isozymes lysozyme evidence species-specific control of egg-white
TRANSCRIPT

Proc. Nat. Acad. Sci. USAVol. 71, No. 5, pp. 1653-1657, May 1974
Isozymes of Lysozyme in Leukocytes and Egg White: Evidencefor the Species-Specific Control of Egg-White Lysozyme Synthesis
(leukocyte granules/bone marrow/hormones/antibacterial mechanisms/oviduct/muramidase)
ALEXANDER HINDENBURG*, JOHN SPITZNAGELt, AND NORMAN ARNHEIMt** Department of Biochemistry, State University of New York, Stony Brook, N.Y. 11790; and t Department of Bacteriology andImmunology, School of Medicine, University of North Carolina, Chapel Hill, N.C. 27514
Communicated by Bentley Glass, February 15, 1974
ABSTRACT Two structurally distinct forms of egg-white lysozyme (EC 3.2.1.17) are known. The egg white ofsome species contains both of these forms, while the eggwhite of other species appears to contain only one or theother ofthem. We have immunological and electrophoreticevidence that the chicken, which has only one lysozymetype in its egg white, contains both types in its poly-morphonuclear leukocytes. Experiments on Embdengoose bone marrow showed that this tissue also containsboth Iysozymes, even though the egg white of this speciescontains only one of them. Our studies suggest that manyavian species have the genetic loci that code for bothforms of lysozyme, but that a species-specific regulatorymechanism controls whether one or the other or both ofthem are expressed during egg white production. The factthat two distinct lysozymes are present in chicken leuko-cytes may be of significance to the antibacterial mecha-nism of these cells, especially in light of the fact that theylack myeloperoxidase, an important leukocyte enzyme inmammals.
Two forms of lysozyme (EC 3.2.1.17) that differ radicallyfrom each other in molecular weight (1, 2), amino-acid com-position (1-4), amino-terminal amino-acid sequence (1, 2, 5),and immunological cross reactivity (6) have been found inavian egg whites. This and other evidence (7) suggest thatthese enzymes are the products of two distinct genetic loci.One of these forms of lysozyme is typified by the enzymefound in chicken egg white ("chick-type" lysozyme), andthe other form by the lysozyme found in the egg white of theEmbden goose (3, 4) ("goose-type" lysozyme).The egg whites of some species appear to contain only a
"chick-type" lysozyme (e.g., chicken and Peking duck), otherspecies only a "goose-type" lysozyme (e.g., Embden goose);still others contain both types of lysozyme (black swan; refs.2 and 6; and Canada goose, ref. 7).We present data that demonstrate that even though a
species may have only one form of lysozyme in its egg white,it can contain both forms of the enzyme in other tissues.Our finding of multiple forms of lysozyme in chicken poly-
morphonuclear leukocyte (PMN) cytoplasmic granules andin Embden goose bone marrow may eventually lead to newinformation about the evolution of regulatory systems invertebrates as well as on the antibacterial mechanisms oflysosomes.
MATERIALS AND METHODSAntiserums and Proteins. Rabbit antiserums directed against
purified Embden goose egg-white lysozyme (gift of Dr. R.
Canfield) and 3X crystalline chicken egg-white lysozyme(Pentex) were the same as those used previously (6). In thosecases where lysozyme activity in a sample was to be measuredin the presence of rabbit antiserum, the antiserum was firsttreated with Bentonite (0.5% final concentration) to removerabbit serum lysozyme.
Gel Diffusion Experiments. These studies were carried outin 1% agar containing 0.15 M NaCl, 0.05 M Tris HCl (pH7.45), and sodium azide as a preservative.
Lysoplate Enzyme Assays. The concentration of lysozymein the large granule fraction from chicken leukocytes was de-termined by either the lysoplate method of Osserman andLawlor (8) or a modification of this technique, where thelysoplates contained 1% Difco agar, 0.033 M sodium phos-phate buffer (pH 6.2), 0.15 M sodium chloride, 0.5 mg/mlof Micrococcus luteus (Worthington), and sodium azide as apreservative.
Antibody Absorption Experiments. Equal volumes of thepreparation of large cytoplasmic granules, or in some casespartially purified lysozyme, and either antibody againstchicken egg-white lysozyme or antibody against Embdengoose egg-white lysozyme were mixed, incubated for one hourat 230, and stored overnight at 4°. The supernatant was col-lected after centrifugation at 1000 X g for 30 min.
Acrylamide Gel Electrophoresis. The "goose-type" and"chick-type" lysozymes were separated in 15% acrylamidegels without a stacking gel at pH 4.5 (9), with the buffersdescribed by Brewer and Ashworth (10). The gels (in tubes13 cm X 0.5 cm) were run for 2.5 hr at a constant currentof 5 mA per gel. After electrophoresis, they were fractionatedinto 0.2-ml samples in a Savant Auto-Gel Divider, with am-monium acetate buffer (0.1 M acetate, pH 9.45), and sub-jected to further tests.
Bone Marrow was obtained from the leg bones of a goose ofthe Embden and Toulouse variety. The marrow was sonicatedin ammonium acetate buffer (0.1 M, pH 9.45), lyophilized,and stored at -20°.
A Large Cytoplasmic Granule Fraction from Chicken PMNWas Prepared according to the method of Brune and Spitznagel(9). Briefly, 6.68 X 108 peritoneal exudate cells from chickens(66% PMN, 17% lymphocytes, 9% monocytes, 8% eosino-phils) were homogenized in 10 ml of 0.32 M sucrose with 3units of heparin per ml at 4°. Cells were homogenized witha Kontes type D glass homogenizer fitted with a Teflon pestle
1653
Abbreviation: PMN, polymorphonuclear leukocyte.t To whom reprint requests should be sent.
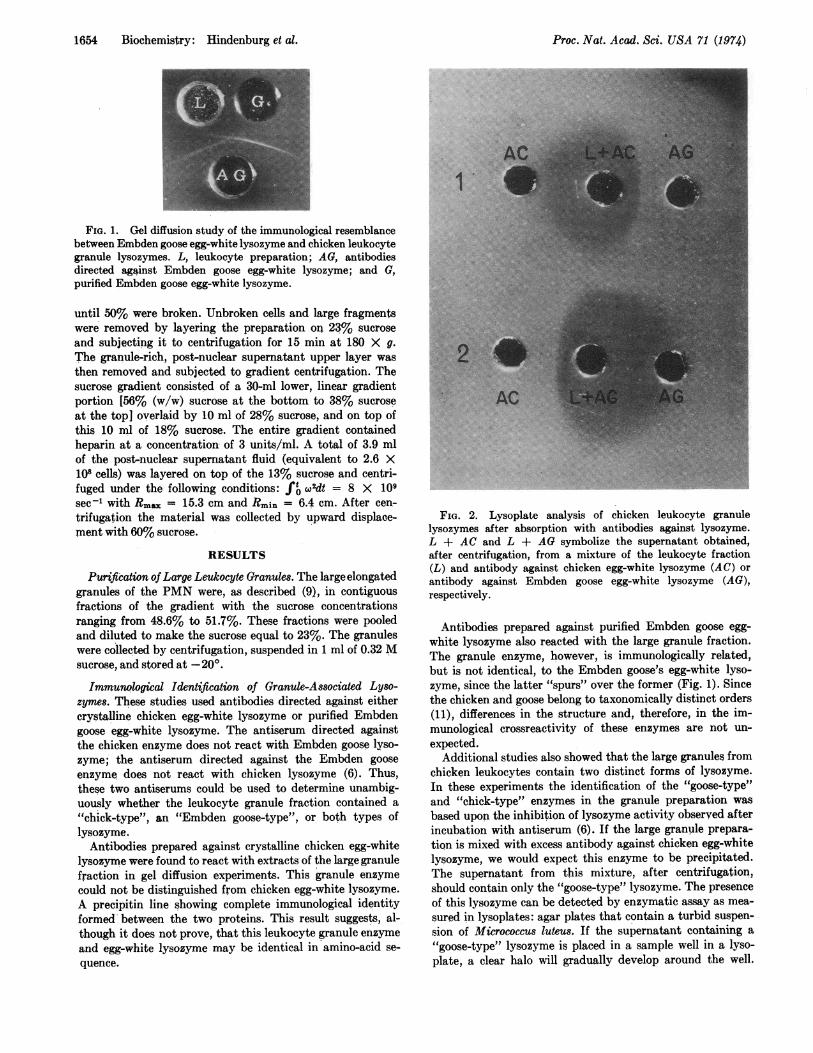
1654 Biochemistry: Hindenburg et al.
FIG. 1. Gel diffusion study of the immunological resemblancebetween Embden goose egg-white lysozyme and chicken leukocytegranule lysozymes. L, leukocyte preparation; AG, antibodiesdirected against Embden goose egg-white lysozyme; and G,purified Embden goose egg-white lysozyme.
until 50% were broken. Unbroken cells and large fragmentswere removed by layering the preparation on 23% sucroseand subjecting it to centrifugation for 15 min at 180 X g.The granule-rich, post-nuclear supernatant upper layer wasthen removed and subjected to gradient centrifugation. Thesucrose gradient consisted of a 30-ml lower, linear gradientportion [56% (w/w) sucrose at the bottom to 38% sucroseat the top] overlaid by 10 ml of 28% sucrose, and on top ofthis 10 ml of 18% sucrose. The entire gradient containedheparin at a concentration of 3 units/ml. A total of 3.9 mlof the post-nuclear supernatant fluid (equivalent to 2.6 X108 cells) was layered on top of the 13% sucrose and centri-fuged under the following conditions: fotw2dt = 8 X 109sec1 with Rma = 15.3 cm and Rmin = 6.4 cm. After cen-trifugation the material was collected by upward displace-ment with 60% sucrose.
RESULTS
Purification ofLarge Leukocyte Granules. The large elongatedgranules of the PMN were, as described (9), in contiguousfractions of the gradient with the sucrose concentrationsranging from 48.6% to 51.7%. These fractions were pooledand diluted to make the sucrose equal to 23%. The granuleswere collected by centrifugation, suspended in 1 ml of 0.32 Msucrose, and stored at -20°.
Immunological Identification of Granule-Associated Lyso-zymes. These studies used antibodies directed against eithercrystalline chicken egg-white lysozyme or purified Embdengoose egg-white lysozyme. The antiserum directed againstthe chicken enzyme does not react with Embden goose lyso-zyme; the antiserum directed against the Embden gooseenzyme does not react with chicken lysozyme (6). Thus,these two antiserums could be used to determine unambig-uously whether the leukocyte granule fraction contained a"chick-type", an "Embden goose-type", or both types oflysozyme.
Antibodies prepared against crystalline chicken egg-whitelysozyme were found to react with extracts of the large granulefraction in gel diffusion experiments. This granule enzymecould not be distinguished from chicken egg-white lysozyme.A precipitin line showing complete immunological identityformed between the two proteins. This result suggests, al-though it does not prove, that this leukocyte granule enzymeand egg-white lysozyme may be identical in amino-acid se-quence.
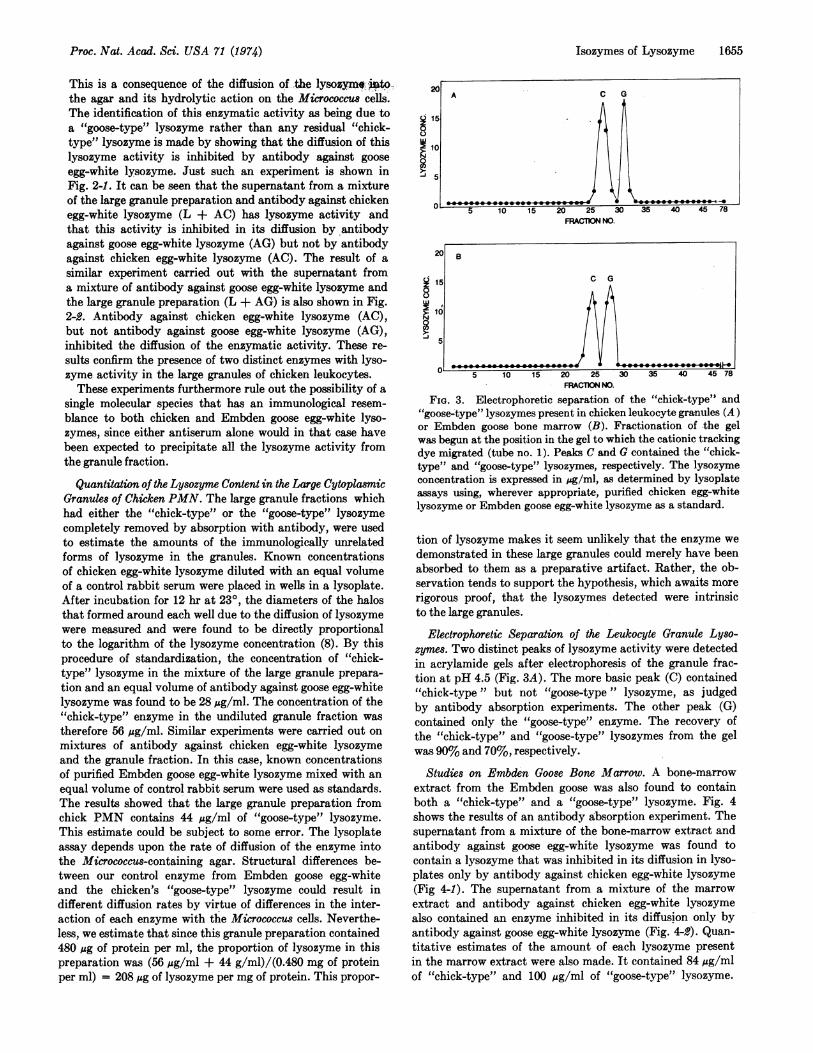
FIG. 2. Lysoplate analysis of chicken leukocyte granulelysozymes after absorption with antibodies against lysozyme.L + AC and L + AG symbolize the supernatant obtained,after centrifugation, from a mixture of the leukocyte fraction(L) and antibody against chicken egg-white lysozyme (AC) orantibody against Embden goose egg-white lysozyme (AG),respectively.
Antibodies prepared against purified Embden goose egg-white lysozyme also reacted with the large granule fraction.The granule enzyme, however, is immunologically related,but is not identical, to the Embden goose's egg-white lyso-zyme, since the latter "spurs" over the former (Fig. 1). Sincethe chicken and goose belong to taxonomically distinct orders(11), differences in the structure and, therefore, in the im-munological crossreactivity of these enzymes are not un-expected.
Additional studies also showed that the large granules fromchicken leukocytes contain two distinct forms of lysozyme.In these experiments the identification of the "goose-type"and "chick-type" enzymes in the granule preparation wasbased upon the inhibition of lysozyme activity observed afterincubation with antiserum (6). If the large granule prepara-tion is mixed with excess antibody against chicken egg-whitelysozyme, we would expect this enzyme to be precipitated.The supernatant from this mixture, after centrifugation,should contain only the "goose-type" lysozyme. The presenceof this lysozyme can be detected by enzymatic assay as mea-sured in lysoplates: agar plates that contain a turbid suspen-sion of Micrococcus luteus. If the supernatant containing a
"goose-type" lysozyme is placed in a sample well in a lyso-plate, a clear halo will gradually develop around the well.
Proc. Nat. Acad. Sci. USA 71 (1974)
Proc. Nat. Acad. Sci. USA 71 (1974)
This is a consequence of the diffusion of the lysozym. mtg-the agar and its hydrolytic action on the Micrococcus cells.The identification of this enzymatic activity as being due toa "goose-type" lysozyme rather than any residual "chick-type" lysozyme is made by showing that the diffusion of thislysozyme activity is inhibited by antibody against gooseegg-white lysozyme. Just such an experiment is shown inFig. 2-1. It can be seen that the supernatant from a mixtureof the large granule preparation and antibody against chickenegg-white lysozyme (L + AC) has lysozyme activity andthat this activity is inhibited in its diffusion by antibodyagainst goose egg-white lysozyme (AG) but not by antibodyagainst chicken egg-white lysozyme (AC). The result of asimilar experiment carried out with the supernatant froma mixture of antibody against goose egg-white lysozyme andthe large granule preparation (L + AG) is also shown in Fig.2-2. Antibody against chicken egg-white lysozyme (AC),but not antibody against goose egg-white lysozyme (AG),inhibited the diffusion of the enzymatic activity. These re-sults confirm the presence of two distinct enzymes with lyso-zyme activity in the large granules of chicken leukocytes.These experiments furthermore rule out the possibility of a
single molecular species that has an immunological resem-blance to both chicken and Embden goose egg-white lyso-zymes, since either antiserum alone would in that case havebeen expected to precipitate all the lysozyme activity fromthe granule fraction.
Quantitation of the Lysozyme Content in the Large CytoplasmicGranules of Chicken PMN. The large granule fractions whichhad either the "chick-type" or the "goose-type" lysozymecompletely removed by absorption with antibody, were usedto estimate the amounts of the immunologically unrelatedforms of lysozyme in the granules. Known concentrationsof chicken egg-white lysozyme diluted with an equal volumeof a control rabbit serum were placed in wells in a lysoplate.After incubation for 12 hr at 23°, the diameters of the halosthat formed around each well due to the diffusion of lysozymewere measured and were found to be directly proportionalto the logarithm of the lysozyme concentration (8). By thisprocedure of standardization, the concentration of "chick-type" lysozyme in the mixture of the large granule prepara-tion and an equal volume of antibody against goose egg-whitelysozyme was found to be 28 sg/ml. The concentration of the"chick-type" enzyme in the undiluted granule fraction wastherefore 56 ,g/ml. Similar experiments were carried out onmixtures of antibody against chicken egg-white lysozymeand the granule fraction. In this case, known concentrationsof purified Embden goose egg-white lysozyme mixed with anequal volume of control rabbit serum were used as standards.The results showed that the large granule preparation fromchick PMN contains 44 /g/ml of "goose-type" lysozyme.This estimate could be subject to some error. The lysoplateassay depends upon the rate of diffusion of the enzyme intothe Micrococcus-containing agar. Structural differences be-tween our control enzyme from Embden goose egg-whiteand the chicken's "goose-type" lysozyme could result indifferent diffusion rates by virtue of differences in the inter-action of each enzyme with the Micrococcus cells. Neverthe-less, we estimate that since this granule preparation contained480 gg of protein per ml, the proportion of lysozyme in thispreparation was (56 ,g/ml + 44 g/ml)/(O.480 mgofproteinper ml) = 2084yg of lysozyme per mg of protein. This propor-
1500w
1o
-5 1
5 10 15 20 25 30 35 40 45 78FRACTON NO.
20 B
15 ~~~~CG
-J5
0 5 10 15 20 25 30 35 40 45 78
FRACTON NO.
FIG. 3. Electrophoretic separation of the "chick-type" and"goose-type" lysozymes present in chicken leukocyte granules (A)or Embden goose bone marrow (B). Fractionation of -the gelwas begun at the position in the gel to which the cationic trackingdye migrated (tube no. 1). Peaks C and G contained the "chick-type" and "goose-type" lysozymes, respectively. The lysozymeconcentration is expressed in ug/ml, as determined by lysoplateassays using, wherever appropriate, purified chicken egg-whitelysozyme or Embden goose egg-white lysozyme as a standard.
tion of lysozyme makes it seem unlikely that the enzyme wedemonstrated in these large granules could merely have beenabsorbed to them as a preparative artifact. Rather, the ob-servation tends to support the hypothesis, which awaits morerigorous proof, that the lysozymes detected were intrinsicto the large granules.
Electrophoretic Separation of the Leukocyte Granule Lyso-zymes. Two distinct peaks of lysozyme activity were detectedin acrylamide gels after electrophoresis of the granule frac-tion at pH 4.5 (Fig. 3A). The more basic peak (C) contained"chick-type " but not "goose-type " lysozyme, as judgedby antibody absorption experiments. The other peak (G)contained only the "goose-type" enzyme. The recovery ofthe "chick-type" and "goose-type" lysozymes from the gelwas 90% and 70%, respectively.
Studies on Embden Goose Bone Marrow. A bone-marrowextract from the Embden goose was also found to containboth a "chick-type" and a "goose-type" lysozyme. Fig. 4shows the results of an antibodyabsorption experiment. Thesupernatant from a mixture of the bone-marrow extract andantibody against goose egg-white lysozyme was found tocontain a lysozyme that was inhibited in its diffusion in lyso-plates only by antibody against chicken egg-white lysozyme(Fig 4-1). The supernatant from a mixture of the marrowextract and antibody against chicken egg-white lysozymealso contained an enzyme inhibited in its diffusion only byantibody against goose egg-white lysozyme (Fig. 4-2). Quan-titative estimates of the amount of each lysozyme presentin the marrow extract were also made. It contained 84 tg/mlof "chick-type" and 100ug/ml of "goose-type" lysozyme.
Isozymes of Lysozyme 1655

1656 Biochemistry: Hindenburg et al.
FIG. 4. Lysoplate analysis of goose bone-marrow lysozymesafter absorption with antibodies against lysozyme. M + AC andM + AG symbolize the sqpernatant obtained, after centrifuga-tion, from a mixture of the marrow extract (M) and antibodyagainst chicken egg-white lysozyme (AO) and antibody againstEmbden goose egg-white lysozyme (AG), respectively.
Two distinct peaks of lysozyme activity were also observedafter acrylamide gel electrophoresis of the marrow extract(Fig. 3B). Peak C contained only the "chick-type" enzyme
and peak G only the "goose-type" lysozyme, as judged byantibody absorption experiments. The recovery of the "chick-type" and "goose-type" lysozymes from this gel was 67%and 60%, respectively.
DISCUSSION
The observation that the egg whites of the chicken and Pekingduck appear to contain only a "chick-type" lysozyme, thatof the Embden goose only a "goose-type" lysozyme, and thoseof the black swan and Canada goose both types of enzymemay be explained in either of two ways. The absence of a
particular lysozyme in egg white could reflect the absenceof the structural gene for that enzyme in the genome of thespecies. Alternatively, the structural genes for both formsof lysozyme may exist in each species and the presence ofone or the other or both lysozymes in egg white would reflectthe action of a species-specific regulatory mechanism.Our data on granule-associated lysozymes of chicken PMN
and Embden goose bone-marrow lysozymes clearly supportthe latter hypothesis. Additional confirmation comes fromdata on the Peking duck. This species, which has "chick-type" lysozymes in its egg white (summarized in ref. 12),appears to have a "goose-type" enzyme in other tissues (7).The black swan (1, 2, 6) and Canada goose (7), on the otherhand, have both types of lysozyme in their egg white.
It would appear, therefore, that the presence of either or
both forms of lysozyme in the egg white of a particular speciesdepends upon a species-specific regulatory mechanism. In-formation on the development of these species-specific mech-
anisms during the evolution of these birds from their commonancestors will require information on the molecular basis ofthe control of egg-white lysozyme synthesis. Further insightsinto the mechanism of control may come from studies onthe hormonal induction of egg-white protein synthesis in someof these species. Experiments on chicken oviducts have al-ready provided a good deal of information on the hormonalregulation of the synthesis of egg-white proteins. Synthesisof chicken egg-white ovalbumin, ovomucoid, conalbumin,avidin, and lysozyme seems to be initiated in the oviductafter hormonal stimulation (13-15). Ovalbumin and avidinsynthesis has been shown to be a consequence of an increasedsynthesis of their respective messenger RNAs (16-18). It islikely, therefore, that the species specificity of egg-whitelysozyme content is controlled by species-specific regulatorymechanisms acting at the level of transcription or perhapstranslation. Other mechanisms that may affect the presenceof these lysozymes in egg white are less probable but cannotbe rigorously excluded until further experiments are carriedout.
Regardless of the exact molecular mechanisms by which theegg-white lysozyme content of the different specieswas changedduring evolution, the question remains as to what the evolu-tionary advantage was in altering the types of egg-whitelysozyme present in different species. One possibility is re-lated to the function of these enzymes. It seems likely thatlysozyme has an antibacterial role in egg white, althoughadditional functions have been proposed for the enzyme inother tissues (19, 20). Studies on the "goose-type" and "chick-type" enzymes have shown that they are muramidases andbreak down the bacterial cell-wall peptidoglycan by hydrolyz-ing the j3(1, 4) linkage between N-acetylmuramic acid andN-acetylglucosamine (21, 22). The two forms of lysozyme,however, also differ in other catalytic properties (1, 3, 4,22-27). The "goose-type" enzyme has a 3- to 6-fold higherspecific activity and lacks the chitinase activity that is char-acteristic of the "chick-type" enzyme. It has also been sug-gested that the "goose-type" enzyme may have a strongpreference for those N-acetylmuramic-acid residues in thebacterial cell wall that are substituted with a peptide (22).It may be relevant that different species of bacteria varyin the degree of cross-linking and amino-acid compositionof the peptide portion of the peptidoglycan (28 29).A second alternative excludes any direct effect of natural
selection on the presence or absence of each lysozyme typebecause of the enzymatic properties of these proteins. It ispossible that during evolution natural selection acted to affectthe appearance of many different gene products in the eggwhites of different species. The presence or absence of eitherform of lysozyme might therefore be a fortuitous consequenceof such an event based, for example, upon a close linkage toother genetic loci.The presence of lysozyme activity in chicken PMN leuko-
cytes deserves comment from the point of view of the anti-bacterial function of these cells. Brune and Spitznagel (9)found two electrophoretically distinct forms of lysozyme inlarge cytoplasmic granules of these cells. We have identifiedthem as "chick-type" and "goose-type" enzymes. Bruneand Spitznagel (9) also observed that these cells lack theenzyme myeloperoxidase, which serves an important anti-bacterial function in mammalian leukocytes. The absenceof this enzyme in chicken PMN leukocytes may be com-
Proc. Nat. Acad. Sci. USA 71 (1974)

Proc. Nat. Acad. Sci. USA 71 (1974)
pensated for by a broader spectrum of antibacterial-activitiesproduced by the different catalytic properties of the "goose-type" and "chick-type" lysozymes. A recent report on humanPMN leukocytes indicates that, in addition to the one mur-
amidase that has been found in those cells, a proteolytic en-
zyme (elastase), which is able to digest the cell walls of somebacterial species, is also present (30). Further studies on avianand mammalian leukocytes will perhaps provide more in-formation on the antibacterial agents of these cells and theirorganelles.
Finally, it will be especially interesting to investigate thelysozyme content of avian macrophages in light of a recentproposal that lysozyme may be a mediator of the antitumorfunctions of macrophages in mammals (20).
This work was supported by N.S.F. Grant B036007 (N.A.),N.I.H. Grant 5-RoI-AI0243 (J.S.), and A.E.C. Grant AT-(40-1)-3628 (J.S.).
1. Morgan, F. J. & Arnheim, N. (1973) in Lysozyme, eds.Osserman, E. F., Canfield, R. E. & Beychok, S. (AcademicPress, New York), in press.
2. Arnheim, N., Hindenburg, A., Begg, G. S. & Morgan, F. J.(1973) J. Biol. Chem. 248, 8036-8042.
3. Canfield, R. E. & McMurry, S. (1967) Biochem. Biophys.Res. Commun. 26, 38.
4. Dianoux, A. C. & Jollbs, P. (1967) Biochim. Biophys. Acta133, 472-479.
5. Canfield, R. E., Kammermann, S., Sobel, J. H. & Morgan,F. J. (1971) Nature New Biol. 232, 16-17.
6. Arnheim, N. & Steller, R. (1970) Arch. Biochem. Biophys.141, 656-661.
7. Arnheim, N. (1973) in Lysozyme, eds. Osserman, E. F.,Canfield, R. E. & Beychok, S. (Academic Press, New York),in press.
8. Osserman, E. & Lawlor, D. P. (1966) J. Exp. Med. 124,921-952.
9. Brune, K. & Spitznagel, J. (1973) J. Infec. Dis. 1-27, 84-94.10. BeBrwer, J. M. & Ashworth, R. B. (1969) J. Chem. Educ. 46,
41-45.11. Delacour, J. (1954) "The Waterfowl of the World," Country
Life, London, Vol. I.12. Prager, E. M. & Wilson, A. C. (1972) Biochem. Genet. 7,
269-272.13. O'Malley, B. W., McGuire, W. L., Kohler, P. O. & Koren-
man, S. G. (1969) Recent Progr. Horm. Res. 25, 105-160.14. Palmiter, R. D. (1972) J. Biol. Chem. 247, 6450-6459.15. Oka, R. & Schimke, R. T. (1969) J. Cell Biol. 43, 123-127.16. Chan, L., Means, A. R., & O'Malley, B. W. (1973) Proc.
Nat. Acad. Sci. USA 70, 1870-1874.17. Means, A. R., Comstock, J. P., Rosenfeld, G. C. & O'Malley,
B. W. (1972) Proc. Nat. Acad. Sci. USA 69, 1146-1150.18. Rhodes, R. E., McKnight, G. S. & Schimke, R. T. (1973)
J. Biol. Chem. 248, 2031-2039.19. Osserman, E. F., Canfield, R. E. & Beychok, S., eds. (1973)
Lysozyme (Academic Press, New York), in press.20. Osserman, E. F., Klockars, M., Halper, J. & Fischel, R. E.
(1973) Nature 243, 331-335.21. Imoto, T., Johnson, L. N., North, A. C. T., -Phillips, D. C. &
Rupley, J. A. (1972) in The Enzymes, ed. Boyer, P. D.(Academic Press, New York), 3rd ed., pp. 665-868.
22. Arnheim, N., Inouye, M., Law, L. & Laudin, A. (1973) J.Biol. Chem. 248, 233-236.
23. McKelvy, J. F., Eshdat, Y. & Sharon, N. (1970) Fed. Proc.29, 532.
24. Jollbs, P., Saint-Blancard, J., Charlemagne, D., Dianoux,A.-C., Jolles, J. & LeBaron, J. L. (1968) Biochim. Biophys.Ada 151, 532-534.
25. Dianoux, A. C. & Jollls, P. (196) Helv. Chim. Acta 52, 611-616.
26. Charlemagne, D. & Jollbs, P. (1967) Bull. Soc. Chim. Biol.49, 1103-1113.
27. Dianoux, A. C. & Jollbs, P. (1969) Bull. Soc. Chim. Biol. 51,1559-1564.
28. Osborn, M. J. (1969) Annu. Rev. Biochem. 38, 501-538.29. Ghuysen, J. M. (1968) Bacteriol. Rev. 32, 425-464.30. Janoff, A. & Blondin, J. (1973) Lab. Invest. 29, 454-457.
Isozymes of Lysozyme 1657