calcium- and calmodulin-regulated phosphorylation ofsoluble
TRANSCRIPT

Plant Physiol. (1984) 76, 359-3650032-0889/84/76/0359/07/$0l .00/0
Calcium- and Calmodulin-Regulated Phosphorylation of Solubleand Membrane Proteins from Corn Coleoptiles'
Received for publication February 9, 1984 and in revised form June 4, 1984
K. VELUTHAMBI AND B. W. POOVAIAH*Department ofHorticulture and Landscape Architecture, Washington State University,Pullman, Washington 99164-6414
ABSTRACT
In vitro phosphorylation ofseveral membrane polypeptides and solublepolypeptides from corn (Zea mays var. Patriot) coleoptiles was promotedby adding Ca2. Ca2e-promoted phosphorylation of the membrne poly-peptides was further increased in the presence of calmodulin. Both Ca2l-stimulated and Ca2"- and calmodulin-stimulated phosphorylations ofmembrane polypeptides were inhibited by chlorpromane, a calmodulinantagonist. Ca2@-stimulated phosphorylation of soluble polypeptides in-creased with increasing Ce2 concentration. The calmodulin antagonistschlorpromazine and trifluoperazine inhibited the Ca2'-promoted phos-phorylation of soluble polypeptides. Added calmodulin promoted theCa2l-dependent phosphorylation of a 98 kilodaltoas polypeptide. BothCa2l-dependent and Ca2l-independent phosphorylations required Mg2+at an optimal concentration of 5 to 10 millimolar. Cyclic AMP was foundto have no stimulatory effect on protein phosphorylation. Sodium molyb-date, an inhibitor of protein phosphatase, increased the net phosphory-lation of several polypeptides. Rapid loss of radioactivity from thephosphorylated polypeptides following incubation in unlabeled ATP in-dicated the presence of phosphoprotein phosphatase activity.
the possible involvement ofcalmodulin in geotropism (3) and invarious hormonal responses (10, 26). Evidence has been obtainedfor the presence of endogenous calmodulin inhibitors in plantswhich could have a role in the in vivo regulation of calmodulinactivity (20). Direct estimation of Ca2' in the alga, Chara,revealed that an increase in free cyctolasmic Ca2+ in response toaction potentials could explain Ca2+-modulated inhibition ofcytoplasmic streaming (35). These evidences suggest a pivotalrole for Ca2+ as a second messenger in plants as in animal cells.
Protein phosphorylation is an important mechanism by whichCa2+ and calmodulin regulate biochemical events inside the cell(8). Protein phosphorylation in higher plants has been reportedfor nuclear proteins (18, 19, 32) chloroplast proteins (2), ribo-somal proteins (11, 15), and for cytokinin-binding protein (23).Recently, the role of Ca2+ and calmodulin in regulating proteinphosphorylation in plants has been demonstrated (12, 22, 27,30, 33). In this paper we provide additional information tosupport the role of Ca2+ and calmodulin in regulating the phos-phorylation of membrane and soluble proteins from corn co-leoptiles. We also present evidence for the existence of phospho-protein phosphatase activity which could be involved in thedephosphorylation of phosphorylated proteins.
Protein phosphorylation catalyzed by cAMP-dependent andCa2+- and calmodulin-activated protein kinases is now recog-nized to be the major general mechanism by which intracellularevents in mammalian tissues are controlled in response to exter-nal stimuli (8). To find the existence of parallel mechanisms inplants, attempts have been made to explore the role ofcAMP asa second messenger in plants. Although convincing evidence wasobtained for the existence ofcAMP in plants (5), all attempts todemonstrate a stimulatory effect of cAMP on phosphorylationofplant proteins have failed (1 1, 15, 18, 19, 30). Therefore, thereis increasing interest in defining the possible role of Ca2' as asecond messenger in plants.Ca+ affects several physiological processes in plants. For ex-
ample, it has been implicated in geotropism (6, 17), cell elonga-tion (21), protoplasmic streaming (35), senescence (25), abscis-sion (24), a-amylase secretion (13), and several other processes.Ca'2 has been shown to regulate various biochemical processesafter binding to calmodulin and related proteins (7). Occurrenceofcalmodulin in plants has been well documented (34) and Ca2+-and calmodulin-mediated ativation of NAD+ kinase (1) Ca2'ATPase (9), and isofloridoside phosphate synthase (14) has beenreported from plants. Studies with calmodulin inhibitors suggest
' Scientific Paper No. SP 6858. College of Agriculture and HomeEconomics, Washington State University, Pullman. Project 0321. Sup-ported in part by National Science Foundation Grant PCM-8208408.
MATERIAIS AND METHODS
Plant Material and Chemicals. Corn seeds (Zea mays var.Patriot) were bought from Rodgers Bros. Seed Company, IdahoFalls, ID. [y-32P]ATP was obtained from New England Nuclear,Boston, MA. Acrylamide, N,N'-methylene-bis-acrylamide, molwt calibration kit, and other chemicals for polyacrylamide gelelectrophoresis were purchased from Bio-Rad Laboratories,Richmond, CA. Bovine brain calmodulin, cAMP, chlorproma-zine HCI, ATP, DTT, disodium EDTA, EGTA,2 and BSA werebought from Sigma. Trifluoperazine HCI was received as a giftfrom Smith, Kline and French, Philadelphia.
Preparation of Soluble Proteins from Corn Coleoptiles. Cornseeds were germinated on vermiculite in dark. Coleoptiles wereharvested after 6 d and homogenized with mortar and pestle withan equal volume of medium 1 containing 50 mm Mes-NaOH(pH 7.0), 5 mm MgCI2, 0.5 mm DTT, 0.25 M sucrose, and 0.2mM EGTA. All operations were performed at 4°C. After centrif-ugation for 10 min at 7,000g, the supernatant was again centri-fuged for 20 min at 130,000g. The supernatant was dialyzedagainst medium 2 (50 mm Mes-NaOH, pH 7.0, 5 mM MgC12,0.1 mm DTT, and 0.2 mm EGTA), centrifuged for 20 min at27,000g and concentrated 5-fold in an Amicon stirred cell witha PM-10 filter. The concentrated extract was centrifuged for 20min at 27,000g and used as a source of soluble proteins. Protein
2Abbreviations: EGTA, ethyleneglycol-bis-(f-aminoethyl ether)-N,N,N,'N'-tetraacetic acid.
359
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021

VELUTHAMBI AND POOVAIAH
content ofthe extracts was determined by Coomassie dye bindingassay (4).Prepation of Membrne Frction. Corn coleoptiles were
homogenized in medium 3 (same as medium 1 except that 5 mM
.' ; ;I ;
+CoP -C +o ..;;
.-Ca +Ca -Ca +Ca -al +(-r BLOC,. +cu
-.'
_3 L
-ab'.
FIG. 1. Phosphorylation ofmembrane proteins. Phosphorylation wasperformed in the presence of 0.2 mm EGTA (-Ca2") or in the presenceof 0.2 mm EGTA + 1 mM CaC12 (+Ca2+). Calmodulin (CM) andchlorpromazine (CP) were added to the reaction mixture of requiredtreatments at 1.6 juM and 200 pM, respectively. The calculated mol wt ofrepresentative phosphorylated polypeptides are indicated.
71K
62K
(1 ~~~~~~35K
Ij~~~Ii~~22KIt~~~~~~Ca+CM
:~~*1IA *~Caf +i~J Ca+CM+CP
._... Kiev...........
FIG. 2. Results ofdensitometer scanning of lanes 1, 2, 6, and 8 oftheautoradiograph in figure 1. Mol wt of representative phosphorylatedpolypeptides are indicated.
FIG. 3. Effect of Ca2" and calmodulin on the phosphorylation ofsoluble polypeptides. Protein phosphorylation was performed in thepresence of 0.2 mm EGTA (-Ca2l) or in the presence of 0.2 mm EGTA+ 1 mM CaCl2 (+Ca2+). Calmodulin (CM) was added at 1.6 jM. Thecalculated mol wt of representative phosphorylated polypeptides areindicated.
EDTA was added instead of 0.2 mM EGTA), homogenate cen-trifuged for 10 min at 7,000g, and the supernatant was centri-fuged at 1 30,000g for 20 min. The pellet was then suspended inthe same medium and centrifuged at 1 30,000g, and washing wasrepeated one more time. The pellet was washed twice more inmedium 1. The final pellet was suspended in medium 1 (1 ml/5g initial fresh weight of coleoptiles) and homogenized in a gasshomogenizer. This membrane preparation was used as a sourceof membrane proteins.
Phosphorylation of Proteins. Phosphorylation of soluble pro-teins was performed according to the method of Schulman andGreengard (31). The reaction mixture for assaying protein phos-phorylation (final volume, 100 jd) contained 0.4 mg protein, 50mM Mes-NaOH (pH 7.0), 5 mM MgCl2, 0.1 mm DTT, 0.2 mMEGTA (for -Ca2? reaction), 0.2 mM EGTA + 0.5 mM CaCl2 (for+Ca2+ reaction), and about 0.25 nmol of ['y-32P]ATP (20-40 Ci/mmol). Changes in Ca2+ concentration made for individualexperiments are indicated in Figure legends. Phosphorylation ofmembrane proteins was performed under similar conditionsexcept that the assay buffer was slightly modified following themethod ofSalimath and Marme (30). The reaction mixture (finalvolume 100 gd) contained about 0.4 mg protein, 50 mM Mes-NaOH (pH 7.0), 5 mM MgCl2, 0.5 mM DTT, and 0.25 M sucrose.Triton X-100 (0.1% w/v) was added to the reaction mixture 15min before starting the assay. To perform the phosphorylation,the reaction mixture was preincubated for 1 min at 30°C andphosphorylation was intitiated by adding [_-32P]ATP. Incubationwas carried out for 1 min at 30°C and the reaction was terminated
360 -Plant Physiol. Vol. 76, 1984
... .. ,.e ? .,.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021
a2+- AND CALMODULIN-REGULATED PROTEIN PHOSPHORYLATION
C P - .200 50 200 500
-Ca +Ca -Ca +Ca +Ca +Co
<- TFP o200 50 200 500-Co +Ca +Ca +Ca MW 3
A B C D E F MW.10 3
-86
-28
~~~~~~~~~~~~~~~~~~~~~~..:. ..
.........:00 * i g . -21:~~~~~~~~~ A -i6
-14
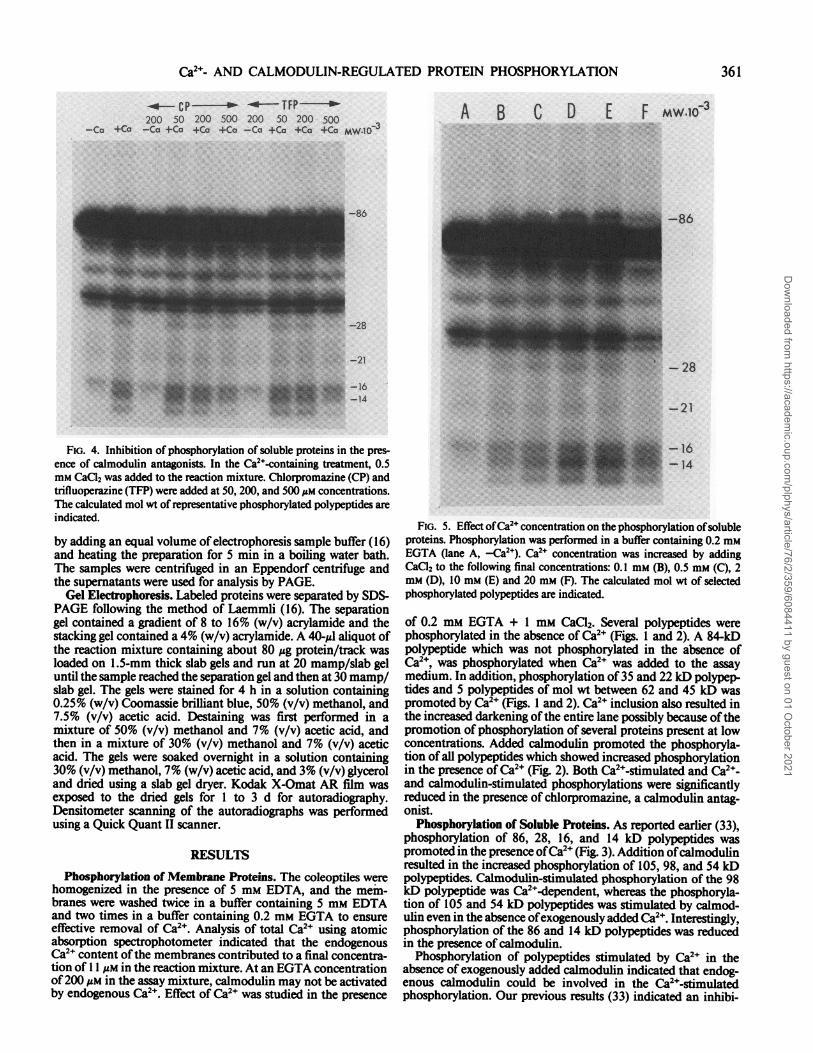
FIG. 4. Inhibition of phosphorylation of soluble proteins in the pres-ence of calmodulin antagonists. In the Ca2l-containing treatment, 0.5mM CaC12 was added to the reaction mixture. Chlorpromazine (CP) andtrifluoperazine (TF1P) were added at 50, 200, and 500 ;tM concentrations.The calculated mol wt of representative phosphorylated polypeptides areindicated.
by adding an equal volume of electrophoresis sample buffer (16)and heating the preparation for 5 min in a boiling water bath.The samples were centrifuged in an Eppendorf centrifuge andthe supernatants were used for analysis by PAGE.Gel Electrophoresis. Labeled proteins were separated by SDS-
PAGE following the method of Laemmli (16). The separationgel contained a gradient of 8 to 16% (w/v) acrylamide and thestacking gel contained a 4% (w/v) acrylamide. A 40-,g aliquot ofthe reaction mixture containing about 80 gg protein/track wasloaded on 1.5-mm thick slab gels and run at 20 mamp/slab geluntil the sample reached the separation gel and then at 30 mamp/slab gel. The gels were stained for 4 h in a solution containing0.25% (w/v) Coomassie brilliant blue, 50% (v/v) methanol, and7.5% (v/v) acetic acid. Destaining was first performed in amixture of 50% (v/v) methanol and 7% (v/v) acetic acid, andthen in a mixture of 30% (v/v) methanol and 7% (v/v) aceticacid. The gels were soaked overnight in a solution containing30% (v/v) methanol, 7% (w/v) acetic acid, and 3% (v/v) glyceroland dried using a slab gel dryer. Kodak X-Omat AR film wasexposed to the dried gels for 1 to 3 d for autoradiography.Densitometer scanning of the autoradiographs was performedusing a Quick Quant II scanner.
RESULTSPhosphorylation of Membrane Proteins. The coleoptiles were
homogenized in the presence of 5 mM EDTA, and the mem-branes were washed twice in a buffer containing 5 mm EDTAand two times in a buffer containing 0.2 mM EGTA to ensureeffective removal of Ca2l. Analysis of total Ca2" using atomicabsorption spectrophotometer indicated that the endogenousCa2" content ofthe membranes contributed to a final concentra-tion of 1 1 Mm in the reaction mixture. At an EGTA concentrationof200 Mm in the assay mixture, calmodulin may not be activatedby endogenous Ca2". Effect of Ca2l was studied in the presence
-86
'A -~~~~~28
-21
ElFS:_l ~~-16N;-N: ~~~14
FIG. 5. Effect ofCa2l concentration on the phosphorylation ofsolubleproteins. Phosphorylation was performed in a buffer containing 0.2 mMEGTA (lane A, -Ca2"). Ca2l concentration was increased by addingCaCl2 to the following final concentrations: 0.1 mM (B), 0.5 mm (C), 2mM (D), 10 mm (E) and 20 mm (F). The calculated mol wt of selectedphosphorylated polypeptides are indicated.
of 0.2 mm EGTA + 1 mm CaCl2. Several polypeptides werephosphorylated in the absence of Ca2' (Figs. 1 and 2). A 84-kDpolypeptide which was not phosphorylated in the absence ofCal+, was phosphorylated when Ca2' was added to the assaymedium. In addition, phosphorylation of 35 and 22 kD polypep-tides and 5 polypeptides of mol wt between 62 and 45 kD waspromoted by Ca2` (Figs. 1 and 2). Ca2' inclusion also resulted inthe increased darkening ofthe entire lane possibly because ofthepromotion of phosphorylation of several proteins present at lowconcentrations. Added calmodulin promoted the phosphoryla-tion of all polypeptides which showed increased phosphorylationin the presence of Ca2' (Fig. 2). Both Ca2'-stimulated and Ca2t-and calmodulin-stimulated phosphorylations were significantlyreduced in the presence of chlorpromazine, a calmodulin antag-onist.
Phosphorylation of Soluble Proteins. As reported earlier (33),phosphorylation of 86, 28, 16, and 14 kD polypeptides waspromoted in the presence ofCa2` (Fig. 3). Addition ofcalmodulinresulted in the increased phosphorylation of 105, 98, and 54 kDpolypeptides. Calmodulin-stimulated phosphorylation of the 98kD polypeptide was Ca24-dependent, whereas the phosphoryla-tion of 105 and 54 kD polypeptides was stimulated by calmod-ulin even in the absence ofexogenously added Ca2'. Interestingly,phosphorylation of the 86 and 14 kD polypeptides was reducedin the presence of calmodulin.
Phosphorylation of polypeptides stimulated by Ca24 in theabsence of exogenously added calmodulin indicated that endog-enous calmodulin could be involved in the Ca24-stimulatedphosphorylation. Our previous results (33) indicated an inhibi-
361
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021

VELUTHAMBI AND POOVAIAH
A H !MW i r
FIG. 6. Effect of increasing concentrations of exogenous calmodulinon the phosphorylation of soluble proteins. Phosphorylation was per-formed in the presence of 0.2 mM EGTA (A, -Ca2") or in the presenceof 0.2 mM EGTA + I mM CaCI2 (B, C, D, E, and F, +Ca2+). Bovinebrain calmodulin was added at 0.16 Mm (C), 0.8 pM (D), 1.6 AM (E), and3.2 uM (F). The calculated mol wt of representative phosphorylatedpolypeptides are indicated.
tion of Ca2+-stimulated protein phosphorylation in presence ofchlorpromazine, a calmodulin antagonist. The role of endoge-nous calmodulin on Ca2-stimulated phosphorylation was inves-tigated further with the second antagonist, trifluoperazine. Bothchlorpromazine and trifluoperazine significantly inhibited Ca2+-stimulated phosphorylation at 500 Mm concentration (Fig. 4).Ca2+-independent phosphorylation was not inhibited by the cal-modulin antagonists.
Effect of Cal on Phosphorylation. The effect of increasingconcentrations of Cae on protein phosphorylation was studiedin the presence of 0.2 mm EGTA (Fig. 5). Ca2'-dependentphosphorylation was promoted as the Ca' concentration wasincreased. Maximum phosphorylation occurred in the presenceof2 mM Ca2l. Both Ca2'-dependent and Ca2l-independent phos-phorylations were inhibited at 20 mM Ca'+ concentration.
Effect of Canlodulin on Phosphorylation. Phosphorylation ofseveral polypeptides was promoted by Ca2l in the absence ofexogenously added calmodulin. However, phosphorylation ofthe 98 kD polypeptide was promoted by calmodulin in a Ca?+dependent manner (Fig. 3). This indicated that even if endoge-nous calmodulin was present in the extract it could be present ata limiting concentration. Therefore, the effect of increasing con-centrations ofexogenous calmodulin on protein phosphorylationwas investigated (Fig. 6). The 98-kD polypeptide was phospho-rylated only when the exogenous calmodulin concentrationreached 0.8 MM. Phosphorylation of 105 and 54 kD polypeptidesalso increased with increasing amounts of calmodulin. Stimula-tion of phosphorylation of 105 and 54 kD polypeptides bycalmodulin in the absence of added Ca2" raised the question of
FIG. 7. Dependence of Mg2e for protein phosphorylation. To removeMg2+, the soluble protein fraction was dialyzed in medium 2 containing0.2 mm EDTA. Phosphorylation of proteins was performed in medium2 containing 0.2 mM EDTA + 1 mM CaCl2. MgCI2 was not added to thereaction mixture of lane A. MgCI2 was added to the reaction mixtures tothe final concentrations of 0.5 mM (C), 1.0 mm (D), 5 mm (B, E), and 10mM (F). The calculated mol wt of representative phosphorylated poly-peptides are indicated.
whether the calmodulin preparation had contaminated proteinkinase(s) and substrates. Phosphorylation was, therefore, per-formed in the calmodulin preparation without the plant extract.Proteins were not phosphorylated in the absence or in the pres-ence of Ca2` (results not shown). Thus, it is unlikely that theCa2+-independent but calmodulin-dependent phosphorylation ofproteins from corn extract was due to any contaminating proteinin the calmodulin preparation.Mg2+ Requirement for Phosphorylation. Since Mg2+ is an
important cofactor for kinases, its requirement for protein phos-phorylation was studied. Both Ca2+-dependent and Ca2+-inde-pendent phosphorylations were strongly dependent upon Mg2+(Fig. 7). Phosphorylation was almost absent in the absence ofMg2+. Phosphorylation reached a high level at 5 mm and in-creased only slightly at 10 mM. The effect of Mg2+ on thephosphorylation of various polypeptides was similar.
Effect of cAMP on Phosphorylfion. Cyclic AMP-activatedprotein kinase phosphorylates a variety of enzymes in animalsystems (8). Occurrence of cAMP in plants has been demon-strated (5) but cAMP-dependent protein kinase activity couldnot be demonstrated in plants. Addition of 10 to 50 #M cAMPto corn extracts did not reveal the presence of any cAMP-dependent protein kinase activity (Fig. 8).
Evidence for Dephosphorylation. Phosphorylation of proteinsobserved in corn extracts reflect only net phosphorylation sinceboth protein phosphorylation and dephosphorylation could oc-
362 Plant Physiol. Vol. 76, 1984
...,IPI
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021

Ca2+- AND CALMODULIN-REGULATED PROTEIN PHOSPHORYLATION
Mo Mo-Ca +Ca MW10 3
-86
-28
-16
-14
FIG. 8. Phosphorylation of soluble proteins in the presence ofcAMP.Phosphorylation was performed in the presence of0.2 mm EGTA. CyclicAMP was not added to the reaction mixture in lane A. Cyclic AMP wasadded to the other reaction mixtures to final concentrations of 10 jtM (B)and 50 jtM (C). Mol wt of standard marker proteins are indicated.
cur simultaneously. Consequently, if protein phosphatase activ-ity is inhibited by phosphatase inhibitors, it would result in anincrease in the net phosphorylation. Therefore, the effects oftwoknown phosphatase inhibitors, NaF and Na2MoO4 (14), wereinvestigated. Addition of Na2MoO4 at 10 mm concentration tothe phosphorylation assay mixture resulted in increased amountof phosphorylated polypeptides of 86, 28, 16, and 14 kD (Fig.9). NaF inhibited the phosphorylation of several polypeptides(data not shown) and was not used as a phosphatase inhibitor.More direct evidence for dephosphorylation was obtained as
follows: Phosphorylation was performed for 1 min in the pres-ence of 2.5 LLM ['y-32P]ATP. At the end of 1 min, unlabeled ATPwas added to 250 uM and the assay was continued for 2 or 4min. If dephosphorylation occurs, the labeled phosphate will beremoved from the proteins and rephosphorylation would resultin the addition of more unlabeled phosphate to the proteins.Therefore, the loss of label from the previously labeled proteinscan be taken as a measure of dephosphorylation activity (Fig.10). Net phosphorylation ofthe polypeptides apparently reacheda maximum within 1 min, since extended phosphorylation for 3and 5 min did not result in an increase in phosphorylation.Phosphorylation of 86, 28, 16, and 14 kD polypeptides wassignificantly reduced when the phosphorylation was performedfor 5 min, indicating the presence ofphosphatase activity. Whenphosphorylation was performed for 1 min in the presence of 2.5MM [y-32P]ATP, followed by phosphorylation in 250 ,uM unla-beled ATP for 2 or 4 min, radioactivity of the polypeptidesmarkedly decreased indicating a great extent of phosphataseactivity. Proteins phosphorylated in a Ca2--independent mannerwere also dephosphorylated.
FIG. 9. Promotion ofphosphorylation of soluble proteins in the pres-ence of sodium molybdate, a phosphatase inhibitor. Phosphorylation ofproteins was performed in the presence of 0.2 mM EGTA (-Ca2") or inthe presence of 0.2 mM EGTA + 0.5 mM CaC12 (+Ca2+). Sodiummolybdate (Mo) was added to a final concentration of 10 mm.
DISCUSSION
Ca2+ stimulated the phosphorylation of several polypeptidesof EGTA-washed membranes from corn coleoptiles (Fig. 1). It isnot clear whether it reflects Ca24-dependent or Ca2l- and cal-modulin-dependent protein kinase activity. In the latter case,calmodulin may have been trapped inside the membrane vesiclesduring homogenization and the complete removal ofcalmodulinmay not have been achieved, with EDTA or EGTA. Determi-nation ofcalmodulin in the Triton X-I00-solubilized membranesusing the phosphodiesterase method (20) revealed the presenceof about 49 ng calmodulin per mg protein. Calmodulin couldalso be present as a subunit of a protein kinase as in the case ofphosphorylase kinase (8). The presence of endogenous calmod-ulin in the membranes seems more likely since the Ca2'-pro-moted phosphorylation was reduced in the presence ofchlorpro-mazine (Fig. 1). Interestingly, similar observations were made inboth the plant membrane systems studied previously (12, 30).Added exogenous calmodulin promoted the Ca2'-stimulatedphosphorylation which was, in turn, inhibited by chlorproma-zine.
Phosphorylation ofseveral soluble polypeptides was stimulatedby Ca2' (Fig. 3). Here again, it is not known whether Ca2-dependent or Ca2+- and endogenous calmodulin-dependent pro-tein kinase was activated. Determination of calmodulin by thephosphodiesterase method (20) showed the presence of about0.85 ,g calmodulin/mg protein. Besides, the inhibition of Ca2+-stimulated phosphorylation by chlorpromazine and trifluopera-zine (Fig. 4) also indicated that endogenous calmodulin might
A B C 7-3MW.10 -Ca +Ca
-93-66
-45
-31
-22
-14
363
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021

VELUTHAMBI AND POOVAIAH
thW AOba B
.86- ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~..FIG. 10. Rapid dephosphorylation ofphosphorylated soluble proteins
in the presence of excess unlabeled ATP. Phosphorylation of proteinswas performed in the presence of 0.2 mM EGTA + 0.5 mM CaCI2.Phosphorylation was performed in the presence of 2.5 AM ['y-32P]ATPfor I min (A), 3 min (B), and 5 min (D). Unlabeled ATP was added toa final concentration of 250 uM at the end of I min (C and E) and thereaction stopped at 3 min (C) and at 5 min (E). The calculated mol wtof representative phosphorylated polypeptides are indicated.
be involved in the stimulation of phosphorylation by Ca'2. Lackof inhibition of Ca2+-independent phosphorylation in the pres-ence ofup to 500 MM chlorpromazine or trifluoperazine suggestedthat the inhibition of Ca2+-dependent phosphorylation may bespecific. However, as reported in other plants (12, 30) a ratherhigh concentration (S00Mm) of calmodulin antagonists was re-quired to inhibit the phosphorylation and their nonspecific ef-fects cannot be ruled out.Our attempts to separate calmodulin from the rest of the
soluble proteins by filtration through PM-30 filters (from AmiconCorporation, mol wt cut off-30,000) following repeated dilutions,did not remove calmodulin activity (unpublished results). Allthese results suggest a possibility that calmodulin could be asubunit of the protein kinase, as in the case of phosphorylasekinase (8). Addition of exogenous calmodulin promoted thephosphorylation of the 98-kD polypeptide in a Ca2'-dependentmanner (Fig. 3). Salimath and Marme (30) reported that cal-modulin concentrations required to stimulate phosphorylationmay vary depending upon the protein substrate. Accordingly,phosphorylation of the 98-kD polypeptide may require a highercalmodulin concentration, whereas the other polypeptides couldbe phosphorylated in the presence of a relatively lower concen-tration of endogenous calmodulin. In zucchini membranes, thephosphorylation of a 180-kD polypeptide was strongly reducedby Ca2+ (30). In many of our experiments, we found that theexogenously added calmodulin reduced the phosphorylation of
86 and 14 kD polypeptides (Fig. 3). The importance ofreductionof phosphorylation of proteins by calmodulin is not clear. Thesignificance ofthe calmodulin-stimulated phosphorylation ofthe105- and 54-kD polypeptides in the absence of added Ca2" isalso not clear. The Ca2+ content of the calmodulin preparationwas less than 2 ,M which is not sufficient to activate calmodulinin the presence of 200 AM EGTA.
Increased stimulation of phosphorylation ofpolypeptides withincreasing concentrations of Ca2+ (Fig. 5) strengthens the regu-latory significance of Ca2` in protein phosphorylation. Obligaterequirement ofMg2+ for protein phosphorylation and maximumphosphorylation at 5 to 10 mm Mg2+ concentration (Fig. 7)conforms to the results ofprevious reports (18, 19, 30). AlthoughcAMP has been shown to be present in plants (5), evidence couldnot be obtained for its role in promoting protein phosphorylation(11, 15, 18, 19, 30). Accordingly, cAMP did not stimulatephosphorylation of proteins from corn coleoptiles (Fig. 8).
Protein phosphorylation can function as an efficient regulatorymechanism only when it is reversible. The cell should havephosphoprotein phosphatases by which the phosphorylated pro-teins can be dephosphorylated and converted back to theirinactive forms. Indirect evidence has been obtained for thereversible phosphorylation and dephosphorylation of plant en-zymes such as pyruvate dehydrogenase (28), hydroxymethylglu-taryl CoA-reductase (29), and quinatet NAD+ oxidoreductase(27). A phosphoprotein phosphatase from soybean has beenpurified and characterized (18). Reduced phosphorylation ofseveral polypeptides during extended phosphorylation providedsuggestive evidence for the presence of phosphatase activity inpea shoot membranes (12) and zucchini hypocotyl membranes(30). Promotion of phosphorylation of proteins from corn co-leoptiles in the presence of sodium molybdate (Fig. 9), a phos-phatase inhibitor, suggests the presence of phosphatase activityin the extracts. Loss ofradioactivity from the labeled polypeptidesfollowing incubation in unlabeled ATP (Fig. 10), also providessupportive evidence for the existence ofdephosphorylating mech-anisms in corn coleoptiles.
In light of the recent reports on the effect of Ca2` and calmod-ulin on protein kinase activity (12,22,27, 30, 33), the importanceof protein phosphorylation and dephosphorylation in the regu-lation ofCa2'-dependent enzyme activities in plants is beginningto be understood. However, many questions remain to be an-swered. In all the plant systems studied, phosphorylation ofseveral polypeptides was stimulated by Ca2+ in the absence ofexogenously added calmodulin. These results suggest two possi-bilities: (a) Ca2' alone could stimulate protein kinase activity or(b) endogenous calmodulin remaining bound to the proteins ortrapped in membrane vesicles was involved in the activation. Inthe latter case, it is also possible that calmodulin could be asubunit of the protein kinase as in the case of phosphorylasekinase (8). Future research on protein phosphorylation in plantsshould help in answering some of these questions.
Acknowledgments.-We thank Dr. L. A. Hadwiger, Dr. G. Paliyath, and Dr. J.Bollinger for valuable discussions.
LITERATURE CITED
1. ANDERSON JM, MH CORMIER 1978 Calcium-dependent regulator of NADkinase in higher plants. Biochem Biophys Res Commun 84: 595-602
2. BENNEr J 1977 Phosphorylation of chloroplast membrane polypeptides.Nature 269: 344-346
3. BIRo RL, CC HALE II, OR WEIGAND, SJRoux 1982 Effects ofchlorpromazineon gravitropism in Avena cokoptils Ann Bot 50: 737-745
4. BRADFORD MM 1976 A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein dye-binding. Anal Biochem 72: 248-254
5. BROWN EG, RP NEwTON 1981 Cyclic AMP and higher plants. Phytochemistry20: 2453-2463
6. CHANDRA S, JF CHABar, GH MORRISON, AC LEOPOLD 1982 Localization ofcalcium in amyloplasts of root-cap cells using ion microscopy. Science 216:
364 Plant Physiol. Vol. 76, 1984
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021

Ca2+ AND CALMODULIN-REGULATED PROTEIN PHOSPHORYLATION
1221-12237. CHEUNG WY 1980 Calmodulin plays a pivotal role in cellular regulation.
Science 207: 19-278. COHEN P 1982 The role of protein phosphorylation in neural and hormonal
control of cellular activity. Nature 296: 613-6199. DIETER P, D MARME 1981 A calmodulin-dependent, microsomal ATPase from
corn (Zea mays L.). FEBS Lett 125:245-24810. ELLIOTF DC, SM BATCHELOR, RA CASSAR, NG MARINOS 1983 Calmodulin-
binding drugs affect responses to cytokinin, auxin and gibberellic acid. PlantPhysiol 72: 219-224
1 1. GOWDA S, DTN PILLAY 1980 Phosphorylation of ribosomal proteins by cyclicAMP-independent protein kinase (ribosomal casein kinase) from soybeancotyledons (Glycine max L.). Plant Cell Physiol 21: 1357-1365
12. HETHERINGTON A, A TREWAVAS 1982 Calcium-dependent protein kinase inpea shoot membrane. FEBS Lett 145: 67-71
13. JONEs RL, JV JACOBSEN 1983 Calcium regulation ofthe secretion ofa-amylaseisoenzymes and other proteins from barley aleurone layers. Planta 158: 1-9
14. KAuss H 1983 Volume regulation in Poterioochromonas. Involvement ofcalmodulin in the Ca2+-stimulated activation of isofloridoside phosphatesynthase. Plant Physiol 71; 169-172
15. KEATES RAB, AJ TREWAVAS 1974 Protein kinase activity associated withisolated ribosomes from peas and Lemna. Plant Physiol 54: 95-99
16. LAEMMLI UK 1970 Cleavage of structural proteins during the assembly of thehead ofbacteriophage T4. Nature 227: 680-685
17. LEE JS, TJ MULKEY, ML EVANS 1983 Reversible loss of gravitropic sensitivityin maize roots after tip application of calcium chelators Science 220: 1375-1376
18. LIN PPC, T MORI, JL KEY 1980 Phosphoprotein phosphatase of soybeanhypocotyls. Plant Physiol 66:368-374
19. MURRAY MG, JL KEY 1978 2,4-Dichlorophenoxyacetic acid-enhanced phos-phorylation of soybean nuclear proteins. Plant Physiol 61: 190-198
20. PALIYATH G, BW POOVAIAH 1984 Calmodulin inhibitor in senescing applesand its physiological and pharmacological significance. Proc Natl Acad SciUSA 81: 2065-2069
21. PICKARD BG 1970 Comparison of calcium and lanthanon ions in the Avena-coleoptile growth test. Planta 91: 314-320
22. POLYA GM, JR DAVIES 1982 Resolution of Ca2+-calmodulin-activated proteinkinase from wheat term. FEBS Lett 150: 167-171
23. POLYA GM, JR DAviEs 1983 Resolution and properties of a protein kinasecatalyzing the phosphorylation of a wheat germ cytokinin-binding protein.Plant Physiol 71: 482-488
24. POOVAIAH BW, AC LEOPOLD 1973 Inhibition of abscission by calcium. PlantPhysiol 51: 848-851
25. POOVAIAH BW, AC LEOPOLD 1973 Deferal of leaf senescence with calcium.Plant Physiol 52: 236-239
26. RAGHOTHAMA KG, Y MIZRAHI, BW POOVAIAH 1983 Effect of calmodulinantagonists on auxin-induced coleoptile elongation. Plant Physiol 72: S-824
27. RANJEVA R, G REFENO, AM BOUDET, D MARME 1983 Activation of plantquinate: NAD' 3-oxidoreductase by Ca2l and calmodulin. Proc Natl AcadSci USA 80: 5222-5224
28. RAo KP, DD RANDALL 1980 Plant pyruvate dehydrogenase complex: Inacti-vation and reactivation by phosphorylation and dephosphorylation. ArchBiochem Biophys 200: 461 466
29. RUSSELL DW,WD Dix 1981 In vitro inactivation and activation ofpea seedlingmicrosomal HMG-CoA reductase. Plant Physiol 67: S-854
30. SALIMATH BP, D MARME 1983 Protein phosphorylation and its regulation bycalcium and calmodulin in membrane fractions from zucchini hypocotyls.Planta 158: 560-568
31. SCHULMAN H, P GREENGARD 1978 Stimulation of brain membrane proteinphosphorylation by calcium and an endogenous heat-stable protein. Nature271: 478-479
32. TREwAvAs A 1979 Nuclear phosphoproteins in germinating cereal embryosand their relationship to the control of mRNA synthesis and the onset ofcell division. In DL Laidman, RG Wyn Jones, eds, Recent Advances in theBiochemistry of Cereals. Academic Press, New York, pp 175-208
33. VELUTHAMBI K, BW POOVAIAH 1984 Calcium-promoted protein phosphory-lation in plants. Science 223: 167-169
34. WAl-FERsON DM, DB IVERSON, LJ VAN ELDIK 1980 Spinach calmodulin:Isolation, characerization and comparison with vertebrate calmodulins.Biochemistry 19: 5762-5768
35. WILLIAMsoN RE, CC ASHLEY 1982 Free Ca2' and cytoplasmic streaming inthe algae Chara. Nature 296: 647-651
365
Dow
nloaded from https://academ
ic.oup.com/plphys/article/76/2/359/6084411 by guest on 01 O
ctober 2021