熊本大学学術リポジトリ kumamoto university repository...
TRANSCRIPT

熊本大学学術リポジトリ
Kumamoto University Repository System
Title 糖尿病におけるアミノ酸プロファイルの経時的変化
Author(s) 持田, 太賀
Citation
Issue date 2011-03-25
Type Thesis or Dissertation
URL http://hdl.handle.net/2298/25168
Right

1
1.目次
1. 目次 P 1-3
2. 要旨 P 4-5
3. 学位論文の骨格となる発表論文、および過去の発表論文リスト P 6
4. 謝辞 P 7
5. 略語一覧 P 8-9
6. 研究の背景および目的 P 10-19
6-(1) 糖尿病および生活習慣病の血漿中アミノ酸濃度に対する影響 P 10-13
6-(2) AKITA マウス P 13-16
6-(3) アミノ酸分析の発展 P 17-18
6-(4) 本研究の目的 P 18-19
Table 1 P 12
Table 2 P 13
Figure 1 P 16
7. 実験方法 P 20-25
7-(1) 実験対象動物および飼育環境 P 20
7-(2) 食餌摂取量の測定 P 20

2
7-(3) 血漿中インスリン濃度の測定 P 20-21
7-(4) 遺伝子型決定 P-21
7-(5) 体重測定および血液採取プロトコール P 21-22
7-(6) アミノ酸分析 P 22
7-(7) 統計学的分析 P 22-23
Figure 2 P 24-25
8. 実験結果 P 26-42
8-(1) 遺伝型決定 P 26
8-(2) AKITA マウスにおける血糖の推移 P 26
8-(3) AKITA マウスの自然経過 P 26-27
8-(4) AKITA マウスにおける血漿中アミノ酸濃度の推移 P 27
8-(5) 糖尿病の発症早期に濃度変化を認めるアミノ酸 P 28
Table 3 P 29
Figure 3-13 P 30-43
9. 考察 P 44-47
9-(1) 実験系 P 44
9-(2) 血漿中アミノ酸濃度に対する食餌摂取量の影響 P 45
9-(3) 血漿中アミノ酸濃度に対するインスリンの影響 P 45
9-(4) 糖尿病発症時のアラニンの代謝変化 P 45-46
9-(5) 糖尿病発症時の分枝鎖アミノ酸の代謝変化 P 46
9-(6) 糖尿病発症時のプロリンの代謝変化 P 46
9-(7) 血漿中アミノ酸濃度に対する糖尿病合併症の影響 P 46-47

3
Figure 14-16 P 48-50
10. 結語 P 51
11. 参考文献 P 52-56

4
2. 要旨
【目的】臨床において、アミノ酸分析は先天代謝異常症の診断を目的として行われる事が
多い。だが、研究レベルにおいては疾患特異的アミノ酸プロファイルを検証した
報告が多数存在する。我々は、疾患特異的アミノ酸プロファイルについて継時的
なデータを供給することで、アミノ酸分析が様々な疾患の病期判定に有用なツー
ルとなる可能性があると考えた。我々は新たなアミノ酸分析の有用性を見出すた
め、疾患モデル動物を用いて経時的なアミノ酸プロファイルの変化を検証した。1
型糖尿病に着目し、アミノ酸プロファイルに対する糖尿病発症の影響について検
証することを本研究の目的とした。
【方法】AKITA マウスは Insulin2 遺伝子にミスセンス変異を持つ。Insulin2 遺伝子の制限
酵素断片長多型分析を行って、遺伝型ごとにグループ分けをした。3週齢から 20
週齢まで週 1 回のペースで検体採取を行った。採血は 2 時間絶食後に行った。
HPLC-ESI-MS によるアミノ酸分析により、同一動物の経時的なアミノ酸プロファイ
ルの変化を検証した。グループ間のアミノ酸濃度の推移を比較検討するため二元
配置分散分析を行った。さらに、糖尿病発症早期にグループ間で濃度差を認める
アミノ酸を検証する目的でクラスター分析を行い、血糖と同じグループに分類さ
れたアミノ酸について、健常状態から高血糖状態になった際におこる血漿中濃度
の相対的変化を評価した。
【結果】ホモの Insulin2 遺伝子変異マウス(Ins2 C96Y / C96Y)は、3 週齢の時点で高血糖で
あったため研究対象から除外し、ヘテロの Insulin2 遺伝子変異マウス(Ins2WT/C96Y)
と野生型マウス(Ins2WT/WT)で比較検討した。Ins2WT/WT と比較し、高血糖状態にな
る Ins2WT/C96Y において、血漿中のバリン、ロイシン、イソロイシン、総分枝鎖ア
ミノ酸、アラニン、シトルリン、プロリンの各濃度が Ins2WT/C96Y において有意に

5
高値であった。両群の差は時間経過とともに徐々に大きくなった。クラスター分
析の結果、血糖上昇に連動して濃度が上昇するアミノ酸として、バリン、ロイシ
ン、イソロイシン、総分枝鎖アミノ酸、アラニンが示唆された。血漿中アミノ酸
濃度の相対変化を評価した結果、健常状態から高血糖状態になる際に最も早く二
群間の差が大きくなる可能性があるアミノ酸としてアラニンが考えられた。ほと
んどのアミノ酸は高血糖状態になった後に変化を認めた。
【考察】両群間で明確な差が出る時期は、血糖値や血漿中インスリン濃度と比較すると、
ほとんどのアミノ酸で遅かった。この結果は、血糖異常、血漿中インスリン濃度
の低下以外の要因も関与することを示唆している。血糖異常、血漿中インスリン
濃度低下以外の要因として代謝酵素活性の変化、高血糖による合併症の影響、尿
中へのアミノ酸排泄量の変化などが考えられる。これらを明確に評価するために
は更なる実験が必要である。
【結語】アミノ酸プロファイルは高血糖の影響をうけるが、ほとんどのアミノ酸は血糖上
昇の後に変化を認めた。さらに、糖尿病を発症した後も時間依存性にアミノ酸プ
ロファイルの変化は続いた。インスリン活性、代謝酵素活性の変化、組織や臓器
における微小環境の変化などさまざまな因子が血漿中のアミノ酸濃度に影響を及
ぼす可能性を有していると考えられた。経時的なアミノ酸分析は、病期や合併症
の評価に有用である可能性が示唆された。

6
3. 学位論文の骨格となる発表論文および過去の発表論文リスト
参考論文
①関連論文
1. Taiga Mochida, Takayuki Tanaka, Yasuko Shiraki, Hiroko Tajiri, Shirou
Matsumoto, Kazutaka Shimbo, Toshihiko Ando, Kimitoshi Nakamura, Masahiro
Okamoto, Fumio Endo. Time-dependent changes in the plasma amino acid
concentration in diabetes mellitus. Molecular Genetics and Metabolism 103 (2011)
406-409.

7
4. 謝辞
本研究を行うにあたり、全面的にご指導頂きました熊本大学大学院生命科学研究部小児科学
分野教授、 遠藤 文夫教授に深く感謝申し上げます。
アミノ酸測定技術および測定データの統計解析について、多大なるご指導、ご協力頂きました
味の素株式会社、安東 敏彦氏、新保 和高氏、田中 孝幸氏に厚く御礼申し上げます。
測定データの統計解析についてご助言頂きました九州大学大学院農学研究院生命機能科学
部門生物機能制御学部門、岡本 正宏教授に厚く御礼申し上げます。
大学院入学時より実験計画、実験手技、論文作成についてご指導いただきました熊本大学大
学院生命科学研究部小児科学分野、中村 公俊先生、松本 志郎先生、白木 恭子先生に深く
感謝申し上げます。
大学院生活の中で様々な場面でご協力いただいた、佐藤 歩先生、服部 希世子先生、上野
靖先生、西原 卓宏先生、緒方 陽先生、福永 啓文先生、森 博子先生、中村 賢二先生、城戸
淳先生、木川 和英先生、田村 博先生に深く感謝申し上げます。
また、研究を進める中で様々な場面でご助力いただきました技術補助員の皆様に深く感謝申し
上げます。

8
5. 略語一覧
ABA:α amino butyric acid
Ala:Alanine
Arg:Arginine
Asn:Asparagine
Asp:Aspartate
BCAA:Branched-chain amino acid
BS:Blood sugar
Cit:Citrulline
Cys2:Cystine
DM:Diabetes mellitus
ESI:Electrospray ionization
Gln:Glutamine
Glu:Glutamate
His:Histidine
HPLC:High performance liquid chromatography
Ile:Isoleucine
Leu:Leucine
Lys:Lysine
Met:Methionine
MODY:Maturity Onset Diabetes of Young
MS:Mass spectrometry
Orn:Ornithine
Phe:Phenylalanine
Pro:Proline

9
Ser:Serine
Tau:Taurine
Thr:Threonine
Trp:Tryptophane
Tyr:Tyrosine
Val:Valine

10
6. 研究の背景および目的
6-(1) 糖尿病および生活習慣病の血漿中アミノ酸濃度に対する影響
臨床の場において、アミノ酸分析は主に先天代謝異常症の診断、肝機能障害を評価する
手段として用いられている(Schulze, Lindner et al. 2003; Wilcken, Wiley et al. 2003;
Morgan, Milsom et al. 1978)。研究レベルにおいては、腎機能障害、敗血症(Vente, von
Meyenfeldt et al. 1989)、悪性腫瘍(Norton, Gorschboth et al. 1985)など疾患特異的ア
ミノ酸プロファイルを検証した報告が多数存在する(Table 1)。これらの報告は先天代謝
異常に限らず、様々な疾患の診断や重症度の評価にアミノ酸分析が有用であることが示唆
している。また、疾患特異的アミノ酸プロファイルについて継時的なデータを供給するこ
とにより、アミノ酸分析は様々な疾患の病期判定に有用なツールとなりうるといえる。
糖尿病はインスリン機能不全に伴い血糖異常をきたす疾患である。国際糖尿病連合(IDF)
の推計によると、2011 年現在の糖尿病患者は、世界で 3 億 6600 万人にのぼるとされている
(IDF Diabetes Atlas)。糖尿病は大きく以下の4つに分類される。
1 型糖尿病は、ランゲルハンス島のβ細胞が破壊されることが原因で発症する糖尿病であ
る。β細胞の破壊に伴い、インスリン分泌機能が障害されるため、1型糖尿病に罹患したヒ
トは、ほとんどインスリンを産生することができない。自己抗体が検出される自己免疫性
の 1 型糖尿病と原因が特定できない特発性の 1 型糖尿病が存在し、後者では前述のような
現象がなぜ起こるかまだ完全には判明していない。1 型糖尿病はどの年代でも起こりうるが、
通常は小児や若年の成人が発症する事が多い。1 型糖尿病では血糖コントロールのために毎
日のインスリン治療が必須であり、インスリンを投与されない場合は命を落とすこともあ
りうる。
2 型糖尿病は、インスリン抵抗性と相対的インスリン不足によって特徴づけられる糖尿病
である。2型糖尿病では 2 型糖尿病は 40 歳以降で発症する事が多く、しばしば肥満や高脂
血症を合併しているケースが認められる。検査で偶然に血糖異常や尿糖を認める、あるい
は合併症がおこって初めて診断に至るケースもしばしば存在し、発症後数年にわたり診断

11
されないこともある。2型糖尿病の治療は、食事療法、運動療法などによる体重コントロー
ルがその中核であり、インスリン投与は必須ではない。
妊娠糖尿病(GDM)は、妊娠中に高血糖を認める場合を指す。世界ではおよそ 25 妊娠に 1
件の割合で起こっている。GDM では出産後症状は軽快するが、GDM を発症した女性およびそ
の子はのちに 2 型糖尿病を発症するリスクが高い。およそ半数は 5年から 10 年以内に 2 型
糖尿病を発症する。
そのほかに、膵炎や膵臓がんなど他の病気が原因で起こる二次性糖尿病や、遺伝子異常
で起こる糖尿病などがあり、これらはその他の糖尿病として分類されている。
インスリンは血糖の調節をつかさどるだけでなく、タンパク合成にも関与しているため、
糖尿病はアミノ酸代謝に影響を及ぼす。それでは、糖尿病の発症によって、アミノ酸代謝
にはどのような変化が生じるのであろうか。過去の報告において、インスリン投与で血漿
中総アミノ酸濃度が減少する(James Murray Luck 1928)ことが知られている。また、糖尿
病モデル(alloxan-diabetes, partial pancreatectomy, administration of anti-insulin
serum)ラットで糖尿病の影響により、血漿中総アミノ酸濃度は増加する傾向にあった
(Scharff and Wool 1966)ことが報告されている。これらの現象は、インスリンの作用の低
下に伴い、蛋白合成が低下し異化は亢進するためであると考えらている。
さらに、糖尿病発症におけるアミノ酸プロファイルについては多くの論文で論じられて
いる。糖尿病に罹患したヒト(Vannini, Marchesini et al. 1982; Marchesini, Bianchi et
al. 1991)や糖尿病モデル動物(Crandall and Fernstrom 1983; Prior and Smith 1983)に
おいて、血漿中の分枝鎖アミノ酸濃度が上昇することはよく知られている。分枝鎖アミノ
酸の血漿濃度の上昇は、インスリン抵抗性の存在するヒトにおいても確認されている
(Felig, Marliss et al. 1969; Chevalier, Burgess et al. 2006)
近年、分枝鎖アミノ酸の代謝について、筋肉のみならず脂肪組織も大きく関与している
ことが分かってきた(Herman, She et al. 2010)。She らは、肥満モデルのげっ歯類におけ
る分枝鎖アミノ酸代謝の変化について報告している。彼らは、肥満モデルで、肥満に伴い
脂肪組織におけるミトコンドリア分枝鎖アミノ酸トランスフェラーゼ(BCATm)活性の低下

12
が認められることを示している。さらに She らは、手術後の侵襲により体重減少したヒト
において、血漿中分枝鎖アミノ酸濃度の低下と BCATm の活性上昇が認められることも報告
している(She, Van Horn et al. 2007)。これらの報告から、肥満に伴う脂肪組織での分枝
鎖アミノ酸代謝の変化が、糖尿病における分枝鎖アミノ酸濃度上昇の一要因であることが
示唆される。しかし、肥満を来さない 1 型糖尿病でも、同様に血漿中の分枝鎖アミノ酸濃
度の上昇を認め、脂肪組織における代謝変化以外の要因の存在が示唆されている。
糖尿病の血漿中アラニン濃度に対する影響についても多くの報告がある (Vannini,
Marchesini et al. 1982; Robert, Beaufrere et al. 1985; Consoli, Nurjhan et al. 1990;
Oresic, Simell et al. 2008)。分枝鎖アミノ酸、アラニンのほかにも、糖尿病において濃
度変化を認めるアミノ酸について論じた報告は多々あるが、その内容は実に様々である
(Table 2)。しかし、高血糖下での長期的なアミノ酸変化についての報告は、いまだ試さ
れていない。
Table1 疾患特異的アミノ酸プロファイルの変化

13
Table 2 糖尿病発症に伴うアミノ酸プロファイルの変化
6-(2) AKITA マウス
AKITA マウスは 1997 年に小泉らによって発見された常染色体優性遺伝を示す糖尿病モデ
ルマウスである(Yoshioka, Kayo et al. 1997)。その病型は、遺伝形式、発症時期、肥満
の有無、インスリン抵抗性の有無などの点において、ヒト MODY(Maturity Onset Diabetes
of Young)と類似する。その責任遺伝子は D7Mit258 への強い連鎖を認め、当時は Mody と
命名されたが(Kayo and Koizumi 1998)、その後 7 番染色体に存在する Insulin2 遺伝子で
ある事が明らかになった(Wang, Takeuchi et al. 1999)。
AKITA マウスにおける糖尿病発症の要因は主にβ細胞の破壊に伴うインスリンの絶対的
不足によるものと考えられる。ヘテロの Insulin2 遺伝子変異マウス(Ins2WT/C96Y) におい
ては、4 週齢ですでにランゲルハンス島の明らかなβ細胞密度の低下が認められる(Figure
1)(Yoshioka, Kayo et al. 1997; Wang, Takeuchi et al. 1999)。しかし糖尿病発症前か
ら発症後の全経過において、ランゲルハンス島へのリンパ球の浸潤や炎症反応を示す所見

14
が認められない点で、同じβ細胞の減少を認める 1 型糖尿病とは発症の機序が異なる。β
細胞密度の低下については雄と雌では差がはっきりしており、インスリン陽性を示すβ細
胞領域の減少は、雄ではより著明である。
電子顕微鏡で AKITA マウスのランゲルハンス島を観察すると、Ins2WT/C96Y において既に出
生時からβ細胞のミトコンドリアの膨張や、小胞体・ゴルジ体の増加が認められ、2 週齢の
時点でその傾向がより強くなっている。また、同時に分泌細胞の部分的脱顆粒を認める。
一方α細胞については、グルカゴンの含量や微細構造について糖尿病完成以後においても
正常に保たれ、Ins2WT/WT と比較して明瞭な差は認めない。
このような AKITA マウスにおけるβ細胞減少は、アポトーシスによって起こる。AKITA
マウスでは、Inslin2 遺伝子の Cys 96 Tyr 変異が認められるが、この残基はプロインスリ
ン 2 の 2 か所のジスルフィド結合のうちの1つを司っている。遺伝子変異により、プロイ
ンスリン2のジスルフィド構造が変化し、構造異常を認めるプロインスリン2が産生され
る。このプロインスリン2が小胞体に保持されることによって、小胞体ストレスが惹起さ
れ、その結果β細胞のアポトーシスが生じると考えられている。実際に、親泊らは小胞体
シャペロンであるBipと小胞体ストレス関連のアポトーシス因子であるChopが AKITAマウ
スの膵臓で誘導されること、Chop 遺伝子を破壊することで AKITA マウス(Ins2WT/C96Y)のβ
細胞の破壊が抑制され、糖尿病発症が遅くなることを確認した(Oyadomari, Koizumi et al.
2002)。また Allen らは、AKITA マウスの膵臓において、小胞体ストレスが生じる際に、小
胞体の蛋白分解システムを構成する HRD1 が上方制御されることを示した。この変化は、変
異蛋白の分解を促進することによってβ細胞を過度の小胞体ストレスおよび細胞死から守
るためであると考えられている。(Allen, Nguyen et al. 2004)。
以上のように、AKITA マウスは、β細胞数の減少によるインスリン分泌不全により、ヒト
MODY に類似した病型を示す糖尿病モデルである。したがって肥満を来さずして糖尿病を発
症する。糖尿病発症前後の体重変化をみてみると、18 週齢以降 Ins2WT/C96Y は体重増加を認
めなくなり、30 週齢までに体重減少を認めるようになる(Yoshioka, Kayo et al. 1997) 。
血漿中のインスリン濃度は雄、雌ともに正常同胞に比較し、7 週齢の時点で著しい低値を示

15
す(Yoshioka, Kayo et al. 1997)。Hong らは 13 週齢の AKITA マウスを用いて、インスリン
抵抗性が生じることを報告している(Hong, Jung et al. 2007)。
このようなAKITAマウスの病態はその遺伝子型、性別により異なることが分かっている。
ホモの Insulin2 遺伝子変異マウス(Ins2-/-)は、生後 2 週までには明らかな高血糖を示し、
成長不全をきたして生後 3 ヵ月までに死亡する。組織学的には生後直後にすでにランゲル
ハンス島の明らかなβ細胞減少が認められる(Kayo and Koizumi 1998; Oyadomari, Koizumi
et al. 2002)。一方、ヘテロの Insulin2 遺伝子変異マウス(Ins2WT/C96Y)では 3 から 4週齢
で高血糖や多飲多尿(Breyer, Bottinger et al. 2005)、13 週齢で心筋肥大(心室中隔およ
び左心室後壁の肥厚)(Hong, Jung et al. 2007)、20 週齢の時点で糖尿病性の瀰漫性の糸
球体硬化像、クレアチニンクリアランスの低下などの腎機能異常といった合併症を生じる
(Haseyama, Fujita et al. 2002)が、その 50%生存率は雄で 305 日(野生型マウス(Ins2WT/WT)
の雄では約 690 日)とホモに比して長い。これらの症状は一般的に雄の方が雌よりも重症
であり、雄では 5 週齢の時点で血糖値が 16.7mmol/L を超える高血糖を示す事がほとんどで
ある。一方、雌では耐糖能低下があるものの 30 週齢の時点でも高血糖をきたさない個体も
まれではなく、(日本エスエルシー株式会社、2003 動物実験データ集)生後 370 日までのフ
ォローでの 50%生存率はほぼ 100%と正常同胞と有意差はない(Yoshioka, Kayo et al.
1997)。糸球体の組織学的変化および臨床検査とも雄と比較して軽症である(Haseyama,
Fujita et al. 2002)。
このように AKITA マウスは肥満を合併せずに糖尿病を発症すること、アミノ酸代謝に影響
を及ぼすインスリンがβ細胞減少によって減少することから、糖尿病の発症前後および糖
尿病の進行によるアミノ酸変化を検出するのにふさわしい疾患モデルであるといえる。
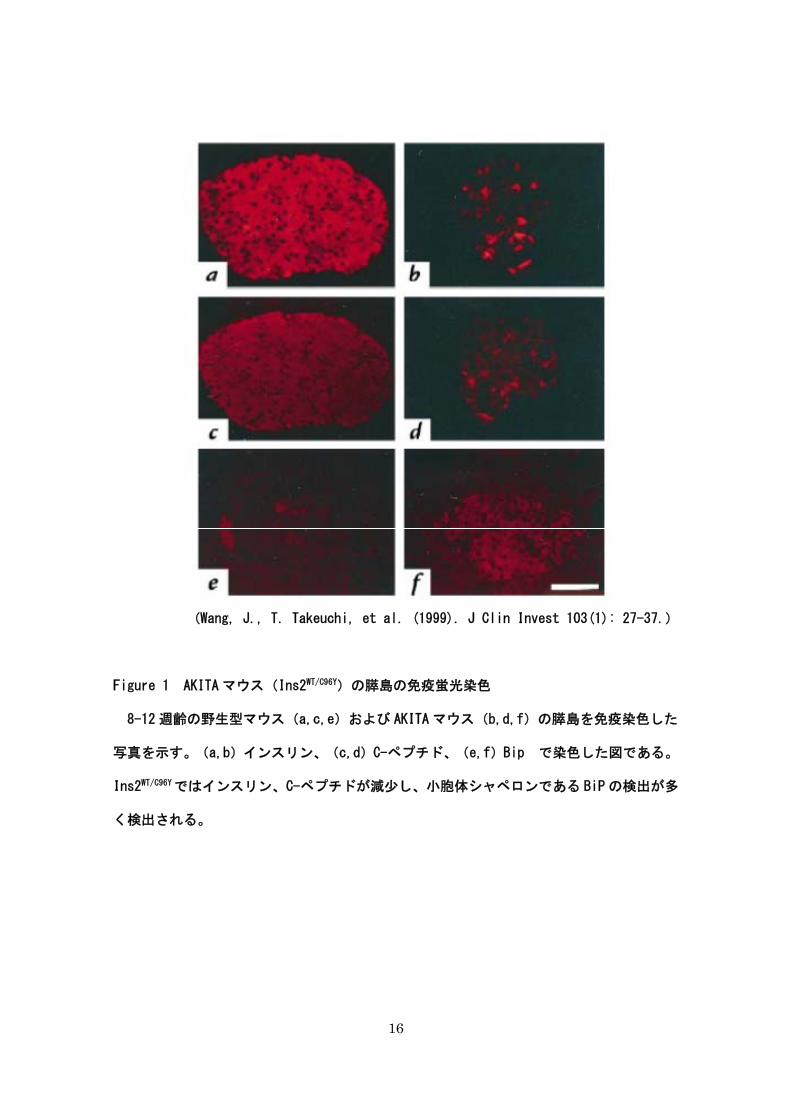
16
(Wang, J., T. Takeuchi, et al. (1999). J Clin Invest 103(1): 27-37.)
Figure 1 AKITA マウス(Ins2WT/C96Y)の膵島の免疫蛍光染色
8-12 週齢の野生型マウス(a,c,e)および AKITA マウス(b,d,f)の膵島を免疫染色した
写真を示す。(a,b)インスリン、(c,d)C-ペプチド、(e,f)Bip で染色した図である。
Ins2WT/C96Yではインスリン、C-ペプチドが減少し、小胞体シャペロンである BiP の検出が多
く検出される。

17
6-(3) アミノ酸分析の発展
1958 年、Stein や Moore らによって自動アミノ酸分析システムが確立された。アミノ酸
はアミノ基とカルボキシル基を同時に持つ物質であるため、溶離液の pH を変化させること
で、イオン交換樹脂に対する選択性を容易に変えることができる。Stein らはこの性質を利
用し、イオン交換樹脂カラムを用いて、溶出時間の差によるアミノ酸の分離に成功した。
分離されたアミノ酸を、ニンヒドリン試薬を用いて発色させ、比色法によって定量するこ
とができるようになった。その後、高速液体クロマトグラフィー(HPLC)のポンプ機能の
漸進的な改善やより効果的な溶出能をもつイオン交換樹脂の開発も進み、現在では体液中
のアミノ酸の測定が可能なものになっている(Shimbo, Oonuki et al. 2009)。
陽イオン交換樹脂カラム固定相とニンヒドリン発色を利用したポストカラム誘導体化
HPLC法は、アミノ酸分析の中で最も代表的な方法の一つで、臨床検査において最も普及し
ている方法である。ニンヒドリン比色法は、至適濃度内では優れた定量精度を発揮する。
その一方で、至適濃度が高く、その範囲も狭いため、試料中に様々な濃度のアミノ酸が含
まれる場合に、様々な希釈倍率の試料を用意する必要がある。さらに、アミノ酸分析に必
要な分析時間が最低でも110分と非常に長い(Japanese Patent A-2002-71660)。
アミノ酸の高感度分析を行う場合には、イオン交換樹脂カラムを用いて試料中のアミノ
酸を分離する前に、分析可能な蛍光誘導体にアミノ酸変換するプレカラム誘導体化HPLC法
を用いる。この方法で用いられる誘導体化試薬の開発は1970年代後半頃から盛んに行われ、
代表的なものに、6-アミノキノリル-N-ヒドロキシスクシニルイミド(AQC)、9-フルオレ
ニルメチルクロロギ酸(FMOC-Cl)、7-フルオロ-4-ニトロベンゾ-2-オキサ-1,3-ジアゾー
ル(NBD-F)などがある。誘導体化試薬が開発された事で、ニンヒドリン比色法と比較して
検出感度が高くなったため、微量な検体でもアミノ酸の定量が可能となった。しかし、こ
の方法でも分析時間は大きく変化することはなかった。
2003年、Piraudらは先天代謝異常診断に必要となるアミノ酸を含む79種類の分子を、
ESI-MS/MSを用いて分析することに成功した(Piraud, Vianey-Saban et al. 2003)。しかし、
この方法では同じ分子量の物質(例えばロイシンとイソロイシン)を分離して別々に測定

18
することが困難であった。その後、2005年には移動相にイオンペア試薬としてペルフルオ
ロカルボン酸類を使用し、逆相高速液体クロマトグラフィーおよびESI-MS/MSを用いて測定
する方法が開発された。この方法では、76種類のアミノ酸を15分で測定することに成功し
ている。しかし、アミノ酸を溶出した後は次の検体を測定するために流路の洗浄を行わな
ければならず、さらに15分ほど時間が必要であった(Piraud, Vianey-Saban et al. 2005)。
2009年、新保らは、さらに迅速に多種のアミノ酸分析を可能にするために新たな誘導化
試薬を開発した。彼らは、3 -アミノピリジル-N - ヒドロキシスクシンイミジルカルバメ
ート(APDS)によるプレカラム誘導体化を行い、HPLC-ESI-MS/MSを用いて100種類以上のアミ
ノ酸を10分程度の時間で測定することに成功した(Shimbo, Oonuki et al. 2009)。この方
法を用いることでこれまでよりも微量検体での測定が可能となった。
このようにアミノ酸分析にはさまざまな方法があるが、現在臨床の場で最も使用されて
いるのはニンヒドリン比色法である。しかし、必要とする検体量も多く対象となる疾患が
限られるため、実際には測定する機会はさほど多くないのが現状である。今後、
HPLC-ESI-MS/MSのように質量分析計を用いたアミノ酸分析が普及することで、わずかな負
担で疾病罹患の可能性や臓器障害の程度を把握することが可能になるなど、より膨大な情
報を得ることが期待できる。
6-(4) 本研究の目的
これまで述べてきたように、インスリン活性の変化が糖代謝だけでなく蛋白質やアミノ
酸代謝にも影響を及ぼすことが知られている(Hebert and Nair 2010)。また、アミノ酸プ
ロファイルに対する糖尿病発症の影響について過去にも多く論じられているが、一貫した
結果が得られていないのが現状である。また、過去の報告の中には、疾患特異的アミノ酸
プロファイルの変化を継時的に検証したものはほとんど認められていない。
したがって、本研究では、1 型糖尿病のアミノ酸プロファイルに焦点を置き、1 型糖尿病
モデルマウスを用いて、継時的なアミノ酸プロファイルの変化を追跡し、1 型糖尿病におけ

19
るアミノ酸変化の正確な評価を行うことを目的とした。さらに、糖尿病の進行に伴い変化
するアミノ酸を見出すことにより、アミノ酸分析を用いた糖尿病の疾患評価の可能性を示
すことを目的に実験を行った。具体的には、AKITA マウスの雄を対象として、血糖と血漿中
アミノ酸濃度を同時に測定し、健常状態から高血糖状態に至る時期、そして合併症を引き
起こす時期までのアミノ酸プロファイルの変化を検証した。HPLC-ESI-MS によるアミノ酸
分析を行い、マウスを犠牲にすることなく 3から 20 週齢までのアミノ酸プロファイルの変
化を追跡した。

20
7. 実験方法
7-(1) 実験対象動物および飼育環境
実験対象は雄の AKITA マウスとした。AKITA マウス(C57BL/6-Ins2 AKITA /J)は Jackson
Laboratory から購入し、熊本大学生命資源研究・支援センターの動物資源開発研究施設に
おいて繁殖を行った。対象動物への実験手技については、熊本大学の動物資源開発研究部
門の認可を得た。マウスの飼育は、床敷きに木くずを用いたケージの中で行った。飼育室
は、湿度は 50-70%、室温は 20-24℃、照明 12 時間(7:00-19:00)で維持された。マウ
スは 4 週齢で離乳した。飼料(CE-2; Clea Japan, Inc., Tokyo, Japan)、水分は自由摂取
とした。CE-2 は、12.6%の脂肪、29.3%の蛋白質、58.3%の可溶無窒素物、100g当たり
343.1kcal のエネルギーを含有する。
7-(2) 食餌摂取量の測定
マウス飼育用のケージを使用し、対象となるマウスがケージあたり 1 匹となるよう管理
した。ケージに補給されている飼料の重量を 24 時間間隔で 2 回測定し、その差を 1 日の食
餌摂取量とした。同一マウスでの継時的フォローは行っていない。
7-(3) 血漿中インスリン濃度の測定
EDTA-2Na (37.5mg/mL)溶液を少量含んだキャピラリーチューブ(Ringcaps; Hirschmann
Laborgeraete GmbH & Co. KG., Germany)を用いて、tail bleeding による血液採取を行
った。採取した血液量は 100μL で、EDTA-2Na の終濃度が 1.5 mg/mL となるよう設定した。
インスリン濃度測定のために、採取した血液を 1500g、10 分、4℃の条件で遠心分離し血漿
を採取した。採取した血漿は、インスリン濃度測定を行うまで-80℃下で冷凍保存した。イ
ンスリン濃度測定は、Insulin Mouse ELISA Kit Ultrasensitive (Mercodia, Uppsala,
Sweden) を用いて、既存のプロトコールに準じて行った。同一マウスでの継時的フォロー
は行っていない。

21
7-(4) 遺伝子型決定
マウスの遺伝型決定は以前の報告に準じて行った(Wang, Takeuchi et al. 1999)。概要
を以下に示す。マウスの尾を採取し、フェノール・クロロホルムを用いて DNA 抽出を行っ
た。採取した DNA を使用して、Insulin2 遺伝子の制限酵素断片長多型分析を行った。PCR
(polymerase chain reaction)を行って Insulin2 遺伝子を増幅する際に、以下のプライ
マーを用いた。
forward: 5’-TGCTGATGCCCTGGCCTGCT-3’
reverse: 5’-TGGTCCCACATATGCACATG-3’
制限酵素 Fnu 4HI を用いて生成された PCR 産物を切断処理した。次に、電気泳動を行い、
エチジウムブロマイド(Nacalai Tesq)染色で可視化した。この結果で、ホモの Insulin2
遺伝子変異マウス(Ins2C96Y/C96Y)、ヘテロの Insulin2 遺伝子変異マウス(Ins2WT/C96Y)、野生
型マウス(Ins2WT/WT)というように、マウスのグループ分けを行った。
7-(5) 体重測定および血液採取プロトコール
体重、血糖、血漿中アミノ酸濃度の測定は、同一マウスを用いて経時的に行った。Ins2WT/C96Y
13 匹、Ins2WT/WT 12 匹を対象に体重測定および血液採取を 3週齢から 1週間間隔で行い、20
週齢まで継続した(Figure 2)。
毎回の血液採取は 2 時間絶食後、11:00 から 12:00 の間に行った。血液採取時のマウス
の苦痛の軽減および迅速な血液採取を行うために、3%イソフルレンを用いてマウスに対し
吸入麻酔を行った。麻酔後、ただちにマウス用の体重測定器(Sartorius Mechatronics,
Tokyo, Japan)を使用して体重測定を行い、tail bleeding にて血糖測定および検体採取を
行った。まず、固定化酵素電極法(Glutest Ace R; Arkray Factory, Inc., Japan)を用
いて血糖を測定した。次に、EDTA-2Na (37.5mg/mL)溶液を少量含んだキャピラリーチュ
ーブ(Ringcaps; Hirschmann Laborgeraete GmbH & Co. KG., Germany)を用いて血液を採
取し、直ちに氷冷した。貧血を防ぐために、採取する血液量を 5 週齢以上のマウスでは 100

22
μL、4 週齢以下のマウスでは 75μL と設定した。EDTA-2Na の 終濃度が 1.5 mg/mL となる
ように検体採取を行った。アミノ酸濃度測定のために、採取した血液を 1500g、10 分、4℃
の条件で遠心分離し、血漿のみをアミノ酸分析を行うまで、-80℃下で冷凍保存した。
7-(6) アミノ酸分析
アミノ酸分析は新保らによって確立された方法を用いて行った(Shimbo, Oonuki et al.
2009)。概要を以下に示す。サンプルの調整の前に、測定対象となる血漿は50%アセトニト
リル下で除蛋白した。3 -アミノピリジル-N - ヒドロキシスクシンイミジルカルバメート
による誘導体化に引き続き、血漿中アミノ酸濃度の測定をHPLC-ESI-MSを用いて行った。MSQ
Plus LC/MS system (Thermo Fischer Scientific, Waltham,MA, USA)については、選択イ
オン検出法としてpositive ionization モードを使用した。データの収集および解析につ
いてはXcalibur (Salek, Maguire et al.) version 1.4 SR1 software を使用して行われ
た。HPLCはL-2100 (ポンプ)、L-2200 (autosampler)および L-2300 (カラムオーブン)
(Hitachi High-Technologies Corporationn, Tokyo,Japan)で構成された。 カラムは
Inertsil C8-3 column (100 mm × 2.1 mm, 3 μm) (GL Sciences Inc., Tokyo, Japan) を
使用した。移動相はA液(25 mM 蟻酸アンモニウム溶液)およびB液(超純水:アセトニト
リル=40:60)を使用した。
7-(7) 統計学的分析
体重、血糖値、血漿中アミノ酸濃度は平均±標準誤差で表記した。グループ間の経時的
変化を比較するため二元配置分散分析を採用し、多重比較は Bonferroni 補正を行った。P <
0.05 をもって有意差ありと判定した。血糖値の上昇に連動して血漿中濃度が変化するアミ
ノ酸を検証するために、ピアソンの相関係数を距離尺度として群平均法を用いてクラスタ
ー分析を行った(Noguchi, Sakai et al. 2003)。

23
クラスター分析の結果抽出されたアミノ酸について、健常状態を基準とした高血糖状態
における血漿中アミノ酸濃度の評価を行うために、3 週齢時のアミノ酸濃度に対する各週齢
のアミノ酸濃度のパーセント比を表示した。

24
Figure 2 AKITA マウスにおける血漿中アミノ酸分析のプロトコール

25
Figure 2 AKITA マウスにおける血漿中アミノ酸分析のプロトコール
(A)血液採取のプロトコール
血液採取は 11 時から 12 時までの間に行った。血液採取の前に 2 時間の絶食を行った。
血液採取を安定かつ迅速に行うため、対象マウスに対し 3%イソフルレンを用いて吸入麻酔
を行った。体重測定後、tail bleeding による血液採取のためにメスで尾を切った。その
後、素早く血糖を測定した。次に、EDTA-2Na 溶液を少量含んだキャピラリーチューブを用
いて血液を採取し、エッペンドルフチューブに血液を移して4℃に保管した。血漿を採取
するため、1500g、10 分、4℃の条件で遠心分離を行った。採取された血漿は、HPLC-ESI-MS
アミノ酸分析を行うまで、-80℃下で冷凍保存された。
(B)血液採取のスケジュール
血糖上昇前から濃度変化がおこるアミノ酸を検証するため、3 週齢の時点でマウスの血液
採取を開始した。貧血を防ぐため、100μL の血液を週に1回採取した。糖尿病発症後のア
ミノ酸プロファイルの変化を検証するため、20 週齢まで血液採取を継続した。

26
8. 実験結果
8-(1) 遺伝型決定
対象となる全てのマウスに、Insulin2 遺伝子の制限酵素断片長多型分析を行った。変異
遺伝子の PCR 産物は制限酵素 Fnu 4HI を用いても切断されず、280bp で検出された。一方、
野生型の遺伝子は切断されて140bpで検出された。この結果をもとに、Ins2C96Y/C96Y、Ins2WT/C96Y、
Ins2WT/WTのグループに分けた(Figure 3)。
8-(2) AKITA マウスにおける血糖の推移
Ins2C96Y/C96Y、Ins2WT/C96Y、Ins2WT/WTの 3 つのグループにおける血糖の推移の差を検証した。
全ての Ins2C96Y/C96Y は 3 週齢の時点で高血糖であった。血糖上昇前の血漿中アミノ酸濃度
を測定することができなかったため、Ins2C96Y/C96Yは研究対象から除外した。
3 週齢時、Ins2WT/C96Y と Ins2WT/WT の間で、血糖値の有意差を認めなかった。Ins2WT/C96Y に
おいて、4 週齢までに血糖上昇が始まり、7 週齢頃まで持続した。その後、血糖値の変化は
あまり認められず、25 から 30mmol/L 程で推移した。一方、Ins2WT/WT は 20 週齢まで血糖値
の変化はほとんどなかった。4 週齢以降は、Ins2WT/WT と比較して Ins2WT/C96Y において有意に
血糖値が高かった(Figure 4)。
8-(3) AKITA マウスの自然経過
肥満があればインスリン抵抗性を生じる可能性があることを考慮し、Ins2WT/WT と
Ins2WT/C96Y で体重を比較した。幼齢では、Ins2WT/WT と Ins2WT/C96Y の体重に差は認められなか
った。体重の差は徐々に大きくなり、Ins2WT/C96Y において 15 週齢以降では有意に軽量であ
った(Figure 5)。
次に Ins2WT/WT と Ins2WT/C96Y において生存率を比較した。Ins2WT/WT は 20 週齢まですべての
マウスが生存したが、Ins2WT/C96Y では 13 匹中 4匹のマウスが死亡した(Figure 6)。

27
体重に対する影響がありアミノ酸摂取に関する重要な因子ともなるため、食餌摂取量の
比較を行った。食事摂取量も、幼齢では Ins2WT/WT と Ins2WT/C96Y の間に差を認めなかった。
しかし、高血糖が持続した Ins2WT/C96Y では食事摂取量は減少していた(Figure 7)。
インスリンはタンパク質合成を促進する作用があり、アミノ酸代謝に影響を及ぼす。血
漿中インスリン濃度は、Ins2WT/WT と比較して Ins2WT/C96Y では大変低い値で推移した(Figure
8)。
8-(4) AKITA マウスにおける血漿中アミノ酸濃度の推移
血液採取が困難なため血漿分離ができない、対象マウスが死亡したなどの理由で一部血
液採取ができなかった。分析可能であった各週齢での検体数を Table 3に示す。
糖尿病の発症が及ぼす血漿中アミノ酸濃度への影響を検証するため、Ins2WT/WT と
Ins2WT/C96Yの血漿中アミノ酸の濃度推移の差を検証した。糖尿病を発症しない Ins2WT/WT にお
いても、一部のアミノ酸における血漿中濃度は週齢に影響された(Figure 9,10)。週齢の
影響を考慮し、Ins2WT/WT と Ins2WT/C96Y の血漿中アミノ酸の濃度推移を比較するため、二元
配置分散分析を行って遺伝子型と週齢の交互作用がある(P < 0.01, age-by-genotype
interaction)と認められるアミノ酸を、二群間で推移に有意差のあるアミノ酸として抽出
した (Figure 10)。
Ins2WT/WT と比較して、血漿中のバリン、ロイシン、イソロイシン、総分枝鎖アミノ酸、
アラニン、シトルリン、プロリンの各濃度が Ins2WT/C96Y において有意に高値であった。3 週
齢の時点では、上記アミノ酸の血漿中濃度は Ins2WT/WT と Ins2WT/C96Y を比較して差を認めな
かった。高血糖が持続することによって、Ins2WT/WT と Ins2WT/C96Y の上記アミノ酸の血漿中
濃度の差は徐々に大きくなっていった(Figure 10)。
また、糖尿病に罹患した場合に低下するパラメータである血漿トリプトファン比
(Trp/(Ile+Leu+Val+Phe+Tyr))についても二群間の推移の差を検証した。遺伝子型と週齢の
交互作用がある(P < 0.01, age-by-genotype interaction)と認められ、Ins2WT/C96Yにお
いて有意に低値であった(Figure 11)。

28
8-(5) 糖尿病の発症早期に濃度変化を認めるアミノ酸
糖尿病発症早期に濃度変化を認めるアミノ酸を見出すため、血糖の上昇に連動して濃度
変化するアミノ酸を検証した。本研究では、血糖とアミノ酸の各パラメータ間におけるピ
アソンの相関係数を距離尺度としたクラスター分析を選択した。得られたデンドログラム
の結果、バリン、ロイシン、イソロイシン、総分枝鎖アミノ酸、アラニンが血糖と同じグ
ループに分類された。これらのアミノ酸は、血糖上昇に連動して濃度上昇がおこるアミノ
酸である可能性が示唆された。
次に、バリン、ロイシン、イソロイシン、総分枝鎖アミノ酸、アラニンの各アミノ酸に
ついて、健常状態と高血糖状態でおこる血漿中濃度の相対的変化を検証した。3 週齢では、
対象となったすべてのマウスで血糖は正常であった。そのため、3週齢時の各アミノ酸濃度
を「基礎値」と定義した。また、アミノ酸間で比較を行うための標準化の手段として、「基
準値」を基準に各週齢のアミノ酸濃度のパーセント比を算出する方法を用いた。その結果、
5 週齢の時点で、血漿中のアラニン濃度は、Ins2WT/C96Y において「基礎値」より増加し、
Ins2WT/WT において「基礎値」より減少した。Ins2WT/WT と Ins2WT/C96Y を比較して差が最も大
きいのはアラニンであった(Figure 13)。
さらに、バリン、ロイシン、イソロイシン、総分枝鎖アミノ酸では、明確に差が出てく
るのは 11 週齢(血糖が上昇した後、25-30mmol/L 程度の高血糖が維持される時期)以降で
あった。また、プロリンとシトルリンもほぼ同時期に Ins2WT/WT と Ins2WT/C96Y の間で差が大
きくなった(Figure 14)。このように、ほとんどのアミノ酸は血糖上昇が認められてから
濃度変化を認めると結論付けられた。
29
Table 3 各週齢でアミノ酸分析を行った検体数
血漿中アミノ酸濃度を測定した検体数を示した。13匹の Ins2WT/C96Y と 12 匹の Ins2WT/C96Y
を対象として本研究は行われた。検体数の減少は、対象マウスが死亡した、あるいは血液
採取を失敗した事に起因する。

30
Figure 3 制限酵素断片長多型分析を用いた Insulin2 遺伝子の遺伝型決定
遺伝型決定のために、Insulin2 遺伝子における制限酵素断片長多型分析を行った。変異
遺伝子の PCR 産物(280bp)は Fnu4HI によって切断されない。一方、正常遺伝子(140bp)
は Fnu4HI によって切断される。制限酵素断片長多型分析によって、対象マウスを Ins2WT/WT、
Ins2C96Y/C96Y および Ins2WT/C96Y のグループに分けた。レーン 1 が Ins2WT/WT、 レーン 2 が
Ins2C96Y/C96Y、 レーン 3 が Ins2WT/C96Yである。
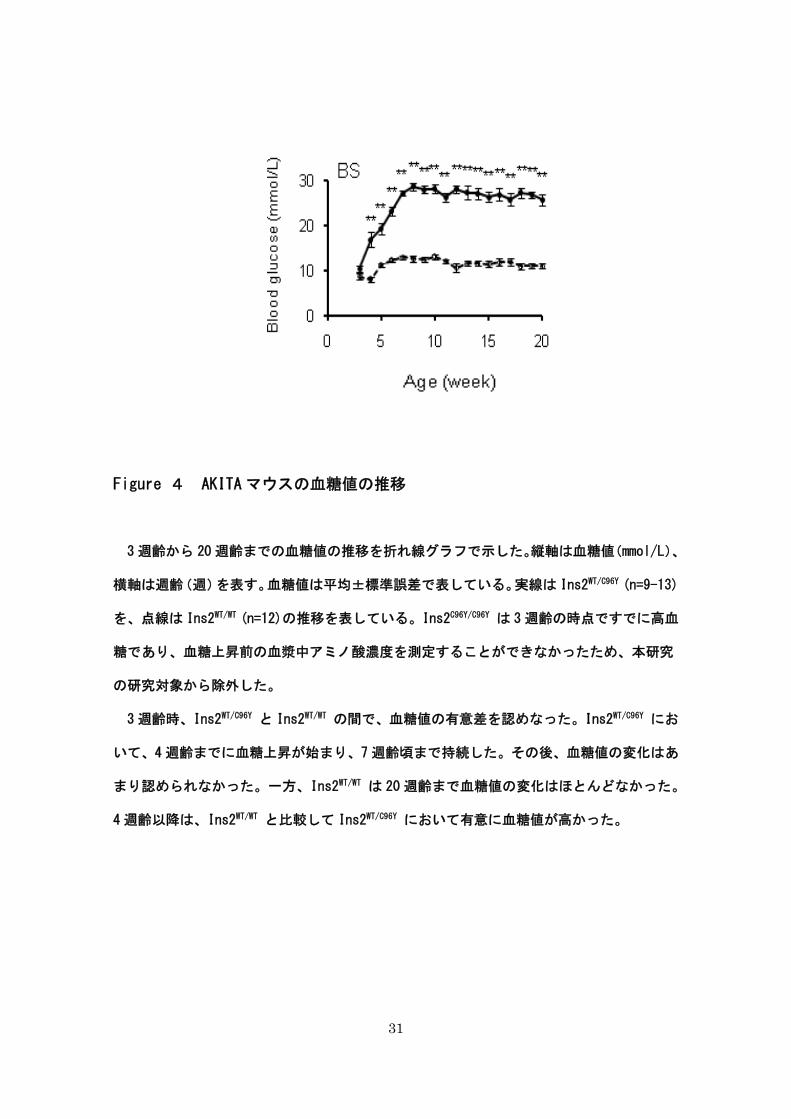
31
Figure 4 AKITA マウスの血糖値の推移
3 週齢から 20 週齢までの血糖値の推移を折れ線グラフで示した。縦軸は血糖値(mmol/L)、
横軸は週齢(週)を表す。血糖値は平均±標準誤差で表している。実線は Ins2WT/C96Y (n=9-13)
を、点線は Ins2WT/WT (n=12)の推移を表している。Ins2C96Y/C96Y は 3 週齢の時点ですでに高血
糖であり、血糖上昇前の血漿中アミノ酸濃度を測定することができなかったため、本研究
の研究対象から除外した。
3 週齢時、Ins2WT/C96Y と Ins2WT/WT の間で、血糖値の有意差を認めなった。Ins2WT/C96Y にお
いて、4 週齢までに血糖上昇が始まり、7 週齢頃まで持続した。その後、血糖値の変化はあ
まり認められなかった。一方、Ins2WT/WT は 20 週齢まで血糖値の変化はほとんどなかった。
4 週齢以降は、Ins2WT/WT と比較して Ins2WT/C96Y において有意に血糖値が高かった。

32
Figure 5 AKITA マウスの体重の推移
3 週齢から 20 週齢までの体重の推移を示した折れ線グラフを示した。縦軸は体重(g)、
横軸は週齢(週)を表す。体重は平均±標準誤差で表している。実線は Ins2WT/C96Y (n=9-13)
を、点線は Ins2WT/WT (n=12)の推移を表している。3 週齢の時点では、Ins2WT/C96Y と Ins2WT/WT
の間で体重の差はほとんど認められない。体重差は週齢を経るごとに徐々に広がっていっ
た。Ins2WT/WT と比較して、Ins2WT/C96Y は 15 週齢以降で有意に軽量であった。 ** P<0.01、
Ins2WT/C96Y vs. Ins2WT/WT。

33
Figure 6 Ins2WT/C96Y および Ins2WT/WT の生存率
20 週齢までの生存率をカプラン-マイヤー曲線で表した。縦軸は生存率(%)、横軸は週齢
(週)を示す。実線は Ins2WT/C96Y (n=13)の推移を示し、点線は Ins2WT/WT (n=12)の推移を示
す。全ての Ins2WT/WT は 20 週齢まで生存した。一方 Ins2WT/C96Y は 20 週齢までに 31%が死
亡した。
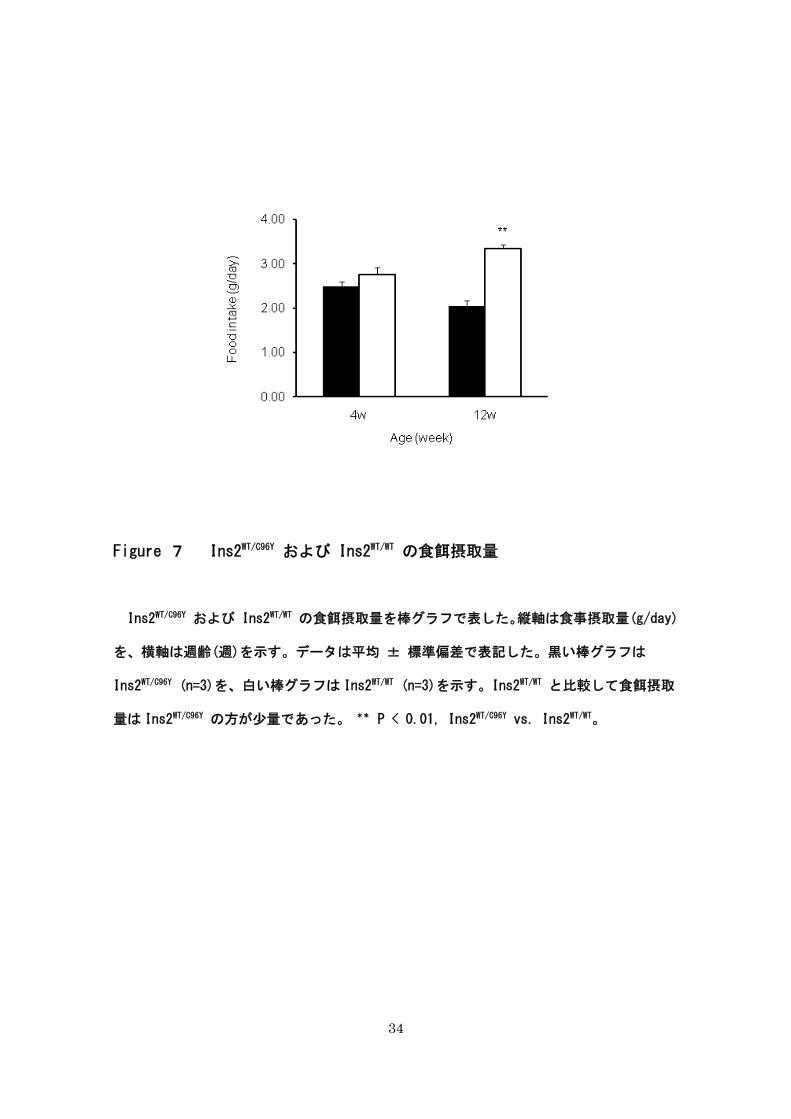
34
Figure 7 Ins2WT/C96Y および Ins2WT/WT の食餌摂取量
Ins2WT/C96Y および Ins2WT/WT の食餌摂取量を棒グラフで表した。縦軸は食事摂取量(g/day)
を、横軸は週齢(週)を示す。データは平均 ± 標準偏差で表記した。黒い棒グラフは
Ins2WT/C96Y (n=3)を、白い棒グラフは Ins2WT/WT (n=3)を示す。Ins2WT/WT と比較して食餌摂取
量は Ins2WT/C96Y の方が少量であった。 ** P < 0.01, Ins2WT/C96Y vs. Ins2WT/WT。
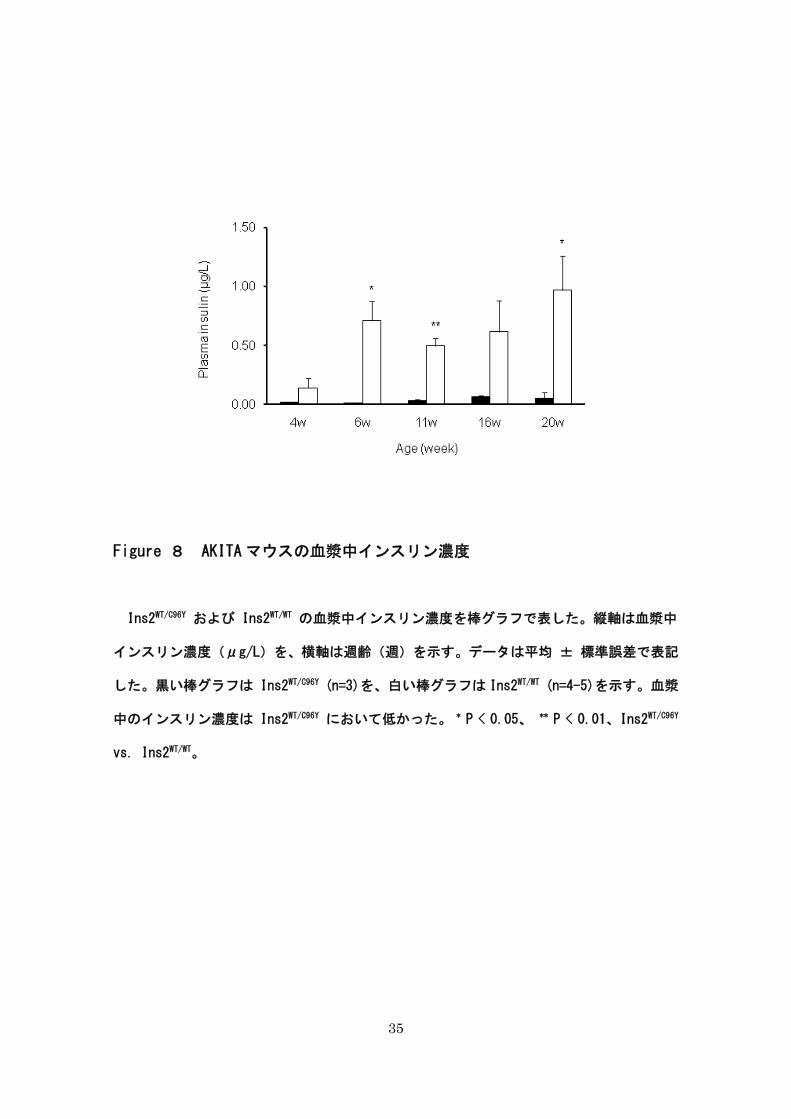
35
Figure 8 AKITA マウスの血漿中インスリン濃度
Ins2WT/C96Y および Ins2WT/WT の血漿中インスリン濃度を棒グラフで表した。縦軸は血漿中
インスリン濃度(μg/L)を、横軸は週齢(週)を示す。データは平均 ± 標準誤差で表記
した。黒い棒グラフは Ins2WT/C96Y (n=3)を、白い棒グラフは Ins2WT/WT (n=4-5)を示す。血漿
中のインスリン濃度は Ins2WT/C96Y において低かった。 * P < 0.05、 ** P < 0.01、Ins2WT/C96Y
vs. Ins2WT/WT。
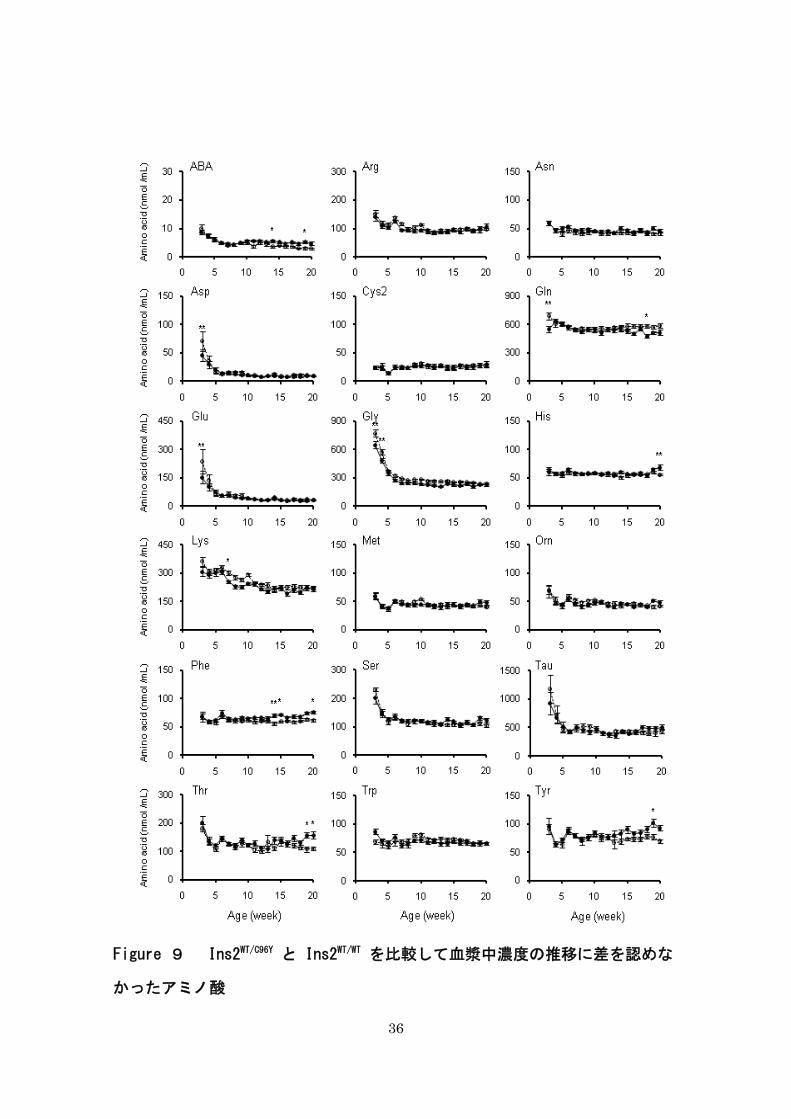
36
Figure 9 Ins2WT/C96Y と Ins2WT/WT を比較して血漿中濃度の推移に差を認めな
かったアミノ酸

37
Figure 9 Ins2WT/C96Y と Ins2WT/WT を比較して血漿中濃度の推移に差を認めな
かったアミノ酸
3 週齢から 20 週齢までの血漿中アミノ酸濃度の変化を折れ線グラフで表した。Ins2WT/C96Y
および Ins2WT/WT の血漿中アミノ酸濃度の変化を比較するために、二元配置分散分析を行っ
た。縦軸は血漿中アミノ酸濃度(nmol/mL)を、横軸は週齢(週)を示す。実線は Ins2WT/C96Y
(n=9-13)の推移を示し、点線は Ins2WT/WT (n=8-12)の推移を示す。データは平均 ± 標準誤
差で表記した。* P < 0.05、 ** P < 0.01、 Ins2WT/C96Y vs. Ins2WT/WT。

38
Figure 10 Ins2WT/C96Y と Ins2WT/WT を比較して血漿中濃度の推移に差を認めた
アミノ酸

39
Figure 10 Ins2WT/C96Y と Ins2WT/WT を比較し血漿中濃度の推移に差を認めたア
ミノ酸
3 週齢から 20 週齢までの血漿中アミノ酸濃度の変化を折れ線グラフで表した。(血糖が上
昇する時期と血漿中アミノ酸濃度の変化を観察するため Figure を併記している。)
Ins2WT/C96Y および Ins2WT/WT の血漿中アミノ酸濃度の変化を比較するために、二元配置分散
分析を行った。遺伝子型と週齢の交互作用がある(P < 0.01, age-by-genotype interaction)
と認められるアミノ酸を、Ins2WT/C96Y および Ins2WT/WT の間で推移に有意差のあるアミノ酸
として抽出した。縦軸は血漿中アミノ酸濃度(nmol/mL)を、横軸は週齢(週)を示す。実
線は Ins2WT/C96Y (n=9-13)の推移を示し、点線は Ins2WT/WT (n=8-12)の推移を示す。データは
平均 ± 標準誤差で表記した。* P < 0.05、 ** P < 0.01、 Ins2WT/C96Y vs. Ins2WT/WT。
Ins2WT/WT と比較して、血漿中のバリン、ロイシン、イソロイシン、総分枝鎖アミノ酸、
アラニン、シトルリン、プロリンの各濃度が Ins2WT/C96Y において有意に高値であった。3 週
齢の時点では、上記アミノ酸の血漿中濃度は Ins2WT/WT と Ins2WT/C96Y を比較して差を認めな
かった。11 週齢以降で統計学的に有意差を認め、Ins2WT/WT と Ins2WT/C96Y の血漿中濃度の差
は徐々に大きくなっていった。

40
Figure 11 Ins2WT/C96Y と Ins2WT/WT における血漿トリプトファン比の推移
3 週齢から 20 週齢までの Trp ratio の変化を折れ線グラフで表した。Ins2WT/C96Y および
Ins2WT/WT の血漿中アミノ酸濃度の変化を比較するために、二元配置分散分析を行った。血
漿トリプトファン比については、遺伝子型と週齢の交互作用がある(P < 0.01,
age-by-genotype interaction)と認められた。縦軸は血漿トリプトファン比を、横軸は週
齢(週)を示す。実線は Ins2WT/C96Y (n=9-13)の推移を示し、点線は Ins2WT/WT (n=8-12)の推
移を示す。データは平均 ± 標準誤差で表記した。* P < 0.05、 ** P < 0.01、 Ins2WT/C96Y vs.
Ins2WT/WT。Ins2WT/WT と比較して有意に低値であった。
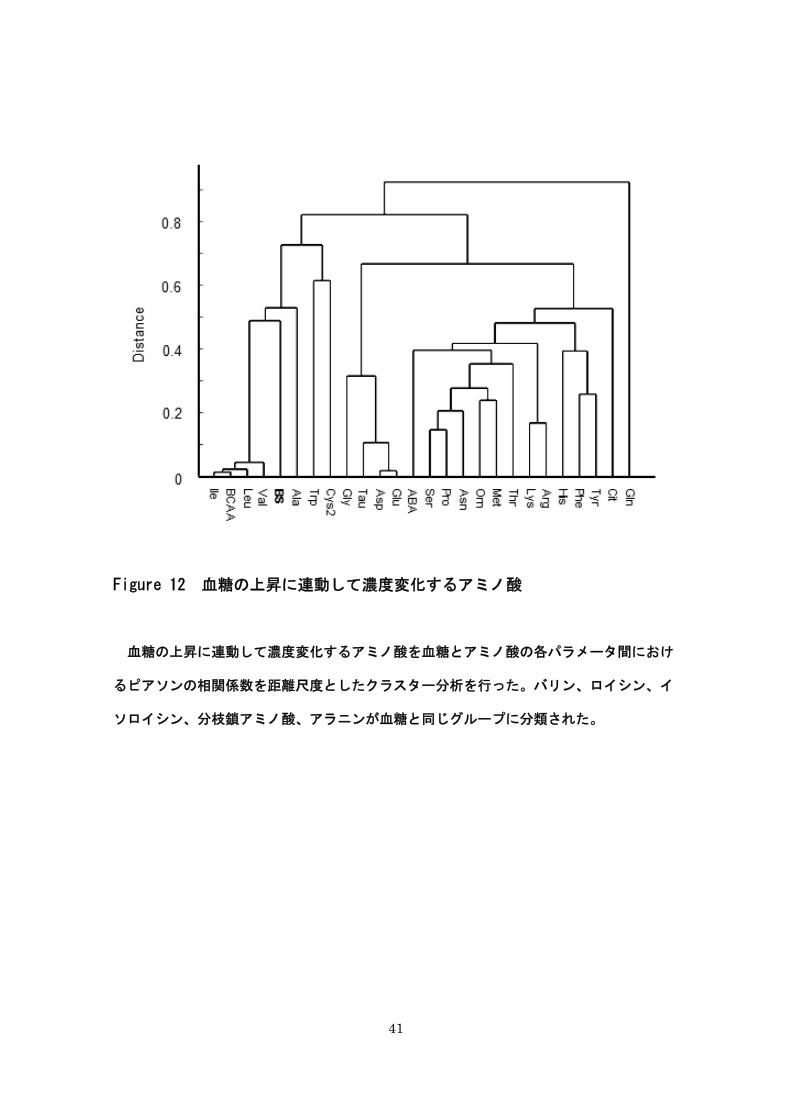
41
Figure 12 血糖の上昇に連動して濃度変化するアミノ酸
血糖の上昇に連動して濃度変化するアミノ酸を血糖とアミノ酸の各パラメータ間におけ
るピアソンの相関係数を距離尺度としたクラスター分析を行った。バリン、ロイシン、イ
ソロイシン、分枝鎖アミノ酸、アラニンが血糖と同じグループに分類された。
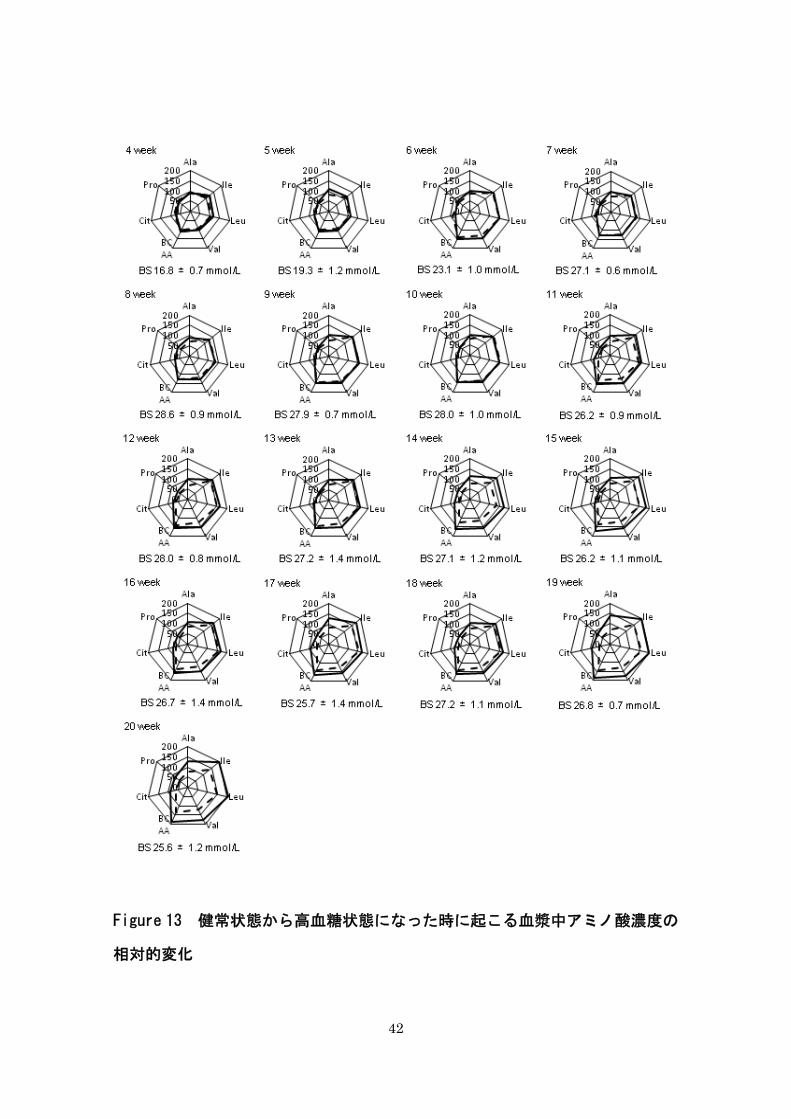
42
Figure 13 健常状態から高血糖状態になった時に起こる血漿中アミノ酸濃度の
相対的変化

43
Figure 13 健常状態から高血糖状態になった時に起こる血漿中アミノ酸濃度の
相対変化
まず、すべてのマウスで正常血糖を確認できた 3 週齢時のデータを「基礎値」と定義し
た。そして、4 週齢以降の各マウスの各アミノ酸のデータを「基礎値」で除し、得られたパ
ーセント比を相対変化の指標とした。
4 週齢から 20 週齢の血漿中アミノ酸の相対変化をレーダーチャートで表した。実線は
Ins2WT/C96Y (n=9-13)を示し、点線は Ins2WT/WT (n=8-12)を示す。Ins2WT/WT と Ins2WT/C96Y を比
較して、アラニンが 5週齢以降で差が大きくなっていった。一方、バリン、ロイシン、イ
ソロイシン、分枝鎖アミノ酸において、両グループの差が明確になるのは 11 週齢以降であ
った。

44
9. 考察
これまで、糖尿病発症によっておこる血漿中アミノ酸濃度変化については多数の報告が
ある。しかし、報告によって結果は一致していなかった。我々は、AKITA マウスを用いて健
常状態から高血糖状態に至るまでの血漿中アミノ酸濃度を継時的に検証し、血漿中アミノ
酸濃度に対する糖尿病の影響についてより詳細なデータを得ることができた。
糖尿病発症によって濃度変化のおこるアミノ酸はバリン、ロイシン、イソロイシン、総
分枝鎖アミノ酸、アラニン、シトルリン、プロリンであり、ほとんどのアミノ酸は血糖上
昇が認められてから濃度変化が起こると考えられた。
9-(1) 実験系
まず糖尿病の発症によっておこる血漿中アミノ酸濃度の変化をより詳細に検証するため、
糖尿病モデルマウスを対象マウスとして、同一マウスから定期的に採血を行って継時的に
血糖と血漿中アミノ酸濃度を測定した。本研究では AKITA マウスを採用したが、その理由
として遺伝型ごとに明確に臨床像が異なる事、Ins2WT/C96Y では 4-8 週齢と比較てき早期に
糖尿病を発症する事、肥満をきたさずインスリン分泌不足によって糖尿病を発症する事、
20 週齢までに様々な糖尿病による合併症がおこりうる事が挙げられる。つまり、比較的短
時間で血漿中のアミノ酸濃度に対する糖尿病の影響を観察することができ、血糖上昇前後
から高血糖状態、さらには合併症を発症する時期に至るまでの各病期でのアミノ酸プロフ
ァイルを検証することができると考えた。ただし AKITA マウスの場合、糖尿病発症の機序
から主に 1型糖尿病に類似した経過をたどる。
本研究で用いられた AKITA マウスは全て近交系である。さらに、継時的な採血について
もほぼ同条件で行われた。本研究で得られた結果は、過去の報告と比較しても血漿中アミ
ノ酸濃度に対する糖尿病の影響をより正確に評価したものと考えられる。

45
9-(2) 血漿中アミノ酸濃度に対する食餌摂取量の影響
食事摂取量は、高血糖が持続している Ins2WT/C96Y で少ないことが分かった。Ins2WT/WT と
Ins2WT/C96Y を比較して徐々に体重差が広がっている事実に合致する結果である。しかしこの
事実は、Ins2WT/C96Y でアミノ酸摂取量が減少していることを示す結果である。Ins2WT/C96Y に
おける血漿中アミノ酸濃度の上昇は、アミノ酸摂取量の増加に起因したものではないと考
えられた。
9-(3) 血漿中アミノ酸濃度に対するインスリンの影響
過去の報告によると、7週齢の時点で Ins2WT/WT と Ins2WT/C96Y の間で血漿中インスリン濃
度に有意差があることが示されている(Yoshioka, Kayo et al. 1997)。本研究においても、
血漿中のインスリン濃度は Ins2WT/C96Y において低い事が示された。インスリンは蛋白合成
を促進する働きがある。血漿中アミノ酸濃度が上昇している要因について、血漿中インス
リン濃度が低いため蛋白の異化が亢進している事が可能性の一つとして考えられる。この
事実を明確に確かめるには、トリメチルリジンのような異化に関連するマーカーを測定す
る必要があった。
なお、Ins2WT/WT と Ins2WT/C96Y の間で血漿中アミノ酸濃度に有意差が出てくるのは 11 週齢
以降であったことを考慮すると、インスリン濃度の低下による異化亢進のみでは血漿中の
アミノ酸濃度上昇について説明できないと考えられる。
9-(4) 糖尿病発症時のアラニンの代謝変化
アラニンは糖新生における主要な前駆体としてよく知られている。これまでの論文では、
血漿中アラニン濃度の変化に関する結論は報告ごとに一致していなかった(Vannini,
Marchesini et al. 1982; Robert, Beaufrere et al. 1985; Oresic, Simell et al. 2008)。
本研究の結果では、血漿中アラニン濃度は糖尿病発症により上昇し、すべてのアミノ酸の
中で血糖の影響を最も受けやすいことが示唆された。過去に、インスリン濃度よりも血糖
自体がアラニン代謝に対して重要な因子となることが報告されている(Shulman, Lacy et al.

46
1980; Robert, Beaufrere et al. 1985)。本研究の結果は、それを改めて支持する結果と
なった。
アラニンの血漿中濃度の上昇については、インスリンの不足による蛋白異化亢進や
Pyruvate dehydrogenase 活性の低下、分枝鎖アミノ酸のアミノ基転移の増加が、メカニズ
ムの一部として考えられる(Figure 14)。
9-(5) 糖尿病発症時の分枝鎖アミノ酸の代謝変化
1 型糖尿病で血漿中のバリン、ロイシン、イソロイシンの濃度が上昇することが知られて
いる(Vannini, Marchesini et al. 1982)。本研究でも同様の結果が認められた。過去の報
告では、1型糖尿病で血漿中の分枝鎖αケト酸濃度も上昇すること(Hutson and Harper
1981)、高ロイシン血症に対する分枝鎖αケト酸デヒドロゲナーゼの活性化が抑えられるこ
とが示されている(Aftring, Miller et al. 1988)。このメカニズムが血漿中の分枝鎖アミ
ノ酸濃度が上昇する要因の一つになりうると考えられた。また、インスリン不足による蛋
白異化亢進も濃度上昇の要因と考えられる(Figure 15)。
9-(6) 糖尿病発症時のプロリンの代謝変化
糖尿病では血清中の乳酸濃度が上昇する。乳酸は、Proline oxidase の活性を抑える働き
がある(Kowaloff, Phang et al. 1977)。さらに、糖尿病では細胞質の[NAD+]/ [NADH] 比
が低下するという報告がある(Ido, Kilo et al. 1997)。これにより、Pyrroline-5-
carboxylate reductase の活性が上昇する可能性が考えられる。糖尿病発症時における血漿
中のプロリン濃度の上昇については、少なくとも上記のメカニズムによって説明されると
考えられる(Figure 16)。
9-(7) 血漿中アミノ酸濃度に対する糖尿病合併症の影響
シトルリンは腎機能障害や尿細管機能障害における有用なマーカーとして報告されてい
る(Levillain, Parvy et al. 1997)。また、AKITA マウスでは 20 週齢までにクレアチニン

47
クリアランスの低下が起こることが知られている(Haseyama, Fujita et al. 2002)。本研
究において Ins2WT/C96Y で確認されたシトルリンの上昇について、腎機能障害や尿細管機能
障害を反映している可能性が考えられた。ただし明確に結論付けるには、腎機能に関する
マーカー(尿酸、クレアチニン、クレアチニンクリアランスなど)の観察が必要であった。
また、アミノ酸の血中濃度に影響を及ぼす因子として、尿中への排泄量が考えられる。
ケトアシドーシス状態で分枝鎖アミノ酸の排泄が亢進する(Szabo, Kenesei et al. 1991)。
この機序によって、血漿中の分枝鎖アミノ酸濃度が上昇する時期が遅くなっている可能性
が考えられる。
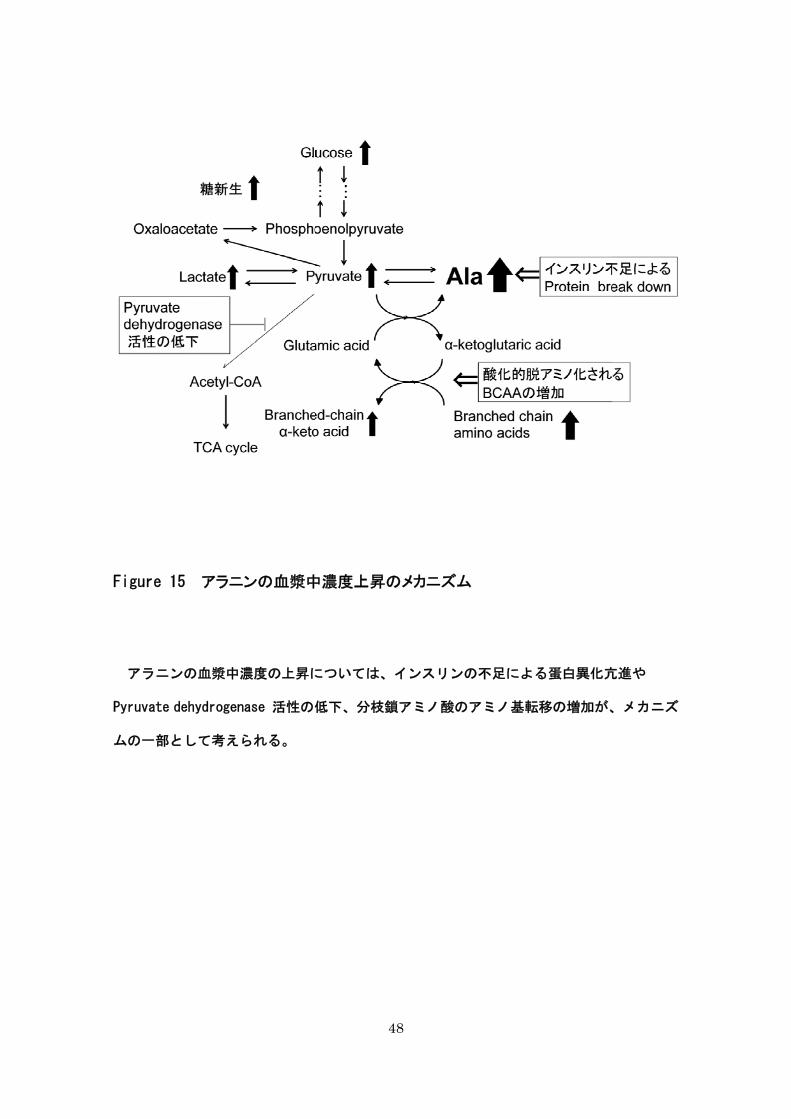
Figure 15 アラニンの血漿中
アラニンの血漿中濃度の上昇に
Pyruvate dehydrogenase 活性の低
ムの一部として考えられる。
48
中濃度上昇のメカニズム
については、インスリンの不足による蛋白異化亢
低下、分枝鎖アミノ酸のアミノ基転移の増加が、
亢進や
、メカニズ
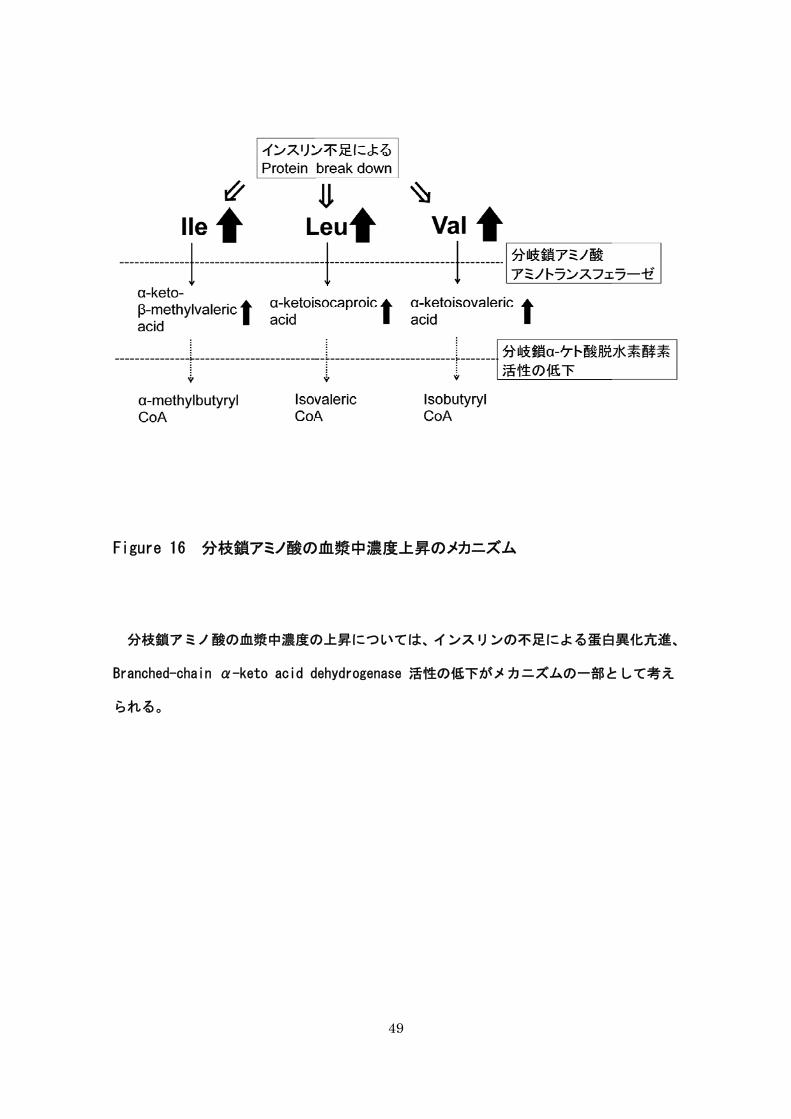
Figure 16 分枝鎖アミノ酸の
分枝鎖アミノ酸の血漿中濃度の
Branched-chain α-keto acid d
られる。
49
の血漿中濃度上昇のメカニズム
の上昇については、インスリンの不足による蛋白
dehydrogenase 活性の低下がメカニズムの一部と
白異化亢進、
として考え
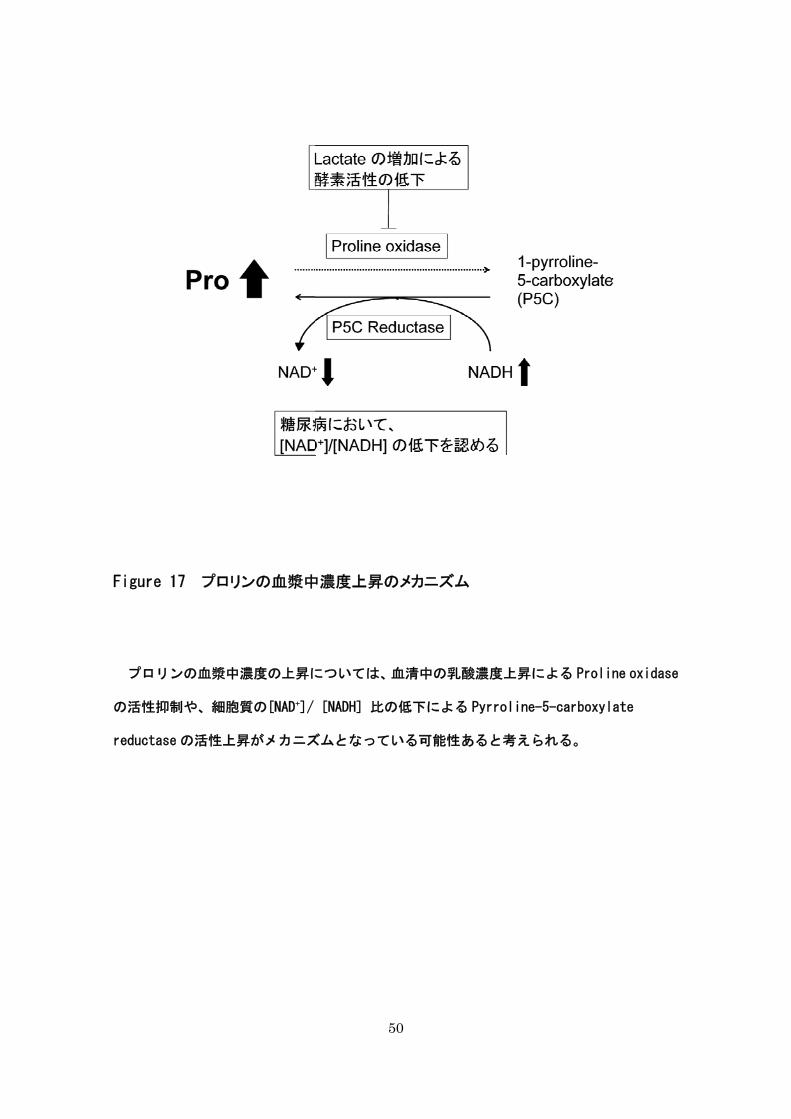
Figure 17 プロリンの血漿中
プロリンの血漿中濃度の上昇に
の活性抑制や、細胞質の[NAD+]/
reductase の活性上昇がメカニズ
50
中濃度上昇のメカニズム
については、血清中の乳酸濃度上昇による Proli
[NADH] 比の低下による Pyrroline-5-carboxyl
ズムとなっている可能性あると考えられる。
ne oxidase
ate

51
10. 結語
本研究で、糖尿病における時間依存性のアミノ酸プロファイルの変化を示した。アミノ
酸プロファイルは健常状態から高血糖状態へと推移する過程で変化した。ほとんどのアミ
ノ酸は血糖上昇の後に変化を認めた。さらに、糖尿病を発症した後も時間依存性にアミノ
酸プロファイルの変化は続いた。血漿中のアミノ酸濃度はインスリン活性のみならず、高
血糖の持続に関連した代謝酵素活性の変化、組織や臓器における微小環境の変化も影響し
ている可能性があると考えられた。経時的なアミノ酸分析は、病期や合併症の評価に有用
である可能性が示唆された。

52
11. 参考文献
Aftring, R. P., W. J. Miller, et al. (1988). "Effects of diabetes and starvation on skeletal
muscle branched-chain alpha-keto acid dehydrogenase activity." Am J Physiol 254(3 Pt 1):
E292-300.
Allen, J. R., L. X. Nguyen, et al. (2004). "High ER stress in beta-cells stimulates
intracellular degradation of misfolded insulin." Biochem Biophys Res Commun 324(1):
166-170.
Breyer, M. D., E. Bottinger, et al. (2005). "Mouse models of diabetic nephropathy." J
Am Soc Nephrol 16(1): 27-45.
Chevalier, S., S. C. Burgess, et al. (2006). "The greater contribution of gluconeogenesis
to glucose production in obesity is related to increased whole-body protein catabolism."
Diabetes 55(3): 675-681.
Consoli, A., N. Nurjhan, et al. (1990). "Mechanism of increased gluconeogenesis in
noninsulin-dependent diabetes mellitus. Role of alterations in systemic, hepatic, and
muscle lactate and alanine metabolism." J Clin Invest 86(6): 2038-2045.
Crandall, E. A. and J. D. Fernstrom (1983). "Effect of experimental diabetes on the levels
of aromatic and branched-chain amino acids in rat blood and brain." Diabetes 32(3):
222-230.
Felig, P., E. Marliss, et al. (1969). "Plasma amino acid levels and insulin secretion
in obesity." N Engl J Med 281(15): 811-816.
Haseyama, T., T. Fujita, et al. (2002). "Complications of IgA nephropathy in a
non-insulin-dependent diabetes model, the Akita mouse." Tohoku J Exp Med 198(4): 233-244.
Hebert, S. L. and K. S. Nair (2010). "Protein and energy metabolism in type 1 diabetes."
Clin Nutr 29(1): 13-17.

53
Herman, M. A., P. She, et al. (2010). "Adipose tissue branched chain amino acid (BCAA)
metabolism modulates circulating BCAA levels." J Biol Chem 285(15): 11348-11356.
Hong, E. G., D. Y. Jung, et al. (2007). "Nonobese, insulin-deficient Ins2Akita mice develop
type 2 diabetes phenotypes including insulin resistance and cardiac remodeling." Am J
Physiol Endocrinol Metab 293(6): E1687-1696.
Hutson, S. M. and A. E. Harper (1981). "Blood and tissue branched-chain amino and
alpha-keto acid concentrations: effect of diet, starvation, and disease." Am J Clin Nutr
34(2): 173-183.
Ido, Y., C. Kilo, et al. (1997). "Cytosolic NADH/NAD+, free radicals, and vascular
dysfunction in early diabetes mellitus." Diabetologia 40 Suppl 2: S115-117.
James Murray Luck, G. M., and Leonard Fisk Wilbur (1928). "The effect of insulin on the
amino acid content of blood " J. Biol. Chem. 77: 151-156.
Kayo, T. and A. Koizumi (1998). "Mapping of murine diabetogenic gene mody on chromosome
7 at D7Mit258 and its involvement in pancreatic islet and beta cell development during
the perinatal period." J Clin Invest 101(10): 2112-2118.
Kowaloff, E. M., J. M. Phang, et al. (1977). "Regulation of proline oxidase activity by
lactate." Proc Natl Acad Sci U S A 74(12): 5368-5371.
Leroux, L., P. Desbois, et al. (2001). "Compensatory responses in mice carrying a null
mutation for Ins1 or Ins2." Diabetes 50 Suppl 1: S150-153.
Levillain, O., P. Parvy, et al. (1997). "Amino acid handling in uremic rats: citrulline,
a reliable marker of renal insufficiency and proximal tubular dysfunction." Metabolism
46(6): 611-618.
Marchesini, G., G. P. Bianchi, et al. (1991). "Elimination of infused branched-chain
amino-acids from plasma of patients with non-obese type 2 diabetes mellitus." Clin Nutr
10(2): 105-113.

54
Morgan, M. Y., J. P. Milsom, et al. (1978). "Plasma ratio of valine, leucine and isoleucine
to phenylalanine and tyrosine in liver disease." Gut 19(11): 1068-1073.
Noguchi, Y., R. Sakai, et al. (2003). "Metabolomics and its potential for assessment of
adequacy and safety of amino acid intake." J Nutr 133(6 Suppl 1): 2097S-2100S.
Norton, J. A., C. M. Gorschboth, et al. (1985). "Fasting plasma amino acid levels in cancer
patients." Cancer 56(5): 1181-1186.
Oresic, M., S. Simell, et al. (2008). "Dysregulation of lipid and amino acid metabolism
precedes islet autoimmunity in children who later progress to type 1 diabetes." J Exp
Med 205(13): 2975-2984.
Oyadomari, S., A. Koizumi, et al. (2002). "Targeted disruption of the Chop gene delays
endoplasmic reticulum stress-mediated diabetes." J Clin Invest 109(4): 525-532.
Piraud, M., C. Vianey-Saban, et al. (2005). "Ion-pairing reversed-phase liquid
chromatography/electrospray ionization mass spectrometric analysis of 76 underivatized
amino acids of biological interest: a new tool for the diagnosis of inherited disorders
of amino acid metabolism." Rapid Commun Mass Spectrom 19(12): 1587-1602.
Piraud, M., C. Vianey-Saban, et al. (2003). "ESI-MS/MS analysis of underivatised amino
acids: a new tool for the diagnosis of inherited disorders of amino acid metabolism.
Fragmentation study of 79 molecules of biological interest in positive and negative
ionisation mode." Rapid Commun Mass Spectrom 17(12): 1297-1311.
Prior, R. L. and S. B. Smith (1983). "Role of insulin in regulating amino acid metabolism
in normal and alloxan-diabetic cattle." J Nutr 113(5): 1016-1031.
Robert, J. J., B. Beaufrere, et al. (1985). "Whole body de novo amino acid synthesis in
type I (insulin-dependent) diabetes studied with stable isotope-labeled leucine, alanine,
and glycine." Diabetes 34(1): 67-73.
Salek, R. M., M. L. Maguire, et al. (2007). "A metabolomic comparison of urinary changes
in type 2 diabetes in mouse, rat, and human." Physiol Genomics 29(2): 99-108.

55
Scharff, R. and I. G. Wool (1966). "Effect of diabetes on the concentration of amino acids
in plasma and heart muscle of rats." Biochem J 99(1): 173-178.
Schulze, A., M. Lindner, et al. (2003). "Expanded newborn screening for inborn errors
of metabolism by electrospray ionization-tandem mass spectrometry: results, outcome, and
implications." Pediatrics 111(6 Pt 1): 1399-1406.
She, P., C. Van Horn, et al. (2007). "Obesity-related elevations in plasma leucine are
associated with alterations in enzymes involved in branched-chain amino acid metabolism."
Am J Physiol Endocrinol Metab 293(6): E1552-1563.
Shimbo, K., T. Oonuki, et al. (2009). "Precolumn derivatization reagents for high-speed
analysis of amines and amino acids in biological fluid using liquid
chromatography/electrospray ionization tandem mass spectrometry." Rapid Commun Mass
Spectrom 23(10): 1483-1492.
Shulman, G. I., W. W. Lacy, et al. (1980). "Effect of glucose, independent of changes
in insulin and glucagon secretion, on alanine metabolism in the conscious dog." J Clin
Invest 65(2): 496-505.
Szabo, A., E. Kenesei, et al. (1991). "Changes in plasma and urinary amino acid levels
during diabetic ketoacidosis in children." Diabetes Res Clin Pract 12(2): 91-97.
Vannini, P., G. Marchesini, et al. (1982). "Branched-chain amino acids and alanine as
indices of the metabolic control in type 1 (insulin-dependent) and type 2
(non-insulin-dependent) diabetic patients." Diabetologia 22(3): 217-219.
Vente, J. P., M. F. von Meyenfeldt, et al. (1989). "Plasma-amino acid profiles in sepsis
and stress." Ann Surg 209(1): 57-62.
Wang, J., T. Takeuchi, et al. (1999). "A mutation in the insulin 2 gene induces diabetes
with severe pancreatic beta-cell dysfunction in the Mody mouse." J Clin Invest 103(1):
27-37.

56
Wilcken, B., V. Wiley, et al. (2003). "Screening newborns for inborn errors of metabolism
by tandem mass spectrometry." N Engl J Med 348(23): 2304-2312.
Yoshioka, M., T. Kayo, et al. (1997). "A novel locus, Mody4, distal to D7Mit189 on
chromosome 7 determines early-onset NIDDM in nonobese C57BL/6 (Akita) mutant mice."
Diabetes 46(5): 887-894.
IDF Diabetes Atlas. http://www.idf.org/
2003 動物実験データ集. http://www.jslc.co.jp/disease/akita.pdf