characteristics of oat (avena sativa l.) seed globulins
TRANSCRIPT

Journal of Experimental Botany, Vol. 34, No. 147, pp. 1320-1332, October 1983
Characteristics Of Oat (Avena sativa L.) Seed Globulins
S. R. BURGESS1, P. R. SHEWRY1, G. J. MATLASHEWSKI2,I. ALTOSAAR2 AND B. J. MIFLIN1-3
1 Biochemistry Department, Rothamsted Experimental Station, Harpenden, Herts.AL5 2JQ, UJC.
2 Department of Biochemistry, University of Ottawa, Ottawa, Ontario, Canada KIN 9B4
Received 4 March 1983
ABSTRACTThe major storage proteins of the oat grain are globulins. These have been extracted and characterized.Sucrose density gradient ultracentrifugation separated the globulins into three main fractions withsedimentation coefficients of about 3S, 7S and 12S, of which the latter was the major component.The 12S globulin, when analysed by sodium dodecylsulphate polyacrylamide gel electrophoresis(SDS-PAGE), was found to consist chiefly of proteins of about 55 000 molecular weight. This majorprotein was purified and when reduced gave rise to two classes of subunit. The larger subunits hadmolecular weights between 35 000 and 42 000 and isoelectric points between pH 5-0 and 7-0 whereasthe smaller were between 19 000 and 25 000 molecular weight with isoelectric points of pH 8-0 to 9-0.The large subunits were poor in methionine and not all of them were cleaved by cyanogen bromide. Incontrast to the 12S proteins, the migration on SDS-PAGE of the components of the 7S and 3Sglobulins was not affected by reduction. The major components of the 7S fraction were polypeptides ofabout 55 000 molecular weight. It is suggested from these results that the oat storage globulins havecertain similarities to the globulins present in the seeds of legumes and other dicotyledonous plants.
Key words: Oats; Storage protein; Globulins; Purification; Characterization.
I N T R O D U C T I O NThe major storage proteins in the grain of most cereal species are the alcohol-solubleprolamins (Miflin and Shewry, 1979). In oats, however, the prolamins are only minorcomponents accounting for less than 15% of the total grain N (Peterson and Smith, 1976;Kim, Charbonnier, and Mosse, 1978); the major storage proteins are globulins (Peterson,1978). The globulins of oats were stated by Danielsson (1949) to have sedimentationcoefficients of 2-6 and 8-1, however, Peterson (1978) showed that the major globulin had asedimentation coefficient of 12-1 and a molecular weight of 320 000. The 12S protein wasreported to consist of two types of subunits of molecular weights of 21 700 and 31 700respectively; a model of six of each subunit type per molecule of globulin was proposed. Inthese respects oat globulin resembles the US globulins (legumin) of legumes which havesimilar molecular weights and are composed of 6 subunits of 40 000 molecular weight and 6of 20 000 (Derbyshire, Wright, and Boulter, 1976). The 12S oat globulin also resembles thelegume 1 IS proteins in that the two types of subunit are associated into dimers stabilized bydisulphide bonds (Derbyshire et al., 1976; Matlashewski, Adeli, Altosaar, Shewry, and
3 To whom correspondence should be addressed.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

Burgess et al.—Oat Seed Globulins 1321
Miflin, 1982; Brinegar and Peterson, 1982a) and are synthesized in vitro as a single precursorpolypeptide which is probably cleaved in vivo to give a subunit pair (Sengupta, DeLuca,Bailey, and Verma, 1981; Matlashewski et al., 1982; Walburg and Larkins, 1982; Brinegarand Peterson, 19826).
In this paper we report further studies of the structure and properties of the 12S globulinstorage protein and preliminary studies of the other globulins of oats.
M A T E R I A L S A N D M E T H O D SProtein extractionSeeds of spring oat, cv. Maris Tabard, were dehusked by soaking for 3 h in 50% (v/v) H2SO4. Afterextensive washing the seeds were air dried. Germination tests showed that this procedure did not affectthe viability. The seeds were milled in a Glen Creston hammer mill to pass a 0-5 mm sieve. Meal (400g) was mixed with 100 g Polyclar AT and defatted by stirring twice widi 2-5 1 acetone for 1 h at 4 °C.The albumins and globulins were then extracted by stirring twice for 1 h at 4 °C with 2-5 1 of 0-05 MTris/HCl buffer, pH 8-5, containing 1-0 M NaCl and, in some experiments, also 1-0 mMphenylmethylsulphonyl fluoride (PMSF). The supernatants after centrifugation were combined anddialysed against distilled water at 4 °C. The precipitated globulins were removed by centrifugation,washed with distilled water and finally lyophilized.
Purification of the holoprotelnsGel filtration: Five ml of a solution of the crude globulin in either 0-05 M Tris/HCl buffer, pH 8-5 or
0-05 M sodium phosphate buffer, pH 8-0, both containing 1-0 M NaCl, was applied to a 90 cm x 2-2cm column of Sephacryl S300 equilibrated and eluted with the same buffer.
Zonal isoelectric precipitation: Five ml of a solution of the globulin in 0-05 M sodium phosphatebuffer, pH 8-0, containing 1-0 M NaCl was applied to a 22 cm x 2-2 cm column of Sephadex G50.The column was equilibrated in 0-05 M Na citrate buffer, pH 4-7, containing 1-0 M NaCl and elutedwith downward flow using the same buffer (modified from Wright and Boulter, H)74).
Preparative isoelectric precipitation: Defatted meal was stirred at 20 °C for 2 h with 8-0 ml g"1 of0-1 M sodium phosphate buffer, pH 7-5, containing 1-0 M NaCl, 0-02% sodium azide and 1-0 mMPMSF. After centrifugation the supernatant was dialysed for 18 h at 20 °C against 3 changes of 0-1 Msodium dihydrogen phosphate, pH 4-8, containing 0-2 M NaCl. The precipitated protein was removedby centrifugation and lyophilized. The supernatant, containing mainly the smaller globulins, wasdialysed for 18 h at 4 °C against 3 changes of distilled water. The dialysate was centrifuged and theresulting supernatant and precipitate lyophilized.
Sucrose density gradient ultracentrifugation: Sedimentation coefficients were determined essentiallyas described by Martin and Ames (1961). Globulin preparations (100 mg) were dissolved in 3-0 ml0-05 M Tris/HCl pH 8-0, 1 -0 M NaCl and 1 -0 ml was layered on top of a 5-20% w/w linear sucrosegradient made up in the same buffer.
Marker proteins: E. coli ^-galactosidase (E.C. 3.2.1.23, S2Ow = 16-l,mol. wt 520 000), bovine livercatalase (E.C. 1.1.1.6, 520w = 11-3, mol. wt. 232 000), yeast alcohol dehydrogenase (E.C. 1.1.1.1, S ^ .= 7-4, mol. wt. 150 000) and horse heart myoglobin (S1Ov = 2-0, mol. wt. 18 500) were added to theglobulin extract layered on the gradient. The gradients were centrifuged at 27 000 rev min"1 for 24 h or40 h at 4 °C and fractionated into 1-2 ml aliquots. The position of the oat globulin proteins wasdetermined by their absorbance at 280 nm and sodium dodecylsulphate polyacrylamide gelelectrophoresis (SDS-PAGE). The positions of the marker proteins alcohol dehydrogenase andcatalase were determined as described by Martin and Ames (1961). Myoglobin was measured by itsabsorption at 415 nm and ^-galactosidase assayed using o-nitrophenyl ^-D-galactopyranoside assubstrate.
Purification ofsubunits of the 12S globulinThe reduced total globulin preparation was separated by preparative SDS-PAGE at pH 8-9 using a
Birchover Macro-PAGE apparatus (see Brownstone, 1969, 1976 for a description of the design andperformance of the apparatus). Fractions containing the subunits of the 12S globulin were combined,dialysed against distilled water and lyophilized. Bound SDS was removed from the preparations bywashing with a freshly prepared mixture of anhydrous acetone:triethylamine:acetic acid:water(85:5:5:5 v/v/v/v) (Henderson, Oroszlan, and Konigsberg, 1979). They were then dissolved in 0-05M Tris/HCl buffer, pH 7-0, containing 6-0 M guanidine hydrochloride and separated on a 60 cm x
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

1322 Burgess et al.—Oat Seed Globulins
1-0 cm column of Sephacryl S200 in the same buffer. The fractions containing the subunits were againcombined, dialysed against distilled water and lyophilized.
ElectrophoresisFor separation under non-reducing conditions the proteins were dissolved in 8-0 M urea, 65 mM
Tris pH 6-8, containing 4% (w/v) SDS and separated by SDS-PAGE at pH 8-9 using a modifiedLaemmli (1970) gel system (Forde, Kreis, Bahramian, Matthews, Miflin, Thompson, Bartels, andFlavell, 1981). For separation under reducing conditions the proteins were dissolved in the same buffercontaining 1% (v/v) 2-mercaptoethanol. In some experiments 2-0 mM dithiothreitol was alsoincorporated into the gel. The gels were fixed and stained with 0-25% Coomassie BBR 250 in 40%methanol, 10% trichloroacedc acid (TCA) and destained in 10% TCA. Apparent mol. wts. bySDS-PAGE were determined as described previously (Shewry, Lew, and Kasarda, 1981).
For isoelectric focusing (IEF) the proteins were dissolved in 10 mM glycine, pH adjusted to 8-0 withTris, containing 6-0 M urea. This buffer contained 1% (v/v) 2-mercaptoethanol for separations underreducing conditions and 1% (v/v) Triton X-100 when used to dissolve the subunits prepared bySDS-PAGE. Trie latter was to ensure complete removal of any residual bound SDS. IEF in 5%acrylamide slab gels with pH 3-5-10 ampholyte was as described by Shewry, Ellis, Pratt, and Miflin(1978). The slabs were fixed for 4 h in 10% (w/v) TCA, 5% (w/v) sulphosalicylic acid and washed for12 h with 2 changes of 10% TCA before staining with 0-002% (w/v) Coomassie BBR 250 in 10%(w/v) TCA. Destaining was not necessary. For the determination of isoelectric points, gel trackscontaining separated proteins were cut into strips, which were macerated with distilled water, and, afterstanding for 30 min, the pH was measured. Adjacent tracks containing the same proteins were fixedand stained. The isoelectric points determined in this way were not adjusted for the presence of urea.
AlkylationThe purified protein subunits were reduced and pyridylethylated as described by Friedman, Krull,
and Cavins (1970).
Cyanogen bromide cleavageProteins were dissolved at a concentration of 4-0 mg ml"1 in 70% (v/v) formic acid containing 1%
(w/v) cyanogen bromide (CNBr). After shaking for 16 h in the dark at 20 °C the solutions were dilutedwith distilled water and lyophilized. Control samples were treated in the same way but with omission ofthe CNBr.
Amino acid analysisOne mg protein was hydrolysed for 21 h at 110 °C under N2 with 2-0 ml of 5-7 N HC1 containing
0-1% 2-mercaptoethanol. The amino acids were then determined using an LKB 4400 amino acidanalyser.
RESULTS(7) SDS-PA GE of total globulinsSDS-PAGE of the total globulin preparation under non-reducing conditions gave an intenselystained background with few discrete bands (results not shown). With reduction (Fig. lb) thebackground staining was eliminated and several groups of bands were observed. Although thesample separated in Fig. lb was pyridylethylated, similar results were observed when thesamples were reduced but not alkylated (cf. Fig. 4). The two most intensely-stained groupshad apparent mol. wts. of between 19 and 25 kilodaltons (kd) and 35 and 42 kd. Thesecorrespond to the subunits of the 12-IS globulin described by Peterson (1978) and will becalled the small and large subunits respectively. Two other groups of bands had apparent mol.wts. of around 50-55 kd and 59-65 kd.
(2) Preparation of holoproteins by sucrose density gradient ultracentrifugationSucrose density gradient ultracentrifugation of the total globulin preparation showed a
major peak and two minor ones (Fig. 2a). An enriched preparation of the material in thelower two peaks was also centrifuged Fig. 2b). The sedimentation coefficients (S2Ow) were
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

Burgess et al.—Oat Seed Glnhulinx 1323
apparent ^f!Mr
[4 5 -
t
2 9 5 -
184- f ;
Fio. 1. SDS-PAGE, after reduction and pyridylethylation, of globulin fractions prepared bysedimentation equilibrium ultracentrifugation. (a) mol. wt markers (bovine serum albumin, Mr = 66 kd,ovalbumin, Mr = 45 kd, carbonic anhydrase, Mr = 29-5 kd, ^-lactoglobulin, Mr = 18-4 kd); (b) (g)total globulins; (c) 12S fraction (area 3 from Fig. 2a); (d) 7S fraction (area 2 in Fig. 2a); (e) 3Sfraction (area 1 in Fig. 2a); ( 0 7S fraction (area 4 from Fig. 2b). The brackets in b indicate-four groupsof bands which are discussed in tfie text These have apparent M,s or 59-65 kd, 5CP55 kd, 35-42 kd
and 19-25 kd respectively.
calculated on the basis of the method of Martin and Ames (1961) using Fig. 2a for the largestpeak and Fig. 2b for the smaller two. The partial specific volumes calculated from the aminoacid composition showed that only the middle peak had a value significantly different fromthe standard value of 0-725 assumed by Martin and Ames (1961). This value of 0-605 wasused to correct the apparent S1Ov value—no correction was applied to the other two peaks.Based on these assumptions we found the 520w values of the peak to be about 2-75, 7-1 and12-6S. However, because there is a degree of uncertainty about the figure after the decimalpoint we will refer to the peaks as 3S, 7S and 12S globulin fractions.
SDS-PAGE of the 12S peak, after reduction, showed the presence of the small and largegroups of subunits, although trace amounts of other polypeptides, notably those of highermol. wts. were also present (Fig. lc). Similar analysis of the 3S peak showed two diffusebands with apparent mol. wts. of below 20 kd (Fig. le) while the 7S peak continuedpredominantly 55 kd polypeptides, although smaller amounts of the 12S subunits and ofother polypeptides were also present (Fig. Id). A purer preparation of the 7S globulin wasobtained by sucrose density gradient ultracentrifugation (Fig. 2b) of a preparation which hadbeen enriched in 7S and 3S globulins by precipitation of the 12S globulin at pH 4-8.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

1324 Burgess et al.—Oat Seed Globulins
15 FRACTIONNUMBER
20FRACTIONNUMBER
Fio. 2. Sucrose density gradient ultracentrifugation of total oat globulin, (a) crude preparation; (b)fraction enriched for smaller globulins. The S ^ values were estimated from the diagonal line betweenthe molecular weight markers also run on the gradient. The markers used were horse heart myoglobin(S^w = 2-0), yeast alcohol dehydrogenase (SJO, = 7-4), bovine liver catalase (5,0, = 11-3) and E. coll^-galactosidase (S^ = 16-1). SDS-PAGE separations of proteins from areas 1—4 are shown in Fig. 1.
SDS-PAGE of this preparation (Fig. If) showed that it contained a higher proportion of55 kd polypeptides, although small amounts of other polypeptides were still present. Themigration of the major components of the 3S and 7S globulins on SDS-PAGE was notaffected when the separations were carried out under non-reducing conditions (results notshown).
Amino acid compositions of the 3S, 7S and 12S globulins prepared by ultracentrifugationare given in Table 1. The composition of the 12S fraction is similar to that reported byPeterson (1978). The 3S and 7S fractions both contained high glycine (over 12 mole %), butthe 7S components contained more glutamate + glutamine and arginine.
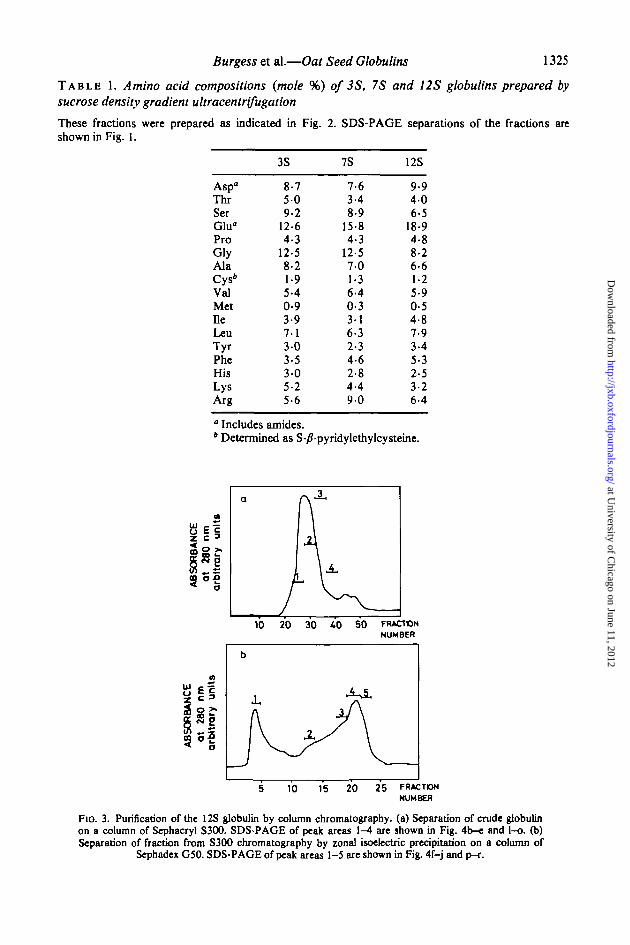
(J) Purification of the 12S holoprotein by column chromatographyLarger quantities of the 12S holoprotein were prepared by column chromatography. The
crude globulin fraction was chromatographed on a column of Sephacryl S300 in 1 0 M NaClat pH 8-5 and a single major peak was eluted followed by two smaller peaks (Fig. 3a).SDS-PAGE, under reducing conditions, showed that fractions taken across the major peakcontained mainly the large and small groups of subunits, although components with highermol. wts. were also present (Fig. 4). When SDS-PAGE was carried out without prior
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from
Burgess et al.—Oat Seed Globulins 1325
T A B L E 1. Amino acid compositions (mole %) of 3S, 7S and 12S globulins prepared by
sucrose density gradient ultracentrifugation
These fractions were prepared as indicated in Fig. 2. SDS-PAGE separations of the fractions areshown in Fig. 1.
Asp"ThrSerGlua
ProGlyAlaCys"ValMetHeLeuTyrPheHisLysArg
3S
8-75 09-2
12-64-3
12-58-21-95-40-93-97 13-03-53-05-25-6
7S
7-63-48-9
15-84-3
12-57-01-36-40-33-16-32-34-62-84-49 0
12S
9-94 06-5
18-94-88-26-61-25-90-54-87-93-45-32-53-26-4
° Includes amides.b Determined as S-/?-pyridylethylcysteine.
8 E !z c 3
is-
a
}10 20 30 40 50 FRACTON
NUMBER
2 0 25 FRACTONNUMBER
Fio. 3. Purification of the 12S globulin by column chromatography. (a) Separation of crude globulinon a column of Sephacryl S300. SDS-PAGE of peak areas 1-4 are shown in Fig. 4b-e and l-o. (b)Separation of fraction from S300 chromatography by zonal isoelectric precipitation on a column of
Sephadex G50. SDS-PAGE of peak areas 1-5 are shown in Fig. 4f-j and p-r.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

1326 Burgess et al.—Oat Seed Globulins
apparent
mm*
29 5 - * *
18-4- f
1 1 7 - *i
a b c d e f g h i j k I m n o p q r
FIG. 4. SDS-PAGE of purified oat globulin fractions, a-k after reduction; \-r without reduction; a, kmol. wt. markers; b-e and l-o, fractions I-4 respectively from S300 chromatography (Fig. 3a); f-jand p-r, are fractions from G50 chromatography (Fig. 3b), f and p are fraction 1; g is fraction 2; hand q are fraction 3; i is fraction 4 and j and r are fraction 5. The arrows in 1 indicate two high mol. wtbands of 80 000 kd which are eliminated by reduction (see text). MoL wL markers are bovine serumalbumin (Mr = 66 kd) carbonic anhydrase (Mr = 29-5 kd) /Mactoglobulin (Mr =18-4 kd), cytochrome
c ( M r = l l - 7 k d ) .
reduction of the samples, the large and small groups of subunits were not observed, a group ofbands with apparent mol. wts. of 51-55 kd being present instead. These bands havepreviously been observed in unreduced separations of a crude 12S globulin fraction(Matlashewski et al., 1982; Brinegar and Peterson, 1982a) and it was suggested that theywere dimers consisting of one large and one small subunit. This is consistent with theirapparent mol. wts. while the effects of reducing agents indicate that the dimers are stabilizedby disulphide bonds. SDS-PAGE under non-reducing conditions also showed the presence oftwo diffuse high molecular weight bands (arrowed in Fig. 4, 1) instead of the bands withapparent mol. wts. between 59 kd and 65 kd which were present under reducing conditions(indicated in Fig. lb).
To purify the 12S globulin further, the peak fractions from S300 chromatography werebulked and separated by zonal isoelectric precipitation on a column of Sephadex G50. Thisprocedure was developed by Shutov and Vaintraub (1965) for the purification of legumin andit is based on the observation that legumin is precipitated at its isoelectric point (around pH4-7). Preliminary investigations showed that the 12S globulin of oats was precipitated at thispH, and so the procedure of Wright and Boulter (1974) was used, modified by the addition of1 0 M NaCl. The first peak eluted from the column (Fig. 3b) contained only a small amountof protein and SDS-PAGE, after reduction, showed that it was enriched in 45-70 kdpolypeptides (Fig. 4f). The second broad peak contained the precipitated 12S globulin andSDS-PAGE under reducing conditions (Fig. 4g-j) showed a lower proportion ofcontaminating polypeptides.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

Burgess et al.—Oat Seed Globulins 1327
(4) Purification and analysis of the subunits of the 12S globulinThe large and small groups of subunits were purified by preparative SDS-PAGE followed
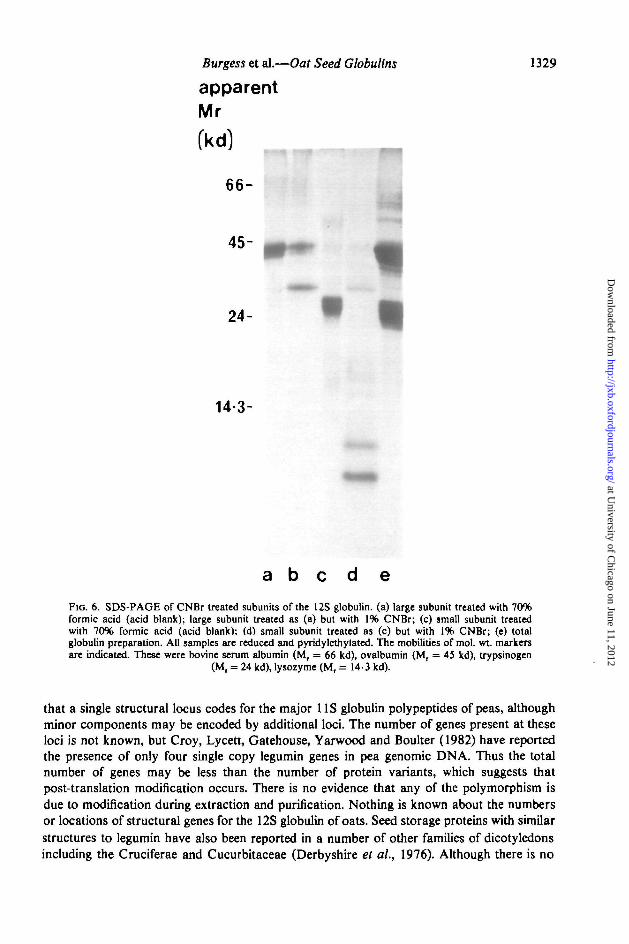
by gel filtration in 6-0 M guanidine on Sephacryl S200. SDS-PAGE (Fig. 5a-d) and IEF(Fig. 5e, f) showed minimal cross-contamination of the two preparations. The IEF separ-ations showed that the large subunit preparation (Fig. 5e) contained about 20 majorbands with pis between 5 and 7, the small subunit preparation 4 or 5 major bands with pis of8-9 (Fig. 5f). The two groups of subunits also differed in their amino acid compositions,notably the large subunits were richer in glutamate + glutamine and the small subunits inaspartate + asparagine and in lysine (Table 2). Both groups contained about 1 mole %cysteine, but methionine was not detected in the large subunits and only about 0-2 mole %(which corresponds to less than one residue per mole) was detected in the small subunits.However, because of the possible destruction of methionine during acid hydrolyses thesevalues may be underestimates. The subunit preparations were also tested for the presence ofmethionine residues by cleavage with CNBr. Blank reactions, in which CNBr was omittedfrom the 70% formic acid solvent, did not give any cleavage of either preparation (Fig. 6a, c)
apparentMr
(kd)
66-HI
36-5-
29-5-
f
13-7-*
11-7-1
a b e d e f
FIG. 5. SDS-PAGE (a-d) and IEF (pH 3-5-10) (e, f) of reduced and pyridylethylated globulinfractions, (a) mol. wt. markers; (b, e) large subunit of 12S globulin; (c, f) small subunit of 12S globulin;(d) total globulin preparation. Mol. wt. markers were bovine serum albumin (Mr = 66 kd), lactatedehydrogenase (Mr = 36-5 kd), carbonic anhydrase (M, = 29-5 kd), ribonuclease (Mr = 13-7 kd),
cytochrome c (M, = 11-7 kd).
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

1328 Burgess et aL—Oat Seed Globulins
T A B L E 2. Amino acid compositions (mole %) of the purified groups ofsubunits of the 12Sglobulin
SDS—PAGE and IEF analyses of these groups ofsubunits are shown in Fig. 5.
Asp"ThrSerGlu"ProGlyAlaCys*ValMetHeLeuTyrPheHisLysArg
Largesubunits
8-33-36-6
23-75-19-96-10-85-80-04-27-73-05-52-12-55-5
" Includes amides.* Determined
Smallsubunits
12-44-47-0
14-95-28-37-51-06-10-2\5-37-53 14-43-03-76-0
as S-/?-pyridylethylcysteine.
indicating the absence of acid-labile peptide bonds. CNBr treatment of the small subunitpreparation gave complete cleavage resulting in two bands with apparent mol. wts. of between5 kd and 7 kd (Fig. 6d). When the large subunit preparation was treated in the same way anadditional band with an apparent mol. wL of about 27 000 was produced, but other bandswith similar mobilities to those of the uncleaved subunits were also present (Fig. 6b). Thismay indicate the presence of methionine in some of the large subunit polypeptides only, but noother bands were observed to account for the other products of the cleavage.
D I S C U S S I O NThe 3S and 7S globulins reported here probably correspond to the a (2-6S) and y (8-IS)globulins of Danielsson (1949), although he showed the presence of a much higher proportionof the y component He was unable to show the presence of a 12S globulin, although a lowconcentration of a 12S component (called S globulin) was present in similar preparationsfrom barley grain. This may be due to the loss of the oat 12S globulin during preparation.Although he initially extracted the globulins with 1-0 M NaCl they were subsequentlydissolved in 0-2 M NaCl at pH 7-0, conditions under which the 12S globulin is only sparinglysoluble (Peterson, 1978).
The 12S globulin of oats is clearly similar in many respects to the US or legumin-typestorage proteins of legumes. Thus both have mol. wts. between 300 kd and 400 kd and arecomposed of disulphide-stabilized dimers, each consisting of a small basic subunit (20-24 kdin legumin) and a larger acidic subunit (27-37 kd in legumin) (Derbyshire et al., 1976). Thesubunits of US legume proteins also show extensive polymorphism when analysed byisoelectric focusing (Casey, 1979; Staswick, Hermodson, and Nielsen, 1981). The origin ofthis polymorphism is not completely understood. Matta and Gatehouse (1982) have shown
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from
Burgess et al.—Oat Seed Globulins 1329
apparentMr
(kcfl
66-
45-
24-
14-3-
a b c d e
FIG. 6. SDS-PAGE of CNBr treated subunits of the 12S globulin, (a) large subunit treated with 70%formic acid (acid blank); large subunit treated as (a) but with 1% CNBr; (c) small subunit treatedwith 70% formic acid (acid blank): (d) small subunit treated as (c) but with 1% CNBr; (e) totalglobulin preparation. All samples are reduced and pyridylethylated. The mobilities of mol. wt markersare indicated. These were bovine serum albumin (Mr = 66 kd), ovalbumin (Mr = 45 ltd), trypsinogen
(M, = 24 kd), lysozyme (Mr = 14-3 kd).
that a single structural locus codes for the major 1 IS globulin polypeptides of peas, althoughminor components may be encoded by additional loci. The number of genes present at theseloci is not known, but Croy, Lycett, Gatehouse, Yarwood and Boulter (1982) have reportedthe presence of only four single copy legumin genes in pea genomic DNA. Thus the totalnumber of genes may be less than the number of protein variants, which suggests thatpost-translation modification occurs. There is no evidence that any of the polymorphism isdue to modification during extraction and purification. Nothing is known about the numbersor locations of structural genes for the 12S globulin of oats. Seed storage proteins with similarstructures to legumin have also been reported in a number of other families of dicotyledonsincluding the Cruciferae and Cucurbitaceae (Derbyshire et al., 1976). Although there is no
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

1330 Burgess et al.—Oat Seed Globulins
evidence that the 12S rf-globulin of barley grain has a legumin-type structure or a storagefunction, Dieckert and Dieckert (1976) have suggested that cocosin, a major storage proteinof coconut endosperms (a monocotyledonous species) is a homologue of legumin. Also, themajor storage protein of rice resembles legumin in that it is synthesized as a 57 kd precursorwhich is subsequently cleaved into 22-23 kd and 37-39 kd subunits (Yamagata, Tanaka, andKasai, 1982; Yamagata, Sugimoto, Tanaka, and Kasai, 1982) although it is not soluble insalt solutions and is classified as a glutelin. This can perhaps be regarded as a more extremedevelopment of the low solubility of the 12S globulin of oats.
The 7S globulin of oats has some structural resemblances to the second major storageprotein type of legumes, the 7S vicilins. Many vicilins have major subunits with mol. wts.between 50 kd and 60 kd and, as in the 7S globulin of oats, these do not appear to beassociated by disulphide bonds (Derbyshire et al, 1976). However, the 7S globulin of oatscontains considerably more glycine than any reported vicilins. Also, the 7S vicilins have arestricted distribution even within the legumes and have never been conclusively identified inother families (Derbyshire et al., 1976). However, it has been shown that the 7-8S globulin ofcoconut endosperms (called t^concocosin) (Dieckert and Dieckert, 1976) and the 7S yglobulin of rice (Morita and Yoshida, 1968; Sawai and Morita, 1970a, b) have structuralsimilarities and that a wide range of species have salt-soluble proteins sedimenting at about 7S(Youle and Huang, 1981). The y-globulins of rice (Morita and Yoshida, 1968) and barley(Danielsson, 1949) are concentrated in the embryos, but similar studies have not beenreported for oats.
Legume seeds have been shown to have minor polypeptides of higher molecular weight(about 80 000) which give rise to two smaller polypeptides upon reduction (Derbyshire andBoulter, 1976; Khan, Gatehouse, and Boulter, 1980). The behaviour of the bands arrowed inFig. 4,1 is similar.
Globulins with S ^ , values between 2S and 4S also occur in the seed of legumes and otherdicotyledonous plants and in some cases these appear to be protease inhibitors (Koshiyama,Kikuchi, Harada, and Fukushima, 1981; Youle and Huang, 1981). However, widely differentamino acid compositions have been reported (Derbyshire et al, 1976; Youle and Huang,1981) and it is not possible to make any direct comparisons with the 3S globulins of oats.
During the completion of this study the authors became aware of a detailed study of thesubunit composition of the 12S globulin of oats by Brinegar and Peterson (1982a). In mostrespects the two studies are consistent
A C K N O W L E D G E M E N T SWe wish to thank Susan Smith for the amino acid analyses. Part of this work was supportedby an Agricultural Canada research contract (DSS 01 5U.01531-1-0808). The authors alsothank Drs. D. M. Peterson and A. C. Brinegar for communicating their results prior topublication and for helpful discussions.
LITERATURE CITEDBRINEGAR, A. C, and PETERSON, D. M., 1982a. Separation and characterization of oat globulin
polypeptides. Archives of Biochemistry and Biophysics, 219,71-9.19826. Synthesis of oat globulin precursors. Analogy to legume US storage protein
synthesis. Plant Physiology, 70,1767-9.BROWNSTONE, A. D., 1969. A versatile system for preparative electrophoresis in acrylamide gel.
Analytical Biochemistry, 27,25-46.1976. Further development of a versatile system for preparative electrophoresis in acrylamide
gel. Ibid. 70,572-91.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

Burgess et al.—Oat Seed Globulins 1331
CASEY, R., 1979. Immunoafflnity chromatography as a means of purifying legumin from Pisum (pea)seeds. Biochemical Journal, 177,509-20.
CROY, R. R. D., LYCETT, G. W., GATEHOUSE, J. A., YARWOOD, J. N., and BOULTER, D., 1982.
Cloning and analysis of cDNAs encoding plant storage protein precursors. Nature, 295,76-9.DANIELSSON, C. E., 1949. Seed globulins of the Gramineae and Leguminosae. Biochemical Journal,
44,387-400.DERBYSHIRE, E., and BOULTER, D., 1976. Isolation of legumin-like protein from Phaseolus aureus and
Phaseolus vulgaris. Phytochemistry, 15,411-14.WRIGHT, D. J., and BOULTER, D., 1976. Legumin and vicilin. Storage proteins of legume seeds.
Ibid. 15,3-24.DIECKERT, J. W., and DDECKERT, M. C , 1976. The chemistry and cell biology of the vacuolar proteins
of seeds. Journal of Food Science, 41,475-82.FORDE, B. G., KREIS, M., BAHRAMIAN, M. B., MATTHEWS, J. A., MIFUN, B. J., THOMPSON, R. D.,
BARTELS, D., and FLAVELL, R. B., 1981. Molecular cloning and analysis of cDNA sequencesderived from pory A+ RNA from barley endosperms: identification of B hordein related clones.Nucleic Acids Research, 9,6689-707.
FRIEDMAN, M., KRULL, L. H., and CAVINS, J. F., 1970. The chromatographic determination of cystineand cysteine residues in proteins as S-/?(4-pyridylethyl)cysteine. Journal of Biological Chemistry,245,3868-71.
KHAN, M. R. I., GATEHOUSE, J. A., and BOULTER, D., 1980. The seed proteins of cowpea {Vignaunguiculata L. Walp.). Journal of Experimental Botany, 31,1599—611.
HENDERSON, L. E., OROSZLAN, S., and KONIGSBERG, W., 1979. A micromethod for complete removalof dodecyl sulphate from proteins by ion-pair extraction. Analytical Biochemistry, 93,153-7.
KIM, S. I., CHARBONNIER, L., and MOSSE, J., 1978. Heterogeneity of avenin, the oat prolamin.Fractionation, molecular weight and amino acid composition. Biochimica et biophysica acta,537,22-30.
KOSHTYAMA, I., Kncucm, M., HARADA, K., and FUKUSHIMA, D., 1981. 2S globulins of soybean seeds.1. Isolation and characterization of protein components. Journal of Agriculture and FoodChemistry, 29,336-40.
LAEMMLI, U. K., 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature, 227,681-5.
MARTIN, R. G., and AMES, B. N., 1961. A method for determining the sedimentation behaviour ofenzymes: Application to protein mixtures. Journal of Biological Chemistry, 236,1372-9.
MATLASHEWSKL G. J., ADELI, K., ALTOSAAR, I., SHEWRY, P. R., and MIFLIN, B. J., 1982. In vitro
synthesis of oat globulin. FEBS Letters, 145,208-12.MATTA, W. A., and GATEHOUSE, J. A., 1982. Inheritance and mapping of storage protein genes in
Pisum sativum L. Heredity, 48,383-92.MIFLIN, B. J., and SHEWRY, P. R., 1979. The synthesis of proteins in normal and high lysine barley
seeds. In Recent advances in the biochemistry of cereals. Eds D. L. Laidman and R. G. WynJones. Academic Press, London. Pp. 239-73.
MORITA, Y., and YOSHIDA, C , 1968. Studies on y globulin of rice embryo. Part 1. Isolation andpurification of y globulin from rice embryo. Agricultural and Biological Chemistry, 32,664-70.
PETERSON, D. M., 1978. Subunit structure and composition of oat seed globulin. Plant Physiology, 62,506-9.and SMITH, D., 1976. Changes in nitrogen and carbohydrate fractions in developing oat groats.
Crop Science, 16,67-71.SAWAI, H., and MORITA, Y., 1970a. Studies on y-globulin of rice embryo. Part II. Separation of three
components of y-globulin by ion exchange chromatography. Agricultural and BiologicalChemistry, 34,53-60.
19706. Studies on y-globulin of rice embryo. Part III. Molecular dimension and chemicalcomposition of yt globulin. Ibid. 34,61-7.
SENGUPTA, C , DELUCA, J., BAILEY, D., and VERMA, D. P. S., 1981. Biosynthesis of soybean storageproteins: Post-translational processing of 7S and US components. Plant Molecular Biology, 1,19-34.
SHEWRY, P. R., ELUS, J. R. S., PRATT, H. M., and MIFUN, B. J., 1978. A comparison of methods for
the extraction and separation of hordein fractions from 29 barley varieties. Journal of the Scienceof Food and Agriculture, 29,433-41.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from

1332 Burgess et al.—Oat Seed Globulins
LEW, E. J-L., and KASARDA, D. D., 1981. Structural homology of storage proteins coded by theHor-l locus of barley (Hordeum vulgare L.). Planta, 153,246-53.
SHUTOV, A. D., and VAJNTRAUB, I. A., 1965. Die Anwendung der Zonensedimentation fur dieAuftrennung der Globuline aus den Samen der Bohne (russ.). Ukraine Biochemistry Journal, 37,177-81.
STASWICK, P. E., HERMODSON, M. A., and NIELSEN, N. C , 1981. Identification of the acidic and basicsubunit complexes of glycinin. Journal of Biological Chemistry, 256,8752-5.
WALBURG, G., and LARKINS, B., 1982. Structure and synthesis of 1 IS globulin from oat seeds. PlantPhysiology, 69,S-123.
WRIGHT, D. J., and BOULTER, D., 1974. Purification and subunit structure of legumin of Viciafaba L.(broad bean). Biochemical Journal, 141,413-18.
YAMAGATA, H., TANAKA, K., and KASAL, Z., 1982. Evidence for a precursor form of rice glutelinsubunits. Agricultural and Biological Chemistry, 46, 321-2.SUGMOTO, T., TANAKA, K., and KASAI, Z., 1982. Biosynthesis of storage proteins in developing
rice seeds. Plant Physiology, 70,1094-1100.YOULE, R. J., and HUANG, A. H. C , 1981. Occurrence of low molecular weight and high cysteine
containing albumin storage proteins in oil seeds of diverse species. American Journal of Botany,68,44-8.
at University of C
hicago on June 11, 2012http://jxb.oxfordjournals.org/
Dow
nloaded from