cestodes change the isotopic signature of brine shrimp, artemia, hosts: implications for aquatic...
TRANSCRIPT

International Journal for Parasitology 43 (2013) 73–80
Contents lists available at SciVerse ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
Cestodes change the isotopic signature of brine shrimp, Artemia, hosts: Implicationsfor aquatic food webs
Marta I. Sánchez ⇑, Nico Varo, Cristina Matesanz, Cristina Ramo, Juan A. Amat, Andy J. GreenDepartment of Wetland Ecology, Estación Biológica de Doñana, CSIC, C/Américo Vespucio s/n, E-41092 Sevilla, Spain
a r t i c l e i n f o
Article history:Received 1 August 2012Received in revised form 5 November 2012Accepted 6 November 2012Available online 3 December 2012
Keywords:C and N stable isotopesArtemiaCestodesIntermediate hostParasitismTrophically transmitted parasite
0020-7519/$36.00 � 2012 Australian Society for Parahttp://dx.doi.org/10.1016/j.ijpara.2012.11.003
⇑ Corresponding author. Tel.: +34 954232340; fax:E-mail addresses: [email protected], m
Sánchez).
a b s t r a c t
To reach the final host (greater flamingos), the cestode Flamingolepis liguloides alters the behaviour of itsintermediate host, the brine shrimp, Artemia parthenogenetica, causing it to spend more time close to thewater surface. During summer 2010, we showed that the prevalence of this cestode was consistentlyhigher at the top of the water column in the Odiel salt pans in south-western Spain. We used stable nitro-gen (N) and carbon (C) isotopic analysis to test the hypothesis that cestodes also alter resource use byArtemia. In early summer, we compared stable isotopes in infected hosts at the surface with those fromuninfected hosts at the bottom of the water column. In late summer, we compared infected and unin-fected Artemia from the bottom. d15N was consistently enriched in infected individuals compared withuninfected hosts, especially in Artemia with multiple infections of F. liguloides (family Hymenolepididae)and those with mixed infections of F. liguloides and cestodes of the family Dilepididae. Infected individ-uals from the surface were enriched in d13C compared with uninfected ones from the bottom, but theopposite was found when comparing uninfected and infected Artemia from the same depth. This maybe caused by the increase in lipid concentration in infected Artemia. Isolated cysticercoids of F. liguloideswere significantly enriched in d13C compared with cysticercoids in infected hosts, but surprisingly werenot enriched in N. Our findings illustrate the way cestodes can alter food webs and highlight the impor-tance of considering the parasitic status of prey in studies of trophic ecology in saline wetlands.
� 2012 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Parasites have often been ignored in aquatic food web studies(Lafferty et al., 2008). However recent theoretical and empiricaldata suggest that parasites affect trophic interactions and energyflow within ecosystems in diverse ways (Thompson et al., 2005;Lafferty et al., 2008). Trophically transmitted parasites, which de-pend on predation of their intermediate hosts to complete their lifecycle, have the potential to strongly impact trophic interactionsand food webs. Many parasites using trophic transmission haveevolved sophisticated mechanisms to modify the behaviour andmorphology of their intermediate hosts to increase the probabilityof predation by the final hosts (Moore, 2002; Poulin, 2007).However, the ecological consequences of such manipulations atdifferent trophic levels remain largely unexplored.
Artemia spp. (Branchiopoda; Anostraca) are keystone speciesand dominant macrozooplancton in hypersaline ecosystems(Wurtsbaugh and Gliwicz, 2001) and can control phytoplanktondensity via grazing (Lenz, 1987; Wurtsbaugh, 1992; Gliwicz et al.,2010). Many species of aquatic birds depend on brine shrimps as
sitology Inc. Published by Elsevier
+34 [email protected] (M.I.
a food source (Sánchez et al., 2007a; Varo et al., 2011; Vest andConover, 2011). Moreover, Artemia is the intermediate host for atleast 15 species of cestodes which depend on predation by avianhosts (Georgiev et al., 2005, 2007). Brine shrimps become infectedby ingesting cestode eggs released into the water with the faecesof the avian definitive host. The newly hatched oncosphere (tape-worm embryo) penetrates into the haemocoel, where it developsinto a cysticercoid (the larval stage). Birds become infected via tro-phic transmission when they feed on infected Artemia. Adult tape-worms develop and reproduce in the small intestine of birds,completing their life cycle. A particularly abundant cestode in brineshrimps is Flamingolepis liguloides (Cyclophyllidea, Hymenolepidi-dae), a parasite of flamingos. Flamingolepis liguloides alters the size,fecundity, colouration, physiology and behaviour of Artemia (Amatet al., 1991; Sánchez et al., 2006a, 2007b, 2009a; Amarouayacheet al., 2009). Previous studies show that behavioural changes (posi-tive phototactism) cause alterations in the spatial distribution ofArtemia, with infected shrimps being more abundant at the surfaceof the water and uninfected ones mainly occupying the bottom ofthe water column (Gabrion et al., 1982; Thiéry et al., 1990; Sánchezet al., 2007b).
The aim of the present work was to explore potential trophicconsequences of infection by cestodes in Artemia using C and N sta-ble isotopic analysis (hereafter d13C and d15N, respectively); the
Ltd. All rights reserved.

74 M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80
signature of d15N can provide information about the trophic posi-tion, owing to enrichment through the trophic levels; the signatureof d13C provides information about the dietary source of carbon.Stable isotopes have been increasingly used in studies of avian diet(Hobson and Clark, 1992a,b; Thompson and Furness, 1995;Podlesak and McWilliams, 2006; Gómez-Díaz and Figuerola,2010) and constitute a powerful tool for studies of food webs (Post,2002). However to date, studies of host–parasite relationshipsusing C and N stable isotopes are still scarce (Doi et al., 2008; Du-bois et al., 2009; Gómez-Díaz and González-Solís, 2010) and haveusually suggested that parasites have no effect on the isotopic sig-nature of their hosts (but see Miura et al., 2006).
In this paper we tested the following hypotheses: (i) parasite-induced microhabitat segregation affects d15N and d13C values ofinfected compared with uninfected Artemia. (ii) The extent of thismodification depends on parasite load and taxa. We consideredthe effect of dilepidid cestodes (Cyclophyllidea, Dilepididae) in co-infections with the dominant F. liguloides. Given that both groupsof parasites show different site preferences within the host’s bodywhich likely reflect different feeding strategies (unpublished data),and may cause different behavioural and physiological changes intheir host, we tested whether d13C and d15N signatures were differ-ent between individuals infected with F. liguloides and those co-in-fected with dilepidids. Owing to the dominance of F. liguloides, wewere unable to study Artemia infected only with dilepidids. (iii) Iso-lated F. liguloides cysticercoids will be enriched in d15N comparedwith hosts because parasites are assumed to feed on host tissue.
2. Material and methods
Artemia sampling was conducted in a saltpan of the Odielmarshes (Huelva, southwestern Spain, 37�1502900N, 6�5802500W), asite of international importance for water birds (Sánchez et al.,2006b). The study area (1,118 ha) is a complex of ponds throughwhich the seawater is circulated and evaporated until salt precip-itation occurs. The Odiel brine shrimps are one of many clonal pop-ulations in the Old World, often grouped under the binomenArtemia parthenogenetica (Amat et al., 2005; Muñoz et al., 2010).Samples were collected in an evaporation pond of moderate salin-ity (E18, salinity = 110–120 g/l) during summer 2010. This pond(labelled as I6 in Sánchez et al., 2006c), of constant depth (24–27 cm), is a simple environment with no structure from e.g. rocksor macrophytes (absent owing to the high salinity).
2.1. Distribution of infected and uninfected Artemia in the watercolumn
In order to examine the proportion of infected and uninfectedArtemia at the surface and at depths in the water column, we col-lected samples (between 0900 h and 1,200 h) at 0–5 cm from thesurface and 0–5 cm from the bottom during five different daysfrom 30 June to 3 August 2010 at a fixed point 140 m from theshoreline. Samples were taken with an aquarium net (0.5 mmmesh) at a constant, slow speed for 1 min at depth (higher density)and 5 min at the surface. In the laboratory, each individual wasmounted in a temporary glycerol mount and examined under amicroscope for parasites (after Georgiev et al., 2005).
2.2. Early summer sampling for isotopic analysis: infected from thesurface versus uninfected from the bottom
In order to test whether changes in the spatial distribution ofArtemia induced by parasites had trophic consequences, we col-lected samples for isotopic analysis in early summer (five occasionsfrom 14 June to 5 July 2010 to provide an adequate sample size).
No variations in food availability and biochemical composition ofArtemia likely to affect isotopic composition were expected to oc-cur during such a short sampling period. We collected infectedArtemia from within 5 cm of the surface and uninfected ones fromwithin 5 cm of the bottom with a 0.1 mm mesh plankton net. In thelaboratory, brine shrimps were immediately placed in a tank filledwith filtered (45 lm sieve) water from the same pond for 24 h toremove all food particles and to allow digestion of previously in-gested food. Individual Artemia were then inspected under the ste-reomicroscope for the presence of parasites. More than 500specimens where checked on each sampling date so as to reach aminimum of 475 individuals infected only with F. liguloides (mixedinfection with other cestodes is very frequent; Georgiev et al.,2007). We always selected individual adult shrimps of the samesize range to minimise age variation (8–10 mm, measured fromthe anterior margin of the head to the end of caudal furca).
Four categories were selected for isotopic analysis on the basis oftheir abundance in the field: no parasites NP (n = 18 samples), in-fected with one F. liguloides (1FL, n = 7 samples), infected with morethan one F. liguloides (>1FL, 2-14 cysticercoids per shrimp, n = 32samples) and infected with both F. liguloides + Dilepidiae (FL + DIL1-2 cysticercoids of F. liguloides and 1-2 cysticercoids of Dilepididae,n = 12 samples). Each sample was necessarily made from 10–20individuals so as to provide enough material for isotopic analysis.It was not practical to divide the Artemia with multiple infectionsof F. liguloides (between two and 14 cysticercoids) into more catego-ries, owing to the very low number of individuals of intermediatecategories recorded, and the need to pool several individuals toachieve an acceptable weight for isotopic analysis (25 mg). Wechose the categories of 1FL and >1FL because our unpublished datasuggest that host phenotype is affected by an increase from one totwo F. liguloides cysticercoids, but not by a further increase (e.g. redcolouration becomes significantly more intense with an increasebetween one and two cysticercoids, but not with a further increase).
In a field sample collected near the surface on 30 June and 5 July(n = 259) and quantified, the proportions of Artemia in the fourstudy categories were 29% NP, 18.1% 1FP, 32% >1FL and 4.2%FL + DIL; 16.6% corresponded to other categories of infectedshrimps (i.e. presence of additional cestode species, and 74.4% ofthose individuals were co-infected with F. liguloides). We werenot able to distinguish between the three dilepidid species infect-ing Artemia (Eurycestus avoceti, Anomotaenia tringae and Anomotae-nia microphallos) because this would require dissection whichwould cause a loss of haemolymph. Cysticercoids of F. liguloidesare easily recognisable without dissection (Georgiev et al., 2005).
From samples taken on the same dates, we also isolated cystic-ercoids of F. liguloides from the body of Artemia (n = 10 samples,each sample containing 2,000 cysticercoids). These cysticercoidswere extracted from many samples of Artemia from the top ofthe water column, which were then minced and sieved to extractthe relatively hard cysticercoids. However, hosts were not con-served after cysticercoid removal as most haemolymph was lostin the dissection. Although a given sample may have containedmultiple cysticercoids from the same individual host, each hostcould not be represented in more than one sample, thus avoidingnon-independence. The isotopic signatures of cysticercoids werecompared with those of intact, uninfected and infected hosts.Cysticercoids of Dilepididae are much smaller and show a lowprevalence in Artemia (Georgiev et al., 2005, 2007) so isolation ofan adequate quantity was not feasible.
2.3. Late summer sampling for isotopic analysis: infected from thebottom versus uninfected from the bottom
In late summer (8 September 2010) we collected an additionalsample of Artemia from the bottom in order to compare C and N
M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80 75
ratios of infected and uninfected individuals at the same depth. Theaim was to test the consequences of parasitism for isotopic signa-ture irrespective of those caused by microhabitat segregation. Hostcategories were similar to early summer except that there wereinsufficient individuals of the FL + DIL category for analysis. Hencewe obtained NP (n = 25 samples, each made of five individuals), FL(n = 11 samples) and >1FL (n = 10 samples). In the laboratory thesamples were processed as described in Section 2.2.
Prior to isotopic analysis, no lipid extraction was carried out.Each sample was dried at 60 �C for 24 h and ground to a fine pow-der using a vibratory ball mill at 30 s�1 for 25 s. See SupplementaryData S1 for detailed information on methods for isotopic analysis.
2.4. Statistical analysis
We used Chi-square tests to compare the proportions of Artemiainfected with F. liguloides between the surface and the bottom ofthe water column (vertical distribution).
Comparisons of d15N between uninfected and different catego-ries of infected Artemia were performed with one-way ANOVA, fol-lowed by Tukey’s HSD (Honestly Significant Difference) post hoctests. Kruskall Wallis tests followed by Mann–Whitney U-testswere used to compare d13C between categories in early summerwhen normality assumptions were not met, even after log andother data transformations. Similarly, comparisons of isolatedcysticercoids and Artemia were made with t-tests for d15N, andMann–Whitney U-tests for d13C. P values were always adjustedfor multiple comparisons via false discovery rate (FDR; Benjaminiand Hochberg, 1995). All statistical analyses were conducted usingStatistica 6.0 (StatSoft 2001). P values less that 0.05 were consid-ered statistically significant.
3. Results
3.1. Vertical distribution analysis
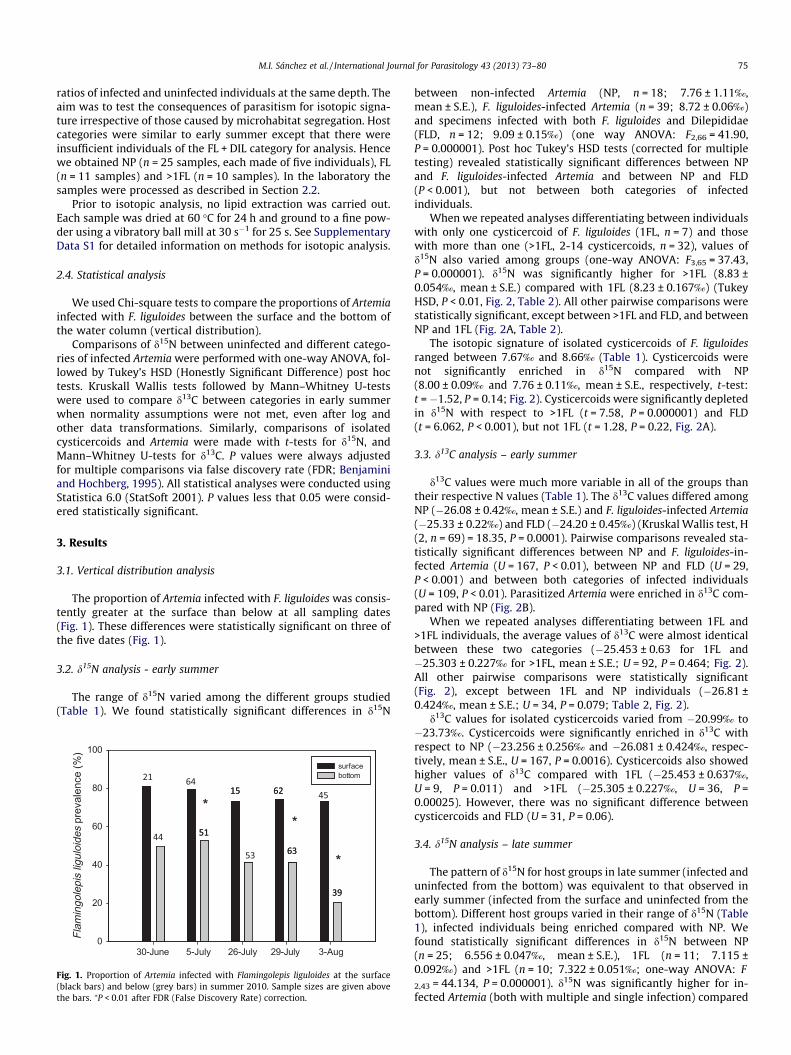
The proportion of Artemia infected with F. liguloides was consis-tently greater at the surface than below at all sampling dates(Fig. 1). These differences were statistically significant on three ofthe five dates (Fig. 1).
3.2. d15N analysis - early summer
The range of d15N varied among the different groups studied(Table 1). We found statistically significant differences in d15N
100surface bottom
%)
80
*
nce
(
60 *reva
lede
sp
40 *ligul
oile
pis
20
min
goFl
a
30-June 5-July 26-July 29-July 3-Aug0
Fig. 1. Proportion of Artemia infected with Flamingolepis liguloides at the surface(black bars) and below (grey bars) in summer 2010. Sample sizes are given abovethe bars. ⁄P < 0.01 after FDR (False Discovery Rate) correction.
between non-infected Artemia (NP, n = 18; 7.76 ± 1.11‰,mean ± S.E.), F. liguloides-infected Artemia (n = 39; 8.72 ± 0.06‰)and specimens infected with both F. liguloides and Dilepididae(FLD, n = 12; 9.09 ± 0.15‰) (one way ANOVA: F2,66 = 41.90,P = 0.000001). Post hoc Tukey’s HSD tests (corrected for multipletesting) revealed statistically significant differences between NPand F. liguloides-infected Artemia and between NP and FLD(P < 0.001), but not between both categories of infectedindividuals.
When we repeated analyses differentiating between individualswith only one cysticercoid of F. liguloides (1FL, n = 7) and thosewith more than one (>1FL, 2-14 cysticercoids, n = 32), values ofd15N also varied among groups (one-way ANOVA: F3,65 = 37.43,P = 0.000001). d15N was significantly higher for >1FL (8.83 ±0.054‰, mean ± S.E.) compared with 1FL (8.23 ± 0.167‰) (TukeyHSD, P < 0.01, Fig. 2, Table 2). All other pairwise comparisons werestatistically significant, except between >1FL and FLD, and betweenNP and 1FL (Fig. 2A, Table 2).
The isotopic signature of isolated cysticercoids of F. liguloidesranged between 7.67‰ and 8.66‰ (Table 1). Cysticercoids werenot significantly enriched in d15N compared with NP(8.00 ± 0.09‰ and 7.76 ± 0.11‰, mean ± S.E., respectively, t-test:t = �1.52, P = 0.14; Fig. 2). Cysticercoids were significantly depletedin d15N with respect to >1FL (t = 7.58, P = 0.000001) and FLD(t = 6.062, P < 0.001), but not 1FL (t = 1.28, P = 0.22, Fig. 2A).
3.3. d13C analysis – early summer
d13C values were much more variable in all of the groups thantheir respective N values (Table 1). The d13C values differed amongNP (�26.08 ± 0.42‰, mean ± S.E.) and F. liguloides-infected Artemia(�25.33 ± 0.22‰) and FLD (�24.20 ± 0.45‰) (Kruskal Wallis test, H(2, n = 69) = 18.35, P = 0.0001). Pairwise comparisons revealed sta-tistically significant differences between NP and F. liguloides-in-fected Artemia (U = 167, P < 0.01), between NP and FLD (U = 29,P < 0.001) and between both categories of infected individuals(U = 109, P < 0.01). Parasitized Artemia were enriched in d13C com-pared with NP (Fig. 2B).
When we repeated analyses differentiating between 1FL and>1FL individuals, the average values of d13C were almost identicalbetween these two categories (�25.453 ± 0.63 for 1FL and�25.303 ± 0.227‰ for >1FL, mean ± S.E.; U = 92, P = 0.464; Fig. 2).All other pairwise comparisons were statistically significant(Fig. 2), except between 1FL and NP individuals (�26.81 ±0.424‰, mean ± S.E.; U = 34, P = 0.079; Table 2, Fig. 2).
d13C values for isolated cysticercoids varied from �20.99‰ to�23.73‰. Cysticercoids were significantly enriched in d13C withrespect to NP (�23.256 ± 0.256‰ and �26.081 ± 0.424‰, respec-tively, mean ± S.E., U = 167, P = 0.0016). Cysticercoids also showedhigher values of d13C compared with 1FL (�25.453 ± 0.637‰,U = 9, P = 0.011) and >1FL (�25.305 ± 0.227‰, U = 36, P =0.00025). However, there was no significant difference betweencysticercoids and FLD (U = 31, P = 0.06).
3.4. d15N analysis – late summer
The pattern of d15N for host groups in late summer (infected anduninfected from the bottom) was equivalent to that observed inearly summer (infected from the surface and uninfected from thebottom). Different host groups varied in their range of d15N (Table1), infected individuals being enriched compared with NP. Wefound statistically significant differences in d15N between NP(n = 25; 6.556 ± 0.047‰, mean ± S.E.), 1FL (n = 11; 7.115 ±0.092‰) and >1FL (n = 10; 7.322 ± 0.051‰; one-way ANOVA: F2,43 = 44.134, P = 0.000001). d15N was significantly higher for in-fected Artemia (both with multiple and single infection) compared

Table 1Range of d15N and d13C (‰) in non-infected Artemia (NP), Artemia infected with one Flamingolepis liguloides (1FL), Artemia infected with more than one F. liguloides (>1FL), infectedby both F. liguloides and Dilepididae (FLD) and isolated cysticercoids of F. liguloides (Cy FL) in early summer (June–July). n = number of pooled samples, each of which is made of10–20 individuals.
Early summer Late summer
n d15N range (‰) d13C range (‰) n d15N range (‰) d13C range (‰)
NP 18 (6.97, 8.32) (�27.61, �21.79) 25 (6.07, 7.01) (�27.72, �25.30)1FL 7 (7.55, 8.65) (�26.80, �21.80) 11 (6.81, 7.84) (�29.01, �25.13)>1FL 32 (8.26, 9.65) (�26.65, �22.28) 10 (7.11, 7.54) (�28.83, �26.68)FLD 12 (8.41, 10.03) (�27.61, �21.79) . . . . . . . . .
Cy FL 10 (7.67, 8.66) (�23.73, �20.99) . . . . . . . . .
(A)
NP 1FL >1FL FL+DIL C
15N
(‰)
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
(B)
NP 1FL >1FL FL+DIL C
13C
(‰)
-28
-27
-26
-25
-24
-23
-22
Fig. 2. Mean (±S.E.) values for (A) d15N and (B) d13C in early and late summer fornon-infected Artemia (NP), those infected with one cysticercoid of Flamingolepisliguloides (1FL), infected with more than one cysticercoid of F. liguloides (>1FL),infected with both F. liguloides and Dilepididae (FL + DIL), and isolated cysticercoidsof F. liguloides (C). Squares are samples from early summer, circles from latesummer. Solid symbols are samples from the surface; empty symbols are samplesfrom the bottom.
76 M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80
with NP (Tukey HSD, P < 0.001; Fig. 1A, Table 2) but not between1FL and >1FL.
3.5. d13C analysis – late summer
Contrary to the pattern in early summer (when infected Artemiacame from the surface) individuals infected with >1FL showed themost negative d13C values (�27.438 ± 0.210‰, mean d S.E.), fol-lowed by 1FL (�26.982 ± 0.398‰) and NP (�26.462 ± 0.118‰,mean ± S.E.) (Table 1, Fig. 2B). The differences between categorieswere statistically significant (one-way ANOVA: F 2,43 = 5.261,P = 0.009). Pairwise comparisons revealed statistically significantdifferences between NP and >1FL (Table 2).
4. Discussion
The present study revealed strong differences in the isotopicsignatures of C and N between infected and uninfected Artemia.Using C isotopes, Miura et al. (2006) found that trematode infec-tion changed the habitat use and trophic ecology of marine gastro-pods, which moved to the lower intertidal zone, where they weremore susceptible to predation by their final hosts (fish). However,other studies have reported no differences in stable isotopes ofinvertebrate hosts when infected by trematodes (Doi et al., 2008;Dubois et al., 2009).
We interpret our results in a similar manner to Miura et al.(2006), and suggest that infection by cestodes has direct trophicconsequences for their brine shrimp host, at least partly becausethe parasite induces movement towards the water surface. As ex-pected from existing literature, prevalence of infected Artemia(with F. liguloides) was higher at the surface of the water. Artemiaparthenogenetica undergo diel vertical migrations and typicallyspend most of the daytime in the bottom 25% of the water columnand most of the night in the other 75% (Britton et al., 1986). Eachindividual does not stay at a fixed depth throughout the daily cycle,and cestode infection changes the proportion of time spent at dif-ferent depths (Sánchez et al., 2007b). The change in Artemia trophicecology appears to be one of a suite of interdependent host charac-teristics changed by infection, including colour, behaviour and lipidaccumulation (Sánchez et al., 2006a, 2007b, 2009a). The differ-ences found between infected and uninfected shrimps collected to-gether from the bottom of the water column can most likely beexplained by the infected shrimps having spent more of their timeat the top of the water column than the neighbouring uninfectedshrimps (and hence have consumed different proportions of avail-able food items), even though they were close together at the mo-ment of sampling.
An alternative explanation for the link between infection andisotopic signature would be that it is an effect of inherent differ-ences between individual shrimps in trophic ecology, not a cause;i.e. that some individuals feed at a higher trophic level in such away that they are more likely to ingest cestode eggs and becomeinfected. We cannot eliminate this possibility without conductingexperimental infections in the laboratory, but we consider it unli-kely. In any case, by comparing infected and uninfected shrimpsfrom the same depth, we have shown that infection status is amore important determinant of isotopic signature than positionin the water column at the time of sampling.
Stable isotopic ratios of a parasite–host system can potentiallyshift due to the direct effect of parasite biomass (‘mass-balance’shift, Dubois et al., 2009). Changes in isotopic ratios betweenhealthy and parasitized hepatopancreas of freshwater snails havebeen attributed to the high proportion of trematode biomass (upto 3.4%) within this organ (Doi et al., 2008). Given our inability toremove cysticercoids from Artemia without losing host fluids, weestimated the contribution of the parasite to the total host biomassto evaluate whether the presence of parasites has an important
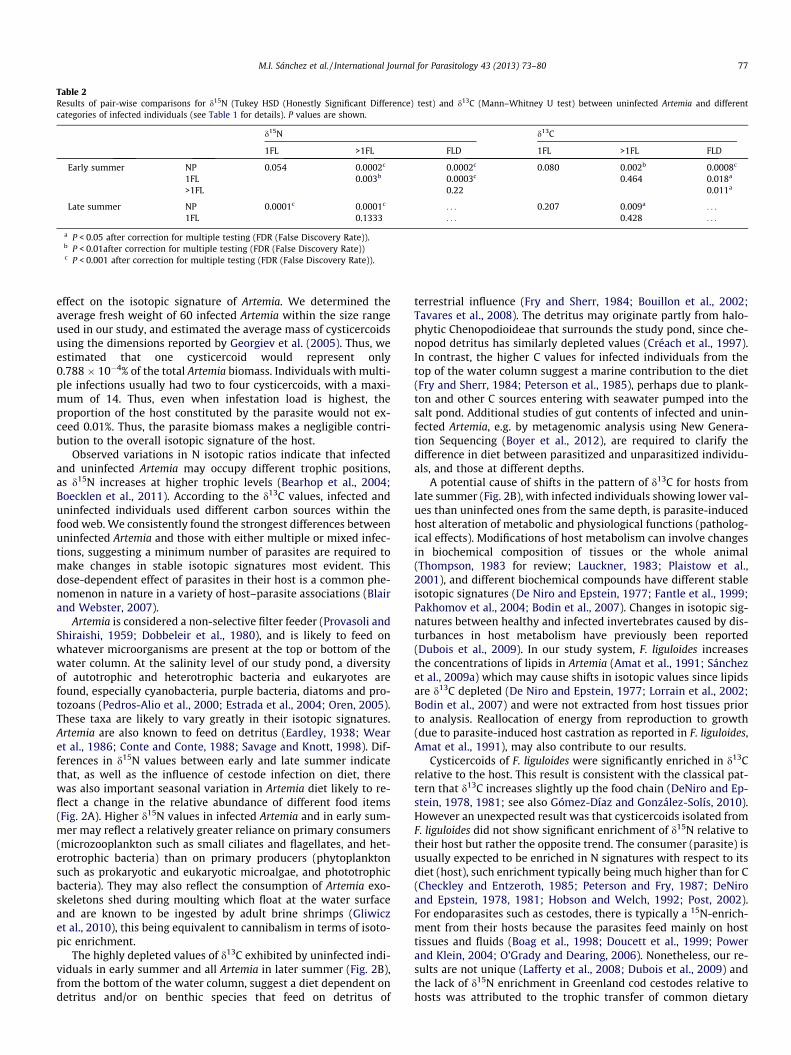
Table 2Results of pair-wise comparisons for d15N (Tukey HSD (Honestly Significant Difference) test) and d13C (Mann–Whitney U test) between uninfected Artemia and differentcategories of infected individuals (see Table 1 for details). P values are shown.
d15N d13C
1FL >1FL FLD 1FL >1FL FLD
Early summer NP 0.054 0.0002c 0.0002c 0.080 0.002b 0.0008c
1FL 0.003b 0.0003c 0.464 0.018a
>1FL 0.22 0.011a
Late summer NP 0.0001c 0.0001c . . . 0.207 0.009a . . .
1FL 0.1333 . . . 0.428 . . .
a P < 0.05 after correction for multiple testing (FDR (False Discovery Rate)).b P < 0.01after correction for multiple testing (FDR (False Discovery Rate))c P < 0.001 after correction for multiple testing (FDR (False Discovery Rate)).
M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80 77
effect on the isotopic signature of Artemia. We determined theaverage fresh weight of 60 infected Artemia within the size rangeused in our study, and estimated the average mass of cysticercoidsusing the dimensions reported by Georgiev et al. (2005). Thus, weestimated that one cysticercoid would represent only0.788 � 10�4% of the total Artemia biomass. Individuals with multi-ple infections usually had two to four cysticercoids, with a maxi-mum of 14. Thus, even when infestation load is highest, theproportion of the host constituted by the parasite would not ex-ceed 0.01%. Thus, the parasite biomass makes a negligible contri-bution to the overall isotopic signature of the host.
Observed variations in N isotopic ratios indicate that infectedand uninfected Artemia may occupy different trophic positions,as d15N increases at higher trophic levels (Bearhop et al., 2004;Boecklen et al., 2011). According to the d13C values, infected anduninfected individuals used different carbon sources within thefood web. We consistently found the strongest differences betweenuninfected Artemia and those with either multiple or mixed infec-tions, suggesting a minimum number of parasites are required tomake changes in stable isotopic signatures most evident. Thisdose-dependent effect of parasites in their host is a common phe-nomenon in nature in a variety of host–parasite associations (Blairand Webster, 2007).
Artemia is considered a non-selective filter feeder (Provasoli andShiraishi, 1959; Dobbeleir et al., 1980), and is likely to feed onwhatever microorganisms are present at the top or bottom of thewater column. At the salinity level of our study pond, a diversityof autotrophic and heterotrophic bacteria and eukaryotes arefound, especially cyanobacteria, purple bacteria, diatoms and pro-tozoans (Pedros-Alio et al., 2000; Estrada et al., 2004; Oren, 2005).These taxa are likely to vary greatly in their isotopic signatures.Artemia are also known to feed on detritus (Eardley, 1938; Wearet al., 1986; Conte and Conte, 1988; Savage and Knott, 1998). Dif-ferences in d15N values between early and late summer indicatethat, as well as the influence of cestode infection on diet, therewas also important seasonal variation in Artemia diet likely to re-flect a change in the relative abundance of different food items(Fig. 2A). Higher d15N values in infected Artemia and in early sum-mer may reflect a relatively greater reliance on primary consumers(microzooplankton such as small ciliates and flagellates, and het-erotrophic bacteria) than on primary producers (phytoplanktonsuch as prokaryotic and eukaryotic microalgae, and phototrophicbacteria). They may also reflect the consumption of Artemia exo-skeletons shed during moulting which float at the water surfaceand are known to be ingested by adult brine shrimps (Gliwiczet al., 2010), this being equivalent to cannibalism in terms of isoto-pic enrichment.
The highly depleted values of d13C exhibited by uninfected indi-viduals in early summer and all Artemia in later summer (Fig. 2B),from the bottom of the water column, suggest a diet dependent ondetritus and/or on benthic species that feed on detritus of
terrestrial influence (Fry and Sherr, 1984; Bouillon et al., 2002;Tavares et al., 2008). The detritus may originate partly from halo-phytic Chenopodioideae that surrounds the study pond, since che-nopod detritus has similarly depleted values (Créach et al., 1997).In contrast, the higher C values for infected individuals from thetop of the water column suggest a marine contribution to the diet(Fry and Sherr, 1984; Peterson et al., 1985), perhaps due to plank-ton and other C sources entering with seawater pumped into thesalt pond. Additional studies of gut contents of infected and unin-fected Artemia, e.g. by metagenomic analysis using New Genera-tion Sequencing (Boyer et al., 2012), are required to clarify thedifference in diet between parasitized and unparasitized individu-als, and those at different depths.
A potential cause of shifts in the pattern of d13C for hosts fromlate summer (Fig. 2B), with infected individuals showing lower val-ues than uninfected ones from the same depth, is parasite-inducedhost alteration of metabolic and physiological functions (patholog-ical effects). Modifications of host metabolism can involve changesin biochemical composition of tissues or the whole animal(Thompson, 1983 for review; Lauckner, 1983; Plaistow et al.,2001), and different biochemical compounds have different stableisotopic signatures (De Niro and Epstein, 1977; Fantle et al., 1999;Pakhomov et al., 2004; Bodin et al., 2007). Changes in isotopic sig-natures between healthy and infected invertebrates caused by dis-turbances in host metabolism have previously been reported(Dubois et al., 2009). In our study system, F. liguloides increasesthe concentrations of lipids in Artemia (Amat et al., 1991; Sánchezet al., 2009a) which may cause shifts in isotopic values since lipidsare d13C depleted (De Niro and Epstein, 1977; Lorrain et al., 2002;Bodin et al., 2007) and were not extracted from host tissues priorto analysis. Reallocation of energy from reproduction to growth(due to parasite-induced host castration as reported in F. liguloides,Amat et al., 1991), may also contribute to our results.
Cysticercoids of F. liguloides were significantly enriched in d13Crelative to the host. This result is consistent with the classical pat-tern that d13C increases slightly up the food chain (DeNiro and Ep-stein, 1978, 1981; see also Gómez-Díaz and González-Solís, 2010).However an unexpected result was that cysticercoids isolated fromF. liguloides did not show significant enrichment of d15N relative totheir host but rather the opposite trend. The consumer (parasite) isusually expected to be enriched in N signatures with respect to itsdiet (host), such enrichment typically being much higher than for C(Checkley and Entzeroth, 1985; Peterson and Fry, 1987; DeNiroand Epstein, 1978, 1981; Hobson and Welch, 1992; Post, 2002).For endoparasites such as cestodes, there is typically a 15N-enrich-ment from their hosts because the parasites feed mainly on hosttissues and fluids (Boag et al., 1998; Doucett et al., 1999; Powerand Klein, 2004; O’Grady and Dearing, 2006). Nonetheless, our re-sults are not unique (Lafferty et al., 2008; Dubois et al., 2009) andthe lack of d15N enrichment in Greenland cod cestodes relative tohosts was attributed to the trophic transfer of common dietary

78 M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80
sources (Power and Klein, 2004). The inability of some endopara-sites to synthesise amino acids and the need to take them fromtheir host (Deudero et al., 2002) has also been proposed to explainslight or no d15N trophic enrichment of parasites (Dubois et al.,2009). No enrichment in N may also be explained by the abbrevi-ated metabolic pathways of parasites and the low rate of excretion,tegument diffusion and respiration (Dubois et al., 2009). All ofthese explanations may apply to our system. Owing to the needto pool large numbers of cysticercoids from different hosts to haveenough samples for analysis, we do not know the exact infectiontypes to which these cysticercoids belonged (but see Section 2for the relative abundance of different infection types). Hence,although they were significantly depleted compared with Artemiawith multiple and mixed infections, it remains unclear whetherthey were depleted compared with their hosts on an individualbasis.
We also found substantial differences in the level of enrichmentof C and N among Artemia infected with F. liguloides and those co-infected with metacestodes from three species of the family Dile-pididae. The isotopic enrichment of Artemia with mixed infection(F. liguloides + Dilepididae) compared with individuals infectedwith F. liguloides may be due to a dose-dependent effect since ahost with a mixed infection harbours at least two parasites. Never-theless, it is possible that cestodes from different families usesdifferent resources in their host or alter host metabolism in a dif-ferent way. While dilepidid cysticercoids are mainly found in theadult thorax, F. liguloides cysticercoids are predominantly locatedin the abdomen of adult Artemia (unpublished data; Amarouayacheet al., 2009; Redón et al., 2011).
Changes in spatial distribution of Artemia induced by parasitesappeared to change resource consumption. Given the abundanceof Artemia and their importance as a food source for birds, cestodeparasites have the potential to alter food web properties, energyand mass flows in hypersaline ecosystems. On the other hand, asmajor consumers of phytoplankton and organic particulate matterin hypersaline waters, alterations of the diet in Artemia can poten-tially cause strong cascading effects through alteration of grazingeffects on phytoplankton and bacterioplankton. Feeding by Artemiainfluences phytoplankton density and composition (Mohebbi,2010), although there is no information on the influence of para-sites on grazing. Higher grazing rates by infected individuals atthe surface of the water may decrease phytoplankton abundance,thus increasing water clarity (Lenz, 1987; Wurtsbaugh, 1992;Gliwicz et al., 2010). Augmentation of light passing through thewater column is likely to affect the algal–cyanobacteria commu-nity at the bottom of the water column. For example, it might fa-vour phototrophic bacteria which are common beneath thecyanobacterial layers in hypersaline microbial mats (DasSarmaand Arora, 2001). Laboratory grazing experiments with infectedand uninfected Artemia would provide further insight into theecological implications of parasite-induced changes in the diet ofArtemia.
This study shows that cestodes can induce strong changes inisotopic signatures of their intermediate hosts, and that infectedand uninfected invertebrates are not equivalent from a trophicpoint of view. The parasitic status of invertebrates should bechecked and incorporated into aquatic food web research and stud-ies of trophic ecology and diet of birds. Given that stable isotopescan be used to assess the importance of specific prey in the dietof birds (Alexander et al., 1996; Tavares et al., 2009), this techniquecan evaluate the importance of infected and uninfected prey as afood source for birds. On the other hand, in studies of avian ecol-ogy, invertebrates are typically sampled as potential bird preywithout considering the presence of helminth parasites and theirpotential effects on isotopic signatures. This may be an importantsource of error when determining the part that each invertebrate
taxon plays in an avian diet. As a consequence of their manipula-tions of host colour and behaviour, birds are particularly likely toingest invertebrates that have been parasitized (Sánchez et al.,2009b). The prevalence of parasites is likely to vary between thoseinvertebrates ingested by birds and those collected by biologists aspotential food items, since the sampling methods may be very dif-ferent. Our work shows it is very important to take into accountthe parasite communities in those taxa, and how they can influ-ence the isotopic ratios of avian prey.
Acknowledgements
We thank Javier Díaz Real for help collecting samples, and M.G.Forero and R. Álvarez Alonso for helpful discussion. Two anony-mous reviewers made helpful suggestions that allowed us to im-prove the manuscript. Enrique Martínez, Director of ParajeNatural Marismas del Odiel, provided permission to work in thesalt ponds. M.I. Sánchez was supported by a Juan de la Cierva post-doctoral contract from the Spanish Ministry of Science and Innova-tion (MICINN). This study was funded by project P07-CVI-02700from the Consejería de Innovación, Ciencia y Empresa (Junta deAndalucía) and project CGL2010-16028 from MICINN.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.ijpara.2012.11.003.
References
Alexander, S.A., Hobson, K.A., Gratto-Trevor, C., Diamond, A.W., 1996. Conventionaland isotopic determinations of shorebird diets at an inland stopover: theimportance of invertebrates and Potamogeton pectinatus turbers. Can. J. Zool. 74,1057–1068.
Amarouayache, M., Farid, D., Hichem, K.M., 2009. The parasitism of Flamingolepisliguloides (Gervais, 1847) (Cestoda, Hymenolepididae) in Artemia salina(Crustacea, Branchiopoda) in two saline lakes in Algeria. Acta Parasitol. 54,330–334.
Amat, F., Gozalbo, A., Navarro, J.C., Hontoria, F., Varó, I., 1991. Some aspects ofArtemia biology affected by cestode parasitism. Hydrobiologia 212, 39–44.
Amat, F., Hontoria, F., Ruiz, O., Green, A.J., Sánchez, M.I., Figuerola, J., Hortas, F., 2005.The American brine shrimp Artemia franciscana as an exotic invasive species inthe Western Mediterranean. Biol. Inv. 7, 37–47.
Bearhop, S., Adams, C.E., Waldron, S., Fuller, R.A., Macleod, H., 2004. Determiningtrophic niche width: a novel approach using stable isotope analysis. J. Anim.Ecol. 73, 1007–1012.
Benjamini, Y., Hochberg, Y., 1995. Controlling the false discovery rate: a practicaland powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 57, 289–300.
Blair, L., Webster, J.P., 2007. Dose-dependent schistosome-induced mortality andmorbidity risk elevates host reproductive effort. J. Evol. Biol. 20, 54–61.
Boag, B., Neilson, R., Robinson, D., Scrimgeour, C.M., Handley, L.L., 1998. Wild rabbithost and some parasites show trophic level relationships for d13C and d15N: afirst report. Isot. Environ. Health. Stud. 34, 81–85.
Bodin, N., Le Loc’h, F., Hily, C., 2007. Effect of lipid removal on carbon and nitrogenstable isotope ratios in crustacean tissues. J. Exp. Mar. Biol. Ecol. 341, 168–175.
Boecklen, W.J., Christopher, T.Y., Cook, B.A., James, A.C., 2011. On the use of stableisotopes in trophic ecology. Ann. Rev. Ecol. Evol. Syst. 42, 411–440.
Bouillon, S.N., Raman, A.V., Dauby, P., Dehairs, F., 2002. Carbon and nitrogen stableisotope ratios of subtidal benthic invertebrates in an estuarine mangroveecosystem (Andhra Pradesh, India). Estuar. Coast Shelf Sci. 54, 901–913.
Boyer, S., Brown, S.D.J., Collins, R.A., Cruickshank, R.H., Lefort, M.-C., Malumbres-Olarte, J., 2012. Sliding window analyses for optimal selection of mini-barcodes,and application to 454-pyrosequencing for specimen identification fromdegraded DNA. PLoS One 7, e38215. http://dx.doi.org/10.1371/journal.pone.0038215.
Britton, R.H., de Groot, E.R., Johnson, A.R., 1986. The daily cycle of feeding activity ofthe Greater Flamingo in relation to the dispersion of the prey Artemia. Wildfowl37, 151–155.
Checkley, D.M., Entzeroth, L.C., 1985. Elemental and isotopic fractionation of carbonand nitrogen by marine, planktonic copepods and implications to the marinenitrogen cycle. J. Plankton Res. 7, 553–568.
Conte, F., Conte, P., 1988. Abundance and spatial distribution of Artemia salina inLake Abert, Oregon. Hydrobiologia 158, 167–172.

M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80 79
Créach, V., Schrecke, M.T., Bertru, G., Mariotti, A., 1997. Stable isotopes and gutanalyses to determine feeding relationships in saltmarsh macroconsumers.Estuar. Coast Shelf Sci. 44, 599–611.
DasSarma, S., Arora, P., 2001. Halophiles. Encyclopedia of Life Sciences. MacmillanPress, London.
De Niro, M.J., Epstein, S., 1977. Mechanism of carbon isotope fractionationassociated with lipid synthesis. Science 197, 261–263.
DeNiro, M.J., Epstein, S., 1978. Influence of diet on the distribution of carbonisotopes in animals. Geochim. Cosmochim. Acta 42, 495–506.
DeNiro, M.J., Epstein, S., 1981. Influence of diet on the distribution of nitrogenisotopes in animals. Geochim. Cosmochim. Acta 45, 341–351.
Deudero, S., Pinnegar, J.K., Polunin, N.V.C., 2002. Insights into fish host–parasitetrophic relationships revealed by stable isotope analysis. Dis. Aquat. Org. 52,77–86.
Dobbeleir, J., Adam, N., Bossuyt, E., Bruggeman, E., Sorgeloos, P., 1980. New aspectsof the use of inert diets for high density culturing of brine shrimp. In: Persoone,G., Sorgeloos, P., Roels, O., Jaspers, E. (Eds.), The Brine Shrimp Artemia. Ecology,Culturing, Use in Aquaculture, vol. 3. Universa, Wetteren Belgium, pp. 165–174.
Doi, H., Yurlova, N.I., Vodyanitskaya, S.N., Kikuchi, E., Shikano, S., Yadrenkina, E.N.,Zuykova, E.I., 2008. Parasite-induced changes in nitrogen isotope signatures ofhost tissues. J. Parasitol. 94, 292–295.
Doucett, R.R., Giberson, D.J., Power, G., 1999. Parasitic association of Nanocladius(Diptera: Chironomidae) and Pteronarcys biloba (Plecoptera: Pteronarcyidae):insights from stable-isotope analysis. J. North Am. Benthol. Soc. 18, 514–523.
Dubois, S.Y., Savoye, N., Sauriau, P.-G., Billy, I., Martinez, P., de Montaudouin, X.,2009. Digenean trematodes–marine mollusc relationships: a stable isotopestudy. Dis. Aquat. Organ. 84, 65–77.
Eardley, A.J., 1938. Sediments of Great Salt lake. Utah. Bull. Am. Soc. Petrol. Geol. 22,1344–1346.
Estrada, M., Henriksen, P., Gasol, J.M., Casamayor, E.O., Pedros-Alio, C., 2004.Diversity of planktonic photo auto trophic microorganisms along a salinitygradient as depicted by microscopy, flow cytometry, pigment analysis andDNA-based methods. FEMS Microbiol. Ecol. 49, 281–293.
Fantle, M.S., Dittel, A.I., Scwalm, S.M., Epifanio, Ch.E., Fogel, M.L., 1999. A food webanalysis of the juvenile blue crab, Callinectes sapidus, using stable isotopes inwhole animals and individual amino acids. Oecologia 120, 416–426.
Fry, B.M., Sherr, E.B., 1984. 13C measurements as indicators of carbon flow in marineand freshwater ecosystems. Contrib. Mar. Sci. 27, 13–47.
Gabrion, C., MacDonald-Crivelli, G., Boy, V., 1982. Dynamique des populationslarvaires du cestode Flamingolepis liguloides dans une population d’Artemia enCamargue. Acta Oecol. 3, 273–293.
Georgiev, B.B., Sánchez, M.I., Green, A.J., Nikolov, P.N., Vasileva, G.P., Mavrodieva,R.S., 2005. Cestodes from Artemia parthenogenetica (Crustacea, Branchiopoda) inthe Odiel Marshes, Spain: a systematic survey. Acta Parasitol. 50, 105–117.
Georgiev, B.B., Sánchez, M.I., Vasileva, G.P., Nikolov, P.N., Green, A.J., 2007. Cestodeparasitism in invasive and native brine shrimps (Artemia spp.): can it explainthe rapid invasion of A. franciscana in the Mediterranean region? Parasitol. Res.101, 1647–1655.
Gliwicz, Z.M., Wursbaugh, W.A., Szymanska, E., 2010. Absence of predationeliminates coexistence. experience from the fish–zooplankton interface.Hydrobiologia 653, 103–117.
Gómez-Díaz, E., Figuerola, J., 2010. New perspectives in tracing vector-borneinteraction networks. Trends Parasitol. 26, 470–476.
Gómez-Díaz, E., González-Solís, J., 2010. Trophic structure in a seabird host–parasitefood web: insights from stable isotope analyses. Plos One 5, e10454.
Hobson, K.A., Welch, H.E., 1992. Determination of trophic relationships within ahigh Arctic marine food web using 13C and 15N analysis. Mar. Ecol. Prog. Ser. 84,9–18.
Hobson, K.A., Clark, R.G., 1992a. Assessing avian diets using stable isotopes I:turnover of 13C in tissues. Condor 94, 181–188.
Hobson, K.A., Clark, R.G., 1992b. Assessing avian diets using stable isotopes II:factors influencing diet-tissue fractionation. Condor 94, 189–197.
Lafferty, K., Allesina, S., Arim, M., Briggs, C., De Leo, G., Dobson, A., Dunne, J., Johnson,P., Kuris, A., Marcogliese, D.J., Martinez, N.D., Marquet, P.A., McLaughlin, J.P.,Mordecai, E.A., Pascual, M., Poulin, R., Thieltges, D.W., 2008. Parasites in foodwebs: the ultimate missing links. Ecol. Lett. 11, 533–546.
Lauckner, G., 1983. Diseases of Mollusca: Bivalvia. In: Kinne, O. (Ed.), Diseases ofMarine Animals, Introduction, Bivalvia to Scaphopoda, vol. II. BiologischeAnstalt Helgoland, Hamburg, pp. 829–830.
Lenz, P.H., 1987. Ecological studies on Artemia: a review. In: Sorgeloos, P., Bengston,D.A., Decleir, W., Jaspers, E. (Eds.), Artemia Research and its. Applications, 3Ecology, Culturing, Use in Aquaculture. Universal Press, Wetteren, Belgium.
Lorrain, A., Paulet, Y.M., Chauvaud, L., Savoye, N., Donval, A., Saout, C., 2002.Differential d13C and d15N signatures among scallop tissues: implications forecology and physiology. J. Exp. Mar. Biol. Ecol. 275, 47–61.
Miura, O., Kuris, A.M., Torchin, M.E., Hechinger, R.F., Chiba, S., 2006. Parasites alterhost phenotype and may create a new ecological niche for snail hosts. Proc. R.Soc. Lond. B Biol. Sci. 273, 1323–1328.
Mohebbi, F., 2010. The Brine Shrimp Artemia and hypersaline environmentsmicroalgal composition: a mutual interaction. Int. J. Aquat. Sci 1, 19–27.
Moore, J., 2002. Parasites and the Behaviour of Animals. Oxford University Press,Oxford, UK, pp. 315.
Muñoz, J., Gómez, A., Green, J., Figuerola, J., Amat, F., Rico, C., 2010. Evolutionaryorigin and phylogeography of the diploid obligate parthenogen Artemiaparthenogenetica (Branchiopoda: Anostraca). PLoS One 5, e11932.
O’Grady, S.P., Dearing, M.D., 2006. Isotopic insight into host–endosymbiontrelationships in Liolaemid lizards. Oecologia 150, 355–361.
Oren, A., 2005. Microscopic examination of microbial communities along a salinitygradient in saltern evaporation ponds: a ‘halophilic safari’. In: Gunde-Cimerman, N., Oren, A., Plemenitaš, A. (Eds.), Adaptation to Life at High SaltConcentrations in Archaea, Bacteria and Eukarya. Springer, Dordrecht, pp. 41–57.
Pakhomov, E.A., McClelland, J.W., Bernard, K., Kaehler, S., Montoya, J.P., 2004. Spatialand temporal shifts in stable isotope values of the bottom-dwelling shrimpNauticaris marionis at the sub-Antarctic archipelago. Mar. Biol. 144, 317–325.
Pedros-Alio, C., Calderon-Paz, J.I., MacLean, M.H., Medina, G., Marrase, C., Gasol, J.M.,Guixa-Boixereu, N., 2000. The microbial food web along salinity gradients. FEMSMicrobiol. Ecol. 32, 143–155.
Peterson, B.J., Howarth, R.W., Garritt, R.H., 1985. Multiple stable isotopes used totrace the flow of organic matter in estuarine food webs. Science 277, 1361–1363.
Peterson, B.J., Fry, B., 1987. Stable isotopes in ecosystem studies. Annu. Rev. Ecol.Syst. 18, 293–320.
Plaistow, S.J., Troussard, J.P., Cézilly, F., 2001. The effect of the acanthocephalanparasite Pomphorhynchus laevis on the lipid and glycogen content of itsintermediate host Gammarus pulex. Int. J. Parasitol. 31, 346–351. http://dx.doi.org/10.1016/S0020-7519(01)00115-1.
Podlesak, D., McWilliams, S.R., 2006. Metabolic routing of dietary nutrients in birds:effects of diet quality and macronutrient composition revealed using stableisotopes. Physiol. Biochem. Zool. 79, 534–549.
Post, D.M., 2002. Using stable isotopes to estimate trophic position: models,methods, and assumptions. Ecology 83, 703–718.
Power, M., Klein, G.M., 2004. Fish host–cestode parasite stable isotope enrichmentpatterns in marine, estuarine and freshwater fishes from northern Canada. Isot.Environ. Health Stud. 40, 257–266.
Poulin, R., 2007. Evolutionary Ecology of Parasites, 2nd ed. Princeton Univ. Press.,Princeton, New Jersey.
Provasoli, L., Shiraishi, K., 1959. Axenic cultivation of the brine shrimp Artemiasalina. Biol. Bull. 117, 347–355.
Redon, S., Amat, F., Hontoria, F., Vasileva, G.P., Nikolov, P.N., Georgiev, B.B., 2011.Participation of metanauplii and juvenile individuals of Artemiaparthenogenetica (Branchiopoda) in the circulation of avian cestodes. Parasitol.Res. 108, 905–912.
Sánchez, M.I., Georgiev, B.B., Nikolov, P.N., Vasileva, G.P., Green, A.J., 2006a. Red andtransparent brine shrimps (Artemia parthenogenetica): comparative study oftheir cestode infections. Parasitol. Res. 100, 111–114.
Sánchez, M.I., Green, A.J., Castellanos, E.M., 2006b. Spatial and temporal fluctuationsin presence and use of chironomid prey by shorebirds in the Odiel saltpans,south–west Spain. Hydrobiologia 567, 329–340.
Sánchez, M.I., Green, A.J., Castellanos, E.M., 2006c. Temporal and spatial variation ofan aquatic invertebrate community subjected to avian predation at the Odielsalt pans (SW Spain). Arch. Hydrobiol. 166, 199–223.
Sánchez, M.I., Green, A.J., Amat, F., Castellanos, E.M., 2007a. Internal transport ofbrine shrimps by migratory waders: dispersal probabilities depend on diet andseason. Mar. Biol. 151, 1407–1415.
Sánchez, M.I., Georgiev, B.B., Green, A.J., 2007b. Avian cestodes affect the behaviourof their intermediate host Artemia parthenogenetica: an experimental study.Behav. Proc. 74, 293–299.
Sánchez, M.I., Thomas, F., Perrot-Minnot, M.J., Bertrand-Michel, J., Missé, D., 2009a.Neurological and physiological disorders in Artemia harboring manipulativecestodes. J. Parasitol. 95, 20–24.
Sánchez, M.I., Hortas, F., Figuerola, J., Green, A.J., 2009b. Sandpipers select red brineshrimps rich in carotenoids but also parasites. Ethology 115, 196–200.
Savage, A., Knott, B., 1998. Artemia parthenogenetica in Lake Hayward, WesternAustralia. II. Feeding biology in a shallow, seasonally stratified, hypersaline lake.Int. J. Salt Lake Res. 7, 13–24.
Tavares, P.C., Kelly, A., Maia, R., Lopes, R.J., Serrao Santos, R., Pereira, M.E., Duarte,A.C., Furness, R.W., 2008. Variation in the mobilization of mercury into Black-winged Stilt Himantopus himantopus chicks in coastal saltpans, as revealed bystable isotopes. Estuar. Coast Shelf. Sci. 77, 65–76.
Tavares, P.C., McGill, R., Newton, J., Pereira, E., Duarte, A., Furness, R.W., 2009.Relationships between carbon sources, trophic level and mercury exposure ingeneralist shorebirds revealed by stable isotope ratios in chicks. Waterbirds 32,311–321.
Thiéry, A., Robert, F., Gabrion, C., 1990. Distribution des populations d’Artemia et deleur parasite Flamingolepis liguloides (Cestoda, Cyclophyllidea), dans les salinsdu littoral méditerranéen français. Can. J. Zool. 68, 2199–2204.
Thompson, S.N., 1983. Biochemical and physiological effects of metazoanendoparasites on their host species. Comp. Biochem. Physiol. Part B Comp.Biochem. 74 (2), 183–211.
Thompson, D.R., Furness, R.W., 1995. Stable isotope ratios of carbon and nitrogen infeathers indicate seasonal dietary shifts in Northern Fulmars. Auk 112, 493–498.
Thompson, R.M., Mouristen, K., Poulin, R., 2005. Importance of parasites and theirlife cycle characteristics in determining the structure of a large marine foodweb. J. Anim. Ecol. 74, 77–85.
Varo, N., Green, A.J., Sánchez, M.I., Ramo, C., Gómez, J., Amat, J.A., 2011. Behaviouraland population responses to changing availability of Artemia prey by moultingblack-necked grebes, Podiceps nigricollis. Hydrobiologia 664, 163–171.

80 M.I. Sánchez et al. / International Journal for Parasitology 43 (2013) 73–80
Vest, J.L., Conover, M.R., 2011. Food habits of wintering waterfowl on the Great SaltLake, Utah. Waterbirds 34, 40–50.
Wear, R.G., Haslett, S.J., Alexander, N.L., 1986. Effects of temperature and salinity onthe biology of Artemia franciscana Kellogg from lake Grassmere, New Zealand. 2.Maturation, fecundity, and generation times. J. Exp. Mar. Biol. Ecol. 98, 167–183.
Wurtsbaugh, W.A., 1992. Food- web modification by an invertebrate predator in theGreat Salt Lake (USA). Oecologia 89, 168–175.
Wurtsbaugh, W.A., Gliwicz, Z.M., 2001. Limnological control of brine shrimppopulation dynamics and cyst production in the Great Salt Lake, Utah.Hydrobiologia 466, 119–132.