changes in orientation of attritional wear facets with...
TRANSCRIPT

Changes in Orientation of Attritional Wear Facets WithImplications for Jaw Motion in a Mixed LongitudinalSample of Propithecus edwardsi From RanomafanaNational Park, Madagascar
Stephanie Blatch,1* Doug M. Boyer,1,2 Stephen J. King,3 Jonathan M. Bunn,1,4
Jukka Jernvall,4,5 and Patricia C. Wright5,6
1Interdepartmental Doctoral Program in Anthropological Sciences, Stony Brook University,Stony Brook, NY 11794-43642Department of Anthropology and Archaeology, Brooklyn College, City University of New York, Brooklyn, NY 112103Department of Anthropology, University of Massachusetts. Amherst, MA 010034Developmental Biology program, Institute of Biotechnology, Viikinkaari 9, 00014 University of Helsinki, Helsinki,Finland5Department of Ecology and Evolution, Stony Brook University, Stony Brook, NY 11794-52456Department of Anthropology, Stony Brook University, Stony Brook, NY 11794-4364
KEY WORDS molar wear; power stroke; mastication; ontogeny; senescence
ABSTRACT In many mammalian species, the pro-gressive wearing down of the teeth that occurs over anindividual’s lifetime has the potential to change dentalfunction, jaw movements, or even feeding habits. Theorientation of phase-I wear facets on molars reveals thedirection of jaw movement during the power stroke ofmastication. We investigated if and how molar wear fac-ets change with increasing wear and/or age by examin-ing a mixed longitudinal dataset of mandibular toothmolds from wild Propithecus edwardsi (N 5 32 individu-als, 86 samples). Measurements of the verticality of wearfacets were obtained from three-dimensional digital mod-els generated from lCT scans. Results show that verti-cality decreases over the lifetime of P. edwardsi, achange that implies an increasingly lateral translation ofthe jaw as the teeth move into occlusion. A more trans-
verse phase-I power stroke supports the hypothesis thatthese animals chew to maximize longevity and function-ality of their teeth, minimizing the ‘‘waste’’ of enamel,while maintaining sharp shearing crests. Results of thisstudy indicate that wear facet verticality is more closelycorrelated with age than overall amount of tooth wear,measured as area of exposed dentin, suggestingthat age-related changes in cranial morphology may bemore responsible for adjustments in jaw motion over thelifetimes of Propithecus than wear-related changesin the shape of occluding teeth. Finally, the rate ofdecrease in wear facet verticality with age is greaterin males than in females suggesting differences indevelopment and/or access to resources between thesexes in this species. Am J Phys Anthropol 000:000–000,2011. VVC 2011 Wiley-Liss, Inc.
Tooth shape affects an animal’s ability to break downfood and the concomitant force necessary to do so (Evansand Sanson, 1998, 2006). Therefore, tooth shape has adirect impact on what types of food an animal is able toprocess and the nutrition it will be able to extract fromthose foods. Teeth have been analogized to tools that aredifferently specialized to particular dietary pressures asan adaptation to optimize feeding efficiency (e.g., Lucas,1982; Lucas and Luke, 1983; Evans and Sanson, 2003).Studies have tended to focus on the links between molarmorphology and dietary preference in living species (e.g.,Rosenberger and Kinzey, 1976; Kay, 1978; Seligsohn andSzalay, 1978) and have often applied the findings to inferdiet in fossil taxa (e.g., Kay, 1977; Anthony and Kay,1993; Meldrum and Kay, 1997).These earlier studies of primate tooth function focused
on the morphology of unworn specimens. Most dentalspecimens available for study are worn to some degree,however, which limits the applicability of this work. Therate of tooth wear and changes in tooth shape thataccompany wear over the course of a mammal’s lifeinfluence which foods the animal is able to process andhow long it is able to efficiently extract nutrients from
its diet with advancing age (e.g., Lanyon and Sanson,1986; Skogland, 1988; Kojola et al., 1998; Ozaki et al.,2010). Therefore, it has been argued that the process oftooth wear should also be under natural selection, favor-ing adaptations that maintain mechanical efficiency forprocessing food (e.g., Teaford, 1983; M’Kirera and Ungar,2003; Ungar and M’Kirera, 2003; Dennis et al., 2004).Hence, patterns of wear on teeth have the capacity toreveal a great deal of information about feeding dynam-
Grant sponsors: Evolving Earth, NSF GRFP, Academy of Finland,National Science Foundation; Grant numbers: BCS-0622544, BCS-0721233.
*Correspondence to: Stephanie Blatch, Department of Anthropol-ogy, Stony Brook University, Stony Brook, NY 11794-4364, USA.E-mail: [email protected]
Received 19 August 2010; accepted 3 May 2011
DOI 10.1002/ajpa.21565Published online in Wiley Online Library
(wileyonlinelibrary.com).
VVC 2011 WILEY-LISS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 000:000–000 (2011)

ics and their associated evolutionary pressures. As sug-gested by Ungar (2002), further quantitative analysis ontooth wear in primates needs to be performed to unlockthis potential. Recently, studies have begun to assesshow wear affects overall tooth shape and the implica-tions of species-specific changes with wear (Jernvall andSelanne, 1999; Ungar and Williamson, 2000; M’Kireraand Ungar, 2003; Ungar and M’Kirera, 2003; Dennis etal., 2004; Ulhaas et al., 2004; King et al., 2005). Thesestudies have mainly examined changes in overall toothshape by considering the molar surface as a topographi-cal landscape and examining topographic features suchas relief or average slope. Observations of species-specificchanges in tooth morphology with increasing wear havebeen primarily based on sequences of variably worn mu-seum specimens with the implicit assumption that indi-viduals of the same species will wear their teeth in simi-lar ways. To address this assumption, Dennis et al.(2004) used topographical analysis to examine changesin molar morphology in a longitudinal sample from awild population of Alouatta palliata. In that sample,molar crown slope and occlusal relief decreased overtime while crown angularity (average change of slopeover a surface) remained constant. These findings sug-gest that members of this species do follow species-spe-cific patterns of wear for changes in overall molar shapeover time and, therefore, that techniques of topographi-cal analysis may be useful in comparing nonlongitudinalsamples of variably worn teeth. However, even with theexpanding analysis of overall changes in shape of pri-mate molars with increasing wear, there has been littledirect examination of shape and orientation of specificwear patterns and how they change with advancingwear, especially in natural populations.As outlined by Kay and Hiiemae (1974a, 1974b), tooth
wear is generally acknowledged to result from two differ-ent types of masticatory behavior. In the first kind,puncture-crushing, food is softened via several ‘‘puncturecrushing strokes’’ during which there is tooth–substratecontact but no actual tooth–tooth contact. This type ofmasticatory behavior produces a kind of tooth wearknown as abrasion, which consists of blunting and ulti-mately the creation of pits in the tips of the cusps. Thesecond category of masticatory behavior is known simplyas chewing. Chewing produces a kind of wear known asattrition, which results in wear facets. It is the wear fac-ets created by attrition that are the focus of this study.Specifically, chewing refers to jaw movement when
food is already significantly processed to allow for tooth–tooth contact. This tooth–tooth contact forms attritionalwear facets during the power-stoke of the chewing cycle.In his study of Perissodactyla teeth, Butler (1952) dis-cussed the corresponding wear facets on upper and lowerteeth formed by attrition and noted that facets indicatethe orientation of the jaw motion that produced them.The correlation between the direction of jaw movementduring the chewing stroke and wear facet morphologywas later further supported for living selenodont artio-dactyls (Greaves, 1973) and confirmed through the ex-perimental production of wear facets on artificial teeth(Costa and Greaves, 1981). Additionally, investigations ofprimate wear facets have also supported the idea thatmovement between upper and lower molars as theycome into contact is guided by crown morphology (e.g.,Kay and Hiiemae, 1974a; Mills, 1978). Because attritionis the result of moving the upper and lower teeth acrosseach other in a habitual pattern producing repetitive
localized wear, the orientation of attritional facets shouldcorrelate to the direction of jaw movement during thepower stroke (Butler, 1952, 1972; Kay and Hiiemae,1974a). Facet orientation should therefore provide someinformation on how the animal is using the jaw to pro-cess food during chewing and may vary depending onwhether the diet of the animal contains foods thatrequire a more vertical shearing stroke or a more hori-zontal grinding stroke during processing. Comparison ofwear facets among mammal taxa suggests that moresteeply angled facets accompany an insectivorous dietwhile shallower facets are associated with frugivory andomnivory (Butler, 1972; Kay and Hiiemae, 1974a).Patterns of tooth wear alter the shape of occlusal
surfaces. In turn, jaw movement is mutually influentialwith crown morphology, helping to guide the motion ofone tooth across another during chewing and with repet-itive jaw motions, eventually creating attritional facetson the occlusal surfaces that correspond to the orienta-tion of jaw motion. Animals eating foods with differentmechanical properties use substantially different pat-terns of jaw movement to process those foods (Agrawalet al., 2000; Lucas, 2004). Altering the pattern of jawmovement used for processing a given food type wouldpresumably result in decreased effectiveness in fragmen-tation of that food. Diminished ability to fragment foodeffectively has been shown to result in decreased abilityto extract nutrients from those foods (Lanyon and San-son, 1986). Thus, if senescent animals are modifying thejaw motions they use to process foods, this is likely toaffect their ability to fragment the food and obtainnutrients from it. Here, we examine wear facet orienta-tion as a proxy for the corresponding jaw motion overthe lifetime of Propithecus edwardsi (Milne-Edwards’sifaka) using a mixed longitudinal sample (Huggins andLoesch, 1998) of wild-caught animals.P. edwardsi is an arboreal, female-dominant member of
the family Indriidae (Pochron et al., 2003). Although thisspecies is primarily folivorous and an anatomical folivore(Hill, 1953), its diet is seasonally variable and includes aselection of seeds and fruits (Meyers and Wright, 1993;Hemingway, 1996, 1998). P. edwardsi reach full adultbody mass at approximately 6 years of age, and maximumlifespan of this species in the wild has been estimated asgreater than 30 years (Pochron et al., 2004). It has beenshown that molar teeth in this species maintain functiondespite tooth wear, even with advancing wear, by exposingcompensatory shearing blades until the onset of dental se-nescence at approximately 18 years of age (King et al.,2005). When dental senescence is reached, the occlusalsurface is essentially a dentin basin encircled by a band ofenamel. Despite diminished shearing capacity at thisstage of wear, King et al. (2005) showed that individualsof advanced age are able to survive. However, they areclearly hindered by compromised feeding ability as theirreproductive success becomes severely environmentallymitigated (in arid years they are unable to successfullyrear an infant). Because dentally senescent individualsare no longer able to shear foliage with the same effi-ciency, it seems that they are using an alternate strategyfor obtaining the nutrition necessary to support reproduc-tion. One such possible strategy would involve boostingthe efficiency of their food processing by adding anincreased horizontal component, or grinding motion, totheir chewing to partially compensate for the decreasingfunctionality of dentition at dental senescence. By theonset of dental senescence, the occlusal surface of the
2 S. BLATCH ET AL.
American Journal of Physical Anthropology

tooth is fairly flat. A more horizontal chewing motion maybe able to make better use of the flatter tooth morphologyto break down food. Specifically, we hypothesize that olderanimals depend more on milling to process food once theirshearing crests have been worn down. It is therefore pre-dicted that a change in jaw movement and wear facet ori-entation to incorporate a more horizontal component offood grinding should coincide with the onset of dental se-nescence.Given that chewing efficiency declines at dental senes-
cence, it is beneficial for animals to delay this event, if pos-sible. Seligsohn (1977) presented a biomechanical modelfor changes in chewing patterns that maximizes toothfunctionality and longevity. He showed that, to keepshearing crests sharp while wasting as little enamel aspossible (thereby extending the life of the tooth), thephase-I of the power stroke should become increasinglytransverse. In his study, he measured these facets in theupper molars, using cross-sectional samples of mostlemuriform species. He found changes in Lepilemur alone.However, the study of Seligsohn (1977) was done 1) with-out controlling for individual differences in wear facet ori-entation (i.e., it was not a longitudinal study), 2) withoutcontrolling for populational differences in wear rates andjaw motion (i.e., different populations may exhibit suchdifferences simply due to idiosyncrasies of food availabil-ity in their particular home range), and 3) using methodshaving much lower levels of precision than those affordedby the methods of this study (see below).
MATERIALS AND METHODS
Study animals
We examined worn molars of P. edwardsi from Ranoma-fana National Park in Madagascar where data collectionon this species was initiated in 1986 and has continued fornearly 25 years (e.g., Wright, 1995; 1998; Dew and Wright,1998; Pochron et al., 2002, 2003, 2004, 2005).In the long-term study of P. edwardsi at Ranomafana
National Park, study animals are habituated to humanobservation and each animal has been fitted with acolor-coded collar to allow for identification of all individ-uals from a distance. Some animals in the populationhave been born into the study group since the studybegan and therefore have exact known ages. For allother individuals, age has been estimated based on stageof development at first sighting and amount of overalltooth wear. Study animals were periodically anestheti-cally darted (following Glander et al., 1991), duringwhich time dental impressions of right mandibular toothrows were obtained from captured animals beforerelease. Specifics of this study site along with demo-graphics of the animal groups, life history information,habitat descriptions, and animal handling procedureshave been described in detail elsewhere (Wright, 1995;Pochron et al., 2004; King et al., 2005). Dental molds col-lected during field seasons from 1993 to 2008 represent-ing 86 moldings from a total of 32 individuals wereavailable to us for this study. Not every individual couldbe molded every season and numbers of molds per indi-vidual vary from one to seven instances.
Specimen preparation
Techniques of casting, lCT scanning, and digital surfacereconstruction follow Boyer (2008). To create high-resolu-tion casts from the molds taken in the field, grey-tinted
EPO-TEK 301 epoxy was poured into the molds and thencentrifuged to eliminate bubbles that can obscure occlusalmorphology. Once hardened, lower second molars werecut out of the tooth rows and mounted in standardized ori-entations on foam disks of 36 mm diameter in preparationfor scanning. A ScancoMedical brand lCT 40 machine(http://www.scanco.ch) was used to scan 10–16 teeth atonce with an 18 lm resolution. This scanning techniqueyielded segmented stacks of two-dimensional (2D) slicesroughly parallel with the plane of occlusion, which weresaved in a DICOM format for further processing.
Digital surface reconstruction
The stacked slices of individual teeth were digitallycropped from the scan disks using ImageJ. To createmeasurable surface images, the resulting segmentedDICOM files of individual teeth were opened using thesoftware Avizo (http://www.vsg3d.com). Avizo was usedto evaluate the threshold between epoxy and air in thescans and to reconstruct three-dimensional (3D) digitalsurface images suitable for measurement. As the recon-structed surfaces from segmented files produce unrealis-tically stepped morphology in Avizo, the tooth surfaceswere smoothed using 20 iterations of the ‘‘surfacesmooth’’ function prior to measurement.
Data collection/measurement protocols
Orientation of isolated teeth. The most critical stepprior to measurement of wear facet angle was orienta-tion of teeth in 3D space. To obtain a consistent, compa-rable orientation for every tooth, all surface files werevisualized simultaneously using Avizo. They were theniteratively adjusted to the same orientation in three or-thogonal views (occlusal, lingual, and distal/coronal)until corresponding profiles of unworn surfaces in eachview matched as closely as possible for all teeth (Fig. 1).Additionally, three further conditions were met as wellas possible: 1) the chord connecting the metaconid andentoconid was aligned with the mesiodistal axis in occlu-sal view; 2) the unworn buccal and lingual aspects of thetrigonid formed the same/similar angles with respect tothe parasagittal axis in mesial (coronal) view; 3) the dis-tal and mesial halves of the entoconid notch formed asimilar/the same angle with the dorsoventral axis in lin-gual view. The advantage of this technique is that itallows for the consideration and comparison of allunworn morphologies of each tooth while ensuring thatunusual variation in any one region of the tooth (due tomold distortion, bubbles, breakage, dirt or saliva onparts of the tooth) does not affect its overall alignment.For the oriented teeth, the XY plane approximates theocclusal plane as closely as possible while buccal and dis-tal orthogonal views approximate coronal and parasagit-tal planes, respectively. The measurements on each toothshould therefore be considered as oriented with respectto the occlusal plane.
Wear facet selection and measurement. We usedaspects of the orientation of two wear facets (those onthe paracristid and cristid obliqua) formed during phase-I of the power stroke of mastication to infer the directionof motion of the lower jaw with respect to the upper jaw.For as many of the teeth in our sample (Table 1) as pos-sible, we also used the wear facet formed on the post-vallid surface of the trigonid that often connects the par-acristid and cristid obliqua wear facets (we call this the
3WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

‘‘post-paracristid facet’’; see Fig. 2). The central, planarparts of all identifiable facets of interest were croppedfrom the tooth surface. Each facet was saved as a sepa-rate surface file (*.surf in ascii format in Avizo). Thecoordinate data were extracted from the surf files usingthe program notepad and imported into Microsoft excel.Excel worksheets were opened in SPSS. SPSS was usedto determine the least squares plane modeling the facetof the form (z) 5 B1(x) 1 B2(y) 1 C where B1, B2, and Care constants. We also recorded the adjusted correlationcoefficient of each plane, as a check on the quality ofcoordinate data. Each wear facet was usually repre-
sented by between 200 and 500 points for the regressionequation and r2 values for least squares planes fit tothem ranged from 0.892 to 0.998.It has long been recognized that the vector of intersec-
tion of two nonparallel facets formed by the same phaseof the power stroke of mastication would be parallel tothe direction of jaw motion relative to the teeth (Butler,1972; Kay and Hiiemae, 1974b). Because the post-para-cristid facet is significantly different in orientation fromthose on the paracristid and cristid obliqua (which them-selves are similarly oriented), and because it is formedby the same phase of the power stroke, whenever it
Fig. 1. Method for orienting teeth in three-dimensional space. The program Avizo was used to manually orient digital models ofteeth to predefined set of anatomical axes corresponding to the x-, y-, and z-axes of the coordinate space (see Materials and Methodstext). Four large boxes represent four initial steps in orientation process. Three rows of teeth represent three components of eachstep. The box surrounding each tooth represents its local coordinate axes. The same four teeth are depicted for each step of the pro-cess. Note that a key component of this procedure is the standardization of orientation of each tooth by comparing it to essentiallyevery other tooth in the sample at each step of the process. A: Beginning point in orientation processes: tooth models are unorien-tated. Note that local coordinate axes are parallel among different teeth, meaning the local axes are aligned with global axes. B:Local axes are rotated so that anatomical axes of teeth match better in global space. C: Local axes and coordinates of teeth are resetto global axes. After orientation in occlusal view (x–y plane) is complete, the teeth are viewed at 908 to this [x–z (step 2) or y–z plane(step 3)]. Parts A–C of the procedure are repeated each time. Once three orthogonal views have been adjusted, original view (step4) is checked again. Rotations in perpendicular planes may have changed the perspective on the x–y plane requiring further adjust-ment. The x–z plane is then checked again, and so on until orientation in all views is consistent and stable. This typically takes sixto nine steps (i.e., two to five beyond the four depicted).
4 S. BLATCH ET AL.
American Journal of Physical Anthropology

could be identified we were able to use it in combinationwith the paracristid and/or cristid obliqua to determinethe exact direction of jaw motion while the teeth were incontact during phase-I of the power stroke of mastica-tion. The coefficients on the x, y, and z variables of theleast squares plane also describe a vector that is perpen-dicular to the plane (the pole of the plane). By takingthe cross-product of the poles of the least squares planeof each wear facet, we were able to determine the vectorof intersection of the two wear facets (Fig. 2). Finally, wedetermined the coordinates for two points on this vectorof intersection to assist in calculation of other valueslater. We found the first common point by rewriting theequations for each plane in the form (21)C 5 B1(x) 1B2(y) 1 B3(z), in which B3 is simply 21. For the firstpoint we set x 5 0 and then solved the system of tworesulting equations [(21)C 5 B2(y) 1 B3(z)] in terms ofy. We then input the value we determined for y andsolved for z. We found the second point by adding thevector entries of the cross-product to the coordinates ofthe first point. Using the coordinates of these two pointswe were able to use trigonometry to describe specificcomponents of jaw motion in terms of its angle from ana-tomical planes and axes.We determined two components of the motion
described by the vector of intersection between wear fac-ets: 1) the orientation of that vector in the occlusal plane(in other words, the path the lower tooth takes as itapproaches the upper tooth from a more buccal positionand then proceeds to a more lingual position). We referto this component as the transverse component of thejaw motion; and 2) the angle by which that vector devi-
ates from the occlusal plane, or the verticality of the vec-tor. This value could theoretically range from 08 (5 par-allel to occlusal) to 908 (5 perpendicular to occlusal). Inthe latter case, the orientation of the transverse compo-nent described above would technically be indefinable.For the transverse component, we hypothesized that thepath would be roughly parallel to the buccolingual axis(i.e., it would form an angle of 08 to the buccolingualaxis) based on studies by Butler (1972) and Kay andHiiemae (1974b). We found instead that for teeth inwhich the post-paracristid facet was observable andquantifiable, the angle with respect to the paracristidaveraged 7.268 from parallel to buccolingual (N 5 25;standard deviation 5 4.878), meaning the tooth pro-gressed mesially (anteriorly), as it moved lingually withrespect to the upper teeth. The cristid obliqua showed aslightly (although not significantly) different path (N 525; x 5 6.458; standard deviation 5 4.648). We found nosignificant correlation between this value and tooth age(i.e., age of individual when tooth was molded) in oursample (paracristid: Spearman’s rank correlation coeffi-cient 5 20.24, P 5 0.241; cristid obliqua: Spearman’srank correlation coefficient 5 20.18, P 5 0.386).Post-paracristid facets could not be identified for more
than two-thirds of teeth in the sample, meaning that thetrue vector of interest could not be explicitly calculated.Therefore, we assumed that this vector would have atransverse direction that matched the average of thosein which it was actually determinable (7.268 for paracris-tid and 6.458 for cristid obliqua). We tested what effectthis would have on the verticality measures by usingthis technique on 26 teeth for which we had been able to
TABLE 1. Angular directions of two components (transverse and vertical) of phase-I of the power stroke for all teeth in which itcould be calculated using intersection of post-paracristid with other facets
Tooth
Paracristid Cristid obliqua
Transverse Vertical Vertical (7.26) Transverse Vertical Vertical (6.45)
57-2002 3.76 31.91 29.29 7.15 34.83 35.3864-2006 25.42 33.44 25.09 25.80 32.94 24.0562-2008 23.13 33.13 25.91 0.75 36.22 31.9126-1994 0.96 34.55 30.30 0.47 34.01 30.042-1995 6.83 26.55 26.29 7.44 27.22 27.9860-2002 7.98 33.74 34.44 12.62 38.07 42.7526-1995 6.10 35.65 34.71 5.77 35.32 34.8529-2002 6.78 30.52 30.15 14.98 35.93 41.565-1993 8.06 23.57 24.23 na na na21-1995 8.44 35.35 36.58 5.96 33.10 32.7042-2002 4.92 38.36 37.03 1.94 35.34 32.1441-2002 6.60 23.96 23.61 4.55 21.95 20.9345-2002 8.28 29.08 30.05 7.90 28.75 29.9342-2006 10.94 20.13 22.70 11.80 21.05 24.8731-1995 14.08 33.62 39.00 3.52 22.71 20.4931-2002 5.69 25.60 24.46 7.98 27.57 28.8066-2006 10.47 26.05 29.33 9.21 24.85 27.3466-2004 13.83 38.05 46.12 11.00 36.06 39.9114-1995 5.46 29.32 27.60 3.84 27.63 24.6636-2003 6.99 30.46 30.26 5.69 28.92 28.4222-1994 12.88 35.33 40.31 5.81 29.45 29.0736-2004 9.88 31.16 32.98 6.03 27.05 26.8236-2005 6.64 29.65 29.19 7.01 30.00 30.4336-2006 na na na 4.54 38.81 37.5636-2008 8.26 28.99 29.58 5.71 26.67 26.2075-2008 16.16 21.50 30.29 15.34 20.96 29.63Mean 7.26 30.39 30.78 6.45 30.22 30.34Standard deviation 4.87 4.96 5.76 4.55 5.35 5.73
‘‘Vertical (7.26)’’ and ‘‘Vertical (6.45)’’ give the vertical component of the power stroke calculated if the transverse component isassumed to be 7.268 and 6.458, respectively.
5WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

calculate the exact orientation of the motion vector dueto a visible post-paracristid facet (Table 1). Comparingthe approximated verticality to actual verticalityrevealed that there was no significant difference in aver-age angle formed (paracristid paired t-test: t 5 0.50, P 50.622; cristid obliqua paired t-test: t 5 0.168, P 5 0.868).Furthermore, we checked how this affected the correla-tion between tooth age and verticality. For the true verti-cality of the paracristid in this subset, we found a signif-icant correlation with age (Spearman’s rank correlationcoefficient 5 20.48, P 5 0.016). Likewise when we usedthe verticality measures in the average transverse direc-tion of 7.268, the results were similar (Spearman’s rankcorrelation coefficient 5 20.56, P 5 0.004). The same
test for the cristid obliqua resulted in nonsignificancewhen using the true angle (Spearman’s rank correlationcoefficient 5 20.17, P 5 0.42); this remained nonsignifi-cant when using verticality in the average transversedirection (Spearman’s rank correlation coefficient 520.20, P 5 0.33). We can think of no reason why usingthe average angle of transverse motion (essentially add-ing a component of random error to our verticality meas-ures) should improve the correlation between verticalityand age; therefore, we assume the fact that probabilitiesof no correlation become lower using average angles inthese particular cases was coincidental. Thus the mostimportant revelation of this test is that in the case of theparacristid, the correlation coefficients were similar and
Fig. 2. Determination of direction of horizontal (transverse) component of jaw motion during phase-I of the power stroke of mas-tication revealed by vector of intersection of two phase-I wear facets. Verticality of wear facets relative to occlusal plane must bemeasured parallel to this vector of intersection. A: Original tooth scan in occlusal view. B: Tooth scan shown transparent with facetshighlighted. C: ‘‘ppc’’ facet translated to intersect PC and CO facets. D: Inset of C with dashed line showing orientation of line ofintersection of two sets of facets. Note they are almost parallel to each other and to the buccolingual axis of the tooth, which is per-pendicular to a line containing the metaconid and entoconid facets of the tooth in occlusal view. In this particular specimen, theline indicates the tooth moved slightly distally (retraction) as it moved lingually relative to the upper teeth during phase-I of thepower stroke. However, on average, the horizontal component of the teeth we measured suggests slight (7.268 for paracristid, 6.458degrees for cristid obliqua) mesial translation (protraction) during phase I. Abbreviations: CO, cristid obliqua; PC, paracristid; PPC,post-paracristid. Scale bars 5 1 mm.
6 S. BLATCH ET AL.
American Journal of Physical Anthropology

relationships were significant using true and averagetransverse directions, while both methods produced non-significant results for the cristid obliqua.Therefore, we used the average angles found for the
transverse components (7.268 and 6.458) to calculate thevertical components of wear facets on the remainingteeth and assume that it will not significantly changethe average verticality angles determined or the patternof statistical significance in our subsequent results (seeAppendix Tables A1 and A2 for all regression parame-ters, correlation coefficients, common points, and com-puted angles).Additionally, for each tooth, data were collected on
total 2D dentin exposure. To collect dentin exposurearea, occlusal views were exported from Avizo as JPEGimages. Total area of dentin exposure per tooth wasmeasured using Sigma Scan Pro 5.0. We used 2D meas-ures instead of 3D measures here because 1) the exposedareas of dentin are typically quite flat and we do notbelieve we are losing much information and 2) we foundit to be impractical to collect accurate 3D informationdue to various imperfections in casts and resulting scansof teeth (bubbles, dirt, and saliva added spurious tex-tural information in most cases).
Data analysis
Resulting measurements of the verticality of wearfacet orientation on the paracristid and cristid obliquawere analyzed separately. Due to the non-normal distri-bution of the angular data, nonparametric tests with theleast possible number of assumptions were used to ana-lyze the data whenever possible. Spearman’s rank corre-lation coefficient was used to assess correlation betweenwear facet angle and age. Spearman’s rank correlationcoefficients were also calculated to assess possible corre-lation between wear facet angle and the square roottransformed dentin exposure area for individuals withknown age only. Trends in the change of facet angles forindividual animals were assessed visually and using the
nonparametric sign test for changes over 1-, 2-, 3-, 4-,and 51-year periods for animals with at least threeinstances of capture. Differences in changes of facetangle with age were also assessed by sex by performinga quick test of sex-specific residuals above and below aLOESS line with 50% tension. The distribution of theseresiduals was assessed using a Fisher’s exact test toevaluate association between wear angle and sex ofadult animals before the onset of dental senescence. Ani-mals beyond the age of dental senescence were excludeddue to the fact that only one sex (female) of advancedage is represented in the data.
RESULTS
Wear facet angle and age
For both the paracristid and the cristid obliqua, theangle of verticality of the facet shares a negative rela-tionship with age (Figs. 3–5). The relationship betweenfacet angle and age on the paracristid (Spearman’s rankcorrelation coefficient 5 20.41, P \ 0.001) was moretightly correlated than that on the cristid obliqua (Spear-man’s rank correlation coefficient 5 20.21, P 5 0.055)with the correlation reaching statistical significance atthe 0.05 level for the paracristid facet and approachingsignificance on the cristid obliqua indicating that thereis considerable decrease in facet angle with increasingage (Fig. 3). Visual examination of the trends in individ-ual animals with age (Fig. 4) follows the same pattern ofa generally decreasing facet angle with age, with most ofthe change seemingly taking place early in the animal’slifetime.Despite correlation between age and decreasing verti-
cality of the facets, there is not a monotonic decrease.Between certain intervals, for certain individuals wearfacets become steeper with age. The change of wear facetverticality is plotted as measured over 1-, 2-, 3-, 4-,and 51-year intervals for individual animals (Fig. 5).Over 1-, 2-, and 3-year intervals, sign tests indicate that
Fig. 3. Scatterplot of age versus wear facet angle of all animals measured. On both the paracristid (Spearman’s r 5 20.41, P <0.001) and the cristid obliqua (Spearman’s r 5 20.21, P 5 0.055), facet angle is negatively correlated with age.
7WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology
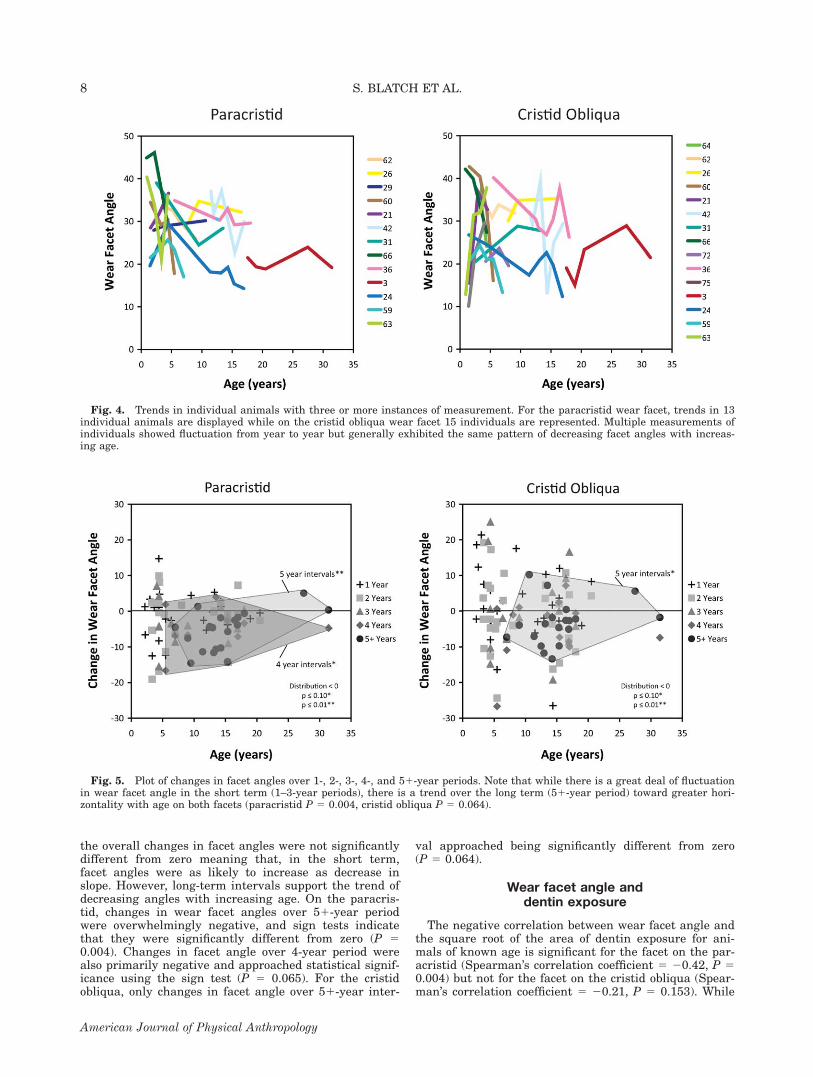
the overall changes in facet angles were not significantlydifferent from zero meaning that, in the short term,facet angles were as likely to increase as decrease inslope. However, long-term intervals support the trend ofdecreasing angles with increasing age. On the paracris-tid, changes in wear facet angles over 51-year periodwere overwhelmingly negative, and sign tests indicatethat they were significantly different from zero (P 50.004). Changes in facet angle over 4-year period werealso primarily negative and approached statistical signif-icance using the sign test (P 5 0.065). For the cristidobliqua, only changes in facet angle over 51-year inter-
val approached being significantly different from zero(P 5 0.064).
Wear facet angle anddentin exposure
The negative correlation between wear facet angle andthe square root of the area of dentin exposure for ani-mals of known age is significant for the facet on the par-acristid (Spearman’s correlation coefficient 5 20.42, P 50.004) but not for the facet on the cristid obliqua (Spear-man’s correlation coefficient 5 20.21, P 5 0.153). While
Fig. 4. Trends in individual animals with three or more instances of measurement. For the paracristid wear facet, trends in 13individual animals are displayed while on the cristid obliqua wear facet 15 individuals are represented. Multiple measurements ofindividuals showed fluctuation from year to year but generally exhibited the same pattern of decreasing facet angles with increas-ing age.
Fig. 5. Plot of changes in facet angles over 1-, 2-, 3-, 4-, and 51-year periods. Note that while there is a great deal of fluctuationin wear facet angle in the short term (1–3-year periods), there is a trend over the long term (51-year period) toward greater hori-zontality with age on both facets (paracristid P 5 0.004, cristid obliqua P 5 0.064).
8 S. BLATCH ET AL.
American Journal of Physical Anthropology

dentin exposure is a component in predicting wear facetangle, it is a less powerful predictor than absolute agefor the same individuals (paracristid Spearman’s correla-tion coefficient 5 20.52, P \ 0.001; cristid obliquaSpearman’s correlation coefficient 5 20.27, P 5 0.063).
Differences in wear facet anglebetween the sexes
Although wear facet angles in juvenile animals appearto be indistinguishable between the sexes, facet anglesin animals that have obtained full adult body mass buthave not yet reached the onset of dental senescenceappear to differ by sex (Fig. 6). On the paracristid, adultfemales maintained significantly steeper facets for theirage compared to males (P 5 0.003, one-tailed Fisher’sexact test). However, no statistical difference betweenthe sexes was found on the cristid obliqua (P 5 0.192,one-tailed Fisher’s exact test).
DISCUSSION
Tempo, mode, and possible mechanismsof wear facet orientation change
The results of this study indicate that there is similar-ity in wear facet development among different P.edwardsi individuals. The negative correlation betweenwear facet angle and age for both the sample populationas a whole and for trends within the lifetimes of individ-ual animals demonstrates the tendency of wear facets tobecome shallower with increasing age. This result is notsurprising. In essence, shallower facets contribute to anoverall flatter occlusal morphology, which one mightexpect for aging teeth with reduced shearing crests andcrown height. This flattening of occlusal morphology anddecrease of overall tooth slope with increasing wear hasbeen noted in several other primate species including
gorillas, chimpanzees (M’Kirera and Ungar, 2003; Ungarand M’Kirera, 2003), orangutans (Ungar and Taylor,2005), howler monkeys (Dennis et al., 2004), western redcolobus, the king colobus, the collared mangabey, andCampbell’ guenon (Bunn and Ungar, 2009). However, nostudy has documented such detailed changes that explic-itly suggest differences in occlusal patterns.Although the larger pattern within the population is
clear, trends within individuals benefit from a more in-depth examination. In P. edwardsi, the pattern ofdecreasing facet angle with increasing age is most irreg-ular among measurements of the youngest individuals.As the measurements of these young individuals essen-tially represent completely unworn teeth, the variabilityof wear facet patterns among this subset of the popula-tion may be a consequence of incipient wear facets notyet being fully formed through attrition of the enamel.Over short time intervals, the changes in orientation ofwear facets exhibit considerable variability. Over 1- or 2-year intervals, facets are as likely to become steeper asto become shallower. However, if changes in facet anglesfor individual animals are examined over longer inter-vals, they evince a significant decrease that coincideswith the pattern observed at the level of the sample pop-ulation.It is worth noting that, consistently, variables investi-
gated in this study were correlated more closely on theparacristid than the same variables on the cristid obli-qua. Although the relative heights of the trigonid andtalonid are not greatly different in this species, the com-paratively elevated trigonid should encounter the upperteeth before the talonids come into occlusion. Therefore,the orientation of the wear facet on the paracristid mayrepresent changes in jaw motion in a relatively sensitiveway, while the motion that forms the wear facet on thecristid obliqua, as the talonids come into occlusion, maybe influenced by other factors such as the surface mor-phology of the trigonids whose contact guides the talo-
Fig. 6. Scatterplot of age versus wear facet angle broken down by sex of the animal. A LOESS line at 50% tension has been fit-ted through the data. Wear facet angles in adult, nonsenescent individuals (ages 6–18 years) are significantly different between thesexes.
9WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

nids into occlusion, and random factors that accumulateover the duration of each chewing cycle. Additionally,the talonid forms after the trigonid during tooth ontog-eny, and similarly to later forming molars along thetooth row, later forming parts within the crown are morevariable due to developmentally accumulated variation(Kavanagh et al., 2007; Salazar-Ciudad and Jernvall,2010). The unilaterally more ambiguous results on thecristid obliqua suggest that the wear facet on the para-cristid may be more honestly representing the signal ofchange in jaw movement.Gradual changes in wear facet angle, indicating grad-
ual changes in jaw motion over lifetimes of individual P.edwardsi, run contrary to the hypothesis that moretransverse milling motions replace more vertical shear-ing as the modal chewing process specifically at theonset of dental senescence. Instead, these results aremore consistent with the hypothesis that power strokemodification happens over the lifetime of the animal tomaximize functionality and longevity of the tooth as itwears down, as proposed by Seligsohn (1977). However,this hypothesis also predicts that degree of tooth wearshould be the most important factor in determining ori-entation of jaw movement, and this is weakly contra-dicted by our data, which show absolute age to be some-what more strongly correlated with wear facet anglethan overall amount of tooth wear as quantified by areaof dentin exposure.Because our results show that dentally senescent ani-
mals do not dramatically change the way they movetheir jaws as the teeth move into centric occlusion, thebenefits conferred by food grinding may be limited. How-ever, our results may also reflect biomechanical or neu-romotor constraints on the sifaka’s ability to change itsjaw motion regardless of the optimal mode of food proc-essing for teeth of a given wear state. If this is the case,then older, dentally senescent animals may be limited inhow finely they can process their food compared toyounger animals and thereby limited in their ability toobtain sufficient nutrition from the same diet that sus-tained them when they were younger (Lanyon and San-son, 1986). A lack of ability of dentally senescent ani-mals to obtain maximal nutrition from their food resour-ces would be consistent with the conclusions of King etal. (2005) that dentally senescent sifakas are only able tomaintain levels of reproductive success in rainy yearswhen resources appear to be more abundant.Because wear facet angle and jaw motion do not change
dramatically or solely with the onset of dental senescence,but rather are changing during maturation and earlyadulthood, an important possibility to consider is that thispattern of wear facet orientation change with age is due toage-related changes in skull and mandible morphologyrather than changes necessitated by efficiency of food proc-essing. This goes against the hypothesis of Seligsohn(1977). Although no studies of developmental changes inskull shape have been undertaken for sifakas, it is likelythat different aspects of the skull and jaw develop at differ-ent rates. The rapid changes in wear facet angles in juve-nile P. edwardsi could be a consequence of changing forcevectors as the attachment sites of the muscles of mastica-tion alter relative to the skull, dental arcades, and eachother in 3D space. A recent study of ontogenetic change inskull development of the spotted hyena (Crocuta crocuta)demonstrated that although changes in skull morphologyslow after puberty, the shape of the skull continues tochange into adulthood (Tanner et al., 2009). If a similar
pattern of skull growth occurs in P. edwardsi, it would con-cur with the apparently diminished but continued changesin wear facet angles after adult body mass is reached. Thisalternative explanation for the changes in wear facet anglewas evaluated with regression analyses. If Seligsohn(1977) is correct, and observed changes in wear facet orien-tation reflect jaw movement changes meant to keep thetooth maximally functional with the lowest rate of attri-tion at a given wear state or tooth shape, then facet angleshould be closely correlated with the amount of overallwear on the tooth, whereas changes in jaw movementsassociated with ontogenetic development should moreclosely correlate with the age of the individual. There wasa significant correlation between the square root of dentinexposure and wear facet angle, but it was not as strong asthe correlation between age and wear facet angle. It seemsthat both amount of wear and the age of the individualhave power in predicting wear facet angle, but ageaccounts for a greater amount of the variability in wearfacet angles than does the overall amount of wear in thatindividual. The significance of age (which should estimatedevelopmental stage) in predicting wear facet angle fur-ther implies the possible importance of skull growth inaffecting jaw motion in this species.It should be noted, however, that these results are far
from definitive, and Seligsohn’s explanation for wear facetangle changes is still more appealing than the idea thatjaw movement changes are simply an indirect effect of cra-nial maturation and senescence. To further test these twopossibilities, future research should measure wear facetson additional species with specimens for each speciesexhibiting a large range of wear states. If wear facet orien-tation changes are meant to keep shearing blades sharpwith minimal enamel waste, as suggested by Seligsohn(1977), then primates that do not emphasize shearing(cheirogaleiids, Lemur catta, etc.) should not exhibit jawmovement changes with increasing wear or age. Alterna-tively, if age-related changes in cranial shape are responsi-ble for jaw movement changes, then dietary preferencesand modal wear type should have no bearing on whethersuch changes in jaw movement occur, and the animalsmentioned above should exhibit patterns similar to thoseof P. edwardsi. Of course, the difficulty in executing thisstudy would be finding a way to measure jaw movement intaxa that do not develop prominent phase-I facets.
Sex differences in facet orientation change
Interestingly, the results of this study also suggestthat patterns of change in wear facet orientation differbetween the sexes. It seems that adult females are main-taining steeper facets for longer periods than their malecounterparts. This difference could be connected to dif-ferential development rates of skull morphology in malesand females. Additionally, the rapidity of change in wearfacet orientation for male sifakas could indicate thatthey are wearing their teeth down faster than thefemales. In many primate species, dominance is linkedto priority of access to preferred food sources and there-fore can be beneficial to rates of reproductive success(e.g., Jolly, 1984; Young et al., 1990, Bercovitch andStrum, 1993; Pusey et al. 1997). Because P. edwardsi isa female-dominant species (Pochron et al., 2003), it ispossible that females are obtaining preferential access tohigh quality food sources and that their higher qualitydiet is driving the marked difference in wear facet orien-tation between the sexes.
10 S. BLATCH ET AL.
American Journal of Physical Anthropology

Implications for reconstructing jawmotion in extinct animals
Finally, this study has implications for using wearfacet angle to reconstruct species-specific occlusal pat-terns in extinct animals. The facet angles in this studyrange between 46.31 and 7.108 with a mean 6 standarddeviation of 27.65 6 7.758. This range is larger than thatencompassed by all 14 taxa in Butler’s (1972) study,which went from 6281 (Triconodon) to 358 (Protungu-latum) for lower teeth, as well as the range (60–368)encompassed by the four taxa of Kay and Hiiemae’s(1974a) study of lower second molars. Therefore, cautionshould be taken when making dietary inferences fromsuch data in fossils.
CONCLUSIONS
Wear facet orientation suggests that there is a change indirection of jaw movement over the lifetime of individualsof P. edwardsi. Individuals may be modifying jaw move-ment to preserve dental function as wear increases or,alternatively, the changes in jaw movement may be a con-sequence of changes in skull morphology related to matu-ration and growth; however, these changes do not appearto be a focused effect of dental senescence. Although theresults presented here indicate that a species-specificwear pattern of decreasing facet angle with increasing ageexists for P. edwardsi, further research is needed to inves-tigate the predominating forces driving these changes.Although changes in tooth morphology with increasingwear is a complex subject that contains a great deal of var-iability based on individual behavior and dietary differen-ces, future investigation of changes in wear facet orienta-tion over time will benefit from including the study of age-related changes in morphology, individual dietary data,and kinematic data on animal chewing. These approacheswill help to clarify what factors most directly affect thepatterns of wear developed over an animal’s lifetime andmay further illuminate the evolutionary pressures actingon worn teeth to maintain functional efficiency withincreasing wear.
ACKNOWLEDGMENTS
We thank Bill Jungers for his statistical expertise andhelpful comments in the preparation of this manuscript.We also thank Joe Groenke, David Krause, and MaureenO’Leary for access to the Stony Brook University Verte-brate Paleontology Lab and Clint Rubin and Stefan Judexfor access to lCT scanners in the Stony Brook UniversityCenter for Biotechnology. Discussions with Stacey Tecot,Jennifer Verdolin, Ashley Gosselin-Ildari, Stevie Carna-tion, and Stephanie Maiolino greatly improved the qualityof this project. We also thank two anonymous reviewersfor their constructive comments and suggestions. Fieldresearch on live animals conformed to the laws of Mada-gascar and was in accordance with the requirements ofIACUC and Stony Brook University.
APPENDIX
TABLE
A1.Allteethin
whichjaw
motionis
directlydeterminable
Too
thAge
Paracristid
Post-paracristid
CrP
XCrP
YCrP
Z
Twopoints
onlineof
intersection
betwee
nPC
andPPC
orCO
andPPC
Transverse
compon
ent
Vertical
compon
ent
R2
B1
B2
CR2
B1
B2
Ccm
px 1
cmpy 1
cmpz 1
cmpx 2
cmpy2
cmpz 2
57-2002
5.5
0.96
0.69
20.95
225.64
0.99
0.54
1.30
23.72
2.25
0.15
1.40
0.00
221.95
24.74
2.25
221.81
23.34
3.76
31.91
64-2006
4.4
0.97
0.58
20.86
213.20
0.99
0.84
1.81
41.97
2.67
20.25
1.77
0.00
220.64
4.57
2.67
220.90
6.35
25.42
33.44
62-2008
9.0
0.99
0.60
20.90
219.48
0.98
0.72
1.16
21.19
2.07
20.11
1.35
0.00
219.69
21.74
2.07
219.80
20.39
23.13
33.13
26-1994
8.0
0.96
0.70
20.90
20.49
0.99
0.66
1.64
40.74
2.54
0.04
1.75
0.00
27.98
27.68
2.54
27.93
29.42
0.96
34.55
2-1995
12.5
0.99
0.58
20.68
7.88
1.00
0.33
1.45
72.36
2.12
0.25
1.07
0.00
230.38
28.41
2.12
230.13
29.47
6.83
26.55
60-2002
1.5
1.00
0.86
21.33
215.91
0.99
0.46
1.53
73.48
2.86
0.40
1.93
0.00
231.29
25.59
2.86
230.89
27.51
7.98
33.74
26-1995
9.5
0.99
0.84
21.12
16.70
1.00
0.56
1.51
35.09
2.63
0.28
1.90
0.00
26.98
24.52
2.63
26.70
26.42
6.10
35.65
29-2002
10.6
0.98
0.71
20.97
6.64
0.93
0.47
1.05
63.23
2.02
0.24
1.20
0.00
228.03
33.71
2.02
227.79
34.91
6.78
30.52
5-1993
31.6
0.99
0.57
20.92
25.16
0.99
0.33
0.79
95.47
1.71
0.24
0.75
0.00
258.98
48.98
1.71
258.74
49.73
8.06
23.57
21-1995
4.5
0.99
0.94
21.48
246.23
0.98
0.51
1.41
104.63
2.89
0.43
2.07
0.00
252.16
31.07
2.89
251.73
33.15
8.44
35.35
42-2002
11.5
0.99
0.86
20.82
236.46
0.99
0.65
1.63
56.30
2.45
0.21
1.94
0.00
237.91
25.46
2.45
237.70
23.51
4.92
38.36
41-2002
23.5
0.97
0.51
20.57
23.16
1.00
0.31
1.19
168.31
1.77
0.20
0.79
0.00
282.17
70.29
1.77
281.97
71.08
6.60
23.96
45-2002
5.5
0.99
0.73
21.17
231.17
1.00
0.39
1.21
171.83
2.38
0.35
1.34
0.00
285.17
68.67
2.38
284.82
70.01
8.28
29.08
42-2006
15.4
0.97
0.52
20.74
2.92
1.00
0.13
1.26
155.72
2.00
0.39
0.75
0.00
276.55
59.19
2.00
276.16
59.94
10.94
20.13
31-1995
2.5
0.99
0.95
21.06
234.73
0.99
0.33
1.41
164.67
2.47
0.62
1.69
0.00
280.79
50.98
2.47
280.17
52.67
14.08
33.62
31-2002
9.5
0.97
0.56
20.82
28.98
0.99
0.37
1.13
143.28
1.95
0.19
0.94
0.00
278.07
55.20
1.95
277.87
56.14
5.69
25.60
66-2006
4.4
0.99
0.72
21.21
239.88
0.99
0.27
1.23
202.46
2.44
0.45
1.21
0.00
299.46
80.35
2.44
299.01
81.56
10.47
26.05
66-2004
2.2
0.97
1.31
22.04
2127.83
1.00
0.50
1.24
208.27
3.28
0.81
2.64
0.00
2102.57
81.24
3.28
2101.76
83.89
13.83
38.05
14-1995
8.5
0.98
0.68
21.16
245.97
0.99
0.43
1.39
225.56
2.56
0.24
1.44
0.00
2106.20
77.44
2.56
2105.95
78.89
5.46
29.32
36-2003
13.0
0.98
0.70
20.91
26.15
1.00
0.39
1.62
314.92
2.54
0.31
1.50
0.00
2126.53
109.59
2.54
2126.22
111.10
6.99
30.46
1Note that these angles have been transformed from the way theywere reported in the original papers. For us, a horizontal facet iszero, where as for Butler (1972) and Kay and Hiiemae (1974a) avertical one is zero. Therefore, to report results from Butler in away that is comparable to ours we subtracted 288 reported for Trico-nodon by Butler (1972) from 908 to get 628.
11WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

APPENDIX
TABLE
A1.(C
ontinued
)
Too
thAge
Paracristid
Post-paracristid
CrP
XCrP
YCrP
Z
Twopoints
onlineof
intersection
betwee
nPC
andPPC
orCO
andPPC
Transverse
compon
ent
Vertical
compon
ent
R2
B1
B2
CR2
B1
B2
Ccm
px 1
cmpy 1
cmpz 1
cmpx 2
cmpy2
cmpz 2
22-1994
3.0
0.99
1.02
21.27
249.95
0.99
0.44
1.26
291.81
2.52
0.58
1.83
0.00
2135.54
121.63
2.52
2134.96
123.47
12.88
35.33
36-2004
14.3
0.95
0.76
20.86
21.62
0.99
0.36
1.46
293.70
2.33
0.41
1.43
0.00
2126.86
108.08
2.33
2126.45
109.51
9.88
31.16
36-2005
15.4
0.98
0.68
20.90
27.84
1.00
0.42
1.29
272.61
2.19
0.25
1.25
0.00
2128.27
106.99
2.19
2128.02
108.24
6.64
29.65
36-2008
18.0
0.96
0.66
20.70
16.14
0.99
0.38
1.22
265.64
1.92
0.28
1.07
0.00
2130.16
107.18
1.92
2129.89
108.26
8.26
28.99
75-2008
4.05
0.98
0.73
21.10
244.37
0.99
0.17
0.84
222.53
1.94
0.56
0.79
0.00
2137.72
107.24
1.94
2137.16
108.04
16.16
21.50
Too
thAge
Cristid
obliqua
Post-paracristid
CrP
XCrP
YCrP
Z
Twopoints
onlineof
intersection
betweenPC
andPPC
orCO
andPPC
Transverse
compon
ent
Vertical
compon
ent
R2
B1
B2
CR2
B1
B2
Ccm
px 1
cmpy 1
cmpz 1
cmpx 2
cmpy 2
cmpz 2
64-2006
4.4
0.99
0.56
20.94
211
.92
0.99
0.84
1.81
41.97
2.75
20.28
1.79
0.00
219.57
6.51
2.75
219.85
8.31
25.80
32.94
62-2008
9.0
0.99
0.75
21.06
222.20
0.98
0.72
1.16
21.19
2.23
0.03
1.63
0.00
219.48
21.49
2.23
219.45
0.14
0.75
36.22
57-2002
5.5
0.98
0.83
21.06
229.65
0.99
0.54
1.30
23.72
2.36
0.30
1.65
0.00
222.63
25.61
2.36
222.33
23.96
7.15
34.83
26-1994
8.0
1.00
0.68
20.89
23.17
0.99
0.66
1.64
40.74
2.53
0.02
1.70
0.00
26.96
29.35
2.53
26.94
31.05
0.47
34.01
26-1995
9.5
0.99
0.81
20.96
21.00
1.00
0.56
1.51
35.09
2.47
0.25
1.76
0.00
25.70
26.46
2.47
25.45
28.22
5.77
35.32
29-2002
10.6
1.00
1.00
20.92
10.28
0.93
0.47
1.05
63.23
1.97
0.53
1.48
0.00
226.87
34.93
1.97
226.35
36.40
14.98
35.93
2-1995
12.5
0.99
0.63
20.89
3.91
1.00
0.33
1.45
72.36
2.34
0.31
1.21
0.00
229.30
29.97
2.34
229.00
31.18
7.44
27.22
60-2002
1.5
0.98
1.06
21.15
28.87
0.99
0.46
1.53
73.48
2.68
0.60
2.15
0.00
230.73
26.45
2.68
230.12
28.61
12.62
38.07
21-1995
4.5
1.00
0.77
21.09
220.20
0.98
0.51
1.41
104.63
2.50
0.26
1.64
0.00
249.89
34.28
2.50
249.63
35.92
5.96
33.10
42-2002
11.5
0.98
0.74
20.98
237.95
0.99
0.65
1.63
56.30
2.61
0.09
1.85
0.00
236.15
22.60
2.61
236.07
20.75
1.94
35.34
42-2006
15.4
1.00
0.55
20.76
3.10
1.00
0.13
1.26
155.72
2.02
0.42
0.80
0.00
275.37
60.68
2.02
274.95
61.47
11.80
21.05
31-1995
2.5
0.99
0.47
20.84
3.33
0.99
0.33
1.41
164.67
2.25
0.14
0.94
0.00
271.78
63.65
2.25
271.65
64.59
3.52
22.71
31-2002
9.5
0.98
0.66
20.95
218.83
0.99
0.37
1.13
143.28
2.08
0.29
1.10
0.00
278.02
55.25
2.08
277.73
56.34
7.98
27.57
41-2002
23.5
0.98
0.45
20.58
23.49
1.00
0.31
1.19
168.31
1.77
0.14
0.72
0.00
281.64
70.93
1.77
281.49
71.65
4.55
21.95
45-2002
5.5
1.00
0.69
20.98
211
.60
1.00
0.39
1.21
171.83
2.19
0.30
1.21
0.00
283.77
70.36
2.19
283.47
71.57
7.90
28.75
66-2006
4.4
1.00
0.64
21.04
220.57
0.99
0.27
1.23
202.46
2.27
0.37
1.06
0.00
298.28
81.81
2.27
297.91
82.87
9.21
24.85
66-2004
2.2
0.99
0.98
21.23
240.40
1.00
0.50
1.24
208.27
2.47
0.48
1.83
0.00
2100.85
83.37
2.47
2100.37
85.20
11.00
36.06
14-1995
8.5
1.00
0.62
21.37
261.69
0.99
0.43
1.39
225.56
2.76
0.19
1.45
0.00
2104.03
80.47
2.76
2103.84
81.92
3.84
27.63
36-2003
13.0
0.99
0.64
20.81
9.16
1.00
0.39
1.62
314.92
2.43
0.24
1.35
0.00
2125.74
110.88
2.43
2125.49
112.23
5.69
28.92
36-2004
14.3
0.98
0.58
20.65
25.79
0.99
0.36
1.46
293.70
2.12
0.22
1.09
0.00
2126.52
108.58
2.12
2126.29
109.67
6.03
27.05
36-2005
15.4
0.99
0.70
20.93
28.78
1.00
0.42
1.29
272.61
2.22
0.27
1.29
0.00
2126.82
108.86
2.22
2126.55
110.15
7.01
30.00
36-2006
16.4
0.92
0.88
20.98
217.93
1.00
0.67
1.67
320.78
2.65
0.21
2.14
0.00
2127.86
107.67
2.65
2127.65
109.81
4.54
38.81
36-2008
18.0
0.99
0.58
20.75
13.43
0.99
0.38
1.22
265.64
1.96
0.20
0.99
0.00
2128.51
109.20
1.96
2128.31
110.19
5.71
26.67
75-2008
4.1
1.00
0.70
21.09
238.40
0.99
0.17
0.84
222.53
1.92
0.53
0.76
0.00
2135.69
108.94
1.92
2135.16
109.70
15.34
20.96
22-1994
3.0
0.98
0.64
20.73
20.33
0.99
0.44
1.26
291.81
1.99
0.20
1.13
0.00
2136.60
120.30
1.99
2136.40
121.43
5.81
29.45
Too
th:individualand
instance
iden
tifier;age:
ageof
individualattimeof
capture
and
gen
eration
oftooth
mold
instance;R2:adjusted
squared
correlation
coefficien
tforleast
squaresplanarregression;B1:coefficien
ton
xcoordinate;B2:coefficien
ton
ycoordinate;C:constantin
planarregression;CrP
X(Y,Z):x(y,z)
valueof
cross-product
oftw
oplanes
specified
inresp
ectiverow;cm
ptx1(y
1,z 1):x(y,z)
valueof
thefirstpointin
common
betwee
ntw
oplanes
specified
inresp
ectiverow;cm
ptx 2
(y2,z 2):x(y,z)
valueof
thesecond
pointin
common
betwee
ntw
oplanes
specified
inresp
ectiverow;transverse
compon
ent:
theangle
ofdev
iation
from
thebuccolingualaxis
ofthevectorof
intersection
betweentw
oplanes
specified
inresp
ectiverow;verticalcompon
ent:
theangle
ofdev
iation
from
parallel
totheocclusa
lplaneof
thevectorof
intersection
betweentw
oplanes
specified
inresp
ec-
tiverow.
12 S. BLATCH ET AL.
American Journal of Physical Anthropology

TABLE
A2.Allteeth
Too
thAge
Paracristid
Twopoints
onfacetalongalinewith7.268angle
Vertical
compon
ent
R2
B1
B2
Cx 1
y 1z 1
x 2y 2
z 2
64-2005
3.34
0.99
0.80
21.05
29.14
0.00
10.00
219.64
10.00
11.27
213.00
33.37
57-2002
5.49
0.96
0.69
20.95
225.64
0.00
10.00
235.16
10.00
11.27
229.51
29.29
64-2006
4.39
0.97
0.58
20.86
213.20
0.00
10.00
221.81
10.00
11.27
217.09
25.09
62-2004
5.25
0.99
0.78
21.07
220.77
0.00
10.00
231.49
10.00
11.27
225.06
32.56
62-2005
6.37
0.99
0.73
21.21
224.81
0.00
10.00
236.88
10.00
11.27
231.07
29.96
62-2008
9.00
0.99
0.60
20.90
219.48
0.00
10.00
228.49
10.00
11.27
223.59
25.91
62-2002
3.49
0.99
0.81
21.11
231.30
0.00
10.00
242.44
10.00
11.27
235.78
33.44
26-1994
7.97
0.96
0.70
20.90
20.49
0.00
10.00
11.48
10.00
11.27
17.37
30.30
11-1995
11.52
0.98
0.51
20.75
7.39
0.00
10.00
20.12
10.00
11.27
4.03
22.34
2-1995
12.52
0.99
0.58
20.68
7.88
0.00
10.00
1.12
10.00
11.27
6.10
26.29
60-2002
1.52
1.00
0.86
21.33
215.91
0.00
10.00
229.17
10.00
11.27
222.26
34.44
60-2004
3.27
0.99
0.72
21.26
215.16
0.00
10.00
227.81
10.00
11.27
222.18
29.16
26-1995
9.52
0.99
0.84
21.12
16.70
0.00
10.00
5.50
10.00
11.27
12.48
34.71
26-2002
16.49
0.99
0.72
20.67
15.99
0.00
10.00
9.26
10.00
11.27
15.59
32.15
18-1995
6.59
0.99
0.50
20.78
13.43
0.00
10.00
5.67
10.00
11.27
9.68
21.68
29-1994
2.08
1.00
0.72
21.44
20.12
0.00
10.00
214.50
10.00
11.27
29.16
27.92
29-1995
3.64
0.99
0.68
20.98
1.93
0.00
10.00
27.82
10.00
11.27
22.26
28.88
29-2002
10.61
0.98
0.71
20.97
6.64
0.00
10.00
23.02
10.00
11.27
2.84
30.15
11-1994
9.97
0.99
0.58
20.86
5.38
0.00
10.00
23.18
10.00
11.27
1.49
24.89
60-2005
4.41
0.99
0.74
21.20
214.02
0.00
10.00
226.06
10.00
11.27
220.19
30.20
65-2005
3.38
1.00
0.77
21.31
224.05
0.00
10.00
237.17
10.00
11.27
231.11
31.04
5-1993
31.56
0.99
0.57
20.92
25.16
0.00
10.00
214.34
10.00
11.27
29.80
24.23
73-2005
1.39
0.98
0.70
21.39
276.74
0.00
10.00
290.60
10.00
11.27
285.37
27.43
61-2005
2.30
0.99
1.01
21.12
224.93
0.00
10.00
236.11
10.00
11.27
227.41
40.79
69-2004
1.25
0.98
0.73
21.09
215.50
0.00
10.00
226.39
10.00
11.27
220.46
30.46
60-2006
5.47
1.00
0.47
21.10
215.16
0.00
10.00
226.20
10.00
11.27
222.96
17.83
21-1993
1.55
0.99
0.79
21.89
253.79
0.00
10.00
272.68
10.00
11.27
267.21
28.48
21-1994
2.96
0.99
0.80
21.36
232.25
0.00
10.00
245.86
10.00
11.27
239.60
31.83
21-1995
4.51
0.99
0.94
21.48
246.23
0.00
10.00
261.06
10.00
11.27
253.57
36.58
65-2003
0.94
0.99
0.90
21.24
242.30
0.00
10.00
254.72
10.00
11.27
247.29
36.38
42-2002
11.50
0.99
0.86
20.82
236.46
0.00
10.00
244.63
10.00
11.27
237.03
37.03
41-2002
23.50
0.97
0.51
20.57
23.16
0.00
10.00
17.42
10.00
11.27
21.83
23.61
27-1994
10.97
0.98
0.52
20.79
266.30
0.00
10.00
274.21
10.00
11.27
270.00
22.65
45-2002
5.49
0.99
0.73
21.17
231.17
0.00
10.00
242.89
10.00
11.27
237.06
30.05
42-2003
11.95
0.99
0.73
20.84
20.13
0.00
10.00
28.55
10.00
11.27
22.32
31.74
42-2004
13.25
0.97
0.82
20.50
25.35
0.00
10.00
20.33
10.00
11.27
27.93
37.04
42-2006
15.44
0.97
0.52
20.74
2.92
0.00
10.00
24.44
10.00
11.27
20.22
22.70
42-2008
17.00
0.97
0.68
20.75
22.11
0.00
10.00
29.62
10.00
11.27
23.81
29.96
31-1995
2.51
0.99
0.95
21.06
234.73
0.00
10.00
245.34
10.00
11.27
237.18
39.00
31-2002
9.50
0.97
0.56
20.82
28.98
0.00
10.00
217.20
10.00
11.27
212.61
24.46
31-2006
13.50
0.99
0.63
20.63
2.66
0.00
10.00
23.65
10.00
11.27
1.80
28.39
83-2006
3.49
1.00
0.88
21.14
228.68
0.00
10.00
240.06
10.00
11.27
232.70
36.13
72-2006
6.44
1.00
0.54
21.06
222.03
0.00
10.00
232.63
10.00
11.27
228.54
22.07
72-2008
8.02
0.99
0.58
21.04
220.75
0.00
10.00
231.12
10.00
11.27
226.68
23.77
36-1995
5.52
1.00
0.84
21.07
225.46
0.00
10.00
236.20
10.00
11.27
229.17
34.88
36-2002
12.49
0.99
0.72
20.97
218.26
0.00
10.00
227.98
10.00
11.27
222.04
30.53
66-2006
4.39
0.99
0.72
21.21
239.88
0.00
10.00
251.96
10.00
11.27
246.30
29.33
66-2004
2.17
0.97
1.31
22.04
2127.83
0.00
10.00
2148.21
10.00
11.27
2137.72
46.12
66-2003
0.87
0.98
1.18
21.36
264.13
0.00
10.00
277.74
10.00
11.27
267.71
44.87
13WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

APPENDIX
TABLE
A2.(C
ontinued
)
Too
thAge
Paracristid
Twopoints
onfacetalongalinewith7.268angle
Vertical
compon
ent
R2
B1
B2
Cx1
y 1z 1
x 2y 2
z 2
14-1995
8.52
0.98
0.68
21.16
245.97
0.00
10.00
257.59
10.00
11.27
252.31
27.60
36-2003
12.95
0.98
0.70
20.91
26.15
0.00
10.00
215.29
10.00
11.27
29.41
30.26
22-1994
2.95
0.99
1.02
21.27
249.95
0.00
10.00
262.61
10.00
11.27
254.05
40.31
36-2004
14.25
0.95
0.76
20.86
21.62
0.00
10.00
210.26
10.00
11.27
23.72
32.98
36-2005
15.37
0.98
0.68
20.90
27.84
0.00
10.00
216.79
10.00
11.27
211
.16
29.19
36-2008
18.00
0.96
0.66
20.70
16.14
0.00
10.00
9.14
10.00
11.27
14.87
29.58
75-2005
1.41
0.99
0.57
21.05
230.07
0.00
10.00
240.58
10.00
11.27
236.23
23.34
75-2008
4.05
0.98
0.73
21.10
244.37
0.00
10.00
255.38
10.00
11.27
249.49
30.29
24-1993
1.45
0.99
0.51
21.21
20.72
0.00
10.00
212.80
10.00
11.27
29.20
19.63
3-1994
18.95
0.99
0.45
20.77
22.71
0.00
10.00
210.40
10.00
11.27
26.85
19.39
3-1995
20.50
0.99
0.43
20.64
22.16
0.00
10.00
28.51
10.00
11.27
25.06
18.89
3-2002
27.47
0.99
0.50
20.43
21.57
0.00
10.00
25.87
10.00
11.27
21.40
23.92
3-2006
31.43
0.92
0.38
20.25
21.80
0.00
10.00
24.27
10.00
11.27
20.75
19.22
24-1995
4.41
0.99
0.72
21.15
0.34
0.00
10.00
211
.14
10.00
11.27
25.43
29.52
24-2002
11.38
0.98
0.43
20.78
21.06
0.00
10.00
28.82
10.00
11.27
25.51
18.17
24-2004
13.14
0.99
0.42
20.75
0.10
0.00
10.00
27.36
10.00
11.27
24.10
17.94
24-2005
14.27
0.99
0.42
20.53
20.44
0.00
10.00
25.77
10.00
11.27
22.25
19.24
24-2006
15.33
0.98
0.36
20.65
20.56
0.00
10.00
27.06
10.00
11.27
24.29
15.38
24-2008
16.91
0.99
0.37
20.85
0.25
0.00
10.00
28.23
10.00
11.27
25.66
14.31
3-1993
17.55
0.99
0.51
20.84
21.98
0.00
10.00
210.42
10.00
11.27
26.44
21.52
63-2003
0.93
0.98
1.01
21.17
0.42
0.00
10.00
211
.26
10.00
11.27
22.69
40.37
63-2004
2.21
0.99
0.83
21.24
1.98
0.00
10.00
210.40
10.00
11.27
23.66
33.77
63-2005
3.33
0.98
0.55
21.24
0.57
0.00
10.00
211
.81
10.00
11.27
27.89
21.28
63-2006
4.40
0.98
0.91
21.40
1.64
0.00
10.00
212.32
10.00
11.27
24.99
36.00
59-2002
1.50
0.99
0.56
21.24
0.96
0.00
10.00
211
.40
10.00
11.27
27.42
21.53
59-2004
3.26
0.98
0.59
21.17
2.10
0.00
10.00
29.62
10.00
11.27
25.17
23.83
59-2005
4.39
0.98
0.63
21.16
0.79
0.00
10.00
210.80
10.00
11.27
25.96
25.64
59-2006
5.46
0.99
0.58
21.10
1.50
0.00
10.00
29.51
10.00
11.27
25.14
23.43
59-2008
7.01
1.00
0.42
20.85
0.72
0.00
10.00
27.76
10.00
11.27
24.68
17.01
Too
thAge
Cristid
Obliqua
Twopoints
onfacetalongalinewith6.458angle
Vertical
compon
ent
R2
B1
B2
Cx1
y 1z 1
x 2y 2
z 2
64-2005
3.34
0.98
0.84
20.94
22.95
0.00
10.00
212.39
10.00
11.13
25.01
36.26
64-2004
2.20
0.99
0.65
20.88
28.16
0.00
10.00
216.95
10.00
11.13
211
.43
28.74
64-2006
4.39
0.99
0.56
20.94
211
.92
0.00
10.00
221.34
10.00
11.13
216.84
24.05
62-2004
5.25
0.99
0.72
21.00
215.66
0.00
10.00
225.70
10.00
11.13
219.67
30.90
62-2005
6.37
0.99
0.81
21.19
221.59
0.00
10.00
233.49
10.00
11.13
226.76
33.80
62-2008
9.00
0.99
0.75
21.06
222.20
0.00
10.00
232.84
10.00
11.13
226.57
31.91
62-2002
3.49
0.98
0.85
21.07
227.92
0.00
10.00
238.64
10.00
11.13
231.38
35.83
57-2002
5.49
0.98
0.83
21.06
229.65
0.00
10.00
240.28
10.00
11.13
233.13
35.38
26-1994
7.97
1.00
0.68
20.89
23.17
0.00
10.00
14.30
10.00
11.13
20.12
30.04
26-1995
9.52
0.99
0.81
20.96
21.00
0.00
10.00
11.41
10.00
11.13
18.42
34.85
26-2002
16.49
0.99
0.80
20.78
17.29
0.00
10.00
9.47
10.00
11.13
16.59
35.30
18-1995
6.59
0.99
0.54
20.89
15.13
0.00
10.00
6.24
10.00
11.13
10.66
23.73
18-1993
3.64
0.99
0.32
20.70
16.00
0.00
10.00
8.98
10.00
11.13
11.33
13.18
29-1995
3.64
0.98
0.74
21.09
3.08
0.00
10.00
27.83
10.00
11.13
21.69
31.38
29-2002
10.61
1.00
1.00
20.92
10.28
0.00
10.00
1.11
10.00
11.13
10.03
41.56
14 S. BLATCH ET AL.
American Journal of Physical Anthropology

APPENDIX
TABLE
A2.(C
ontinued
)
Too
thAge
Cristid
Obliqua
Twopoints
onfacetalongalinewith6.458angle
Vertical
compon
ent
R2
B1
B2
Cx1
y 1z 1
x 2y 2
z 2
11-1994
9.97
1.00
0.58
20.79
10.03
0.00
10.00
2.10
10.00
11.13
7.02
26.04
11-1995
11.52
0.99
0.45
20.73
10.24
0.00
10.00
2.93
10.00
11.13
6.58
19.94
2-1995
12.52
0.99
0.63
20.89
3.91
0.00
10.00
24.98
10.00
11.13
0.37
27.98
60-2002
1.52
0.98
1.06
21.15
28.87
0.00
10.00
220.36
10.00
11.13
211
.06
42.75
60-2004
3.27
0.99
0.98
21.07
210.14
0.00
10.00
220.84
10.00
11.13
212.26
40.44
60-2005
4.41
0.98
0.76
21.07
23.51
0.00
10.00
214.18
10.00
11.13
27.78
32.44
60-2006
5.47
0.99
0.37
20.74
4.31
0.00
10.00
23.08
10.00
11.13
20.18
16.08
21-1993
1.55
0.98
0.38
20.87
0.11
0.00
10.00
28.56
10.00
11.13
25.77
15.47
21-1994
2.96
0.99
0.88
21.11
217.10
0.00
10.00
228.17
10.00
11.13
220.63
36.84
21-1995
4.51
1.00
0.77
21.09
220.20
0.00
10.00
231.13
10.00
11.13
224.66
32.70
71-2005
7.37
0.99
0.46
20.58
8.84
0.00
10.00
3.00
10.00
11.13
6.99
21.60
65-2003
0.94
0.99
0.76
20.94
219.28
0.00
10.00
228.68
10.00
11.13
222.14
33.03
65-2005
3.38
0.99
0.53
20.99
24.79
0.00
10.00
214.71
10.00
11.13
210.53
22.56
73-2005
1.39
0.89
0.65
21.11
256.50
0.00
10.00
267.63
10.00
11.13
262.36
27.66
61-2005
2.30
0.98
1.16
20.97
214.35
0.00
10.00
224.01
10.00
11.13
213.48
46.31
69-2004
1.25
0.99
0.81
21.01
212.91
0.00
10.00
223.05
10.00
11.13
216.13
34.50
42-2002
11.50
0.98
0.74
20.98
237.95
0.00
10.00
247.72
10.00
11.13
241.40
32.14
42-2003
11.95
0.99
0.64
20.66
15.13
0.00
10.00
8.54
10.00
11.13
14.15
29.14
42-2004
13.25
0.97
0.92
20.85
2.60
0.00
10.00
25.93
10.00
11.13
2.34
39.40
42-2005
14.38
0.99
0.28
20.40
31.74
0.00
10.00
27.73
10.00
11.13
30.03
12.89
42-2006
15.44
1.00
0.55
20.76
3.10
0.00
10.00
24.53
10.00
11.13
0.13
24.87
42-2008
17.00
0.99
0.64
20.67
7.01
0.00
10.00
0.28
10.00
11.13
5.96
29.44
31-1995
2.51
0.99
0.47
20.84
3.33
0.00
10.00
25.08
10.00
11.13
21.32
20.49
31-2002
9.50
0.98
0.66
20.95
218.83
0.00
10.00
228.33
10.00
11.13
222.79
28.80
31-2006
13.50
0.99
0.61
20.71
20.12
0.00
10.00
27.20
10.00
11.13
21.92
27.70
41-2002
23.50
0.98
0.45
20.58
23.49
0.00
10.00
17.68
10.00
11.13
21.52
20.93
27-1994
10.97
0.99
0.67
20.77
262.45
0.00
10.00
270.16
10.00
11.13
264.28
30.29
45-2002
5.49
1.00
0.69
20.98
211
.60
0.00
10.00
221.39
10.00
11.13
215.59
29.93
20-1994
12.97
0.99
0.32
21.08
211
.72
0.00
10.00
222.49
10.00
11.13
220.47
11.37
83-2006
3.49
0.99
0.91
21.18
228.59
0.00
10.00
240.40
10.00
11.13
232.63
37.69
66-2006
4.39
1.00
0.64
21.04
220.57
0.00
10.00
230.99
10.00
11.13
225.78
27.34
66-2004
2.17
0.99
0.98
21.23
240.40
0.00
10.00
252.67
10.00
11.13
244.25
39.91
66-2003
0.87
0.99
1.05
21.19
241.58
0.00
10.00
253.48
10.00
11.13
244.37
42.16
14-1994
6.97
0.98
0.26
21.23
237.93
0.00
10.00
250.23
10.00
11.13
248.98
7.10
14-1995
8.52
1.00
0.62
21.37
261.69
0.00
10.00
275.36
10.00
11.13
270.74
24.66
72-2004
4.25
0.96
0.46
20.74
8.10
0.00
10.00
0.75
10.00
11.13
4.53
20.61
72-2006
6.44
0.97
0.52
20.72
18.99
0.00
10.00
11.82
10.00
11.13
16.21
23.58
72-2008
8.02
1.00
0.46
20.87
20.71
0.00
10.00
29.43
10.00
11.13
25.85
19.56
36-1995
5.52
0.98
0.96
20.95
27.24
0.00
10.00
216.73
10.00
11.13
28.23
40.17
36-2002
12.49
0.99
0.69
20.89
25.54
0.00
10.00
214.42
10.00
11.13
28.51
30.40
36-2003
12.95
0.99
0.64
20.81
9.16
0.00
10.00
1.07
10.00
11.13
6.52
28.42
36-2004
14.25
0.98
0.58
20.65
25.79
0.00
10.00
19.25
10.00
11.13
24.33
26.82
36-2005
15.37
0.99
0.70
20.93
28.78
0.00
10.00
218.06
10.00
11.13
212.15
30.43
36-2006
16.44
0.92
0.88
20.98
217.93
0.00
10.00
227.75
10.00
11.13
220.01
37.56
36-2008
18.00
0.99
0.58
20.75
13.43
0.00
10.00
5.98
10.00
11.13
10.93
26.20
75-2005
1.41
1.00
0.28
20.87
0.44
0.00
10.00
28.23
10.00
11.13
26.43
10.10
75-2006
2.47
0.97
0.49
20.66
22.76
0.00
10.00
16.12
10.00
11.13
20.27
22.43
15WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

APPENDIX
TABLE
A2.(C
ontinued
)
Too
thAge
Cristid
Obliqua
Twopoints
onfacetalongalinewith6.458angle
Vertical
compon
ent
R2
B1
B2
Cx 1
y 1z 1
x 2y 2
z 2
75-2008
4.05
1.00
0.70
21.09
238.40
0.00
10.00
249.26
10.00
11.13
243.53
29.63
22-1994
2.95
0.98
0.64
20.73
20.33
0.00
10.00
13.01
10.00
11.13
18.61
29.07
24-2993
1.45
0.99
0.65
21.24
20.55
0.00
10.00
212.91
10.00
11.13
27.84
26.75
24-1995
4.41
0.99
0.57
21.02
0.44
0.00
10.00
29.77
10.00
11.13
25.19
24.44
24-2002
11.38
0.99
0.40
20.80
20.76
0.00
10.00
28.72
10.00
11.13
25.57
17.37
24-2004
13.14
0.99
0.46
20.71
0.36
0.00
10.00
26.72
10.00
11.13
22.88
20.90
24-2005
14.27
0.97
0.48
20.56
20.20
0.00
10.00
25.84
10.00
11.13
21.66
22.59
24-2006
15.33
1.00
0.45
20.78
20.30
0.00
10.00
28.13
10.00
11.13
24.50
19.83
24-2008
16.91
0.99
0.31
20.77
0.32
0.00
10.00
27.36
10.00
11.13
25.16
12.34
3-1993
17.55
0.99
0.44
20.85
21.75
0.00
10.00
210.22
10.00
11.13
26.75
19.05
3-1994
18.95
0.98
0.33
20.54
21.90
0.00
10.00
27.34
10.00
11.13
24.64
15.03
3-1995
20.50
0.98
0.48
20.45
21.78
0.00
10.00
26.24
10.00
11.13
21.91
23.29
3-2002
27.47
1.00
0.61
20.46
21.53
0.00
10.00
26.09
10.00
11.13
20.54
28.88
3-2006
31.43
1.00
0.44
20.37
21.81
0.00
10.00
25.55
10.00
11.13
21.59
21.49
63-2003
0.93
0.98
0.30
20.60
0.36
0.00
10.00
25.63
10.00
11.13
23.34
12.83
63-2004
2.21
0.98
0.70
20.77
1.40
0.00
10.00
26.32
10.00
11.13
20.17
31.43
63-2005
3.33
0.95
0.76
21.12
0.40
0.00
10.00
210.79
10.00
11.13
24.49
32.03
63-2006
4.40
0.98
0.89
20.93
1.24
0.00
10.00
28.10
10.00
11.13
20.29
37.81
59-2002
1.50
0.99
0.48
20.89
0.57
0.00
10.00
28.29
10.00
11.13
24.52
20.57
59-2004
3.26
0.99
0.57
21.00
1.90
0.00
10.00
28.05
10.00
11.13
23.52
24.23
59-2005
4.39
0.99
0.48
20.84
0.56
0.00
10.00
27.81
10.00
11.13
23.92
21.12
59-2006
5.46
1.00
0.49
20.86
1.21
0.00
10.00
27.40
10.00
11.13
23.51
21.17
59-2008
7.01
0.99
0.34
20.88
1.20
0.00
10.00
27.58
10.00
11.13
25.20
13.31
Too
th:individualand
instance
iden
tifier;age:
ageof
individualattimeof
capture
and
gen
eration
oftooth
mold
instance;R2:adjusted
squared
correlation
coefficien
tforleast
squaresplanarregression;B1:coefficien
ton
xcoordinate;B2:coefficien
ton
ycoordinate;C:constantin
planarregression.
16 S. BLATCH ET AL.
American Journal of Physical Anthropology

LITERATURE CITED
Agrawal KR, Lucas PW, Bruce IC. 2000. The effects of food frag-mentation index on mandibular closing angle in human mas-tication. Arch Oral Biol 45:577–584.
Anthony RL, Kay RF. 1993. Tooth form and diet in Ateline andAlouattine primates: reflections on the comparative method.Am J Sci A 293:356–382.
Bercovitch FB, Strum SC. 1993. Dominance rank, resourceavailability, and reproductive maturation in female savannababoons. Behav Ecol Sociobiol 33:313–318.
Boyer DM. 2008. Relief index of second mandibular molars is acorrelate of diet among prosimian primates and other euarch-ontan mammals. J Hum Evol 55:1118–1137.
Bunn JM, Ungar PS. 2009. Dental topography and diets of fourOld World monkey species. Am J Primatol 71:466–477.
Butler PM. 1952. The milk-molars of Perissodactyla, with remarksonmolar occlusion. Proc Zool Soc Lond 121:777–817.
Butler PM. 1972. Some functional aspects of molar evolution.Evolution 26:474–483.
Costa RL, Greaves WS. 1981. Experimentally produced toothwear facets and the direction of jaw motion. J Paleontol55:635–638.
Dennis JC, Ungar PS, Teaford MF, Glander KE. 2004. Dentaltopography and molar wear in Alouatta palliata from CostaRica. Am J Phys Anthropol 125:152–161.
Dew JL, Wright PC. 1998. Frugivory and seed dispersal by fourspecies of primates in Madagascar’s eastern rain forest. Bio-tropica 30:425–437.
Evans AR, Sanson GD. 1998. The effect of tooth shape on thebreakdown of insects. J Zool Lond 246:391–400.
Evans AR, Sanson GD. 2003. The tooth of perfection: functionaland spatial constraints on mammalian tooth shape. Biol JLinn Soc 78:173–191.
Evans AR, Sanson GD. 2006. Spatial and functional modeling ofcarnivore and insectivore molariform teeth. J Morphol 267:649–662.
Glander KE, Fedigan LM, Fedigan L, Chapman C. 1991. Fieldmethods for capture and measurement of three monkey spe-cies in Costa Rica. Folia Primatol 57:70–82.
Greaves WS. 1973. The inference of jaw motion from tooth wearfacets. J Paleontol 47:1000–1001.
Hemingway CA. 1996. Morphology and phenology of seeds andwhole fruit eaten by Milne-Edwards’ sifaka, Propithecus dia-dema edwardsi, in Ranomafana National Park, Madagascar.Int J Primatol 17:637–659.
Hemingway CA. 1998. Selectivity and variability in the diet ofMilne-Edwards’ sifakas (Propithecus diadema edwardsi):implications for folivory and seed-eating. Int J Primatol 19:355–377.
Hill WCO. 1953. Primates: comparative anatomy and taxonomy.I. Strepsirhini. Edinburgh: University Press.
Huggins RM, Loesch DZ. 1998. On the analysis of mixed longi-tudinal growth data. Biometrics 54:583–595.
Jernvall J, Selanne L. 1999. Laser confocal microscopy and geo-graphic information systems in the study of dental morphol-ogy. Palaeontol Electron 2:18.
Jolly A. 1984. The puzzle of female feeding priority. In: SmallM, editor. Female primates: studies by women primatologists.New York: Alan R. Liss. p 197–215.
Kavanagh KD, Evans AR, Jernvall J. 2007. Predicting evolu-tionary patterns of mammalian teeth from development. Na-ture 449:427–432.
Kay RF. 1977. The evolution of molar occlusion in the Cercopi-thecidae and early catarrhines. Am J Phys Anthropol 46:327–352.
Kay RF. 1978. Molar structure and diet in extant Cercopithecidae.In: Butler PM, Joysey KA, editors. Development, function andevolution of teeth. New York: Academic Press. p 21–53.
Kay RF, Hiiemae KM. 1974a. Jaw movement and tooth use inrecent and fossil primates. Am J Phys Anthropol 40:227–256.
Kay RF, Hiiemae KM. 1974b. Mastication in Galago crassicau-datus: a cinefluorographic and occlusal study. In: Martin RD,
Doyle GA, editors. Prosimian biology. London: Duckworth. p501–530.
King SJ, Arrigo-Nelson SJ, Pochron ST, Semprebon GM, God-frey LR, Wright PC, Jernvall J. 2005. Dental senescence in along-lived primate links infant survival to rainfall. Proc NatlAcad Sci USA 102:16579–16583.
Kojola I, Helle T, Huhta E, Niva A. 1998. Foraging conditions,tooth wear and herbivore body reserves: a study of femalereindeer. Oecologia 117:26–30.
Lanyon JM, Sanson GD. 1986. Koala (Phascolarctos cinereus)dentition and nutrition. II. Implications of tooth wear innutrition. J Zool Lond 209:169–181.
Lucas PW. 1982. Basic principles of tooth design. In: Kurten B,editor. Teeth: form, function and evolution. New York: Colum-bia University Press. p 154–162.
Lucas PW. 2004. Dental functional morphology: how teeth work.Cambridge: Cambridge University Press.
Lucas PW, Luke DA. 1983. Methods for analyzing the break-down of food in human mastication. Arch Oral Biol 28:813–819.
Meldrum DJ, Kay RF. 1997. Nuciruptor rubiciae, a new pithe-ciin seed predator from Miocene of Colombia. Am J PhysAnthropol 102:407–428.
Meyers DM, Wright PC. 1993. Resource tracing: food availabil-ity and Propithecus seasonal reproduction. In: Kappeler PM,Ganzhorn JU, editors. Lemur social systems and their ecologi-cal basis. New York: Plenum Press. p 179–192.
Mills JRE. 1978. The relationship between tooth patterns andjaw movements in the Hominoidea. In: Butler PM, JoyseyKA, editors. Development, function and evolution of teeth.New York: Academic Press. p 341–353.
M’Kirera F, Ungar PS. 2003. Occlusal relief changes with molarwear in Pan troglodytes troglodytes and Gorilla gorilla gorilla.Am J Primatol 60:31–41.
Ozaki M, Kaji K, Matsuda N, Ochiai K, Asada M, Ohba T, HosoiE, Tado H, Koizumi T, Suwa G, Takatsuki S. 2010. The rela-tionship between food habits, molar wear and life expectancyin wild sika deer populations. J Zool 280:202–212.
Pochron ST, Fitzgerald J, Gilbert CC, Lawrence D, Grgas M,Rakotonirina G, Ratsimbazafy R, Mirakotosoa R, Wright PC.2003. Patterns of female dominance in Propithecus diademaedwardsi of Ranomafana National Park, Madagascar. Am JPhys Anthropol 61:173–185.
Pochron ST, Morelli TL, Scirbona J, Wright PC. 2005. Sex dif-ferences in scent marking in Propithecus edwardsi of Rano-mafana National Park, Madagascar. Am J Phys Anthropol66:97–110.
Pochron ST, Tucker WT, Wright PC. 2004. Demography, life his-tory, and social structure in Propithecus diadema edwardsifrom 1986–2000 in Ranomafana National Park, Madagascar.Am J Phys Anthropol 125:61–72.
Pusey A, Williams J, Goodall J. 1997. The influence of domi-nance rank on the reproductive success of female chimpan-zees. Science 277:828–831.
Pochron ST, Wright PC, Schaentzler E, Ippolito M, RakotonirinaG, Ratsimbazafy R, Rakotosoa R. 2002. Effect of season andage on the gonadosomatic index of Milne-Edwards’ sifakas(Propithecus diadema edwardsi) in Ranomafana NationalPark, Madagascar. Int J Primatol 23:355–364.
Rosenberger AL, Kinzey WG. 1976. Functional patterns ofmolar occlusion in platyrrhine primates. Am J Phys Anthro-pol 45:281–298.
Salazar-Ciudad I, Jernvall J. 2010. A computational model ofteeth and the developmental origins of morphological varia-tion. Nature 464:583–586.
Seligsohn D. 1977. Analysis of species-specific molar patterns instrepsirhine primates. Contrib Primatol 11:1–116.
Seligsohn D, Szalay FS. 1978. Relationship between naturalselection and dental morphology: tooth function and diet inLepilemur and Hapalemur. In: Butler PM, Joysey KA, editors.Development, function and evolution of teeth. New York: Aca-demic Press. p 289–307.
Skogland T. 1988. Tooth wear by food limitation and its life-his-tory consequences in wild reindeer. Oikos 51:238–242.
17WEAR FACET ORIENTATION IN Propithecus edwardsi
American Journal of Physical Anthropology

Tanner JB, Zelditch ML, Lundrigan BL, Holekamp KE. 2009. Onto-genetic change in skull morphology and mechanical advantage inthe spotted hyena (Crocuta crocuta). J Morphol 271:353–365.
Teaford MF. 1983. The morphology and wear of the lingual notchin macaques and langurs. Am J Phys Anthropol 60:7–14.
Ulhaas L, Kullmer O, Schrenk F, Henke W. 2004. A new 3-dapproach to determine functional morphology of cercopithe-coid molars. Ann Anat 186:487–493.
Ungar PS. 2002. Reconstructing the diets of fossil primates. In:Plavcan JM, Kay RF, Jungers WL, van Schaik CP, editors.Reconstructing behavior in the primate fossil record. NewYork: Kluwer Academic Press. p 261–296.
Ungar PS, M’Kirera F. 2003. A solution to the worn tooth co-nundrum in primate functional anatomy. Proc Natl Acad SciUSA 100:3874–3877.
Ungar PS, Taylor SR. 2005. Dental topographic analysis:tooth wear and function. Am J Phys Anthropol Suppl40:210.
Ungar PS, Williamson M. 2000. Exploring the effects of toothwear on functional morphology: a preliminary study usingdental topographic analysis. Paleontol Electron 3:1–18.
Wright PC. 1995. Demography and life history of free-rangingPropithecus diadema edwardsi in Ranomafana National Park,Madagascar. Int J Primatol 16:835–854.
Wright PC. 1998. Impact of predation risk on the behaviour ofPropithecus diadema edwardsi in the rain forest of Madagas-car. Behaviour 135:483–512.
Young AL, Richard AF, Aiello LC. 1990. Female dominance andmaternal investment in strepsirrhine primates. Am Nat135:473–488.
18 S. BLATCH ET AL.
American Journal of Physical Anthropology