compaction and root modifications of soil aeration
TRANSCRIPT

Compaction and Root Modifications of Soil AerationG. H. Asady and A. J. M. Smucker*
ABSTRACTSoil compaction and the incipient aeration and mechanical imped-
ance stresses are known to reduce root growth and function. Littleis known, however, regarding the influence of root accumulationsadjacent to a compacted area upon the aeration status of the soilprofile below the root mass. This study was designed to measureplant root and soil aeration interactions with soil compaction. A claysoil was compacted to bulk densities in 1.1,1.4 and 1.7 Mg m~3 insoil columns planted to dry edible bean (Phaseolus vitlgaris L.). Me-chanical resistances of these soil compaction treatments were 0.43,2.14, and 5.50 MPa, respectively. Oxygen diffusion rates (ODR)were below the critical level for the 1.7 Mg nr1 treatment at allplant growth stages measured. As root accumulations increased atthe 0.10- to 0.25-m depths, ODR dropped below critical values forall bulk densities. Lower O, concentrations above the soil surfacedecreased ODR both earlier and at shallower depths. Root pluggingof 5% or more of the aeration pores significantly reduced the dif-fusion of O2 into and CO, out of the soil regions below and adjacentto areas with the greatest accumulation of roots.
GREATER MECHANICAL RESISTANCE and the de-creased aeration of compacted soils limit both
the length and function of plant roots. These limitingsoil conditions are generally synergistic and difficultto separate. Several have attempted to separate themechanical impedance and aeration effects upon rootsby measuring root elongation responses to artificialmedia (Abdalla et al., 1969; Russell and Goss, 1974)and by measuring granular media responses to artifi-cial roots (Richards and Greacean, 1986). However,there have been no reports of measurements thatquantify the adverse effects of compaction-modifiedroot systems on the aeration properties of a com-pacted soil. Limitations of root growth by compactedconditions at soil depths below the tillage pan com-bined with the greater fluctuations in water contentand temperature of the surface horizon may result ingreater crop vulnerability to aeration as well as nu-trient and water stresses (Kladivko et al., 1986, Mackayet al., 1987). This study was designed to quantify rootresponses to soil compaction and measure the changesG.H. Asady, Dep. of Agronomy, Univ. of Georgia, Coastal PlainExp. Stn., P.O. Box 748, Tiftpn, GA 31793-0748; and A.J.M.Smucker, Dep. of Crop and Soil Sciences, Michigan State Univ.,East Lansing, MI 48824. Published as Michigan Agric. Exp. Stn.Journal Article no. 12141. Received 31 Dec. 1987. 'Correspondingauthor.
Published in Soil Sci. Soc. Am. J. 53:251-254 (1989).
in soil aeration attributed to the presence or absenceof roots.
MATERIALS AND METHODSTopsoil from the Ap horizon of a Charity clay (Illitic,
calcareous, mesic, Aerie Haplaquept) was compressed intosections of schedule 80 polyyinyl chloride (PVC) tubes 7.6cm in length and diam. Gas-tight access ports, 0.5-cm diam.,were drilled into the sides and at the center of each sectionand sealed by rubber serum stoppers. Soil was compressedto the desired bulk density in each of the three sections be-fore assembly, by the methods reported earlier (Asady et al.,1985). Penetrometer measurements were made on three ad-ditional replications using the dynamic penetrometer at theUniversity of Minnesota. Soil at the top and bottom of eachsection was lightly abraided by a wire brush to eliminate anysealing effects the steel press may have had on the surfaceof each core. An empty PVC section (7.6-cm diam., 2.5-cmlength) and the three compacted soil sections were assem-bled into a soil column and secured by plastic impregnatedduct tape, 5-cm wide (Fig. 1). One layer each of filter paper(Whatman no. 1) and cheesecloth were secured to the baseof each soil column by a rubber band. Water continuity wasestablished in each assembled soil column by submergingthe columns to within 1 cm of the surface soil. Soil columnswere saturated for 48 h before transferring them to a blotterpaper tension table (Hillel, 1982) and equilibrated to a ma-tric soil water potential of —0.006 MPa at the base of eachcolumn. Tension tables were covered with black plastic film(3-mm thickness), each containing 18 gas- and water-tightports for the soil columns (Fig. 1). Matric potential of thetension table and water supply was maintained by a Mari-onette burette reservoir, which was also used to measurewater use.
Seeds of dry edible bean (cv. Seafarer) were surface ster-ilized with 0.05% sodium hypochlorate solution for 10 minand thoroughly rinsed with distilled water. Seeds were ger-minated on wet cheesecloth and paper towels at 25 °C for4 d. Uniform seedlings were selected and one transplantedinto the top empty PVC ring of each soil column, coveredwith loose Charity clay soil and irrigated to saturation toestablish water continuity with the soil column below (Fig.1). Although several columns had been packed with soilsranging in bulk densities from 1.1 to 1.7 Mg m~3 (0.18 gra-vimetric) and soil measurements taken, time and space lim-itations reduced plant experiments to soils having bulk den-sities of 1.1, 1.4, and 1.7 Mg m~3. Four days aftertransplanting, the top of each soil column was isolated fromthe atmosphere by a plastic impregnated cardboard con-tainer cap that was secured to the columns by duct tape andsealed around the plant by foam plugs impregnated withparaline (paraffin and vaseline mixed at a ratio of 7:1). Hy-podermic needles (18 gauge and 3.8 cm in length) were usedto control the flow rates of compressed air and N2 gases at

252 SOIL SCI. SOC. AM. J., VOL. 53, JANUARY-FEBRUARY 1989
Fig 1. Schematic view of the root chamber and tension table ap-paratus, which includes the chamber for surface soil (A), threesections of the experimentally compacted soil column (B), accessports for gas sampling (C), soil moisture tension table (D), plasticgas and water barrier (E), and the applied matric potential head(H).
the rate of 67 X 10^3 L s~'. This arrangement provided headspace O2 concentrations of 0.21 and 0.14 mol mol~' for theair and N2 treatments, respectively. Hoagland's nutrient so-lution was applied at 25% full strength and distributed equallythrough each soil column at 7-d intervals following the sec-ond week after planting. Greenhouse day/night tempera-tures were 24/18 "C and a 16-h photoperipd was controlledby supplemental fluorescent tubes providing photosynthet-ically active radiation (PAR) light intensities of 0.640 (cloudy)and 1.012 ianol m~2 s~' (full sun). Relative humidity wasmaintained at 65 ± 5%.
Soil oxygen diffusion rates (ODR) were measured by theplatinum microelectrode method of Lemon and Erickson(1955). Electrodes were inserted through the access ports andmeasured 3.8 cm into each of the three sections of the soilcolumn. Soil temperatures were measured through the rub-ber septum access ports using a portable digital microther-mocouple (Service Tectonics Instruments type K modelA2501, Detroit). Prior to assembly, diffusion rates throughthe bulk soil (D/D0) were measured on the 7.6-cm soil cores,equilibrated to a mid core Stm of —0.006 MPa, by the dif-fusion chamber method of Rolston (1986).
Gas concentrations of the soil were sampled periodically,during the plant growth period, by extracting 0.001-L sam-ples via the access ports using a hypodermic syringe (Tu-berculin type B-D and 18-gauge needles, 3.8 cm in length,Becton, Dickinson Co., Rutherford, NJ). Concentrations ofp2 and CO2 at the three soil depths were determined byinjecting 0.001-L samples into a gas-liquid chromatograph(Carle Instruments, Model 8500, Fullerton, CA) equippedwith a 0.2- by 0.0032-m (i.d.) stainless steel column of Po-rapak QS (Supelco Inc., Bellfonte, PA) connected in parallelwith a molecular sieve column having the same dimensions.Column oven temperatures were 40 °C, the He carrier gasflow rate was 0.0005 L s~~' and the thermal conductivitydetector temperature was at the low setting. Resulting peakswere compared to standard gases with a computing integra-tor.
Morphological plant analyses included the destructivesampling and oven drying at 70 °C for fresh weight and dryweight measurements of the roots and shoots. Root lengthsfor each soil depth were determined by washing roots freeof soil with the hydropneumatic elutriation method (Smuckeret al., 1982) and manually counting root lengths by the line-intercept method of Tennant (1975). Volume of roots within
each compartment were determined by the water displace-ment method and root-filled porosity established as the ratioof volume of roots to total soil volume at a given waterpotential.
Experimental design was a four factor factorial, random-ized complete block with four replications. The main effectswere: (i) three levels of soil bulk density (1.1, 1.4, and 1.7Mg m~3 dry weight compacted at 0.18 gravimetric watercontents), (ii) two head space gas treatments (air and N2),(iii) three harvest dates (10,20, and 30 d after transplanting),and (iv) three soils depths (6, 14, and 21 cm at mid-points).Statistical analysis included standard analysis of varianceusing SAS Institute procedures (1985).
RESULTS AND DISCUSSIONSoil Aeration
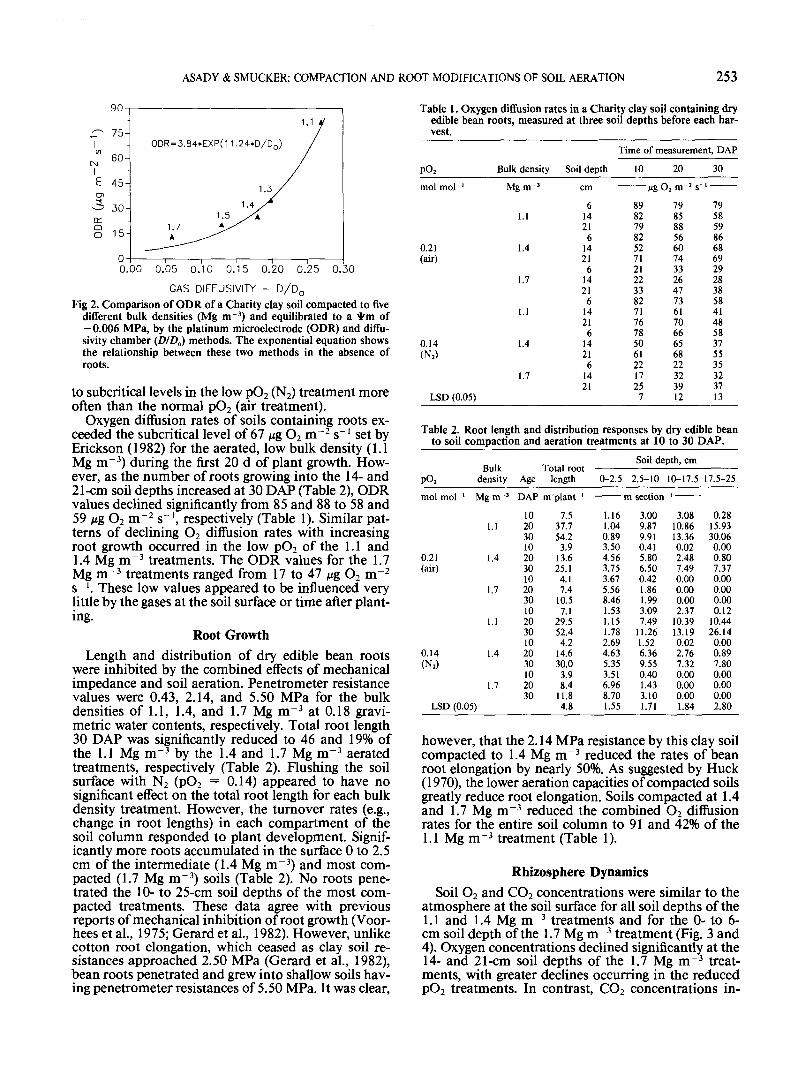
Relatively small increases in the bulk density of theCharity clay greatly reduced the diffusion rates of soiloxygen (Fig. 2). Platinum microelectrode (ODR) mea-surements of soil without roots were reduced more(58%) than the diffusivity CD/A,) method (22%), whenbulk densities were increased from 1.1 to 1.3 Mg m~3.Since O2 diffusion to the surface of microelectrodes isprimarily controlled by film widths of the liquid phase,small increases in soil density (e.g., 1.1. to 1.3 Mg m~3)increased the relative proportion of saturated micro-pores resulting in greater resistances to O2 diffusionthrough the liquid phase (ODR). Although these in-cremental increases in bulk density reduced the num-ber and continuity of macropores somewhat, O2 dif-fusion through the gaseous phase (D/D0) appeared tobe less affected. Consequently reductions of ODR weremuch greater than D/D0. Further reductions in soilbulk density, however, reduced D/D0 values more thanODR (Fig. 2) due to the loss of macropores and greatertortuosities of gaseous diffusion. The ODR appearedto change very little at the higher bulk densities, pre-sumably because film thickness at 1.4 Mg m~3 wasgreat enough to limit O2 diffusion rates to below thecritical level of 33 Mg m~2 s"1 (Erickson, 1982). Theexponential relationship between ODR and DJD0 fora clay soil compacted to multiple bulk densities in-dicates that the microelectrode method is more sen-sitive to changes in O2 at lower bulk densities whilegas diffusivity measurements are more sensitive tochanges in the O2 status of soils compacted to higherbulk densities. However, since it was impossible tonondestructively measure O2 diffusivity through thegas phase at each of the three depths of the soil columnby the chamber method, we chose the less disruptivein situ microelectrode method.
Oxygen diffusion rates within soil columns contain-ing plant roots appeared to be reduced to rates belowthe critical level of 33 \t% O2 m~2 s"1 (Erickson, 1982)by soil compaction and lower O2 concentrations at thesoil surface (Table 1). Increasing bulk density from 1.1to 1.7 Mg m~3 resulted in significant decreases in theODR for all soil depths and ages of the plant reportedexcept for the 14- and 21-cm depths at 30 d after plant-ing (DAP), under low pO2 (N2 treatment). Increasingbulk densities from 1.1 to 1.4 Mg m~3 resulted inmixed and transitional reductions in ODR with sig-nificant reductions occurring when air at the soil sur-face was replaced by N2. The ODR generally dropped
ASADY & SMUCKER: COMPACTION AND ROOT MODIFICATIONS OF SOIL AERATION 253
90-
^ 75-
E 45-|
§ 15
ODR=3.B4*EXP(1 1.24*D/DO)
Table I. Oxygen diffusion rates in a Charity clay soil containing dryedible bean roots, measured at three soil depths before each har-vest.
00.00 0.05 0.10 0.15 0.20 0.25 0.30
GAS DIFFUSIVITY - D/DO
Fig 2. Comparison of ODR of a Charity clay soil compacted to fivedifferent bulk densities (Mg m~3) and equilibrated to a *m of-0.006 MPa, by the platinum microelectrode (ODR) and diffu-sivity chamber (D/D0) methods. The exponential equation showsthe relationship between these two methods in the absence ofroots.
to subcritical levels in the low pO2 (N2) treatment moreoften than the normal pO2 (air treatment).
Oxygen diffusion rates of soils containing roots ex-ceeded the subcritical level of 67 /*g O2 m~2 s~' set byErickson (1982) for the aerated, low bulk density (1.1Mg m~3) during the first 20 d of plant growth. How-ever, as the number of roots growing into the 14- and21-cm soil depths increased at 30 DAP (Table 2), ODRvalues declined significantly from 85 and 88 to 58 and59 /tg O2 m"2 s~', respectively (Table 1). Similar pat-terns of declining O2 diffusion rates with increasingroot growth occurred in the low pO2 of the 1.1 and1.4 Mg m~3 treatments. The ODR values for the 1.7Mg m~3 treatments ranged from 17 to 47 /ig O2 m~2
s~'. These low values appeared to be influenced verylittle by the gases at the soil surface or time after plant-ing.
Root GrowthLength and distribution of dry edible bean roots
were inhibited by the combined effects of mechanicalimpedance and soil aeration. Penetrometer resistancevalues were 0.43, 2.14, and 5.50 MPa for the bulkdensities of 1.1, 1.4, and 1.7 Mg m~3 at 0.18 gravi-metric water contents, respectively. Total root length30 DAP was significantly reduced to 46 and 19% ofthe 1.1 Mg m~3 by the 1.4 and 1.7 Mg m~3 aeratedtreatments, respectively (Table 2). Flushing the soilsurface with N2 (pO2 = 0.14) appeared to have nosignificant effect on the total root length for each bulkdensity treatment. However, the turnover rates (e.g.,change in root lengths) in each compartment of thesoil column responded to plant development. Signif-icantly more roots accumulated in the surface 0 to 2.5cm of the intermediate (1.4 Mg m~3) and most com-pacted (1.7 Mg m~3) soils (Table 2). No roots pene-trated the 10- to 25-cm soil depths of the most com-pacted treatments. These data agree with previousreports of mechanical inhibition of root growth (Voor-hees et al., 1975; Gerard et al., 1982). However, unlikecotton root elongation, which ceased as clay soil re-sistances approached 2.50 MPa (Gerard et al., 1982),bean roots penetrated and grew into shallow soils hav-ing penetrometer resistances of 5.50 MPa. It was clear,
Time of measurement, DAP
pO2 Bulk density Soil depth
mol mol ' Mg
1
0.21 1(air)
1
1
0.14 1(N2)
1
LSD (0.05)
Table 2. Root lengthto soil compaction
m J
.1
.4
.7
.1
.4
.7
andand
cm
61421
614216
1421
61421
61421
61421
distribution
10
US (898279825271212233827176785061221725
7
responses byaeration treatments at 10
20)2 m-2 s
79858856607433264773617066656822323912
30
79585986686929283858414858375535323713
dry edible beanto 30 DAP.
Soil deoth. cmBulk
pO2 density Age
mol mol ' Mg m 3 DAP
1.1
0.21 1.4(air)
1.7
1.1
0.14 1.4(N2)
1.7
LSD (0.05)
102030102030102030102030102030102030
Total roui -length
m 'plant
7.537.754.23.9
13.625.14.17.4
10.57.1
29.552.44.2
14.630.0
3.98.4
11.84.8
-' -
0-2.5
1.161.040.893.504.563.753.675.568.461.531.151.782.694.635.353.516.968.701.55
2.5-10
m section
3.009.879.910.415.806.500.421.861.993.097.49
11.261.526.369.550.401.433.101.71
10-17.5
3.0810.8613.360.022.487.490.000.000.002.37
10.3913.190.022.767.320.000.000.001.84
17.5-25
0.2815.9330.060.000.807.370.000.000.000.12
10.4426.140.000.897.800.000.000.002.80
however, that the 2.14 MPa resistance by this clay soilcompacted to 1.4 Mg m~3 reduced the rates of beanroot elongation by nearly 50%. As suggested by Huck(1970), the lower aeration capacities of compacted soilsgreatly reduce root elongation. Soils compacted at 1.4and 1.7 Mg m~3 reduced the combined O2 diffusionrates for the entire soil column to 91 and 42% of the1.1 Mg m~3 treatment (Table 1).
Rhizosphere DynamicsSoil O2 and CO2 concentrations were similar to the
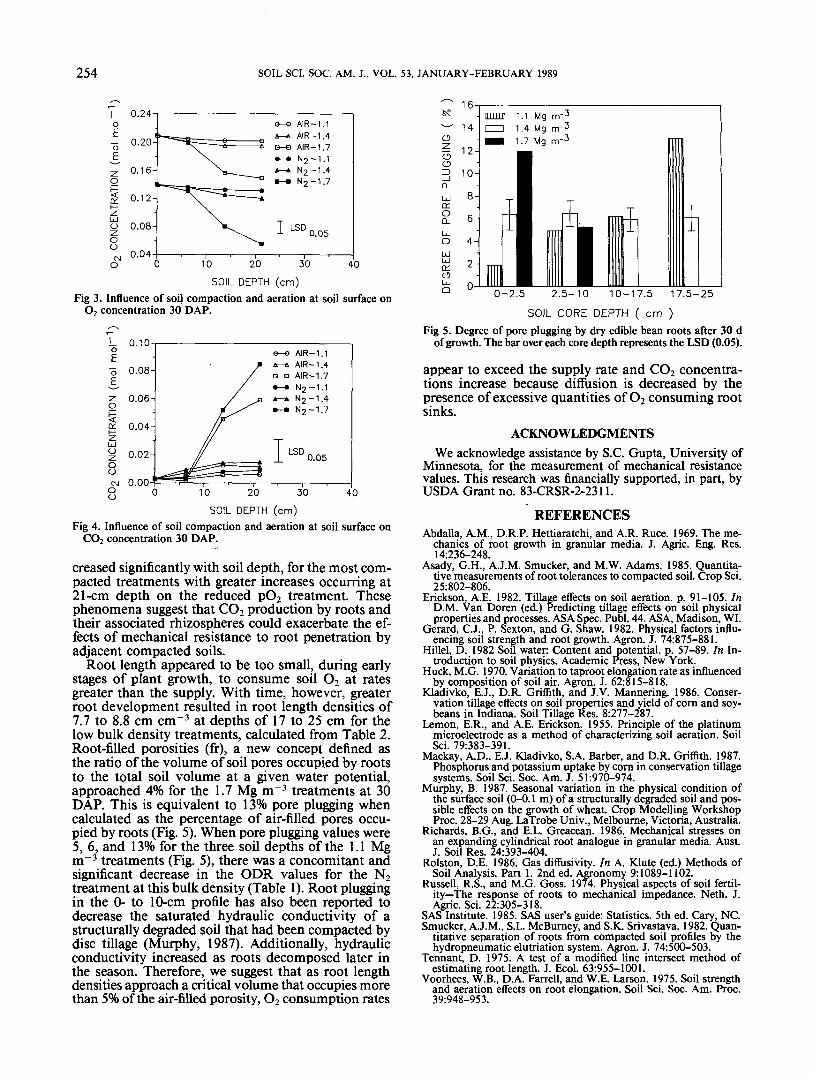
atmosphere at the soil surface for all soil depths of the1.1 and 1.4 Mg m~3 treatments and for the 0- to 6-cm soil depth of the 1.7 Mg m~3 treatment (Fig. 3 and4). Oxygen concentrations declined significantly at the14- and 21-cm soil depths of the 1.7 Mg m~3 treat-ments, with greater declines occurring in the reducedpO2 treatments. In contrast, CO2 concentrations in-
254 SOIL SCI. SOC. AM. J., VOL. 53, JANUARY-FEBRUARY 1989
I 0.24 HTTTTTI 1.1 Mg rrr-5
1.4 Mg rrr3
1.7 Mg m-3
10 20 30
SOIL DEPTH (cm)
Fig 3. Influence of soil compaction and aeration at soil surface onO2 concentration 30 DAP.
L 0.10oE-5 0.08-
<QL
OO
<NOO
0.06-
0.04-
0.02-
0.0020
SOIL DEPTH (cm)Fig 4. Influence of soil compaction and aeration at soil surface on
CO2 concentration 30 DAP.
creased significantly with soil depth, for the most com-pacted treatments with greater increases occurring at21-cm depth on the reduced pO2 treatment. Thesephenomena suggest that CO2 production by roots andtheir associated rhizospheres could exacerbate the ef-fects of mechanical resistance to root penetration byadjacent compacted soils.
Root length appeared to be too small, during earlystages of plant growth, to consume soil O2 at ratesgreater than the supply. With time, however, greaterroot development resulted in root length densities of7.7 to 8.8 cm cm~3 at depths of 17 to 25 cm for thelow bulk density treatments, calculated from Table 2.Root-filled porosities (fr), a new concept defined asthe ratio of the volume of soil pores occupied by rootsto the total soil volume at a given water potential,approached 4% for the 1.7 Mg m~3 treatments at 30DAP. This is equivalent to 13% pore plugging whencalculated as the percentage of air-filled pores occu-pied by roots (Fig. 5). When pore plugging values were5, 6, and 13% for the three soil depths of the 1.1 Mgm"3 treatments (Fig. 5), there was a concomitant andsignificant decrease in the ODR values for the N2treatment at this bulk density (Table 1). Root pluggingin the 0- to 10-cm profile has also been reported todecrease the saturated hydraulic conductivity of astructurally degraded soil that had been compacted bydisc tillage (Murphy, 1987). Additionally, hydraulicconductivity increased as roots decomposed later inthe season. Therefore, we suggest that as root lengthdensities approach a critical volume that occupies morethan 5% of the air-filled porosity, O2 consumption rates
tfl
0-2.5 2.5-10 10-17.5 17.5-25
SOIL CORE DEPTH ( cm )
Fig 5. Degree of pore plugging by dry edible bean roots after 30 dof growth. The bar over each core depth represents the LSD (0.05).
appear to exceed the supply rate and CO2 concentra-tions increase because diffusion is decreased by thepresence of excessive quantities of O2 consuming rootsinks.
ACKNOWLEDGMENTSWe acknowledge assistance by S.C. Gupta, University of
Minnesota, for the measurement of mechanical resistancevalues. This research was financially supported, in part, byUSDA Grant no. 83-CRSR-2-2311.