comparative analysis of the plasmodium falciparum histidine-rich proteins hrp-i, hrp-ii and hrp-iii...
TRANSCRIPT

Parasitologyhttp://journals.cambridge.org/PAR
Additional services for Parasitology:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
Comparative analysis of the Plasmodium falciparum histidinerich proteins HRPI, HRPII and HRPIII in malaria parasites of diverse origin
E. P. Rock, K. Marsh, A. J. Saul, T. E. Wellems, Diane W. Taylor, W. L. Maloy and R. J. Howard
Parasitology / Volume 95 / Issue 02 / October 1987, pp 209 227DOI: 10.1017/S0031182000057681, Published online: 06 April 2009
Link to this article: http://journals.cambridge.org/abstract_S0031182000057681
How to cite this article:E. P. Rock, K. Marsh, A. J. Saul, T. E. Wellems, Diane W. Taylor, W. L. Maloy and R. J. Howard (1987). Comparative analysis of the Plasmodium falciparum histidinerich proteins HRPI, HRPII and HRPIII in malaria parasites of diverse origin. Parasitology,95, pp 209227 doi:10.1017/S0031182000057681
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/PAR, IP address: 139.184.30.131 on 12 Sep 2012

Parasitology (1987), 95, 209-227 209
With 6 figures in the text
Comparative analysis of the Plasmodium falciparum histidine-richproteins HRP-I, HRP-II and HRP-III in malaria parasites of
diverse origin
E. P. ROCK1, K. MARSH2, A. J. SAUL5, T. E. WELLEMS5,DIANE W. TAYLOR3, W. L. MALOY4 and R. J. HOWARD5*1 Howard Hughes Medical Institute, Bethesda, Maryland 20814, USA
2 Medical Research Council Laboratories, F ajar a, The Gambia3 Department of Biology, Georgetown University, Washington, D.C. 20057, USA
4 Laboratory of Immunogenetics, National Institute of Allergy and Infectious Disease,National Institutes of Health
5 Laboratory of Parasitic Diseases, National Institute of Allergy and InfectiousDiseases, National Institutes of Health, Bethesda, Maryland 20892, USA
(Accepted 12 March 1987)
SUMMARYPlasmodium falciparum-inkcted erythrocytes (IRBC) synthesize 3 histidine-rich proteins:
HRP-I or the knob-associated HRP, HRP-II and HRP-III or SHARP. In order to dis-tinguish these proteins immunochemically we prepared monoclonal antibodies which reactwith HRP-I, HRP-II and HRP-III, and rabbit antisera against synthetic peptides derivedfrom the HRP-II and HRP-III sequences. A comparative analysis of diverse P. falciparumparasites was made using these antibodies and immunoprecipitation or Western blotting.HRP-I (Mr 80000-115000) was identified in all knob-positive P. falciparum parasites inclu-ding isolates examined directly from Gambian patients. However, this protein was of lowerabundance in these isolates and in 6 knob-positive, culture-adapted parasites compared toAolus monkey-adapted parasites or culture-adapted parasites studied previously. HRP-II(Mr 60000-105000) was identified in all P'. falciparum parasites regardless of knob-pheno-type, and was recovered from culture supernatants as a secreted water-soluble protein.Within IRBC, HRP-II was found as a complex of several closely spaced bands. Cell surfaceradio-iodination of IRBC from several isolates and immunoprecipitation with a rabbitantiserum against the HRP-II repeat sequence identified HRP-II as a surface-exposedprotein. Like HRP-I, the abundance of HRP-II was lower in the Gambian isolates than withAolus monkey-adapted parasites studied earlier. Neither HRP-I nor HRP-II were identifiedin a knob-positive isolate of P. malariae collected from a Gambian patient. Analogues ofthese HRP were also absent from asexual parasites of diverse primate and murine malariaspecies screened with this panel of antibodies. HRP-III (MT 40000-55000) was distinguishedby its lower apparent size and by specific reaction with rabbit antibody against its 5-merrepeat sequence. HRP-III was of lowest abundance compared with the other two HRP.These antibody reagents and distinguishing properties should prove useful in studies on theseparate functions of the 3 P. falciparum HRP.
INTRODUCTION
Histidine generally represents a small proportion of protein amino acids, for example,an average value of 0-7 mole% in Escherichia coli proteins (Schulz & Schirmer, 1979).Histidine-rich proteins (HRP) are rare and when they occur they are of interest for the
* Reprint requests to: Dr Russell J. Howard, NIH, Building 5, Room 112, Bethesda, Maryland20892, USA.

210 B. P. BOCK AND OTHERS
particular structural and functional properties they may possess due to a high contentof imidazole groups. Asexual blood-stage parasites of the avian malaria, Plasmodiumlophurae, synthesize a remarkable protein containing 72 mole% histidine (Kilejian,1974) and tandemly repeated sequences of several contigious histidine residues (Ra-vetch, Feder, Pavlovec & Blobel, 1984; Howard, Raum, Maloy, Kao & Colligan, 1984).The P. lophurae HRP is located in cytoplasmic granules of unknown function (Kilejian,1974), and its biochemical function remains unknown.
At least 3 HRP are made by asexual blood-stage P. falciparum, denoted here asHRP-I, HRP-II and HRP-III . HRP-I (PfHRP-I, the knob-associated HRP) hasan Mr of 80000-110000 in differ-ent isolates (Kilejian, 1979; Hadley, Leech, Green,Daniel, Wahlgren, Miller & Howard, 1983; Leech, Barnwell, Aikawa, Miller & Howard,1984; Vernot-Hernandez & Heidrich, 1985; Rock, Saul, Taylor, Leech, Sherwood &Howard, manuscript in preparation) and is associated phenotypically with expressionof knob-like protrusions on the surface membrane of infected erythrocytes (IRBC)(Kilejian, 1979). A cDNA clone encoding the amino terminal half of HRP-I has beenshown to encode a histidine-rich domain containing 30 histidine residues within a spanof 50 amino acids (Kilejian, Sharma, Karoui & Naslund, 1986). Laboratory-derivedP. falciparum variants lacking knobs (K—) do not synthesize this HRP (Kilejian, 1979;Hadley et al. 1983; Leech et al. 1984). The correlation between expression of knobs onIRBC and synthesis of HRP-I, together with the localization of this molecule to purifiedmembranes (Vernot-Hernandez & Heidrich, 1984) or erythrocyte cytoskeletons fromIRBC (Leech et al. 1984), led to the concept that HRP-I plays a role in either thestructure or function of the knob protrusions. Recently, we used a monoclonal antibody(McAb) specific for HRP-I (McAb 89) and immunoelectron microscopy to localize thisprotein at the submembrane electron-dense material of knobs (Taylor, Parra, Stearns,Chapman, Aikawa, Uni, Panton & Howard, 1987). Human antibodies affinity purifiedon a cDNA expression clone encoding HRP-I have also been used to localize this proteinto knobs, particularly on the cytoplasmic side of the IRBC surface membrane(Culvenor, Langford, Crewther, Saint, Coppel, Kemp, Anders & Brown, 1987).
HRP-II (PfHRP-II) is expressed by both K + and K - IRBC (Leech et al. 1984;Howard et al. 1986). I t is released by K + IRBC as a soluble extracellular protein andis presumed to interact with host cells and/or proteins in plasma (Howard, Uni, Aikawa,Aley, Leech, Lew, Wellems, Rener & Taylor, 1986). Sequencing of genomic DNA hasshown that HRP-II contains 34% histidine, as well as relatively high contents ofalanine and aspartic acid (37 and 10%, respectively) (Wellems & Howard, 1986). HRP-II contains multiple contiguous tri- and hexapeptide repeats, predominantly AHH andAHHAAD, which comprise roughly 80% of the protein sequence. The nucleotidesequence of the two exons encoding HRP-II encodes a protein of MT ~ 35000, muchlower than its apparent MT of 60-80000 on SDS-polyacrylamide gel electrophoresis(Wellems & Howard, 1986).
HRP-III (PfHRP-III, SHARP) is a third HRP expressed by asexual blood-stageparasites. Sequence analysis of the gene encoding HRP-III in two different P. falci-parum isolates has been reported (Stahl et al. 1985; Wellems & Howard, 1986) and hasdemonstrated polymorphism in the repeats of the gene. HRP-III contains 28% his-tidine, 28% alanine and 12% asparagine. There is a high degree of sequence homologybetween HRP-III, and HRP-II, implying that these two proteins have evolved byduplication and divergence from a common ancestral gene (Wellems & Howard,1986).
We have prepared monoclonal antibodies (McAb) against HRP-I and HRP-II. We

P. falciparum histidine-rich proteins 211
also prepared rabbit antisera to peptides within the sequences of HRP-II or HRP-IIIwhich react specifically to immunoprecipitate the appropriate HRP. These reagents areused here to compare and distinguish the three HRP in various P. falciparum isolates,including two from Gambian malaria patients. The properties of each HRP are sum-marized and a property of HRP-II is described: exposure on the surface of intact IRBC.We also used these antibodies to screen for the presence of HRPs in a P. malariaesample from a Gambian patient and in diverse primate and murine malaria species.
MATERIALS AND METHODS
ParasitesThe derivation of the uncloned K + or K— Malayan Camp and K + or K— St Lucia
strains of P. falciparum has been described (Leech et al. 1984). These parasites wereobtained from infected Aotus monkeys and cryopreserved (Meryman & Hornblower,1972) at 10-50 % parasitaemia. A K -I- clone, C-5, derived from the FCR-3/FMG Africanstrain of P'. falciparum and a K— clone, C-6, from the same strain, were provided byDr T. J. Green (Green, Gadsden, Seed, Jacobs, Morhardt & Brackett, 1985) and grownin human erythrocytes (RBC). The K + P. falciparum clone 7G8, also grown in humanRBC, was derived from the It strain of Brazilian origin (Burkot, Williams & Schneider,1984). Derivation, culture and cryopreservation of the four culture-adapted P. falci-parum strains of diverse geographic origin have been described (Udeinya, Graves,Carter, Aikawa & Miller, 1983). The country of origin of these isolates was as follows:T2 (T) from Thailand; Nl (N) from Nigeria; L from Liberia; Kl (K) from Kenya(Udeinya et al. 1983). Clone 3D7 was derived by limiting dilution of isolate NF54 fromthe Netherlands (Walliker, Carter, Quakyi, Wellems, McCutchan & Szarfmann, 1986).In vitro culture was in human O-positive erythrocytes with AB serum. Parasitized RBCused for antigen analysis were simultaneously tested in the in vitro cytoadherence assayfor binding to amelanotic melanoma cells (Roberts, Sherwood, Spitalnik, Panton, How-ard, Dixit, Frazier, Miller & Ginsburg, 1985). As originally reported (Udeinya et al.1983) infected cells from T, K, L and N isolates bound very poorly or not at all to themelanoma cell line. Electron microscopy was used to confirm the knob phonotype(Hadley et al. 1983; Rock et al. manuscript in preparation). Primary isolates ofP. falciparum-infected human blood (isolates 52, 89, 104 and 133 of 5, 6, 1 and 2%parasitaemia, respectively) and an isolate of P. malariae (0-5% parasitaemia) werecollected from naturally infected Gambian children with acute malaria, who wereattending the oupatient clinic of the Medical Research Council Laboratories in Fajara.Blood was collected by venipuncture after parental consent; chloroquine was admin-istered after bleeding. Prior to in vitro culture these samples were passed through acolumn of CF-11 cellulose (Whatman Ltd, Maidstone, Kent) to remove leucocytes(Grant & Fulton, 1957).
Culture, metabolic labelling and detergent extractionThawing (Meryman & Hornblower, 1972), synchronization (Lambros & Vanderberg,
1979) and culture (Trager & Jensen, 1976) of cryopreserved IRBC or culture-adaptedparasites were performed by standard protocols. The thawing procedure lysed leucocytesand the majority of trophozoite- or schizont-infected RGB, yielding parasitaemias of5-30% consisting entirely of ring-stage parasites. When culture supernatants were to becollected and analysed, the thawed RBC were immediately fractionated on a Percoll

212 E. P. BOCK AND OTHEBS
gradient containing sorbitol (Aley, Sherwood & Howard, 1984) such that debris fromleucocytes or mature parasitized RBC was removed. The purified, ring-infected RBCwere cultured in RPMI-1640 containing 10% of the normal concentration of histidine,leueine, isoleucine or methionine. The medium was supplemented with 10% human ABserum (for culture of clone 7G8 or isolates in human RBC) or 10% horse serum (forculture of parasites in Aotus RBC), 30 IDM HEPES, 027% sodium bicarbonate, 10 fig/ml hypoxanthine (Sigma), 2 mg/ml D-glucose and 50 /^g/ml gentamicin (ScheringCorp., Kenilworth, NJ). Radioactive amino acids were added at 100/iCi/ml to culturemedium deficient in the appropriate amino acid. In radio-isotope labelling experimentsthe concentration of parasitized erythrocytes was 2-3 x 107 IRBC/ml, yielding haem-atocrits of 0-6-6%. The cells were harvested after 20-26 h when the majority ofparasites were mature trophozoites. The supernatant fractions from high-speed cen-trifugation of biosynthetically labelled cultures and Triton X-100 or SDS extracts ofwashed cells were collected as described previously (Howard et al. 1986).
Cell surface radioiodination^4o<M5-infected RBC containing late-trophozoites and early schizonts were purified on
Percoll/sorbitol gradients and cell-surface radiolabelled by the Na125I-H2O2-lacto-peroxidase method (Aley, Sherwood & Howard, 1984). The labelled cells were extractedsequentially with Triton X-100 and SDS (Howard, Kao & Barnwell, 1984).
Production of monoclonal antibodies to HRP-I and HRP-IIMcAb 89, specific for HRP-I (Taylor et al. 1987) and McAb 87, specific for HRP-II
(Howard et al. 1986), were prepared as mouse ascites. Two other McAb which specificallyimmunoprecipitate HRP-II , 1D6 and 2G12 were produced in fusions using spleen cellsfrom mice immunized with cytoskeletal preparations from K + PRBC. Ouchterlonyanalysis of culture supernatants and immunoelectrophoresis of ascites showed that eachof these McAbs was homogeneous and of the IgGyj isotype. Control McAb IA2 (IgG)specific for the major surface glycoprotein of P. falciparum schizonts was generouslyprovided by Dr F. Klotz of the National Institutes of Health.
AntiseraSerum was collected from a Gambian patient (20-year-old female) 3 weeks after drug-
cure of acute P. falciparum malaria. Rabbit antiserum 6211 against the HRP ofP. lophurae was prepared by immunization with 0-3 mg of protein (Howard et al. 1984).The protein was emulsified in Freund's Incomplete Adjuvant and administered intra-muscularly on day 0, 6 weeks and 6 months. Serum was collected 1 month after thelast injection. Rabbit antiserum no. MC 1 against HRP-I was prepared by immuni-zation with HRP-I cut from a preparative gel (Rock et al. 1986). Rabbit antiseraagainst the peptides AHHAHHAADAHHAAD (a HRP-II) and GAHHDDAHH-DGAHHD (a HRP-III) were generated as described (Wellems, Walliker, Smith, doRosario, Maloy, Howard, Carter & McCutchan, 1987).
Immunofluorescence, immunoprecipitation and Western blot analysisThe binding of monoclonal antibodies or rabbit serum antibodies to acetone-fixed
infected cells was examined by indirect immunofluorescence (Howard et al. 1986).

P. falciparum histidine-rich proteins 213
Immunoprecipitation. SDS-PAGE and fluorographj' were as previously performed(Howard et al. 1986). IRBC or uninfected cells were washed twice in PBS and the pelletsolubilized directly in SDS-sample buffer (Howard & Barnwell, 1984) (100/tl/2x 107
total RBC). Samples were stored at —90 °C. Electrophoretic transfer to nitrocellulosewas performed by the method of Towbin, Staehlin & Gordon (1979).
Transfer conditions, quenching, incubation and washing have previously been des-cribed (Howard et al. 1986) except that McAb 89 was purified on a Protein A columnand radio-iodinated (6/^Ci 125I//*g antibody) by the Iodogen method (Fraker & Speck,1978). Labelled McAb 89 was stored as a stock solution of 40 fig/ml in 1 % bovine serumalbumin in PBS plus 0-05 % sodium azide and incubated with blots at a 1:800 dilutionof the stock (300000 c.p.m./ml). Control McAb 2D2C5 (IgG) was incubated with blotsat a dilution of 1:100 of ascites and probed by incubation with a 1:1000 dilution ofrabbit anti-mouse IgG followed by 125I-Protein A (Taylor et al. 1987).
RESULTS
Identification of HRP-I and HRP-II in Aotus-adapted parasites by biosyntheticlabelling and detergent extraction
K + and K— IRBC of Malayan Camp parasites in Aotus erythrocytes were culturedwith different radiolabelled amino acids from the early ring-stage to late trophozoitesand early schizonts. Washed IRBC were solubilized sequentially with 1 % Triton X-100and 2% SDS and the detergent extracts compared by SDS-PAGE and fluorography.The knob-associated HRP (HRP-I) was identified as a protein of Mr 92000 that wasstrongly radiolabelled by uptake of [3H]histidine with K + parasites but not with K —parasites (Leech et al. 1984). This protein was not labelled by uptake of [3H]isoleucinenor was it solubilized by Triton X-100 in isotonic solution. A second HRP (HRP-II),at Mr 72000, was strongly radiolabelled by [3H]histidine but only weakly by[3H]leucine and not at all by [3H]isoleucine. In contrast to HRP-I, this HRP was pres-ent with both K + and K— Malayan Camp parasites. In some experiments, HRP-IIwas completely extracted with Triton X-100, while in others up to 30% of the totalHRP-II associated with washed IRBC remained insoluble in this detergent. HRP-IIof Malayan Camp Strain parasites characteristically migrated as a broad band at MT
72000 with a sharp trailing edge and diffuse leading edge extending to approximatelyMt 58000. With St Lucia parasites HRP-II was a broad band at Mr 79000-74000.Table 1 summarizes these results.
Detection of HRP-I in culture-adapted strainsFour knob-positive culture-adapted P. falciparum strains of geographically diverse
origin were radiolabelled with [3H]histidine and [3H]isoleucine and their HRP analysedas for the >4o<ws-adapted parasites. Bands comparable to HRP-I could not be identifiedunequivocally with any of these culture-adapted parasites. There was no band pref-erentially labelled with [3H]histidine in the Triton X-100 insoluble fraction that was notalso in the Triton X-100 extract. McAb 89 was used in an attempt to immunoprecipitateHRP-I from the biosynthetically labelled cell fractions of the four culture-adaptedstrains. We failed to identify any specifically immunoprecipitated [3H]histidine- or[3H]isoleucine-labelled antigens using McAb 89 and the Triton X-100 or SDS-detergentextracts. The same McAb 89 ascites reacted with these parasites by immunofluorescenceand Western blotting (Rock et al. 1986) and specifically immunoprecipitated HRP-Ifrom [3H]histidine-labelled extracts of K + Malayan Camp and K + St Lucia parasites

214 E. P. BOCK ASD OTHERS
Table 1. Molecular weights (Mr) of HEP-I and HRP-II in diverse Plasmodiumfalciparum parasites
ParasiteMalayan Camp
St Lucia
FMGFVOLKNT7G8 cloneFCR-3 clone, C5FCR-3 clone, C6
Host*Aotus
AotusAotusAotusAotusHuRBCHuRBCHuRBCHuRBCHuRBCHuRBCHuRBC
PhenotypefK 4 B +K -KH-B +K -K + B-l-K + B-l-K - f B -K + B -K + B -K + B -K-t-B-K+B +K -
M r (>HRP-I
92§—
108—
N.D.N.D.
92104104
N.D.N.D.
92§925
< 10"3)tHRP-II
7272797974748897
104104837263
* Parasites were either in Aotus trivirgatus RBC (Aotus) which were cultured in vitro for less than30 h, or in human RBC (HuRBC) adapted to long-term culture.
f K + B +, knob-positive and eytoadherence/binding positive; K + B —, knob-positive and bindingnegative; K —, knob-negative and binding negative.
% Mr as determined by SDS-PAGE on 5-10% acrylamide gradient gels under reducing conditions.For HRP-I, which migrates as a single broad band, the MT is calculated for the centre of the band.For HRP-II, which migrates as a single sharp band from culture supernatants, or as a multiplet ofbands extending from this band to lower Mr with P-RBC, the Mr corresponds to the slowest-movingHRP-II band. These MT values are different if the samples are electrophoresed on gels of differentacrylamide or cross-linker concentrations.
§ Based on an Mr of 92500 for the marker protein phosphorylase a, we originally calculated Mr89000 for HRP-I of these parasites (Hadley et al. 1983). Using the revised Mr of 97400 for phos-phorylase a, the Mr of this HRP-I is now calculated as 92000.
N.D., Not determined.
from Aotus monkeys. The Mr of HRP-I in these parasites, as identified by Westernblotting with McAb 89, a.re given in Table 1. Thus, although HRP-I is present in eachof the four K + culture adapted parasites, it cannot be identified using [3H]histidine-labelled extracts. HRP-I may represent a much smaller proportion of total parasiteproteins compared with the Aotus-adapted parasites or could have a lower content ofhistidine.
Detection of HRP-I in parasite isolates collected directly from Gambian patients
Isolates of knob-positive P. malariae and P. falciparum were collected directly fromGambian patients, cultured in vitro for less than 24 h, and mature IRBC purified onPercoll density gradients. The IRBC (greater than 85 % parasitaemia) were solubilizedin SDS-sample buffer, electrophoresed, transferred to nitrocellulose, and incubated withantibody reagents specific for HRP-I (McAb 89, Rabbit aP l HRP, and Rabbit a HRP-I), followed by 125I-Protein A.
No analogues of HRP-I were detected in P. malariae IRBC under conditions whichgave strong reactions with HRP-I of two K 4- Aotus-a.da.pted parasites (Malayan Camp,MC and St Lucia SL) (Fig. 1 shows results for McAb 89). In contrast, a broad band ofMv 112000 was detected in two P'. falciparum isolates from Gambian patients (133 and104) using the same three antibodies (Fig. 1). The specificity of this reaction was

P. falciparum histidine-rich proteins 215
108-
92-
r(X 1 0 "
ooa<r
200
97-4
68
112
— 43 i
_ 25-7 I
— 18-4 • f
Fig. 1. Probing Plasmodium falciparum isolates and a P. malariae isolate from Gambianpatients for HRP-I by Western blotting with McAb89 and rabbit antisera to HRP. Ring-stage P. falciparum parasites were cryopreserved, thawed and cultured for less than 30 huntil parasites were mature trophozoites. P. malariae-infected blood (predominantly ringsand young trophozoites) was cultured 20 h until the majority of parasites were maturetrophozoites and schizonts. IRBC were purified (> 90% parasitaemia) on Percoll/sorbitolgradients and sequentially extracted with Triton X-100 and SDS. Extracts were mixed withequal volumes of double-strength electrophoresis sample buffer, electrophoresed on 5-10%aerylamide gradient gels, transferred electrophoretically to nitrocellulose and probed withMcAb 80, rabbit antisera to HRP-I or PIHRP followed by 125I-Protein A. Results are shownfor SDS extracts probed with McAb 89. Left panel: positive and negative controls of St Lucia(SL) K + , Malayan Camp (MC) K-t- and K— P. falciparum parasites in Aotus RBC and anegative control of uninfected human RBC (H RBC) compared with P. malariae (P mal).Right panel: H RBC compared with IRBC of isolates 133 and 104 of P. falciparum. TritonX-100 extracts were negative with all antibody probes. Rabbit antisera to HRP-I andPIHRP gave identical results to McAb 89 with the SDS extracts. Molecular weight standardsand HRP are indicated by dashes.
demonstrated by the fact that uninfected RBC did not react with these antibodies, nordid K— IRBC from an ^4o<tts-adapted P. falciparum strain (Fig. 1). Six P. falciparumisolates collected directly from patients, including 133 and 104, were biosyntheticallylabelled, sequentially extracted with Triton X-100 and SDS, and immunoprecipitatedwith McAb 89. Although HRP-I could be identified in these isolates by immunofluor-escence (not shown) and Western blotting using McAb 89 (Fig. 1), it could not beidentified by immunoprecipitation of [3H]histidine-labelled extracts.
Effect of urea on electrophoretic mobility of HRP-I
We tested whether addition of urea to the 5% SDS-sample buffer used in SDS-PAGE would affect the electrophoretic mobility of HRP-I and HRP-II. With the

216 E. P. ROCK AND OTHERS
a be
Mr M, (X10-
• 200
•116-25
• 97-4
66-2
•45
B0B
Fig. 2. SDS-PAGE of HRP from K + Malayan Camp strain Plasmodium faldparum in thepresence of urea. Infected blood (38 % parasitaemia) was radio-isotope labelled with [3H]-histidine and extracted sequentially with Triton X-100 and SDS. The SDS extract wasmixed with an equal volume of double strength SDS-sample buffer (containing 10% SDS)and crystalline urea added to portions of this material ((a) no urea; (b) 2 M urea; (c) 8 M urea).After 10 min at 60 °C the samples were analysed by SDS-PAGE and fluorography. Molecularweight standards are indicated by dashes. HRP-II (MT 72000) and HRP-I (MT 92000-114000) are marked by asterisks. B0B, bromphenol blue.
addition of 2 M urea, the mobility of HRP-I decreased to an apparent MT of 105000(Fig. 2). The mobility decreased further to an apparent Mr of 114000 with sample buffercontaining 8 M urea. In contrast, the mobility of HRP-II was not affected by theaddition of 8 M urea to the sample buffer.

P. falciparum histidine-rich proteins 217
Identification of HRP-II in Aotus-adapted and cultured parasitesHRP-II has been identified in ^4o£ws-adapted P. falciparum parasites by biosynthetic
labelling with [3H]histidine and [3H]isoleucine and examination of culture supernatantsfor the presence of a single sharp band that is labelled preferentially by [3H]histidine(Howard et al. 1986). The same experiment was performed with the four K + culture-adapted parasites that were screened for expression of HRP-I. For each parasite, awater-soluble extracellular protein was identified that was labelled by [3H]histidineuptake but not by [3H]isoleucine. Table 1 summarizes the MT of HRP-II for theseparasites.
HRP-II can also be identified by specific reaction with McAbs: McAb 87, an IgM(Howard et al. 1986), or the IgG McAbs 2G12 and 1D6. IRBC of diverse parasites werelabelled by biosynthetic uptake of [3H]histidine and these antibodies reacted byimmunoprecipitation with the supernatant fraction from high-speed centrifugation,the 1 % Triton X-100 extract of washed cells and 2% extract of Triton X-100 insolublematerial.
Fig. 3 shows the results for three strains of ^4o^s-adapted parasites and McAb 87.Immunoprecipitation of culture supernatants revealed one major [3H]histidine-labelledantigen which comigrated with HRP-II. This extracellular antigen was a single sharpband of MT 79000 with K + St Lucia parasites or Mr 72000 with K + or K - MalayanCamp parasites. [3H]histidine-labelled antigens of the same size with each strain (Mr
79000 or 72000) were also the major components specifically immunoprecipitated byMcAb 87 from the Triton X-100 and SDS-extracts (Fig. 3). In the detergent extractsthe major antigen was a broad band with a sharp trailing edge and diffuse leading edge.Identical results were obtained with IgG McAb's 2D12 and 1D6.
Other minor antigens were also specifically immunoprecipitated from [3H]histidine-labelled material by these McAbs: a band of MT 85000 in Triton X-100 extracts fromeach of these three parasites; in SDS extracts, a doublet at Mr 138000 and 127000 forK + St Lucia parasites and a doublet at Mr 121000 and 116000 for K + orK— MalayanCamp parasites. Another minor [3H]histidine-labelled antigen of MT 56000 wasimmunoprecipitated from SDS extracts of the Malayan Camp parasites (Fig. 3). Theseminor antigens were not labelled with [3H]isoleucine (not shown).
We also examined reactivity of the McAb's with P'. falciparum strains of diversegeographical origin by Western blotting. Fig. 4 shows the results for McAb 87. Thethree McAb's did not react with antigens of uninfected Aotus RBC. The antigensspecifically recognized by this McAb with K + and K — IRBC of Malayan Camp strainparasites in Aotus RBC were identical: four bands of MT 72000, 63000, 58000 and49000. Longer film exposure revealed bands at MT > 300000 that were weakly reactivewith McAb 87. K + and K - IRBC of cloned parasites from the FCR-3/FMG strain ofP. falciparum, cultured in human RBC, also gave identical patterns of reactivity: majorantigens of MT 72000 and 63000 with a smear of radioactivity extending to MT 49000.K + IRBC of the St Lucia strain in Aotus RBC showed 2 bands reactive with McAb 87&tMr 79000 and 74000. K + IRBC of the 7G8 clone in human RBC had only 1 reactiveband at Mr 83000. All three McAb's also reacted by Western blotting with antigens inthe culture supernatant. Fig. 4 shows that a doublet, Mt 72000 and Mt 63000, reactedin the supernatant fraction of K— FCR-3 parasites, while only a MT 63000 band wasreactive in the supernatant fraction from K + FCR-3 parasites. Thus, by Westernblotting, McAbs 87, 2G12, and 1D6 define a group of antigens of diverse MT in the range

218 E. P. ROCK AND OTHERS
a b c
r{x i c r 3 )
B0B — - B<t>B
Culture supplement TX-100 extract SDS extract
Fig. 3. Identification of HRP-II by immunoprecipitation of [3H]histidine-labelled proteinswith McAb 87 and analysis by SDS-PAGE and fluorography. Lanes b-d show results forMcAb 87, Lane a shows results for normal mouse serum. K+ St Lucia (Lane b), K— MalayanCamp (Lane c) or K+ Malayan Camp (Lane d) parasites (parasitaemias respectively 18, 40and 35%) were cultured from rings to the late trophozoite stage with [3H]histidine. Im-munoprecipitation was performed with culture supernatants, 1% Triton X-100 extracts ofwashed cells, or 1 % SDS extracts of Triton X-100-insoluble material. Identical results wereobtained with McAbs 1D6 and 2G12. Antigens discussed in the text are labelled with anasterisk. Molecular weight markers are indicated by dashes. B0B, bromphenol blue.
50000-80000 which occur in both K + and K— parasites, and which have the sameMT for each parasite as HRP-II.
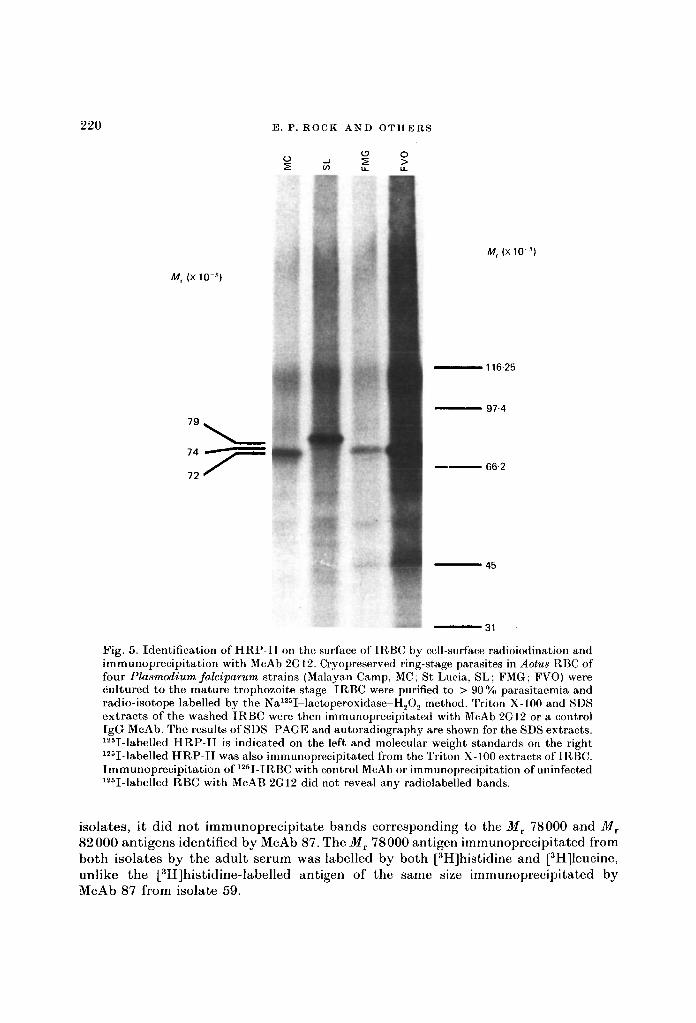
Cell-surface radio-iodination of HRP-IITo determine whether HRP-II is exposed on the cell surface purified IRBC from four
different ^lotes-adapted strains of P. falciparum were radio-iodinated by the Na125I-H202-lactoperoxidase method and detergent extracts immunoprecipitated with McAbsID6 and 2G12. Fig. 5 shows the results for McAb 2G12. With each parasite strain,125I-labelled antigens corresponding in Mr to [3H]histidine-labelled HRP-II werespecifically immunoprecipitated from the SDS extracts. The Mr of 125I-labelledHRP-I I was MT 72000 with Malayan Camp strain parasites, MT 79000 with St Luciastrain and Mt 74000 with FMG and FVO strains of P'. falciparum (Fig. 5).

P. falciparum histidine-rich proteins 219
M, (X1CT
200 .
c'c
03
3 co
CO
d:o
COir
ou.
Wr(X10'3)
*72,*63-*58 »•49'
83*• 79*'74*
Cells
Fig. 4. Identification of HRP-II by Western blotting and probing with McAb 87. WashedRBC from cultures containing late-trophozoite and early-schizont stages of K + or K —Malayan Camp, or K+ St Lucia strain parasites (in Aotus RBC; parasitaemias in order 30,45 and 15%),K+ clone 7G8, K + clone C-5 (labelled K + FCR-3)andK- clone C-6 (labelledK— FCR-3) (in human RBC; parasitaemias in order, 10, 8 and 22%) or uninfected AotusRBC, were solubilized in 5% SDS-sample buffer, fractionated by SDS-PAGE and electro-phoretically transferred to nitrocellulose. Supernatants from culture were also analysed(labelled 'Supt'). After incubation with McAb 87 and rabbit-anti-mouse IgM, immune com-plexes were identified by reactivity with 125I-Protein A and autoradiography. Identicalresults were obtained with McAbs 2G12 and 1D6 followed by 125I-Protein A. Antigens dis-cussed in the text are identified by an asterisk and their molecular weights indicated.
Identification of HRP-II with P. falciparum isolates from Gambian patients
Two isolates of ring-stage P. falciparum parasites collected directly from Gambianpatients were cultured in vitro with [3H]histidine or [3H]leucine. At the late trophozoitestage (26 h culture) the washed blood was solubilized in 1% Triton X-100 and theextracted proteins compared by SDS-PAGE. The patterns of [3H]leucine-labelled pro-teins of isolates 59 and 82 were almost identical, the only obvious differences being inquantitatively minor proteins of Mr ^ 200000. However, each isolate had a quanti-tatively major [3H]histidine-labelled band that appeared unique to the particularisolate: Mr 78000 with isolate 59 versus MT 82000 with isolate 82. There were noanalogous [3H]leucine-labelled proteins of different MT for each isolate.
The Mr 78000 and 82000 [3H]histidine-labelled bands were shown to be HRP-II bytheir specific immunoprecipitation with McAb 87 (Fig. 6). McAb 87 failed to irnmuno-precipitate radiolabelled antigens of this size from [3H]leucine-labelled extracts of thesame parasites. Immunoprecipitation was also performed with a serum from an adultGambian who has presumably had numerous infections with P. falciparum (Lane a,Fig. 6). Although the adult serum immunoprecipitated several antigens from both
220 E. P. ROCK AND OTHERS
M, (XKT 1 )
M, (X1(T
79
72
• 116-25
• 97-4
• 66-2
•45
•31
Fig. 5. Identification of HRP-II on the surface of TRBC by cell-surface radioiodination andimmunoprecipitation with McAb 2G12. Cryopreserved ring-stage parasites in Aotus RBC offour Plasmodium falciparum strains (Malayan Camp, MC; St Lucia, SL; FMG; FVO) werecultured to the mature trophozoite stage. IRBC were purified to > 90% parasitaemia andradio-isotope labelled by the Na125I-lactoperoxidase-H202 method. Triton X-100 and SDSextracts of the washed IRBC were then immunoprecipitated with McAb 2GI2 or a controlIgG McAb. The results of SDS PAGE and autoradiography are shown for the SDS extracts.125I-labelled HRP-II is indicated on the left and molecular weight standards on the right125I-labelled HRP-II was also immunoprecipitated from the Triton X-100 extracts of IRBC.Immunoprecipitation of 125I-IRBC with control McAb or immunoprecipitation of uninfected125I-labelled RBC with McAB 2G12 did not reveal any radiolabelled bands.
isolates, it did not immunoprecipitate bands corresponding to the Mr 78000 and MT
82000 antigens identified by McAb 87. The MT 78000 antigen immunoprecipitated fromboth isolates by the adult serum was labelled by both [3H]histidine and [3H]leucine,unlike the [3H]histidine-labelled antigen of the same size immunoprecipitated byMcAb 87 from isolate 59.

P. falciparum histidine-rich proteins 221
H]histidine
M, (X10"3)
200
116-25 —
[3H]leucine
t ~ \
a b c c b a
— •78 — I
45
«*»•
45
Isolate: 82 59 82 59
Fig. 6. Identification of HRP-IT in Plasmodium falciparum isolates radio-isotope labelleddirectly after collection from Gambian patients. Isolates 59 and 82 were collected as ringstages from infected Gambian children and, after removal of leucocytes, cultured with [3H]-histidine or [3HJleucine until parasites matured to late-trophozoites and early schizonts.The washed cells were extracted with Triton X-100 and the extracts immunoprecipitatedusing Protein A-Sepharose as solid-phase adsorbent. Immunoprecipitation was with serumfrom a Gambian adult (Lane a), normal mouse serum (Lane b) or McAb 87 (Lane c). Proteinsdiscussed in the text are labelled with an asterisk. Molecular weight standards are indicatedby dashes.
Identification of HRP-III in laboratory-adapted parasitesRabbit antiserum a HRP-III has been prepared against the peptide GAHHDDA-
HHDGAHHD derived from the HRP-III sequence. This peptide corresponds to partof the HRP-III sequence which contains pentapeptide repeats lacking from thesequence of HRP-II (Wellems & Howard, 1986). Rabbit antiserum a HRP-II has alsobeen prepared against the peptide AHH(AHHAAD)2, derived from the sequence ofHRP-II. These antisera have been used to distinguish and characterize HRP-II andHRP-III.
Asynchronous cultures of several P. falciparum parasites were biosynthetically radio-labelled for 24 h by addition of [3H]histidine, [3H]alanine or [3H]isoleucine and theculture supernatant, Triton X-100 extract and SDS extract of Triton X-100-insolublematerial immunoprecipitated. With a Triton X-100 extract of clone 7G8, the rabbita HRP-III serum immunoprecipitated a single [3H]histidine-labelled band, MT 41000,that was not immunoprecipitated by control rabbit sera. This band was also stronglylabelled by uptake of [3H]alanine but was not detected after immunoprecipitation of[3H]isoleucine-labelled extracts. The rabbit a HRP-II serum also failed to immuno-

222 E. P. ROCK AND OTHERS
precipitate this .Mr41000 band, although it did immunoprecipitate an Mr 83000[3H]histidine-labelled band which corresponds in size with HRP-II of the 7G8 clone.Similar results were obtained with clone 3D7 in human RBC and the Malayan Campstrain in Aotus RBC. Rabbit a HRP-III serum specifically immunoprecipitated a[3H]histidine- or [3H]alanine-labelled band of Mr 51000 from 3D7 and MT 42000 fromMalayan Camp parasites. Rabbit a HRP-II serum specifically immunoprecipitatedonly the corresponding HRP-II proteins of Mr 80000 and 72000 from 3D7 and MalayanCamp parasites. None of these bands were labelled by uptake of [3H]isoleucine. Thus,the two antisera raised against peptides that are unique to HRP-II or HRP-III reactspecifically with the corresponding HRP.
The amount of [3H]histidine-labelled HRP-III was significantly less than the amountof HRP-II with all P. falciparum parasites we examined. This difference was evidentfrom comparison of the level of 3H radioactivity at the corresponding Mr withoutimmunoprecipitation, as well as comparison of the radioactivity in these two HRP afterimmunoprecipitation with each specific rabbit antiserum. With the above-mentionedthree parasites HRP-III was immunoprecipitated by the rabbit a HRP-III serum as asingle sharp band present predominantly in the Triton X-100 extract of washed P-RBC,but also in low levels in the culture supernatant and SDS extract of Triton X-100-insoluble material. Since cultures were asynchronous, the extracellular HRP-III couldhave arisen by low level secretion from intact infected erythrocytes or release at thetime of cell rupture and merozoite reinvasion. The rabbit a HRP-II serum immuno-precipitated a single sharp band from culture supernatants, and a smear of radioactivityextending from the major HRP-II band to bands of faster mobility (roughly Mr 5000-10000 lower in apparent size) with the Triton X-100 extracts. With SDS extracts therabbit a HRP-I I serum immunoprecipitated doublets or triplets of bands correspondingto the slowest moving HRP-II band and bands of slightly faster mobility.
Comparison of these results with the specific rabbit antisera with those obtainedusing McAb selected for reactivity with HRP-II revealed cross-reactivity with all theMcAb's. McAb 2G12 immunoprecipitated HRP-II at Mt 83000 for 7G8 parasites, asmear of radioactivity in several bands extending from HRP-II to HRP-III, and adistinct HRP-III band at Mr 41000. Each McAb which immunoprecipitated HRP-IIalso immunoprecipitated HRP-III, although it should be noted that the proportion oftotal 3H radioactivity immunoprecipitated in HRP-III was less than 5 % of the total.Similar results were obtained with clone 3D7 and the Malayan Camp strain ofP. falciparum.
DISCUSSION
In order to study the biological and immunological relevance of the three HRP ofP. falciparum it is essential to have criteria which distinguish them particularly sincethey are all expressed simultaneously by some parasites and since the size ranges ofH R P I and HRP-II overlap on SDS-PAGE (Mt 80000-115000 and Mt 60000-105000respectively). Table 2 summarizes the specificities of the panel of antibody reagentsshown here to distinguish HRP-I and HRP-II, together with a variety of additionalcriteria for their identification. HRP-I can be identified unambiguously by its reactionwith several antibody reagents. McAb 89 reacts specifically with the HRP on bothimmunoprecipitation and Western blotting. Rabbit antiserum against HRP-I cut froma preparative gel (Rabbit MCI), immunoprecipitates both HRP-I and HRP-II butreacts by immunoblotting only with HRP-I. Rabbit antiserum 6211, raised against

P. falciparum histidine-rich proteins 223
Property
Immunoreactivity*McAbs87, 2G12 and 1D6McAb 89Rabbit a-AHH(AHHAAD)2Rabbit a. HRP-IRabbit a-Plophurae HRP
Amino acid compositionUptake of [3H]histidineUptake of [3H]isoleucine
Mr (x 10"3)tRelationship to knobsSubcellular location
Release from infected cellsas a soluble protein
Initiation of synthesis
Effect of urea an SDS-PAGEmobility
Table 2. Comparison of HRP-I and HRP-II
HRP-I HRP-II
+
80-110K+ onlyElectron-dense materialunder knobs, aggregatesin host RBC cytoplasm
Early rings
65-85K+ a n d K -Parasite cytoplasm,concentrated' packets' in hostRBC cytoplasm
Early tomid-trophozoites
* Immunoreactivity by immunoprecipitation and Western blotting. Reactivities shown in par-entheses show results of Western blotting different from those of immunoprecipitation.
f Apparent Mr on SDS-PAGE (reducing conditions) after heating sample for 10 min at 60 °C with5% (w/v) SDS and 5% (v/v) 2-mercaptoethanol in electrophoresis sample buffer.
P. lophurae HRP (Pl-HRP), is similar to MCI in that it immunoprecipitates bothHRP-I and HRP-II but binds only HRP-I on immunoblots. Both surfaceimmunofluorescence assays and immunoelectronmicroscopy using McAb 89 havedemonstrated that the appropriate epitope of HRP-I is not exposed at the cell surface(Taylor et al. 1987). In addition, neither McAb 89 nor MCI have an effect on in vitrocy toadheren ce.
Our results show considerable variation in the levels of HRP-I synthesis amongP. falciparum isolates. In ^otos-adapted Malayan Camp and St Lucia strain parasites(Leech et al. 1983), or a K + clone of the FCR-3 strain in human RBC (Hadley et al. 1983)HRP-I was detected simply by comparison of [3H]histidine- and [3H]isoleucine labelledproteins separated by SDS-PAGE. HRP-I is also readily detected in these parasites byimmunoprecipitation using McAb 89. By contrast, with the culture-adapted parasitestested here and isolates taken directly from Gambian patients, HRP-I was not detectedby biosynthetic labelling or immunoprecipitation methods. However, Western blottingusing McAb 89, rabbit a HRP-I or rabbit a P1HRP, did identify HRP-I in all knob-positive parasites tested including those from Gambian patients (Fig. 1; Rock et al.1987; Taylor et al. 1987).
The inclusion of urea in the 5% SDS-sample buffer used for polyacrylamide gelelectrophoresis had an unexpected effect on the mobility of HRP-I (Fig. 2). In thepresence of 2 M and 8 M urea, HRP-I exhibited a progressive decrease in electrophoreticmobility, while the mobility of HRP-II was unchanged. This suggests that HRP-Iretains secondary protein structure even after solubilization in 5 % SDS at 60 °C.

224 E. P. ROCK AND OTHERS
Additional unfolding induced by urea would presumably account for the higher ap-parent MT as the urea concentration was increased.
HRP-II has been shown in this report to be released as a soluble extracellular proteinfrom both K + and K— P. falciparum parasites. Multiple bands reactive with McAb87, McAb 1D6, and McAb 2G12 have been identified in several parasites. Some parasitescontain only 1 or 2 reactive bands while others contain up to 5 bands spanning roughlyMr 10000, in apparent size. The major HRP-II band consistently gives a diffuse leadingedge on SDS-PAGE. In all cases the extracellular form of HRP-II was of the sameapparent MT as the slowest moving intracellular form, suggesting that the extracellularform is not produced by proteolytic cleavage of the other HRP-II bands found insidewashed IRBC. The extracellular form of HRP-II always migrated as a sharp band.
HRP-II bears epitopes that appear to be unique compared with the other two HRP,and also shares epitopes found on HRP-I or HRP-III . The IgM McAb 87 and twoIgG McAbs, 1D6 and 2G12 react very strongly with HRP-II on both immunopreci-pitation and Western blotting, do not react with HRP-I by either technique, and reactweakly with HRP-III by both methods. Rabbit antisera developed against the sequenceAHH(AHHHAAD)2 react only with HRP-II. Rabbit antisera developed against HRP-1 (MCI) or the P. lophurae HRP (6211) react with HRP-1 and HRP-II on immuno-precipitation, while they only react with HRP-I on Western blotting. Previous im-munoprecipitation studies with McAbs raised against the P. lophurae HRP alsoidentified cross-reactive epitopes on HRP-I, HRP-II and the P. lophurae HRP. Thesame McAbs would react only with HRP-I or the P. lophurae protein on Westernblotting (Kilejian, 1980; Kilejian & Rosenbaum, 1985).
Immunoelectronmicroscopy using McAb 87 demonstrated that HRP-II occurs indiscrete areas within the cytoplasm of the intracellular malaria parasite, as electron-dense 'packets' within the cytoplasm of the host cell, and as peripheral clusters at andjust exterior to the IRBC surface membrane (Howard et al. 1986). Consistent with theresults of immunoelectronmicroscopy, HRP-II was identified for the first time as asurface membrane constituent of infected erythrocytes (Fig. 5). The 125I-labelled proteinwas specifically immunoprecipitated by the McAbs 87, 2G12 or 1D6 from both theTriton X-100 and sequential SDS extracts of 125I-lactoperoxidase-labelled IRBC. Thesame ikfr and size diversity was seen for the 125I-labelled HRP-II as for the corresponding[3H]histidine-labelled HRP-II from each of the four isolates examined. The McAbs didnot react with any bands from 125I-labelled uninfected RBC.
The 125I-labelled HRP-II band comigrated with the single HRP-II band recoveredfrom culture supernatants after [3H]histidine labelling. Since the faster moving HRP-II bands associated with washed PRBC were not 125I-labelled, we conclude from theseresults that we have radiolabelled the exported form of HRP-II in transit through thePRBC outer membrane. With the same samples of 125I-labelled PRBC, the malarialprotein Pf EMP 2 (P. /afciparwra-erythrocyte-membrane-protein 2) was not125l-labelled. Pf EMP 2 has been localized by immunoelectronmicroscopy using McAbs tothe electron-dense material under knobs (Howard, Lyon, Uni, Saul, Aley, Klotz,Panton, Sherwood, Marsh, Aikawa & Rock, 1987). In addition, spectrin was notlabelled under the conditions which labelled HRP-II . Thus, the 125I-labelled HRP-II isunlikely to be submembranous and is either exposed directly to the cell surface or is notsurface exposed but embedded in the lipid bilayer at a site where activated 125I canpermeate. Surface exposure of HRP-II during its transit is most likely since we havebeen able to show direct binding of McAbs against this protein to non-fixed, intactP-RBC (D. Taylor & R. Howard, unpublished results). The detergent solubility

P. falciparum histidine-rich proteins 225
properties of the 125I-labelled HRP-II suggest a strong association with the outermembrane and/or underlying cytoskeleton.
The function of HRP-II is unknown. Expression of this HRP is not related toexpression of knobs on the IRBC membrane nor to the cytoadherence phenomenonsince levels of synthesis and labelling patterns for the multiple Mr forms were identicalfor two pairs of K + and K — parasites from different P. falciparum strains and verysimilar for a third pair of cloned K + and K — parasites. Synthesis and release of HRP-II continues from early ring to late schizont stage such that at any time with MalayanCamp strain parasites in Aotus RBC roughly 50% of the total HRP-II is extracellular(Howard et al. 1986). This suggests that the function of HRP-II relates to its presencein the extracellular medium. In vivo this would correspond to the blood plasma, asindicated by radioimmunoassays which identify an antigen in malarious plasma or serawhich reacts with McAb 87 (D. Taylor, unpublished observations).
Under the conditions of SDS-PAGE used here the size range of HRP-III (MT 40000-55000) did not overlap HRP-I or HRP-II. In addition, HRP-I I I is a quantitativelyminor parasite protein in comparison to HRP-II. The rabbit antiserum raised againsta peptide sequence unique to HRP-III reacts specifically with HRP-III on immuno-precipitation and should be useful for studies on the specific location and function ofthis HRP.
Western blotting of purified P. malariae IRBC and probing with antibodies whichreact with HRP-I did not reveal an antigenically cross-reactive P. malariae protein.The knobs of P. malariae-infected erythrocytes (Smith & Theakston, 1980) thereforediffer antigenically from those of P. falciparum. P. malariae knobs may lack an analogueof HRP-I or could contain an analogous protein which lacks the epitopes defined by ourpanel of antibodies.
Our panel of antibody reagents against HRP-I and HRP-II , has not identified anyHRP analogues in numerous other malarial species tested by Western blotting (J. W.Barnwell, G. C. Cambell, R. Carter, D. W. Taylor & E. P. Rock, unpublished results).These species included a human malaria (P. vivax); primate malarias of falciparum type{P. fragile), malariae type (P. brasilianum, P. inui, P. reichienowii), and P. knowlesi; anavain malaria (P. gallinaceum); and murine malarias (P. berghei, P. vinckei, P. yoelii);as well as a sample of Babesia microti. Of the above, P. fragile and P . brasilianum bothhave surface membrane knobs (Aikawa, Miller & Rabbege, 1975).
The properties of HRP-I, HRP-II and HRP-III described here emphasize severalfeatures of these unusual proteins that may give clues to their roles for the malariaparasite. The monoclonal antibodies and rabbit antisera serve to define epitopes specificto each molecule and could be used for immunoelectronmicroscopy studies on theirsubcellular location, studies on the occurrence of these antigens and the presence ofantibodies to these proteins in malaria patients, as well as for immunoadsorbentpurification of the appropriate HRP for studies on the function of each molecule.
We are grateful to Vivien Kao, M. Parra and C. B. Evans for excellent technical assistance and toVVilma Davis for editorial assistance. This work was supported in part by grants from the Walter ReedArmy Institute for Medical Research, USPHS Grants AI 10645 and RO1 AI 20917, the UNDP/WorldBank/WHO Special Programme for Research and Training in Tropical Diseases and the US Agencyfor International Development (DPE-0453-A-00-4027). Immunoblot samples of primate malariaswere generously provided by John W. Barnwell and Gary C. Campbell and samples of avian malariagenerously provided by Richard Carter. Edwin Rock is a Research Scholar in the Howard HughesMedical Institute.

226 E. P. ROCK AND OTHERS
REFERENCESAIKAWA, M., MILLER, L. H. & RABBEGE, J. (1975). Caveola-vesicle complexes in the plasmalemma of
erythrocytes infected by Plasmodium vivax and P. cynomolgi. American Journal of Pathology 79,285-94.
ALEY, S. B., SHERWOOD, J. A. & HOWARD, R. J. (1984). Knob-positive and knob-negative Plasmodiumfalciparum differ in expression of a strain-specific malarial antigen on the surface of infectederythrocytes. Journal of Experimental Medicine 160, 1585-90.
BURKOT, T. R., WILLIAMS, J. L. & SCHNEIDER, I. (1984). Infectivity to mosquitoes of Plasmodiumfalciparum clones grown in vitro from the same isolate. Transactions of the Royal Society of TropicalMedicine and Hygiene 78, 339-41.
CULVENOR, J. G., LANGFORD, C. J., CREWTHER, P. E., SAINT, R. B., COPPEL, R. L., KEMP, D. J.,ANDERS, R. F. & BROWN, G. V. (1987). Plasmodium falciparum: Identification and localization ofa knob protein antigen expressed by a cDNA clone. Experimental Parasitology 63, 58-67.
FRAKER, P. J. & SPECK, J. C. (1978). Protein and cell membrane iodinations with a sparingly solublechloramide 1, 3, 4, 6-tetrachloro 3a, 6a-diphenyl glycoluril. Biochemical and Biophysical Research,Communications 80, 849—57.
GRANT, P. T. & FULTON, J. D. (1957). The catabolism of glucose by strains of Trypanosoma rho-desiense. Journal of Biochemistry 66, 242-50.
GREEN, T. J., GADSDEN, G., SEED, T., JACOBS, R., MORHARDT, M. & BRACKETT, R. (1985). Cloning andcharacterization of Plasmodium falciparum FCR-3/FMG strain. American Journal of TropicalMedicine and Hygiene 34, 24-30.
HADLEY, T. J., LEECH, J. H., GREEN, T. J., DANIEL, W. A., WAHLGREN, M., MILLER, L. H. & HOW-ARD, R. J. (1983). Plasmodium falciparum: A comparison of knobby (K-f) and knobless (K —)parasites of two strains. Molecular and Biochemical Parasitology 9, 271—8.
HOWARD, R. J. & BARNWELL, J. W. (1984). Solubilization and immunoprecipitation of 125I-labelledantigens from Plasmodium knowlesi schizont-infected erythrocytes using non-ionic, anionic andzwitterionic detergents. Parasitology 88, 27-36.
HOWARD, R. J., KAO, V. & BARNWELL, J. W. (1984). Protein antigens of Plasmodium knowlesi clonesof different variant antigen phenotype. Parasitology 88, 221-37.
HOWARD, R. J., LYON, J. A., UNI, S., SAUL, A. J., ALEY, S. B., KLOTZ, F., PANTON, L. J., SHERWOOD,J. A., MARSH, K., AIKAWA, M. & ROCK, E. P. (1987). Transport of a Mr ~ 300000 Plasmodiumfalciparum protein (Pf EMP2) from the intraerythrocytic asexual parasite to the cytoplasrhic faceof the host cell membrane. Journal of Cell Biology 104, 1269-80.
HOWARD, R. J., RAUM, M. G., MALOY, W. L., KAO, V. & COLIGAN, J. E. (1984). N-terminal amino acidsequence of the histidine-rich protein from Plasmodium lophurae. Molecular and BiochemidalParasitology 12, 237^6.
HOWARD, R. J., UNI, S., AIKAWA, M., ALBY, S. B., LEECH, J. H., LEW, A.M., WELLEMS, T. E.,RENER, J. & TAYLOR, D. W. (1986). Secretion of a malarial histidine-rich protein (HRP-II) fromPlasmodium falciparum-infccted erythrocytes. Journal of Cell Biology 103, 1269-77.
KILEJIAN, A. (1974). A unique histine-rich polypeptide from the malaria parasite, Plasmodiumlophurae. Journal of Biological Chemistry 249, 4650-5.
KILEJIAN, A. (1979). Characterization of a protein correlated with the production of knob-likeprotrusions on membranes of erythrocytes infected with Plasmodium falciparum. Proceedings of theNational Academy of Sciences, USA 76, 4650-3.
KILEJIAN, A. (1980). Homology between a histidine-rich protein from Plasmodium lophurae and aprotein associated with the knob-like protrusions on membranes of erythrocytes infected withPlasmodium falciparum. Journal of Experimental Medicine 151, 1534-8.
KILEJIAN, A. (1984). The biosynthesis of the knob protein and a 65000 Dalton histidine-rich poly-peptide of Plasmodium falciparum. Molecular and Biochemical Parasitolgy 12, 185-94.
KILEJIAN, A. & ROSENBAUM, S. (1985). Reactivity of a monoclonal antibody produced to the histidine-rich protein of Plasmodium lophurae with Plasmodium falciparum. Molecular and BiochemicalParasitology 17, 155-62.
KILEJIAN, A., SHARMA, Y. D., KAROUI, H. & NASLUND, L. (1986). Histidine-rich domain of the knobprotein of the human malaria parasite Plasmodium falciparum. Proceedings of the National Academyof Sciences, USA 83, 7938-41.
LAMBROS, C. & VANDERBERG, J. P. (1979). Synchronization of Plasmodium falciparum erythrocyticstages in culture. Journal of Parasitology 65, 418-20.

P. falciparum histidine-rich proteins 227
LEECH.J . H., BARNWELL, J. W.. AIKAWA, M., MILLER, L. H. & HOWARD, R. J. (1984). Plasmodiumfalciparum malaria: Association of knobs on the surface of infected erythrocytes with a histidine-rich protein and the erythrocyte skeleton. Journal of Cellular Biology 98, 1256-64.
MERYMAN, H. T. & HORNBLOWER, M. (1972). A method for freezing and washing red blood cells usinga high glycerol concentration. Transfusion 12, 145-56.
RAVETCH, J. V., FEDER, R., PAVLOVEC, A. & BLOBEL, G. (1984). Primary structure and genomicorganization of the histidine-rich protein of the malarial parasite Plasmodium lophurae. Nature,London 312, 616-20.
ROBERTS, D. D., SHERWOOD, J. A., SPITALNIK, S. L., PANTON, L. J., HOWARD, R. J., DIXIT, V. M.,FRAZIER, W. A. MILLER, L. H. & GINSBURG, V. (1985). Thrombospondin binds falciparum para-sitized erythrocytes and may mediate cytoadherence. Nature, London 318, 64-6.
SCHULZ, G. E. & SCHIRMER, R. H. (1979). Principles of Protein Structure. New York and Heidelberg:Springer-Verlag.
SMITH, D. H. & THEAKSTON, R. D. G. (1970). Comments on the ultrastructure of human erythrocytesinfected with Plasmodium malariae. Annals of Tropical Medicine and Parasitology 64, 329-31.
STAHL, H. D., KEMP, D. J., CREWTHER, P. E., SCANLON, D. B., WOODROW, G., BROWN, G. V., BIANCO,A. E., ANDERS, R. F., COPPEL, R. L. (1985). Sequence of a cDNA encoding a small polymorphichistidine and alanine-rich protein from Plasmodium falciparum. Nucleic Acids Research 13, 7837-46.
TAYLOR, D. W., PARRA, M., CHAPMAN, G. B., STEARNS, M. E., RENER, J., AIKAWA, M., UNI, S., ALKY,S. B., PANTON, L. J. & HOWARD, R. J. (1987). Localization of Plasmodium falciparum histidine-richProtein 1 in the erythrocyte skeleton under knobs. Molecular and Biochemical Parasitology (in thePress.)
TOWBIN, H., STAEHLIN, T. & GORDON, J. (1979). Electrophoretic transfer of proteins frompolyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proceedings of theNational Academy of Science USA 76, 4350-4.
TRAGER, W. & JENSEN, J . B. (1976). Human malaria parasites in continuous culture. Science 193,673-5.
UDEINYA, I. J., GRAVES, P.M., CARTER, R., AIKAWA, M. & MILLER L. H. (1983). Plasmodium fal-ciparum: Effect of time in continuous culture on binding to human endothelial cells and amelanoticmelanoma cells. Experimental Parasitology 56, 207-13.
VERNOT-HERNANDEZ, J . P. & HEIDRICH, H. G. (1984). Time-course of synthesis, transport and in-corporation of a protein identified in purified membranes of host erythrocytes infected with a knob-forming strain of Plasmodium falciparum. Molecular and Biochemical Parasitology 12, 337-50.
VERNOT-HERNANDEZ, J. P. & HEIDRICH, H. G. (1985). The relationship to knobs of the 92000 Dprotein specific for knobby strains of Plasmodium falciparum. Zeilschrift fur Parasilenkunde 71,41-51.
WALLIKER, D., CARTER, R., QUAKYI, I. A., WELLEMS, T. E., MCCUTCHAN, T. F. & SZARFMANN, A.(1986). Genetics of Plasmodium falciparum. In Molecular Strategies of Parasitic Invasion, UCLASymposium on Molecular and Cellular Biology, New Series, vol. 42, (ed. N. Agabian, H. Goodmanand N. Noguiera). New York: Alan R. Liss, Inc.
WELLEMS, T. E. & HOWARD, R. J. (1986). Homologous genes encode two distinct histidine-richproteins in a cloned isolate of Plasmodium falciparum. Proceedings of the National Academy ofSciences, USA 83, 6065-9.
WELLEMS, T. E., WALLIKER, D., SMITH, C. L., DO ROSARIO, V. E., MALOY. W. L., HOWARD, R. J.,CARTER, R. & MCCUTCHAN, T. F. (1987). A histidine-rich protein gene marks a linkage groupfavored strongly in a genetic cross of Plasmodium falciparum. Cell (in the Press.)
Printed in Great Britain