cellulose synthesis and cell expansion are regulated … · cellulose synthesis and cell expansion...
TRANSCRIPT

Cellulose Synthesis and Cell Expansion Are Regulated byDifferent Mechanisms in Growing Arabidopsis HypocotylsOPEN
Alexander Ivakov,a,b,1,2,3 Anna Flis,b,1,2 Federico Apelt,b Maximillian Fünfgeld,b Ulrike Scherer,b,4 Mark Stitt,b
Friedrich Kragler,b Kris Vissenberg,c,d Staffan Persson,a,b,5 and Dmitry Suslovc,e,5
a School of Biosciences, University of Melbourne, Parkville, VIC 3010, Melbourne, AustraliabMax-Planck Institute of Molecular Plant Physiology, 14476 Potsdam, GermanycBiology Department, Integrated Molecular Plant Physiology Research, University of Antwerp, 2020 Antwerpen, BelgiumdUASC-TEI, Plant Biochemistry and Biotechnology Lab, Department of Agriculture, School of Agriculture, Food, and Nutrition,Stavromenos, 71 004 Heraklion, Crete, Greecee Saint Petersburg State University, Faculty of Biology, Department of Plant Physiology and Biochemistry, 199034 SaintPetersburg, Russia
ORCID IDs: 0000-0003-4091-1135 (A.I.); 0000-0003-3450-9369 (F.A.); 0000-0002-4900-1763 (M.S.); 0000-0001-5308-2976 (F.K.);0000-0003-0292-2095 (K.V.); 0000-0002-6377-5132 (S.P.); 0000-0002-0070-3182 (D.S.)
Plant growth is sustained by two complementary processes: biomass biosynthesis and cell expansion. The cell wall is crucialto both as it forms the majority of biomass, while its extensibility limits cell expansion. Cellulose is a major component of thecell wall and cellulose synthesis is pivotal to plant cell growth, and its regulation is poorly understood. Using periodic diurnalvariation in Arabidopsis thaliana hypocotyl growth, we found that cellulose synthesis and cell expansion can be uncoupledand are regulated by different mechanisms. We grew Arabidopsis plants in very short photoperiods and used a combinationof extended nights, continuous light, sucrose feeding experiments, and photosynthesis inhibition to tease apart the influencesof light, metabolic, and circadian clock signaling on rates of cellulose biosynthesis and cell wall biomechanics. Wedemonstrate that cell expansion is regulated by protein-mediated changes in cell wall extensibility driven by the circadianclock. By contrast, the biosynthesis of cellulose is controlled through intracellular trafficking of cellulose synthase enzymecomplexes regulated exclusively by metabolic signaling related to the carbon status of the plant and independently of thecircadian clock or light signaling.
INTRODUCTION
Plant growth is sustained by two complementary processes:biomass biosynthesis, resulting in dry matter accumulation, andcell expansion accompanied by water uptake leading to irre-versible increases in size. Both processes are tightly linked to thecellwall, a structure that encasesall plant cells. Amajor proportionof plant biomass is in the cell wall and large fluxes of photosyn-thetic products are directed to cell walls in growing cells (Wanget al., 2013; Boex-Fontvieille et al., 2014; Haigler et al., 2001). Onthe other hand, plant cell expansion is limited by cell wall ex-tensibility (Cosgrove, 1986, 2016a). The cell wall is thus a keyplayer in cell expansion, morphology, and size as well as plantbiomass accumulation.
Cell expansion is controlled by changes in cell wall properties,which have been extensively studied under different conditions(Cosgrove and Li, 1993; Schopfer et al., 2002; Suslov et al., 2009;Peaucelle et al., 2015). More specifically, cell expansion is linkedto cell wall extensibility, which can be defined as the ability ofthe cell wall to increase in surface area irreversibly during growth(Cosgrove, 2016a). Cell wall extensibility is regulated throughselective breakage and formation of links between wall polymersdriven by loosening and tightening proteins secreted by the cell(Cosgrove, 2016b).Bycontrast, howdifferentexternal and internalcues control cell wall synthesis is only starting to be unraveled(Bischoff et al., 2011; Boex-Fontvieille et al., 2014). To protectexpanding cell walls from excessive thinning and failure duringgrowth, their synthesis and expansion need to be coordinated(Voxeur and Höfte, 2016). While several reports have observedthat these processes can occur independently of each other(Edelmann et al., 1989; Edelmann and Fry, 1992; Refrégier et al.,2004; Derbyshireet al., 2007), theprinciplesbehindsuchuncouplingare not clear.Howcell wall synthesis is controlledandwhether it iscoordinated
with cell expansion can be investigated in plants growing in con-ditionsofalternating lightanddarknessduring24-hcycles,wherebylight, temperature,andmetabolic resourcesupplyfluctuate. In theseconditions, dicot plants exhibit diurnal variation in expansiongrowth. Studies on diurnal growth dynamics (Wiese et al., 2007;Yazdanbakhsh et al., 2011; Apelt et al., 2015) have shown that theperiodic patterns are generated through the interplay between light
1Current address: ARC Centre of Excellence in Translational Photosyn-thesis, Research School of Biology, Australian National University, GPOBox 475, Canberra, ACT 2601, Australia.2 These authors contributed equally to this work.3 Address correspondence to [email protected] Current address: Zoological Institute, University of Hamburg, Martin-Luther-King-Platz 3, 20146 Hamburg, Germany.5 Co-senior authors.The author responsible for distribution of materials integral to the findingspresented in this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Alexander Ivakov([email protected]).OPENArticles can be viewed without a subscription.www.plantcell.org/cgi/doi/10.1105/tpc.16.00782
The Plant Cell, Vol. 29: 1305–1315, June 2017, www.plantcell.org ã 2017 ASPB.

signaling and an internal timingmechanism known as the circadianclock (Nozue et al., 2007;Nusinowet al., 2011) and that the patternspersist in constant-light, constant-temperature conditions for ex-tended periods. However, the mechanistic basis for these growthvariations at the level of cell wall synthesis and cell expansion re-mains unclear, and it is unknown towhat extent the synthesis of cellwall varies with changing growth rates and what regulatory inputsimpact on it. In addition, how the cell wall extensibility modulatesthese rapid changes in growth and to what extent cell wall bio-synthesis and cell expansion are coordinated as growth rates varyremain largely unknown.
Anydiurnal patternobserved in aplantmaybe theconfluenceofseparate responses to light, metabolic, and endogenous cues.Untangling these effects requires an experimental system whereall three can be manipulated separately but all remain functional.Cell wall biosynthesis relies on carbon newly fixed through pho-tosynthesisduring thedayand remobilized fromstarch reservesatnight (Smith andStitt, 2007; Stitt andZeeman, 2012; Sulpice et al.,2014). When days are very short, the limited starch reserves ac-cumulated during the day are only sufficient to maintain res-piration at night (Gibon et al., 2009; Sulpice et al., 2014),leaving little for biosynthesis, while still allowing a circadianrhythm to be maintained in a natural diurnal cycle. These con-ditions offer an excellent experimental system to explore the regu-latory inputs thatcontrolgrowthratesat the levelofcellwall synthesisand remodeling.
Cellulose is a major component of plant cell walls. In contrastto other cell wall components, cellulose is produced by plasmamembrane cellulose synthase (CesA) enzyme complexes (CSCs).Fluorescently labeled CSCs can be observed microscopically asmoving foci in the plasmamembrane of plant cells (Paredez et al.,2006). Their movement is thought to be driven by the polymeri-zation and extrusion of cellulose chains into the cell wall, whichpushes theenzymecomplex in theplaneof theplasmamembrane,and this has been used as a proxy for the rate of cellulose syn-thesis. CSCs are also observed in intracellular membrane com-partments thought to be involved in their trafficking and the CSCsare thought to be inactive until they reach the plasma membrane(Paredez et al., 2006; Gutierrez et al., 2009; Crowell et al., 2009).Here,we investigated the regulationof cellulose synthesis andcellwall extensibility in hypocotyls of Arabidopsis thaliana plantsgrown in short diurnal cycles.
RESULTS
CesA Abundance at the Plasma Membrane Exhibits DiurnalFluctuations in Low-Carbon Conditions
To create a strong diurnal alternation between periods of growthand no growth, and to rely purely on photosynthesis-derivedcarbon forcellwall biosynthesis,wegrewArabidopsis seedlings inshort photoperiods (4 h day/20 h night) on sucrose-free media for7 d. It has been shown that seed storage lipids are essentiallydepleted as early as three days after germination (Andre andBenning, 2007; Kelly et al., 2011), and the plantsmust then relyon photosynthesis for their carbon supply, although it is notknown whether the reserves last longer or shorter under certain
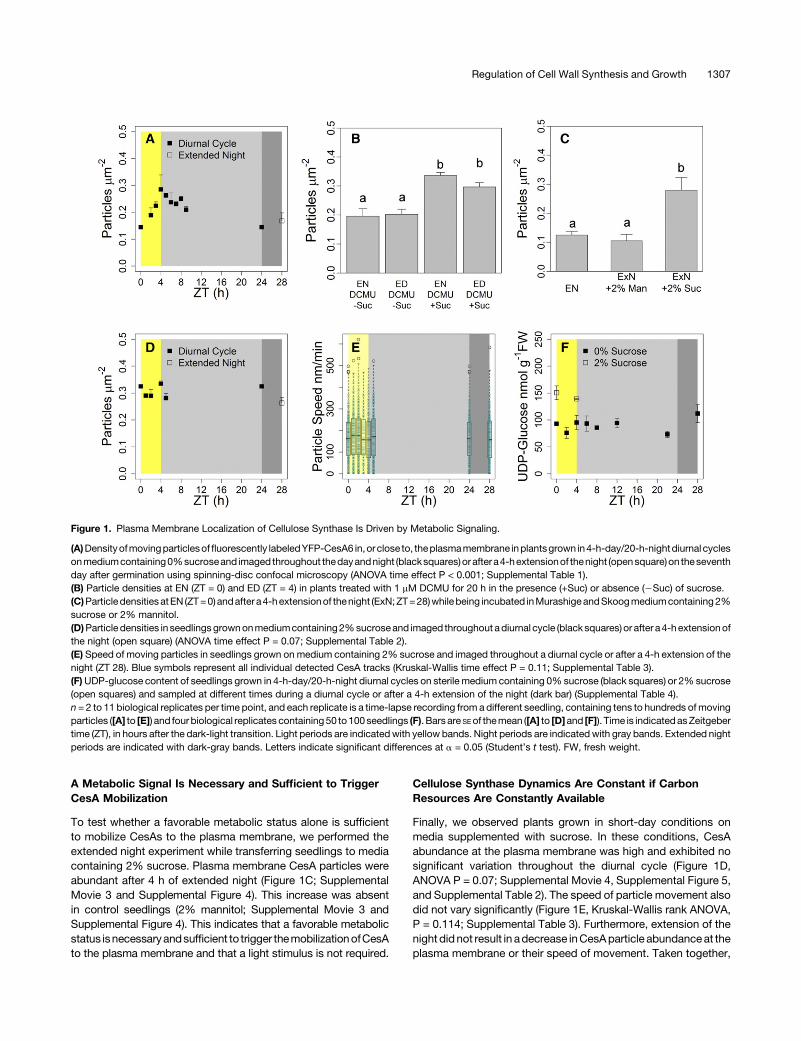
conditions. We observed a fluorescently tagged subunit of theCSC, YFP-CesA6, in hypocotyl cells using spinning disc confocalmicroscopyasan indicator of cellulose synthesis. At theendof thenight, there was a low abundance of CesA particles in the plasmamembrane, with the overwhelming majority associated with in-tracellular membrane compartments (Supplemental Movie 1 andSupplemental Figure 1), referred to as small CesA compartments(SmaCCs;Gutierrez et al., 2009), ormicrotubule-associatedCesAcompartments (MASCs;Crowell et al., 2009),whichare thought tocontain inactive CSCs and to be involved in either exo- or en-docytosis of the CSCs and exhibit erratic, irregular movement.CesA particles began to repopulate the plasmamembrane duringthe light period, while the number and intensity of SmaCCs de-creased. The density of plasmamembrane-based particles variedsignificantlywith time (ANOVAP<0.001) andsteadily increased toa peak at the end of the day (ED; Figure 1A; Supplemental Table 1)and declined thereafter, persisting for several hours into the night.ThisdiurnalpatternofCesAdynamicssuggests that lightsignalingand/or metabolic status and/or endogenous signals may regulatethe secretion of CesA to the plasmamembrane during the diurnalcycle.
Extended Darkness Prevents the Mobilization of CesA to thePlasma Membrane
To assess whether endogenous factors or signals regulated theobserved CesA behavior, we performed an extended night ex-periment where, on one day, plants were kept in the dark for 4 hinstead of in the light and were imaged at end of the subjectiveday. Here, plasma membrane CesA particle abundance did notincrease significantly during the extended night (Figure 1A;SupplementalMovie 1 andSupplemental Figure 1). This suggeststhat the diurnal fluctuations in CesA abundance at the plasmamembrane were not the result of endogenous rhythms and in-dicated that light signaling (i.e., photoreceptors) or the metabolicand/or signaling effects associated with photosynthesis in thelight must be required to mobilize the CesAs to the plasmamembrane.
Inhibition of Photosynthesis in the Light PreventsCesA Mobilization
To separate the roles of light signaling and photosynthesis, wepretreated short-day-grown seedlings with the photosynthesisinhibitor DCMU overnight and then exposed them to light in theabsence or presence of exogenous sucrose to provide a carbonsource independent of photosynthesis. Treatment with DCMUreduced the quantum efficiency of photosystem II to very lowlevelsboth in thepresenceandabsenceof sucrose (SupplementalFigure2). In theabsenceofsucrose,CesAabundanceat theplasmamembrane did not increase significantly during the day after in-hibition of photosynthesis by DCMU (Figure 1B; SupplementalMovie 2 and Supplemental Figure 3). However, in the presence ofDCMUandsucrose,CesAcomplexesweremobilized totheplasmamembrane despite the severe inhibition of photosynthesis. Theseresults suggest thatmetabolic signaling associatedwith the supplyof carbon triggers the mobilization of CesA complexes to theplasma membrane in the light.
1306 The Plant Cell
A Metabolic Signal Is Necessary and Sufficient to TriggerCesA Mobilization
To test whether a favorable metabolic status alone is sufficientto mobilize CesAs to the plasma membrane, we performed theextended night experiment while transferring seedlings to mediacontaining 2% sucrose. Plasma membrane CesA particles wereabundant after 4 h of extended night (Figure 1C; SupplementalMovie 3 and Supplemental Figure 4). This increase was absentin control seedlings (2% mannitol; Supplemental Movie 3 andSupplemental Figure 4). This indicates that a favorable metabolicstatus isnecessaryandsufficient to trigger themobilizationofCesAto the plasma membrane and that a light stimulus is not required.
Cellulose Synthase Dynamics Are Constant if CarbonResources Are Constantly Available
Finally, we observed plants grown in short-day conditions onmedia supplemented with sucrose. In these conditions, CesAabundance at the plasma membrane was high and exhibited nosignificant variation throughout the diurnal cycle (Figure 1D,ANOVA P = 0.07; Supplemental Movie 4, Supplemental Figure 5,and Supplemental Table 2). The speed of particle movement alsodid not vary significantly (Figure 1E, Kruskal-Wallis rank ANOVA,P = 0.114; Supplemental Table 3). Furthermore, extension of thenight did not result in adecrease inCesAparticle abundanceat theplasma membrane or their speed of movement. Taken together,
Figure 1. Plasma Membrane Localization of Cellulose Synthase Is Driven by Metabolic Signaling.
(A)Densityofmovingparticlesoffluorescently labeledYFP-CesA6 in,orclose to, theplasmamembrane inplantsgrown in4-h-day/20-h-nightdiurnal cyclesonmediumcontaining0%sucroseand imaged throughout thedayandnight (blacksquares)orafter a4-hextensionof thenight (opensquare)on theseventhday after germination using spinning-disc confocal microscopy (ANOVA time effect P < 0.001; Supplemental Table 1).(B) Particle densities at EN (ZT = 0) and ED (ZT = 4) in plants treated with 1 mM DCMU for 20 h in the presence (+Suc) or absence (2Suc) of sucrose.(C)Particle densitiesatEN (ZT=0)andafter a4-hextensionof thenight (ExN;ZT=28)whilebeing incubated inMurashigeandSkoogmediumcontaining2%sucrose or 2% mannitol.(D)Particle densities in seedlingsgrownonmediumcontaining2%sucroseand imaged throughout adiurnal cycle (black squares) or after a4-hextensionofthe night (open square) (ANOVA time effect P = 0.07; Supplemental Table 2).(E) Speed of moving particles in seedlings grown on medium containing 2% sucrose and imaged throughout a diurnal cycle or after a 4-h extension of thenight (ZT 28). Blue symbols represent all individual detected CesA tracks (Kruskal-Wallis time effect P = 0.11; Supplemental Table 3).(F)UDP-glucose content of seedlings grown in 4-h-day/20-h-night diurnal cycles on sterile medium containing 0%sucrose (black squares) or 2%sucrose(open squares) and sampled at different times during a diurnal cycle or after a 4-h extension of the night (dark bar) (Supplemental Table 4).n = 2 to 11 biological replicates per time point, and each replicate is a time-lapse recording from a different seedling, containing tens to hundreds ofmovingparticles ([A] to [E]) and four biological replicates containing50 to100seedlings (F). Barsare SEof themean ([A] to [D]and [F]). Time is indicatedasZeitgebertime (ZT), in hours after the dark-light transition. Light periods are indicatedwith yellow bands. Night periods are indicated with gray bands. Extended nightperiods are indicated with dark-gray bands. Letters indicate significant differences at a = 0.05 (Student’s t test). FW, fresh weight.
Regulation of Cell Wall Synthesis and Growth 1307
the unchanging abundance and movement speed of CesA inthese conditions implies that cellulose synthesis rates are con-stant when carbon resources are constantly available.
Substrate Levels Do Not Underlie Diurnal Variation inCellulose Synthase Dynamics
The fluxes through many enzymes are limited by the supply ofsubstrate, and it may be assumed that severemetabolic limitationmay reduce the pool of substrate available to theCesA, limiting itsactivityor affecting its trafficking.To investigate this,wemeasuredthe levels of UDP-glucose, the substrate of CesA, throughoutdiurnal cycles in seedlings grown in 4-h days with and withoutsucrose supplementation and also in extended night conditions.Surprisingly, UDP-glucose levels did not vary significantlywithoutsucrose supplementation (Figure 1F) despite altered CesA locali-zation. Extension of the night by 4 h also did not significantly alterUDP-glucose levels compared with normal short-day conditions.The presence of sucrose in the growth medium significantly in-creased UDP-glucose levels (ANOVA, P < 0.001; SupplementalTable 4),with 48 to 78%higher levels at the endof theday andendof the night in sucrose-grown plants compared with no sucrose,respectively. However, sucrose-grown plants had similar levelsof UDP-glucose at the end of the day and end of the night(Supplemental Table 4). These results suggest that the CesAdynamics are not due to substrate limitation as UDP-glucose ispresent at similar levels during the day, night, and extended nightand implies that a signaling process might be responsible for theobserved changes in CesA trafficking.
Extension Growth Is Independent of Cellulose Synthesis
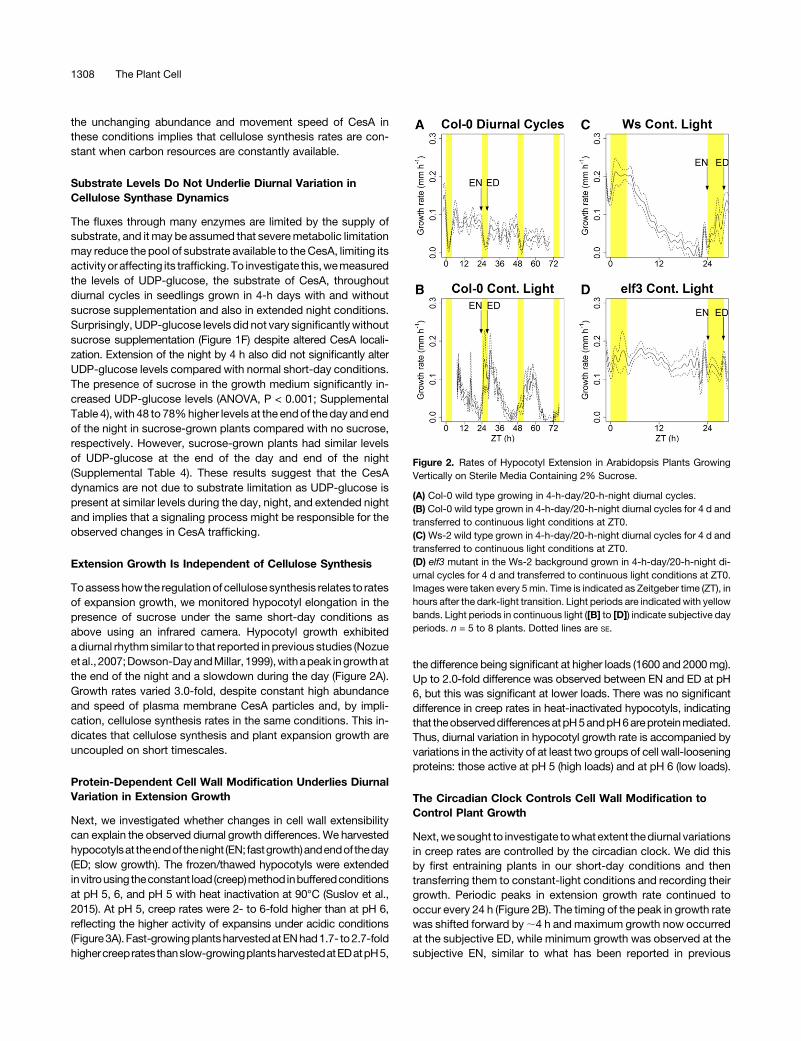
Toassesshowthe regulationofcellulosesynthesis relates to ratesof expansion growth, we monitored hypocotyl elongation in thepresence of sucrose under the same short-day conditions asabove using an infrared camera. Hypocotyl growth exhibitedadiurnal rhythmsimilar to that reported inprevious studies (Nozueetal., 2007;Dowson-DayandMillar, 1999),withapeak ingrowthatthe end of the night and a slowdown during the day (Figure 2A).Growth rates varied 3.0-fold, despite constant high abundanceand speed of plasma membrane CesA particles and, by impli-cation, cellulose synthesis rates in the same conditions. This in-dicates that cellulose synthesis and plant expansion growth areuncoupled on short timescales.
Protein-Dependent Cell Wall Modification Underlies DiurnalVariation in Extension Growth
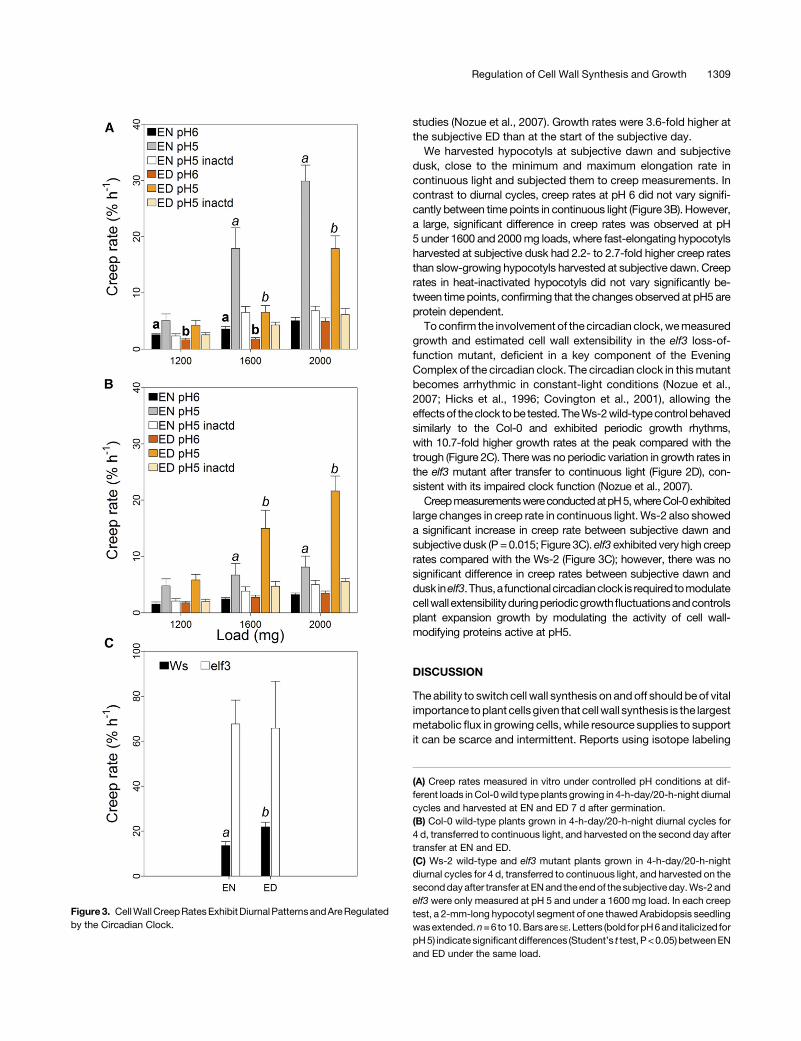
Next, we investigated whether changes in cell wall extensibilitycan explain the observed diurnal growth differences.We harvestedhypocotylsat theendofthenight (EN; fastgrowth)andendof theday(ED; slow growth). The frozen/thawed hypocotyls were extendedinvitrousing theconstant load (creep)method inbufferedconditionsat pH 5, 6, and pH 5 with heat inactivation at 90°C (Suslov et al.,2015). At pH 5, creep rates were 2- to 6-fold higher than at pH 6,reflecting the higher activity of expansins under acidic conditions(Figure3A). Fast-growingplantsharvestedatENhad1.7- to2.7-foldhighercreepratesthanslow-growingplantsharvestedatEDatpH5,
the difference being significant at higher loads (1600 and 2000mg).Up to 2.0-fold difference was observed between EN and ED at pH6, but this was significant at lower loads. There was no significantdifference in creep rates in heat-inactivated hypocotyls, indicatingthat theobserveddifferencesatpH5andpH6areproteinmediated.Thus, diurnal variation in hypocotyl growth rate is accompanied byvariations in the activity of at least two groups of cell wall-looseningproteins: those active at pH 5 (high loads) and at pH 6 (low loads).
The Circadian Clock Controls Cell Wall Modification toControl Plant Growth
Next,wesought to investigate towhatextent thediurnal variationsin creep rates are controlled by the circadian clock. We did thisby first entraining plants in our short-day conditions and thentransferring them to constant-light conditions and recording theirgrowth. Periodic peaks in extension growth rate continued tooccur every 24 h (Figure 2B). The timing of the peak in growth ratewas shifted forward by;4 h and maximum growth now occurredat the subjective ED, while minimum growth was observed at thesubjective EN, similar to what has been reported in previous
Figure 2. Rates of Hypocotyl Extension in Arabidopsis Plants GrowingVertically on Sterile Media Containing 2% Sucrose.
(A) Col-0 wild type growing in 4-h-day/20-h-night diurnal cycles.(B) Col-0 wild type grown in 4-h-day/20-h-night diurnal cycles for 4 d andtransferred to continuous light conditions at ZT0.(C)Ws-2 wild type grown in 4-h-day/20-h-night diurnal cycles for 4 d andtransferred to continuous light conditions at ZT0.(D) elf3 mutant in the Ws-2 background grown in 4-h-day/20-h-night di-urnal cycles for 4 d and transferred to continuous light conditions at ZT0.Images were taken every 5min. Time is indicated as Zeitgeber time (ZT), inhours after the dark-light transition. Light periods are indicated with yellowbands. Light periods in continuous light ([B] to [D]) indicate subjective dayperiods. n = 5 to 8 plants. Dotted lines are SE.
1308 The Plant Cell
studies (Nozue et al., 2007). Growth rates were 3.6-fold higher atthe subjective ED than at the start of the subjective day.We harvested hypocotyls at subjective dawn and subjective
dusk, close to the minimum and maximum elongation rate incontinuous light and subjected them to creep measurements. Incontrast to diurnal cycles, creep rates at pH 6 did not vary signifi-cantly between time points in continuous light (Figure 3B). However,a large, significant difference in creep rates was observed at pH5 under 1600 and 2000mg loads, where fast-elongating hypocotylsharvested at subjective dusk had 2.2- to 2.7-fold higher creep ratesthan slow-growing hypocotyls harvested at subjective dawn. Creeprates in heat-inactivated hypocotyls did not vary significantly be-tween time points, confirming that the changes observed at pH5 areprotein dependent.To confirm the involvement of the circadian clock,wemeasured
growth and estimated cell wall extensibility in the elf3 loss-of-function mutant, deficient in a key component of the EveningComplex of the circadian clock. The circadian clock in thismutantbecomes arrhythmic in constant-light conditions (Nozue et al.,2007; Hicks et al., 1996; Covington et al., 2001), allowing theeffectsof theclock tobe tested. TheWs-2wild-typecontrol behavedsimilarly to the Col-0 and exhibited periodic growth rhythms,with 10.7-fold higher growth rates at the peak compared with thetrough (Figure 2C). There was no periodic variation in growth rates inthe elf3 mutant after transfer to continuous light (Figure 2D), con-sistent with its impaired clock function (Nozue et al., 2007).CreepmeasurementswereconductedatpH5,whereCol-0exhibited
large changes in creep rate in continuous light.Ws-2 also showeda significant increase in creep rate between subjective dawn andsubjective dusk (P =0.015; Figure 3C). elf3 exhibited very high creeprates compared with the Ws-2 (Figure 3C); however, there was nosignificant difference in creep rates between subjective dawn anddusk inelf3.Thus,a functionalcircadianclock is required tomodulatecellwall extensibilityduringperiodicgrowthfluctuationsandcontrolsplant expansion growth by modulating the activity of cell wall-modifying proteins active at pH5.
DISCUSSION
The ability to switch cell wall synthesis on andoff should beof vitalimportance toplant cellsgiven that cellwall synthesis is the largestmetabolic flux in growing cells, while resource supplies to supportit can be scarce and intermittent. Reports using isotope labeling
Figure3. CellWallCreepRatesExhibitDiurnalPatternsandAreRegulatedby the Circadian Clock.
(A) Creep rates measured in vitro under controlled pH conditions at dif-ferent loads inCol-0wild type plants growing in 4-h-day/20-h-night diurnalcycles and harvested at EN and ED 7 d after germination.(B) Col-0 wild-type plants grown in 4-h-day/20-h-night diurnal cycles for4 d, transferred to continuous light, and harvested on the second day aftertransfer at EN and ED.(C) Ws-2 wild-type and elf3 mutant plants grown in 4-h-day/20-h-nightdiurnal cycles for 4 d, transferred to continuous light, and harvested on thesecondday after transfer at ENand the endof the subjective day.Ws-2 andelf3 were only measured at pH 5 and under a 1600 mg load. In each creeptest, a 2-mm-long hypocotyl segment of one thawed Arabidopsis seedlingwasextended.n=6 to10.Barsare SE.Letters (bold forpH6and italicized forpH5) indicate significantdifferences (Student’s t test, P<0.05) betweenENand ED under the same load.
Regulation of Cell Wall Synthesis and Growth 1309

techniques show that the incorporation of glucose units into cellwall polymers decreases during the night inwild-typeArabidopsisplants and is abolished from ;4 h into the night in the starchlesspgm mutant, showing a close temporal relationship to the de-pletion of sucrose in the mutant (Boex-Fontvieille et al., 2014;Ishihara et al., 2015). These labeling experiments are in closeagreement with our results on diurnal changes in cellulose syn-thesis estimated from CSC dynamics. Based on our data and thereport from Ishihara et al. (2015), it is tempting tospeculate that theproductionandsecretionof othercellwall components, likepectinand hemicelluloses, may be regulated in a similar manner. Thiswould perhaps not be surprising as they are also abundantcomponents of cell walls, their synthesis is a significant resource-consuming flux in growing cells, and their deposition likewisedepends on vesicle trafficking and exocytosis (McFarlane et al.,2014; Kim and Brandizzi, 2016).
It has been shown previously that cellulose synthesis is con-trolled by light through the action of the photoreceptor Phyto-chrome B (Bischoff et al., 2011), in contrast to our findingsindicating that light does not play a part in diurnal CesA dynamics.However, that study focuses on and highlights the distinctionbetween etiolated plants growing in complete and constantdarkness and light-grown seedlings growing in a long photope-riod. Furthermore, there were no differences in CesA behaviorbetween light- and dark-grown seedlings in wild-type plants, thedifferences only becoming apparent when a subunit of the CesAcomplex was mutated, resulting in slower CesA speeds in thedark. Taken together, our findings and those of Bischoff et al.(2011) suggest that there may be functional differences in theCesApopulations betweenetiolated seedlings and thosegrowingin the light. However, once growing in the light, the CesA pop-ulation exhibits diurnal dynamics that do not depend on lightdirectly, but respond to the metabolic effects of light on photo-synthesis and carbon assimilation.
The lack of correlation between UDP-glucose levels and ap-parent cellulose synthesis rates is intriguing. However, it has beenproposed, although never proven, that UDP-glucose may un-dergo metabolic channeling to the active site of CesA by sucrosesynthase associated with the CesA rosette as sucrose is brokendown (Haigler et al., 2001). Therefore, the relevant UDP-glucoseconcentration, i.e., that at the active site, may not be apparent inUDP-glucose measurements conducted on whole plant extracts,aswasdonehere.Anotherpossibleexplanation is thatametabolicsignaling process, rather than substrate levels directly, is re-sponsible for the observed CesA dynamics. While the identity ofthe metabolic sensor or signaling pathway responsible for theobserved patterns of CesA trafficking is not known at this point,several candidate sugar signaling pathways have been reported,including sucrose/trehalose-6-phosphate signaling, and severalkinases such as Target of Rapamycin, Sucrose-Nonfermenting(SNF1), andhexokinases (Mooreetal., 2003;Cooksonet al., 2005;Ruan et al., 2010; Dobrenel et al., 2011; Eveland and Jackson,2012; Yadav et al., 2014). It is not known whether any of thesepathways are involved in the regulation of vesicle trafficking;however, they are good candidates to explore. Mechanismslinking cellular metabolic status to the regulation of vesicularsecretion through Golgi-localized lipid binding SEC14 domainproteins have been proposed in animal cells (Bankaitis et al.,
2010). Arabidopsis containsa family of theseproteins and theyareknown to regulate vesicle trafficking (Huang et al., 2016). In-vestigation of the involvement of these pathways in the regulationof CesA trafficking and vesicle trafficking in general may bea promising lead for future research.Superimposed on the accumulation of cell wall material is cell
expansion, resulting in increases in cell and plant size. Regulationof expansion growth rate is known to be one of the chief outputsof the plant circadian clock (Nozue et al., 2007; Dowson-Day andMillar,1999).This regulationappears towork throughtheeffectof theclock on the expression of growth-promoting PHYTOCHROMEINTERACTING FACTOR (PIF) transcription factors, which furthermodulateauxinsignaling (Nozueetal.,2011;Hornitscheketal.,2012;Seatonetal.,2015).However, thefinalstep in this regulatorycascadeis not clear. The exclusive effect of the clock on the activity of cellwall loosening proteins active at pH5 and high loads strongly sug-gests that expansins are the ultimate targets in the regulation ofexpansion growth by the clock (Cosgrove, 2016b). This is also in linewith published observations that several expansin genes haverhythmic, circadian expressionpatterns (Harmer et al., 2000;Bläsinget al., 2005). Some of these have been implicated as targets of PIFtranscription factors (Leivar et al., 2009; Hornitschek et al., 2012), aswell as being directly targeted by the clock transcription factorCIRCADIANCLOCKASSOCIATED1 (Nageletal., 2015). Interestingly,we observed that changes in cell wall extensibility are also partlymediated by proteins active at pH6 and under low loads in shortphotoperiods (Figure 3A). Their activity is not controlled by the clock(Figure3B), butmaybedependent on light signaling and/ormetabolicinputs. Xyloglucan endotransglucosylase/hydrolases (XTHs) may beresponsible for this activity because of their characteristic pH-optima(Maris et al., 2011) and their ability to inducewall extension under lowloads (Miedes et al., 2013).The EN versus ED differences in cell wall extensibility were
abolished by heat inactivation (Figures 3A and 3B), showing thatthey resulted from changes in the activity of cell wall looseningproteins active at pH 5.0 and 6.0 that are present in the wall.However, the diurnal regulation of cell expansion and the con-tribution of these proteins to diurnal growth rhythms (Figure 2)could depend not only on their relative abundance or activity, butalsoonpossible variations in turgor andpH.High turgor stimulatesexpansionby exerting agreater force on thewall, while lowpHhasbeen shown to stimulate expansion by increasing the activity ofexpansins. Higher turgor will favor the contribution of expansins,consistentwith theirmorepronouncedeffects oncreep rate underhigher loads (Miedes et al., 2013; Suslov et al., 2015). Diurnal andcircadian variations in turgor have been demonstrated in spe-cialized cells such as stomata and pulvini (Hennessey and Field,1992). Although it has never been shown to occur in Arabidopsishypocotyls, diurnal changes in turgor there could reflect rhythmicdiurnal changes in the accumulation/mobilization of osmoticallyactive assimilates (Haydonet al., 2011) andvariation inplantwaterstatus brought on by stomatal opening and closing, known toaffect turgor (Ache et al., 2010; Bramley et al., 2015).Changes in cell wall acidification may also play a role in the
regulation of cell expansion. Cell wall acidification mediated byplasma membrane H+-ATPases (Falhof et al., 2016) stimulatesexpansins due to their low-pH optima (Cosgrove, 2016b) and isalso the driving force for solute uptake to generate cell turgor.
1310 The Plant Cell

H+-ATPases aremajorconsumersofATP,and their activitymaybeincreased by light, which elevates the ATP level (Liang et al., 2016).On the other hand, light strongly up- or downregulates H+-ATPaseactivity via phosphorylation (Kinoshita andShimazaki, 1999; Hohmet al., 2014). To our knowledge, no diurnal or circadian rhythms incell wall pH have ever been demonstrated. However, there is clearevidence for circadian rhythms in auxin content in Arabidopsisshoots (Jouve et al., 1999). Auxin is known to promote cell wallacidificationbyactivatingH+-ATPases (Spartz et al., 2014), and it istherefore likely that cell wall pH variation also contributes to diurnalgrowth rhythms. These data show that the contribution of proteinsactive at pH 5.0 and 6.0 could vary depending on the phase of thediurnal growth cycle and external conditions. The role of expansinsmay be prevalent in conditions optimal for growth, i.e., in highlyturgid cells having favorable energetic status. By contrast, thecontribution of proteins active at pH 6.0 may increase in morestressful conditions.
While we use the Arabidopsis hypocotyl as a simple model ofone-directional cellular and organ growth, it is possible that thissystem may be unique in certain aspects and may not reflect theregulationof growth inotherorgans. For example, an investigationof rhythmic patterns of leaf growth and leaf movements in Ara-bidopsis uncovered that light was an essential factor promotingleaf expansion during the day (Dornbusch et al., 2014); however,light suppresses hypocotyl extension (Nozue et al., 2007). Hy-pocotyls and leaves have somewhat opposite growth responses,in that darkness, low light, or short photoperiods stimulate hy-pocotyl extension while inhibiting the growth and development ofleaves in what is known as the etiolation response (Fankhauserand Chory, 1997). This serves to direct the plant’s resources torapid expansion growth to “grow out” of zones of darkness orshade, e.g., when seeds germinate underground. When light issensed by photoreceptors, hypocotyl growth is rapidly suppressedand leafgrowth isstimulated toproducephotosynthetic leafareaandbegins to assimilate carbon. Thus, the regulation of leaf expansiongrowth is rather different to that of the hypocotyl. On the other hand,biosynthesis is inevitably under metabolic constraints in any cell ororgan and must be regulated in accordance with the supply ofavailable resources (Smith and Stitt, 2007). This regulation maydiffer slightly in green organs that fix their own carbon (e.g., leaves)comparedwith non-green carbon-importing cells in hypocotyls androots; however, the constraints remain the same and unregulatedbiosynthesis in times of scarce supply can exhaust the supply ofcarbon and energy and result in death. Therefore, unlike organ-specific requirements for expansion growth in response to envi-ronmental cues, it is unlikely that hypocotyls are uniquewith respectto the regulation of cellular biosynthesis.
This work demonstrates that cellulose biosynthesis and cellexpansion are regulated by different mechanisms, with the en-suing result that they can be uncoupled for short time periods. Ofcourse, there cannot be complete uncoupling between the twoprocesses, as wall expansion cannot proceed limitlessly withoutthe synthesis and addition of new wall material to maintain theintegrity of the cell wall. While we show that plant carbon statusdefines the amount of cellulose the cell produces, Stewart et al.(2011) showed that sucrose also affects hypocotyl expansiongrowth in Arabidopsis. In that study, growth became very slow,while stillmaintainingdiurnal rhythms, in short days in theabsence
of external carbon similar to our conditions, while the addition ofsucrose strongly stimulatedexpansiongrowth. This suggests thaton longer timescales, the rate of cell expansion adjusts to theamount of cell wall material that is able to be produced. This isreminiscentof thecellwall integrity sensingmechanisms thathavebeen shown to inhibit expansion growth in situations where cel-lulose synthesis is inhibited using genetic or chemical means andunder conditions of stress, known to involve several receptor-like-kinases and wall-associated-kinases (Hématy et al., 2007; Wolfet al., 2012). These kinases are worthy of investigation as thepossible missing piece of the puzzle in the regulation of growth inresponse to resource supply.In summary,we show that cell wall synthesis andcell expansion
can be uncoupled in Arabidopsis hypocotyls on short timescales.We conclude that this uncoupling is based on different regulatorymechanisms of cell wall biosynthesis and cell expansion. Thebiosynthesis of cellulose, and possibly other cell wall polymers, isregulated through a relatively simple one-way control mechanisminvolving metabolic signals associated with the carbon status ofthe plant. It is independent of the circadian clock or light signaling.On the other hand, cell expansion is controlled by the plant cir-cadian clock mediating protein-dependent changes in cell wallextensibility. At the moment, these observations are only valid forthe hypocotyl, which is a highly specialized organ, and for a ratherextremephotoperiod. Therefore, amuch closer coupling betweenwall synthesis and cell expansion may exist in other cell types orother conditions. Our finding that the two basic processes un-derlying plant growth are regulated by different mechanismsenables their independent manipulation. This could open upavenues to modulate plant growth and biomass production.
METHODS
Plant Material
YFP-CesA6 expressed under the native CesA6 promoter in the prc1-1background was provided by D. Ehrhardt (Paredez et al., 2006). elf3-4mutant in Ws-2 background was provided by Andrew Millar (Hicks et al.,1996).
Plant Growth
Seedswere sterilizedwith 70%ethanolwith adropof TritonXdetergent for30 min and washed four times with sterile water. Seeds were spread onMurashigeandSkoogagarplateswith0or2%sucroseandstratifiedat4°Cfor 3 d. Plates were transferred to controlled environment chambers(Percival Scientific) running on a 4-h-light/20-h-dark 20°C/18°C photo-period and an irradiance of 100 mmol m22 s21 provided by fluorescentlamps during the day, and grown vertically. Confocal imagingwas done onthe seventh day after stratification. For continuous light treatments, plantswere grown in 4-h photoperiods for 4 d and transferred to continuous lowlight at 20°C for 2 d. Lamps were darkened with plastic shades duringcontinuous light to give an irradiance of 12 mmol m22 s21. For creepmeasurements, plants were harvested and frozen in liquid nitrogen on theseventh day in diurnal cycles and on the second day of continuous light.
Microscopy
For live-cell imaging, seedlings expressing YFP:CesA6 were mounted onglass slides under an agarose gel pad and imaged on aNikon Ti-E inverted
Regulation of Cell Wall Synthesis and Growth 1311

confocal microscope with a CSU-X1 Yokogawa spinning disc head, usinga CFI APO TIRF 3100 N.A. 1.49 oil immersion objective, an evolve CCDcamera (Photometrics Technology), and a 1.23 lens between the spinningdisc and camera. YFPwas excited at 491 nmusing amultichannel dichroicand an ET525/50M band-pass emission filter (Chroma Technology). Im-ages were acquired using Metamorph online premier, version 7.5. Expo-sure times were 400 ms.
Growth Imaging
To image plant growth in real time, a modified Nikon DSLR camera wasused, where the infrared cut-off filter in front of the CCD sensor was re-moved. A long-pass near-infrared filter was fitted to the objective so thatonly infrared radiationentered thecamera, allowing theplants tobe imagedday and night. Plants were continuously illuminated from behind with in-frared light at 900nm, shownnot to stimulateplant photoreceptor systems.Images were taken every 5 min.
Image Analysis
Confocal space-time stacks of fluorescently labeled CesA were analyzedusingcustom-written scripts running in Fiji andR. ForCesAparticle densitymeasurements, one biological replicate was a confocal time-lapse movieof 30 frames and lasting 5 min, with each movie being collected on a dif-ferentplant.Eachmoviewasanalyzedusingourparticle trackingalgorithm,detecting tens tohundredsofCesAparticles ineachmovie.Particledensitywas calculated by dividing the number of detected particles by the areaimaged in the movie in mm2. Multiple movies were collected for each timepoint or treatment, and the means and SE are reported in the figures.
CesA Particle Tracking
The first 20 frames of each stack were used for analysis. The positions ofCesA particles in each frame were detected by applying a Laplacian filterwith a smoothing kernel width of 1.5 pixels. The particles were thendetected by finding local maxima with a noise tolerance of 800.
Tracks were detected in R. The detected particles in all frames wereencoded as three-dimensional x-y-t points. Particles in the top frame (firsttime point) of each movie were used as starting points. Proceeding iter-atively, unit vectors from each starting point to all other points in all frameswere computed. Vectors with a t-direction component of at least 0.9 werethen retained. This removes the majority of vectors which point to CesAparticles in other parts of the image (and thus probably not the same track).
The remaining points are then expected to contain points corre-sponding to the track of interest (the sameCesA particle moving over time)and a number of erroneous points corresponding to other CesAs movinginto the search region, or noise points. Due to the very regularmovement ofplasmamembraneCesAparticles (constant speed, constantdirection), thepoints in a track should have a very consistent direction through the x-y-tspace, looking from the starting point, while the erroneous points woulddeviate from this direction as outliers. A median direction vector wascalculated from the retained points using the L1multivariate median (Vardiand Zhang, 2000), a measure with low sensitivity to outliers. All the pointswere then compared with the median direction using cosine similarity,a measure that expresses the cosine of the angle between two directionvectors. Pointswith acosinesimilarityof at least 0.8 to themediandirectionwere retained. The L1 median was then recalculated on the remainingpoints and points with cosine similarity of at least 0.95 were retained. Thisway,onlypointswithhighlysimilardirectionsareselected.Thesepointsarethen treated as a track. Tracks with fewer than five points were discarded.
Each track was then centered by subtracting its mean and a line of bestfit in the x-y plane was fitted by extracting the first principal component,a line which minimizes the perpendicular distances between all points and
itself. The percentage of explained variance (similar to r2 in least squaresregression) was used as ameasure of linearity for the track and tracks withlinearities below 90% were discarded.
The tracks that satisfy the above criteria were then treated as a singleCesAparticlemoving over time. For these tracks, eachpointwasprojectedonto the line of best fit to infer the “true” position of the particle, treatingdeviations from the line as error. The position of the particle along the linewas then regressed against time and the slope taken as the speed ofmovement along the track. The number of trackswasdividedby the area ofthe region of interest to obtain particle density (CesA/area).
It must be noted that at a given time a proportion of SmaCCs move ina similar manner and with similar speeds to plasma membrane CesAcomplexes and cannot be distinguished from them with certainty in allinstances. Thus, speed and density measurements are only accuratein situations where few SmaCCs are present and the vast majority of allparticles are plasma membrane CesA complexes, or vice versa. In sit-uations where SmaCCs are in high abundance, plasma membrane CesAnumbers are overestimated, while speed distributions are biased towardthe movement properties of SmaCCs.
Analysis of hypocotyl length was done in ImageJ with subsequent dataanalysis inR.The lengthof theplantwasdetermined ineach frame.The rateof extension growth was obtained by fitting cubic splines to the growthcurves and evaluating their first derivative at each time point.
Biomechanics Measurements
Cell wall extensibility was estimated by the creep method in which frozen/thawed hypocotyls were extended under a constant load. On the seventhday after germination, each Arabidopsis thaliana seedling was transferredto a 2-mL Eppendorf test tube, which was closed and plunged into liquidnitrogen. The frozen seedlingswere stored at –20°C and extended in creeptests within 2 weeks after freezing. Freeze/thawing disrupts the cells whileleaving the cell wall intact, making the measurements independent ofturgor variations, while the constant load resembles the action of turgor onthe cell wall and allows the force applied to the wall to be controlled. Cellwall heat inactivation was performed at 90°C for 3 min as described bySuslov et al. (2015). The optimization of the heat inactivation procedure isdetailed inSupplemental Figure6.Foreachcellwall creep (time-dependentdeformation) measurement, one thawed Arabidopsis seedling was se-cured inacustom-built constant-loadextensiometer (SuslovandVerbelen,2006) such that a2-mm-longhypocotyl segment located1.5mmbelow thecotyledonswasplacedbetween theclampsof thesetup.Thissegmentwasfirst preincubated in a buffer (20 mM MES-KOH, pH 6.0, or 20 mMNa-acetate, pH 5.0) in the relaxed state for 2 min, after which a constantload (1200, 1600, or 2000 mg) was applied, and cell wall extension wasrecorded for 15 min in the same buffer as during the preincubation.Conducting themeasurements in buffered conditions eliminates the role ofwall pH variations that may occur in vivo. The pH of buffers used corre-sponds to typical values in the plant apoplastic space (Felle, 2001). Theearly (0–5min after loading) cell wall deformation is known to contain largeviscoelastic components having no direct relation to cell wall extensibility(Suslov and Verbelen, 2006). This deformation was taken into account tocalculate the length of extending cell wall samples at 5 min after loading(Miedes et al., 2013). Theaveragecreep ratewas thencalculatedduring theinterval 5 to 15 min after loading using the formula (ln L15min 2 ln L5min)/T3
100%,where L15min and L5min indicate the length of an extending hypocotylsegment at 15 and5min after loading, respectively, andT is the timeduringwhich the average creep rate is calculated (10 min) (Miedes et al., 2013).
Metabolite Analysis
Plants were grown as described above and harvested at relevant timepoints by quenching in liquid nitrogen. Approximately 200 seedlings were
1312 The Plant Cell

pooled per biological replicate sample. Material was homogenized whilefrozen in a ball mill (Retsch) and aliquoted into Eppendorf tubes. Sampleswere extracted in a 3:7 chloroform/methanol solution. UDP-glucose wasmeasured using reverse-phase liquid chromatography-tandem massspectrometry as described by Arrivault et al. (2009).
Statistical Analysis
All statistical analysis was performed in R. Time courses of particle den-sities were tested for a significant effect of time using one-way parametricANOVA, testing the null hypothesis that particle densities do not vary overtime.TimecoursesofCesAspeedwere tested forasignificanteffectof timeusing the nonparametric Kruskal-Wallis rank test as the CesA speeddistributions were not normally distributed, testing the null hypothesis thatCesA speed does not vary over time. Pairwise comparisons of particledensities and creep rates were conducted using Student’s t test. Nullhypotheses were rejected below the 0.05 P value threshold.
Supplemental Data
Supplemental Figure 1. Inhibition of photosynthesis by DCMU.
Supplemental Figure 2. Heat inactivation of cell wall proteins underdifferent conditions.
Supplemental Figure 3. Cellulose synthase localization follows a di-urnal pattern in living Arabidopsis cells growing in very short dayswithout exogenous sucrose.
Supplemental Figure 4. Cellulose synthase localization to the plasmamembrane in the light requires active photosynthesis or an alternativecarbon source.
Supplemental Figure 5. A favorable metabolic status is necessaryand sufficient to promote localization of cellulose synthase to theplasma membrane.
Supplemental Figure 6. Arabidopsis seedlings synthesize celluloseconstantly under a favorable metabolic status.
Supplemental Figure 7. Speed of moving particles in seedlings grownon medium without exogenous sucrose and imaged throughouta diurnal cycle or after a 4-h extension of the night.
Supplemental Table 1. Variation in CesA particle densities overa diurnal cycle and an extended night in seedlings grown in theabsence of sucrose.
Supplemental Table 2. Variation in CesA particle densities overa diurnal cycle and an extended night in seedlings grown in thepresence of 2% sucrose.
Supplemental Table 3. Variation in CesA particle speed over a diurnalcycle and an extended night in seedlings grown in the presence of 2%sucrose.
Supplemental Table 4. Variation in UDP-glucose content over a di-urnal cycle and an extended night in seedlings grown in the presenceand absence of 2% sucrose.
Supplemental Movie 1. Cellulose synthase localization follows a di-urnal pattern in living Arabidopsis cells growing in very short dayswithout exogenous sucrose.
Supplemental Movie 2. Cellulose synthase localization to the plasmamembrane in the light requires active photosynthesis or an alternativecarbon source.
Supplemental Movie 3. A favorable metabolic status is necessary andsufficient to promote localization of cellulose synthase to the plasmamembrane.
Supplemental Movie 4. Arabidopsis seedlings synthesize celluloseconstantly under a favorable metabolic status.
ACKNOWLEDGMENTS
A.I. was funded by the Max-Planck Gesellschaft and the University ofMelbourne. A.F. was funded by the Max Planck Gesellschaft. S.P. wasfunded by the Max-Planck Gesellschaft and by a R@MAP professorshipat the University of Melbourne and acknowledges funding from anARC Discovery grant (DP150103495), an ARC Future fellowship grant(FT160100218), and a Hermon-Slade grant (Persson HSF 15/4). D.S.acknowledges Saint Petersburg State University for research grants1.38.233.2014and1.42.1025.2016, theResearchPark ofSaintPetersburgState University Center for Molecular and Cell Technologies, and RFBRfor Grant 15-04-04075. K.V. acknowledges the University of Antwerpand the Research Foundation Flanders (FWO-Vlaanderen) for GrantsG.0656.13.N.10, 1.5.091.11.N.00, and G039815N. We acknowledgeRoss Dennis for help with imaging.
AUTHOR CONTRIBUTIONS
A.I., D.S., and A.F. conceived the project and designed experiments. A.I.,D.S., A.F., F.A., M.F., U.S., and F.K. conducted experiments and inter-preted results. A.I., D.S., andM.F. analyzed data. M.S., F.K., K.V., and S.P.provided discussions on experiment design and data interpretation.
ReceivedOctober26,2016; revisedApril 18,2017;acceptedMay24,2017;published May 26, 2017.
REFERENCES
Ache, P., Bauer, H., Kollist, H., Al-Rasheid, K.A.S., Lautner, S.,Hartung, W., and Hedrich, R. (2010). Stomatal action directly feedsback on leaf turgor: new insights into the regulation of the plantwater status from non-invasive pressure probe measurements.Plant J. 62: 1072–1082.
Andre, C., and Benning, C. (2007). Arabidopsis seedlings deficientin a plastidic pyruvate kinase are unable to utilize seed storagecompounds for germination and establishment. Plant Physiol. 145:1670–1680.
Apelt, F., Breuer, D., Nikoloski, Z., Stitt, M., and Kragler, F. (2015).Phytotyping(4D): a light-field imaging system for non-invasive andaccurate monitoring of spatio-temporal plant growth. Plant J. 82:693–706.
Arrivault, S., Guenther, M., Ivakov, A., Feil, R., Vosloh, D., vanDongen, J.T., Sulpice, R., and Stitt, M. (2009). Use of reverse-phase liquid chromatography, linked to tandem mass spectrometry,to profile the Calvin cycle and other metabolic intermediates inArabidopsis rosettes at different carbon dioxide concentrations.Plant J. 59: 826–839.
Bankaitis, V.A., Mousley, C.J., and Schaaf, G. (2010). The Sec14superfamily and mechanisms for crosstalk between lipid metabo-lism and lipid signaling. Trends Biochem. Sci. 35: 150–160.
Bischoff, V., Desprez, T., Mouille, G., Vernhettes, S., Gonneau, M.,and Höfte, H. (2011). Phytochrome regulation of cellulose synthesisin Arabidopsis. Curr. Biol. 21: 1822–1827.
Bläsing, O.E., Gibon, Y., Günther, M., Höhne, M., Morcuende, R.,Osuna, D., Thimm, O., Usadel, B., Scheible, W.R., and Stitt, M.(2005). Sugars and circadian regulation make major contributions to
Regulation of Cell Wall Synthesis and Growth 1313

the global regulation of diurnal gene expression in Arabidopsis.Plant Cell 17: 3257–3281.
Boex-Fontvieille, E., Davanture, M., Jossier, M., Zivy, M., Hodges,M., and Tcherkez, G. (2014). Photosynthetic activity influencescellulose biosynthesis and phosphorylation of proteins involvedtherein in Arabidopsis leaves. J. Exp. Bot. 65: 4997–5010.
Bramley, H., Bitter, R., Zimmermann, G., and Zimmermann, U.(2015). Simultaneous recording of diurnal changes in leaf turgor andstem water status of bread wheat reveal variation in hydraulic mecha-nisms in response to drought. Funct. Plant Biol. 42: 1001–1009.
Cookson, S.J., Van Lijsebettens, M., and Granier, C. (2005). Cor-relation between leaf growth variables suggest intrinsic and earlycontrols of leaf size in Arabidopsis thaliana. Plant Cell Environ. 28:1355–1366.
Cosgrove, D. (1986). Biophysical control of plant cell growth. Annu.Rev. Plant Physiol. 37: 377–405.
Cosgrove, D.J. (2016a). Plant cell wall extensibility: connecting plantcell growth with cell wall structure, mechanics, and the action ofwall-modifying enzymes. J. Exp. Bot. 67: 463–476.
Cosgrove, D.J. (2016b). Catalysts of plant cell wall loosening. F1000Res.5: 119.
Cosgrove, D.J., and Li, Z.-C. (1993). Role of expansin in cell en-largement of oat coleoptiles. Analysis of developmental gradientsand photocontrol. Plant Physiol. 103: 1321–1328.
Covington, M.F., Panda, S., Liu, X.L., Strayer, C.A., Wagner, D.R.,and Kay, S.A. (2001). ELF3 modulates resetting of the circadianclock in Arabidopsis. Plant Cell 13: 1305–1315.
Crowell, E.F., Bischoff, V., Desprez, T., Rolland, A., Stierhof, Y.D.,Schumacher, K., Gonneau, M., Höfte, H., and Vernhettes, S.(2009). Pausing of Golgi bodies on microtubules regulates secretionof cellulose synthase complexes in Arabidopsis. Plant Cell 21:1141–1154.
Derbyshire, P., Findlay, K., McCann, M.C., and Roberts, K. (2007).Cell elongation in Arabidopsis hypocotyls involves dynamic changesin cell wall thickness. J. Exp. Bot. 58: 2079–2089.
Dobrenel, T., Marchive, C., Sormani, R., Moreau, M., Mozzo, M.,Montané, M.H., Menand, B., Robaglia, C., and Meyer, C. (2011).Regulation of plant growth and metabolism by the TOR kinase.Biochem. Soc. Trans. 39: 477–481.
Dornbusch, T., Michaud, O., Xenarios, I., and Fankhauser, C.(2014). Differentially phased leaf growth and movements in Arabi-dopsis depend on coordinated circadian and light regulation. PlantCell 26: 3911–3921.
Dowson-Day, M.J., and Millar, A.J. (1999). Circadian dysfunctioncauses aberrant hypocotyl elongation patterns in Arabidopsis. PlantJ. 17: 63–71.
Edelmann, H., Bergfeld, R., and Schopfer, P. (1989). Role of cell-wall biogenesis in the initiation of auxin-mediated growth in co-leoptiles of Zea mays L. Planta 179: 486–494.
Edelmann, H.G., and Fry, S.C. (1992). Effect of cellulose synthesisinhibition on growth and the integration of xyloglucan into pea in-ternode cell walls. Plant Physiol. 100: 993–997.
Eveland, A.L., and Jackson, D.P. (2012). Sugars, signalling, andplant development. J. Exp. Bot. 63: 3367–3377.
Falhof, J., Pedersen, J.T., Fuglsang, A.T., and Palmgren, M. (2016).Plasma membrane H(+)-ATPase regulation in the center of plantphysiology. Mol. Plant 9: 323–337.
Fankhauser, C., and Chory, J. (1997). Light control of plant de-velopment. Annu. Rev. Cell Dev. Biol. 13: 203–229.
Felle, H.H. (2001). pH: signal and messenger in plant cells. Plant Biol.3: 577–591.
Gibon, Y., Pyl, E.T., Sulpice, R., Lunn, J.E., Höhne, M., Günther, M.,and Stitt, M. (2009). Adjustment of growth, starch turnover, protein
content and central metabolism to a decrease of the carbon supplywhen Arabidopsis is grown in very short photoperiods. Plant CellEnviron. 32: 859–874.
Gutierrez, R., Lindeboom, J.J., Paredez, A.R., Emons, A.M.C., andEhrhardt, D.W. (2009). Arabidopsis cortical microtubules positioncellulose synthase delivery to the plasma membrane and interactwith cellulose synthase trafficking compartments. Nat. Cell Biol. 11:797–806.
Haigler, C.H., Ivanova-Datcheva, M., Hogan, P.S., Salnikov, V.V.,Hwang, S., Martin, K., and Delmer, D.P. (2001). Carbon parti-tioning to cellulose synthesis. Plant Mol. Biol. 47: 29–51.
Harmer, S.L., Hogenesch, J.B., Straume, M., Chang, H.S., Han, B.,Zhu, T., Wang, X., Kreps, J.A., and Kay, S.A. (2000). Orchestratedtranscription of key pathways in Arabidopsis by the circadian clock.Science 290: 2110–2113.
Haydon, M.J., Bell, L.J., and Webb, A.A. (2011). Interactions be-tween plant circadian clocks and solute transport. J. Exp. Bot. 62:2333–2348.
Hématy, K., Sado, P.E., Van Tuinen, A., Rochange, S., Desnos, T.,Balzergue, S., Pelletier, S., Renou, J.P., and Höfte, H. (2007). Areceptor-like kinase mediates the response of Arabidopsis cells tothe inhibition of cellulose synthesis. Curr. Biol. 17: 922–931.
Hennessey, T.L., and Field, C.B. (1992). Evidence of multiple circa-dian oscillators in bean plants. J. Biol. Rhythms 7: 105–113.
Hicks, K.A., Millar, A.J., Carré, I.A., Somers, D.E., Straume, M.,Meeks-Wagner, D.R., and Kay, S.A. (1996). Conditional circadiandysfunction of the Arabidopsis early-flowering 3 mutant. Science274: 790–792.
Hohm, T., Demarsy, E., Quan, C., Allenbach Petrolati, L., Preuten,T., Vernoux, T., Bergmann, S., and Fankhauser, C. (2014). Plasmamembrane H+-ATPase regulation is required for auxin gradientformation preceding phototropic growth. Mol. Syst. Biol. 10: 751.
Hornitschek, P., Kohnen, M.V., Lorrain, S., Rougemont, J., Ljung,K., López-Vidriero, I., Franco-Zorrilla, J.M., Solano, R., Trevisan,M., Pradervand, S., Xenarios, I., and Fankhauser, C. (2012).Phytochrome interacting factors 4 and 5 control seedling growth inchanging light conditions by directly controlling auxin signaling.Plant J. 71: 699–711.
Huang, J., Ghosh, R., and Bankaitis, V.A. (2016). Sec14-like phos-phatidylinositol transfer proteins and the biological landscape ofphosphoinositide signaling in plants. Biochim. Biophys. Acta 1861:1352–1364.
Ishihara, H., Obata, T., Sulpice, R., Fernie, A.R., and Stitt, M. (2015).Quantifying protein synthesis and degradation in Arabidopsis bydynamic 13CO2 labeling and analysis of enrichment in individualamino acids in their free pools and in protein. Plant Physiol. 168:74–93.
Jouve, L., Gaspar, T., Kevers, C., Greppin, H., and Degli Agosti, R.(1999). Involvement of indole-3-acetic acid in the circadian growthof the first internode of Arabidopsis. Planta 209: 136–142.
Kelly, A.A., Quettier, A.L., Shaw, E., and Eastmond, P.J. (2011).Seed storage oil mobilization is important but not essential forgermination or seedling establishment in Arabidopsis. Plant Physiol.157: 866–875.
Kim, S.J., and Brandizzi, F. (2016). The plant secretory pathway forthe trafficking of cell wall polysaccharides and glycoproteins. Gly-cobiology 26: 940–949.
Kinoshita, T., and Shimazaki, Ki. (1999). Blue light activates theplasma membrane H(+)-ATPase by phosphorylation of the C-terminusin stomatal guard cells. EMBO J. 18: 5548–5558.
Leivar, P., Tepperman, J.M., Monte, E., Calderon, R.H., Liu, T.L.,and Quail, P.H. (2009). Definition of early transcriptional circuitryinvolved in light-induced reversal of PIF-imposed repression of
1314 The Plant Cell

photomorphogenesis in young Arabidopsis seedlings. Plant Cell 21:3535–3553.
Liang, C., Cheng, S., Zhang, Y., Sun, Y., Fernie, A.R., Kang, K.,Panagiotou, G., Lo, C., and Lim, B.L. (2016). Transcriptomic,proteomic and metabolic changes in Arabidopsis thaliana leavesafter the onset of illumination. BMC Plant Biol. 16: 43.
Maris, A., Kaewthai, N., Eklöf, J.M., Miller, J.G., Brumer, H., Fry,S.C., Verbelen, J.P., and Vissenberg, K. (2011). Differences inenzymic properties of five recombinant xyloglucan endotransglucosylase/hydrolase (XTH) proteins of Arabidopsis thaliana. J. Exp. Bot. 62: 261–271.
McFarlane, H.E., Döring, A., and Persson, S. (2014). The cell biologyof cellulose synthesis. Annu. Rev. Plant Biol. 65: 69–94.
Miedes, E., Suslov, D., Vandenbussche, F., Kenobi, K., Ivakov, A.,Van Der Straeten, D., Lorences, E.P., Mellerowicz, E.J., Verbelen,J.P., and Vissenberg, K. (2013). Xyloglucan endotransglucosylase/hydrolase (XTH) overexpression affects growth and cell wall mechanics inetiolated Arabidopsis hypocotyls. J. Exp. Bot. 64: 2481–2497.
Moore, B., Zhou, L., Rolland, F., Hall, Q., Cheng, W.H., Liu, Y.X.,Hwang, I., Jones, T., and Sheen, J. (2003). Role of the Arabidopsisglucose sensor HXK1 in nutrient, light, and hormonal signaling.Science 300: 332–336.
Nagel, D.H., Doherty, C.J., Pruneda-Paz, J.L., Schmitz, R.J., Ecker,J.R., and Kay, S.A. (2015). Genome-wide identification of CCA1targets uncovers an expanded clock network in Arabidopsis. Proc.Natl. Acad. Sci. USA 112: E4802–E4810.
Nozue, K., Covington, M.F., Duek, P.D., Lorrain, S., Fankhauser, C.,Harmer, S.L., and Maloof, J.N. (2007). Rhythmic growth explained bycoincidence between internal and external cues. Nature 448: 358–361.
Nozue, K., Harmer, S.L., and Maloof, J.N. (2011). Genomic analysisof circadian clock-, light-, and growth-correlated genes revealsPHYTOCHROME-INTERACTING FACTOR5 as a modulator of auxinsignaling in Arabidopsis. Plant Physiol. 156: 357–372.
Nusinow, D.A., Helfer, A., Hamilton, E.E., King, J.J., Imaizumi, T.,Schultz, T.F., Farré, E.M., and Kay, S.A. (2011). The ELF4-ELF3-LUX complex links the circadian clock to diurnal control of hypo-cotyl growth. Nature 475: 398–402.
Paredez, A.R., Somerville, C.R., and Ehrhardt, D.W. (2006). Visu-alization of cellulose synthase demonstrates functional associationwith microtubules. Science 312: 1491–1495.
Peaucelle, A., Wightman, R., and Höfte, H. (2015). The control ofgrowth symmetry breaking in the Arabidopsis hypocotyl. Curr. Biol.25: 1746–1752.
Refrégier, G., Pelletier, S., Jaillard, D., and Höfte, H. (2004). In-teraction between wall deposition and cell elongation in dark-grownhypocotyl cells in Arabidopsis. Plant Physiol. 135: 959–968.
Ruan, Y.-L., Jin, Y., Yang, Y.-J., Li, G.-J., and Boyer, J.S. (2010).Sugar input, metabolism, and signaling mediated by invertase: rolesin development, yield potential, and response to drought and heat.Mol. Plant 3: 942–955.
Schopfer, P., Liszkay, A., Bechtold, M., Frahry, G., and Wagner, A.(2002). Evidence that hydroxyl radicals mediate auxin-induced ex-tension growth. Planta 214: 821–828.
Seaton, D.D., Smith, R.W., Song, Y.H., MacGregor, D.R., Stewart,K., Steel, G., Foreman, J., Penfield, S., Imaizumi, T., Millar, A.J.,and Halliday, K.J. (2015). Linked circadian outputs control elon-gation growth and flowering in response to photoperiod and tem-perature. Mol. Syst. Biol. 11: 776.
Smith, A.M., and Stitt, M. (2007). Coordination of carbon supply andplant growth. Plant Cell Environ. 30: 1126–1149.
Spartz, A.K., Ren, H., Park, M.Y., Grandt, K.N., Lee, S.H., Murphy,A.S., Sussman, M.R., Overvoorde, P.J., and Gray, W.M. (2014).SAUR inhibition of PP2C-D phosphatases activates plasma-membraneH+-ATPases to promote cell expansion in Arabidopsis. Plant Cell 26:2129–2142.
Stewart, J.L., Maloof, J.N., and Nemhauser, J.L. (2011). PIF genesmediate the effect of sucrose on seedling growth dynamics. PLoSOne 6: e19894.
Stitt, M., and Zeeman, S.C. (2012). Starch turnover: pathways, reg-ulation and role in growth. Curr. Opin. Plant Biol. 15: 282–292.
Sulpice, R., Flis, A., Ivakov, A.A., Apelt, F., Krohn, N., Encke, B.,Abel, C., Feil, R., Lunn, J.E., and Stitt, M. (2014). Arabidopsiscoordinates the diurnal regulation of carbon allocation and growthacross a wide range of photoperiods. Mol. Plant 7: 137–155.
Suslov, D., Ivakov, A., Boron, A.K., and Vissenberg, K. (2015). Invitro cell wall extensibility controls age-related changes in thegrowth rate of etiolated Arabidopsis hypocotyls. Funct. Plant Biol.42: 1068–1079.
Suslov, D., and Verbelen, J.P. (2006). Cellulose orientation determinesmechanical anisotropy in onion epidermis cell walls. J. Exp. Bot. 57:2183–2192.
Suslov, D., Verbelen, J.P., and Vissenberg, K. (2009). Onion epi-dermis as a new model to study the control of growth anisotropy inhigher plants. J. Exp. Bot. 60: 4175–4187.
Vardi, Y., and Zhang, C.H. (2000). The multivariate L1-median andassociated data depth. Proc. Natl. Acad. Sci. USA 97: 1423–1426.
Voxeur, A., and Höfte, H. (2016). Cell wall integrity signaling in plants:“To grow or not to grow that’s the question”. Glycobiology 26: 950–960.
Wang, J., Nayak, S., Koch, K., and Ming, R. (2013). Carbon parti-tioning in sugarcane (Saccharum species). Front. Plant Sci. 4: 201.
Wiese, A., Christ, M.M., Virnich, O., Schurr, U., and Walter, A.(2007). Spatio-temporal leaf growth patterns of Arabidopsis thalianaand evidence for sugar control of the diel leaf growth cycle. NewPhytol. 174: 752–761.
Wolf, S., Hématy, K., and Höfte, H. (2012). Growth control and cellwall signaling in plants. Annu. Rev. Plant Biol. 63: 381–407.
Yadav, U.P., Ivakov, A., Feil, R., Duan, G.Y., Walther, D., Giavalisco,P., Piques, M., Carillo, P., Hubberten, H.M., Stitt, M., and Lunn,J.E. (2014). The sucrose-trehalose 6-phosphate (Tre6P) nexus:specificity and mechanisms of sucrose signalling by Tre6P. J. Exp.Bot. 65: 1051–1068.
Yazdanbakhsh, N., Sulpice, R., Graf, A., Stitt, M., and Fisahn, J.(2011). Circadian control of root elongation and C partitioning inArabidopsis thaliana. Plant Cell Environ. 34: 877–894.
Regulation of Cell Wall Synthesis and Growth 1315

DOI 10.1105/tpc.16.00782; originally published online May 26, 2017; 2017;29;1305-1315Plant Cell
Friedrich Kragler, Kris Vissenberg, Staffan Persson and Dmitry SuslovAlexander Ivakov, Anna Flis, Federico Apelt, Maximillian Fünfgeld, Ulrike Scherer, Mark Stitt,
Arabidopsis HypocotylsCellulose Synthesis and Cell Expansion Are Regulated by Different Mechanisms in Growing
This information is current as of July 20, 2018
Supplemental Data /content/suppl/2017/07/31/tpc.16.00782.DC2.html /content/suppl/2017/05/26/tpc.16.00782.DC1.html
References /content/29/6/1305.full.html#ref-list-1
This article cites 70 articles, 21 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists