characterization of a signaling system in streptococcus ... · s. mitis spar10 alch01000001.1...
TRANSCRIPT

Characterization of a Signaling System in Streptococcus mitisThat Mediates Interspecies Communication with Streptococcuspneumoniae
R. Junges,a K. Sturød,a G. Salvadori,a H. A. Åmdal,a T. Chen,b F. C. Petersena
aInstitute of Oral Biology, Faculty of Dentistry, University of Oslo, Oslo, NorwaybDepartment of Microbiology, Forsyth Institute, Cambridge, Massachusetts, USA
ABSTRACT Streptococcus mitis is found in the oral cavity and nasopharynx andforms a significant portion of the human microbiome. In this study, in silico analysesindicated the presence of an Rgg regulator and short hydrophobic peptide (Rgg/SHP) cell-to-cell communication system in S. mitis. Although Rgg presented greatersimilarity to a repressor in Streptococcus pyogenes, autoinducing assays and geneticmutation analysis revealed that in S. mitis Rgg acts as an activator. Transcriptomeanalysis showed that in addition to shp, the system regulates two other downstreamgenes, comprising a segment of a putative lantibiotic gene cluster that is in a conju-gative element locus in different members of the mitis group. Close comparison to asimilar lantibiotic gene cluster in Streptococcus pneumoniae indicated that S. mitislacked the full set of genes. Despite the potential of SHP to trigger a futile cycle ofautoinduction, growth was not significantly affected for the rgg mutant under nor-mal or antibiotic stress conditions. The S. mitis SHP was, however, fully functional inpromoting cross-species communication and increasing S. pneumoniae surface poly-saccharide production, which in this species is regulated by Rgg/SHP. The activity ofSHPs produced by both species was detected in cocultures using a S. mitis reporterstrain. In competitive assays, a slight advantage was observed for the rgg mutants.We conclude that the Rgg/SHP system in S. mitis regulates the expression of its ownshp and activates an Rgg/SHP system in S. pneumoniae that regulates surface poly-saccharide synthesis. Fundamentally, cross-communication of such systems mayhave a role during multispecies interactions.
IMPORTANCE Bacteria secrete signal molecules into the environment which aresensed by other cells when the density reaches a certain threshold. In this study, wedescribe a communication system in Streptococcus mitis, a commensal species fromthe oral cavity, which we also found in several species and strains of streptococcifrom the mitis group. Further, we show that this system can promote cross-communication with S. pneumoniae, a closely related major human pathogen. Im-portantly, we show that this cross-communication can take place during coculture.While the genes regulated in S. mitis are likely part of a futile cycle of activation, thetarget genes in S. pneumoniae are potentially involved in virulence. The understand-ing of such complex communication networks can provide important insights intothe dynamics of bacterial communities.
KEYWORDS Streptococcus, mutagenesis, quorum sensing, transcriptional regulation,virulence
The mitis group of streptococci comprises a variety of species such as Streptococcuspneumoniae, Streptococcus mitis, Streptococcus oralis, Streptococcus gordonii, Strep-
tococcus sanguinis, and Streptococcus parasanguinis (1, 2). Although most of these
Citation Junges R, Sturød K, Salvadori G, ÅmdalHA, Chen T, Petersen FC. 2019. Characterizationof a signaling system in Streptococcus mitis thatmediates interspecies communication withStreptococcus pneumoniae. Appl EnvironMicrobiol 85:e02297-18. https://doi.org/10.1128/AEM.02297-18.
Editor Donald W. Schaffner, Rutgers, The StateUniversity of New Jersey
Copyright © 2019 American Society forMicrobiology. All Rights Reserved.
Address correspondence to R. Junges,[email protected], or F. C. Petersen,[email protected].
Received 20 September 2018Accepted 25 October 2018
Accepted manuscript posted online 2November 2018Published
GENETICS AND MOLECULAR BIOLOGY
crossm
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 1Applied and Environmental Microbiology
9 January 2019
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

members are often regarded as commensals, they present a wide range of contrastingpathogenic potentials. While S. pneumoniae is a major human pathogen responsible fornoninvasive and invasive diseases, such as otitis media and pneumonia, other species,such as S. mitis, are rarely associated with disease and present mostly a nonvirulentbehavior (3, 4). Interestingly, these two species in particular present a high geneticsimilarity, which raises questions regarding the specific factors that contribute to therelevant pathogenic potential of S. pneumoniae (4, 5).
The bacterial behavioral change from a commensal to a pathogen is complex andlikely dependent on a number of factors and cofactors that involve both the microor-ganism and the host (6, 7). One of these contributing stimuli is thought to be cell-to-cellcommunication in bacteria. By utilizing these systems, bacteria can communicate andestablish group behaviors such as virulence, biofilm formation, and others (8, 9). InGram-positive bacteria, specifically, this communication is often regulated by smallpeptides that act as pheromones (10–12). Regulator/short hydrophobic peptide (Rgg/SHP) systems belong to the RRNPP family of peptide-binding regulators (Rgg, Rap,NprR, PlcR, and Prg) and have been gaining increased attention due to their involve-ment in the regulation of behavioral change in streptococci (11, 12). More importantly,such systems pose as interesting targets for nonantibiotic based strategies to disruptbacterial behavior and thus fight infections (9, 13).
Rgg (regulator gene of glucosyltransferase) was first described in S. gordonii (14)and, at the time, defined as a stand-alone transcriptional regulator. With time, moreRgg-like regulators were discovered in different species of the phylum Firmicutes(15–18). In 2007, Ibrahim et al. described a cyclic peptide (Pep1357C) that formed apotential novel quorum-sensing system in S. thermophilus (19). Ultimately, severalstreptococcal species and strains were scanned for small open reading frames (ORFs)close to Rgg-like regulators (19, 20). These pheromone systems control a variety offunctions in several species of streptococci, such as the production of SpeB and biofilmdevelopment in S. pyogenes (16, 21, 22), bacteriocin production in S. mutans (17), andvirulence regulation in S. agalactiae and S. pyogenes (23–25). Moreover, in the pyogenic,salivarius, bovis, and mutans groups of streptococci, natural competence for transfor-mation is regulated by the ComRS system, which belongs to the Rgg/SHP family (26).In the mitis group of streptococci, a TprA/PhrA quorum-sensing system that regulatesa lantibiotic biosynthesis gene cluster (27) and an Rgg/SHP system that regulatessurface polysaccharide expression (28) have both been recently reported in S. pneu-moniae. While interspecies cross-communication has been detected in some of thesesystems (29–31), it is important to note that the stringency of SHP recognition by Rggproteins across species is variable and, in general, a high specificity of the Rgg and SHPinteraction is expected.
Here, we report the identification of an Rgg/SHP cell-to-cell communication in an S.mitis type strain, which we also found present in the genomes of other oral streptococciof the mitis group. The regulator functions as an activator and controls the transcriptionof a lantibiotic gene cluster segment that is present in a conjugative element locus ofdifferent species and strains of streptococci from the mitis group. Further, we show thatthis system can cross-communicate with S. pneumoniae and trigger an increase inpneumococcal surface polysaccharide expression.
RESULTSScreening of Rgg/SHP cell-to-cell communication systems in oral streptococci
of the mitis group. A recent study characterized a quorum-sensing Rgg/SHP system inS. pneumoniae that increases surface polysaccharide synthesis by upregulation of a sitedistinct from the original capsule locus (28). Considering the genetic similarities be-tween the pathogenic S. pneumoniae and oral streptococci from the mitis group, thisstudy addressed whether other such systems could be identified in these closely relatedoral species as well. Rgg/SHP cell-to-cell communication systems are composed by aregulator (Rgg) and a short hydrophobic peptide (SHP) that are often adjacent in thegenome. As such, homology search was performed using the Rgg0939 sequence from
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 2
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

S. pneumoniae D39 as a reference against relevant mitis group members. The searchwas performed using the expanded Human Oral Microbiome Database (HOMD) (32), acurated and comprehensive database on the bacterial species present in the humanaerodigestive tract. Entries of species belonging to the mitis group of streptococci wereincluded in the search, with the only exclusion being S. pneumoniae since it wasrecently described (28). In total, we examined 15 strains of S. mitis, 1 strain of S. gordonii,9 strains of S. oralis, 13 strains of S. sanguinis, and 7 strains of S. parasanguinis. Theresults are presented in Table 1. The short coding sequences for SHP are often tightlylinked to the regulator gene and contain between 20 and 25 codons. In this analysis,close to 20% of the strains surveyed presented an ORF in the upstream region of Rggputatively coding for shp. In particular, 40% of the included S. mitis strains presentedthe sequence for a potential pheromone in the antisense strand upstream of theregulator. Such findings indicate that the putative cell-to-cell communication system isfrequently present in the oral commensal species of the mitis group. In the speciescontaining the sequence for a potential peptide pheromone, the downstream region toshp differed significantly among them. Further, there were also variations in shp thatmight affect the activation potential of these peptides. Since one of the strongesthomology hits was in the type strain of S. mitis, this system was further investigated inthis study.
The SHP produced by S. mitis acts as an autoinducer. The Rgg in S. mitis
(SM12261_0094) presents 74% identity and 87% similarity to the Rgg0939 in S. pneu-moniae. The predicted mature SHP sequence shows difference in only one isoleucineresidue at position 5: N-DIIIIVGG (S. pneumoniae D39) versus N-DIIIVGG (S. mitis typestrain). As one of the core characteristics of quorum-sensing systems is the positive-feedback loop created by the pheromone autoinducing its own gene, the shp mRNAexpression was assessed in cultures treated or not treated with the synthetic SHP (sSHP)from S. mitis (N-DIIIVGG). An SHP concentration of 4.5 �M was chosen based on studieswith other streptococcal species indicating that this concentration would saturate thesystem. Pellet collection for RNA extraction was performed at 2 h due to previousreports indicating high activation of the pheromone system at this time point (28). Inaddition, since we have been unable to find a chemically defined peptide-free mediumthat works well for S. mitis, we opted to test rich and semidefined media for activationof the system. As shown in Fig. 1A, in treated samples the shp expression was �200-foldhigher, and this induction was slightly higher in C�Y (semidefined medium) comparedto tryptone soya broth (TSB; rich medium). Conversely, the rgg expression remainedunaltered in both media and the treatment group (Fig. 1B). Such findings indicate thatthe Rgg/SHP system in S. mitis is active.
Transcriptome analysis of the response to sSHP. Rgg/SHP quorum-sensing sys-tems have been shown to regulate the expression of a variety of genes with differentfunctions in streptococci (11, 12, 20, 21, 23, 24, 28). RNA sequencing was performed toaddress the gene expression in samples treated or not treated with the sSHP. Thecomplexity of the medium has been shown to play a role in the activation potential ofquorum-sensing systems (24); thus, we assessed the patterns of expression both in TSBand in C�Y. Considering a threshold for an up- or downregulation of 2-fold and astatistically significant differential expression, as measured by DESeq (P � 0.05), therewere 13 genes differently expressed in TSB and 6 genes differently expressed in C�Y(Table 2). Two genes, SM12261_0093 and SM12261_0092, were strongly upregulatedand commonly identified in both conditions (Fig. 2A), and they are located justdownstream of shp, forming a single continuous transcript, as shown in Fig. 2B. In TSB,the rgg was also upregulated 5-fold. Interestingly, in C�Y, a three-gene iron-compoundtransport locus (SM12261_0514, SM12261_0515, and SM12261_0516) was downregu-lated almost 3-fold. Auxiliary quantitative reverse transcription-PCR (qRT-PCR) analysesconfirmed the upregulation of SM12261_0093 and SM12261_0092 while not confirm-ing the differential expression observed for rgg and the iron-compound transport locus(data not shown). Overall, the RNA sequencing data indicate that the main targets of
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 3
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

TAB
LE1
Hom
olog
yse
arch
for
Rgg0
939
inor
alst
rep
toco
cci
ofth
em
itis
grou
pa
Spec
ies
Acc
essi
onn
o.C
hro
mos
ome
Posi
tion
Eva
lue
Posi
tive
s(%
)Po
ten
tial
ph
erom
one
Dow
nst
ream
ofsh
p
Stre
ptoc
occu
sm
itis
S.m
itis
SK10
80A
FQV0
0000
000
cont
ig_N
Z_A
FQV0
1000
017
c637
20-6
2869
3.00
E–17
994
MKK
ISKF
LPIL
VLVM
DIII
IFG
Six
gene
s:tr
ansp
orte
r,hy
pot
hetic
alp
rote
in,
DN
Am
ethy
ltra
nsfe
rase
dom
ain
pro
tein
,C
of-li
kehy
drol
ase,
Cof
-like
hydr
olas
e,Ft
sY
AFQ
V010
0001
3.1
ctg1
1296
4536
6368
7260
-812
34.
00E–
3049
S.m
itis
SK32
1A
EDT0
0000
000
cont
ig_N
Z_A
EDT0
1000
009
c136
76-1
2825
1.00
E–17
794
MKK
ILKN
SLIL
LLVM
DIII
VGG
Two
gene
s:al
pha
-hem
olys
intr
ansl
ocat
ion
ATP
-bin
ding
pro
tein
Hly
B,hy
pot
hetic
alp
rote
inA
EDT0
1000
014.
1co
ntig
2451
20-5
983
2.00
E–32
51
S.m
itisT
CC
UG
3161
1/N
CTC
1226
1A
EDX
0100
0001
.1co
ntig
1047
78-5
626
2.00
E–15
087
MKK
ILKN
SLIL
LLVM
DIII
VGG
Two
gene
s:A
BCtr
ansp
orte
rA
TP-b
indi
ngp
rote
in, p
ossi
bly
mul
tidru
gef
flux,
cons
erve
dhy
pot
hetic
alp
rote
inA
EDX
0100
0024
.1co
ntig
9c1
6598
-157
351.
00E–
2949
S.m
itis
F039
2N
Z_A
FUO
0100
0001
.1co
ntig
16c2
3087
7-23
0029
2.00
E–15
087
MKK
LLKF
T*IL
LLIM
DIII
IVG
GTw
enty
gene
s:p
erm
ease
,D-A
laD
-Ala
ligas
e,se
rine
reco
mb
inas
e,hy
pot
hetic
alp
rote
in,h
ypot
hetic
alp
rote
inN
Z_A
FUO
0100
0001
.1co
ntig
16c3
2100
0-32
0137
8.00
E–31
50N
Z_A
FUO
0100
0001
.1co
ntig
16c1
1197
38-1
1188
872.
00E–
2749
S.m
itis
SK61
6A
ICR0
1000
024.
1co
ntig
0005
7c5
863-
4991
4.00
E–35
56A
ICR0
1000
047.
1c1
8053
-171
90c1
8053
-171
903.
00E–
2949
S.m
itis
ATC
C62
49A
EEN
0100
0012
.1co
ntig
0003
8c5
0935
6-50
8463
2.00
E–34
50S.
miti
sSK
575
AIC
U01
0000
54.1
cont
ig00
002
8196
1-82
824
2.00
E–33
50M
KKQ
KLLP
LLFL
ILG
GIM
IIVVG
Four
gene
sin
the
opp
osin
gst
rand
AIC
U01
0000
84.1
cont
ig00
008
3479
5-35
658
2.00
E–32
50S.
miti
sb
v.2
SK95
AFU
B010
0004
2.1
cont
ig00
028
8824
3-89
106
2.00
E–33
50A
FUB0
1000
005.
1co
ntig
0000
742
095-
4294
64.
00E–
2749
S.m
itis
SK56
4A
EDU
0100
0025
.1co
ntig
35c1
7203
-163
432.
00E–
3250
MKK
QKL
LSLF
FLLF
EGIII
IVVG
Four
gene
slik
ely
form
ing
anop
eron
inth
eop
pos
ing
stra
ndS.
miti
sSK
597
AED
V010
0004
3.1
cont
ig48
c338
94-3
3031
4.00
E–31
50S.
miti
sSK
1073
AFQ
T010
0004
9.1
ctg1
1296
4535
5518
c527
68-5
1905
2.00
E–30
51A
EDU
0100
0008
.1co
ntig
1818
201-
1906
42.
00E–
3049
S.m
itis
SK57
9A
JJL0
1000
008.
1co
ntig
0002
651
24-5
987
5.00
E–30
49S.
miti
sB6
FN56
8063
.11
c116
5957
-116
5094
8.00
E–29
49S.
miti
sSK
569
AFU
F010
0004
2.1
ctg1
1299
1397
7728
1845
0-19
313
2.00
E–28
49S.
miti
sSP
AR1
0A
LCH
0100
0001
.1G
A06
201.
cont
ig.0
5456
1-55
412
2.00
E–26
49
Stre
ptoc
occu
sgo
rdon
iiS.
gord
onii
Cha
llis
CH
1C
P000
725.
11
9287
23-9
2958
63.
00E–
3350
Stre
ptoc
occu
sor
alis
S.or
alis
SK10
0A
JKP0
1000
019.
1ct
g120
0043
8055
212
1367
-122
230
4.00
E–35
50S.
oral
isSK
10A
JKO
0100
0033
.1ct
g120
0050
2459
0c7
8504
-776
416.
00E–
3449
S.or
alis
SK10
74A
ICT0
1000
005.
1co
ntig
0000
1c7
2651
8-72
5655
8.00
E–33
49S.
oral
isSK
255
AFN
M01
0000
59.1
ctg1
1291
5745
0708
5056
4-51
427
5.00
E–33
49S.
oral
isU
o5FR
7206
02.1
192
7277
-928
140
2.00
E–32
49S.
oral
isSK
304
ALJ
N01
0000
16.1
ctg1
2000
8662
248
4179
49-4
1881
28.
00E–
3249
S.or
alis
SK31
3A
FUU
0100
0003
.1ct
g112
9914
0049
2152
350-
5321
33.
00E–
3149
S.or
alis
SK61
0A
JKQ
0100
0008
.1co
ntig
0000
1c6
9626
-687
632.
00E–
3049
S.or
alis
ATC
C35
037
AD
MV0
1000
020.
1co
ntig
0004
8c8
7429
-869
591.
00E–
1954
Stre
ptoc
occu
ssa
ngui
nis
S.sa
ngui
nis
SK36
CP0
0038
7.1
177
9661
-780
524
1.00
E–33
51S.
sang
uini
sVM
C66
AEV
H01
0000
17.1
cont
ig00
017
c305
82-2
9719
9.00
E–34
51S.
sang
uini
sSK
353
AEW
Y010
0000
5.1
cont
ig00
005
c305
87-2
9724
1.00
E–33
51
(Con
tinue
don
next
pag
e)
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 4
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

TAB
LE1
(Con
tinue
d)
Spec
ies
Acc
essi
onn
o.C
hro
mos
ome
Posi
tion
Eva
lue
Posi
tive
s(%
)Po
ten
tial
ph
erom
one
Dow
nst
ream
ofsh
p
S.sa
ngui
nis
SK16
0A
EXZ0
1000
008.
1co
ntig
0000
831
773-
3263
61.
00E–
3351
AEX
Z010
0000
7.1
cont
ig00
007
c162
493-
1616
245.
00E–
2749
S.sa
ngui
nis
SK72
AEX
V010
0000
5.1
cont
ig00
005
3537
88-3
5465
13.
00E–
3351
S.sa
ngui
nis
ATC
C49
296
AEP
O01
0000
13.1
cont
ig00
013
c125
798-
1249
358.
00E–
3348
S.sa
ngui
nis
SK49
AFF
O01
0000
07.1
cont
ig00
007
6492
0-65
783
1.00
E–32
51S.
sang
uini
sSK
150
AEX
Y010
0002
2.1
cont
ig00
022
c321
62-3
1299
6.00
E–32
51S.
sang
uini
sA
TCC
2966
7A
FQA
0100
0025
.1co
ntig
0002
598
032-
9890
41.
00E–
2851
S.sa
ngui
nis
SK34
0A
FQB0
1000
002.
1co
ntig
0000
298
035-
9890
71.
00E–
2851
S.sa
ngui
nis
SK11
5A
EXW
0100
0005
.1co
ntig
0000
567
2552
-673
439
2.00
E–25
49S.
sang
uini
sSK
355
AFF
N01
0000
13.1
cont
ig00
013
c190
313-
1894
445.
00E–
2547
S.sa
ngui
nis
SK33
0A
FBD
0100
0004
.1co
ntig
0000
420
4150
-205
019
9.00
E–25
48
Stre
ptoc
occu
spa
rasa
ngui
nis
IS.
para
sang
uini
sC
C87
KA
ZJD
0100
0007
.1su
per
cont
1.2.
C7
3347
64-3
3561
22.
00E–
150
87M
KKIL
KNSL
ILLL
VMD
IIIVG
GEl
even
gene
san
nota
ted
ashy
pot
hetic
alp
rote
ins,
inte
rrup
ted
atp
ositi
on2
by
asm
all
hyp
othe
tical
pro
tein
AZJ
D01
0000
01.1
sup
erco
nt1.
1.C
123
8609
-239
484
1.00
E–32
52
S.pa
rasa
ngui
nis
ATC
C15
912
CP0
0284
3.1
119
8169
7-19
8082
22.
00E–
4458
MKK
YYQ
IFLL
LLFD
IIIIIG
LYQ
Six
gene
sin
the
opp
osin
gst
rand
toth
esh
pC
P002
843.
11
2322
31-2
3310
31.
00E–
3151
CP0
0284
3.1
110
9867
0-10
9953
01.
00E–
2650
S.pa
rasa
ngui
nis
F044
9A
JMV0
1000
057.
1co
ntig
0000
839
264-
4012
76.
00E–
4558
AJM
V010
0010
1.1
cont
ig00
001
c141
12-1
3240
5.00
E–32
51A
JMV0
1000
091.
1co
ntig
0001
2c4
670-
3810
4.00
E–27
50S.
para
sang
uini
sA
TCC
903
AEV
E010
0001
1.1
cont
ig00
011
5192
5-52
797
3.00
E–34
52A
EVE0
1000
037.
1co
ntig
0003
741
893-
4275
35.
00E–
2751
S.pa
rasa
ngui
nis
FW21
3C
P003
122.
11
7802
89-7
8116
48.
00E–
3451
S.pa
rasa
ngui
nis
SK23
6A
FUC
0100
0013
.1ct
g112
9914
0035
52c1
9935
4-19
8494
3.00
E–25
49
Stre
ptoc
occu
spa
rasa
ngui
nis
IIS.
para
sang
uini
sII
F040
5A
EKM
0100
0006
.1ct
g112
6926
8313
63c1
5156
-142
842.
00E–
3351
aSt
rain
sb
elon
ging
toth
em
itis
grou
pof
stre
pto
cocc
iav
aila
ble
inth
eex
pan
ded
Hum
anO
ral
Mic
rob
iom
eD
atab
ase
(HO
MD
),ex
clud
ing
S.pn
eum
onia
e.
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 5
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
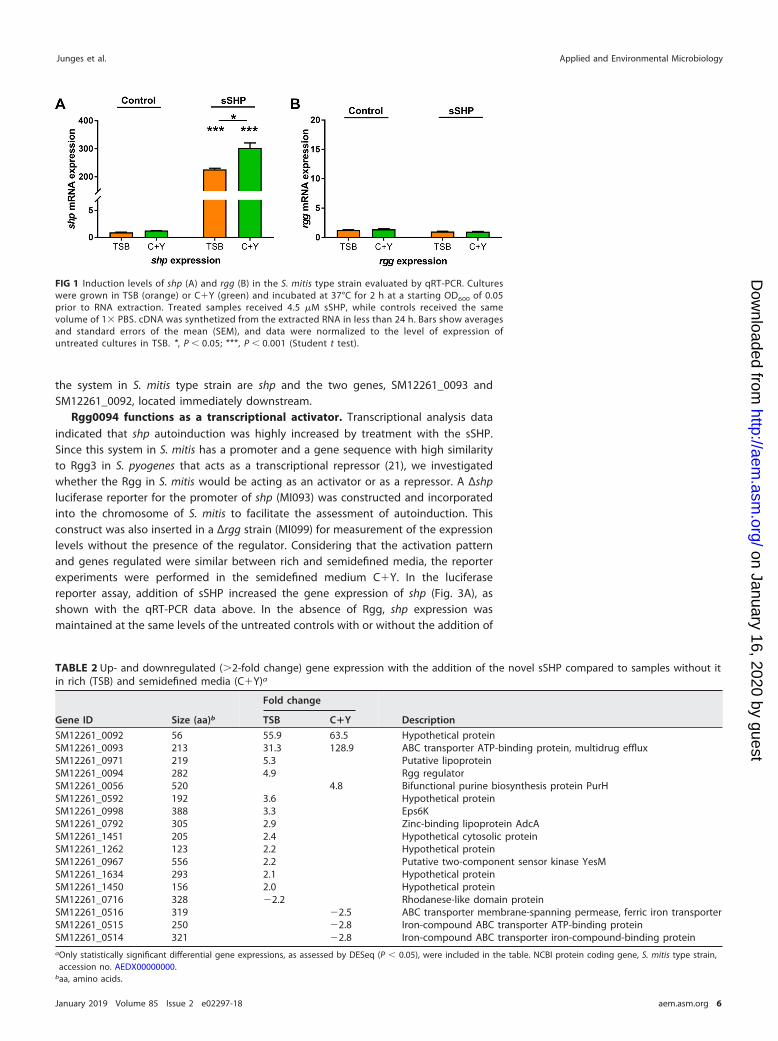
the system in S. mitis type strain are shp and the two genes, SM12261_0093 andSM12261_0092, located immediately downstream.
Rgg0094 functions as a transcriptional activator. Transcriptional analysis dataindicated that shp autoinduction was highly increased by treatment with the sSHP.Since this system in S. mitis has a promoter and a gene sequence with high similarityto Rgg3 in S. pyogenes that acts as a transcriptional repressor (21), we investigatedwhether the Rgg in S. mitis would be acting as an activator or as a repressor. A Δshpluciferase reporter for the promoter of shp (MI093) was constructed and incorporatedinto the chromosome of S. mitis to facilitate the assessment of autoinduction. Thisconstruct was also inserted in a Δrgg strain (MI099) for measurement of the expressionlevels without the presence of the regulator. Considering that the activation patternand genes regulated were similar between rich and semidefined media, the reporterexperiments were performed in the semidefined medium C�Y. In the luciferasereporter assay, addition of sSHP increased the gene expression of shp (Fig. 3A), asshown with the qRT-PCR data above. In the absence of Rgg, shp expression wasmaintained at the same levels of the untreated controls with or without the addition of
FIG 1 Induction levels of shp (A) and rgg (B) in the S. mitis type strain evaluated by qRT-PCR. Cultureswere grown in TSB (orange) or C�Y (green) and incubated at 37°C for 2 h at a starting OD600 of 0.05prior to RNA extraction. Treated samples received 4.5 �M sSHP, while controls received the samevolume of 1� PBS. cDNA was synthetized from the extracted RNA in less than 24 h. Bars show averagesand standard errors of the mean (SEM), and data were normalized to the level of expression ofuntreated cultures in TSB. *, P � 0.05; ***, P � 0.001 (Student t test).
TABLE 2 Up- and downregulated (�2-fold change) gene expression with the addition of the novel sSHP compared to samples without itin rich (TSB) and semidefined media (C�Y)a
Gene ID Size (aa)b
Fold change
DescriptionTSB C�Y
SM12261_0092 56 55.9 63.5 Hypothetical proteinSM12261_0093 213 31.3 128.9 ABC transporter ATP-binding protein, multidrug effluxSM12261_0971 219 5.3 Putative lipoproteinSM12261_0094 282 4.9 Rgg regulatorSM12261_0056 520 4.8 Bifunctional purine biosynthesis protein PurHSM12261_0592 192 3.6 Hypothetical proteinSM12261_0998 388 3.3 Eps6KSM12261_0792 305 2.9 Zinc-binding lipoprotein AdcASM12261_1451 205 2.4 Hypothetical cytosolic proteinSM12261_1262 123 2.2 Hypothetical proteinSM12261_0967 556 2.2 Putative two-component sensor kinase YesMSM12261_1634 293 2.1 Hypothetical proteinSM12261_1450 156 2.0 Hypothetical proteinSM12261_0716 328 �2.2 Rhodanese-like domain proteinSM12261_0516 319 �2.5 ABC transporter membrane-spanning permease, ferric iron transporterSM12261_0515 250 �2.8 Iron-compound ABC transporter ATP-binding proteinSM12261_0514 321 �2.8 Iron-compound ABC transporter iron-compound-binding proteinaOnly statistically significant differential gene expressions, as assessed by DESeq (P � 0.05), were included in the table. NCBI protein coding gene, S. mitis type strain,accession no. AEDX00000000.
baa, amino acids.
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 6
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

sSHP. Altogether, these results indicate that Rgg0094 is an activator protein rather thana repressor. In addition, under these conditions, no other regulator protein seeminglyinteracts with SHP to promote gene activation in the referred promoter site, since theΔrgg mutant showed no differential expression in the locus with or without sSHP.Although the luciferase reporter seemed to be less sensitive than qRT-PCR, the patternof induction was similar.
FIG 2 Transcriptome analysis revealed one gene cluster significantly upregulated by sSHP during growth in rich and semidefined media. (A) Scatter plot ofglobal mRNA expression of S. mitisT with or without the addition of sSHP from S. mitis (SHP0094) (4.5 �M) in both rich medium (TSB, orange) and semidefinedmedium (C�Y, green). Axes show read counts in a log10 scale. (B) In both media, the genes significantly upregulated, SM12261_0093 and SM12261_0092, arelocated immediately downstream of the peptide gene (shp) in the genome. The first pair of rows shows the transcript levels (number of reads – log2) in richmedium TSB after treatment and in the control. The second pair of rows shows the levels in semidefined medium C�Y. All four rows show results in the reversecomplement (RC) strand. Treated and control samples were isolated at 2 h of incubation at 37°C and 5% CO2. The RNA-Seq raw data can be accessed atBioProject accession number PRJNA in the NCBI BioProject database.
FIG 3 Expression of shp promoter reporters in response to synthetic SHP. (A) The synthetic peptidecomplements the Δshp strain, but not the Δrgg and Δopp strains. Luminescence values for shp promoterreporter without or with (�) sSHP0094 (1 �M) at peak expression in 90 min. Precultures at an OD600 of 0.5were washed and resuspended in C�Y. Cultures were then incubated at 37°C, while measuring opticaldensity and luminescence (in RLU) in a plate reader every 30 min up to 5 h. The bars show averages andSEM for samples measured in at least three independent biological experiments. The luminescence resultsof all samples were compared to each other with one-way ANOVA, followed by Tukey’s multiple-comparison test (****, P � 0.0001). (B) To measure the concentration curve response, precultures of theΔshp reporter strain at an OD600 of 0.5 were washed, resuspended in 10 ml of C�Y, and divided into wellswith different concentrations of sSHP0094. The results presented above are reflective of the peak expres-sion in response to sSHP observed at 90 min. For normalization, the control Δshp background values weresubtracted. Each point shows averages and SEM for samples measured in at least three independentbiological experiments.
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 7
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

The interaction of Rgg and the signal peptide SHP has been shown to be intracel-lular and to require the oligopeptide permease (Opp) as an importer of the peptide (19,21, 24). The SHP peptide addressed in this study belongs to the same group as thesystems previously described in S. pyogenes (21), S. agalactiae (24), and S. pneumoniae(28); however, the mature peptide sequence presents one less isoleucine residue thanthe other systems. Thus, it was sought to determine whether Opp would also beresponsible for the import of the peptide. A pshp-luc reporter strain was constructed ina ΔoppD background strain, yielding mutant MI100. As shown in Fig. 3A, without Opp,the sSHP added to cultures had no significant effect on luminescence activity. Thisfinding indicates that Opp is the importer of the SHP in S. mitis as well.
Deletion of rgg does not affect growth under normal or antibiotic stressconditions. The targets for Rgg in S. mitis were shown to only be the shp and adownstream two-gene locus, which is possibly nonfunctional and can trigger a futilecycle. In addition, Rgg systems have been shown to be involved in stress responses(33–35). Thus, we tested whether the presence of Rgg would affect the species abilityto grow in normal and in antibiotic stress conditions. Growth in the presence ofciprofloxacin (DNA-gyrase inhibitor), erythromycin (protein synthesis inhibitor), ampi-cillin (cell wall inhibitor), and nisin (bacterial membrane disruptor) was assessed todetermine the concentration necessary to inhibit the growth of each strain. These drugswere selected since they target different functions in bacteria and can provide a widerassessment of the potential role the system in antibiotic stress response. However, theΔrgg mutant strain showed no differences compared to the type strain control (Fig. 4).To maximize any potential difference, the addition of sSHP was included in the typestrain as well. These results indicate that the deletion of Rgg does not affect S. mitisgrowth under normal or antibiotic stress conditions.
SHP peptides from S. pneumoniae and S. pyogenes interact with Rgg0094 topromote gene transcription. Previous studies have shown that while some Rggregulators promiscuously respond to different SHPs, others are more specific and
FIG 4 The Rgg-regulated transcript showed no effect on growth under normal and antibiotic stress conditions. Strains were inoculatedinto wells containing a range of antibiotic concentrations diluted in rich medium TSB. The plate was incubated overnight at 37°C and 5%CO2. At 20 h, the OD600 was measured in a plate reader. No growth was defined as no increase in OD600 from the initial inoculum(OD600 � 0.1) over the 20 h of the experiment. The synthetic peptides from S. mitis (DIIIVGG) and from S. salivarius (PYFTGCL) wereadded at a final concentration of 1 �M. At least three independent biological experiments were conducted. Each bar represents the meanand SEM of each group.
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 8
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

seemingly respond exclusively to their native signal peptide (29–31). The SHP sequencein S. mitis is unique (DIIIVGG); however, it presents only one different residue in itspredicted mature form compared to other group I peptides (DIIIIVGG or DILIIVGG). Toassess whether the peptides from other streptococcal species could stimulate autoin-duction in S. mitis, the shp reporter strain was used. Initially, to measure the concen-tration of peptide necessary for saturation of the system, the reporter strain wasstimulated with varied concentrations of the S. mitis sSHP. As shown in Fig. 3B,saturated levels of autoinduction are observed for concentrations greater than 500 nM.In addition, concentrations as low as 62.5 nM already showed stimulation of the system.Further, SHPs produced by other streptococci were synthesized and added to culturesof the reporter strain for assessment of the activation potential. Alongside the peptideof S. mitis type strain (DIIIVGG), the following peptides were used: DIIIIVGG from S.pneumoniae and S. pyogenes (21, 28), DILIIVGG from S. agalactiae and S. pyogenes (21,24), ETIIIIGGG from S. mutans (30), EIIIILPFLTNL from S. pneumoniae (20), and PYFTGCLfrom S. salivarius (36). The peptides presenting the greatest similarity (DIIIIVGG andDILIIVGG) to the peptide from S. mitis (DIIIVGG) showed high shp induction (Fig. 5A).This finding is in line with previous cross-communication studies that strongly indicatethe occurrence of cross-communication among streptococcal species through Rgg/SHPsystems (29, 30). As the previous sections of this study have indicated, only one Rgg inS. mitis is responsible for the activations of this locus. As such, to test whether thepeptides from S. pneumoniae, S. agalactiae, and S. pyogenes were interacting with thesame regulator in S. mitis, their activation potential was tested in the Δrgg pshp-lucstrain. None of the SHPs activated autoinduction when Rgg is absent (Fig. 5B). Alto-gether, these findings show that the Rgg/SHP system in S. mitis can be stimulated withthe peptides DIIIIVGG and DILIVGG produced by closely related streptococci. In addi-
FIG 5 Signaling peptides from S. pneumoniae and S. pyogenes stimulate the Rgg/SHP system in the S.mitis type strain. Luminescence expression for S. mitis MI093 grown with or without sSHP (1 �M) wasassessed. (A) Δshp pshp-luc strain (B) Δshp Δrgg pshp-luc strain. Precultures at an OD600 of 0.5 werewashed, resuspended in C�Y, and incubated at 37°C, while measuring the optical density and lumines-cence (in RLU) in a plate reader. The following peptides were added: DIIIVGG (S. mitis), DIIIIVGG (S.pneumoniae and S. pyogenes), DILIIVGG (S. pyogenes and S. agalactiae), ETIIIIGGG (S. mutans), EIIIILPFLTNL(S. pneumoniae), and PYFTGCL (S. salivarius). Each line shows either an untreated culture or a culture withthe addition of one of the peptides. For normalization, the control “no treatment” background valueswere subtracted. The symbols show averages and vertical bars indicate the SEM for samples measuredin at least three independent biological experiments.
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 9
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

tion, it is plausible to conclude that all interactions are coordinated by Rgg0094 in S.mitis type strain CCUG31611 (S. mitisT).
The S. mitis SHP can trigger an increase in surface polysaccharide productionin S. pneumoniae. A recent study shows that the Rgg/SHP system in S. pneumoniae isresponsible for the activation of a cluster of 11 genes that seemingly regulates surfacepolysaccharide production (28). We showed above that the peptide produced by S.pneumoniae could trigger the activation of the system in S. mitis. To test whether thepeptide produced by the S. mitis could stimulate surface polysaccharide production inS. pneumoniae, the D39 derivative unencapsulated strain R36 was cultured and treatedin the presence of the synthetic SHP0094. For assessment of surface polysaccharideproduction, the zone of exclusion of fluorescein isothiocyanate (FITC)-dextran (28,37–39) was measured in the different treatment groups. Treatment with both the S.pneumoniae SHP and the S. mitis SHP significantly increased surface polysaccharideproduction compared to the untreated group (Fig. 6). In addition, as a negative control,treatment with the S. salivarius SHP showed no difference in polysaccharide production.Moreover, it was also observed that the SHP from S. mitis induced the expression of allgenes in the operon similarly to the region induced by the S. pneumoniae SHP (see Fig.S1 in the supplemental material). No effect on the surface polysaccharide expressionwas observed upon treatment of the S. mitis type strain with sSHP (data not shown).These findings show that not only the novel Rgg/SHP system in S. mitis can respond tosignals from other streptococci, but it can also trigger the activation of the system inS. pneumoniae and, more importantly, trigger a phenotypic response characterized byincreased production of surface polysaccharides.
Cross-communication is observed during coculture. The results presented aboveshow that interspecies activation of Rgg-regulated pathways occurs when a syntheticpurified peptide is provided to growing cultures of streptococci. To investigate whethersuch cross-communication occurs under growth in the absence of synthetic peptides,coculture assays were conducted with the S. mitis Δshp strain and the S. mitis type strainor S. pneumoniae D39. First, it was tested whether growth in the presence of mannoseand smaller amounts of glucose would increase the response of the system in S. mitis,as shown in a previous study in S. pyogenes (40). C�Y medium with added mannose(1%) and lower levels of glucose (0.02%) contributed to a slightly higher expressionfrom the shp reporter (Fig. S2). Thus, cocultures were prepared in this medium withequal amounts of each species. Growth and luminescence were monitored over time ina plate reader, and each combination was plated at selected time points to measure themixed bacterial ratio. Cocultures of the S. mitis Δshp luciferase reporter with the S. mitis
FIG 6 The peptide of S. mitis type strain triggers an increased surface polysaccharide production in S.pneumoniae R36. Each dot represents either the light microscopy area or the measurement of theFITC-dextran exclusion area for a single diplococcus. DIIIVGG peptide was synthesized based on thesequence found in S. mitisT. DIIIIVGG was synthesized based on the S. pneumoniae D39 shp sequence.PYFTGCL was synthesized based on the shp sequence of S. salivarius. Each line represents the mean andthe SEM of each group. FITC results from the four samples were compared to one-way ANOVA, followedby Tukey’s multiple-comparison test (ns, not significant; ****, P � 0.0001).
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 10
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

type strain or S. pneumoniae D39 showed activation of the system, which indicates thatthe peptides are indeed produced by both species (Fig. 7). Together, these resultsindicate that cross-communication can readily take place in bacterial communities withmembers of the mitis group of streptococci that share such Rgg/SHP systems.
Absence of Rgg provides a slight competition advantage to S. mitis and S.pneumoniae. Since both S. mitis and S. pneumoniae colonize the human upperrespiratory tract and oral cavity, and Rgg/SHP systems seem to regulate differentbehaviors on the different species and even strains, it is important to understandwhether such cell-to-cell signaling systems can have an impact on competition. Differ-ent combinations of strain pairs, including the S. mitis type strain, S. pneumoniae D39,and their isogenic Δrgg mutants, were inoculated into C�Y and grown for 4 h, afterwhich they were plated for CFU counts. For both species, when Rgg was not present,and the transcriptional target was therefore not activated, the wild type gained a slightadvantage (Fig. 8). This result indicates that Rgg/SHP systems may have an impact oncompetition between the two species.
DISCUSSION
Quorum sensing has been gaining increasing attention due to its ability to coordi-nate microbial behavior. As such, interference with signaling poses as an interestingalternative to disrupt pathological behavior and fight infections. We report here thecharacterization of a cell-to-cell signaling system in the oral commensal S. mitis typestrain that allows for interspecies cross-communication with the pathogen S. pneu-moniae D39. More importantly, this signal can lead to phenotype change in the latter,as observed by an increase in cell surface polysaccharide expression, and may play arole during competition.
In the S. mitis type strain, the main targets of the Rgg/SHP system were identified inthe present study as SM12261_0093, an ABC transporter ATP-binding protein, andSM12261_0092, a conserved hypothetical protein. Previous analyses of ABC transport-ers in S. pneumoniae (41) categorize SM12261_0093 homologues in S. pneumoniae D39,R6, and TIGR4 as “orphan” proteins since they only present the ATPase domain of anABC transporter. A closer analysis of SM12261_0093 homologues in the fully sequencedgenomes of S. pneumoniae revealed the presence of a larger gene coding for the ABCtransporter in the strains JJA, A45, A66, ATCC 700669, INV104, SPN033038, andSPN032672. In these cases, the larger transporter gene also presents a peptidase and apermease domain just upstream of the ATPase domain. In addition, the ABC transporter
FIG 7 Cross-communication is observed during coculture. Bars show relative light units (RLU) per CFU.RLU was measured from a Δshp luciferase reporter strain of S. mitis during coculture with differentspecies or strains, which are listed in the x axis. Strains were added at a ratio of 1:1 in C�Y plus 1%mannose and 0.02% glucose. The OD600 and the RLU were monitored in a plate reader at 37°C at regularintervals. In the first series, sSHP was added at a concentration of 1 �M. At 2, 3, and 4 h, cultures wereplated for the CFU counts. For normalization, the control Δshp background values were subtracted. Theresults are representative of independent biological experiments.
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 11
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

is annotated as a lantibiotic transporter system, and there are other genes in the locusthat are highly similar to modification enzymes and immunity proteins. However, nobacteriocin/lantibiotic prepeptide genes were identified in these referred strains. Re-markably, a previous study (42) has shown that this locus in S. pneumoniae is, in fact,located in an integrative and conjugative element (ICE), representing mobile geneticentities that can be disseminated between bacteria through conjugative transfer. ThisICE-derived genomic island was described as the pneumococcal pathogenicity island 1(PPI-1), which includes virulence genes such as the PezAT toxin-antitoxin (TA) system,the iron uptake locus piaABCD, and others (43–45). Croucher et al. (42) addressed therole of conjugative elements in S. pneumoniae strain ATCC 700669, a member of theserotype 23F ST81 lineage, and showed that the potentially complete lantibiotic locusis present in some strains, while in others only the transporter and the immunityproteins are found (Fig. 9). Moreover, this gene cluster was found to be a two-peptidelantibiotic similar to mersacidin and lichenicidin. As such, it contains genes for twolantibiotic prepeptides, two modification enzymes (LanM), a transporter with a protease
FIG 8 Competitive index values for S. mitisT, S. pneumoniae D39, and its isogenic Δrgg strains. Coculturesof pairs of strains were created by 1:1 inoculation in C�Y and incubated for 4 h at 37°C and 5% CO2 priorto plating. CFU were counted in plain and selective plates to estimate each strain and to calculate theratio just after inoculation and at the end of the incubation period. The ratio in the y axis was calculatedby dividing the final ratio of S. mitis (blue) to S. pneumoniae (red) at the end of incubation by the initialratio at the start of the experiment – the ratio of 1 represents no difference in the competition. CIperformed in C�Y with the addition of 1 �M of sSHP from S. mitis (SHP0094) for activation of the systems.ANOVA, followed by Tukey’s post hoc test, was performed (*, P � 0.05; ***, P � 0.001). At least threeindependent biological experiments were conducted.
FIG 9 A segment of the locus downstream of shp is conserved in S. mitis compared to S. pneumoniae. A black background patternshows the conserved areas in S. pneumoniae and S. mitis, whereas red areas highlight segments involved in lantibiotic production andimmunity that were likely lost in S. mitis type strain. “shp*” indicates an interruption in the mature sequence of the peptide (DIIII).
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 12
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

domain (LanT), and three genes coding for immunity proteins (LanEFG) (46). Similarlantibiotics have been shown be produced by other species and can inhibit a widerange of gram-positive organisms, including methicillin-resistant Staphylococcus aureus(47). The presence of putative relaxases, integrases, mobilization and plasmid replica-tion genes, and ribosomal protein ORFs in the vicinity of the Rgg/SHP locus in S. mitisalso supports the presence of an ICE in this locus (48).
A high variation in the PPI-1 has been observed across different serotypes andstrains (44, 49). In particular, Harvey et al. compared serotype 1 nonvirulent and virulentisolates to identify factors that could alter the outcome of infection (49). Accordingly,in the PPI-1 region, the locus immediately downstream of the Rgg regulator is part ofan accessory region that presents high variability across different strains. More impor-tantly, the partial lantibiotic gene cluster found in strain ATCC 700669 was highlightedas a replacement for a region associated with high virulence in other isolates (49). It isimportant to note, however, that while strains such as ATCC 700669 present a partiallantibiotic locus, the genes encoding immunity are maintained, which could be advan-tageous when competing for resources, in particular against strains producing thereferred lantibiotic (42, 49). However, addressing the question of whether the fulllantibiotic locus found in strain SP23-BS72 provides a competitive advantage againstsusceptible strains and species can lead to a better understanding of the ecologicalrelationship among different strains of the pathogen. In other members of the mitisgroup of streptococci, as shown in this study, the substantial variation of the Rgg locusand putative regulons is also observed, particularly in S. mitis. The current hypothesisindicates that this genetic island originated as an ICE insertion that has either degen-erated or recombined in many strains of S. pneumoniae (42), which may also hold truefor S. mitis given that only a section of the gene cluster is present in the oralcommensal. Interestingly, the mature shp gene is present in S. pneumoniae SP23-BS72and S. mitis type strain, but it is interrupted at position 6 (DIIII) in S. pneumoniae ATCC700669 (28). This indicates that both S. pneumoniae ATCC 700669 and the S. mitis typestrain have likely lost part of the gene cluster in independent events. In the S.pneumoniae strain SP23-BS72, the rgg gene is 95.7% identical to strain D39 (SPD_0939).Further, the locus between rgg and shp containing the promoter sites is also remarkablysimilar, showing 88.5% identity. This includes the sequences of SHP, which in its full23-amino-acid sequence presents only one divergent residue at position 12, and thepredicted mature 8-amino-acid sequence is identical between SP23-BS72 and D39.Fundamentally, the identification of the target locus in an ICE sheds light into thereason for lack of conservation of the Rgg/SHP system in members of the mitis groupcompared to other streptococci, such as S. agalactiae and S. pyogenes (21, 24).
Previous studies show cross-communication between streptococcal species throughRgg/SHP systems (29, 30). Bidirectional signaling is found to activate promoter tran-scription in S. pyogenes, S. agalactiae, and S. dysgalactiae. Further, cross-communicationregulates biofilm formation in S. pyogenes (21, 29). Remarkably, interaction is alsoobserved between different groups of Rgg/SHP systems as a group II reporter systemin S. mutans UA159 responded to both its peptide ETIIIIGGG and the DIIIIVGG peptide,found in the genomes of S. pyogenes and S. pneumoniae D39 (30). In this study, it wasfound that the peptides DIIIIVGG and DILIIVGG from S. pneumoniae, S. agalactiae, andS. pyogenes stimulate the activation of the system in S. mitis, while no response topeptides from other groups was observed. This is in line with the findings fromFleuchot et al. that showed cross-communication response in the S. mutans system,which produces ETIIIIGGG, but not in S. agalactiae, which produces DILIIVGG (30).Moreover, this study showed data for cross-communication in the presence of thesynthetic peptide and during coculture, which strongly suggests that both S. mitisT andS. pneumoniae D39 produce the peptide in an active form. The finding that not allstrains of S. pneumoniae and S. mitis present the complete peptide gene suggests thatcross-communication may play a role in coordinating transcriptional responses in abacterial community.
The direct target of the complex Rgg-SHP in S. mitis type strain was found to be the
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 13
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

two genes immediately downstream of shp. Not all genes that are putatively requiredfor this potential lantibiotic-producing cluster are present in the genome of S. mitis typestrain, which indicates it might not be functional. However, despite ATPases beingoften encoded together with their coupled transporters, it is suggested that an“orphan” ATPase could energize multiple sugar uptake ABC transporters in S. pneu-moniae (50). As such, while this study found no evident effect of the system on thestress response and biology of S. mitisT in the presence of ciprofloxacin, erythromycin,ampicillin, or nisin, the possibility of this gene being involved in other transportpathways should not be ruled out. In addition, through unknown mechanisms, thepartial lantibiotic biosynthesis locus in S. pneumoniae ATCC 700669 and others isassociated with nonvirulent behavior (49), suggesting that this locus might indeed beinvolved in other pathways. Regardless, even if the system in S. mitisT may only presenta role in self-regulation, the production of SHP in itself could have implications incross-communication and potentially during competition.
Since Rgg/SHP systems have been shown to coordinate different genes and func-tions in a variety of streptococcal species, it is puzzling that these signal peptidescross-communicate among different species and strains to coordinate different behav-iors. These systems are likely part of complex networks that rearrange and recombinemore often compared to other parts of the genome, potentially adjusting and adaptingto selective environmental pressures. To place bacteriocin and lantibiotic productionunder the control of quorum-sensing systems is not unusual in streptococci, as exem-plified by the widely studied competence system in S. pneumoniae that relies on a tightconnection between the stimulation of competence and bacteriocin production (51,52). In addition, other lantibiotic biosynthesis gene clusters have been shown to beregulated by quorum sensing both in S. pneumoniae with the TprA/PhrA system (27, 53)and in other bacteria, which includes the lantibiotics nisin by Lactococcus lactis andsubtilin by Bacillus subtilis (54). Interestingly, S. mitis type strain presents the TprA gene,but the PhrA and the lantibiotic biosynthesis cluster downstream are not conserved,which could indicate that the fitness cost for maintaining the lantibiotic-producingmachinery is not optimal. To coordinate the expression of such compounds by cell densitymight be useful since it increases the chance of horizontal gene transfer in a givenpopulation (42). Overall, quorum-sensing signals often regulate mechanisms that providefitness advantages in response to environmental conditions such as nutrient availability andcell density (8, 12, 55). S. mitis has likely evolved with diverse mechanisms that allow it tosuccessfully colonize hostile environments, such as the oral cavity. Fundamentally, theunderstanding of the complex pathways coordinated by cell-to-cell communication mightprovide important insights regarding the host-microbiome relationship.
MATERIALS AND METHODSBacterial strains and media. All used strains and primers are listed in Table 3. Strains were cultured
in tryptone soya broth (TSB; Oxoid, Hampshire, UK) or C�Y (56). For storage, media were supplementedwith 30% glycerol. Agar plates were prepared with blood agar base 2 (Oxoid, Hampshire, England)supplemented with defibrinated sheep blood (5%) (TCS Biosciences, Ltd., Buckingham, United Kingdom).For mutant selection and recovery, antibiotics were used at the following concentrations: kanamycin,500 �g/ml; erythromycin, 10 �g/ml; and spectinomycin, 500 �g/ml. Preculture stocks were prepared asdescribed previously (57). All peptides utilized in the assays were obtained from GenScript (GenScript,Piscataway, NJ) as custom synthetic peptides and stored as sterile 10 mM solutions at �20°C (Table 3).
qRT-PCR. For qRT-PCR, precultures were diluted 1:10 and were allowed to grow in TSB or C�Y underthe specific conditions of each experiment. At specific time points, cells were harvested at 8,000 � g and4°C for 8 min. RNA was extracted with a High-Pure RNA isolation kit (Roche), and cDNA was generatedusing a first-strand cDNA synthesis kit (Thermo Scientific; Fermentas). The primers used are listed in Table4. For qRT-PCR, Maxima SYBR Green/ROX qPCR Master Mix (Thermo Scientific) was utilized according tothe manufacturer’s protocol.
RNA sequencing. Cultures of S. mitis type strain were grown in TSB or C�Y at 37°C and 5% CO2 for20 min. The culture was then divided in two, and sSHP (DIIIVGG) was added to half of the sample.Cultures were grown at 37°C and 5% CO2 for 2 h, and pellets were harvested and stored at �80°C. RNAextraction, enrichment, cDNA synthesis and labeling were performed as recently described (28, 58).Sequencing was performed with Illumina HiSeq at the Norwegian Sequencing Centre (http://www.sequencing.uio.no). Analyses and mapping followed previously described protocols (58, 59). Briefly,from the FASTQ file generated from each samples, Bowtie was used to map the nucleotide reads to
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 14
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

the genome of S. mitis type strain under GenBank accession number NZ_AEDX00000000.1. To mapeach read to the correct strand, SAMtools software was used, and visualization was graphed in theJBrowse viewer. Differential expression analyses in untreated and treated samples were determinedbased on the read profiles using a Perl script. The DESeq package from Bioconductor was used to assessthe differential levels when comparing samples. Significance was set at P � 0.05. The transcriptomesequencing (RNA-Seq) raw data can be accessed at BioProject accession number PRJNA488478 in theNCBI BioProject database.
Contruction of mutants. Primers used for construction of mutants are listed in Table 5. MutantsMI070 and MI075 were constructed by a PCR mutagenesis method described previously (60). Briefly, theflanking regions of the knockout gene were amplified and ligated with a kanamycin resistance cassette.
TABLE 3 List of strains and peptides used
Strain or peptide Description or sequencea
Source orreferenceb
StrainsCCUG31611T Streptococcus mitis type strain CCUGNCTC7466 Streptococcus pneumoniae D39 NCTCNCTC10319 Streptococcus pneumoniae R36A NCTCMI048 CCUG31611T, but pldh-luc; Spcr This studyMI070 CCUG31611T, but Δrgg::Kan; Kanr This studyMI075 CCUG31611T, but ΔSM12261_0093::Kan; Kanr This studyMI083 CCUG31611T, but pshp-luc; Spcr This studyMI091 CCUG31611T, but Δshp 62MI093 MI091, but pshp-luc; Spcr This studyMI097 CCUG31611T, but ΔoppD This studyMI099 MI093, but Δrgg; Spcr Kanr This studyMI100 MI093, but ΔoppD; Spcr This studySP068 NCTC7466, but Δrgg 28
PeptidesS. mitis NH2-DIIIVGG-OH GenScriptS. pneumoniae and S. pyogenes NH2-DIIIIVGG-OH GenScriptS. pyogenes and S. agalactiae NH2-DILIIVGG-OH GenScriptS. pneumoniae NH2-EIIIILPFLTNL-OH GenScriptS. mutans NH2-ETIIIIGGG-OH GenScriptS. salivarius NH2-PYFTGCL-OH GenScript
aKanr, kanamycin resistance; Spcr, spectinomycin resistance.bCCUG, Culture Collection, University of Gothenburg; NCTC, National Collection of Type Cultures (CentralPublic Health Laboratory, London, England).
TABLE 4 Primers used for qRT-PCR
Primer Sequence (5=–3=) Gene
FP369 GCCGTTCGTGGTATGAGTCG S. mitis gyrAFP370 GGTCGCAACTGTGCGCTTAC S. mitis gyrAFP1010 TTTCATCGTGTCCTCCAACCA S. mitis shpFP1011 TGTCGGTGGATAAAATGTTTAGGTG S. mitis shpFP760 TTGCACCAAGTTAGCGAATG S. mitis rggFP761 GATTTGAACGGGGAGAATCA S. mitis rggFP1012 TCCTGTGGAACTACTCCTAAATTTTGAC SM12261_0093FP1013 CAACCGGGGTCAGGAAAAAG SM12261_0093FP1918 GGGGAAAACTCTTTGCACTGG SM12261_0516FP1919 GGCAGTCAGGAGAATGCTAATGG SM12261_0516FP1920 AGGATATTTTGCAAGGCATTTCG SM12261_0515FP1921 TCGATTTTCCAGCACCATTTG SM12261_0515FP1922 CTCACTGCCCTAGTAGCCAGCTT SM12261_0514FP1923 GAGCAGAGCTACTTGACTCCGTCTT SM12261_0514FP367 ACCGAGGCACGTATGAGCAA S. pneumoniae gyrAFP368 AGACCAAGGGTTCCCGTTCA S. pneumoniae gyrAFP1014 ACCGATTTTGTTTTTGGTAATGGA S. pneumoniae shpFP1015 TGTACACCTAAGCATTTTATCCACCA S. pneumoniae shpFP1697 CATTTAGTTTGGGAACACGAGGAA SPD_0944FP1698 ATTCCAACCGCCTTCTCCAAT SPD_0944FP1699 TGGATGGTCTTGGGGAGTTTC SPD_0947FP1700 AAAAGCCCACACTGCAAAAGG SPD_0947FP1701 TGCCAGTTGGGTCTGGATTTT SPD_0950FP1702 CATACCTCTTCAATAGCGTGCTGAAT SPD_0950
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 15
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

For the construction of a luciferase reporter (61), the promoter region of shp was amplified with theprimers FP853 and FP854. After restriction with NheI and BamHI, the amplicon was ligated to pFW5-luc(spectinomycin resistance [Spcr]) and cloned into Escherichia coli as previously described (61, 62). Further,the correct plasmid was purified and transformed into the S. mitisT, being incorporated in the chromo-some by single crossover recombination. To avoid transcriptional changes in the tightly regulated locusof the Rgg/SHP system, the pshp-luc segment was amplified and inserted into an area of the S. mitisgenome that showed no transcription and no predicted ORF, upstream of genes SM12261_0592 andSM12261_0593. In addition, we observed no phenotype for any of the mutants with this construct. Theshp gene was deleted to minimize the background expression levels of the system, resulting in mutant
TABLE 5 Primers used for mutant construction
Primer Sequence (5=–3=)a Location Template Description
FP001 AGGCGCGCCGTTTGATTTTTAATG Kanr gene pSF151 Kan markerFP068 AGGCCGGCCTAGGTACTAAAACAATTCATCCAGTA Kanr gene pSF151 Kan markerFP817 TGCCAGTGCTATCCATTGTC Flanking rgg Type strain Construction of mutant
MI070 (Kanr)FP818 AGGCGCGCCTGATTCTCCCCGTTCAAATC Flanking rgg Type strain Construction of mutant
MI070 (Kanr)FP819 AGGCCGGCCAAAAGCACAGTACCAAGAGCA Flanking rgg Type strain Construction of mutant
MI070 (Kanr)FP820 CGTTCGTGAATTTGAAGTGG Flanking rgg Type strain Construction of mutant
MI070 (Kanr)FP835 GCAATTGATTGCCAATTGTGT Flanking SM12261_0093 Type strain Construction of mutant
MI075 (Kanr)FP836 AGGCGCGCCGGTTGACCGACTAGTGCAA Flanking SM12261_0093 Type strain Construction of mutant
MI075 (Kanr)FP83a AGGCCGGCCAATGGCTCTTGGTGGAGAGT Flanking SM12261_0093 Type strain Construction of mutant
MI075 (Kanr)FP834 TGCTCAAGGCGTTATCTGTG Flanking SM12261_0093 Type strain Construction of mutant
MI075 (Kanr)FP853 ATACAAGCTAGCTTCGCAGGATTGCACTTAAA Flanking shp Type strain Construction of MI083 (Spcr)FP854 ACGCAGGATCCTCTTCATATTTAGACTTCCTTTCA Flanking shp Type strain Construction of MI083 (Spcr)FP1262 ATTGATGATGTCCCCCAAAA Flanking pshp-luc SM12261_0592
and SM12261_0593Type strain Construction of MI093 (Spcr)
FP1263 TCATTCTAATTGGTAAGCGCGTATTAAAGAAGAAAGTTAAT Flanking pshp-luc SM12261_0592and SM12261_0593
Type strain Construction of MI093 (Spcr)
FP1266 GTTGGGAAAACGGGAAAATTTAGACAAGATAAAAAAACCTC Flanking pshp-luc SM12261_0592and SM12261_0593
Type strain Construction of MI093 (Spcr)
FP1267 AAAAACGGCACGAAAACATC Flanking pshp-luc SM12261_0592and SM12261_0593
Type strain Construction of MI093 (Spcr)
FP1264 ATTAACTTTCTTCTTTAATACGCGCTTACCAATTAGAATGA Flanking pshp-luc SM12261_0592and SM12261_0593
MI083 Construction of MI093 (Spcr)
FP1265 GAGGTTTTTTTATCTTGTCTAAATTTTCCCGTTTTCCCAAC Flanking pshp-luc SM12261_0592and SM12261_0593
MI083 Construction of MI093 (Spcr)
FP1268 TGTCAATCGTAAACGCACCT Nested PCR AmpliconFP1269 GAGAATGCGGCTCTTCATTT Nested PCR AmpliconFP1408 GATGGAACCCGGACTATCAA Flanking oppD Type strain Construction of MI097
(markerless)FP1409 TCATGGAGATTTGCAATCACACGCGAGCAGTCAAAATTACA Flanking oppD Type strain Construction of MI097
(markerless)FP1410 TGTAATTTTGACTGCTCGCGTGTGATTGCAAATCTCCATGA Flanking oppD Type strain Construction of MI097
(markerless)FP1411 TTGGCTTTTGTTTGCATTTG Flanking oppD Type strain Construction of MI097
(markerless)FP1412 GCACAAGCTTGGTTGACTGA Nested PCR Amplicon Construction of MI097
(markerless)FP1413 TCATCAAACTCTCGATGATCTCTC Nested PCR Amplicon Construction of MI097
(markerless)FP1414 CCAATTACCGTGCCAAGTTT Screening Mutant Construction of MI097
(markerless)FP1415 TGATGGCACGACCAATAGTT Screening Mutant Construction of MI097
(markerless)FP1416 TTGGGATTCCAGATGCAGAT Screening Mutant Construction of MI097
(markerless)FP1417 CAACAGTCGCACGTTCATTC Screening Mutant Construction of MI097
(markerless)aUnderlined letters represent restriction sites: AscI, AGGCGCGCC; FseI, AGGCCGGCC; NheI, GCTAGC; BamHI, GGATCC.
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 16
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

MI093. The construction of the markerless deletion of shp (MI091) has been recently described (62).MI097 was constructed similarly. Mutants MI099 and MI100 were obtained by natural transformation ofMI093 with amplicons constructed from MI070 and MI097.
Reporter assays. Precultures of the strains were diluted 1:10 in TSB or C�Y in the presence orabsence of different concentrations of sSHP. Aliquots of 200 �l were then distributed among the wellsof 96-well microtiter plates (Nunc Thermo Scientific). Blanks containing pure medium were used. A 10-�lvolume of a 1 mM D-luciferin solution (Synchem, Felsberg-Altenberg, Germany) was added to each well,and cultures were incubated at 37°C. For cross-communication experiments, strains were added 1:1 inC�Y with 1% mannose and 0.02% glucose, followed by incubation for 5 h at 37°C in a plate reader. Atthe selected time points, cultures were plated for CFU count. Relative luminescence units (RLU), and theoptical densities at 600 nm (OD600s) were measured at various time intervals during growth in amicroplate reader (Synergy HT; BioTek, Winooski, VT). The luminescence sensitivity was set to high, andfor data analysis the background values were subtracted.
Surface polysaccharide assessment. The degree of surface polysaccharide production was deter-mined by measuring the zone of exclusion with FITC-dextran (2,000 kDa; Sigma) as described previously(28, 37, 39).
Growth under normal and antibiotic stress conditions. Growth under normal and antibiotic stressassays were performed in 96-well plates with 2-fold serial dilutions of antibiotics: ciprofloxacin, 1 to16 �g/ml; ampicillin, 0.05 to 0.8 �g/ml; erythromycin, 0.05 to 0.8 �g/ml; and nisin, 12.5 to 200 �g/ml. Allexperiments included a control without antibiotics in addition to a blank medium control. Bacterialcultures were prepared by inoculating colonies from an overnight agar plate in TSB until they reachedan OD600 of �0.1. The bacterial culture of the type strain was divided into three parts: the first part withsSHP added, second with a negative-control peptide, and the third part received no treatment. TheMI070 culture was not supplemented with any peptides. The bacterial culture (2� concentration) wasaliquoted into the 96-well plate and mixed with the antibiotic solution (2�). At least three independentbiological experiments were performed. The OD600 was measured at the start of the experiment; theplate was covered with a Top Seal lid to avoid evaporation, and the OD600 was measured again at 20 h.The plate was incubated in 37°C and 5% CO2. Inhibition of growth was defined as no increase in theOD600 from the starting point at 0.1 over the 20 h of the experiment.
CI. Competitive index (CI) was conducted as described previously (28, 63). Briefly, 5 � 106 CFU of eachstrain was added in a 50 �l volume to a total of 1 ml of C�Y medium that was then incubated for 4 hat 37°C and 5% CO2. After inoculation and at the end of incubation, the cultures were plated onto regularand selective media for calculation of the ratio. Different combinations of two competitors from the S.mitis type strain, S. pneumoniae D39, and their isogenic Δrgg mutants were assessed as described in thefigure legends. To allow for one strain to be detectable in selective media, MI048, a pldh-luc reporterstrain containing an Spcr cassette, was used in place of the S. mitis type strain. The CI was calculated bydividing the outcome ratio by the inoculum ratio.
Statistical analysis. To compare two groups, a Student t test was used. For three or more groups,one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test was used. Significance was setat a P value of �0.05. Details of the statistical analyses conducted with the transcriptomic data areavailable in the RNA sequencing section of Materials and Methods.
SUPPLEMENTAL MATERIALSupplemental material for this article may be found at https://doi.org/10.1128/AEM
.02297-18.SUPPLEMENTAL FILE 1, PDF file, 0.1 MB.
ACKNOWLEDGMENTSThis study was partially funded by Norwegian Research Council grant 241011, by the
Norwegian Surveillance System for Antibiotic Resistance in Microbes (NORM), by theNansen funds for research, and by the Faculty of Dentistry at the University of Oslo.
REFERENCES1. Kawamura Y, Hou XG, Sultana F, Liu S, Yamamoto H, Ezaki T. 1995.
Transfer of Streptococcus adjacens and Streptococcus defectivus toAbiotrophia gen. nov. as Abiotrophia adiacens comb. nov. and Abiotro-phia defectiva comb. nov., respectively. Int J Syst Bacteriol 45:798 – 803.https://doi.org/10.1099/00207713-45-4-798.
2. Bek-Thomsen M, Tettelin H, Hance I, Nelson KE, Kilian M. 2008.Population diversity and dynamics of Streptococcus mitis, Streptococ-cus oralis, and Streptococcus infantis in the upper respiratory tracts ofadults, determined by a nonculture strategy. Infect Immun 76:1889 –1896. https://doi.org/10.1128/IAI.01511-07.
3. Mitchell J. 2011. Streptococcus mitis: walking the line between commen-salism and pathogenesis. Mol Oral Microbiol 26:89 –98. https://doi.org/10.1111/j.2041-1014.2010.00601.x.
4. Kilian M, Riley DR, Jensen A, Bruggemann H, Tettelin H. 2014. Parallel
evolution of Streptococcus pneumoniae and Streptococcus mitis to patho-genic and mutualistic lifestyles. mBio 5(4):e01490-14. https://doi.org/10.1128/mBio.01490-14.
5. Kilian M, Poulsen K, Blomqvist T, Håvarstein LS, Bek-Thomsen M, TettelinH, Sørensen UBS. 2008. Evolution of Streptococcus pneumoniae and itsclose commensal relatives. PLoS One 3:e2683. https://doi.org/10.1371/journal.pone.0002683.
6. Casadevall A, Pirofski LA. 2018. What is a host? Attributes of individualsusceptibility. Infect Immun 86:e00636-17. https://doi.org/10.1128/IAI.00636-17.
7. Kadioglu A, Weiser JN, Paton JC, Andrew PW. 2008. The role ofStreptococcus pneumoniae virulence factors in host respiratory colo-nization and disease. Nat Rev Microbiol 6:288 –301. https://doi.org/10.1038/nrmicro1871.
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 17
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

8. Papenfort K, Bassler BL. 2016. Quorum sensing signal-response systemsin Gram-negative bacteria. Nat Rev Microbiol 14:576 –588. https://doi.org/10.1038/nrmicro.2016.89.
9. LaSarre B, Federle MJ. 2013. Exploiting quorum sensing to confusebacterial pathogens. Microbiol Mol Biol Rev 77:73–111. https://doi.org/10.1128/MMBR.00046-12.
10. Morrison DA. 1997. Streptococcal competence for genetic transformation: reg-ulation by peptide pheromones. Microb Drug Resist 3:27–37. https://doi.org/10.1089/mdr.1997.3.27.
11. Cook LC, Federle MJ. 2014. Peptide pheromone signaling in Streptococ-cus and Enterococcus. FEMS Microbiol Rev 38:473– 492. https://doi.org/10.1111/1574-6976.12046.
12. Pérez-Pascual D, Monnet V, Gardan R. 2016. Bacterial cell-cell commu-nication in the host via RRNPP peptide-binding regulators. Front Micro-biol 7:706. https://doi.org/10.3389/fmicb.2016.00706.
13. Defoirdt T. 2017. Quorum-sensing systems as targets for antivirulencetherapy. Trends Microbiol https://doi.org/10.1016/j.tim.2017.10.005.
14. Sulavik MC, Tardif G, Clewell DB. 1992. Identification of a gene, rgg,which regulates expression of glucosyltransferase and influences theSpp phenotype of Streptococcus gordonii Challis. J Bacteriol 174:3577–3586. https://doi.org/10.1128/jb.174.11.3577-3586.1992.
15. Kreikemeyer B, McIver KS, Podbielski A. 2003. Virulence factor regulationand regulatory networks in Streptococcus pyogenes and their impact onpathogen-host interactions. Trends Microbiol 11:224 –232. https://doi.org/10.1016/S0966-842X(03)00098-2.
16. Chaussee MS, Ajdic D, Ferretti JJ. 1999. The rgg gene of Streptococcuspyogenes NZ131 positively influences extracellular SPE B production.Infect Immun 67:1715–1722.
17. Qi F, Chen P, Caufield PW. 1999. Functional analyses of the promoters inthe lantibiotic mutacin II biosynthetic locus in Streptococcus mutans.Appl Environ Microbiol 65:652– 658.
18. Sanders JW, Leenhouts K, Burghoorn J, Brands JR, Venema G, Kok J.1998. A chloride-inducible acid resistance mechanism in Lactococcuslactis and its regulation. Mol Microbiol 27:299 –310. https://doi.org/10.1046/j.1365-2958.1998.00676.x.
19. Ibrahim M, Nicolas P, Bessieres P, Bolotin A, Monnet V, Gardan R. 2007. Agenome-wide survey of short coding sequences in streptococci. Microbiol-ogy 153:3631–3644. https://doi.org/10.1099/mic.0.2007/006205-0.
20. Fleuchot B, Gitton C, Guillot A, Vidic J, Nicolas P, Besset C, FontaineL, Hols P, Leblond-Bourget N, Monnet V, Gardan R. 2011. Rgg proteinsassociated with internalized small hydrophobic peptides: a newquorum-sensing mechanism in streptococci. Mol Microbiol 80:1102–1119. https://doi.org/10.1111/j.1365-2958.2011.07633.x.
21. Chang JC, LaSarre B, Jimenez JC, Aggarwal C, Federle MJ. 2011. Twogroup A streptococcal peptide pheromones act through opposing Rggregulators to control biofilm development. PLoS Pathog 7:e1002190.https://doi.org/10.1371/journal.ppat.1002190.
22. Lyon WR, Gibson CM, Caparon MG. 1998. A role for trigger factor and anrgg-like regulator in the transcription, secretion and processing of thecysteine proteinase of Streptococcus pyogenes. EMBO J 17:6263– 6275.https://doi.org/10.1093/emboj/17.21.6263.
23. Samen UM, Eikmanns BJ, Reinscheid DJ. 2006. The transcriptional regu-lator RovS controls the attachment of Streptococcus agalactiae to humanepithelial cells and the expression of virulence genes. Infect Immun74:5625–5635. https://doi.org/10.1128/IAI.00667-06.
24. Pérez-Pascual D, Gaudu P, Fleuchot B, Besset C, Rosinski-Chupin I, Guillot A,Monnet V, Gardan R. 2015. RovS and its associated signaling peptide forma cell-to-cell communication system required for Streptococcus agalactiaepathogenesis. mBio 6(1):e02306-14. https://doi.org/10.1128/mBio.02306-14.
25. Zutkis AA, Anbalagan S, Chaussee MS, Dmitriev AV. 2014. Inactivationof the Rgg2 transcriptional regulator ablates the virulence of Strep-tococcus pyogenes. PLoS One 9:e114784. https://doi.org/10.1371/journal.pone.0114784.
26. Fontaine L, Goffin P, Dubout H, Delplace B, Baulard A, Lecat-Guillet N,Chambellon E, Gardan R, Hols P. 2013. Mechanism of competenceactivation by the ComRS signaling system in streptococci. Mol Microbiol87:1113–1132. https://doi.org/10.1111/mmi.12157.
27. Hoover SE, Perez AJ, Tsui HC, Sinha D, Smiley DL, DiMarchi RD, WinklerME, Lazazzera BA. 2015. A new quorum-sensing system (TprA/PhrA) forStreptococcus pneumoniae D39 that regulates a lantibiotic biosynthesisgene cluster. Mol Microbiol 97:229 –243. https://doi.org/10.1111/mmi.13029.
28. Junges R, Salvadori G, Shekhar S, Åmdal HA, Periselneris JN, Chen T,Brown JS, Petersen FC. 2017. A quorum-sensing system that regulates
Streptococcus pneumoniae biofilm formation and surface polysaccharideproduction. mSphere 2:e00324-17. https://doi.org/10.1128/mSphere.00324-17.
29. Cook LC, LaSarre B, Federle MJ. 2013. Interspecies communicationamong commensal and pathogenic streptococci. mBio 4(4):e00382-13.https://doi.org/10.1128/mBio.00382-13.
30. Fleuchot B, Guillot A, Mezange C, Besset C, Chambellon E, Monnet V,Gardan R. 2013. Rgg-associated SHP signaling peptides mediate cross-talk in streptococci. PLoS One 8:e66042. https://doi.org/10.1371/journal.pone.0066042.
31. Shanker E, Morrison DA, Talagas A, Nessler S, Federle MJ, Prehna G. 2016.Pheromone recognition and selectivity by ComR proteins among Strep-tococcus species. PLoS Pathog 12:e1005979. https://doi.org/10.1371/journal.ppat.1005979.
32. Chen T, Yu WH, Izard J, Baranova OV, Lakshmanan A, Dewhirst FE. 2010.The Human Oral Microbiome Database: a web accessible resource forinvestigating oral microbe taxonomic and genomic information. Data-base (Oxford) 2010:baq013. https://doi.org/10.1093/database/baq013.
33. Chaussee MA, Callegari EA, Chaussee MS. 2004. Rgg regulates growthphase-dependent expression of proteins associated with secondary me-tabolism and stress in Streptococcus pyogenes. J Bacteriol 186:7091–7099. https://doi.org/10.1128/JB.186.21.7091-7099.2004.
34. Dmitriev AV, McDowell EJ, Kappeler KV, Chaussee MA, Rieck LD,Chaussee MS. 2006. The Rgg regulator of Streptococcus pyogenes influ-ences utilization of nonglucose carbohydrates, prophage induction, andexpression of the NAD-glycohydrolase virulence operon. J Bacteriol188:7230 –7241. https://doi.org/10.1128/JB.00877-06.
35. Pulliainen AT, Hytonen J, Haataja S, Finne J. 2008. Deficiency of the Rggregulator promotes H2O2 resistance, AhpCF-mediated H2O2 decomposi-tion, and virulence in Streptococcus pyogenes. J Bacteriol 190:3225–3235.https://doi.org/10.1128/JB.01843-07.
36. Fontaine L, Boutry C, de Frahan MH, Delplace B, Fremaux C, Horvath P,Boyaval P, Hols P. 2010. A novel pheromone quorum-sensing systemcontrols the development of natural competence in Streptococcus ther-mophilus and Streptococcus salivarius. J Bacteriol 192:1444 –1454. https://doi.org/10.1128/JB.01251-09.
37. Gates MA, Thorkildson P, Kozel TR. 2004. Molecular architecture of theCryptococcus neoformans capsule. Mol Microbiol 52:13–24. https://doi.org/10.1111/j.1365-2958.2003.03957.x.
38. Hathaway LJ, Brugger SD, Morand B, Bangert M, Rotzetter JU, Hauser C,Graber WA, Gore S, Kadioglu A, Muhlemann K. 2012. Capsule type ofStreptococcus pneumoniae determines growth phenotype. PLoS Pathog8:e1002574. https://doi.org/10.1371/journal.ppat.1002574.
39. Weinberger DM, Trzcinski K, Lu Y-J, Bogaert D, Brandes A, Galagan J,Anderson PW, Malley R, Lipsitch M. 2009. Pneumococcal capsular poly-saccharide structure predicts serotype prevalence. PLoS Pathog5:e1000476. https://doi.org/10.1371/journal.ppat.1000476.
40. Chang JC, Jimenez JC, Federle MJ. 2015. Induction of a quorum sensingpathway by environmental signals enhances group A streptococcalresistance to lysozyme. Mol Microbiol 97:1097–1113. https://doi.org/10.1111/mmi.13088.
41. Durmort C, Brown JS. 2015. Streptococcus pneumoniae lipoproteins andABC transporters, p 482. In Brown JS, Hammerschmidt S, Orihuela C (ed),Streptococcus pneumoniae: molecular mechanisms of host-pathogeninteractions. Elsevier, New York, NY.
42. Croucher NJ, Walker D, Romero P, Lennard N, Paterson GK, Bason NC,Mitchell AM, Quail MA, Andrew PW, Parkhill J, Bentley SD, Mitchell TJ.2009. Role of conjugative elements in the evolution of the multidrug-resistant pandemic clone Streptococcus pneumoniae Spain23F ST81. JBacteriol 191:1480 –1489. https://doi.org/10.1128/JB.01343-08.
43. Brown JS, Gilliland SM, Holden DW. 2001. A Streptococcus pneumoniaepathogenicity island encoding an ABC transporter involved in ironuptake and virulence. Mol Microbiol 40:572–585. https://doi.org/10.1046/j.1365-2958.2001.02414.x.
44. Brown JS, Gilliland SM, Spratt BG, Holden DW. 2004. A locus containedwithin a variable region of pneumococcal pathogenicity island 1 con-tributes to virulence in mice. Infect Immun 72:1587–1593. https://doi.org/10.1128/IAI.72.3.1587-1593.2004.
45. Khoo SK, Loll B, Chan WT, Shoeman RL, Ngoo L, Yeo CC, Meinhart A.2007. Molecular and structural characterization of the PezAT chromo-somal toxin-antitoxin system of the human pathogen Streptococcuspneumoniae. J Biol Chem 282:19606 –19618. https://doi.org/10.1074/jbc.M701703200.
46. Begley M, Cotter PD, Hill C, Ross RP. 2009. Identification of a novel
Junges et al. Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 18
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

two-peptide lantibiotic, lichenicidin, following rational genome miningfor LanM proteins. Appl Environ Microbiol 75:5451–5460. https://doi.org/10.1128/AEM.00730-09.
47. Dischinger J, Josten M, Szekat C, Sahl HG, Bierbaum G. 2009. Productionof the novel two-peptide lantibiotic lichenicidin by Bacillus licheniformisDSM 13. PLoS One 4:e6788. https://doi.org/10.1371/journal.pone.0006788.
48. Johnson CM, Grossman AD. 2015. Integrative and conjugative elements(ICEs): what they do and how they work. Annu Rev Genet 49:577– 601.https://doi.org/10.1146/annurev-genet-112414-055018.
49. Harvey RM, Stroeher UH, Ogunniyi AD, Smith-Vaughan HC, Leach AJ, PatonJC. 2011. A variable region within the genome of Streptococcus pneumoniaecontributes to strain-strain variation in virulence. PLoS One 6:e19650.https://doi.org/10.1371/journal.pone.0019650.
50. Marion C, Aten AE, Woodiga SA, King SJ. 2011. Identification of anATPase, MsmK, which energizes multiple carbohydrate ABC transportersin Streptococcus pneumoniae. Infect Immun 79:4193– 4200. https://doi.org/10.1128/IAI.05290-11.
51. Kjos M, Miller E, Slager J, Lake FB, Gericke O, Roberts IS, Rozen DE,Veening JW. 2016. Expression of Streptococcus pneumoniae bacteriocinsis induced by antibiotics via regulatory interplay with the competencesystem. PLoS Pathog 12:e1005422. https://doi.org/10.1371/journal.ppat.1005422.
52. Straume D, Stamsas GA, Håvarstein LS. 2015. Natural transformation andgenome evolution in Streptococcus pneumoniae. Infect Genet Evol 33:371–380. https://doi.org/10.1016/j.meegid.2014.10.020.
53. Majchrzykiewicz JA, Lubelski J, Moll GN, Kuipers A, Bijlsma JJ, Kuipers OP,Rink R. 2010. Production of a class II two-component lantibiotic ofStreptococcus pneumoniae using the class I nisin synthetic machineryand leader sequence. Antimicrob Agents Chemother 54:1498 –1505.https://doi.org/10.1128/AAC.00883-09.
54. Kleerebezem M. 2004. Quorum sensing control of lantibiotic production;nisin and subtilin autoregulate their own biosynthesis. Peptides 25:1405–1414. https://doi.org/10.1016/j.peptides.2003.10.021.
55. Moreno-Gamez S, Sorg RA, Domenech A, Kjos M, Weissing FJ, van Doorn
GS, Veening JW. 2017. Quorum sensing integrates environmental cues,cell density and cell history to control bacterial competence. Nat Com-mun 8:854. https://doi.org/10.1038/s41467-017-00903-y.
56. Martin B, Garcia P, Castanie MP, Claverys JP. 1995. The recA gene ofStreptococcus pneumoniae is part of a competence-induced operon andcontrols lysogenic induction. Mol Microbiol 15:367–379. https://doi.org/10.1111/j.1365-2958.1995.tb02250.x.
57. Junges R, Khan R, Tovpeko Y, Åmdal HA, Petersen FC, Morrison DA. 2017.Markerless genome editing in competent streptococci. Methods Mol Biol1537:233–247. https://doi.org/10.1007/978-1-4939-6685-1_14.
58. Khan R, Junges R, Åmdal HA, Chen T, Morrison DA, Petersen FC. 2017. Apositive feedback loop mediated by Sigma X enhances expression of thestreptococcal regulator ComR. Sci Rep 7:5984. https://doi.org/10.1038/s41598-017-04768-5.
59. Yu WH, Hovik H, Chen T. 2010. A hidden Markov support vector machineframework incorporating profile geometry learning for identifying mi-crobial RNA in tiling array data. Bioinformatics 26:1423–1430. https://doi.org/10.1093/bioinformatics/btq162.
60. Lau PC, Sung CK, Lee JH, Morrison DA, Cvitkovitch DG. 2002. PCRligation mutagenesis in transformable streptococci: application andefficiency. J Microbiol Methods 49:193–205. https://doi.org/10.1016/S0167-7012(01)00369-4.
61. Podbielski A, Spellerberg B, Woischnik M, Pohl B, Lutticken R. 1996.Novel series of plasmid vectors for gene inactivation and expressionanalysis in group A streptococci (GAS). Gene 177:137–147. https://doi.org/10.1016/0378-1119(96)84178-3.
62. Salvadori G, Junges R, Morrison DA, Petersen FC. 2016. Overcoming thebarrier of low efficiency during genetic transformation of Streptococcusmitis. Front Microbiol 7:1009. https://doi.org/10.3389/fmicb.2016.01009.
63. Basavanna S, Chimalapati S, Maqbool A, Rubbo B, Yuste J, Wilson RJ,Hosie A, Ogunniyi AD, Paton JC, Thomas G, Brown JS. 2013. The effectsof methionine acquisition and synthesis on Streptococcus pneumoniaegrowth and virulence. PLoS One 8:e49638. https://doi.org/10.1371/journal.pone.0049638.
Interspecies Activity of Small Hydrophobic Peptides Applied and Environmental Microbiology
January 2019 Volume 85 Issue 2 e02297-18 aem.asm.org 19
on January 16, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from