carbachol stimulates cl− secretion via activation of two distinct apical cl− pathways in...
TRANSCRIPT

Biochimica et Biophysica Acta, 1220 (1994) 241-247 241 © 1994 Elsevier Science B.V. All rights reserved 0167-4889/94/$07.00
BBAMCR 13506
Carbachol stimulates C1- secretion via activation of two distinct apical C1- pathways in cultured human T84
intestinal epithelial monolayers
Gordon T.A. McEwan *, Barry H. Hirst and Nicholas L. Simmons Gastrointestinal Drug Delivery Research Centre, Department of Physiological Sciences, University of Newcastle upon Tyne,
Medical School, Newcastle upon Tyne, NE2 4HH (UK)
(Received 21 April 1993) (Revised manuscript received 29 July 1993)
Key words: Carbachol; T84 cell; Intestine; Epithelial cell; Chloride secretion; (Human)
The mode of action of carbachol in stimulation of transepithelial CI- secretion in intact human intestinal Ta4 epithelial monolayers has been investigated in order to determine whether a DIDS-insensitive exit pathway (via CFTR) coexists with a DIDS-sensitive exit pathway at the apical membrane. Carbachol stimulates a transient inward lsc due to CI- secretion whose magnitude is related to the basal level of inward I~. The inward current responses to both carbachol and hypo-osmotic media are abolished in nominally Ca2+-free media. The action of apical DIDS (100/~M) upon carbachol-stimulated I~ depends on the initial value of the basal Isc. At basal Iu levels < 10/zA cm -2, 100/zM DIDS applied to the apical cell border abolishes the inward I~ following exposure to both carbachol and hypo-osmotic media. In contrast a VIP-stimulated inward Isc is observed in the presence of 100 tzM DIDS. After VIP stimulation of inward Isc, or if spontaneous basal values of Isc were > 10/~A cm -2, the carbachol stimulation of inward I~ was largely insensitive to 100/zM DIDS. The data are consistent with the participation of both DIDS-sensitive and DIDS insensitive pathways for C1- at the apical membrane of human intestinal T84 epithelial cells.
I n t r o d u c t i o n
There is considerable current interest concerning the CI- exit pathways across the apical plasma mem- brane of intestinal enterocytes. To date, the human intestinal cell line, T84, has provided the best studied model for intestinal chloride secretion. Whole cell patch-clamp studies have demonstrated that these cells express at least three pharmacologically and kinetically distinguishable CI- channels which are activated re- spectively by cAMP, Ca 2÷ and cell swelling [1-5]. A number of studies have demonstrated the effectiveness of carbachol and Ca2+-mobilizing stimuli in the stimu- lation of transepithelial CI- secretion in T84 epithelial monolayers [6-10]. However, controversy exists con- cerning the exact mechanism by which such stimuli act.
Wong et al., using simultaneous measurements of cytosolic Ca 2 + and basolateral K + conductance in T84 epithelia following amphotericin permeabilization of the apical membrane [9], concluded that carbachol
* Corresponding author.
SSDI 0167-4889(93)E0155-S
stimulated activation of GK by Ca 2+ is essential to the activation of C1- secretion. In another study in which the basolateral membrane was permeabilized with nys- tatin, Andersen and Welsh [1] have measured apical membrane CI- currents and find no evidence for a Ca2+-activated CI- exit pathway in the apical mem- brane. They conclude that the mechanism of carbachol stimulation of transepithelial CI- secretion must occur by activation of basolateral K ÷ conductance alone, thereby hyperpolarizing the cell and increasing the driving force for C1- exit across pre-existing C1- chan- nels in the apical membrane. This view is supported by the failure of conventional patch-clamp techniques to show activation of a CI- conductance upon carbachol stimulation in these cells [11] despite the evidence for the existence of cae÷-activated C1- conductances in these cells.
Activation of rheogenic CI- secretion by cAMP mediated stimuli is associated with activation of the small conductance (5-10 pS) cAMP-activated C1- channel which is insensitive to inhibition by extracellu- lar DIDS (4,4'-diisothiocyanantostilbene-2,2'-disul- phonic acid) [5,12]. Recently we have shown that expo-

242
sure of T84 epithelia to hypo-osmotic solutions stimu- lates CI- secretion and that this was blocked by extra- cellular DIDS, suggesting that both DIDS-sensitive and DIDS-insensitive C1- pathways may exist in intact T84 epithelia [13]. Since cell swelling was also observed to elevate intracellular Ca 2÷ [13] the possibility exists that Ca 2÷ mobilizing stimuli, such as carbachol, might also access this DIDS-sensitive apical CI- pathway. A recent re-evaluation of carbachol stimulation of whole cell CI- conductances (using the perforated patch- clamp technique to limit intracellular dialysis) has shown that carbachol may, in addition to stimulation of a K ÷ conductance, also activate C1--selective and non-selective cation conductances [14].
As with the swelling-induced C1- channel, activity of the outwardly rectifying C1- channel (30-50 pS) [4,5] is also blocked by extracellular DIDS [4,5]. How- ever although the two channels display similar kinetic characteristics it is still unclear whether there is only one channel which displays different states or whether separate channels exist [4]. The debate concerning the participation of outwardly rectifying channels in C1- secretion has been reopened by the observation that CFTR transfection can restore cAMP activation of outwardly rectifying C1- channels [15]. In addition, Sorscher et al. [16] have identified a membrane protein using antibodies directed against a DIDS-binding pep- tide whose properties are consistent with the outward- rectifier, which migrates from intracellular locations in T84 cells to the apical surface upon forskolin (cAMP) stimulation.
The purpose of the present investigation has been to determine whether carbachol stimulation of CI- secre- tion in intact T84 epithelia is consistent with activation of a single or multiple C1- pathways in the apical membrane. To this end we have utilized the DIDS sensitivity of carbachol-stimulated C1- secretory cur- rents in intact T84 epithelia as a prime criterion.
Materials and Methods
Cell culture T84 cells were obtained from the ATCC (Roclccille,
MA, USA). T84 cells were grown in a 1:1 mixture of Dulbecco's modified Eagle's medium and Ham's F-12 with 5% newborn calf serum and 200 IU m1-1 peni- cillin, 200 /~g m1-1 streptomycin. Confluent monolay- ers were subcultured every 14-16 days, by treatment with 0.05% trypsin and 0.2% EDTA in Ca 2÷- and Mg2+-free phosphate-buffered saline (PBS). The cul- tures were incubated at 37°C in a humidified atmo- sphere of 5% CO2/95% air.
Functional epithelial layers of T84 cells on perme- able supports were prepared essentially as described by Dharmsathaphorn et al. [17] and Simmons [18]. Cells were seeded at high density (1.106 cells m1-1, 0.5
ml/well) into rat-tail collagen coated fiitercups (Nun~ tissue culture inserts, 10 mm diameter, inorganic mere brane matrix). Filtercups were then cultured in 6-wel plates at 37°C, 5% CO 2 for 2 weeks to ! month witI medium replacement every 2-3 days. The formation o: functional epithelial layers was monitored by the devel opment of a significant transepithelial resistance (R T) as measured using a WPI Evometer fitted with 'chop. stick' electrodes to allow transepithelial current pas sage and potential sensing [18]. Cell monolayers werc used when the transepithelial resistance typically ex. ceeded 900 S2 cm 2 (values of resistance for filter cups approx. 300 ~ cm 2 are subtracted from all data).
Measurements of short-circuit current and transepithelia, conductance
Cultured epithelial layers were mounted in Ussing type chambers maintained at 37°C, connected to ar automatic voltage clamp via KC1/agar salt-bridges and reversible electrodes (Ag/AgC1 for current passage calomel for voltage sensing) and measurements ol open-circuit electrical p.d., transepithelial conductance (G x) and short-circuit current (Isc) made in modified Krebs solutions (see below).
Solutions The basic Krebs solution was (all mmol/l) NaCI, 70:
KCI, 5.4; CaC12, 2.8; MgSO4, 1.2; NaH2PO4, 0.3', KH2PO4, 0.3; Hepes, 10; mannitol, 140; glucose, 1£ (pH 7.4 at 37°C) with 1% (v/v) donor horse serum./~ hypotonic solution was prepared as above except man- nitol was omitted. A Cl--free medium was obtained b5 replacement of NaCI and KCI by their methane- sulphonate salts and buffering with Tris-Hepes. Nomi. nally Ca2+-free solutions were prepared as above bul with CaCI2 omitted, and with the addition of ethylene- diaminetetraacetic acid (EDTA; 1 mM).
Statistics Variation in mean data is expressed as the standard
error of the mean (S.E.). Tests of significance of differ- ences between mean values were made using a two- tailed Student's t-test (for paired or unpaired mea~ data), or Mann-Whitney U/Wilcoxan signed rank tesl (for unpaired or paired data, respectively) where ap- propriate.
Materials Salts and biochemicals were obtained from BDI-]
Chemicals (Poole, Dorset) or Sigma and were of AnalaJ grade or equivalent. 4,4'-diisothioeyanostilbene-2,2'di- sulphonic acid (DIDS) and EDTA were from Sigma Bumetanide was a gift from Leo Laboratories (Prince., Risborough, UK). Stock solutions of DIDS anc bumetanide were made in 1 mM Tris.

Results
Carbachol stimulation of I~ is dependent upon magni- tude of initial Is~
Carbachol action (100 /xM) upon inward Is~ in T84
epithelial monolayers is characterized by a rapid rise from basal values of 5.6 + 0.8/~A cm -2 (n = 9) to near maximum values of inward I~ of 20.0 + 3.6 ~ A cm -2 within 0.5 min. Thereafter, I~c declines towards pre- stimulation values over 5 min (Fig. 1). The increased inward I~c with carbachol is accompanied by an in- creased tissue conductance from 1.70 + 0.16 mS cm -2 to 3.48 +__ 1.15 mS cm -2 (n = 9 paired data, P < 0.005) 0.5 min after carbachol addition. There is a significant correlation between the magnitude of the basal I~ and the subsequent response to carbachol (Fig. 1 inset); with low basal values of inward I~ (C1- secretion) the magnitude of the response to carbachol is attenuated. In contrast to the detailed knowledge that the ionic basis of the response to carbachol represents CI- se- cretion [2], basal values of Is~ and net Na and C1- flux are usually small and close to zero [2]. Replacement of medium C1- by methanesulphonate salts effectively abolishes the basal I~c from 2.3 + 0.3 /~A cm -z to
243
0.3 5:0.15/~A cm -2 (n -- 3) after 1 min. The basal Isc is only slightly reduced by bumetanide (from 1.9 _+ 0.1 to 1.6 _+ 0.2 /~A cm -2 ( n - - 5 ) ) at 1 min, but is more sensitive to apical addition of DIDS (100/~M; reduced from 2.0 ___ 0.3 to 0.8 _+ 0.3 /zA cm -2 (n = 5)). These data suggest that the basal Isc, as is carbachol-stimu- lated I~¢, is dependent upon Cl- secretion.
Repeated additions of carbachol (after washing and 20 min recovery) give rise to a further inward Isc response though this is diminished compared to the initial response (from 21.7 + 3.9 to 10.5 _+ 1.49/xAcm -2 (n = 6)). This reduction is accompanied by a parallel reduction in the basal I,c from 5.5 __ 0.8 to 2.8 _+ 0.4 /xA cm -2.
Relation between carbachol and hypotonic stimulated Isc: dependence on extracellular Ca 2 ÷
Cell swelling induced by hypo-osmotic exposure of the basolateral cell surfaces may stimulate an inward Isc resulting from transepithelial CI- secretion in T84 epithelia [13]. Both carbachol and hypo-osmotic expo- sure of the basolateral surfaces stimulate inward Isc in the same batch of monolayers (Fig. 2). When the paired responses to carbachol and hypo-osmotic expo-
¢q I
E tJ
:=L w
O
2 5
20
1 5
1 0
C a r b a c h o l
T
, II
1 2 0
~E 1 0 0
u. 8O
/ . ,--','"." o d "-~'J. "
I .L \ o 5 ,o ,5
I ~ elllnll l.e I/~A cm t)
! I I I I I
0 5 6 7 8 9 10
s ~ - "
s s ~
s
i
2O
T i m e ( m i n ) Fig. 1. Transient st imulation of inward Isc by carbachoi in T84 epithelial monolayers. Data are means 5: S.E. of nine T84 monolayers from three separate batches. Carbachol (100/~M) was added to the basolateral solution at the time indicated by the arrow. Inset: dependence of the size of the response to carbachol upon the initial resting Is: in individual Ts4 monolayers; the dashed line is the linear regression line for the data
y = 4.94x - 1.55 (d.f. = 43, r = 0.91, P < 0.001).
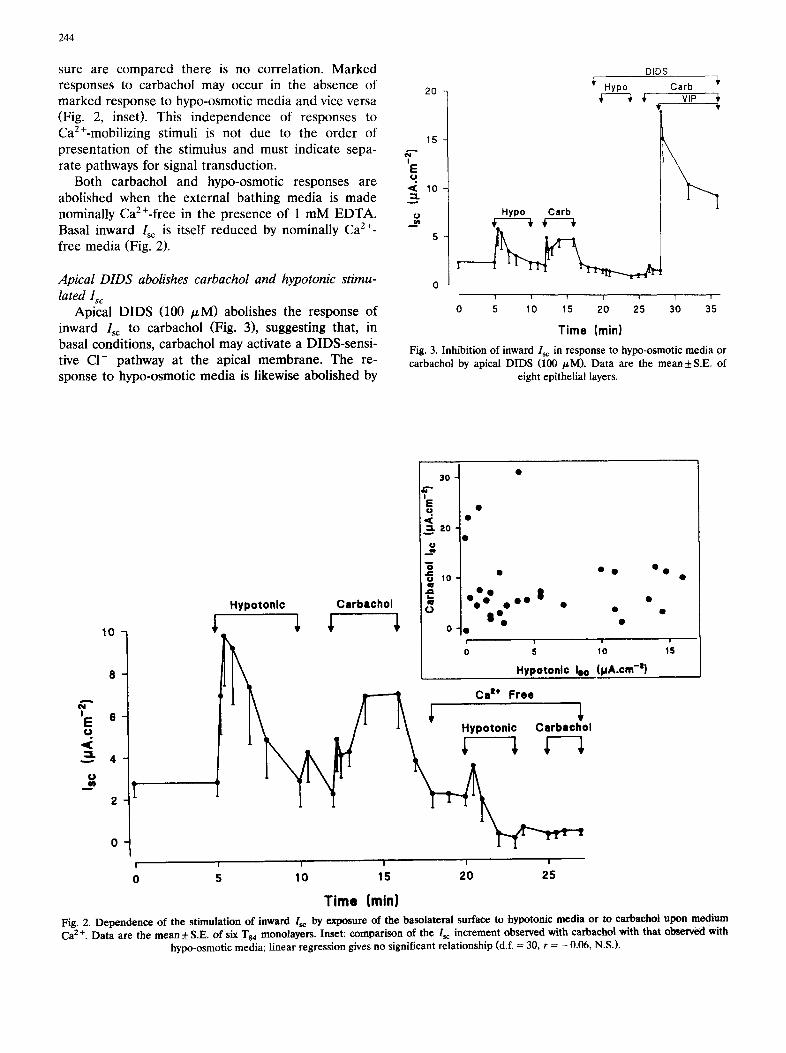
244
sure are compared there is no correlation. Marked responses to carbachol may occur in the absence of marked response to hypo-osmotic media and vice versa (Fig. 2, inset). This independence of responses to Ca2+-mobilizing stimuli is not due to the order of presentation of the stimulus and must indicate sepa- rate pathways for signal transduction.
Both carbachol and hypo-osmotic responses are abolished when the external bathing media is made nominally Ca2+-free in the presence of 1 mM EDTA. Basal inward Isc is itself reduced by nominally Ca z÷- free media (Fig. 2).
Apical DIDS abolishes carbachol and hypotonic stimu- lated Isc
Apical DIDS (100 /zM) abolishes the response of inward Isc to carbachol (Fig. 3), suggesting that, in basal conditions, carbachol may activate a DIDS-sensi- tive C1- pathway at the apical membrane. The re- sponse to hypo-osmotic media is likewise abolished by
20
15
I E ( J
• < 10 : : : L
5
DIDS
Hypo Carb ~ VIP ~,
Hypo Carb
I I i f I I I I
0 5 10 15 20 25 30 35
Time (rain) Fig. 3. Inhibition of inward Isc in response to hypo-osmotic media or carbachol by apical DIDS (100 /~M). Data are the mean :t: S.E. of
eight epithelial layers.
10
I E 6
~ 4 o
2
! I
0 5
Hypo ton l c
3 0 •
. . <= 20 g
~ 10 • ° O ° O
e o~o" o , 8 • Carbschol r~ • • 8oe • • •
o o
o , o
_ Hypotonl¢ leo (tJA.cm -z)
~----~ Cat+Free ~ _ / " A \ Hypo ton l c Ca rbacho l
I I I I
10 15 20 25
Time (min)
r - ' - ' - - ' -
15
Fig. 2. Dependence of the stimulation of inward Isc by exposure of the basolateral surface to hypotonic media or to carbachol upon medium Ca 2+. Data are the mean + S.E. of six T ~ monolayers. Inset: comparison of the /'sc increment observed with earbaehol with that observed with
hypo-osmotic media; linear regression gives no significant relationship (d.f. = 30, r = -0 .06, N.S.).
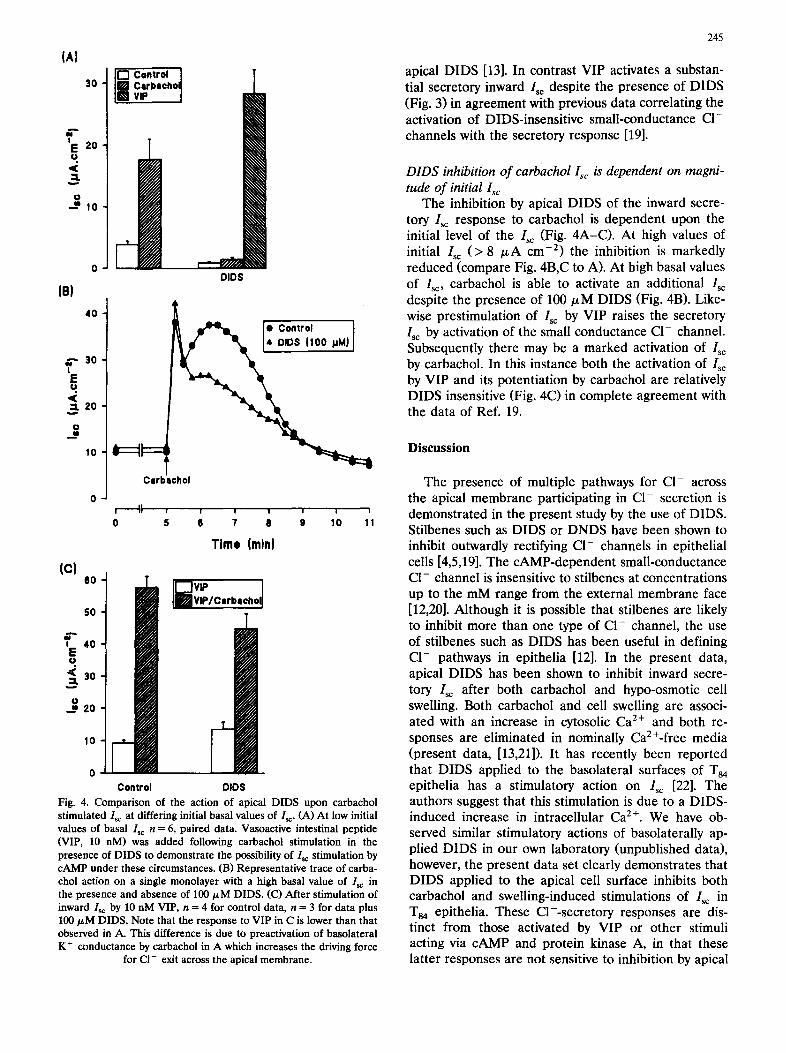
(AI
30 [~ cnntr°~ / Carbechoq vie I
, 0
° t o
0 I - ~/I~N//~
'i I 4O
, o
u
2o
r chol
0 , I I , , , , , , ,
0 5 $ 7 I! 9 10 11
"rime (mini
(cJ . o /il vu' I
V I P / O i r b i e h o l
50
g 2o T
,o I 0
C o n t r o l DID$ Fig. 4. Comparison of the action of apical DIDS upon carbachol stimulated I~ at differing initial basal values of I~. (A) At low initial values of basal Isc n = 6, paired data. Vasoactive intestinal peptide (VIP, 10 nM) was added following carbachol stimulation in the presence of DIDS to demonstrate the possibility of I~ stimulation by cAMP under these circumstances. (B) Representative trace of carba- chol action on a single monolayer with a high basal value of I~ in the presence and absence of 100/zM DIDS. (C) After stimulation of inward Is¢ by 10 nM VIP, n = 4 for control data, n = 3 for data plus 100/~M DIDS. Note that the response to VIP in C is lower than that observed in A. This difference is due to preactivation of basolateral K + conductance by carbachol in A which increases the driving force
for CI- exit across the apical membrane.
245
apical DIDS [13]. In contrast VIP activates a substan- tial secretory inward Is~ despite the presence of DIDS (Fig. 3) in agreement with previous data correlating the activation of DIDS-insensitive small-conductance CI- channels with the secretory response [19].
DIDS inhibition of carbachol Isc is dependent on magni- tude of initial Isc
The inhibition by apical DIDS of the inward secre- tory Isc response to carbachol is dependent upon the initial level of the Isc (Fig. 4A-C). At high values of initial I~c (> 8 /zA cm -2) the inhibition is markedly reduced (compare Fig. 4B,C to A). At high basal values of I~:, carbachol is able to activate an additional Isc despite the presence of 100/xM DIDS (Fig. 4B). Like- wise prestimulation of I~c by VIP raises the secretory I~ by activation of the small conductance C1- channel. Subsequently there may be a marked activation of I~ by carbachol. In this instance both the activation of Isc by VIP and its potentiation by carbachol are relatively DIDS insensitive (Fig. 4C) in complete agreement with the data of Ref. 19.
Discussion
The presence of multiple pathways for Cl- across the apical membrane participating in C1- secretion is demonstrated in the present study by the use of DIDS. Stilbenes such as DIDS or DNDS have been shown to inhibit outwardly rectifying C1- channels in epithelial cells [4,5,19]. The cAMP-dependent small-conductance C1- channel is insensitive to stilbenes at concentrations up to the mM range from the external membrane face [12,20]. Although it is possible that stilbenes are likely to inhibit more than one type of C1- channel, the use of stilbenes such as DIDS has been useful in defining C1- pathways in epithelia [12]. In the present data, apical DIDS has been shown to inhibit inward secre- tory I~ after both carbachol and hypo-osmotic cell swelling. Both carbachol and cell swelling are associ- ated with an increase in cytosolic Ca 2+ and both re- sponses are eliminated in nominally Ca2+-free media (present data, [13,21]). It has recently been reported that DIDS applied to the basolateral surfaces of Ts4 epithelia has a stimulatory action on Isc [22]. The authors suggest that this stimulation is due to a DIDS- induced increase in intracellular Ca 2+. We have ob- served similar stimulatory actions of basolaterally ap- plied DIDS in our own laboratory (unpublished data), however, the present data set clearly demonstrates that DIDS applied to the apical cell surface inhibits both carbachol and swelling-induced stimulations of Isc in T84 epithelia. These C1--secretory responses are dis- tinct from those activated by VIP or other stimuli acting via cAMP and protein kinase A, in that these latter responses are not sensitive to inhibition by apical

246
DIDS. In addition, the activation of CI- secretion upon carbachol or cell swelling alone is transient, whereas the Cl- secretion activated by VIP is sus- tained. The present data are consistent with a recent perforated patch whole cell study which demonstrated a carbachol-activated CI- current [14]. The magnitude of the response to carbachol was usually small com- pared to the VIP-stimulated inward Is~. In addition, when the basal Isc was large or prestimulated with VIP no appreciable DIDS inhibition was observed. These data are thus consistent with the predominant C1- pathway in the apical membrane of T84 being DIDS-in- sensitive in agreement with the data of Tabchararni et al. [12]. In addition, it seems unlikely that after VIP stimulation an additional DIDS-sensitive pathway has been recruited to the apical membrane [15,16].
The finding of a carbachol activated Cl- secretion by an apical C1- pathway separate from CFTR is similar to carbachol activation of Cl- secretion in tracheal epithelium and airway cells [23-25] and is also consistent with the finding in cultured HT29.c119a ep- ithelia that Ca 2 + and cAMP activate different chloride efflux pathways [26,27].
Swelling and Ca2+-activated C1- currents have been extensively characterized in T84 cells [2-4,28]. The lo- cation of such channels has been the subject of debate. Although CaE+-stimulated Cl- currents may be easily observed with whole cell patch-clamp, CaE+-activated O--secretory currents are not evident under condi- tions of basolateral membrane permeabilization and transepithelial Cl- gradients [1]. The location of the CaE+-activated C1- channels might be considered to be the basolateral membrane [6]. In MDCK epithelial layers where a volume activated Cl- conductance oc- curs, activation of this conductance by cell swelling gives rise to a prolonged inhibition of all secretory transepithelial currents [29]. Since no inhibition of cAMP-activated secretion is observed in T84 epithelia upon carbachol or cell swelling, we infer that the location of the C1- conductance activated by cell swelling and /o r Ca 2+ demonstrated in the present study must be predominantly at the apical membrane. The presence of Ca2+-activated CI- pathways at both membrane faces has been inferred in HT29.cl19a cells where Ca2+-mobilizing stimuli do not generate rheogenic CI- secretion [26,27].
Both carbachol and hypotonic media increase cy- tosolic Ca 2+ [21], the increase in Ca 2÷ due to carba- chol being larger than that observed following hypo- tonic exposure. Comparing the responses to carbachol and hypotonic media, it is evident that the tissue re- sponse to these two separate stimuli may be unrelated suggesting that different mechanisms may be operative in stimulation of the DIDS-sensitive C1- pathway(s). Since intracellular Ca 2+ measurements have not been correlated with inward Isc in the present data we
cannot conclude that parallel activation pathways ir addition to Ca 2+ mobilization exist. However, multipk calcium mediated effector mechanisms have beer demonstrated with taurodeoxycholate, histamine, car. bachol and Ca2+-ionophores [30]. The participation 01 a Ca 2 +-calmodulin complex in Ca 2 +-mediated secretior in T84 epithelium has been suggested by calmodulir antagonists and inhibitory peptides [3]. There appears, to be no participation of protein kinase C in carbachol'~ secretory action in T84 cells [31]. In contrast proteir kinase C has been shown to increase open state proba- bility of CFTR appreciably only after prior stimulatio~ by protein kinase A [19].
In addition to providing an exit pathway for C1- across the apical membrane, stimulation of trans- epithelial Cl- secretion by carbachol also requires the carriage of current across the basolateral membrane. In secretory epithelia this is accomplished either by the Na+/K+-ATPase or by Ca2+-activated K + conduc- tances [32]. Exposure to carbachol or cell swelling increases cytosolic Ca 2+ resulting in an increased di- uretic-insensitive K + efflux and K + conductance across the basolateral membrane [9]. After pre-stimulation ot Isc by VIP, it is likely that the augmentation of inward Isc by carbachol and hypotonic swelling is due to baso- lateral K + conductance activation, thereby hyperpolar- izing the cell and increasing the driving force for C1- exit across the apical membrane. Greenwald and Biagi [33] have recently demonstrated in colonic crypts that resting membrane p.d. is close to E~ and that carba- chol hyperpolarization of basolateral membrane p.d. is only pronounced after VIP has depolarized the cell potential. In the present study, the nature of the re- sponse to carbachol differs whether there is a large basal or stimulated inward Isc. The variability in Isc in T84 epithelia has been noted by other groups [34]. However, the stimulus underlying the high basal in- ward Isc is not known. Since cultured renal and airway epithelia are known to produce endogenous prosta- glandins [35-37] and since these are known to stimu- late Cl- secretion in T84 epithelia [38] an endogenous prostaglandin production may account for the variable levels of basal I~ noted in the present study. The data are both consistent with DIDS-sensitive and DIDS-in- sensitive Cl- pathways existing in parallel at the apical membrane of Ts4 cells, and with the absolute amount of transepithelial C1- secretion being determined by the electrochemical gradient for C1- across the apical membrane. The Cl- electrochemical gradient will be determined primarily by the Ca2+-dependent K + con- ductance(s) and the electroneutral cotransporter pres- ent at the basolateral membrane. It is possible, there- fore, that intestinal cells possess alternative pathways for the mediation of transepithelial C1- secretion which are independent of the cAMP-activated DIDS-insensi- tive pathway.

Acknowledgements
Supported under the LINK programme in Selective Drug Delivery and Targeting funded by SERC/ MRC/DTI/Industry (SERC Grant GR/F 09747). We wish to thank Maxine Geggie for excellent technical assistance.
References
1 Andersen, M.P. and Welsh, M.J. (1991) Proc. Natl. Acad. Sci. USA 88, 6003-6007.
2 Cliff, W.H. and Frizell, R.A. (1990) Proc. Natl. Acad. Sci. USA 87, 4956-4960
3 Worrell, R.T. and Frizzell, R.A. (1991) Am. J. Physiol. 260, C877-C882.
4 Solc, C.K. and Wine, J.J. (1991) Am. J. Physiol. 261, C658-C674. 5 Widdicombe, J.H. and Wine, J.J. (1991) Trends Biochem. Sci. 16,
474-477 6 Bolton, J.E. and Field, M. (1977) J. Membr. Biol. 35, 159-173. 7 Dharmsathaphorn, K. and Pandol, S.J. (1986) J. Clin. Invest. 77,
348-354. 8 Reinlib, L., Mikkelsen, R., Zahniser,, D., Dharmsathaphorn, K.
and Donowitz, M. (1989) Am. J. Physiol. 257, G950-G960. 9 Wong, S.M.E., Tesfaye, A., DeBell, M.C. and Chase, H.E. (1990)
J. Gen. Physiol. 96, 1271-1285. 10 Cartwright, C.A., MeRoberts, J,A., Mandel, K.G. and Dharm-
sathaphorn, K. (1985) J. Clin. Invest. 76, 1837-1842. 11 Devor, D.C., Simasko, S.M. and Duffey, D.E. (1990) Am. J.
Physiol. 258, C318-C326. 12 Tabcharani, J.A., Low, W., Elie, D. and Hanrahan, J.W. (1990)
FEBS Lett. 270, 157-164. 13 McEwan, G.T.A., Brown C.D.A., Hirst, B.H. and Simmons, N.L.
(1992) Biochim. Biophys. Acta 1135, 180-183. 14 Devor, D.C. and Duffey, M.E. (1992) Am. J. Physiol. 263, C780-
C787 15 Egan, M., Flotte, T., Aflone, S., Solow, R., Zeitlin, P.L. Carter,
B.J. and Guggino, W.B. (1992) Nature 358, 581-584. 16 Sorscher, E.J., Fuller, C.M., Bridges, R.J., Tousson, A., Mar-
247
chase, R.B., Brinkley, B.R., Frizzell, R.A. and Benos, D.J. (1992) Am. J. Physiol. 262, C136-C147.
17 Dharmsathaphorn, K., McRoberts, J.A., Mandel, K.G., Tisdale, L.D. and Masui, H. (1984) Am. J. Physiol. 246, G204-G208.
18 Simmons, N.L. (1990) Methods Enzymol. 191, 426-437. 19 Tabcharani, J.A., Chang, X.-B., Riordan, J.R. and Hanrahan,
J.W. (1991) Nature 352, 628-631. 20 Sullivan, S., Schick, R., Gregory, R., Paul, S., Agellon, L. and
Field, M. (1992) Pediat. Pulmonol. Suppl. 8, 127. 21 MeEwan, G.T.A., Brown, C.D.A., Hirst, B.H. and Simmons, N.L.
(1993) Pfliigers Arch. 423, 213-220. 22 Brayden, D.J., Krouse, M.E., Law, T. and Wine, J.J. (1993) Am.
J. Physiol. 264, G325-G333. 23 McCann, J.D. and Welsh, M.J. (1989) J. Gen. Physiol. 94, 1015-
1036. 24 Wagner, J.A., Cozens, A.L., Schulman, H., Gruenert, D.C., Stryer,
L. and Gardner, P. (1991) Nature 28, 793-796. 25 Chan, H.-S., Goldstein, J. and Nelson, D.J. (1992) Am. J. Physiol.
262, C1273-C1283. 26 Vaandrager, A.B., Bajnath, R., Groot, J.A., Bot, A.G.M. and De
Jonge, H.R. (1991) Am. J. Physiol. 261, G958-965. 27 Bajnath, R.B., Vaandrager, A.B., Dekker, K., De Jonge, H.R.
and Groot, J.A. (1992) J. Membr. Biol. 127, 81-94. 28 Worrell, R.T., Butt, G.A., Cliff, W.H. and Frizzell, R.A. (1989)
Am. J. Physiol. 256, Cl111-Cl119. 29 Simmons, N.L (1991) Pfliigers Arch. 419, 572-578. 30 Dharmsathaphorn, K., Cohn, J. and Beuerlein, G. (1988) Am. J.
Physiol. 256, C1224-C1230. 31 Lindeman, R.P. and Chase, H.S. (1992) Am. J. Physiol. 263,
C140-C146 32 Petersen, O.H. and Maruyama, Y. (1984) Nature 23, 693-696. 33 Greenwald, L. and Biagi, B.A. (1992) Am. J. Physiol. 262, G940-
G944. 34 Field, M. and Semrad, C~E. (1993) Annu. Rev. Physiol. 55,
631-655. 35 Simmons N.L. (1991) J. Physiol. 432, 459-472. 36 Aboolian, A., Vander Molen, M. and Nord, E.P. (1989) Am. J.
Physiol. 256, Fl135-Fl143 37 Smith, J.J., McCann, J.D. and Welsh, M.J. (1990) Am. J. Physiol.
258, L369-L377. 38 Weymer, A., Huott, P., Liu, W., McRoberts, J.A. and Dharm-
sathaphorn, K. (1985) J. Clin. Invest. 76, 1828-1836.