cloning and characterization of a variant surface glycoprotein
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, Aug. 1986, p. 2950-2956 Vol. 6, No. 80270-7306/86/082950-07$02.00/0Copyright © 1986, American Society for Microbiology
Cloning and Characterization of a Variant Surface GlycoproteinExpression Site from Trypanosoma equiperdumANNA RAIBAUD,l GREG BUCK,'t THEO BALTZ,2 AND HARVEY EISEN1*t
Immunoparasitologie Unit, Institut Pasteur, 75015 Paris,' and University of Bordeaux II, Bordeaux,2 France
Received 21 February 1986/Accepted 1 May 1986
Variant surface glycoprotein (VSG) genes of African trypanosomes are expressed when they are inserted intoone of several telomere-linked expression sites. We cloned and characterized an 1 1-kilobase (kb) DNA fragmentlocated upstream of an expressed VSG gene. A DNA sequence of 1.8 kb that is located immediately upstreamof the inserted VSG gene contains sequences homologous to the 76-base-pair repeats described as beingupstream of VSG genes in Trypanosoma brucei (D. A. Campbell, M. P. Van Bree, and J. C. Boothroyd,Nucleic Acids Res. 12:2759-2774). There are no such sequences elsewhere in the 11-kb cloned region. Southernblot analysis using probes from the cloned region revealed multiple unlinked copies of the same or very similarregions. At least three of these are located near telomeres, and two have been shown to be used for theexpression of known Trypanosoma equiperdum VSG genes. Like VSG genes, the upstream sequences themselvescan be duplicated and deleted. The choice of expression site to be used by a duplicated VSG gene is nonrandom;the site used for expression of the parental VSG gene is strongly favored for use in the daughter variant.Furthermore, even when the parental expression site is not used, the VSG gene occupying it is replaced. Thus,an active expression site is a preferential target for gene conversion in the next variation event.
African trypanosomes alternately present different VSGs(see Materials and Methods for definitions of abbreviationsand a trypanosome glossary) in the mammalian host. Thisprocess of antigenic variation is presumed to aid the para-sites in evading the immune system of the host by causing itto repeatedly respond to new surface antigens (19). Althoughthe actual number of VSG genes in the trypanosome reper-toire is not known, over 100 immunologically distinct vari-ants have been characterized in Trypanosoma equiperdum(6). Activation of VSG genes is a complex process involvingat least two steps. First, silent BC genes are duplicated andtransposed to one of several telomere-linked ES's to formELCs (2, 9, 10, 15). Although this gene conversion event isnecessary for VSG gene expression, it is not sufficient sinceduplicated genes in ES's are further regulated in situ. ActiveVSG genes in ES's can be inactivated without furthergenomic rearrangements and can remain as R-ELCs (3).These inactivated R-ELCs can then be reactivated by anonduplicative event (3, 21, 23).The structure of a typical ES is shown in Fig. lc. The
DNA situated 3' to the inserted ELC contains no restrictionsites and constitutes a chromosome telomere (8, 16, 22). Theregion located 5' to the ELC is also very poor in restrictionsites. This 5' barren region has been shown to be composedof tandem repeats of a 70- to 76-base-pair (bp) sequencewhich contains multiple repeats of the simple sequence TAA(5). The length of these 5' barren regions is not fixed butvaries when the VSG gene inserted into the ES is changed.Similar or identical tandem repeats are also found 5' of someBCs (5). The DNA situated upstream of the 5' barren regionsin the ES's appears to be normal with respect to restriction
* Corresponding author.t Present address: Department of Microbiology and Immunology,
Medical College of Virginia, Richmond, VA 23298.t Present address: Fred Hutchinson Cancer Research Center,
Seattle, WA 98104.
sites. Thus, the restriction map of the DNA upstream of the5' barren region is used to characterize a given ES.The molecular mechanisms involved in VSG gene regula-
tion are not understood, in part because little is known aboutthe actual structure of the ES's. Recently it has been shownthat a gene situated immediately upstream of the 5' barrenregion in Trypanosoma brucei ES's is coordinately regulatedwith the inserted VSG gene (7). This supports the idea thatthe regulation of ES activation occurs 5' to the barrenregions.We report here the cloning and characterization of a
genomic fragment containing the 5' end of the expressedVSG-1 gene of T. equiperdum BoTat-178 and 11 kilobases(kb) of ES sequences located 5' to the inserted gene. A9.2-kb segment of the cloned ES DNA is situated 5' to the 5'barren region. We term this region, located upstream of the5' barren region, the 1.78 ES. There are multiple apparentlyunlinked similar or identical copies of the 1.78 ES. InBoTat-1, which is the T. equiperdum VAT from which allother variants of the BoTat serodeme were derived, three ofthese copies are located near telomeres, and two of themhave been shown to accept VSG genes for expression. Thesetelomere-linked copies can be duplicated and deleted, as canthe inserted VSG genes. Other intact or partial copies of the1.78 ES exist both near and remote from telomeres.We also studied the fate of the ES's during antigenic
variation by using hybridization probes specific for the siterather than for the inserted VSG genes. Our results show thefollowing. (i) Replacement of the VSG gene occupying an ESprobably occurs only during antigenic variation. (ii) Thepreviously active VSG gene ELC is usually replaced in anES regardless of whether that ES is used for expression ofthe next VSG gene. (iii) VSG genes can be duplicated andtransposed into an ES independently of their expression. (iv)The active ES is preferentially used for expression of thenext VSG gene when non-VSG-1-expressing cells vary. (v)Variation of VSG-1-expressing trypanosomes results in the
2950

TRYPANOSOME VSG GENE EXPRESSION SITE 2951
S H P Xb H A H1B A20 Ps p SI_ I 1X P1P2HoM Rs
-4
B HoM
1 kb1IEG 1.78-1
p EG 1.78-5pEG 1.784pEG 1.78-6pEG 1.78-2pEG 1.78-3pEG 1.78-1
PsVI - \l I
BC I
R
cRSPS P Xb BgA1 H BA20 Rs R R tI III I I \'%XI I 1 _
X pip242'4P BR ELC17
B$g R S P Xib I H B A2O Ps SHA2PB t
I I I I I \ B1I
IR y
P17 AXo\ SR/ 1 1x Pi P2HoMPs DnSs Ho0 M
201
P Xb BA H B A20rl \ I I
X PzHoM BRHIa A2 Pl r l
M
FIG. 1. Restriction maps of 1.78 ES's. (a) Region of the 1.78 ES C site cloned in lambda EG1.78-1 and its subclones. (b) Map of the VSG-1BC in BoTat-178. (c) Maps of the three 1.78 ES A, B (in BoTat-1), and C (in BoTat-178) sites. Abbreviations: BR, 5' barren region; t, telomere;Al, AlaI; A,, Aiall; B, BamHI; Bg, BglII; D, DdeI; H, HindIlI; Ha, HaeIII; M, MspI; P, PstI; P1, Pvul; P,, P1'iiI1; R, EcoRl; Rs, Rsal: Ss,Sstl; X, XhoI; Xb, XbaI.
in situ inactivation of the VSG-1 ELC and the activation ofa different ES in the daughter variant.
MATERIALS AND METHODS
Abbreviations and glossary. VSG, Variable surface glyco-protein; the unique glycoprotein that coats the bloodstreamforms of African trypanosomes. BC, Silent basic copy of aVSG gene. ELC, Expression-linked copy of a VSG gene,i.e., the duplicated gene in an ES. R-ELC, Residual expres-sion-linked copy of a VSG gene; refers to a duplicated VSGgene, in an ES, which has been inactivated in situ. ES,Expression site; refers to the specific site in which VSGgenes can be expressed. VAT, Variant antigen type; refersto any trypanosome clone expressing a given VSG. BoTat,Bordeaux trypanozoon antigen type; includes all clonallyisolated VATs in the Bordeaux serodeme (6).
Trypanosomes. The relationships among the variants usedhere are shown in Fig. 2A. The number after BoTat- desig-nates the VSG expressed by the trypanosome clone. Thesuperscript numbers represent the parental clone. All of thetrypanosomes used in this work were of the BoTat serodeme(6) and were clonally derived from a single BoTat-1 parasite.Genomic mapping. Restriction endonucleases were from
commercial sources and were used as indicated by thesuppliers. Digested DNA was fractionated in agarose gelsand transferred to nitrocellulose paper by the method ofSouthern (17). Hybridization to 32P-labeled plasmids wasdone as previously described (10). Oligonucleotide probeswere labeled with T4 polynucleotide kinase (11). Hybridiza-tion with oligonucleotides was done at 42°C in 6x SSPE (1 xSSPE is 0.18 M NaCI-1 mM EDTA-10 mM NaPO4 [pH7])-50% formamide, followed by washings in 2x SSPE atroom temperature and then at 60°C.BAL 31 exonuclease digestion. An 80-,ug amount of try-
panosome DNA was treated with 140 U of BAL 31 exonu-
clease as described previously (16). After various times, thereactions were terminated by the addition of EDTA to a finalconcentration of 50 mM and were heated to 65°C for 15 min.The incubation mixtures were then dialyzed against theappropriate buffer for subsequent restriction endonucleasedigestion.
Molecular cloning. DNA fragments, size fractionated inlow-melting-point agarose gels, were extracted from the gelswith phenol and precipitated with ethanol. The DNA wasthen ligated into appropriately digested plasmid pUC9 (20)and used for transformation of Escherichia coli 1046.Ampicillin-resistant colonies were screened by colony hy-bridization (11) with the 32P-labeled insert from the VSG-1-specific cDNA plasmid pTel.612 (10) as probe. Appropriatesubclones were prepared in pUC9, pSP64, or pSP65 (12). Alibrary of Sall-digested BoTat-178 DNA was constructed inlambda EMBL-3 as described previously (13). The librarywas screened with 32P-labeled pTel.612 and pEG1.78-2.Recombinant phage which hybridized with both probes were
kept. Phage were grown, and DNA was extracted as de-scribed by Maniatis et al. (11).
RESULTS
Molecular cloning of the 1.78 ES. The cloning of the DNAlocated 5' to the duplicated VSG gene ELCs has beenhindered by the presence of long regions of tandem repeti-tions of an A-T-rich 76-base-pair sequence (5). These 5'barren regions are very poor in restriction endonucleasesites and are often very long (>25 kb). Therefore, we soughtan ELC-containing ES which had a short 5' barren region.Among the VSG gene ES's which we have characterized,the one containing the VSG-1 ELC in trypanosome cloneBoTat-178 was found to have only 1.8 kb of barren regionDNA 5' to the VSG-1 ELC (Fig. 1) (10). This region, the 1.78ES, was also of interest because it is used for VSG gene
( a )
( b )
(c )
,P2 t
A§s R1 ? 0 t
VOL. 6, 1986

2952 RAIBAUD ET AL.
B 2 3 4 14P 78 F r20 28
A1
2013134-142 12b
i2c12d12a
78-178- 2011.78
28 128aL128b
23.3-
A _ma __ _ __
9.5- pie
6.4-
4.2-B _t,i4.
i..E.;!:.
-1'1 4
c -
2.2-1.8 _ _ _ ___dwd
-20
FIG. 2. Multiple copies of the 1.78 ES and observed rearrangements in different trypanosome clones. (A) Genealogy of trypanosomevariants. All of the T. equiperdum variants were derived either directly or indirectly from BoTat-1 as described by Capbern et al. (6) (B)Southern blot of DdeI-digested DNA from trypanosome clones (indicated at the top of each lane) hybridized to pEG1.78-2. Molecular sizesare marked in kilobases. The bands marked A and B are fragments known to arise from ES's. Asterisks mark bands which have also beenshown to contain active ELCs.
expression more frequently than the other sites that we haveanalyzed (3, 4). Restriction maps of the BoTat-178 ELC andof the corresponding VSG-1 BC gene are shown in Fig. 1. Itcan be seen that the regions 5' to the VSG-1 BC and ELCgenes diverge somewhere between the 5'-most EcoRI site inthe transposed region and the RsaI site located 1.8 kbupstream from it in the ES.We first cloned the 2.9-kb fragment extending from the
BamHI site located 5' to the barren region to the Sall sitewithin the VSG-1 gene. After cleavage of BoTat-178 DNAwith BamHI and SalI, 3.8- to 4-kb-long fragments werepurified and cloned into BamHI-SalI-digested plasmidpUC9. Transformed E. coli colonies were screened with apurified insert from the VSG-1 cDNA clone pTel.612 (10).Recombinant plasmids from positive colonies were furthercharacterized by restriction endonuclease analysis. PlasmidpEG1.78-1 contained all of the predicted restriction sites(Fig. 1). We then prepared two subclones, pEG1.78-2 andpEG1.78-3, containing sequences specific to the 1.78 ES butnot to the VSG-1 region (Fig. 1).Having succeeded in obtaining a 1.78 ES genomic clone
which spanned the barren region, we then isolated a longerclone in bacteriophage lambda. In this case, the 11.7-kbSall-SalI fragment was cloned in EMBL-3 (13). Plaqueswere scored for hybridization with both pTel.612 andpEG1.78-2. Those phage that were positive for both probeswere further examined for the presence of predicted restric-
tion endonuclease sites. Restriction maps of lambdaEG1.78-1 and of its subclones are shown in Fig. 1. All of thesubclones containing the PstI-XhoI fragment were found tobe rearranged with respect to their in vivo configuration(data not shown). Nevertheless, clone pEG1.78-5 (PstI-BamHI) was used for Southern blot analysis (see below).
Characterization of the cloned 1.78 ES region. We firstexamined the cloned 1.78 ES for the presence of the 76-bprepeat sequences reported to make up the barren regionlocated 5' to a VSG gene in a T. brucei ES (5). DNA fromlambda EG1.78-1 was digested with RsaI and EcoRI andanalyzed by Southern blotting using as probe a 42-basesynthetic oligonucleotide corresponding to the 3' end of the76-bp repeat sequence. The oligomer hybridized only to the1.8-kb fragment corresponding to the expected barren regionin the 1.78 ES clone (data not shown).There are multiple 1.78 ES's. T. equiperdum contains
multiple telomere-linked ES's for VSG genes (10), and wehave assumed that, when not in use, an ES contains aninactive VSG gene. Having clones corresponding to the ESrather than to the inserted VSG gene allowed us to examinethe 1.78 ES as a function of whether it was active in a givenvariant. Therefore, we performed Southern blot analysis onDNAs from variants the ELCs of which were previouslydetermined to be either present in or absent from the 1.78ES. The lineage of the VATs used here is shown in Fig. 2A.The genomic DNAs were digested with enzymes that do not
do
MOL. CELL. BIOL.

TRYPANOSOME VSG GENE EXPRESSION SITE 2953
TABLE 1. Use and fate of ES's during antigenic variation
VariationES used by: Fate ofMode of pVaration ES used by: parent":activation (parenter)daughter) Parent Daughter ELC ES
Duplicative 1 - 2 1 1.78 B R U1 -4 1 1.78A R U1 3 1 NDb L ND1 -13 1 ND L ND2 > 12d 1.78 B 1.2d L C2 d 12c 1.78 B 1.78 B L C4 14 1.78A 1.78 A L C78-> 17 1.78 C 1.78 C L C28 >128 1.28 1.28 L C28 > 12" 1.28 1.28 L C
Nonduplicative 17 -. 201178 1.78 C 1.78 B R U2 1'a 1.78B 1 L C2 a 2'b 1.78 B 1 L C2 b 12c 1.78 B 1 L C
a Abbreviations: R, Residual ELC; L, lost; U, unchanged; C, converted.b ND, Not determined.
cut within the 5' barren regions. Thus, the generated frag-ments would have one terminus upstream of the 5' barrenregion in the 1.78 ES and the other terminus within theVSG-transposed region occupying the ES. Thus, it wasreasoned that different inserted VSG genes would give riseto fragments of various sizes since the cutting sites forvarious enzymes would be different. In variants the ELCs ofwhich are in the 1.78 ES, the variable fragments shouldcorrespond to the ELC-containing fragments revealed byprobing with VSG-specific cDNA clones.DNA from 11 variants was digested with DdeI and hybrid-
ized to 32P-labeled pEG1.78-2. The results are shown in Fig.2B. Bands marked with an asterisk were also recognized bycDNA probes specific for the ELC of the given variant.
Three types of fragments were revealed by the ES-specificprobe. First, there are invariant fragments. Second, thereare two fragments, labeled A and B, which are missing insome variants and replaced by other bands. Finally, theDNAs from BoTat-178 and its parent, BoTat-78, contain asupplementary fragment. Hybridization with the VSG-1cDNA probe pTel.612 revealed that this supplementaryfragment in BoTat-178 contains the ELC (data not shown).This fragment will be subsequently referred to as fragmentC. Thus, fragment C arises from the 1.78 ES in BoTat-178.We have previously characterized other T. equiperdum
variants, BoTat-14 (4), BoTat-201 (1), BoTat-2 (3), BoTat-4(9a), BoTat-78 (9a), and BoTat-20 (unpublished data), theELCs of which are in ES's with restriction maps closelyresembling that of the 1.78 ES. The ELC for each variantcorresponds either to the fragment replacing A or B or tofragment C (Fig. 2B). The ES's used by the variants theDNA of which was examined in Fig. 2B are listed in Table 1.The 1.78 ES A site is used by BoTat-4 and BoTat-14, the Bsite is used by BoTat-2, BoTat-201'178, and BoTat-20, andthe C site is used by BoTat-78 and BoTat-178. These resultsindicate that BoTat-1 contains at least two similar copies ofthe 1.78 ES and that both can act as acceptors for duplicatedVSG genes. Furthermore, in BoTat-78 and BoTat-178, thereare at least three potentially competent 1.78 ES's.Because in BoTat-1 the active VSG-1 ELC is not in any of
the 1.78 ES's (10), it was of interest to characterize the A andB 1.78 ES's in this variant to determine whether they containinactive VSG genes. The restriction map of the A site and ofthe gene occupying it in BoTat-1 was determined by com-paring BoTat-1 DNA with that of BoTat-14, in which theactive VSG-1 gene occupies the 1.78 ES A site. GenomicDNAs from the two variants were digested with enzymeswhich do not cut in the barren regions and were compared onSouthern blots probed with the 1.78 ES-specific probepEG1.78-2. Because in BoTat-14 the VSG-1 gene has re-placed the gene that occupied the 1.78 A site in BoTat-1,
B A\23.35 -
9.5C *§6.4- to
4.2-
_ *4
2.2-1.8-
A S P X B RsRsI I IA
5 3 2 6 5 4
FIG. 3. (A) Restriction map of the cloned 1.78 ES and the various subclones used as probes. (B) Southern blot of EcoRl-digested BoTat-178DNA hybridized to various 1.78 ES subclones. The numbers under the lanes represent the subclone used as a probe. Molecular sizes aremarked in kilobases.
VOL. 6, 1986

2954 RAIBAUD ET AL.
fragments of different sizes would be expected to be gener-ated. Thus, fragments which were present only in the digestsof BoTat-1 DNA were assumed to arise from the 1.78 A ES.Similarly, the configuration of the B site in BoTat-1 wasdetermined by comparison with DNA from BoTat-20, inwhich it is rearranged and contains the VSG-20 ELC. Thededuced restriction maps of the 1.78 ES A and B sites inBoTat-1 are shown in Fig. 1. It can be seen that therestriction map of the 1.78 ES B site is identical to that of thecloned C site, suggesting that the C site arose by duplicationof the B site. Whereas the restriction map of the A site isvery similar to that of the B and C sites, it is not identical.We have identified the gene in the B site in BoTat-1 as theVSG-201 gene which was activated by a nonduplicativeevent when BoTat-178 was grown in culture (1). BoTat-2011.78 contains a residual, inactive VSG-1 ELC (R-ELC)(1). The A, B, and C sites are associated with telomeres inBoTat-1 DNA since they were preferentially digested byexonuclease BAL 31 (see below).To further characterize the 1.78 ES, Southern blots of
EcoRI-digested BoTat-178 DNA were analyzed with varioussubclones of lambda EG1.78 as hybridization probes. Theresults are shown in Fig. 3. The fragments containing the A,B, and C sites, to which all of the subclones hybridized, aremarked. One other fragment, marked X, also hybridized toall of the probes. A similar fragment which hybridized to allof the subclones was seen when the genomic DNA wasdigested with Sall and with PstI (see Fig. 4). Thus, it appearsthat the X fragment contains sequences related to all of theregion of the 1.78 ES. It is also apparent that the regionhybridizing to subclones pEgl.78-2, pEG1.78-3, pEG1.78-5,and pEG1.78-6 exists in at least three other copies within thegenome. It is also evident that the 5'-most subclonepEG1.78-4 recognizes a family of sequences not recognizedby the other subclones.
Telomere linkage. Restriction enzyme mapping and previ-ous studies have shown that the 1.78 A, B, and C ES's aretelomere linked (3, 4, 10). However, it was of interest toknow whether the other 1.78 ES-related sequences are alsolocated near telomeres. To determine this, BoTat-1 DNAwas digested for various times with the exonuclease BAL 31.It was then further digested with various restrictionendonucleases and subjected to Southern blot analysis withpEG1.78-2 DNA as probe. Depending upon the restrictionendonuclease used to digest the DNA, the 1.78 ES's A, B,and C are located in regions which are sensitive to BAL 31digestion (Fig. 4). Thus, they are associated with chromo-some telomeres, as expected. Two other 1.78 ES-relatedsequences were also localized near BAL 31-sensitive re-gions. Region X, which appears to contain an entire copy ofthe 1.78 ES, is telomere linked (Fig. 4C and D). However, inboth experiments, the X fragment appeared as an intenseband, only approximately half of which was sensitive to theexonuclease. Similar results were found with different DNApreparations and also with different restriction endonucle-ases. A fifth sequence, marked Y, is also telomere linked.This fragment does not contain all of the 1.78 ES sequencessince it does not hybridize to pEG1.78-4 (data not shown).
Fate of ES's during antigenic variation. The results shownin Fig. 2B and Table 1 demonstrate several additional pointsconcerning the fate of ES's during antigenic variation. First,the genes occupying the ES's can remain unchangedthroughout the entire cycle of variation. For example, nei-ther the 1.78 ES A site nor the B site was altered in thesequence of variations leading from BoTat-1 to BoTat-28 (alate variant) and back to BoTat-128. Similarly, the A site
AA-A-~BXG
C--
,.W 411....
.~* is .e
- - - -
- -!,- --AM a b c d e
M a b c d e f
M a b c d e f
BA-
C- v;-
abc de f
DA--_Ip,
a b cd e fFIG. 4. BAL 31 exonuclease sensitivity of regions containing
1.78 ES homologous sequences. BoTat-178 DNA was treated withBAL 31 as described in Materials and Methods. Treatment was for(lanes) a, 0; b, 5; c, 15; d, 25; e, 40; and f, 60 min. M, Molecular sizestandards (HindIll fragments). Lanes: a, 23.3; b, 9.5; c, 6.4; d, 4.2;e, 2.2; and f, 1.8 kb. After BAL 31 digestion, the DNA was treatedwith (A) PstI; (B) HindlIl; (C) Sall; (D) EcoRI. Southern blotanalysis was performed, and the blots were probed with pEG1.78-2.
remained unchanged in the variations leading from BoTat-1to the late variant BoTat-20. Since both BoTat-28 andBoTat-20 are very late variants, these results indicate thatES's can be very stable. In contrast, once an ES is activated,it becomes highly unstable in subsequent variations. The1.78 ES A and B sites were first used for VSG geneexpression in VAT-4 and VAT-2, respectively. The 1.78 ESC site, which probably arose by duplication of the B site,first appeared in BoTat-78, where it contains the activeVSG-78 ELC. In all of the variants derived from theseVATs, the parental ES has been altered. In other words, thegenes occupying the previously active ES's were replaced.Furthermore, in BoTat-14, BoTat-178, BoTat-128a, andBoTat-128b the parental ES was chosen for expression of theVSG-1 ELC.To better understand how the fate of active ES's is
determined during variation in T. equiperdum, we analyzedthe DNA from four independently derived VSG-1-ex-pressing isolates derived from BoTat-2 (3). We have previ-ously shown that BoTat-2 contains an R-ELC of the VSG-1
MOL. CELL. BIOL.
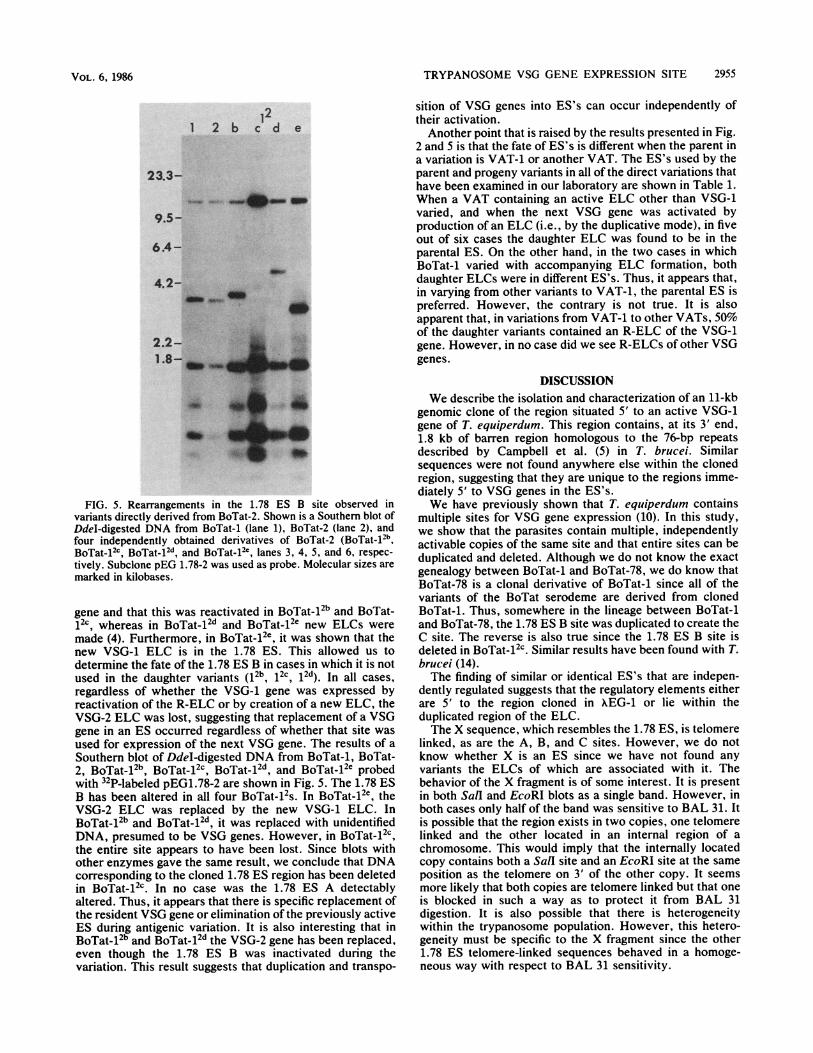
TRYPANOSOME VSG GENE EXPRESSION SITE 2955
l 2 b cd e
23.3-
9.5-
6A-
4.2-
2.2-1.8-
FIG. 5. Rearrangements in the 1.78 ES B site observed in
variants directly derived from BoTat-2. Shown is a Southern blot of
DdeI-digested DNA from BoTat-1 (lane 1), BoTat-2 (lane 2), and
four independently obtained derivatives of BoTat-2 (BoTat-12b,BoTat-1Zc, BoTat.12d and BoTat.12c, lanes 3, 4, 5, and 6, respec-
tively. Subclone pEG 1.78-2 was used as probe. Molecular sizes are
marked in kilobases.
gene and that this was reactivated in BoTat-12b and BoTat-
12c whereas in BoTat-12d and BoTat-12e new ELCs were
made (4). Furthermore, in BoTat-12e, it was shown that the
new VSG-1 ELC is in the 1.78 ES. This allowed us to
determine the fate of the 1.78 ES B in cases in which it is not
used in the daughter variants (12b, 12c, 12d). In all cases,
regardless of whether the VSG-1 gene was expressed by
reactivation of the R-ELC or by creation of a new ELC, the
VSG-2 ELC was lost, suggesting that replacement of a VSG
gene in an ES occurred regardless of whether that site was
used for expression of the next VSG gene. The results of a
Southern blot of DdeI-digested DNA from BoTat-1, BoTat-
2, BoTat-12b BoTat-12c BoTat-12d and BoTat-12e probed
with 12P-labeled pEG1.78-2 are shown in Fig. 5. The 1.78 ES
B has been altered in all four BoTat-12S. In BoTat-12e, the
VSG-2 ELC was replaced by the new VSG-1 ELC. In
BoTat-12b and BoTat-12d it was replaced with unidentified
DNA, presumed to be VSG genes. However, in BoTat-12c,
the entire site appears to have been lost. Since blots with
other enzymes gave the same result, we conclude that DNA
corresponding to the cloned 1.78 ES region has been deleted
in BoTat512c. In no case was the 1.78 ES A detectably
altered. Thus, it appears that there is specific replacement ofthe resident VSG gene or elimination of the previously active
ES during antigenic variation. It is also interesting that in
BoTat-12b and BoTat-12d the VSG-2 gene has been replaced,even though the 1.78 ES B was inactivated during the
variation. This result suggests that duplication and transpo-
sition of VSG genes into ES's can occur independently oftheir activation.Another point that is raised by the results presented in Fig.
2 and 5 is that the fate of ES's is different when the parent ina variation is VAT-1 or another VAT. The ES's used by theparent and progeny variants in all of the direct variations thathave been examined in our laboratory are shown in Table 1.When a VAT containing an active ELC other than VSG-1varied, and when the next VSG gene was activated byproduction of an ELC (i.e., by the duplicative mode), in fiveout of six cases the daughter ELC was found to be in theparental ES. On the other hand, in the two cases in whichBoTat-1 varied with accompanying ELC formation, bothdaughter ELCs were in different ES's. Thus, it appears that,in varying from other variants to VAT-1, the parental ES ispreferred. However, the contrary is not true. It is alsoapparent that, in variations from VAT-1 to other VATs, 50%of the daughter variants contained an R-ELC of the VSG-1gene. However, in no case did we see R-ELCs of other VSGgenes.
DISCUSSIONWe describe the isolation and characterization of an 11-kb
genomic clone of the region situated 5' to an active VSG-1gene of T. equiperdum. This region contains, at its 3' end,1.8 kb of barren region homologous to the 76-bp repeatsdescribed by Campbell et al. (5) in T. brucei. Similarsequences were not found anywhere else within the clonedregion, suggesting that they are unique to the regions imme-diately 5' to VSG genes in the ES's.We have previously shown that T. equiperdum contains
multiple sites for VSG gene expression (10). In this study,we show that the parasites contain multiple, independentlyactivable copies of the same site and that entire sites can beduplicated and deleted. Although we do not know the exactgenealogy between BoTat-1 and BoTat-78, we do know thatBoTat-78 is a clonal derivative of BoTat-1 since all of thevariants of the BoTat serodeme are derived from clonedBoTat-1. Thus, somewhere in the lineage between BoTat-1and BoTat-78, the 1.78 ES B site was duplicated to create theC site. The reverse is also true since the 1.78 ES B site isdeleted in BoTat-12c. Similar results have been found with T.brucei (14).The finding of similar or identical ES's that are indepen-
dently regulated suggests that the regulatory elements eitherare 5' to the region cloned in AEG-1 or lie within theduplicated region of the ELC.The X sequence, which resembles the 1.78 ES, is telomere
linked, as are the A, B, and C sites. However, we do notknow whether X is an ES since we have not found anyvariants the ELCs of which are associated with it. Thebehavior of the X fragment is of some interest. It is presentin both SalI and EcoRI blots as a single band. However, inboth cases only half of the band was sensitive to BAL 31. Itis possible that the region exists in two copies, one telomerelinked and the other located in an internal region of achromosome. This would imply that the internally locatedcopy contains both a SalI site and an EcoRI site at the sameposition as the telomere on 3' of the other copy. It seemsmore likely that both copies are telomere linked but that oneis blocked in such a way as to protect it from BAL 31digestion. It is also possible that there is heterogeneitywithin the trypanosome population. However, this hetero-geneity must be specific to the X fragment since the other1.78 ES telomere-linked sequences behaved in a homoge-neous way with respect to BAL 31 sensitivity.
.Idub
VOL. 6, 1986
Am mop-amt 40
.-vm

2956 RAIBAUD ET AL.
We also demonstrate that individual sequences within the1.78 ES are repeated several times in the T. equiperdumgenome and that not all of these homologous sequencesappear to be located at chromosome telomeres. We arepresently isolating genomic clones containing these se-quences to determine whether they are associated withbarren regions or with VSG genes.
It has been reported that there are frequent, large-scalechromosomal rearrangements involving chromosome endsin African trypanosomes (18). Our finding that the clonedDNA sequences located 5' to the barren region in the 1.78ES are repeated several times in the genome offers anexplanation for these rearrangements. Reciprocal recombi-nation between a segment of the ES and a homologoussequence located at an internal region of another chromo-some would result in the transfer of the ES to anotherchromosome.The fate of an ES during the process of antigenic variation
appears to be very different depending on whether it wasused by the last ELC. When an ES is used for expression ofVSG genes other than VSG-1, it is always (in nine out of ninecases observed) converted in the next variation. In most ofthe cases that we observed, the previously expressed ELCwas replaced, even when the newly incorporated gene wasnot used in the next variant. However, in one variation(BoTat-2 to BoTat-12c) the entire ES was lost.
It is also apparent that an active ES has a high probabilityof being used for the next variant (in five out of six casesobserved). On the other hand, we show here that if an ES isnot activated it has a very low probability of having the VSGgene in it replaced. These findings indicate that there is afundamental difference between an active and an inactive ESin that the active ES is a better target for the gene conversionevents which change the VSG genes in the ES's. This mightbe due to a different chromatin configuration or to the factthat the active VSG gene is transcribed.The exceptions to the finding that the active parental ES is
always converted in the subsequent variant are those varia-tions in which VAT-1 is the parent. In the three such casesexamined (Table 1), the ELCs in the progeny VATs were allin new ES's. Furthermore, of the five early variants exam-ined, three contain residual VSG-1 ELCs. We did notobserve an R-ELC of any other VSG gene. Thus, it appearsthat variation from VAT-1 allows for both the activation ofnew ES's and the inactivation without removal of the VSG-1ELC.
ACKNOWLEDGMENTS
This work was supported by grants from the Centre National de laRecherche Scientifique and the Institut National de la Sante et de larecherche Mddicale, France, and from the European community.We are grateful for discussions with S. Longacre and C. Roth. We
also thank G. Smith and M. Groudine for criticism of the manu-script.
LITERATURE CITED1. Baltz, T., C. Giroud, D. Baltz, C. Roth, A. Raibaud, and H.
Eisen. 1986. Stable expression of two variable surface glycopro-teins by cloned Trypanosoma equiperdum. Nature (London)319:602-604.
2. Borst, P., and G. A. M. Cross. 1982. Molecular basis fortrypanosome antigenic variation. Cell 29:291-303.
3. Buck, G. A., C. Jacquemot, T. Baltz, and H. Eisen. 1984.Re-expression of an inactivated variable surface glycoproteingene in Trypanosome equiperdum. Gene 32:329-336.
4. Buck, G. A., S. Longacre, A. Raibaud, U. Hibner, C. Giroud, T.
Baltz, D. Baltz, and H. Eisen. 1984. Stability of expression-linked surface antigen gene in Trypanosoma equiperdum. Na-ture (London) 307:563-565.
5. Campbell, D. A., M. P. Van Bree, and J. C. Boothroyd. 1984.The 5'-limit of transposition and upstream barren region of atrypanosome VSG gene: tandem 76 base-pair repeats flanking(TAA)90. Nucleic Acids Res. 12:2759-2774.
6. Capbern, A., C. Giroud, T. Baltz, and P. Mattern. 1977.Trypanosoma equiperdum: etude des variations antigeniques aucours de la trypanosomose experimentale du lapin. Exp.Parasitol. 42:6-13.
7. Cully, D., H. Ip, and G. Cross. 1985. Coordinate transcription ofvariant surface glycoprotein genes and an expression site asso-ciated gene family in Trypanosoma brucei. Cell 42:173-182.
8. De Lange, T., and P. Borst. 1982. Genetic environment of theexpression-linked extra copies of genes for surface antigens ofTrypanosoma brucei resembles the end of a chromosome.Nature (London) 299:451-453.
9. Hoeijmarkers, J. H. J., A. C. C. Frasch, A. Bernards, P. Borst,and G. A. M. Cross. 1980. Novel expression-linked copies of thegenes for variant surface antigens in trypanosomes. Nature(London) 284:78-80.
9a.Longacre, S., and H. Eisen. 1986. Expression of whole andhybrid genes in Trypanosoma equiperdum antigenic variation.EMBO J. 5:1057-1063.
10. Longacre, S., U. Hibner, A. Raibaud, H. Eisen, T. Baltz, C.Giroud, and D. Baltz. 1983. DNA rearrangements and antigenicvariation in Trypanosoma equiperdum: multiple expression-linked sites in independent isolates of trypanosomes expressingthe same antigen. Mol. Cell. Biol. 3:399-409.
11. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning, a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
12. Melton, D., P. Krieg, M. Rebagliati, T. Maniatis, K. Zinn, andM. Green. 1984. Efficient in Oitro synthesis of biologically activeRNA and RNA hybridization probes from plasmids containing abacteriophage sp6 promoter. Nucleic Acids Res. 12:7035-7056.
13. Murray, N. E. 1983. Phage lambda and molecular cloning, p.395-432. In R. W. Hendrix, J. W. Roberts, F. W. Stahl, andR. A. Weisberg (ed.), Lambda ll. Cold Spring Harbor Labora-tory, Cold Spring Harbor, N.Y.
14. Pays, E., M. F. Delauw, S. Van Assel, M. Laurent, T. Vervoort,N. Van Meirvenne, and M. Steinert. 1983. Modifications of aTrypanosoma brucei antigen gene repertoire by different DNArecombinational mechanisms. Cell 35:721-731.
15. Pays, E., N. Van Meirvenne, D. Le Ray, and M. Steinert. 1981.Gene duplication and transposition linked to antigenic variationin Trypanosoma brucei. Proc. Natl. Acad. Sci. USA78:2673-2677.
16. Raibaud, A., C. Gaillard, S. Longacre, U. Hibner, G. Buck, G.Bernardi, and H. Eisen. 1983. Genomic environment of variantsurface antigen genes of Trypanosoma equiperdum. Proc. Natl.Acad. Sci. USA 80:4306-4310.
17. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by electrophoresis. J. Mol. Biol.98:503-517.
18. Van der Ploeg, L. H. T., A. W. C. A. Cornelissen, P. A. M.Michels, and P. Borst. 1984. Chromosome rearrangements inTrypanosoma brucei. Cell 39:213-221.
19. Vickerman, K. 1978. Antigenic variation in trypanosomes. Na-ture (London) 273:613417.
20. Vieira, J., and J. Messing. 1982. The pUC plasmids, an M13mp7-derived system for insertion mutagenesis and sequencingwith synthetic universal primers. Gene 19:259-268.
21. Williams, R. O., J. R. Young, and P. A. 0. Majiwa. 1979.Genomic rearrangements correlated with antigenic variation inTrypanosoma brucei. Nature (London) 282:847-849.
22. Williams, R., J. Young, and P. Majiwa. 1982. Genomic environ-ment of T. brucei VSG genes: presence of a minichromosome.Nature (London) 299:417-421.
23. Young, J. R., E. N. Miller, R. 0. Williams, and M. J. Turner.1983. Are there two classes of VSG genes in Trypanosomabrucei? Nature (London) 306:196-198.
MOL. CELL. BIOL.