gluconeogenesis the synthesis of glucose from noncarbohydrate precursors mainly in the liver
TRANSCRIPT

GLUCONEOGENESIS
The synthesis of glucose from noncarbohydrate precursors
Mainly in the liver

IMPORTANT PRINCIPLES OF BIOSYNTHESIS
The synthesis pathway is usually different from the degradation pathway.
The two opposing pathways may share many reversible reactions.
There is always at least one unique enzymatic step to each pathway.
BECAUSEIf this wasn’t the case, then the flow of carbon
through the two pathways would be dictated by the mass action and not the cellular changing
needs for energy, precursors or macromolecules.

IMPORTANT PRINCIPLES OF BIOSYNTHESIS
Corresponding anabolic and catabolic pathways are controlled by different regulatory enzymes in a reciprocal manner.
If one pathway is stimulated, then the opposite is inhibited.
Biosynthetic pathways are usually regulated at their initial steps.
BECAUSETo prevent wasting precursors to make unneeded intermediates.

IMPORTANT PRINCIPLES OF BIOSYNTHESIS
The energy-requiring biosynthetic processes are coupled to the energy-yielding hydrolysis of ATP
The overall process is essentially irreversible in vivo. The total amount of energy from ATP (and
NAD(P)H) is larger than the minimum energy needed to convert the precursor into the biosynthetic product.
THUSThe resulting large negative free energy for the overall process will assure that it will take place even when the concentrations of the precursors are relatively low.
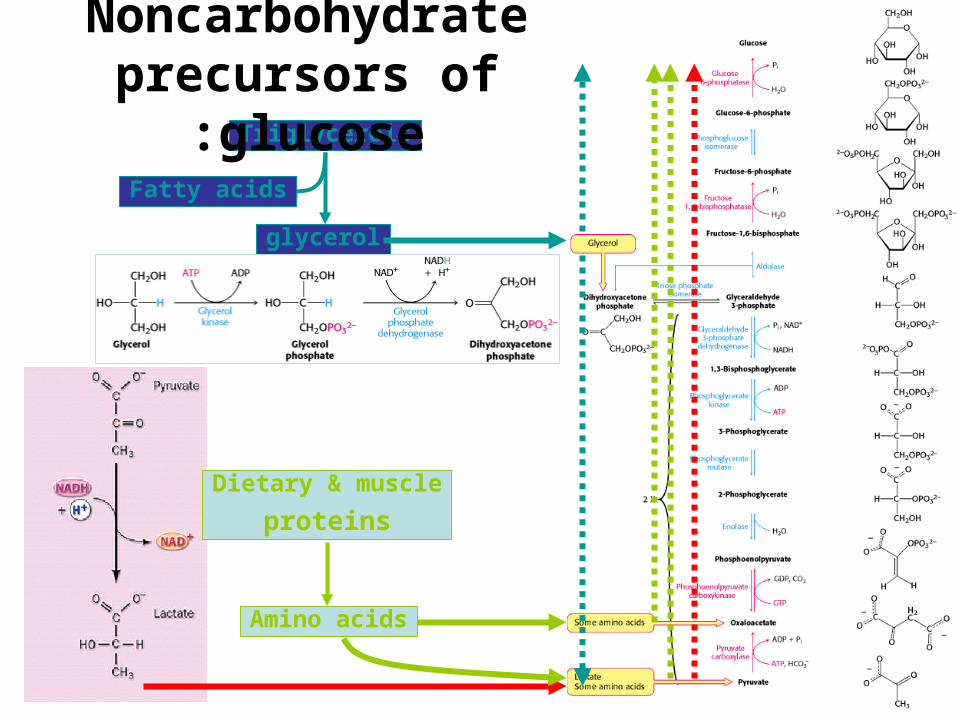
Dietary & muscle
proteins
Amino acids
Triglycerols
glycerol
Fatty acids
Noncarbohydrate precursors of glucose:

Main sites of gluconeogenesis:
•Major site: Liver.•Minor site: Kidney.•Very little:
–Brain.–Muscle (skeletal and heart).
gluconeogenesis in the liver and kidney helps to maintain the glucose level in the blood so that
brain and muscle can extract sufficient glucose from it to meet their metabolic demands.

Gluconeogenesis Is Not a Reversal of Glycolysis:
•Seven steps are shared by glycolysis and gluconeogenesis.
•However, three essentially irreversible steps in glycolysis shift the equilibrium far
in the side of glycolysis.
•Most of the decrease in free energy in glycolysis takes place in these steps.


In gluconeogenesis the three reactions are bypassed by a set
of separate enzymes.Phosphoenolpyruvate is formed from pyruvate:
.2Fructose 6-phosphate is formed from fructose 1,6-bisphosphate:
.3Glucose is formed by hydrolysis of glucose 6-phosphate:

PYRUVATEPHOSPHOENOLPYROVATE
A
B

PYROVATE CARBOXYLASE
Mitochondrial enzyme

.ACARBOXYLATION OF PYROVATE:
2
3
1
HCO3-, the aqueous form of CO2 is activated to
carboxyphosphate.
it is subsequently activated by binding to the N-1 atom of the biotin ring to form the carboxybiotin-enzyme intermediate. The G°´ for its cleavage is -20 kJ mol-1.
The activated carboxyl group is then transferred from carboxybiotin to pyruvate to form oxaloacetate.


•The long, flexible link between biotin and the
enzyme enables this prosthetic group to rotate
from one active site of the enzyme (the ATP-bicarbonate site) to the other (the pyruvate
site).•To be carboxylated, biotin
needs the enzyme to be allosterically activated by
Acetyl CoA.

.BDECARBOXYLATION AND
PHOSPHORYLATION OF OXALOACETATE:
•Oxaloacetate, is reduced to malate inside the mitochondrion
for transport to the cytosol.–The reduction is accomplished by
an NADH-linked malate dehydrogenase.
•When malate has been transported across the
mitochondrial membrane, it is reoxidized to oxaloacetate by an
NAD+-linked malate dehydrogenase in the cytosol.

•Oxaloacetate is simultaneously decarboxylated and phosphorylated by
phosphoenolpyruvate carboxykinase in the cytosol.
–The CO2 that was added to pyruvate by pyruvate carboxylase comes off in this step.
–The formation of the unstable enol is driven by decarboxylation, and trapped by the addition
of a phosphate to carbon 2 from GTP .

Pyruvate
Pyruvate
Oxaloacetate
Malate
Malate
Oxaloacetate PEP
CO2
NADHH++NAD+
NADH + H+
NAD+ CO2
Pyruvate CARBOXYLASE
Cytosolic PEP CARBOXYKINASE
Cytosolic Malate DEHYDROGENASE

•This pathway predominates when pyruvate or alanine is the glucogenic precursor.
•The carboxylation-decarboxylation represents a way of activating pyruvate.
–the decarboxylation of Oxaloacetate facilitates PEP formation.
•The cytosolic NADH is consumed by other gluconeogenesis reactions, and has to be
regenerated in order to proceed with the process.
•This is accomplished by transporting Malate outside the mitochondria.

NADH + H+
NAD+
Lactate
Pyruvate
Oxaloacetate
PEP
Pyruvate
Pyruvate
Oxaloacetate
Malate
Malate
Oxaloacetate PEP
CO2
CO2
CO2
NADHH++NAD+
NADH + H+
NAD+ CO2
Pyruvate CARBOXYLASE
Pyruvate CARBOXYLASE
Mitochondrial PEP CARBOXYKINASE
Cytosolic PEP CARBOXYKINASE
Lactate DEHYDROGENASE
Cytosolic Malate DEHYDROGENASE
Pyruvate

•This pathway predominates when lactate is the precursor.
•The conversion of lactate to pyruvate in the hepatocyte cytosole yields NADH.
•Thus no Malate transport is needed any more for this purpose.
•The mitochondrial and cytosolic PEP CARBOXYKINASE enzymes are encoded by
separate nuclear genes. (two different enzymes catalyzing the same reaction in different
localizations)

FRUCTOSE 1,6-BISPHOSPHATEFRUCTOSE 6-PHOSPHATE
•The enzyme responsible for this step is fructose 1,6-bisphosphatase.
•Like its glycolytic counterpart, it is an allosteric enzyme that participates in the regulation of
gluconeogenesis.

GLUCOSE 6-PHOSPHATEGLUCOSE
•This hydrolytic reaction is catalyzed by glucose-6-phosphatase in the Endoplasmic
reticulum of the hepatocytes..•In most tissues, free glucose is not generated;
–the glucose 6-phosphate is processed in some other fashion, notably to form glycogen.
•Unlike free glucose, glucose 6-phosphate cannot diffuse out of the cell.

•To keep glucose inside the cell, the generation of free glucose is controlled in
two ways:.1The enzyme responsible for the conversion of
glucose 6-phosphate into glucose, glucose 6-phosphatase, is regulated.
.2The enzyme is present only in tissues whose metabolic duty is to maintain blood-glucose
homeostasis tissues that release glucose into the blood (the liver and to a lesser extent the
kidney).

•Several endoplasmic reticulum (ER) proteins play a role in the generation of
glucose from glucose 6-phosphate.–T1 transports glucose 6-phosphate into the
lumen of the ER.–T2 and T3 transport Pi and glucose,
respectively, back into the cytosol.–Glucose 6-phosphatase is stabilized by a
Ca2+-binding protein (SP).









REGULATION OF CARBOHYDRATE
METABOLISM

•The rate of conversion of glucose into pyruvate is regulated to meet
two major cellular needs:.1The production of ATP, generated by
the degradation of glucose.
.2The provision of building blocks for synthetic reactions, such as the
formation of fatty acids.

It is all in the enzymes
•Enzymes can enhance the rates of metabolic (or other) reactions by many orders of magnitude.
•A rate enhancement of 1017 means that what would occur in 1 second with an enzyme’s help,
would otherwise require 31,710,000,000 years to take place.
•So essentially without enzymes such reactions don’t take place.
•Thus, regulation of enzymatic activity is in a sense, regulation of metabolism, or any other
cellular process.

Regulation and control of enzyme activity
•Substrate level control.•Allosteric effectors•Covalent modification•Enzyme concentration: increased synthesis•Enzyme concentration: generation of active
enzyme by processing•Substrate cycles

Substrate level control
•Since most often [S] > Km, the change in substrate concentration does not
change the reaction rate appreciably.
•Thus, controlling a metabolic flux is not
normally achieved by varying substrate
concentrations.

Allosteric effectors
•Noncovalently bind and regulate the enzyme.
•The effector may be stimulatory or inhibitory.
•The substrate and effector usually occupy
different specific binding sites.

Allosteric enzymes kinetics:
•Sigmoid kinetic behavior is seen.
•K0.5 represents the substrate concentration
at which the enzyme velocity is half Vmax.
• (-)and (+) respectively indicate inhibitory and
stimulatory effectors.

Covalent modification

Enzyme concentration: increased synthesis

Enzyme concentration: generation of active enzyme by
processing

Regulation of the flux through multistep pathways occurs at steps that are enzyme limited:
•RATE-LIMITING STEP: the rate of at least one reaction in
every metabolic pathway depends on the activity of the enzyme (ENZYME-LIMITED),
and is not limited to the substrate availability.

.1Any enzyme that catalyzes the 1st step in a pathway is a potential control point since it shows
“commitment” to the pathway.–Phosphofructokinase is the obvious point in glycolysis.
.2Any enzyme that is working slowly (small Vmax) is
obviously a bottle-neck in the reaction.–Therefore activation of a slow enzyme can increase
the flux of the entire pathway.–In heart muscle glycolysis the slowest enzymes are:
•Hexokinase.•Phosphofructokinase.•Aldolase.•Enolase.

.3Irreversible reactions in glycolysis (rate-limiting) are potential sites of control.
–Hexokinase–Phosphofructokinase–pyruvate kinase .
•Their activities are regulated by:–the reversible binding of allosteric effectors–by covalent modification.–the amounts of these important enzymes are
varied by the regulation of transcription to meet changing metabolic needs .

Gluconeogenesis and Glycolysis Are Reciprocally Regulated
•The amounts and activities of the distinctive enzymes of each pathway are controlled so that both pathways are not
highly active at the same time.•The interconversion of fructose 6-
phosphate and fructose 1,6-bisphosphate is stringently controlled.
–Phosphofructokinase and fructose 1,6-bisphosphatase are reciprocally controlled by
fructose 2,6-bisphosphate in the liver


ALLOSTERIC REGULATORS OF PFK-1 and FBPase-1

PHOSPHOFRUCTOKINASE: The most important control element in the
mammalian glycolytic pathway.•PFK in the liver is a
tetramer of 4 identical subunits.
•The allosteric effectors binding site
is distinct from the catalytic site.
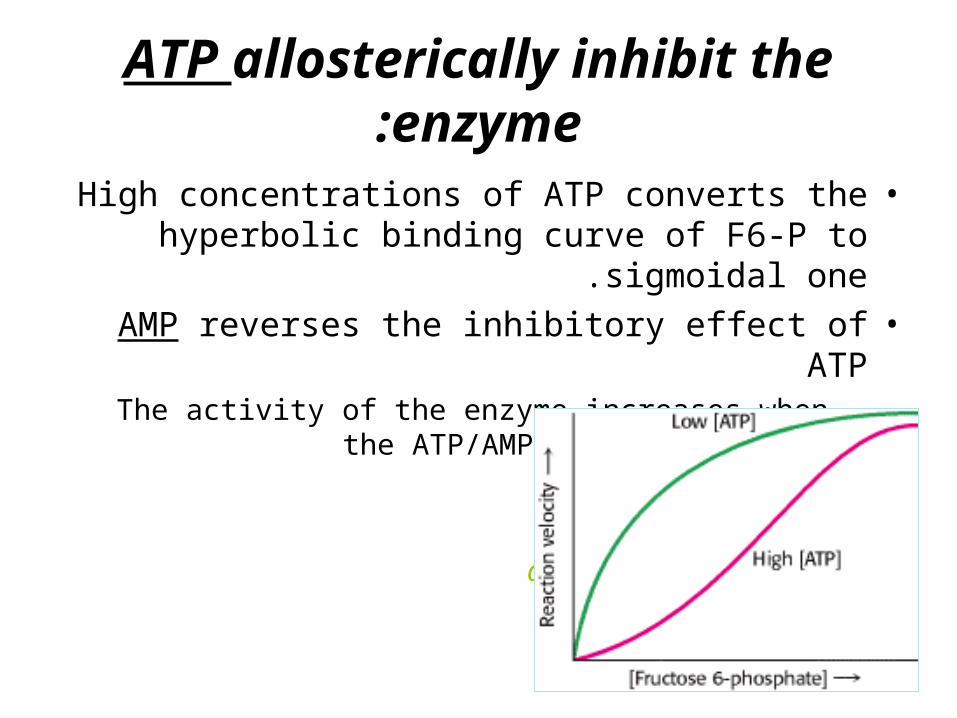
ATP allosterically inhibit the enzyme:
•High concentrations of ATP converts the hyperbolic binding curve of F6-P to sigmoidal
one.•AMP reverses the inhibitory effect of ATP
–The activity of the enzyme increases when the ATP/AMP ratio is lowered
glycolysis is stimulated
as the energy charge falls

WHY is AMP rather than ADP the positive regulator of PFK-1?
•When ATP is utilized rapidly, the enzyme Adenylate Kinase forms ATP and AMP from
ADP:
•AMP becomes the signal for low energy charge.
ADP + ADP ATP + AMP

Citrate inhibit the PFK-1 enzyme
•A high level of citrate means that biosynthetic precursors are abundant and
additional glucose should not be degraded for this purpose.
•Citrate inhibits PFK-1 by enhancing the inhibitory effect of ATP.

F2,6-BP allosterically activates PFK-1 and inhibits FBPase:
•Phosphofructokinase and fructose 1,6-bisphosphatase are reciprocally controlled by
fructose 2,6-bisphosphate in the liver.

.AThe sigmoidal dependence of velocity on substrate concentration becomes hyperbolic.
.BATP acting as substrate initially stimulates the reaction and afterward acts as allosteric
inhibitor–The inhibitory effect of ATP is reverse by F2.6-BP

How is the concentration of F 2,6-BP appropriately controlled?
•F2,6-BP is formed in a reaction catalyzed by Phosphofructokinase-2 (PFK-2)
•It is hydrolyzed to F6-P by Fructose Bisphosphatase-2 (FBPase-2)
•Both PFK-2 and FBPase-2 are part of the same 55Kd polypeptide chain.
•The bifunctional enzyme Exists in 5 isozymic forms.
–L-isoform in liver.–M-isoform in muscle.

The L-isoform help to maintain blood-glucose homeostasis:
Feedforward stimulation

HEXOKINASE
PHOSPHOGLUCOSEISOMERASE
PHOSPFOFRUCTOKINASE
Fructose 6-phosphateaccomulates
glucose 6-phosphateaccomulates
• Glucokinase has ~ 50-fold lower affinity for glucose than does hexokinase.
• The low affinity in the liver gives the brain and muscles first call on glucose when its supply is limited, whereas it ensures that glucose will not be wasted when it is abundant.
Synthesis of glycogen and fatty acids.
GLUCOKINASE
ATP
ADP
In LIVERHexokinase:

Why is phosphofructokinase rather than hexokinase the
pacemaker of glycolysis? •Glucose 6-phosphate is not solely a glycolytic
intermediate.–It can also be converted into glycogen or it can be
oxidized by the pentose phosphate pathway to form NADPH.
•The first irreversible reaction unique to the glycolytic pathway, the committed step, is the
phosphorylation of fructose 6-phosphate to fructose 1,6-bisphosphate .

The interconversion of PEP and pyruvate is precisely regulated.
•gluconeogenesis is favored when the cell is rich in biosynthetic precursors and ATP .

Pyruvate kinase L-isoform• Pyruvate kinase is regulated by
allosteric effectors and covalent modification.
• These hormone-triggered phosphorylations, prevent the liver from consuming glucose when it is more urgently needed by brain and muscle
PHOSPHATASE

GLUCOSE TRANSPORTRSNameTissue locationKm Comments
GLUT1All mammalian tissues
1 mMBasal glucose uptake
GLUT2Liver and pancreatic cells
15-20 mM
In the pancreas, plays a role in regulation of insulin secretion
In the liver, removes excess glucose from the blood
GLUT3All mammalian tissues
1 mMBasal glucose uptake
GLUT4Muscle and fat cells5 mMAmount in muscle plasma membrane increases by insulin and with endurance
training
GLUT5Small intestine Primarily a fructose transporter


HORMONS control the amount and activities of essential
enzymes•Hormones affect gene expression
primarily by:.1changing the rate of transcription
.2regulating the degradation of mRNA.
•Transcriptional control in eukaryotes is much slower than allosteric control;
–it takes hours or days in contrast with seconds to minutes .

Insulin
•Rises subsequent to eating and stimulates the expression of:
–Phosphofructokinase
–pyruvate kinase
–PFK-2/FBPase-2

Glucagon
•Rises during starvation:•inhibits the expression of:
–Phosphofructokinase–pyruvate kinase–PFK-2/FBPase-2.
•stimulates instead the production of two key gluconeogenic enzymes:
–phosphoenolpyruvate carboxykinase–fructose 1,6-bisphosphatase

Hormones work at the promoter level
•The PEP-Carboxykinase promoter approximately 500 bp in length
•Contains regulatory sequences (response elements) that mediate the action of several hormones:
–IRE: insulin response element
–GRE: glucocorticoid response element
–TRE: thyroid hormone response element
–CREI and CREII: cAMP response elements.
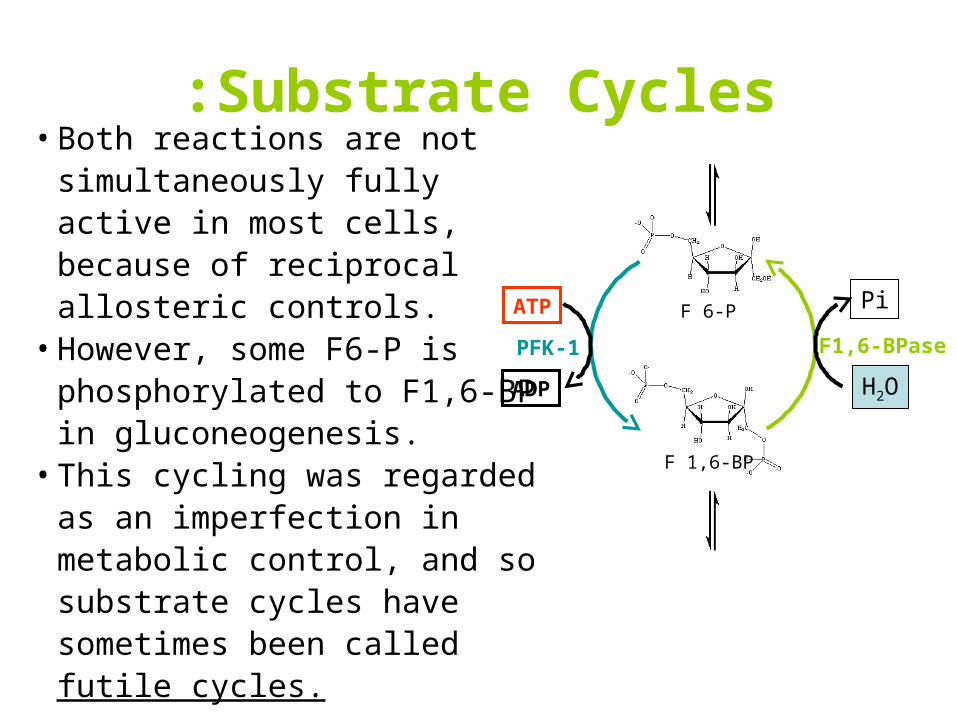
Substrate Cycles:• Both reactions are not
simultaneously fully active in most cells, because of reciprocal allosteric controls.
• However, some F6-P is phosphorylated to F1,6-BP in gluconeogenesis.
• This cycling was regarded as an imperfection in metabolic control, and so substrate cycles have sometimes been called futile cycles.
F 6-P
F 1,6-BP
H2O
Pi
F1,6-BPase
ATP
ADP
PFK-1

Substrate cycles are biologically important:
.1Substrate cycles amplify metabolic signals:
–This amplification is made possible by the rapid hydrolysis of ATP.
If an allosteric effector reciprocally increases A to B and decreases B to A by 20% each
Then a 20% change in the rates of the opposing reactions has led to a 480% (=100x48/10) increase in the net flux.
.2generation of heat produced by the hydrolysis of ATP.

Lactate produced by active skeletal muscle and erythrocytes is a source of
energy for other organs.•The only purpose of the reduction of pyruvate to
lactate is to regenerate NAD+ so that glycolysis can proceed in active skeletal muscle and
erythrocytes.•lactate is a dead end in metabolism.
–It must be converted back into pyruvate before it can be metabolized.

• Both pyrovate and lactate diffuse out of active skeletal muscle into the blood and are carried to the liver.
• Much more lactate than pyruvate is transported out because the high NADH/NAD+ ratio in contracting skeletal muscle favors the conversion of pyruvate into lactate.
• Lactate in the liver is oxidized to pyruvate, a reaction favored by the low NADH/NAD+ ratio in the cytosol of liver cells.
• Pyruvate in the liver is converted into glucose by the gluconeogenic pathway.
• Glucose then enters the blood and is taken up by skeletal muscle.
The Cori cycle

The interplay between glycolysis and gluconeogenesis

Lactate dehydrogenase isozymes
•Tetramer of 2 kinds of 35kd subunits (H, M).H mainly in heartM mainly in skeletal muscle and liver
•5 types of tetramers:
H4
H3M1
H2M2
H1M3
M4
Type Composition Affinity to substrate Reaction
12
3
45
High
Low
Lactate pyrovate
Lactate pyrovate

Soled tumors grow
hypoxia
Hypoxia inducible factor (HIF-1)
High expression of:• glycolytic enzymes • and GLUT1 and GLUT2
Increased tumor aggressiveness and poor prognosis
Increased new tumors growth by inducing the expression of signal molecules like vascular endothlial growth factor VEGF
Increased vascularization
CANCER AND GLYCOLYSIS

Proteins in glucose metabolism encoded by genes regulated by HIF-1:
•GLUT1GLUT3•Hexokinase•Phosphofructokinase•Aldolase•Glyceraldehyde 3-phosphate dehydrogenase•Phosphoglycerate kinase•Enolase•Pyruvate kinase•Lactate dehydrogenase