characteristics of alpha-amylase isozymes in cytologenetically different wheat cultivars
TRANSCRIPT

ISSN 1022�7954, Russian Journal of Genetics, 2014, Vol. 50, No. 7, pp. 722–727. © Pleiades Publishing, Inc., 2014.Original Russian Text © V.P. Netsvetaev, E.D. Badaeva, 2014, published in Genetika, 2014, Vol. 50, No. 7, pp. 825–830.
722
INTRODUCTION
The alpha�amylase of wheat seed endosperm iscontrolled by loci located in the long shoulders ofchromosomes 6 and 7 of the homologous groups ofgenomes A, B and D [1–11]. Genes responsible for theisoenzyme composition of alpha�amylase, which arelocated in the chromosomes of the sixth group, weredesignated as α�Amy A1, α�Amy B1, and α�Amy D1 forhomologs A, B and D respectively [2, 5, 12]. Moreover,genes α�Amy B3, α�Amy B4 and α�Amy B5 [8], as wellas the genetic factor α�Amy B6 [10], were mapped inchromosome 6BL near the locus α�Amy B1. Lociα�Amy A2, α�Amy B2, and α�Amy D2, which controlseveral isoenzymes of alpha�amylase, are located inthe chromosomes of the seventh homologous group[1–5, 11, 12].
The present study was aimed at a complex investi�gation of wheat cultivars carrying different genomeswith an alien translocation in chromosome 7D by viaC�differential chromosome staining and analysis ofthe alpha�amylase isoenzyme spectrum by PAAG�electrophoresis in the Tris�glycine system (pH 8.3).
MATERIALS AND METHODS
The experiments were carried out on samples ofwinter soft wheat (Triticum aestivum, 2n = 6x = 42,
BBAADD) and winter durum wheat (Triticum durum,2n = 4x = 28, BBAA), which were obtained from thecollection of the Belgorod State Research Institute ofAgriculture, Russian Academy of Agricultural Sci�ences. The study also involved the amphidiploidT. durum × Hordeum chilense HT 135 (DH)T�46�08(2n = 6x = 42, BBAAHchHch), which was kindly pro�vided by I.G. Loskutov (All�Russia Research Instituteof Plant Industry, St. Petersburg, Russia), and the softwheat cultivar Dobrynya carrying the couch grasstranslocation in the chromosome 7D (Triticum aesti�vum, 2n = 6x = 42, BBAADD), which was kindly pro�vided by S.N. Sibikeev (Agricultural Research Insti�tute for South�East Region (ARISER), Saratov, Rus�sia).
Grains were first sterilized with KMnO4 solution at65–70°C for 30 s, washed with distilled water, andcouched in the dark at room temperature for 4 days. Toextract the enzyme, germinated grains were put in0.2% NaHCO3 containing 20% sucrose and 0.03%bromphenol blue to the final volume of the extractingsolution 250 μL per grain. Each grain was thoroughlyhomogenized and centrifuged at 10000 rpm for 4 min.Closed tubes containing the supernatant were warmed upat 80°C for 20–25 min, and 2–3 μL of the supernatantwere introduced into the starting wells on the gel.
Characteristics of Alpha�Amylase Isozymes in Cytologenetically Different Wheat CultivarsV. P. Netsvetaeva and E. D. Badaevab
aBelgorod State Research Institute of Agriculture, Russian Academy of Agricultural Sciences, Belgorod, 308001 Russiae�mail: [email protected]
b Vavilov Institute of General Genetics, Russian Academy of Sciences, Moscow, 119991 Russiae�mail: [email protected]
Received December 30, 2013; in final form, February 2, 2014
Abstract—The isoenzyme composition of alpha�amylase is studied by polyacrylamide gel electrophoresis inTris�glycine (pH 8.3) system in wheat cultivars with different genome composition. We show that durumwheat (Triticum durum, 2n = 4x = 28, BBAA) lacks the isoenzymes encoded by 6D and 7D chromosomes thatare present in common wheat zymograms (Triticum aestivum, 2n = 6x = 42, BBAADD). A similar pattern isobserved in a synthetic allohexaploid carrying the BBAA genomes of wheat and the HchHch genome of barley(Hordeum chilense). Our method of electrophoresis fails to reveal additional variants of alpha�amylaseencoded by the barley genome, although C�banding analysis confirms the genomic structure BBAAHchHch ofthis allopolyploid. The electrophoretic spectrum of the spring common wheat cultivar Dobrynya with thewheat�Agropyron translocation 7DL�7AiL contains all of the alpha�amylase isoenzymes typical for commonwheat (2n = 6x = 42, BBAADD) except for the zymotype encoded by the long arm of chromosome 7D. Thisobservation confirms the results of cytogenetic analysis that identified a 7DL�7AiL translocation in this cul�tivar. No additional alpha�amylase isoenzymes encoded by Agropyron chromosome have been observed. Ourdata indicate that analysis of wheat�alien hybrids or introgressive forms should be carried out using a complexof different methods.
DOI: 10.1134/S102279541407014X
PLANT GENETICS
RUSSIAN JOURNAL OF GENETICS Vol. 50 No. 7 2014
CHARACTERISTICS OF ALPHA�AMYLASE ISOZYMES IN CYTOLOGENETICALLY 723
The conditions of electrophoresis were as describedin [13]. Electrophoretic separation was performed inTris�glycine buffer, pH 8.3 at 300 V for 2.5 h. Amylaseswere incubated in 1% hydrolyzed starch solution in0.1 M acetate buffer, pH 5.4, which was preliminarilyboiled at constant stirring [13]. Incubation was per�formed at room temperature for 30 min. After incuba�tion, the gels were washed with running water andstained with iodine solution in potassium iodide(0.5 g KI, 260 mg I2, and 6 g trichloracetic acidbrought up to 100 mL with water).
Cytological analysis of chromosomes was per�formed in cells of the root meristem of germinatinggrains. The standard method of chromosome C�dif�ferential staining was used in the study [14, 15].
RESULTS AND DISCUSSION
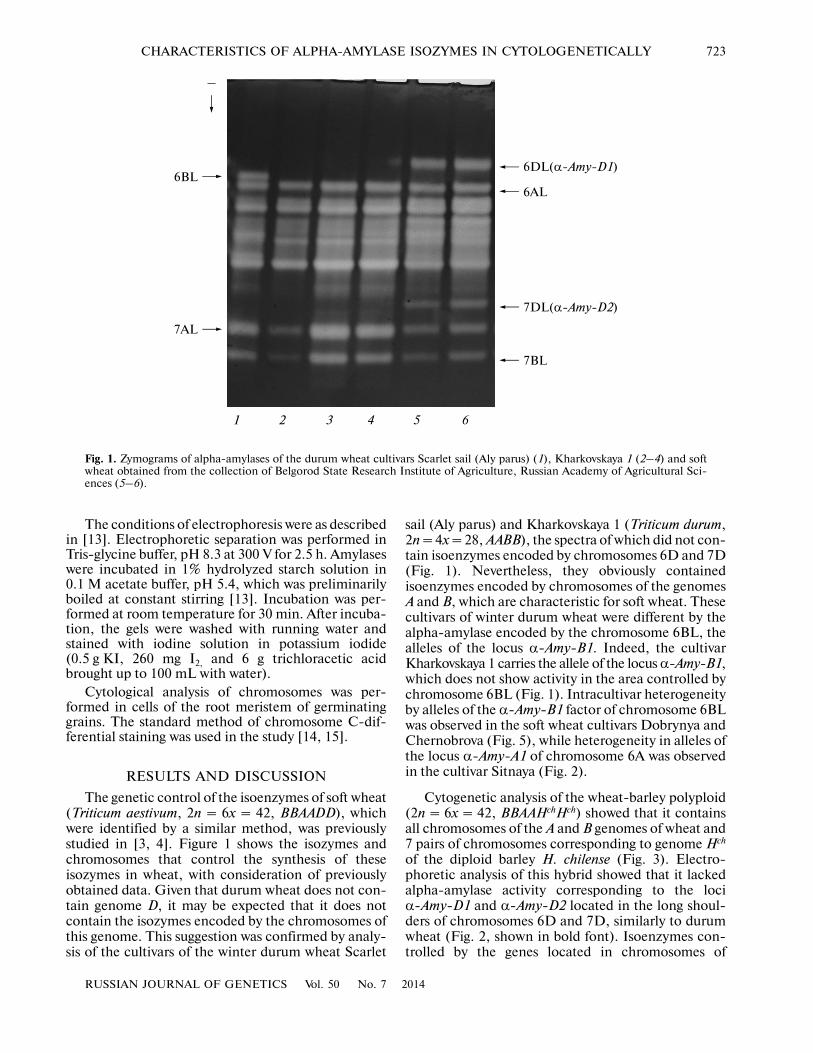
The genetic control of the isoenzymes of soft wheat(Triticum aestivum, 2n = 6x = 42, BBAADD), whichwere identified by a similar method, was previouslystudied in [3, 4]. Figure 1 shows the isozymes andchromosomes that control the synthesis of theseisozymes in wheat, with consideration of previouslyobtained data. Given that durum wheat does not con�tain genome D, it may be expected that it does notcontain the isozymes encoded by the chromosomes ofthis genome. This suggestion was confirmed by analy�sis of the cultivars of the winter durum wheat Scarlet
sail (Aly parus) and Kharkovskaya 1 (Triticum durum,2n = 4x = 28, AABB), the spectra of which did not con�tain isoenzymes encoded by chromosomes 6D and 7D(Fig. 1). Nevertheless, they obviously containedisoenzymes encoded by chromosomes of the genomesA and B, which are characteristic for soft wheat. Thesecultivars of winter durum wheat were different by thealpha�amylase encoded by the chromosome 6BL, thealleles of the locus α�Amy�B1. Indeed, the cultivarKharkovskaya 1 carries the allele of the locus α�Amy�B1,which does not show activity in the area controlled bychromosome 6BL (Fig. 1). Intracultivar heterogeneityby alleles of the α�Amy�B1 factor of chromosome 6BLwas observed in the soft wheat cultivars Dobrynya andChernobrova (Fig. 5), while heterogeneity in alleles ofthe locus α�Amy�A1 of chromosome 6A was observedin the cultivar Sitnaya (Fig. 2).
Cytogenetic analysis of the wheat�barley polyploid(2n = 6x = 42, BBAAHchHch) showed that it containsall chromosomes of the A and B genomes of wheat and7 pairs of chromosomes corresponding to genome Hch
of the diploid barley H. chilense (Fig. 3). Electro�phoretic analysis of this hybrid showed that it lackedalpha�amylase activity corresponding to the lociα�Amy�D1 and α�Amy�D2 located in the long shoul�ders of chromosomes 6D and 7D, similarly to durumwheat (Fig. 2, shown in bold font). Isoenzymes con�trolled by the genes located in chromosomes of
6BL
7AL
6DL(α�Amy�D1)
6AL
7DL(α�Amy�D2)
7BL
–
1 2 3 4 5 6
Fig. 1. Zymograms of alpha�amylases of the durum wheat cultivars Scarlet sail (Aly parus) (1), Kharkovskaya 1 (2–4) and softwheat obtained from the collection of Belgorod State Research Institute of Agriculture, Russian Academy of Agricultural Sci�ences (5–6).

724
RUSSIAN JOURNAL OF GENETICS Vol. 50 No. 7 2014
NETSVETAEV, BADAEVA
genomes A and B were present on the electrophore�grams. These data confirm that this polyploid carriesgenomes A and B but lacks genome D. It is noteworthythat alpha�amylase zymograms of the wheat�barleyallopolyploid lacked the additional bands of enzy�matic activity that are not typical for durum wheat.Apparently, bands of alpha�amylase activities, whichare controlled by barley chromosomes, were located inthe monomorphous area of the wheat zymograms thatmasks them.
Cytogenetic analysis of the spring soft wheat culti�var Dobrynya showed it carrying a normal karyotypeof soft wheat, except for chromosome 7D. The longshoulder of chromosome 7D lacked the distal and sub�terminal blocks that are typical for soft wheat. At thesame time, it revealed a quite bright intercalaryC�band in a position atypical for wheat, and the lengthof the chromosome shoulder itself was longer than thatof the normal chromosome 7D. Based on these traits,it was suggested that the cultivar Dobrynya carried thealien translocation 7DL�7AiL in chromosome 7D(Fig. 4). Comparative analysis of the alpha�amylasecompositions of this cultivar and the typical soft wheat
showed an absence of enzymatic activity associatedwith the locus α�Amy�D2, which is located in the longshoulder of chromosome 7D in the cultivar Dobrynya(Fig. 5). This confirms the idea that the long shoulderof chromosome 7D contained alien genetic material.It is significant that in this case, as in the case with thewheat�barley polyploidy, the zymogram did not revealnew enzyme variations controlled by the fragment ofthe couch grass chromosome.
Therefore, the data of cytological and isoenzy�matic analyses coincide with each other and lead tothe inevitable conclusion that translocation in the cul�tivar Dobrynya affects the long shoulder of chromo�some 7D. At the same time, the cultivars Dobrynyaand Chernobrova appeared to be heterogenic byalpha�amylase in the slow mobility zone of the zymo�gram as a result of the different alleles of the locusα�Amy�B1 located in chromosome 6BL (Fig. 4).
Therefore, the study of alpha�amylase isoenzymesallows the identification of certain genes that controltheir synthesis. Based on these data, it may be prelim�inarily concluded that the studied wheat sampleseither have or lack the genomes or certain chromo�
6BL
7AL
6DL(α�Amy�D1)
6AL(α�Amy�A1)
7DL(α�Amy�D2)
6BL
–
1 2 3 4
7BL
Fig. 2. Zymograms of alpha�amylases of the wheat�barley allopolyploid (BBAAHchHch)—HT 135 (DH)T�46�08 (1, 2) and thesoft wheat cultivar Sitnaya (3, 4).

RUSSIAN JOURNAL OF GENETICS Vol. 50 No. 7 2014
CHARACTERISTICS OF ALPHA�AMYLASE ISOZYMES IN CYTOLOGENETICALLY 725
4B
1Cch
3Cch
7Cch
6B4Cch
2B
1A
3B
2A 7B
1B
7A
4B
5A
3A
1B6B
1A6Cch
7B
4A
2B
1Cch
3A
5A
2A
5Cch
2Cch
3B
5Cch
3Cch
2Cch6A
6Cch
4A6A7A
5B 5B
4Cch
7Cch
Fig. 3. Differential staining of the metaphase plate of the wheat�barley allopolyploid. Chromosomes are numbered in accordancewith the genetic classification of the chromosomes of soft wheat and Hordeum chilense [17].
1 2 3 4 5 6 7
A
B
D
7DT7DL�7AiLDobrynya
Fig. 4. The diploid chromosomal complement of the spring soft wheat cultivar Dobrynya (letters indicate the genomes; numeralsindicate the numbers of chromosomes, arrow indicates the presence of the couch grass translocation). The normal chromosome7D of the soft wheat cultivar Saratovskaya 68 is shown individually.

726
RUSSIAN JOURNAL OF GENETICS Vol. 50 No. 7 2014
NETSVETAEV, BADAEVA
somes. This analysis allowed us to define more pre�cisely the shoulders affected by alien translocationsonce they occur in chromosome 6 or 7 of the homolo�gous groups. However, it does not always help identifythe alien material. The combination of isoenzymaticand cytogenetic analyses is considered to be the onlyway to perform a sufficiently full and adequate assess�ment of the chromosomal and genomic compositionof hybrid wheat forms.
ACKNOWLEDGMENTS
The authors thank Doctor of Science I.G. Losku�tov (All�Russia Research Institute of Plant Industry,Russian Academy of Agricultural Sciences, St. Peters�burg, Russia) and Doctor of Science S.N. Sibikeev(Agricultural Research Institute for South�EastRegion (ARISER), Saratov, Russia) for the kindlyprovided materials.
This work was supported by the Russian Foundationfor Basic Researches, by the Program of the Presidiumof the Russian Academy of Sciences “Wildlife: Contem�porary Condition and Developmental Problems,” andthe subprogram, “The Dynamics and Preservation ofGenofonds.”
REFERENCES
1. Gale, M.D., Law, C.N., Chojecki, A.J., andKempton, R.A., Genetic control of α�amylase produc�tion in wheat, Theor. Appl. Genet., 1983, vol. 64,pp. 309–316.
2. Ainsworth, C.C., Doherty, P., Edwards, K.G.K., et al.,Allelic variation at α�amylase loci in hexaploid wheat,Theor. Appl. Genet., 1985, vol. 70, pp. 400–406.
3. Illichevskii, N.N., Metakovskii, E.V., and Sozinov, A.A.,Polymorphism and genetic control of α�amylaseamong native cultivars of summer bread wheat, Gene�tika (Moscow), 1989, vol. 25, no. 12, pp. 2176–2186.
4. Rybalka, A.I., Bodelan, O.P., and Litvinenko, N.A.,Genetic analysis of α�amylase of the wheat grain, Gene�tika (Moscow), 1989, vol. 25, no. 12, pp. 2187–2198.
5. Nishikawa, K. and Nobuhara, M., Genetic studies onα�amylase isozymes in wheat: 1. Location of genes andvariation in tetra� and hexaploid wheat, Jpn. J. Genet.,1971, vol. 46, pp. 345–358.
6. Nishikawa, K., Chromosome mapping by use of aneu�ploids in wheat, Wheat Inform. Serv., 1991, no. 72,pp. 60–63.
7. Mrva, K. and Mares, D.J., Regulation of high pI alpha�amylase synthesis in wheat aleurone by a gene(s)located on chromosome 6B, Euphytica, 1999, vol. 109,pp. 17–23.
8. Nishikawa, K., Ban, T., and Furuta, Y., Telocentricmapping of alpha�amylase loci in wheat, Wheat Inform.Serv., 1993, no. 77, pp. 39–45.
(α�Amy�A1)
6DL(α�Amy�D1)
6BL(α�Amy�B1)
7DL(α�Amy�D2)
7BL(α�Amy�B2)
–
1 2 3 4 5
6AL
7AL(α�Amy�A2)
Fig. 5. Zymograms of alpha�amylase of the soft wheat cultivar Dobrynya (1–3) and Chornobrova (4, 5).

RUSSIAN JOURNAL OF GENETICS Vol. 50 No. 7 2014
CHARACTERISTICS OF ALPHA�AMYLASE ISOZYMES IN CYTOLOGENETICALLY 727
9. Rybalka, O.I., Yakist’ pshenitsi ta ii polipshennya(Wheat Quality and Its Improvement), Kiiv: Logos,2011, pp. 221–234.
10. Netsvetaev, V.P., Akinshina, O.V., and Bondarenko, L.S.,Genetic control of several α�amylase isozymes in win�ter hexaploid wheat, Russ. J. Genet., 2012, vol. 48,no. 3, pp. 347–349.
11. Mac Gene, Gene Symbols, Gene Classes and Refer�ences. http://shigen.lab.nig.ac.jp/wheat/komugi/genes/macgene/2005/GeneSymbol.pdf
12. Nishikawa, K., Furuta, Y., Hina, Y., and Yamada, T.,Genetic studies of α�amylase isozymes in wheat:4. Genetic analyses in hexaploid wheat, Jpn. J. Genet.,1981, vol. 56, pp. 385–395.
13. Netsvetaev, V.P., Locatization of β�amylase locus (Bmy 1)in the barley chromosome 4, Tsitol. Genet., 1993,vol. 27, no. 5, pp. 74–78.
14. Badaeva, E.D., Badaev, N.S., Gil, B.S., andFilatenko, A.A., Intraspecific karyotype divergence inTriticum araraticum (Poaceae), Plant Syst. Evol., 1994,vol. 192, pp. 117–145.
15. Pukhal’skii, V.A., Solov’ev, A.A., and Badaeva, E.D.,Praktikum po tsitologii i tsitogenetike rastenii: Uchebnoeposobie dlya vuzov (Plant Cytology and Cytogenetics(Practical Course for High School)), Moscow: Kolos,2007.
16. Cabrera, A., Friebe, B., Jiang, J., and Gill, B.S., Char�acterization of Hordeum chilense chromosomes byC�banding and in situ hybridization using highlyrepeated DNA probes, Genome, 1995, vol. 38, no. 3,pp. 435–442.
Translated by M. Bibov