as mean f standard error of the mean With the above inactivated liver preparation, epinephrine and
and are given either per gram of perfused liver or per 100 g of norepinephrine also stimulated lactate gluconeogenesis (Fig. lB,
body weight. The significance of differences between means was Table I), Because the effects of glucagon and the catecholamines established by the Student 1 test. on glycogenolysis is mediated by cyclic AMP (25)) this compound
was tested and proved to be active (Fig. lB, Table I). AMP at RESULTS 1 mM concentration was without effect (Table I).
Effects of Glucagon, Catecholamines, and Cyclic AMP on Glucose The sensitivity of gluconeogenesis to these agents was explored synthesis from Saturating Levels of Lactate in Livers of Fasted as shown in Table I. Half-maximal stimulation of gluconeogene-
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of August 25, 1968 J. H. Exton and C. R. Park 4191
sis was observed with approximately 2 x lo-‘* M glucagon, 2 X lo-* M epinephrine, 1 X lop8 M norepinephrine, and 2 X 10e5 M cyclic AMP.
Factors Affeciing Innaetivation of Gluconeogenesis-Some con- sideration was given to factors which might affect the inactivation of gluconeogenesis during the hour of perfusion without substrate. It was apparent that lactate itself largely prevented inactivat,ion since the rate of glucose production was well maintained during the second hour when lactate was present from the beginning (Fig. 1A).
Gluconeogenesis was reactivated if livers perfused for 1 hour without substrate were then perfused with fresh medium (Table II, Experiment C). The responsible factor in fresh medium has not been characterized but was associated with certain prepara- tions of bovine Fraction V albumin (Experiment D). It was not present in crystalline bovine albumin (Experiment E). The presence of a stimulatory factor in certain preparations of crude albumin had been shown in earlier studies (23). Although re- moval or destruction of this factor by the liver might be involved in the inactivation process, changes in the liver itself also play a significant role inasmuch as a high rate of gluconeogenesis was
TABLE I
Effect of glucagon, epinephrine, norepinephrine, cyclic AMP, and AMP on glucose synthesis from lactate
Livers from rats (90 to 130 g) fasted for 18 to 22 hours were per- fused for 1 hour without substrate. Sodium n-lactate (20 mM),
sodium nn-2-‘e-lactate (10,500 cpm per ml), and the other com- pounds indicated in the table were then added, and the perfusion was continued for 1 hour. Cyclic AMP and AMP were added as single doses; the hormones were infused into the medium flowing to the liver at rates calculated to produce the designated con- centrations. The values for glucose and “C-glucose production are for the second hour.
Additions and concentration
M
None Glucagon
1 x 10-1 1 x 10-g 2 x lo-‘0 1 x lo-10
Epinephrine 1 x 10-S 1 x 10-T 2 x IO-* 1 x 10-a
Norepinephrine 2 x lo-7 5 x lo-8 1 x 10-S 5 x HP
Cyclic AMP 1 x 10-s 1 x 10-d 2 x lo-6 1 x 10-6
5’-AMP 1 x 10-a
-
-
zs*
11
4
Glucose production
pmoles/g liver/ hr
18 f 2
!Bf5 55 f 4 33 f 3 21 f 2
59 f 3 56 i 4 34 f 3 29f2
57 f 4 58f4 35 f 3 26, 28
55 f 4 56 i 5 34 f 6 22, 31
23 f 2
-
I
pwles/g live?
2.4 f 0.6”
0.9 f 0.1
1.1 i 0.5
1.2 f 0.3
3.7 f 0.2
IT-Glucose synthesis
cm x 1lwg liw/hr
12.5 f 1.3
30.9 i 1.8
29.8 f 1.4
26.6 f 4.0
31.1 f 5.4
TABLE II Factors affecting inactivation of gluconeogenesis from lactate As noted in the table, livers from fasted rats were either pre-
pared for the test period by a preliminary perfusion for 1 hour with medium containing no substrate or were used fresh, i.e. without prior perfusion. The test medium had either been perfused for 1 hour through the liver of a fasted rat or was freshly prepared. To it was added 20 mu n-lactate. Where indicated, albumin was added as 5 ml of a solution containing 20 mg per ml to give a final concentration of about 2%. The duration of the test period of perfusion was 60 min.
-
Experi- I ment
-
11 Perfused 10 Fresh 6 Perfused 8 Perfused
4 Perfused
7 Fresh
-
-
Typeof medium GhlCXe production
“y$p
Perfused 18 f 2 Fresh 55f2 Fresh 31 f 2 Perfused, Fraction V albumin 33 f 4
added Perfused, crystalline albumin 15 f 3
added Perfused 41 * 3
observed in fresh livers perfused with medium which had pre- viously been perfused through other livers for 1 hour (Experi- ment F) .
It seemed possible that the low rate of gluconeogenesis seen when lactate was added after perfusion for 1 hour might also be due to a fall in pH. However, in agreement with Hems et al. (26), it was found that glucose production from lactate was unaltered by pH changes over the range 7.2 to 7.7. Although the pH of the medium fell in some instances to below 7.2 when livers were perfused for 1 hour without substrate, adjustment of the pH to 7.4 with NaHC03 or NaOH did not increase the rate of glucose formation from lactate over the next hour.
Efects of Glumgon on Glumneogenesis from 0th.m Substrates- Ross, Hems, and Krebs (27) have reported that in the perfused rat liver, glucagon stimulates gluconeogenesis from lactate and pyruvate but not from alanine, oxalacetate, glycerol, fructose, and dihydroxyacetone. However, Garcia et al. (17) have stated that the hormone increases glucose synthesis from all glucogenic substrates studied. Table III shows the effects of 1 x lo--* M
glucagon on gluconeogenesis from various substrates in liver perfused initially for 1 hour without substrate. As was the case with lactate, formation of glucose plus glycogen from pyruvate or oxalacetate, added at 1 hour, was lower than that observed when these substrates were added at the start of the perfusion. Whereas glucagon largely restored gluconeogenesis from pyru- vate, it only partly restored that from oxalacetate (Table III). Since it seemed possible that gluconeogenesis from these sub- strates was limited by the supply of cytoplasmic NADH, the effects of ethanol were examined. This hydrogen donor stimu- lated gluconeogenesis from pyruvate and oxalacetate added at the start of the perfusion and was more effective than glucagon in restoring gluconeogenesis when added at 1 hour (Table III). The stimulatory effect of glucagon on gluconeogenesis from pyruvate or oxalacetate appeared to be additive with that of ethanol (Table III). With fructose and dihydroxyacetone as substrates, the rate of gluconeogenesis was not reduced by the initial perfusion and glucagon had no effect on glucose production other than to stimulate glycogenolysis (Table III). The effect
0 The glycogen level before addition of lactate was 1 pmole per g of liver.
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4192 Control of Gluconeogenesis in Liver. II Vol. 243, No. 16
TABLE III
Effects of glucagon on gluconeogenesis from pyruvate, oxalacetate, dihydroxyacetone, and fructose
Livers from fasted rats were either perfused for 1 hour without substrate or used fresh, i.e. without prior perfusion. The test period was 1 hour and the substrate concentration was 20 mM. Where indicated, ethanol was added to give a 20 mM concentration and glucagon was infused into the medium flowing to the liver at a rate of 0.5 pg per min.
Substrate and condition of live
Pyruvate Fresh. Fresh. . . Perfused.. Perfused.. Perfused.. Perfused..
Oxalacetate Fresh. . . Fresh. . Perfused.. Perfused.. Perfused.. Perfused..
Dihydroxy- acetone
Fresh . . Perfused. Perfused.
Fructose Fresh Perfused Perfused
-
rE
--
thall01
-
+ -
+ -
+
-
+ -
+ -
+
- -
-
-
- -
Glu- cagon
- -
- -
+
+
-
- - -
+ +
- -
+
-
-
+
\To. of xperi- nents
Carbohydrate synthesis
Glucose Glycogen Total
,mmles/g liver/fir
6 50 f 2 5.2 f 1.7 55 i 2 5 69 f 1 11.2 i 0.3 80 f 1 8 28 f 3 7.6 f 0.5 36 f 3 4 40 f 3 15.2 f 0.9 55 f 3 8 46 f 3 0.9 f 0.2 47 f 3 4 62 f 2 1.0 f 0.1 63 f 2
10 46 f 2 3.8 f 0.5 50 f 1 5 59 f 3 4.6 f 0.5 64 i 2
10 17 f 2 4.5 f 0.5 22 f 1 4 33 f 1 7.5 f 1.4 41 f 1
10 28 f 1 0.9 f 0.3 29 f 1 4 51 f 1 2.1 f 0.5 53 f 1
6 4 4
11 4 4
06 f 9 19 f 3 125 f 5 .Ol f 10 20 f 3 121 f 5 .22 f 10 2 f 1 124 f 6
.06 f 3 16 f 2 122 f 3 92 f 4 22f2 114f3 111 f 8 3fl 114f5
TABLE IV Lactate production and tissue malate levels in livers perfused with
oxalacetate and pyruvate
Livers from fasted rats were perfused for 1 hour without sub- strate; then 20 mM pyruvate or oxalacetate was added with or without 20 mM ethanol, and perfusion was continued for 1 hour more. Lactate production was measured over the second hour, and the malate level was determined in tissue rapidly frozen at the end of the perfusion.
-
Substrate EthaIl01 Lactate production Tissue malate level
wdes/g liver/fir nmoles/g Of liver, dry wt
Pyruvate Absent 71 f 11 0.89 i 0.13 Pyruvate Present 129 f 12 1.91 f 0.08 Oxalacetate......... Absent 12 & 3 0.94 * 0.11 Oxalacetate......... Present 58 f 10 1.58 f 0.13
of glucagon on gluconeogenesis from alanine or glycerol was not tested under these conditions.
Lactate and Malate Formation in Livers Perfused with Oxalace- tate or Pyruvale-In agreement with the observations of Ross et al. (27)) the maximum rate of gluconeogenesis from oxalacetate was approximately the same as that from pyruvate (Table III). This is in striking contrast to other di- and tricarboxylic acids which are ineffective as glucose precursors (23, 27). I f oxalace-
tate were entering the liver cell freely as such, the results of Table III would indicate that the reaction limiting maximum gluconeogenesis from lactate or pyruvate and the site of stimula- tion by cyclic AMP lie beyond oxalacetate in the glucose syn- thetic pathway. However, since oxalacetate is a relatively unstable compound, it was important to determine whether it was being metabolized as such or being broken down to pyruvate and then converted to glucose. In view of the oxidation-reduc- tion state of the NAD system and the high activities of malate and lactate dehydrogenases in the cytoplasm of the liver celI (28), any oxalacetate entering the cell would be expected to be largely converted to malate and any pyruvate formed from oxalacetate would be largely converted to lactate which would pass into the medium. The tissue level of malate and the rate of lactate production were therefore measured in livers perfused with oxalacetate and pyruvate. The results (Table IV) showed substantial lactate formation in livers perfused with oxalacetate and the same tissue level of malate with oxalacetate or pyruvate. There was therefore no evidence that osalacetate was being metabolized as such. As expected from its ability to donate hydrogen, ethanol produced large increases in the rate of lactate production and the tissue level of malate (Table IV).
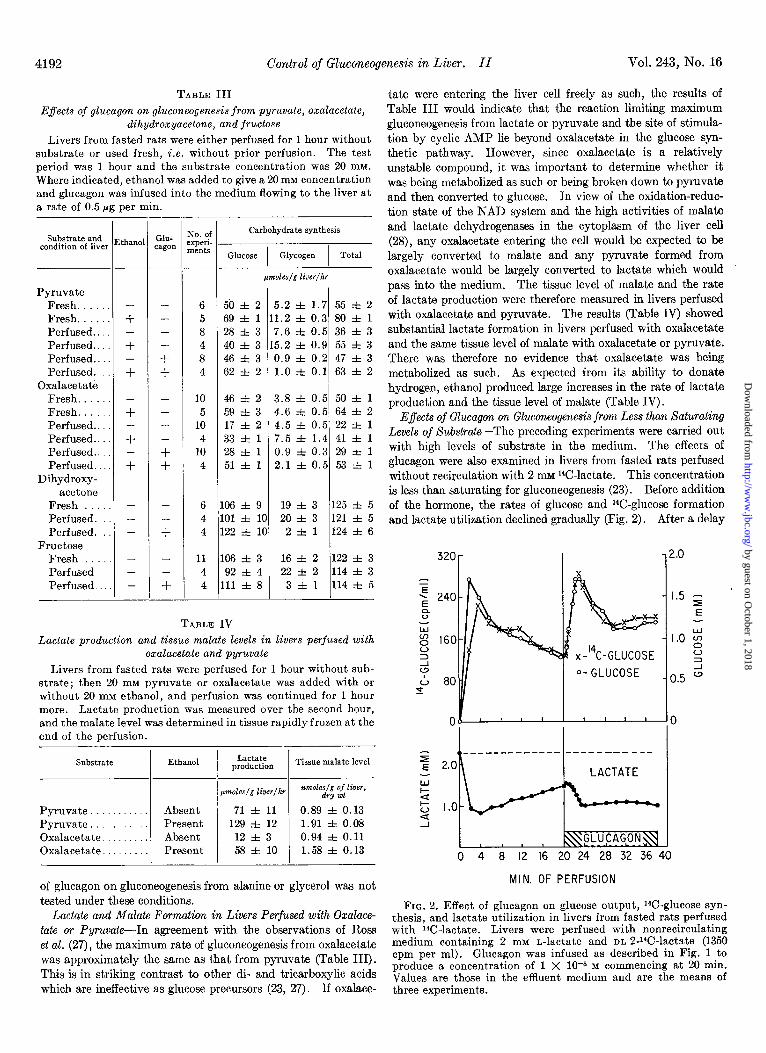
Effects of Glucagon on Gluconeogenesis from Less than Saturating Levels of Substrate-The preceding experiments were carried out with high levels of substrate in the medium. The effects of glucagon were also examined in livers from fasted rats perfused without recirculation with 2 mM 14C-lactate. This concentration is less than saturating for gluconeogenesis (23). Before addition of the hormone, the rates of glucose and W-glucose formation and lactate utilization declined gradually (Fig. 2). After a delay
320
_________ ---- ---- ---- - --
LACTATE
-I ~uJAG$N~
0 4 8 12 16 20 24 28 32 36 40
MIN. OF PERFUSION
FIG. 2. Effect of glucagon on glucose output, r4C-glucose syn- thesis, and lactate utilization in livers from fasted rats perfused with ‘S-lactate. Livers were perfused with nonrecirculating medium containing 2 mM n-lactate and m-2-%-lactate (1350 cpm per ml). Glucagon was infused as described in Fig. 1 to produce a concentration of 1 X 10-S M commencing at 20 min. Values are those in the effluent medium and are the means of three experiments.
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of August 25, 1968 J. H. Exton and C. R. Park 4193
of approximately 1 min, glucagon caused rapid increases in glu- cose and W-glucose production which then declined to steady rates which were 50% higher than those observed prior to addi- tion of the hormone. Because part of the initial increase in 14C-glucose could have been due to the breakdown of glycogen labeled during the period before glucagon addition, the precise onset of the stimulation of gluconeogenesis cannot be known. However, the rapid large increase in lactate utilization occurring almost simultaneously with the acceleration of glucose produc- tion suggests rapid activation of gluconeogenesis.
In similar experiments, glucagon did not affect gluconeogenesis from glycerol, dihydroxyacetone, or fructose. A stimulation of glucose formation from a mixture of amino acids at blood con- centrations was observed (29) and will be reported separately.
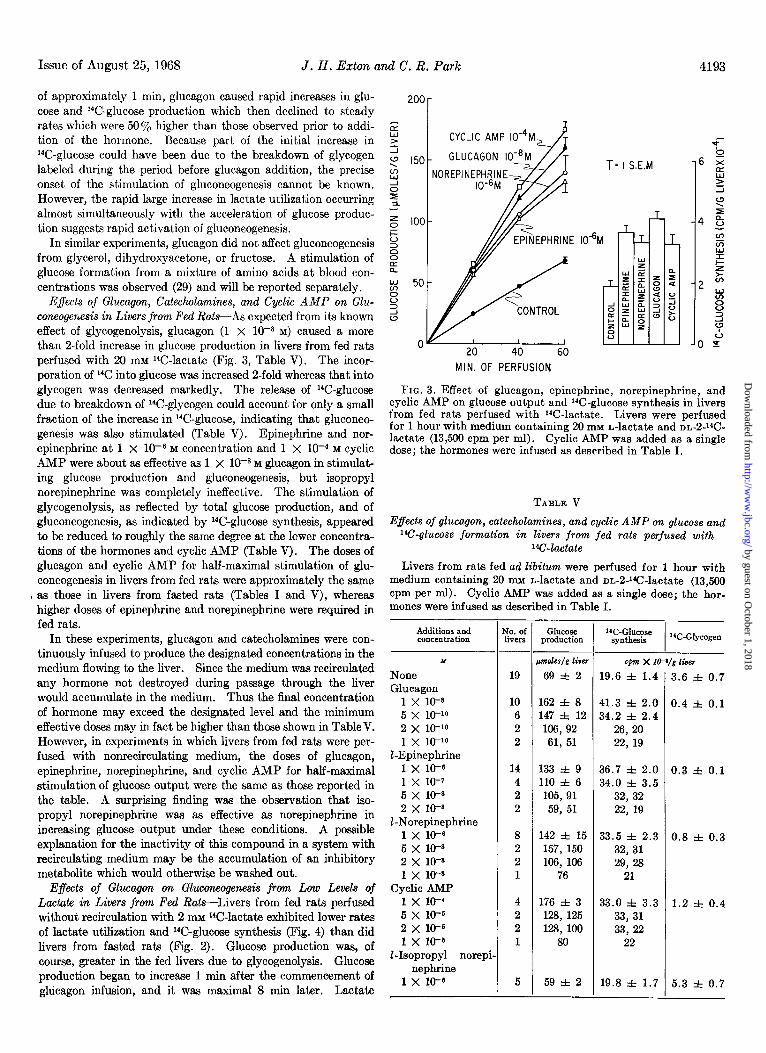
E$ects of Glucagon, Catecholamines, and Q&c AMP on Glu- coneogenesis in Livers from Fed RatsAs expected from its known effect of glycogenolysis, glucagon (1 X 10v8 M) caused a more than 2-fold increase in glucose production in livers from fed rats perfused with 20 mM 14C-lactate (Fig. 3, Table V). The incor- poration of 14C into glucose was increased a-fold whereas that into glycogen was decreased markedly. The release of 14C-glucose due to breakdown of 14C-glycogen could account for only a small fraction of the increase in 14C-glucose, indicating that gluconeo- genesis was also stimulated (Table V). Epinephrine and nor- epinephrine at 1 X 10m6 M concentration and 1 X 1W4 M cyclic AMP were about as effective as 1 x lWs M glucagon in stimulat- ing glucose production and gluconeogenesis, but isopropyl norepinephrine was completely ineffective. The stimulation of glycogenolysis, as reflected by total glucose production, and of gluconeogenesis, as indicated by 14C-glucose synthesis, appeared to be reduced to roughly the same degree at the lower concentra- tions of the hormones and cyclic AMP (Table V). The doses of glucagon and cyclic AMP for half-maximal stimulation of glu- coneogenesis in livers from fed rats were approximately the same
I as those in livers from fasted rats (Tables I and V), whereas higher doses of epinephrine and norepinephrine were required in fed rats.
In these experiments, glucagon and catecholamines were con- tinuously infused to produce the designated concentrations in the medium flowing to the liver. Since the medium was recirculated any hormone not destroyed during passage through the liver would accumulate in the medium. Thus the final concentration of hormone may exceed the designated level and the minimum effective doses may in fact be higher than those shown in TableV. However, in experiments in which livers from fed rats were per- fused with nonrecirculating medium, the doses of glucagon, epinephrine, norepinephrine, and cyclic AMP for half-maximal stimulation of glucose output were the same as those reported in the table. A surprising finding was the observation that iso- propyl norepinephrine was a*4 effective as norepinephrine in increasing glucose output under these conditions. A possible explanation for the inactivity of this compound in a system with recirculating medium may be the accumulation of an inhibitory metabolite which would otherwise be washed out.
Eflects of Glucagon an Gluconeogenesis from Low Levels of Lactate in Livers from Fed Rats-Livers from fed rats perfused without recirculation with 2 mu 14C-lactate exhibited lower rates of lactate utilization and 14C-glucose synthesis (Fig. 4) than did livers from fasted rats (Fig. 2). Glucose production was, of course, greater in the fed livers due to glycogenolysis. Glucose production began to increase 1 min after the commencement of glucagon infusion, and it was maximal 8 min later. Lactate
El > i y 150
22 NOREPINEPHRINE
z 5.
0 20 40 60
MIN. OF PERFUSION
FIG. 3. Effect of glucagon, epinephrine, norepinephrine, and cyclic AMP on glucose output and 14C-glucose synthesis in livers from fed rats perfused with 1°C~lactate. Livers were perfused for 1 hour with medium containing 20 mM L-lactate and DL-2J4C- lactate (13,500 cpm per ml). Cyclic AMP was added as a single dose; the hormones were infused as described in Table I.
TABLE V
E$ects of glucagon, catecholamines, and cyclic AMP on glucose and W-glucose formation in livers from fed rats perfused with
W-lactate
Livers from rats fed ad libitum were perfused for 1 hour with medium containing 20 mu L-lactate and DL-2-‘C-hCt&e (13,500 cpm per ml). Cyclic AMP was added as a single dose; the hor- mones were infused as described in Table I.
Additions and concentration
M
None Glucagon
1 x lo-8 5 x lo-‘0 2 x lo-‘0 1 x NY’0
I-Epinephrine 1 x 10-e 1 x 10-1 5 x 10-S 2 x 10-a
I-Norepinephrine 1 x 10-G 5 x lo-8 2 x lo-* 1 x lo-8
Cyclic AMP 1 x lo-’ 6 x w-6 2 x 10-s 1 x 10-h
l-Isopropyl norepi nephrine
1 x 10-e
- GlUCOSe
production “C-Glucose
synthesis “C-Glycogen
cp?n x lo-‘/g liver
19 mboles/g liver
69 i 2 19.6 f 1.4
10 162 f 8 41.3 * 2.0 6 147 f 12 34.2 f 2.4 2 106,92 26, 20 2 61, 51 22, 19
14 133 i 9 36.7 i 2.0 4 110 i 6 34.0 f 3.5 2 105, 91 32, 32 2 59, 51 22, 19
8 142 i 15 33.5 i 2.3 2 157, 150 32, 31 2 106,106 29, 28 1 76 21
176 f 3 33.0 f 3.3 128, 125 33, 31 128, 100 33, 22
80 22
59 f 2 19.8 i 1.7
3.6 f 0.7
0.4 f 0.1
0.3 * 0.1
0.8 i 0.3
1.2 * 0.4
5.3 * 0.7
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
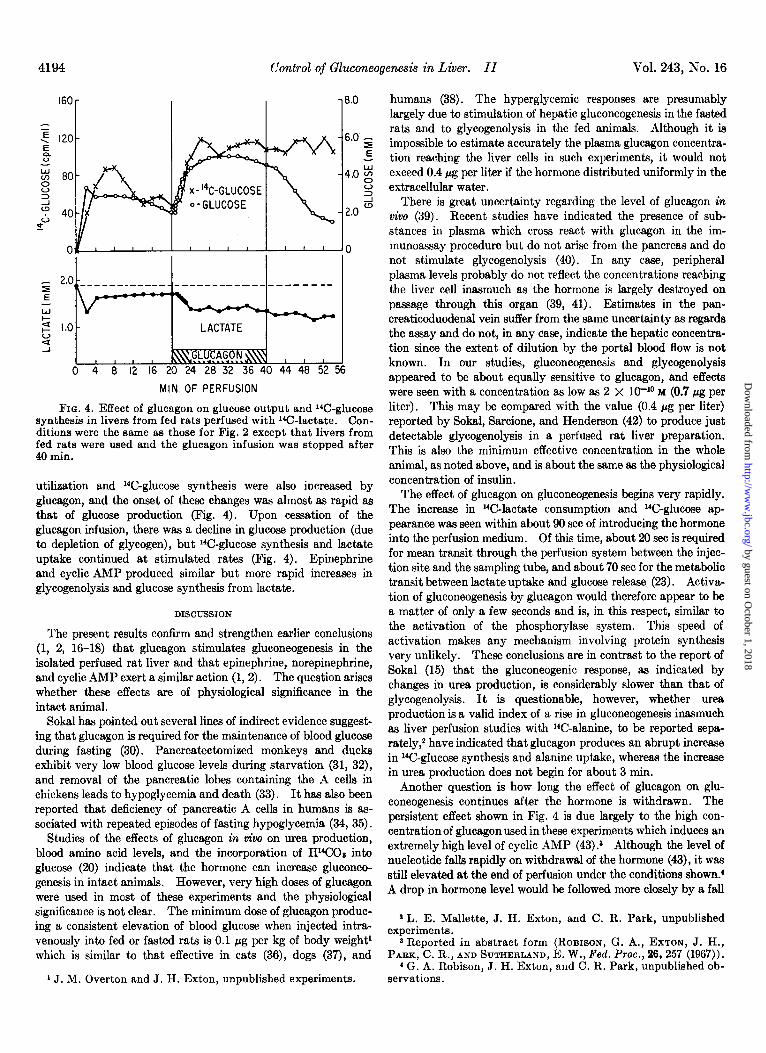
4194 Control of Gluconeogenesis in Liver. II Vol. 243, No. 16
LACTATE :: _)
, , , , ,,w, , ,
0 4 8 12 16 20 24 26 32 36 40 44 48 52 ! MIN. OF PERFUSION
-6
56
FIG. 4. Effect of glucagon on glucose output and 14C-glucose synthesis in livers from fed rats perfused with 14C-lactate. Con- ditions were the same as those for Fig. 2 except that livers from fed rats were used and the glucagon infusion was stopped after 40 min.
utilization and r4C-glucose synthesis were also increased by glucagon, and the onset of these changes was almost as rapid as that of glucose production (Fig. 4). Upon cessation of the glucagon infusion, there was a decline in glucose production (due to depletion of glycogen), but 14C-glucose synthesis and lactate uptake continued at stimulated rates (Fig. 4). Epinephrine and cyclic AMP produced similar but more rapid increases in glycogenolysis and glucose synthesis from lactate.
There is great uncertainty regarding the level of glucagon in v&o (39). Recent studies have indicated the presence of sub- stances in plasma which cross react with glucagon in the im- munoassay procedure but do not arise from the pancreas and do not stimulate glycogenolysis (40). In any case, peripheral plasma levels probably do not reflect the concentrations reaching the liver cell inasmuch as the hormone is largely destroyed on passage through this organ (39, 41). Estimates in the pan- creaticoduodenal vein suffer from the same uncertainty as regards the assay and do not, in any case, indicate the hepatic concentra- tion since the extent of dilution by the portal blood flow is not known. In our studies, gluconeogenesis and glycogenolysis appeared to be about equally sensitive to glucagon, and effects were seen with a concentration as low as 2 x 10-l” M (0.7 pg per liter). This may be compared with the value (0.4 fig per liter) reported by Sokal, Sarcione, and Henderson (42) to produce just detectable glycogenolysis in a perfused rat liver preparation. This is also the minimum effective concentration in the whole animal, as noted above, and is about the same as the physiological concentration of insulin.
DISCUSSION
The present results confirm and strengthen earlier conclusions (1, 2, 16-18) that glucagon stimulates gluconeogenesis in the isolated perfused rat liver and that epinephrine, norepinephrine, and cyclic AMP exert a similar action (1,2). The question arises whether these effects are of physiological significance in the intact animal.
Sokal has pointed out several lines of indirect evidence suggest- ing that glucagon is required for the maintenance of blood glucose during fasting (30). Pancreatectomized monkeys and ducks exhibit very low blood glucose levels during starvation (31, 32), and removal of the pancreatic lobes containing the A cells in chickens leads to hypoglycemia and death (33). It has also been reported that deficiency of pancreatic A cells in humans is as- sociated with repeated episodes of fasting hypoglycemia (34, 35).
The effect of glucagon on gluconeogenesis begins very rapidly. The increase in 14C-lactate consumption and 14C-glucose ap- pearance was seen within about QO set of introducing the hormone into the perfusion medium. Of this time, about 20 set is required for mean transit through the perfusion system between the injec- tion site and the sampling tube, and about 70 set for the metabolic transit between lactate uptake and glucose release (23). Activa- tion of gluconeogenesis by glucagon would therefore appear to be a matter of only a few seconds and is, in this respect, similar to the activation of the phosphorylase system. This speed of activation makes any mechanism involving protein synthesis very unlikely. These conclusions are in contrast to the report of Sokal (15) that the gluconeogenic response, as indicated by changes in urea production, is considerably slower than that of glycogenolysis. It is questionable, however, whether urea production is a valid index of a rise in gluconeogenesis inasmuch as liver perfusion studies with 14C-alanine, to be reported sepa- rately,* have indicated that glucagon produces an abrupt increase in X!-glucose synthesis and alanine uptake, whereas the increase in urea production does not begin for about 3 min.
Studies of the effects of glucagon in wivo on urea production, blood amino acid levels, and the incorporation of H”COa into glucose (20) indicate that the hormone can increase gluconeo- genesis in intact animals. However, very high doses of glucagon were used in most of these experiments and the physiological significance is not clear. The minimum dose of glucagon produc- ing a consistent elevation of blood glucose when injected intra- venously into fed or fasted rats is 0.1 pg per kg of body weight? which is similar to that effective in cats (36), dogs (37), and
Another question is how long the effect of glucagon on glu- coneogenesis continues after the hormone is withdrawn. The persistent effect shown in Fig. 4 is due largely to the high con- centration of glucagon used in these experiments which induces an extremely high level of cyclic AMP (43) .8 Although the level of nucleotide falls rapidly on withdrawal of the hormone (43)) it was still elevated at the end of perfusion under the conditions shown? A drop in hormone level would be followed more closely by a fall
*L. E. Mallette, J. H. Exton, and C. R. Park, unpublished experiments.
*Reported in abstract form (ROBISON, G. A., EXTON, J. H., PARK, C. R., AND SUTHERLAND, E. W., Fed. Proc., 26,257 (1967)).
4 G. A. Robison, J. H. Exton, and C. R. Park, unpublished ob- 1 J. M. Overton and J. H. Exton, unpublished experiments. servations.
humans (38). The hyperglycemic responses are presumably largely due to stimulation of hepatic gluconeogenesis in the fasted rats and to glycogenolysis in the fed animals. Although it is impossible to estimate accurately the plasma glucagon concentra- tion reaching the liver cells in such experiments, it would not exceed 0.4 fig per liter if the hormone distributed uniformly in the extracellular water.
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of August 25, 1968 J. H. Exton and C. R. Park 4195
in gluconeogenesis if physiological levels of glucagon were used. An important additional factor in the termination of gluconeo- genesis in wivo may be insulin secretion which can cause a rapid lowering of cyclic AMP in the liver (44).
The possibility that hepatic glucose production is regulated by blood levels of catecholamines seems unlikely on the basis of the present results and those reported by Sokal (37, 42). The mini- mum effective concentration of epinephrine for stimulation of glucose output by the perfused liver was 2 X lop8 M (4 pg per liter) and that of norepinephrine was 1 X 10-* M (2 pg per liter). These are considerably higher than the resting plasma levels of epinephrine or norepinephrine in man or dogs and above those found during insulin hypoglycemia (4548). The question remains, however, whether hepatic glucose output is controlled by sympathetic nervous activity within the liver. The classical observations of Claude Bernard on piqtire hyperglycemia suggest such a possibility, and there is considerable direct and indirect support for it. Electrical stimulation of the ventromedial hypo- thalamic nuclei or puncture of the floor of the fourth ventricle causes hyperglycemia and hepatic glycogenolysis in fed rabbits (49-51)) and stimulation of the splanchnic nerve increases hepatic phosphorylase activity and glucose production (50, 52). Stimu- lation of nerve fibers in the hepatic pedicle increases the glucose level in the blood of the vena cava and section of nerves in the hepatic pedicle abolishes the hyperglycemic response to stimula- tion of the splanchnic nerves (50).
The above considerations suggest that release of catecholamine from hepatic nerve endings does have a physiological role in the regulation of glucose output by the liver. In general, the cir- cumstances which lead to the secretion of catecholamine or glucagon are quite uncertain owing to the difficulties of estimating concentrations of these substances in vivo at the hepatic cell, as discussed above. Increased gluconeogenesis is well known in diabetes and starvation, and increased levels of cyclic AMP have been observed in the liver in these conditions (44).4 We have proposed that the elevation of the nucleotide is due to an altera- tion in the balance between glucagon and sympathetic nervous activity on the one hand and insulin on the other (2,44). Accord- ing to this concept, the presence of glucagon and/or catechola- mine is required for increased gluconeogenesis, but the levels of these hormones need not be increased because diminished insulin secretion could have the same result.
It appears likely that catecholamine secretion accelerates the removal of blood lactate and its conversion to glucose during exercise, or in times of increased sympathetic activity during acute stress. This would constitute another facet of catechola- mine action in the Cori cycle which would supplement the known effects of these hormones on lactate production in muscle and glycogenolysis.
The observation that exogenous cyclic AMP stimulates glu- coneogenesis in the perfused liver suggests that this nucleotide mediates the action of glucagon and catecholamines on glu- coneogenesis (1, 2, 43). The concentration of exogenous cyclic AMP required to produce effects on gluconeogenesis was the same as that needed for glycogenolysis, but was about lo-fold higher than the intracellular concentration (1.5 X 10m6 M) found with minimally effective doses of glucagon or catecholamines (43). This discrepancy presumably arises from the slow penetration of exogenous cyclic AMP into intact cells in vitro or in viva and its rapid intracellular degradation by the adenosine 3’) 5’-phosphate diesterase. Consequently, large doses (13 pmoles per kg) have
to be administered to dogs to produce hyperglycemia (53) and are required to stimulate lipolysis in isolated fat cells (54). Menahan and Wieland (55) have recently shown that dibutyryl cyclic AMP, a biologically active analogue of the natural com- pound, stimulates gluconeogenesis. Cyclic AMP causes the same changes in the cellular levels of the intermediates along the gluconeogenic pathway as do glucagon and the catecholamines (2, 3), and glucagon and epinephrine cause very rapid increases in intracellular cyclic AMP (43) .4
As reported in preceding papers (2, 23), the step limiting the maximum rate of gluconeogenesis from lactate appears to be located in the sequence of reactions converting pyruvate to P-pyruvate. Cyclic AMP must accelerate this limiting reaction and perhaps other reactions. An effect of cyclic AMP in the gluconeogenic pathway between pyruvate and P-pyruvate is consistent with the changes in intermediary metabolites reported in tivo after glucagon (19) and found in livers pe:fused with lac- tate (1, 2) or amino acids2 It also agrees with the observation that glucagon stimulates gluconeogenesis from lactate and pyruvate but not from dihydroxyacetone and fructose which enter the gluconeogenic pathway mainly at the level of the triose phosphates (56, 57).
The results with oxalacetate need special comment. As referred to earlier, the finding of the same maximum rate of glu- coneogenesis with oxalacetate, pyruvate, and lactate should not be interpreted as indicating that the rate-limiting step for lactate gluconeogenesis lies beyond oxalacetate in the pathway. This is because it is questionable whether oxalacetate is metabolized as such by the liver. Although oxalacetate rapidly disappears from the medium perfusing livers (26), it does not increase the tissue concentration of malate above that found with pyruvate (Table IV). There is, therefore, no evidence that it can enter the intact liver cell, and the lactate and glucose formation observed by Ross et al. (26) and in this study (Tables III and IV) may have been caused by extracellular breakdown of oxalacetate to pyruvate. It would be surprising if oxalacetate entered the liver cell freely in view of the poor penetration of other di- and tricarboxylic anions (23, 26). In view of the foregoing considera- tions, the effect of glucagon on gluconeogenesis from oxalacetate probably reflects the action of glucagon on glucose synthesis from a limited concentration of pyruvate.
The stimulatory effect of ethanol on gluconeogenesis from pyruvate (Table III) is in marked contrast to its inhibitory action on glucose synthesis from lactate, alanine, and endogenous sub- strates (58-63). The inhibitory effect of ethanol on gluconeo- genesis is usually ascribed to an increase in the NADH-NAD+ ratio of the cytoplasm (58-63). Such a change would explain the increased lactate production and malate levels observed in the present study (Table IV) and might account for the increased gluconeogenesis from pyruvate. It is possible that with the high concentration of pyruvate used in these experiments, rapid formation of lactate caused a heavy drain on cytoplasmic NADH, limiting gluconeogenesis at the level of the 3-phosphoglyceralde- hyde dehydrogenase reaction, and that the limitation was relieved by ethanol. It is highly improbable that NADH limits glu- coneogenesis in vivo inasmuch as the pyruvatc level of the blood is normally 0.1 mM and lactate-pyruvate ratios less than 8:l have not been reported.
The proposal that glucagon acts on gluconeogenesis by increas- ing the cytoplasmic NADH level (19, 20) is not supported by several observations. First, the acceleration of carbohydrate
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4196 Control of Gluconeogenesis in Liver. II Vol. 243, No. 16
synthesis from pyruvate by glucagon was not diminished in the presence of ethanol (Table III). Second, the effects of ethanol and glucagon on gluconeogenesis from lactate are in opposite directions. Third, glucagon does not produce increases in the tissue lactate-pyruvate or glycerol-l-P-dihydroxyacetone-P ratios in livers perfused with lactate or alanine (3, 43).2* 6
The mechanism by which perfusion without substrate inacti- vates gluconeogenesis is not entirely clear. We suggest, how- ever, that inactivation results from the conversion of the principal rate-limiting enzyme to a less active form consequent to a fall in tissue cyclic AMP. Presumably, the liver rapidly inactivates glucagon and catecholamines carried over from the animal (or present in certain batches of crude albumin), and the tissue cyclic AMP has been found to decline during perfusion.’ Another factor must also be involved, however, since inactivation can be prevented by high levels of lactate. This may be an example of protection of enzyme activity by substrate in an intact cell.
The phenomenon of inactivation in. vitro may reflect an aspect of the regulation of gluconeogenesis in uivo in which the liver changes over from the active gluconeogenic state during fasting to the less active state associated with feeding. The inactivation in both circumstances would be due in part to loss or diminution in the secretion of glucagon and catecholamines. Consistent with such an interpretation is the observation that the level of cyclic AMP in vivo is reduced by feeding.
AcknowZedgmentsWe wish to thank T. B. Miller, Jr., J. G. Corbin, and J. P. Brineaux for their skilled and conscientious technical assisitance.
1. 2.
3. 4. 5.
6.
7. 8.
EXTON, J. H., AND PARK, C. R., Pharmacol. Rev., 18,181 (1966). EXTON, J. H., JEFFERSON, L. S., BUTCHER, R. W., AND PARK,
C. R., Amer. J. Med., 40, 709 (1966). TYBERGHEIN, J. M., Arch. Int. Physiol., 61, 104 (1953). KALANT, N., Proc. Sot. Ezp. Biol. Med., 86, 617 (1954). ROSE, S., AND NELSON, J., Australian J. Exp. Biol. Med. Sci.,
84, 33 (1956). SALTER, J. M., DAVIDSON, I. W. F., AND BEST, C. H., Diabetes,
6, 248 (1957). Izzo, J. L., AND GLASSER, S. R., Endocrinology, 68, 189 (1961). SHOEMAKER, W. C., AND VAN ITALLIE, T. B., Endocrinology,
66, 260 (1960). 9.
10. KALANT, N., Arch. Biochem. Biophys., 66, 469 (1956). CURRY, D. M., AND BEATON, G. H., Endocrinology, 63, 252
(1958). 11. 12. 13.
14.
15. 16.
MILLER, L. L., Nature, 186, 248 (1960). MILLER; L. L., Recent Progr. Hormone Res., 17, 539 (1961). WILLIAMSON, J. R., GARCIA, A., RENOLD, A. E., AND CAHILL,
G. F., JR., Diabetes, 16, 183 (1966). PENHOS, T. C., WV, C. H., DAUNAS, J., REITMAN, M., AND
LEVINE. R., Diabetes, 16, 740 (1966). SOKAL, J.’ E.; Endocrinology, 78; 538 (1966). SCHIMASSEK, H., AND MITZKAT, H. J., Biochem. Z., 337, 510
(1963). 17. GARCIA, A., WILLIAMSON, J. R., AND CAHILL, G. F., JR.,
Diabetes, 16, 188 (1966). 18. STRUCK. E.. ASHMORE. J.. AND WIELAND. 0.. Biochem. Z..
343, 107 (i965). ’ ’ I I
19. 20. 21.
WILLIAMSON, J. R., Biochem. J., 101, llc (1966). WILLIAMSON, J. R.. Advance. Enzume Reaulat.. 6. 229 (1967). WILLIAMSON; J. R.‘, WRIGHT, P. h., MALAIsSE,-W. J:, AND
ASHMORE, J., Biochem. Biophys. Res. Commun., 24, 765 (1966).
22. MILLER, L. L., Fed. Proc., 24, 737 (1965). 23. EXTON, J. H., AND PARK, C. R., J. Biol. Chem., 242,2622 (1967).
6 J. H. Exton and C. R. Park, unpublished observations.
REFERENCES
24.
25.
26.
27.
28.
29.
30. 31.
32. 33. 34.
35.
36.
37. 38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48. 49.
50.
51.
52. 53.
54.
55.
56.
57.
58.
59.
60.
61.
62. 63.
HOHORST, H. J., in H. U. BERG~XEYER enzymatic analysis, Academic Press, 328.
SUTHERLAND, E. W., AND RALL, T. W., 265 (1960).
(Editor), Methods of New York, 1963, p.
Pharmacol. Rev., 12,
HEMS, R., Ross, B. D., BERRY, M. N., AND KREBS, H. A., Biochem. J., 101, 284 (1966).
Ross, B. D., HEMS, R., AND KREHS, H. A., Biochem. J., 102, 942 (1967).
HOHORST, H. J., KREUTZ, F. H., AND BUCHER, T., Biochem. Z., 332, 18 (1959).
MALLETTE, L. E., EXTON, J. H., AND PARK, C. R., Fed. Proc., 26, 563 (1967).
SOKAL, J. E., Amer. J. Med., 41, 331 (1966). COLLIP, J. B., SELYE, H., AND NEUFELD, A., Amer. J. Physiol.,
119, 289 (1937). MIALHE, P., Acta Endocrinol., 28 (Suppl. 36), 134 (1958). MIKAMI, S. I., AND ONO, K., Endocrinology, 71, 464 (1962). MCQUARRIE, I., BELL, E. T., ZIMMERMANN, B., AND WRIGHT,
W. S., Fed. Proc., 9, 337 (1950). GROLLMAN, A., MCCALEB, W. E., AND WHITE, F. N., Metab.
Clin. Exp., 13, 686 (1964). STAUB, A., SINN, L., AND BEHRENS, 0. K., J. Biol. Chem., 214,
619 (1955). EZDINLI, E. Z., AND SOKAL, J. E., Endowinology, 78,47 (1966). BUTTERFIELD, W. J. H., FRY, I. K., AND WHICHELOM~, M. J.,
Guy’s Hosp. Rep., 109, 95’ (196Oj. SOKAL. J. E.. AND EZDINLI. E. Z.. J. Clin. Invest.. 46. 778
(196j). ’ UNGER; R. H., OHNEDA, O., VALVERDE, I., EISENTRAUT, A.
M.. AND EXTON. J. H.. J. Clin. Invest.. 47. 48 (1968). BERS~N, S. A., YA~o\v, k. S., AND VOLK, k. w., J.~ Lab: Clin.
Med.,‘49, 33i (1957).’ SOKAL. J. E.. SARCIONE. E. J.. AND HENDERSON. A. &I.. Endo-
crindlogy, j4, 930 (1964). ’ EXTON, J. H., AND PARK, C. It., Advance. Enzyme Regulat., 6,
391 (1968). JEFFERSON, L. S., EXTON, J. H., BUTCHER, R. W., SUTHER-
LAND, E. W., AND PARIC, C. R., J. Biol. Chem., 243, 1031 (1968).
HOLZBAUER, M., AND VOGT, M., Bril. J. Pharmacol., 9, 249 (1954).
GOLDFIEN, A., ZILELI, M. S., DESPOINTES, R. H., AND BE- THUNE, J. E., Endocrinology. 62, 749 (1958).
GOLDFIE~, A., MOORE, R., ZI&I, M.,HA~ENs, L. L., BOLING, L.. AND THORN. G. W.. J. Clin. Endowinol. Metab.. 21. 296 (1961). ’ ’
VENDSALU,.A., Acta Physiol. &and., 173 (Suppl.), 33 (1960). BERNARD, C., Lecons sur le diabkte et la glycogenbse animale,
J. B. Baillibre et fils, Paris, 1877. MACLEOD, J. J. R., Carbohydrate metabolism and insulin,
Lonemans. Green and Co.. Ltd.. London. 1926. SHIMA&, T:, FUKUDA, A., XND BAN, T., ‘Nature, 210, 1178
(1966). SHIMAZU, T., AND FUKUDA, A., Science, 160, 1607 (1965). HENION, W. F., SUTHERLAND, E. W., AND POSTERNAK, T.,
Biochim. Biophys. Acta, 148, 106 (1967). BUTCHER, R. W., Ho, R. J., MENG, H. C., AND SUTHERLAND,
E. W., J. Biol. Chem., 240, 4515 (1965). MENAHAN, L., AND WIELAND, O., Biochem. Biophys. Res. Com-
mm., 29, 880 (1967). HERS, H. G., Le metabolism du jructose, Editions Arscia,
Brussels, 1957. RAUSCHENBACH, P., AND LAMPRECHT, W., Hoppe-Seyler’s Z.
Physiol. Chem., 339, 277 (1964). FIELD, J. B., WILLIAMS, H. E., AND MORTIMORE, G. E., J.
Clin. Znvest., 42, 497 (1963). FREINKEL, N., COHEN, A. K., ARKY, It. A., AND FOSTER,
A. E., J. Clin. Endocrinol Metab., 26, 76 (1965). FORSANDER, 0. A., R;~IH& N., SALASPURO, M., AND MXENPKK,
P., Biochem. J., 94, 259 (1965). LOCHNER, A., WULFF, J., AND MADISON, L. I,., MetaD. Clin.
Exp., 16, 1 (1967). KREBS, H. A., Advance. Enzyme Regulal., in press. KREISRERG, R. A., Diabetes, 16, 784 (19671.
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. H. Exton and C. R. ParkGLUCONEOGENESIS IN THE PERFUSED RAT LIVER
CATECHOLAMINES, AND ADENOSINE 3',5'-MONOPHOSPHATE ON Control of Gluconeogenesis in Liver: II. EFFECTS OF GLUCAGON,
1968, 243:4189-4196.J. Biol. Chem.
http://www.jbc.org/content/243/16/4189Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/16/4189.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on October 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from