closely related symbiodinium spp. differ in relative
TRANSCRIPT

MARINE ECOLOGY PROGRESS SERIESMar Ecol Prog Ser
Vol. 284: 147–161, 2004 Published December 21
INTRODUCTION
The development and employment of moleculargenetic tools has initiated a new era in ecological, evo-lutionary and systematic discovery. This is especiallytrue for microscopic and morphologically cryptic orga-
nisms such as dinoflagellate endosymbionts (zooxan-thellae) that promote the survival and growth of stonycorals, soft corals, and related species that constructand/or thrive on tropical reefs (Rowan & Powers 1991,Rowan et al. 1997, LaJeunesse 2001, 2002, Baker 2003).Ribosomal gene sequence comparisons indicate that
© Inter-Research 2004 · www.int-res.com*Email: [email protected]
Closely related Symbiodinium spp. differ in relativedominance in coral reef host communities across
environmental, latitudinal and biogeographicgradients
T. C. LaJeunesse1, 2, 6,*, R. Bhagooli3, M. Hidaka3, L. deVantier4, T. Done4, G. W. Schmidt1, W. K. Fitt2, O. Hoegh-Guldberg5
1Department of Plant Biology, Plant Sciences Building, 2Institute of Ecology, University of Georgia, Athens, Georgia 30602, USA3Department of Chemistry, Biology and Marine Sciences, University of the Ryukyus, Nishihara, Okinawa 903-0213, Japan
4Australian Institute of Marine Science, Private Mail Box 3, Townsville, Queensland 4810, Australia5Centre for Marine Studies, University of Queensland, St. Lucia, Queensland 4072, Australia
6Present address: Department of Biology, Florida International University, University Park Campus, OE 245 11200SW 8th Street, Miami, Florida 33199, USA
ABSTRACT: The diversity and community structures of symbiotic dinoflagellates are described fromreef invertebrates in southern and central provinces of the Great Barrier Reef (GBR), Australia, andZamami Island, Okinawa, Japan. The symbiont assemblages from region to region were dominatedby Clade C Symbiodinium spp. and consisted of numerous host-specific and/or rare types (special-ists), and several types common to many hosts (generalists). Prevalence in the host community amongcertain host–generalist symbionts differed between inshore and offshore environments, across latitu-dinal (central versus southern GBR) gradients, and over wide geographic ranges (GBR versusOkinawa). One particular symbiont (C3h) from the GBR had a dramatic shift in dominance. Its pre-valence ranged from being extremely rare, or absent on high-latitude reefs to dominating thescleractinian diversity on a mid-latitude inshore reef. These changes occurred among coral faunawhose larvae must acquire symbionts from environmental sources (horizontal symbiont acquisition).Such differences did not occur among ‘vertical transmitters’ such as Porites spp., Montipora spp. andpocilloporids (corals that directly transmit symbionts to their offspring) or among those hosts display-ing ‘horizontal acquisition’, but that associate with specific symbionts. Most host-specialized typeswere found to be characteristic of a particular geographic region (i.e. Okinawa versus Central GBRversus Southern GBR). The mode of symbiont acquisition may play an important role in howsymbiont composition may shift in west Pacific host communities in response to climate change.There is no indication that recent episodes of mass bleaching have provoked changes in host–symbiont combinations from the central GBR.
KEY WORDS: Symbiodinium · Zooxanthellae biodiversity · Coral symbiosis · Phylogeography ·Community structure
Resale or republication not permitted without written consent of the publisher

Mar Ecol Prog Ser 284: 147–161, 2004
coral endosymbionts, in the genus Symbiodinium, arehighly diverse and evolutionarily old (Trench 1997,Rowan 1998). This revelation is not especially remark-able given the diverse environments, biological com-plexity and ages of the ecosystems where these sym-bioses are dominant. Nevertheless, our growingunderstanding of this diversity and its ecological sig-nificance has direct implications to addressing issuesregarding the impact of global climate change onnature conservation (Hoegh-Guldberg 1999).
Improved awareness of symbiont diversity, physio-logy and host-specificity contributes to hypotheses onhow corals respond to climate change (Buddemeier &Fautin 1993, Baker 2001). Differences in partner com-binations across latitudinal, longitudinal and environ-mental (e.g. irradiance) gradients have been reportedfor a number of host taxa (Baker & Rowan 1997, Rowanet al. 1997, LaJeunesse & Trench 2000, Rodriquez-Lanetty et al. 2001, Van Oppen et al. 2001, Burnett2002). Such patterns could be related to symbiontswith different sensitivities to thermal stress (Iglesias-Prieto et al. 1992, Iglesias-Prieto & Trench 1997,Jones et al. 1998, Warner et al. 1999), irradiance(Chang et al. 1983, Iglesias-Prieto et al. 2004), and/orhost-specificity (Colley & Trench 1983, LaJeunesse2001). These findings underlie the hypothesis thatpartner-switching creates new partnerships that arebetter adapted to changes in physical–environmentalconditions (Rowan & Powers 1991, Buddemeier &Fautin 1993, Baker 2001). Can the formation of newpartnerships (i.e. ‘switching’) take place rapidly overecological time scales or is this capacity limited byslower evolutionary processes that may not keep pacewith the current rate of environmental change (Hoegh-Guldberg et al. 2002)?
Further description of the variability between hostsand their symbionts over environmental, latitudinaland geographic ranges should help define the spatialand temporal limitations governing the extent to whichthese systems may respond and possibly adjust tofuture climate change. To this end, large-scale surveysof Symbiodinium spp. diversity and ecology have beeninitiated on stony corals and related host groups fromthe southern Great Barrier Reef (GBR), Hawaii, andCaribbean regions (Loh et al. 1998, Baker 1999, La-Jeunesse 2002, LaJeunesse et al. 2003, 2004).
The genus Symbiodinium is partitioned systemati-cally into a number of major phylogenetic divisions orclades. To date, most Pacific reef cnidarians are foundto harbor Symbiodinium spp. from Clade C, whereashost assemblages in the Caribbean associate com-monly with Symbiodinium spp. from Clades A and(especially) B (Baker & Rowan 1997, LaJeunesse et al.2003). The shared ecological dominance in theCaribbean of Clade B with Clade C Symbiodinium
spp. resulted possibly from environmental change inthis region during the Pliocene–Pleistocene transition(Jackson 1994, Collins et al. 1996, Budd 2000, Baker2003, LaJeunesse et al. 2003). Harsher (e.g. colder)physical–environmental conditions in the region dur-ing this time may have promoted the partial ecologicaldisplacement of Clade C by Clade B (Baker 2003,LaJeunesse et al. 2003).
Each Symbiodinium clade comprises a yet undeter-mined number of closely related ‘types’ or species thatcluster at the end of long diverging branches (Rowan1998, LaJeunesse 2002, Baker 2003). Molecular geneticidentification of ecologically different Symbiodiniumspp. within each major clade has been achieved bycomparing internal transcribed spacer region (ITS 1and 2) sequences of ribosomal RNA genes (Baillie et al.2000, LaJeunesse 2001, 2002, Van Oppen et al. 2001.These genetically distinct types possess unique envi-ronmental (depth zonation), ecological (host range)and geographic distributions. Some types have widegeographic distributions (Baillie et al. 2000, LaJeunesse2001) and are found in numerous host taxa (LaJeunesse2002). Most others display limited geographic rangesand/or associate specifically with a particular hostgenus or species. Because the host is a major axis ofresource (habitat) utilization, each type is defined asbeing a ‘generalist’ or ‘specialist’ depending on theirrelative capacity to associate with different host taxa(LaJeunesse et al. 2003).
The world’s largest contiguous reef system, the GreatBarrier Reef (GBR), off the east coast of Australia,offers an appropriate setting to explore host–symbiontdynamics because communities from different lati-tudes (a total range of nearly 15°), physical environ-ments (warm turbid inshore versus colder clear off-shore waters), and those with different bleachinghistories (including frequency and severity) can beexamined. Systematic surveys were first begun in themost southern region of the GBR, Heron Island of theCapricorn bunker group (LaJeunesse et al. 2003), afterinitial studies by Loh et al. (1998). The majority of hosttaxa from the southern GBR associate with one ofseveral generalist symbionts from the Clade C lineage.While these types were the most ecologically common,the greatest proportion of Symbiodinium spp. diversityconsisted of host-specific and/or rare types, special-ists, also from Clade C (LaJeunesse et al. 2003). Thelog normal distribution of symbiont abundance and/orprevalence (few highly prevalent generalists and manyrare specialists) is consistent with reefs surveyed fromthe Caribbean (LaJeunesse 2002).
Among other remaining questions, it is not known ifthe patterns described above hold for GBR reefs sys-tems closer to the equator, where many of the mid-shelf and inshore reefs suffered greater bleaching and
148

LaJeunesse et al.: West Pacific zooxanthellae communities
mortality than those in the southern GBR (Berkelmans& Oliver 1999, Skirving & Guinotte 2001). Moreover,there may be an inherent latitude-related pattern inhost–symbiont partnerships across coral communities,similar to that already documented for several host taxa(LaJeunesse & Trench 2000, Loh et al. 2001, Rodriquez-Lanetty et al. 2001). Finally, we sought to determineif the relatively low host-to-symbiont diversity in thesouthern GBR is consistent with lower-latitude GBRreefs. The coral diversity in the southern province ofthe GBR is substantially lower than in more northernareas, forming a high-latitude subset of the total GBRhost-species pool (Veron 1995, 2000).
We also conducted surveys at Zamami Island, partof the Kerama Islands, located 20 to 40 km west ofOkinawa, in the northern hemisphere, for purposes ofcomparing GBR Symbiodinium spp. diversity with thaton a reef from another region in the western Pacific.Unlike Okinawa to the immediate east, Zamami Islandwas not severely impacted by bleaching and representsa more ‘pristine’ reef analogous to Heron Island at thesame relative latitude in the opposite hemisphere.
MATERIALS AND METHODS
In May 2003, symbiotic invertebrates were collectedby SCUBA on reefs from the central Great BarrierReef. Collections were conducted in late March 2002 atZamami Island, of the Kerama Islands, approximately30 km west of the south end of Okinawa Island, Japan.
As with previous studies attempting to quantify thegeneral diversity of symbionts from a reef system (La-Jeunesse 2002, LaJeunesse et al. 2003, 2004), anemphasis was placed on sampling from a diverse rangeof hosts (over 75 genera, 154 species) consisting ofhard corals, soft corals, gorgonians, anemones, zoan-thids, corallimorphs, tridacnid clams and nudibranchs.Importantly, members of a host taxon found in a partic-ular reef environment and geographic region usuallypossess the same symbiont type (Baker 1999, La-Jeunesse 2002), with some exceptions (cf. Loh et al.2001) (i.e. 80 to 100% of individuals of Coral C sam-pled at Depth D on Reef R will have Symbiont S). Dif-ferences in host diversity between various reef habitatsand locations made sampling from a proportional num-ber of species difficult. For example, half of the hostgenera collected at Rib and Feather Reefs weresampled at the Curaçao Island fringing reef. The lowgeneric host diversity at this inshore location andlimited field time made collections at this reef unpro-portional to those of the mid-shelf reefs.
Collections from shallow (1 to 4 m) and deeper(>10 m) reef zones were made to obtain corals livingunder different irradiances. Host taxa distributed at
both depths were collected to identify possible ‘poly-morphisms’. Sampling colonies from deep and shal-low habitats increased the probability of identifyingcoral species that associate with more than 1 symbiont(Baker 1999). This work did not attempt to quantify thecomplete diversity of symbionts with which a particu-lar host associates, but rather represents a ‘snapshot’ ofthe symbionts across a wide array of hosts in a particu-lar community. A study of complete diversity wouldrequire exhaustive sampling of the host from everyenvironment in which it is found.
Before sampling, each host individual was photo-graphed (overall morphology and close-up) using aNikon Cool-Pix 5000 digital camera in an underwaterhousing (Subal, Netherlands) for later identification.The images obtained were compared with taxonomicreferences and identification guides (e.g. Veron 2000,Fabricius & Alderslade 2001), and most hosts wereidentified to the genus and species level.
Fragments representative of the host colony or indi-vidual were collected and processed to separate sym-bionts from host tissues, as previously described (La-Jeunesse et al. 2003). The resulting algal pellet waspreserved in 20% DMSO, 0.25 M EDTA in NaCl-saturated water (Seutin et al. 1991). Algal pellets fromZamami Island were preserved in CHAOS solution(4M guanidine thiocyanate, 0.5% sarkosyl, 2.5 mMTris-HCL (pH 8.0) and 0.1 M β-mercaptoethanol).
The Wizard DNA preparation protocol by Promega,modified by LaJeunesse et al. (2003), was used toextract nucleic acids. Approximately 30 mg of materialwas placed into 1.5 ml microcentrifuge tubes with250 µg 0.5 mm glass beads and 600 µl nuclei lysisbuffer (Promega) and bead-beaten for 2 min at 800 × gin a Biospec Mini-Beadbeater. The lysate was thenincubated with 0.1 mg ml–1 Proteinase K for 1 to 2 h at65°C, followed by incubation with 6 µg ml–1 RNase at37°C for 10 min. Protein precipitation buffer (250 µl)was then added and the extract incubated on ice for 10to 15 min. After centrifugation for 5 min at 24 000 × g,600 µl of supernatant was transferred to a second1.5 ml tube containing 700 µl isopropanol 100% and50 µl NaAc (3 M, pH 5.6). Following incubation on icefor 10 min, the precipitated DNA was centrifuged andthe pellet washed with 70% EtOH. The DNA was cen-trifuged again for 5 min, dried, and resuspended in70 µl H2O and 4 µl 10× Tris-EDTA.
While numerous molecular markers are now employedfor the study of Symbiodinium diversity (Baker 2003),the internal transcribed spacer region (ITS) methodappears to provide adequate resolution between eco-logically distinctive forms while being sufficientlyconserved to enable assessment of different types tobe compared among communities from distant geo-graphic areas. For each DNA extract, the ITS 2 region
149

Mar Ecol Prog Ser 284: 147–161, 2004
was amplified using primers ‘ITS 2 clamp’ and ‘ITSint-for 2’ (LaJeunesse & Trench 2000) with the touch-downthermal cycle given in LaJeunesse (2002). Productsfrom these PCR reactions were electrophoresed for9.5 h on denaturing gradient gels (45 to 80%) using aCBScientific system. Gels stained with Sybergreen(Molecular Probes) for 30 min were photographedusing a Kodak digital imaging system or with standardblack-and-white 677 polaroid film.
The PCR-DGGE fingerprint signatures from eachsample were compared with profiles from earlier datasets (LaJeunesse et al. 2003, 2004). The identificationof new symbiont types was verified by excising brightlystained bands from the denaturing gel. The DNA waseluted in 500 µl HOH for several days, re-amplifiedusing the same primer set without the guanine-cytosine-rich clamp extension in a standard PCR ther-mal cycle profile (annealing of 52°C for 40 cycles), andsequenced as previously described (LaJeunesse 2002).
To demonstrate the reproducibility of the PCR reac-tion by this method, 2 samples that showed a similarprofile involving 2 co-dominant ITS variants, but withdifferent relative band intensities, were repeatedlyamplified and compared. This verified whether cleardifferences in band intensity were the product of PCRartifact or reflected true differences in copy numberbetween variant sequences within a genome.
Maximum parsimony under the default settings ofPAUP* 4.0b10 (Swofford 1999) was employed for infer-ring a phylogeny. Sequences were edited and alignedmanually using Sequence Navigator Version 1.0 soft-ware (ABI, Division of Perkin Elmer). Clade C pos-sesses ITS 2 sequence types distinguished by a lownumber of base substitutions and/or insertion/dele-tions. With maximum parsimony (MP), informativesequence gaps as a 5th character state were included,delayed-transformation (DELTRAN) was chosen forcharacter state optimization, and no model of molecu-lar evolution was assumed. Sister lineages to Clade Crepresented by Fr1 (sensu Pochon et al. 2001) and F1in Clade F (Symbiodinium kawagutii; LaJeunesse2001) were used as outgroups (GenBank SequencesAJ291520 and AF333515). Phylogenetic analyses,neighbor-joining (NJ) and Bayesian inference of phy-logeny were also performed and the resulting treetopologies compared to MP. Baysian analyses wereimplemented using the software MrBayes Version3.0b4 (Huelsenbeck & Ronquiest 2001). We ran 500 000generations under the HKY85 models of sequence evo-lution, beginning with an unspecified tree topology,and no defined prior probabilities. The log probabilityreached stationarity between 50 000 and 75 000 gener-ations. This burn-in was not discarded and thereforethe posterior probabilities presented below are con-servative estimates. A bootstrap resampling was also
conducted for 500 replicates to assess relative branchsupport. Given that there are a small number of criticalbase substitutions (out of 321 aligned characters, 230were invariable and 32 were parsimony-informative),bootstrapping probably underestimates the actual sup-port for sub-clade radiations. A second bootstrappingwas performed with the resampling value doubled(642 characters). An unrooted phylogenetic analysis ofClade C sequences was also performed and topologywas compared with the rooted one.
Finally, symbiont community similarity from eachregion was assessed statistically using the Sørensencoefficient. It is preferred here to the Jaccard methodbecause it weights species, or types that are commonbetween regions over types found in only 1 area.
RESULTS
Symbiodinium spp. diversity
We identified 32 symbiont ‘types’ from biopsies of 74different host genera sampled at the 3 reefs surveyedfrom the central GBR (Table 1) and 20 types from 31host genera from Zamami island (Table 2). PCR-DGGEfingerprint profiles representing ‘new’ types from theCentral GBR and Okinawa are shown in Fig. 1a,brespectively. All belong to Clade C, the group mostprevalent among host taxa surveyed at all 3 regionallocations (Tables 1 & 2). New Clade C types character-ized were C1j (AY589732), C1k (AY589733), C31a(AY589746), C31b (AY589767), C40 (AY258485), C40a(AY589747), C40b (AY589748), C55 (AY589759), C56(AY589760), C57 (AY589761), C58 (AY589762), C59(AY589763), C60 (AY589764), C61 (AY589765), C62(AY589766), C64 (AY589768), C65/65a (AY589769/AY589770), C67 (AY686647), C68 (AY589772), C69/C69a (AY589773/AY686648), C70 (AY589774), andC71/C71a (AY589775/AY589776). A nexus alignmentof all Clade C ITS types from the west Pacific isavailable upon request from the corresponding author.
Due to the dominance of Clade C Symbiodiniumspp., the diversity and prevalence of symbionts fromother clades was low. Types A3 and A6 (AY686646)were identified from tridacnid clam tissues from thecentral GBR and Okinawa respectively. Clade B mem-bers were absent from our collections, except Type B1in the soft coral genus Nephthia from the southernGBR (LaJeunesse et al. 2003).
Only 3 ITS types in Clade D were identified (Fig. 2).Type D1a was found sporadically among faviids, mus-sids and oculinids from the central GBR, and agaricidsand fungiids from Zamami Island in typically shallowhabitats (1 to 4 m). Its fingerprint is known fromhosts from the western Indian Ocean, central Pacific
150

LaJeunesse et al.: West Pacific zooxanthellae communities
Ocean, and Caribbean Sea (LaJeun-esse 2002, LaJeunesse et al. 2005, A.Baker & T. LaJeunesse unpubl.). TypeD2 (AY686649) was found only amongsome Acropora spp., indicating that itmay be specialized to this particulargenus. Type D3 (AY686650) wasfound at only 1 location and in 1 hosttaxon (Clavularia sp. collected at 6 m).The rarity and probable host-specificnature of this type indicates thatClade D also consists of geographi-cally widespread host-generalists andmore localized, host-specific and/orrare forms.
PCR-DGGE analysis of Type C3h
The ITS 2 PCR-DGGE fingerprintprofiles of Type C3h from 19 host gen-era are illustrated in Fig. 3a. C3h wasnever identified in acroporids, how-ever. The genome of this particularSymbiodinium sp. contains 2 co-dominant ITS 2 sequences across theribosomal repeat array. Essentially it isintermediate between Types C3 andC21, a genome either caught in theprocess of concerted evolution fromthe ancestral sequence of C3 tothe more derived sequence of C21, orthe result of sexual recombination be-tween Types C3 and C21. The de-signation C3h was therefore given tothis fingerprint profile to distinguish itfrom Types C3 and C21. The upperC21/C3h band is consistently brighterthan the lower and suggests that therelative copy number between each
sequence is fixed in the genome of this organism. Therewas one exception; the C3 band from Goniastrea pecti-nata (Feather Reef, 15 m) was brighter than theC21/C3h band and the faint banding pattern foundhigher up in that lane differed from the other profiles.
To test whether this difference was a random artifactof the amplification process, 8 PCR replicate reactionswere conducted on the DNA extract from Goniastreapectinata that gave rise to this variant and on a repre-sentative that produced the normal profile. There wasno discernible fingerprint variation from reaction toreaction and the profiles from each example remainedconsistent with original analyses (Fig. 3b). The C3hprofile from G. pectinata is therefore different from theother C3h profiles. Because these similar fingerprint
151
Fig. 1. Symbiodinium spp. (a) Representative PCR-DGGE ITS 2 fingerprints (pro-files) of Lineage C types observed in hosts from the Central Great Barrier Reef.Identities, given as alphanumeric designations, and diagnostic band(s) to whicheach refers are compiled for those species newly reported: uppercase letters indi-cate lineage or clade, numbers represent ITS type, and lowercase letters denote arDNA paralog, when one is present within the genome and diagnostic of theentire fingerprint. (b) PCR-DGGE ITS 2 fingerprints of endosymbionts fromZamami Island, Okinawa Japan. Profiles from common symbionts C3, C1, and D1were pooled and run in the marker (MKR) lane. Examples of heteroduplexes areindicated; they are artifacts of the DGGE-PCR process present in fingerprints
of genomes with more than 1 dominant ITS 2 sequence
a
b
Fig. 2. Symbiodinium spp. PCR-DGGE ITS 2 fingerprints of Clade Didentified from the western Pacific.D1a was found sporadically amongfaviid, mussid and oculinid genera,usually related to shallow environ-ments but not completely dependenton depth. This fingerprint is found inhosts from the western Indian Ocean,central Pacific and Caribbean (La-Jeunesse 2002, A. Baker & T. La-Jeunesse unpubl.). Type D2 wasfound only in Acropora spp. from thecentral GBR and may represent a host-specialized symbiont. Type D3 wasfound at Curaçao Island in octocoralClavularia sp., but questions about itsprevalence and host-specificity remain
Cade D ‘types’
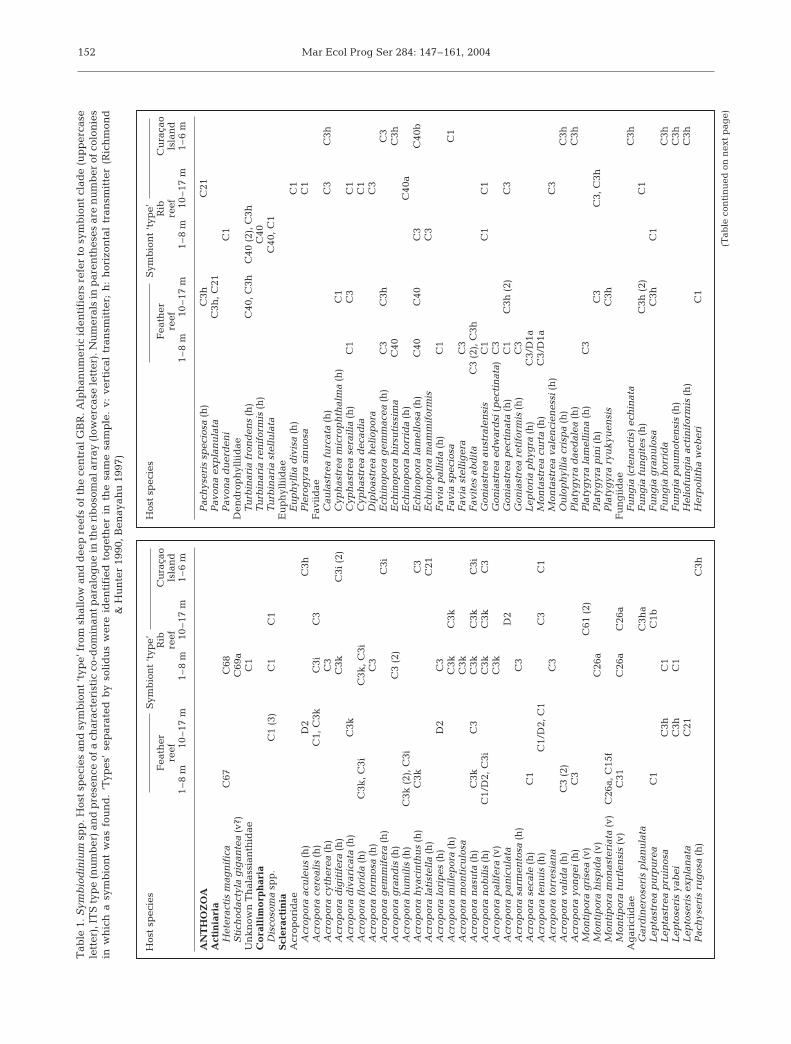
Mar Ecol Prog Ser 284: 147–161, 2004152
Tab
le 1
. Sym
bio
din
ium
sp
p. H
ost
spec
ies
and
sym
bio
nt
‘typ
e’ f
rom
sh
allo
w a
nd
dee
p r
eefs
of
the
cen
tral
GB
R. A
lph
anu
mer
ic id
enti
fier
s re
fer
to s
ymb
ion
t cl
ade
(up
per
case
lett
er),
IT
S t
ype
(nu
mb
er)
and
pre
sen
ce o
f a
char
acte
rist
ic c
o-d
omin
ant
par
alog
ue
in t
he
rib
osom
al a
rray
(lo
wer
case
lett
er).
Nu
mer
als
in p
aren
thes
es a
re n
um
ber
of
colo
nie
sin
wh
ich
a s
ymb
ion
t w
as f
oun
d.
‘Typ
es’
sep
arat
ed b
y so
lid
us
wer
e id
enti
fied
tog
eth
er i
n t
he
sam
e sa
mp
le.
v: v
erti
cal
tran
smit
ter;
h:
hor
izon
tal
tran
smit
ter
(Ric
hm
ond
&
Hu
nte
r 19
90, B
enay
ahu
199
7)
AN
TH
OZ
OA
Act
inia
ria
Het
erac
tis
mag
nif
ica
C67
C68
Sti
chod
acty
la g
igan
tea
(v?)
C69
aU
nk
now
n T
hal
assi
anth
idae
C1
Co
rall
imo
rph
aria
Dis
coso
ma
spp
.C
1 (3
)C
1C
1S
cler
acti
nia
Acr
opor
idae
Acr
opor
a ac
ule
us
(h)
D2
C3h
Acr
opor
a ce
real
is (
h)
C1,
C3k
C3i
C3
Acr
opor
a cy
ther
ea (
h)
C3
Acr
opor
a d
igit
ifer
a (h
)C
3kC
3i (
2)A
crop
ora
div
aric
ata
(h)
C3k
Acr
opor
a fl
orid
a (h
)C
3k, C
3iC
3k, C
3iA
crop
ora
form
osa
(h)
C3
Acr
opor
a g
emm
ifer
a (h
)C
3iA
crop
ora
gra
nd
is (
h)
C3
(2)
Acr
opor
a h
um
ilis
(h
)C
3k (
2), C
3iA
crop
ora
hya
cin
thu
s (h
)C
3kC
3A
crop
ora
lati
stel
la (
h)
C21
Acr
opor
a lo
rip
es (
h)
D2
C3
Acr
opor
a m
ille
por
a (h
)C
3kC
3kA
crop
ora
mon
ticu
losa
C3k
Acr
opor
a n
asu
ta (
h)
C3k
C3
C3k
C3k
C3i
Acr
opor
a n
obil
is (
h)
C1/
D2,
C3i
C3k
C3k
C3
Acr
opor
a p
alif
era
(v)
C3k
Acr
opor
a p
anic
ula
taD
2A
crop
ora
sarm
ento
sa (
h)
C3
Acr
opor
a se
cale
(h
)C
1A
crop
ora
ten
uis
(h
)C
1/D
2, C
1C
3C
1A
crop
ora
torr
esia
na
C3
Acr
opor
a va
lid
a (h
)C
3 (2
)A
crop
ora
yon
gei
(h
)C
3M
onti
por
a g
rise
a (v
)C
61 (
2)M
onti
por
a h
isp
ida
(v)
C26
aM
onti
por
a m
onas
teri
ata
(v)
C26
a, C
15f
Mon
tip
ora
turt
len
sis
(v)
C31
C26
aC
26a
Ag
aric
iid
aeG
ard
iner
oser
is p
lan
ula
taC
3ha
Lep
tast
rea
pu
rpu
rea
C1
C1b
Lep
tast
rea
pru
inos
aC
3hC
1L
epto
seri
s ya
bei
C3h
C1
Lep
tose
ris
exp
lan
ata
C21
Pac
hys
eris
ru
gos
a (h
)C
3h
Hos
t sp
ecie
sS
ymb
ion
t ‘t
ype’
Fea
ther
Rib
Cu
raça
ore
efre
efIs
lan
d1
–8
m10
–17
m1
–8
m10
–17
m1
–6
m
Pac
hys
eris
sp
ecio
sa (
h)
C3h
C21
Pav
ona
exp
lan
ula
taC
3h, C
21P
avon
a d
uer
den
iC
1D
end
rop
hyl
liid
aeT
urb
inar
ia f
ron
den
s (h
)C
40, C
3hC
40 (
2), C
3hT
urb
inar
ia r
enif
orm
is (
h)
C40
Tu
rbin
aria
ste
llu
lata
C40
, C1
Eu
ph
ylli
idae
Eu
ph
ylli
a d
ivis
a (h
)C
1P
lero
gyr
a si
nu
osa
C1
Fav
iid
aeC
aula
stre
a fu
rcat
a (h
)C
3C
3hC
yph
astr
ea m
icro
ph
thal
ma
(h)
C1
Cyp
has
trea
ser
aili
a (h
)C
1C
3C
1C
yph
astr
ea d
ecad
iaC
1D
iplo
astr
ea h
elio
por
aC
3E
chin
opor
a g
emm
acea
(h
)C
3C
3hC
3E
chin
opor
a h
irsu
tiss
ima
C40
C3h
Ech
inop
ora
hor
rid
a (h
)C
40a
Ech
inop
ora
lam
ello
sa (
h)
C40
C40
C3
C40
bE
chin
opor
a m
amm
ifor
mis
C3
Fav
ia p
alli
da
(h)
C1
Fav
ia s
pec
iosa
C1
Fav
ia s
tell
iger
aC
3F
avit
es a
bd
ita
C3
(2),
C3h
Gon
iast
rea
aust
rale
nsi
sC
1C
1C
1G
onia
stre
a ed
war
dsi
(p
ecti
nat
a)C
3G
onia
stre
a p
ecti
nat
a (h
)C
1C
3h (
2)C
3G
onia
stre
a re
tifo
rmis
(h
)C
3L
epto
ria
ph
ygra
(h
)C
3/D
1aM
onta
stre
a cu
rta
(h)
C3/
D1a
Mon
tast
rea
vale
nci
enes
si (
h)
C3
Ou
lop
hyl
lia
cris
pa
(h)
C3h
Pla
tyg
yra
dae
dal
ea (
h)
C3h
Pla
tyg
yra
lam
elli
na
(h)
C3
Pla
tyg
yra
pin
i (h
)C
3C
3, C
3hP
laty
gyr
a ry
uk
yuen
sis
C3h
Fu
ng
iid
aeF
un
gia
(cte
nac
tis)
ech
inat
aC
3hF
un
gia
fu
ng
ites
(h
)C
3h (
2)C
1F
un
gia
gra
nu
losa
C3h
C1
Fu
ng
ia h
orri
da
C3h
Fu
ng
ia p
aum
oten
sis
(h)
C3h
Hel
iofu
ng
ia a
ctin
ifor
mis
(h
)C
3hH
erp
olit
ha
web
eri
C1
Hos
t sp
ecie
sS
ymb
ion
t ‘t
ype’
Fea
ther
Rib
Cu
raça
ore
efre
efIs
lan
d1
–8
m10
–17
m1
–8
m10
–17
m1
–6
m
(Tab
le c
onti
nu
ed o
n n
ext
pag
e)

LaJeunesse et al.: West Pacific zooxanthellae communities 153
Lit
hop
hyl
lon
un
du
latu
mC
3hP
odab
acia
cru
stac
eaC
3hC
3hP
olyh
pyl
lia
talp
ina
(h)
C1
C3h
/C1
San
dal
olit
ha
rob
ust
a (h
)C
3hM
eru
lin
idae
Hyd
nop
hor
a ex
esa
C3h
(2)
, C3
C3h
Mer
uli
na
amp
liat
a (h
)C
1C
3C
3(2)
Mer
uli
na
scab
ricu
laC
1 (2
)M
uss
idae
Aca
nth
astr
ea e
chin
ata
(h)
D1a
Cyn
arin
a la
crym
alis
C3h
Lob
oph
ylli
a h
emp
rich
ii (
h)
C3h
C3
(2),
C3h
aC
3hS
coly
mia
au
stra
lis
C1
Sym
ph
ylli
a ag
aric
iaC
3S
ymp
hyl
lia
rad
ian
sC
40C
40S
ymp
hyl
lia
rect
aC
40O
culi
nid
aeG
alax
ea h
orre
sen
s (G
. acr
hel
ia)
D1a
Gal
axea
ast
reat
a (h
)C
1G
alax
ea f
asci
cula
ris
(h)
C1,
D1a
D1a
Pec
tin
iid
aeE
chin
oph
ylli
a as
per
a (h
)C
3ha
C3h
(2)
Ech
inop
hyl
lia
ech
inat
aC
3hE
chin
oph
ylli
a ec
hin
opor
oid
esC
1E
chin
oph
ylli
a or
ph
een
sis
(h)
C3h
Myc
ediu
m e
lep
han
totu
s (h
)C
3h (
2), C
3C
3h, C
1C
40O
xyp
ora
gla
bra
(h
)C
3hC
3ha
Oxy
por
a la
cera
(h
)C
3hC
3hP
ecti
nia
lac
tuca
(h
)C
3hP
ecti
nia
pae
onia
(h
)C
3hP
ecti
nia
sp
.C
3hP
ocil
lop
orid
aeP
ocil
lop
ora
dam
icor
nis
(v)
C1j
C1j
(2)
C1j
Poc
illo
por
a ey
dou
xi (
v)C
1cP
ocil
lop
ora
mea
nd
rin
a (v
)C
1cP
ocil
lop
ora
verr
uco
sa (
v)C
1cC
1cC
1cC
1cS
eria
top
ora
hys
trix
(v)
C3
C3
Sty
lop
hor
a p
isti
llat
a (v
)C
8C
8P
orit
idae
Alv
eop
ora
fen
estr
ata
C1
C31
b(2
)G
onio
por
a d
jib
outi
ensi
sC
1G
onio
por
a m
inor
C1
C1
Gon
iop
ora
ten
uid
ens
C1
Por
ites
an
nae
(v)
C15
(2)
C60
Por
ites
cyl
ind
rica
(v)
C15
C15
C15
C60
Por
ites
lic
hen
(v)
C15
C15
C15
C15
Por
ites
mas
sive
(v)
C15
C15
C15
C15
(2)
C15
Por
ites
nig
resc
ens
(v)
C15
Por
ites
ru
s (v
)C
15P
orit
es v
aug
han
i (v
)C
60C
15C
15
Hos
t sp
ecie
sS
ymb
ion
t ‘t
ype’
Fea
ther
Rib
Cu
raça
ore
efre
efIs
lan
d1
–8
m10
–17
m1
–8
m10
–17
m1
–6
m
Sid
eras
trei
dae
Cos
cin
arae
a co
lum
na
C1
C1
C1
(2)
Psa
mm
ocor
a co
nti
gu
aC
1P
sam
moc
ora
dig
itat
aC
1C
1P
sam
moc
ora
pro
fun
dac
ella
C1
Zo
oan
thid
eaP
alyt
hoa
sp
. (h
)C
1C
1, C
3Z
oan
thu
s sp
. (h
?)C
62 (
3)C
62C
62A
lcyo
nac
eaA
lcyo
nii
dae
Kly
xum
sp
.C
64L
obop
hyt
um
sp
p. (
h)
C1
C1
(2)
C1
Sar
cop
hyt
on s
pp
. (h
)C
1 (4
)C
65(2
)C
1S
inu
lari
a sp
. (h
)C
65 (
3)C
1cC
65 (
2)C
1 (2
)A
nth
oth
elid
aeIc
ilog
org
ia s
p.
C1
Bri
arei
dae
Bri
areu
m s
p.
C3
Cla
vula
rid
aeC
lavu
lari
a sp
. (h
?)D
3G
org
onid
aeH
ick
son
ella
exp
ansa
C65
Ru
mp
hel
la s
p.
C1
Ifal
uk
elli
dae
Plu
mig
org
ia s
p.
C1
Isid
idae
Isis
sp
.C
23T
ub
ipor
idae
Tu
bip
ora
mu
sica
C1b
Xen
iid
aeA
nth
elia
sp
. (v)
C64
Xen
ia s
p. (
v)C
64C
64C
64C
1kC
64H
eter
oxen
ia s
p. (
v)C
64C
64 (
2),C
15C
15e
Hel
iop
ora
cea
Hel
iop
ora
coer
ule
aC
1 (2
)
HY
DR
OZ
OA
Hyd
roid
aA
gla
oph
enia
sp
.C
15C
15M
ille
por
a ex
aesa
C1
C1
Mil
lep
ora
ten
ella
C15
e
MO
LL
US
CA
Tri
dac
nid
aeT
rid
acn
a d
eras
a (h
)C
1T
rid
acn
a g
igas
(h
)C
1, A
3T
rid
acn
a m
axim
a (h
)C
1 (3
)N
ud
ibra
nch
Pte
raeo
lid
ia i
anth
ina
(h)
C1
Hos
t sp
ecie
sS
ymb
ion
t ‘t
ype’
Fea
ther
Rib
Cu
raça
ore
efre
efIs
lan
d1
–8
m10
–17
m1
–8
m10
–17
m1
–6
m
Tab
le 1
(co
nti
nu
ed)

Mar Ecol Prog Ser 284: 147–161, 2004
profiles lack additional bands that could be sequencedto distinguish them on phylogenetic grounds, theywere both conservatively scored as the same type.Genetic structure occurs at all levels of a lineage downto the individual (Avise 2000). It is not surprising thatthe recent use of more variable markers indicates thatthese ITS lineages may be further subdivided (Santoset al. 2004).
Clade C phylogeny
The genetic relatedness of western Pacific Clade CSymbiodinium spp. is presented in Fig. 4. Internal
topology remained consistent under MP, NJ orBaysian inference of phylogeny methods, both rootedand unrooted. The exact point at which the outgroupbranch joins Clade C varied slightly, dependent onwhether Fr1 (Clade H; Pochon et al. 2005) and/orF1 (Clade F; LaJeunesse 2001) were used separatelyor together. The effect of long-branch attraction maysometimes supplant the outgroup connection withthe most ancestral sequence of this clade. However,a combination of ecological (host–generalist), biogeo-graphic (pandemic distribution), and phylogenetic(ancestral sequence to a radiation of numerous host-specific and/or rare types) evidence suggest TypesC3 or C1 are probably the most ancestral. The posi-
154
Table 2. Symbiodinium spp. Host species and symbiont ‘type collected’ at 1 to 10 m depth on reefs off Zamami Island, Okinawa, Japan. Further details as in Table 1
ANTHOZOA
ActiniariaUnkown anemone C70
ScleractiniaAcroporidae
Acropora abrolhosensis C1Acropora aspera C3 (2)Acropora cerealis C3 (2)Acropora copiosa C3Acropora digitifera (h) C3 (2)Acropora divaricata C3, C1 (2)Acropora donei C3 (3), C1Acropora exquisita C3Acropora florida (h) C3Acropora listeri C3Acropora microphthalma (h) C3Acropora nobilis (h) C1, C3Acropora palifera C1 (2)Acropora secale C3 (2)Acropora selago C3 (2)Acropora subglabra C3Acropora tenuis (h) C3 (2)Acropora valida (h)) C3, C3iAcropora verweyi C1Astreopora myriophthalma C1 (3)Montipora danae (v) C31Montipora efflorescens (v) C30Montipora mollis (v) C58 (2)Montipora venosa (v) C31a, C31Montipora verrucosa (v) C31Montipora spp. (v) C31, C31a, C31c, C1 (2)
AgariciidaePachyseris rugosa C1 (3)Pachyseris speciosa C27Pavona varians D1a
AstrocoeniidaePalauastrea ramosa C1 (3)
FaviidaeCaulastrea chalcidicum C1Cyphastrea japonica C1Cyphastrea sp. C21a (2)Echinopora lamellosa (h) C1, C3Echinopora pacificus C3Favia matthaii C3Favia stelligera C3Favites halicora C21aPlatygrya sp. (h) C55
FungiidaeFungia danai (h) C27Fungia scutaria (h) C1 (2)Fungia spp. (h) C27, C1Sandalolitha robusta (h) C1/D1a
MerulinidaeHydnophora exesa C21a (2), C27Hydnophora rigida C21a (2), C1
MussidaeLobophyllia robusta C21aSymphyllia sp. (h) C21aSymphyllia radians C21a (3)
OculinidaeGalaxea fascicularis (h) C21a
PectiniidaeEchonophyllia sp. C21aPectinia alcicornis C21aPectinia sp. C21a
PocilloporidaePocillopora damicornis (v) C1c (2)Pocillopora eydouxi (v) C1c (3)Seriatopora hystrix (v) C59Stylophora pistillata (v) C1 (4)
PoritidaeAlveopora sp. C27 (2)Porites cylindrica (v) C56a (2)Porites lichen (v) C56 (2)Porites lutea (v) C15 (3)Porites massive (v) C15 (3)Porites sp. (v) C56 (2)Porites rus (v) C15Porites silimaniana (v) C15
SiderastreidaeCoscinaraea sp. C1 (3)Coscinaraea exesa C1/C27
ZooanthideaZoanthus sp. (h) C1 (2)
AlcyonaceaAlcyoniidae
Sarcophyton sp. (h) C71a
HYDROZOAMillepora sp. C57 (6)
MOLLUSCATridacnidae
Tridacna sp. (h) A6
Host taxon Symbiont ‘type’ Host taxon Symbiont ‘type’

tion of Type C61 was unstable and certain branchconnections within the C21 sub-clade varied slightly.Several well-developed sub-cladal lineages contain-ing multiple types (viz. C15 and C21 and their off-shoots) correspond with host genera that transfertheir symbionts directly from generation to genera-tion (Fig. 4b). Some host-specialized and/or raresymbionts, especially those identified from species
of Porites, Montipora and Pocillopora, werefound at all locations, but many othersexhibited slight sequence differences thatdistinguished them regionally (Fig. 4b).
Similarity of symbiont communities anddifferences in relative dominance
Type C1 dominated most host communi-ties. Several other types were found at eachlocation, but the majority of diversity con-sisted of less common, geographicallyrestricted, rare and/or host-specific types.Reefs from distant geographic regions ex-hibited marked differences in their symbiontcommunities (Fig. 5). The Sørensen percent-age of dissimilarity estimates betweenZamami Island and Heron Island, ZamamiIsland and the central GBR, and the centralGBR and Heron Island were 68, 68, and 60%respectively. The 3 reefs surveyed from thecentral GBR region shared essentially thesame community composition of symbionttypes (dissimilarity estimates calculated be-tween Rib and Feather Reefs were 21%;Fig. 5). This valuation of community differ-ence is probably an overestimate because ofthe presence or absence of undersampledrare types and limited, uneven, host taxasampling at each reef. Because only half ofthe host genera collected a Rib and FeatherReefs were sampled at Curaçao Island, sta-tistical comparisons of their community com-positions were not made. Clearly, the sym-biont community at Curaçao is a subset ofthat found on offshore reefs.
Differences among the relative dominanceof some common symbiont types were ob-served from reef to reef (Fig. 5). C3h differedin its relative abundance between the mid-shelf reefs of Rib and Feather and the inshorereef at Curaçao Island (within 150 km of eachother). It was the most common type at therelatively turbid, inshore site at Curaçao,where it occurred in more than 50% of hostgenera surveyed. Often found in certain scle-
ractinian hosts sampled below 15 m on mid-shelf reefs(Fig. 3a), it occured in more genera on Feather Reef(approx. 30%) than on Rib Reef (approx. 8%). In con-trast, C3h was rare at the southern GBR location ofHeron Island, being found in 1 sample from Pavonamaldivensis.
As was the case for the outer GBR reefs, C1 was themost prevalent symbiont at the Zamami Island loca-
LaJeunesse et al.: West Pacific zooxanthellae communities 155
Fig. 3. Symbiodinium spp. (a) PCR-DGGE ITS2 fingerprints of Type C3h,collected from 19 different scleractinian genera from deep or low-lightreefs on the central GBR. Rare in the southern GBR, it achieved its highestprevalence in the host community surveyed from the turbid inshorelocation at Curaçao Island, central GBR. Relative intensities of each bandare similar from sample to sample and indicate that these sequences areintragenomic variants. It is postulated that ribosomal array contains 2 co-dominant ITS 2 sequences (C3 and C21) and represents an intermediatecondition of concerted evolution from C3 to C21. Alternatively, it couldrepresent a recombinant of these 2 distinctive species. (b) Repeatability ofthe PCR-DGGE method. The sample that gave rise to a distinctive ‘variant’whose C3 band was brighter (hence greater copy number) than theC3h/C21 band was amplified again along with a representative exhibitinga ‘normal’ profile. No discernible variation was detected from reaction toreaction, and fingerprint profiles remained consistent with original ana-lyses. Therefore, C3h profile from Goniastrea pectinata can be viewed asqualitatively different from the others but, because of limits in discerningthese 2 profiles through sequencing, they are both conservatively scored as
the same type
a
b

Mar Ecol Prog Ser 284: 147–161, 2004
tion. But here, Types C21a and C27 were also amongthe most common. The complex PCR-DGGE finger-print observed for Type C21a is thus far unique tothe Okinawa, NW Pacific, region. C27 was previouslyfound to occur in Pavona spp. from Hawaii (La-Jeunesse et al. 2004) and in 1 Pavona sp. specimenfrom Heron Island (LaJeunesse et al. 2003).
DISCUSSION
Variability in symbiont dominance is influenced bylife history of the host
External physical conditions may influence speci-ficity in symbioses that begin anew after each genera-
156
Fig. 4. Symbiodinium spp. Phylogenetic reconstruction of western Pacific Clade C based on ITS 2 and partial 5.8S sequences. Max-imum parsimony phylograms are both (a) rooted and (b) un-rooted; both have very similar topologies. (a) The point of connectionbetween Clade C and outgroup lineages differed depending on whether Fr1 or F1 were used separately or together. Dotted line in-dicates putative connection to Type C3 (bold vertical line) and is based on phylogeographic and ecological evidence for being an-cestral among Clade C types. Values indicated for each internal branch node are bootstrap estimates (first number), bootstrap withresampling doubled to compensate for high proportion of invariant characters (underlined), and Bayesian posterior probabilities(in parentheses); internal nodes that lack posterior probabilities are based on insertion/deletions not assessed by Bayesian methods.(b) Radiations of host-specific and/or rare types from a small number of widely distributed and/or host-generalist types are shownin this unrooted topology. Localization among types to specific geographic regions is indicated by symbols on branch termini:(h) southern GBR; (j) central GBR; (�) both GBR regions; d Zamami Island, Okinawa. Encircled types were found at all locations
a
b

LaJeunesse et al.: West Pacific zooxanthellae communities 157
Fig. 5. Symbiodinium spp. Comparison of diversity and community structures across latitudinal, environmental, and geographicgradients. Percentage of host genera surveyed is presented on y-axes and the different symbiont types in order of prevalenceamong host genera along the x-axes. Each community comprises a few types common to many host taxa and numerous host-specific and/or rare species that characterize each region surveyed. Type C1 is usually most common in each community. Exclud-ing Type C3, each region contains secondary generalists that are prevalent only in their respective regions (e.g. Types C21a andC27 in Zamami Island, C3h on the central GBR, and C21 from the southern GBR). Striped bars show relative prevalence ofType C3h at different locations over the GBR. Number of host genera surveyed at each site is given on upper-right of each graph,
with total numbers of symbiont types (displaying different ITS 2 PCR-DGGE fingerprints) in parentheses

Mar Ecol Prog Ser 284: 147–161, 2004
tion. A majority of western Pacific corals broadcast-spawn eggs and sperm that do not contain symbionts(Richmond & Hunter 1990). Therefore, coral popula-tions from this expansive region may be highly sus-ceptible to shifts in symbiont type from generation togeneration. We found that environment and latitudeaffected the relative dominance of certain Symbio-dinium spp. within hosts that acquire their symbiontsfrom environmental sources (Fig. 5, Tables 1 & 2).Because adult colonies often show long-term stabilitywith a particular symbiont type (Coffroth et al. 2001,Goulet & Coffroth 2003, Iglesias-Prieto et al. 2004,LaJeunesse et al. 2005), the present host–symbiontcommunity structure among broadcast-spawning coraltaxa probably reflects the environmental conditionsunder which these symbioses were initially estab-lished. Over several generations, change in environ-ment could cause significant shifts in the type of sym-biont dominating the host community; however, thisscenario assumes that the extent and rate of changedoes not exceed the physiological capability of eachpotential partner. Because there is the likelihood ofsignificant climatic disturbances in these communitiesover the coming decades (Hoegh-Guldberg 1999,Kleypas et al. 1999), continued monitoring will docu-ment the range of partner flexibility (Baker 2003) anddetermine if symbiont change can ultimately sustainthese reef corals.
Symbiont C1 was dominant on most reefs, but anumber of generalist taxa exhibited great regionalvariability in their relative dominance. Temperatureand light fluctuations associated with decreasing lati-tude may explain differences between the central GBRand southern GBR symbionts (Fig. 5). Within the cen-tral GBR region, higher temperatures and perhapsgreater turbidity at the inshore site of Curaçao Islandmay be key selective factors that explain the offshore-to-inshore gradient of C3h dominance. Also, little isknown about the environmental abundances of Sym-biodinium spp. If C3h is particularly abundant in theplanktonic or benthic assemblages nearer to shore, thismay in part explain differences in its host communitydominance (Baker & Rowan 1997).
Specialized and/or rare symbionts that displayedlimited ranges in geographic and host distributionaccounted for major differences in symbiont composi-tion from region to region. Many of these symbiontsassociate with hosts that pass on symbionts directly totheir offspring. For example, montiporid corals broad-cast eggs containing symbionts (Richmond & Hunter1990). They predictably associate with one of a numberof closely related types that are a part of the montiporidsub-clade radiating from C21 (Fig. 4b). The majority ofmembers from this sub-clade associate exclusivelywith Montipora spp. They have particular geographic
distributions, and are common in 1 region but rare orabsent in other places. Type C26a occurred rarely inMontipora spp. from the southern GBR, but was com-mon among these corals from the central GBR. It wasabsent from Zamami Island, yet was identified fromM. capitata in Hawaii living at depths below 20 m (La-Jeunesse et al. 2004) and, therefore, appears to have alarge geographic range. Another type, C31, was com-mon among montiporids from Zamami Island but rareon the central GBR. It associates with the commonbrown color morph of M. capitata (at depths above15 m) from Hawaii (LaJeunesse et al. 2004). Thesewidely distributed host-specialists contrast with othermontiporid symbionts surveyed from the NW Pacific,SW Pacific, and Hawaii that appear to have more lim-ited ranges. These distributions, involving widely dis-persed specialized types versus regionally endemictypes, are also observed for poritid and pocilloporidscorals, and among non-scleractinian groups such asthe alcyonarians.
Implications of mass coral bleaching
Many of the central GBR reefs experienced masscoral bleaching in 1998, 2002 and 2004, with mostsevere effects occurring on inshore reefs (Berkelmans& Oliver 1999, Marshall & Baird 2000). Such episodesof stress and reduction in symbiont population densitymay facilitate a shift in the symbiont type that becomesdominant upon recovery, either through uptake fromthe environment (Buddemeier & Fautin 1993, Baker2001) or proliferation of surviving cells remainingwithin the polyps (Baker 2001). For example, symbiontshifts in the population of ecologically dominant Pocillo-pora verrucosa were reported in the eastern Pacificfollowing the 1997 El Niño-related bleaching event(Glynn et al. 2001). It cannot be determined if the in-crease of Clade D (specifically Type D1) in proportionto Clade C (possibly involving different types withinthis clade) among individual coral colonies was due tonatural selection of thermally resistant combinations orwhether symbiont population shifts occurred via pro-liferation of a minor population of Clade D withincolonies as they recovered (Baker 2001).
Symbiodinium spp. from Clade D are often found toassociate with hosts from thermally variable and turbidenvironments and colonies or individuals recoveringfrom bleaching episodes (Baker 2001, Toller et al.2001, Van Oppen et al. 2001, Chen et al. 2003, Iglasias-Prieto et al. 2004). Various Clade D members areconsequently viewed as thermally tolerant, stress-re-sistant, opportunistic Symbiodinium spp. In thepresent study, Curaçao Island was also surveyed to de-termine if the inshore environment favored types from
158

LaJeunesse et al.: West Pacific zooxanthellae communities
Clade D over those from Clade C. Contrary to our ex-pectation that Clade D would dominate the host com-munity on the warmer inshore reefs (cf. Van Oppen etal. 2001), its prevalence was not different from that onreefs further offshore. Questions remain as to whatextent Clade D Symbiodinium spp. may allow coralpopulations to compensate under continued sea--surface warming (Baker 2001, Little et al. 2004). Com-parative physiological analyses will need to be em-ployed to learn what attributes or limitations accountfor the changes in the host community dominance ofthese Symbiodinium spp. These geographic and envi-ronmentally relatable shifts are ultimately of interest indescribing evolutionary processes between host andsymbiont lineages through time (Thompson 1994).
Heron Island and Zamami Island, both high-latitudereefs, have had limited exposures and experienceswith bleaching. While their symbiont community struc-tures are similar, the difference in composition ofClade C Symbiodinium spp. is clearly a product of theirgeographic separation. Thus for the present, the sever-ity or frequency of mass coral bleaching in the centralGBR does not seem to have resulted in a shift in sym-biont population structures that cannot be otherwiseexplained by geographic, latitudinal and normal envi-ronmental factors. These differences in Symbiodiniumspp. distribution probably took many generations tobecome established.
Symbiont diversity relative to host diversity
Many Pacific hosts associate with closely relatedSymbiodinium spp. in Clade C (Baker & Rowan 1997,Loh et al. 1998, Baker 1999, LaJeunesse et al. 2003,2004). Initially, host assemblages from southern GBRreefs were reported to have lower relative symbiont di-versity of ITS types than reefs from the Caribbean (La-Jeunesse et al. 2003). Subsequent work in Hawaii hasalso observed this inverse relationship between hostand symbiont diversity (LaJeunesse et al. 2004). Theenumeration of ITS types found on the central GBR waslow in relative proportion to the number of host generasurveyed. This trend offers further support of an inverserelationship between host and symbiont diversity.Genetic divergence and symbiont speciation throughhost-specialization is common for hosts in which sym-bionts are directly transferred from generation togeneration (Futuyma & Moreno 1988, Douglas 1998) aprocess that is exemplified by poritid, pocilloporid andmontiporid corals and their symbionts. The greatercomposition of hosts in the Caribbean (63% brood outof the 19 coral species investigated) and Hawaii (29%out of the 12 investigated) whose symbionts are verti-cally transferred in comparison to the west Pacific (GBR
6% out of 144, Okinawa 4% out of the 26 coral speciesinvestigated; Richmond & Hunter 1990), explains someof these inverse relations, but not all.
Symbiont-specificity and the presence of host-specificsymbionts were found for hosts that rely upon horizontalsymbiont transmission. Among others, species of Acrop-ora (with Types C3i, C3k and D2) (some Acropora [Iso-pora] brood their larvae) and Zoanthus (with Type C62)(Ryland 1997), possessed Symbiodinium spp. not identi-fied in other host taxa (cf. Van Oppen 2004). Regionswith a high diversity of hosts involved in acquiring sym-bionts from the environment may favor the maintenanceof highly prevalent generalist symbionts (Law 1985).Host rarity would present problems for passivelydispersed symbionts and newly settled aposymbioticlarvae in finding each other within a diverse hostcommunity over effectively vast spatial scales.
Emergent patterns of Symbiodinium spp. biogeography
Phylogeography, relating patterns of geographic dis-tribution with genetic relatedness, is a powerful tool inassessing historical and evolutionary processes (Avise2000). Our current understanding of Symbiodiniumdiversity and geographic distribution remains limited,but as more ITS data are gathered, patterns of dispersaland geographic isolation/connectivity are beginning toemerge. Widely distributed symbiont types tend to behost-generalists and are, phylogenetically, ancestral tosymbionts that are more specialized. Based on ecologi-cal, biogeographic, and phylogenetic grounds, we inter-pret Types C1 and C3 as the ancestral stock from whichnumerous host-specific, regionally endemic and/or raretypes have radiated. Divergent, host-specific and/or rareforms tend to have narrow geographic ranges and areprobably endemic. These patterns of geographic parti-tioning are reinforced by genetic surveys at the ‘popula-tion level’ that have identified clear geographic structurewithin specific ITS lineages (Santos et al. 2003, 2004).
Certain symbionts show exceptions to these basic pat-terns of geographic partitioning. Their host specificityand biogeography leads to hypotheses concerning theevolutionary processes that occur between host andsymbiont lineages. Type C27 is widely distributedthroughout the Pacific but has different host relations indifferent regions. It is rare in the southern GBR (LaJe-unesse et al. 2003), highly specific for the corals in thegenus Pavona from Hawaii (LaJeunesse et al. 2004), yetis a generalist among hosts from Zamami Island reefs.This example demonstrates the capacity for a symbiontto be specialized for 1 particular host in 1 region, butto display more generalized associations or greaterprevalence at other locations. The extent to whichhost–symbiont specificity is expressed in geographically
159

Mar Ecol Prog Ser 284: 147–161, 2004
separate populations is predicted by the geographicmosaic theory of coevolution (Thompson 1994).
Differences in symbiont communities between cen-tral and southern regions indicate that the GBR is bio-geographically partitioned. The similar symbiont com-positions found at Rib, Feather and Curaçao reefs areprobably possessed by other reefs from the centralGBR region. Except for the more common types (e.g.C1, C3, C21, C15), few others were identified in thesouthern GBR survey (LaJeunesse et al. 2003). On thenorthern GBR, Types C1, C40, C3h and C8 wereidentified in corals from Lizard Island (Baker 1999, A.Baker & T. LaJeunesse unpubl.). While these resultsindicate some similarity in symbiont assemblages be-tween northern and central regions, other Symbio-dinium spp. not identified from central or southernregions were also among those identified from LizardIsland (A. Baker & T. LaJeunesse unpubl.). Basedon the presence and/or absence of characteristic host-specific symbionts, the GBR may be divided into anumber of biogeographic provinces. Ayre & Hughes(2000) reported a lack of geographic partitioning, ascoral allozyme variation revealed moderate to highlevels of gene flow along the entire GBR. Trackingthe presence/absence of host-specific symbiont types,not influenced by environment across spatial scalesencompassed by the GBR is potentially a rapid methodfor determining genetic connectivity between andwithin major reef systems, information important toconservation-related decisions.
Acknowledgements. The authors would like to thank thecrew of the ‘Lady Basten’ for their hard work and support.S. Kininmonth produced the maps in Fig. 5 and waitedpatiently on the surface. A. Irikawa helped with coral identifi-cation from Okinawa. M. Wakeford assisted with diving andhelped with host identification on the GBR. D. Fautin advisedon anemone identifications and life-history traits. M. vanOppen and R. Berkelmans hosted our stay in Townsville. R.Berkelmans got us through the ‘Australian’ paper work. Thisresearch was made possible from funding provided by theAustralian Research Council (O.H.-G.: ARCL245G), IOC-UNESCO-World Bank Targeted Working Group on CoralBleaching and Related Ecological Factors, Australian Instituteof Marine Sciences, and the NSF (grant OCE-0137007 toW.K.F. and G.W.S.)
LITERATURE CITED
Avise JC (2000) Phylogeography: the history and formation ofspecies. Harvard University Press, Cambridge, MA
Ayre DJ, Hughes TP (2000) Genotypic diversity and gene flowin brooding and spawning corals along the Great BarrierReef, Australia. Evolution 54:1590–1605
Baillie BK, Belda-Baillie CA, Maruyama T (2000) Con-specificity and Indo-Pacific distribution of Symbiodiniumgenotypes (Dinophyceae) from giant clams. J Phycol 36:1153–1161
Baker AC (1999) The symbiosis ecology of reef-buildingcorals. PhD thesis, University of Miami, Miami
Baker AC (2001) Corals bleach to survive change. Nature 411:765–766
Baker AC (2003) Flexibility and specificity in coral–algalsymbiosis: diversity, ecology and biogeography of Sym-biodinium. Annu Rev Ecol Evol Syst 34:661–689
Baker AC, Rowan R (1997) Diversity of symbiotic dinoflagel-lates (zooxanthellae) in scleractinian corals of the Carib-bean and eastern Pacific. Proc 8th Int Coral Reef Symp2:1301–1306
Benayahu Y (1997) Developmental episodes in reef softcorals: ecological and cellular determinants. Proc 8th IntCoral Reef Sym 2:1213–1218
Berkelmans R, Oliver JK (1999) Large-scale bleaching ofcorals on the Great Barrier Reef. Coral Reefs 18:55–60
Budd A (2000) Diversity and extinction in the Cenozoichistory of Caribbean reefs. Coral Reefs 19:25–35
Buddemeier RW, Fautin DG (1993) Coral bleaching as anadaptive mechanism. BioScience 43:320–326
Burnett WJ (2002) Longitudinal variation in algal symbionts(zooxanthellae) from the Indian Ocean zoanthid Palythoacaesia. Mar Ecol Prog Ser 234:105–109
Chang SS, Prézelin BB, Trench RK (1983) Mechanisms ofphotoadaptation in three strains of the symbiotic dino-flagellate Symbiodinium microadriaticum. Mar Biol 76:219–29
Chen CA, Lam KK, Nakano Y, Tsai W (2003) A stable associ-ation of the stress-tolerant zooxanthellae, Symbiodiniumclade D, with the low-temperature tolerant coral, Oulas-trea crispata (Scleractinia: Faviidae) in subtropical non-reefal coral communites. Zool Stud 42:540–550
Coffroth MA, Santos SR, Goulet TL (2001) Early ontogeneticexpression of specificity in a cnidarian–algal symbiosis.Mar Ecol Prog Ser 222:85–96
Colley NJ, Trench RK (1983) Selectivity in phagocytosis andpersistence of symbiotic algae by the scyphistoma stage ofthe jellyfish Cassiopeia xamachana. Proc R Soc Lond B219:61–82
Collins LS, Budd A, Coates AG (1996) Earliest evolution asso-ciated with closure of the Tropical American Seaway. ProcNatl Acad Sci USA 93:6069–6072
Douglas AE (1998) Host benefit and the evolution of speciali-zation in symbiosis. Heredity 81:599–603
Fabricius K, Alderslade P (2001) Soft corals and sea fans: acomprehensive guide to the tropical shallow water generaof the central-west Pacific, the Indian Ocean and the RedSea. Australian Institute of Marine Science, Townsvilleand New Litho, Surrey Hills, Melbourne
Futuyma DJ, Moreno G (1988) The evolution of ecologicalspecialization. Annu Rev Ecol Syst 19:207–233
Glynn PW, Maté JL, Baker AC, Calderon MO (2001) Coralbleaching and mortality in Panama and Ecuador duringthe 1997–1998 El Niño–southern oscillation event: spatial/temporal patterns and comparisons with the 1982–1983event. Bull Mar Sci 69:79–109
Goulet TL, Coffroth MA (2003) Stability of an octocoral–algalsymbiosis over time and space. Mar Ecol Prog Ser 250:117–124
Hoegh-Guldberg O (1999) Climate change, coral bleachingand the future of the world’s coral reefs. Mar Freshw Res50:839–866
Hoegh-Guldberg O, Jones R, Ward S, Loh W (2002) Ecology—is coral bleaching really adaptive? Nature 415:601–602
Huelsenbeck JP, Ronquist F (2001) MrBays: Bayesian infer-ence of phylogenetic trees. Bioinformatics 17:754–755
Iglesias-Prieto R, Trench RK (1997) Photoadaptation, photo-
160

LaJeunesse et al.: West Pacific zooxanthellae communities
acclimation and niche diversification in invertebrate-dinoflagellate symbioses. Proc 8th Int Coral Reef Symp2:1319–1324
Iglesias-Prieto R, Matta JL, Robins WA, Trench RK (1992)Photosynthetic response to elevated temperature in thesymbiotic dinoflagellate Symbiodinium microadriaticumin culture. Proc Nat Acad Sci USA 89:10302–10305
Iglesias-Prieto R, Beltrán VH, LaJeunesse TC, Reyes-BonillaH, Thomé PE (2004) Different algal symbionts explain thevertical distribution of dominant reef corals in the easternPacific. Proc R Soc Lond B 271:1757–1763
Jackson JBC (1994) Constancy and change of life in the sea.Phil Trans R Soc Lond B 344:55–60
Jones R, Hoegh-Guldberg O, Larkum AWD, Schreiber U(1998) Temperature induced bleaching of corals beginswith impairment of dark metabolism in zooxanthellae.Plant Cell Environ 21:1219–1230
Kleypas JA, Buddemeier RW, Archer D, Gattuso JP, LangdonC, Opdyke BN (1999) Geochemical consequences ofincreased atmospheric carbon dioxide on coral reefs.Science 284:118–120
LaJeunesse TC (2001) Investigating the biodiversity, ecology,and phylogeny of endosymbiotic dinoflagellates in thegenus Symbiodinium using the internal transcribed spacerregion: in search of a ‘species’ level marker. J Phycol 37:866–880
LaJeunesse TC (2002) Diversity and community structure ofsymbiotic dinoflagellates from Caribbean coral reefs. MarBiol 141:387–400
LaJeunesse TC, Trench RK (2000) The biogeography of twospecies of Symbiodinium (Freudenthal) inhabiting theintertidal anemone, Anthopleura elegantissima (Brandt).Biol Bull (Woods Hole) 199:126–134
LaJeunesse TC, Loh WKW, van Woesik R, Hoegh-GuldbergO, Schmidt GW, Fitt WK (2003) Low symbiont diversity insouthern Great Barrier Reef corals relative to those of theCaribbean. Limnol Oceanogr 48:2046–2054
LaJeunesse TC, Thornhill DJ, Cox EF, Stanton FG, Fitt WK,Schmidt GW (2004) High diversity and host-specificityobserved among symbiotic dinoflagellates in reef coralcommunities from Hawaii. Coral Reefs 23:596–603
LaJeunesse TC, Lee S, Bush S, Bruno JF (2005) Persistence ofnon-Caribbean algal symbionts in Indo-Pacific mushroomcorals released to Jamaica 35 years ago. Coral Reefs (inpress)
Law R (1985) Evolution in a mutualistic environment. In:Boucher DH (ed) The biology of mutualism: ecology andevolution. Oxford University Press, New York, p 145–170
Little AF, Van Oppen JH, Willis BL (2004) Flexibility in algalendosymbioses shapes growth in reef corals. Science 304:1492–1494
Loh WK, Carter D, Hoegh-Guldberg O (1998) Diversity ofzooxanthellae from scleractinian corals of One Tree Island(The Great Barrier Reef). In: Greenwood JG, Hall NJ (eds)Proceedings of the Australian Coral Reef Society 75thAnniversary Conference, Heron Island. School of MarineSciences, University of Queensland, Brisbane, p 141–150
Loh WK, Loi T, Carter D, Hoegh-Guldberg O (2001) Geneticvariability of the symbiotic dinoflagellates from the wideranging coral species Seriatopora hystrix and Acroporalongicyathus in the Indo-West Pacific. Mar Ecol Prog Ser222:97–107
Marshall PA, Baird AH (2000) Bleaching of corals on theGreat Barrier Reef: differential susceptibilities amongtaxa. Coral Reefs 19:155–163
Pochon X, Pawlowski J, Zaninetti L, Rowan R (2001) Highgenetic diversity and relative specificity among Symbio-
dinium-like endosymbiotic dinoflagellates in soritid fora-miniferans. Mar Biol 139:1069–1078
Pochon X, LaJeunesse TC, Pawlowski J (2004) Biogeographicpartitioning and host specialization among foraminiferan di-noflagellate symbionts (Symbiodinium, Dinophyta). MarBiol 146:17–27
Richmond RH, Hunter CL (1990) Reproduction and recruitmentof corals: comparisons among the Caribbean, the tropicalPacific and the Red Sea. Mar Ecol Prog Ser 60:185–203
Rodriquez-Lanetty M, Loh W, Carter D, Hoegh-Guldberg O(2001) Latitudinal variability in symbiont specificity withinthe widespread scleractinian coral Plesiastrea versipora.Mar Biol 138:1175–1181
Rowan R (1998) Diversity and ecology of zooxanthellae on coralreefs. J Phycol 34:407–417
Rowan R, Powers DA (1991) A molecular genetic classificationof zooxanthellae and the evolution of animal–algal symbio-sis. Science 251:1348–1351
Rowan R, Knowlton N, Baker A, Jara J (1997) Landscape eco-logy of algal symbionts creates variation in episodes of coralbleaching. Nature 388:265–269
Ryland JS (1997) Reproduction in zoanthidea (Anthozoa: Hexa-corallia). Invertebr Reprod Dev 31:177–188
Santos SR, Gutierrez-Rodriquez C, Lasker HR, Coffroth MA(2003) Symbiodinium sp. associations in the gorgonianPseudopterogorgia elisabethae in the Bahamas: high levelsof genetic variability and population structure in symbioticdinoflagellates. Mar Biol 143:111–120
Santos SR, Shearer TL, Hannes AR, Coffroth MA (2004) Fine-scale diversity and specificity in the most prevalent lineage ofsymbiotic dinoflagellates (Symbiodinium, Dinophyceae) ofthe Caribbean. Mol Ecol 13:459–469
Seutin G, White BN, Boag PT (1991) Preservation of avianblood and tissue samples for DNA analyses. Can J Zool69:82–92
Skirving W, Guinotte J (2001) The sea surface temperature storyon the Great Barrier Reef during the coral bleaching event of1998. In: Wolanski E (ed) Oceanographic processes of coralreefs. Physical and biological links in the Great Barrier Reef.CRC Press, Boca Raton, p 301–313
Swofford DL (1999) PAUP*, phylogenetic analysis usingparsimony (*and other methods), Version 4.0b10. Sinauer,Sunderland, MA
Thompson JN (1994) The coevolutionary process. UniversityChicago Press, Chicago, IL
Toller WW, Rowan R, Knowlton N (2001) Zooxanthellae of theMontastraea annularis species complex: patterns of distrib-ution of four taxa of Symbiodinium on different reefs andacross depths. Biol Bull (Woods Hole) 201:348–359
Trench RK (1997) Diversity of symbiotic dinoflagellates and theevolution of microalgal-invertebrate symbioses. Proc 8th IntCoral Reef Symp 2:1275–1286
Van Oppen MJH (2004) Mode of zooxanthella transmissiondoes not affect zooxanthella diversity in acroporid corals.Mar Biol 144:1–7
Van Oppen MJH, Palstra FP, Piquet AMT, Miller DJ (2001) Pat-terns of coral–dinoflagellate associations in Acropora: signif-icance of local availability and physiology of Symbiodiniumstrains and host-symbiont selectivity. Proc R Soc Lond B268:1759–1767
Veron JEN (1995) Corals in space and time. University of NewSouth Wales Press, Sydney
Veron JEN (2000) Corals of the world, Vol 1–3. Australian Insti-tute of Marine Science, Townsville
Warner EM, Fitt WK, Schmidt GW (1999) Damage to photosys-tem II in symbiotic dinoflagellates: a determinant of coralbleaching. Proc Natl Acad Sci USA 96:8007–8012
161
Editorial responsibility: Otto Kinne (Editor), Oldendorf/Luhe, Germany
Submitted: March 24, 2004; Accepted: July 27, 2004Proofs received from author(s): December 16, 2004