calcification of vascular smooth muscle cell...
TRANSCRIPT

Calcification of Vascular Smooth Muscle Cell CulturesInhibition by Osteopontin
Takeo Wada, Marc D. McKee, Susie Steitz, Cecilia M. Giachelli
Abstract—Calcification of vascular tissue is a common complication in aging, atherosclerosis, diabetes, renal failure, aorticstenosis, and prosthetic valve replacement. Osteopontin is a noncollagenous adhesive protein routinely found at sites ofdystrophic calcification and synthesized at high levels by macrophages in calcified aortic valves and atheroscleroticplaques. In the present study, we have characterized the calcification of bovine aortic smooth muscle cell (BASMC)cultures in vitro and have studied the effects of exogenous osteopontin on mineral deposition. Induction of calcificationin BASMC cultures was alkaline phosphatase–dependent and was characterized by a multilayer cell morphology.Mineral deposition occurred in the basal matrix of multilayered areas as indicated by von Kossa staining, andtransmission electron microscopy and electron diffraction identified the mineral as apatite. Ultrastructural analysis of thecultures showed the presence of extracellular matrix vesicles, calcifying collagen fibrils, and nodular-type calcificationssimilar to those found in calcified heart valves and atherosclerotic plaques. Purified osteopontin (0.05 to 5mg/mL) dosedependently inhibited calcification of BASMC cultures, whereas vitronectin and fibronectin had no effect. In contrastto the inhibitory mechanism of levamisole on mineral deposition, osteopontin did not inhibit alkaline phosphataseactivity or reduce phosphorus levels in the culture medium. Addition of calcium to the cultures overcame the inhibitoryeffect of osteopontin on BASMC culture calcification and resulted in decreased levels of calcium in the culture mediumand increased levels in the cell layer. Moreover, using high-resolution, colloidal-gold immunocytochemistry,osteopontin was found intimately associated with growing apatite crystals. These data indicate that the effect ofosteopontin, although calcium-dependent, was not mediated by simple calcium chelation but most likely by directinteraction of osteopontin with crystal surfaces. These studies suggest that BASMCs can be used to model vascularcalcification in vitro and that soluble osteopontin released near sites of vascular calcification may represent an adaptivemechanism aimed at preventing vascular calcification.(Circ Res. 1999;84:166-178.)
Key Words: osteopontinn vascular calcificationn ultrastructuren smooth muscle celln vascular disease
Dystrophic calcification is defined simply as mineraliza-tion that occurs in tissues other than bones and teeth in
the absence of a systemic mineral imbalance.1 Dystrophiccalcification is a common response to injury in many softtissues and can have devastating consequences when it occursin certain soft tissues including heart valves and arteries. It isalso a common detrimental consequence of implantation ofbiomaterials, prostheses, and other medical devices. In theheart, valve calcification leads to mechanical dysfunction andultimately to valve failure. In fact, dystrophic mineralizationremains the leading cause of stenotic or regurgitant failure innative and bioprosthetic valves.2 In arteries, calcification ispositively correlated with heart disease and increased risk ofmyocardial infarction,3 increased ischemic episodes in pe-ripheral vascular disease,4 and increased risk of dissectionafter angioplasty.5 Importantly, the presence of calcificationin arteries has recently been found to be predictive for deathand myocardial infarction in high-risk, asymptomaticpatients.6,7
The mechanisms regulating dystrophic calcification are notknown. Recent morphological studies have shown that patho-logical calcification of blood vessels shares features withnormal mineralization of bone and cartilage tissue. Matrixvesicles, postulated nucleation sites for formation of apatitemineral in cartilage, and perhaps bone, have been detected incalcified atherosclerotic lesions.8,9 Bone morphogenetic pro-tein type 2, a potent osteogenic differentiation factor, also isexpressed in vascular calcified lesions.10 Additionally,mineral-binding, gamma-carboxy-glutamate (Gla)–contain-ing proteins such as osteocalcin and matrix Gla protein(MGP)1 have been localized to advanced atheroscleroticlesions.11–15 Furthermore, MGP-null mutant mice die withinthe first 2 months of age as a result of arterial rupture andheart failure due to extensive calcification of the large elasticand muscular arteries and the aortic valve.16
Aside from these molecules, we17–19and others13,20–22havereported that osteopontin is abundant at sites of calcification
Received June 29, 1998; accepted October 27, 1998.From Department of Pathology (T.W., S.S., C.M.G.), Faculty of Medicine, University of Washington, Seattle, Washington; Department of Stomatology
(M.D.M.), Faculty of Dentistry and MRC Group in Periodontal Physiology, University of Montreal, Montreal, Quebec, Canada.Correspondence to Cecilia M. Giachelli, PhD, Pathology Department Box 357335, University of Washington, 1959 NE Pacific Street, Seattle, WA
98195. E-mail [email protected]© 1999 American Heart Association, Inc.
Circulation Researchis available at http://www.circresaha.org
166
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

in human atherosclerotic plaques and in calcified aorticvalves but is not found in normal arteries. Osteopontin is anacidic phosphoprotein normally found in mineralized tissuessuch as bones and teeth, as well as in kidney and epitheliallinings of the body (see Reference 23 for review). Inatherosclerotic plaques and calcifying aortic valves, os-teopontin is highly localized to the surfaces of calcifieddeposits. The major source of osteopontin protein in bothlesions is infiltrating macrophages that are intimately associ-ated with the calcified deposits, although smooth muscle andendothelial cells also synthesize osteopontin but at muchlower levels.17–19 These data suggest that osteopontin mightbe an important regulator of vascular calcification.
Although a prominent component of the extracellularmatrix of bone, the function of osteopontin in hard tissueformation, mineralization, and turnover is not yet clear. Thepresence of osteopontin at calcification foci in the osteoid, itsassociation with bone surfaces, and its ability to facilitateadhesion of osteoblast-like cells in vitro have suggested thatosteopontin might function to promote bone formation.24 Onthe other hand, cell-free nucleation and crystal growth studiesin metastable calcium phosphate solutions have suggestedthat osteopontin inhibits apatite formation,25,26and interactionof osteopontin with osteoclastavb3 receptor has been shownto stimulate resorption of bone.27 To determine the potentialrole of osteopontin in vascular calcification, we have used anin vitro model wherein extracellular matrix produced bybovine aortic smooth muscle cells accumulates apatitic min-eral after addition of the organic phosphate donorb-glycerophosphate.28 In the present study, we show thatunder these conditions, apatite is deposited along and withincollagen fibrils in the presence of matrix vesicles, observa-tions previously reported for calcified vascular tissues invivo.8 Using this culture system, we have found that exog-enously added osteopontin associates with apatitic crystalsand is a potent inhibitor of vascular calcification by amechanism that does not involve alkaline phosphatase inhi-bition or calcium chelation/sequestration but likely involvesdirect inhibition of apatite growth by binding to crystalsurfaces. From these data, we hypothesize that osteopontinreleased by various cell types, including macrophages, at sitesof vascular calcification may represent an adaptive mecha-nism aimed at preventing deleterious, ectopic mineralization.
Materials and MethodsCell CultureBovine aortic smooth muscle cells (BASMCs) were obtained by amodification of the explant method originally described by Ross.29
Briefly, medial tissue was separated from segments of bovinethoracic aorta. Small pieces of tissue (1 to 2 mm3) were loosened bya 1-hour incubation in DMEM containing 4.5 g/L glucose supple-mented with 165 U/mL collagenase type I, 15 U/mL elastase type III,and 0.375 mg/mL soybean trypsin inhibitor at 37°C. Partiallydigested tissues were placed in 6-well plates and cultured for severalweeks in DMEM containing 4.5 g/L glucose supplemented with 20%FBS at 37°C in a humidified atmosphere containing 5% CO2. Cellsthat had migrated from the explants were collected and maintained ingrowth medium (DMEM containing 15% FBS and 10 mmol/Lsodium pyruvate supplemented with 100 U/mL of penicillin and 100mg/mL of streptomycin). To confirm that the cells isolated frombovine aortic wall were vascular smooth muscle cells,a-smooth
muscle actin, vimentin, and calponin levels were examined byimmunofluorescence as described below. All 3 antibodies stained.95% of the cells in a filamentous pattern (data not shown),indicating that the cells were of vascular smooth muscle origin. Forall experiments, cells were used between passages 2 and 5.
Adhesive Proteins and Neutralizing AntibodyRat osteopontin was purified from the conditioned medium of ratneonatal smooth muscle cell cultures as described previously.30 Thispreparation was judged to be.95% pure, on the basis of Coomassiestaining and N-terminal sequence analysis. Rat plasma vitronectin(Sigma Immunochemicals) and bovine fibronectin (TELIOS Phar-maceutical Inc) were resuspended in PBS to a concentration of 0.5mg/mL and stored frozen until use. Goat anti-rat osteopontinantibody OP-199 and nonimmune goat serum were prepared, andIgG fractions were purified as previously described.30
Histochemical AnalysisFor immunofluorescence microscopy, BASMCs were cultured on10-well heavy Teflon-coated microscope glass slides (Cel-LineAssociates Inc) for 24 hours, fixed with cold methanol, blocked withPBS containing 2% BSA and 10% normal rabbit serum, and treatedwith monoclonal anti–a-smooth muscle actin antibody (1A4, Sigma)and monoclonal anti-vimentin antibody (V9, Dako) diluted with PBScontaining 2% BSA 1:50 and 1:25, respectively. Monoclonal anti-calponin antibody (CALP, a gift of Maria Frid, University ofColorado, Denver),31 was used without dilution. As a secondaryantibody, FITC-conjugated rabbit anti-mouse IgG was used afterdilution with PBS 1:30. Mouse nonimmune IgG was used as acontrol for the primary antibody. Greater than 95% of the BASMCsstained with all 3 muscle markers, proving their smooth muscleorigin.
The expression of alkaline phosphatase was visualized by incu-bating citrate-acetone-formaldehyde-fixed cells at room temperaturefor 15 minutes with Naphthol AS-BI Alkaline Solution (Sigma).Mineral deposition was assessed by von Kossa staining (30 minutes,5% silver nitrate), as previously described.32
Induction of CalcificationCalcification of BASMC cultures was induced by the method ofShioi et al.28 Briefly, BASMCs were cultured in growth medium for4 days and then switched to calcification medium [calcificationmedium5DMEM (high glucose, 4.5 g/L) containing 15% FBS,10 mmol/L sodium pyruvate, 1027 mol/L insulin, 50mg/mL ascorbicacid, 100 U/mL penicillin, 100mg/mL streptomycin, andb-glycerophosphate (1210 mmol/L as indicated)] for 10 days. Themedium was replaced with fresh medium twice a week. In the timecourse experiments, the beginning day of culture in calcificationmedium was defined as day 0. For immunodepletion studies,medium containing 10mg/mL osteopontin was mixed with 20mg/mL anti-osteopontin (OP-199) and incubated for 1 hour at roomtemperature. Two hundred fifty milligrams of protein A–Sepharosewas added and incubated for 1 hour at room temperature. Theantibody-protein A–Sepharose complexes were removed by centrif-ugation, and the remaining supernatant diluted 20-fold for use incalcification studies.
Quantitation of Calcium DepositionCalcification was assessed by a modification of the method describedby Jono et al.33 Briefly, the cultures were decalcified with 0.6 N HClfor 24 hours. The calcium content of the HCl supernatant wasdetermined colorimetrically by the o-cresolphthalein complexonemethod (Calcium Kit, Sigma). After decalcification, the cultureswere washed with PBS and solubilized with 0.1 N NaOH/0.1% SDS.Total protein content was measured with a Bio-Rad protein assay kit(Bio-Rad). The calcium content of the cell layer was normalized toprotein content.
Wada et al February 5, 1999 167
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
Alkaline Phosphatase, Phosphorus, and CalciumIon AssaysFor cellular alkaline phosphatase activity measurements, cells werewashed 3 times with PBS, and cellular proteins were solubilized with1% Triton X-100 in 0.9% NaCl and centrifuged. Supernatants wereassayed for alkaline phosphatase activity on the basis of the methodof Bessey et al.34 One unit was defined as the activity producing 1nmol of p-nitrophenol within 1 minute. Protein concentrations weredetermined with a Bio-Rad protein assay kit (Bio-Rad). The datawere normalized to the protein content of the cell layer.
Phosphorus and calcium concentrations in the culture mediumwere measured by the phosphomolybdate complex method (Phos-phorus Kit, Sigma) and the o-cresolphthalein complexone method(Calcium Kit, Sigma), respectively.
Ultrastructure Analysis andMineral CharacterizationFor ultrastructural examination of BASMC cultures by transmissionelectron microscopy, cells grown on plastic were fixed overnight inan aldehyde solution containing 1% glutaraldehyde and 1% parafor-maldehyde buffered with 0.1 mol/L sodium cacodylate buffer at pH7.2. The cultures were then washed with 0.1 mol/L sodium cacody-late buffer alone, dehydrated in a graded series of ethanol solutions,and infiltrated and embedded in either Taab epoxy resin or LR Whiteacrylic resin (Marivac). The resins were polymerized for 2 days at55°C. Samples destined for epoxy embedding were also post-fixedwith potassium ferrocyanide–reduced 4% osmium tetroxide to pro-vide additional membrane contrast in the electron microscope. Formineral analyses by selected-area electron diffraction, other cultureswere treated nonaqueously by fixing only with 100% ethanol,followed by direct embedding in resin without further processing.One micrometer-thick survey sections were prepared from variousregions of the cultures and stained with Toluidine blue for exami-nation by light microscopy. Thin sections (80 to 100 nm) of selectedregions were then cut using a diamond knife on a Reichert UltracutE microtome and placed on 5% dichloroethane–coated nickel gridsevaporated with carbon. Grid-mounted sections were stained brieflywith ethanolic uranyl acetate and lead citrate and examined using aJEOL JEM 1200EX transmission electron microscope operating at60 kV. Anhydrously treated samples left unstained were used forselected-area electron diffraction using a 100mmol/L diffractionaperture and a camera length of 80 cm. Diffraction patterns wereanalyzed and compared with synthetic apatite standards and powderdiffraction files as previously reported for bone mineral.35
Osteopontin Immunogold LabelingBASMCs were cultured in calcification medium for 7 days to allowmineralization to begin. Purified rat osteopontin (0.5mg/mL) wasthen added until day 10. Cultures were preserved as described aboveusing aldehyde fixative followed by embedding in LR White acrylicresin for immunocytochemistry. Postembedding immunolabelingwas performed using osteopontin antibody (OP-199) and proteinA–colloidal gold complex as described previously.36 Briefly, thin(80-nm) sections of the cultures were placed on nickel grids andincubated for 5 minutes with 1% ovalbumin in PBS, followed byincubation with primary antibody for 1 hour, rinsing with PBS,blocking again with ovalbumin, and then exposure to protein A–goldcomplex for 30 minutes. After final rinsing with distilled water, gridswere air-dried and conventionally stained with uranyl acetate andlead citrate and viewed by transmission electron microscopy. Thespecificity of the OP-199 antibody has been shown previously byWestern blotting30 and by incubations performed in the present studywith use of preimmune serum and protein A–gold complex alone.
Statistical AnalysisUnpaired Studentt test was used to compare groups. A value ofP,0.05 was considered significant.
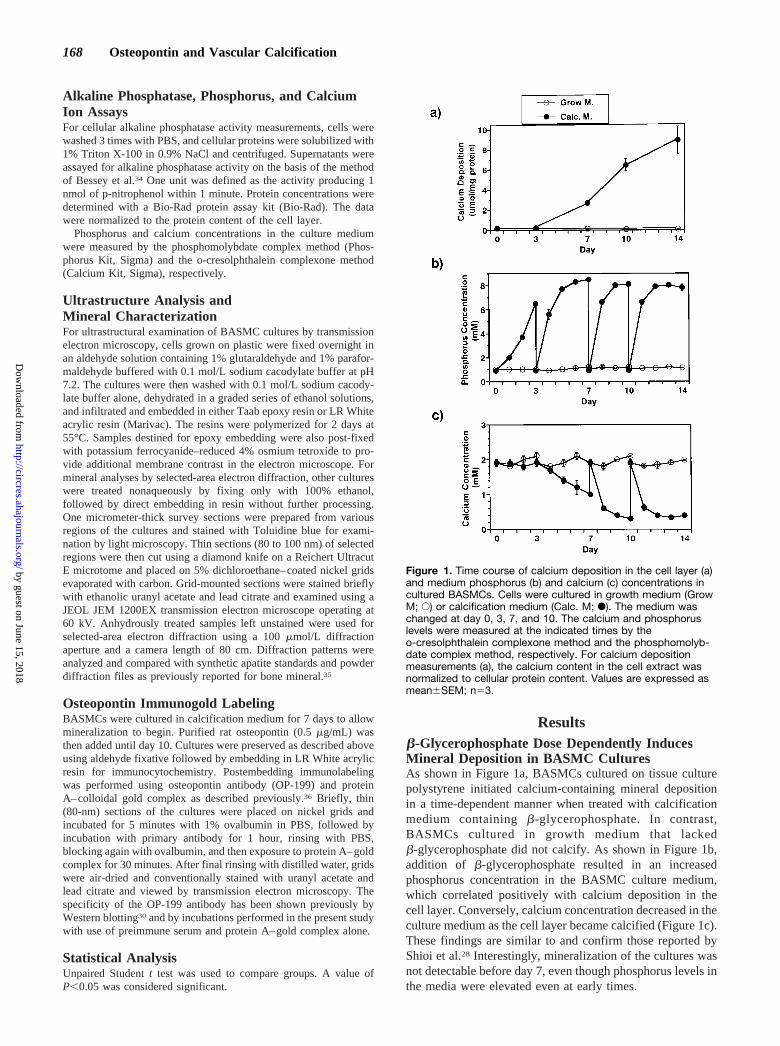
Resultsb-Glycerophosphate Dose Dependently InducesMineral Deposition in BASMC CulturesAs shown in Figure 1a, BASMCs cultured on tissue culturepolystyrene initiated calcium-containing mineral depositionin a time-dependent manner when treated with calcificationmedium containingb-glycerophosphate. In contrast,BASMCs cultured in growth medium that lackedb-glycerophosphate did not calcify. As shown in Figure 1b,addition of b-glycerophosphate resulted in an increasedphosphorus concentration in the BASMC culture medium,which correlated positively with calcium deposition in thecell layer. Conversely, calcium concentration decreased in theculture medium as the cell layer became calcified (Figure 1c).These findings are similar to and confirm those reported byShioi et al.28 Interestingly, mineralization of the cultures wasnot detectable before day 7, even though phosphorus levels inthe media were elevated even at early times.
Figure 1. Time course of calcium deposition in the cell layer (a)and medium phosphorus (b) and calcium (c) concentrations incultured BASMCs. Cells were cultured in growth medium (GrowM; E) or calcification medium (Calc. M; F). The medium waschanged at day 0, 3, 7, and 10. The calcium and phosphoruslevels were measured at the indicated times by theo-cresolphthalein complexone method and the phosphomolyb-date complex method, respectively. For calcium depositionmeasurements (a), the calcium content in the cell extract wasnormalized to cellular protein content. Values are expressed asmean6SEM; n53.
168 Osteopontin and Vascular Calcification
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
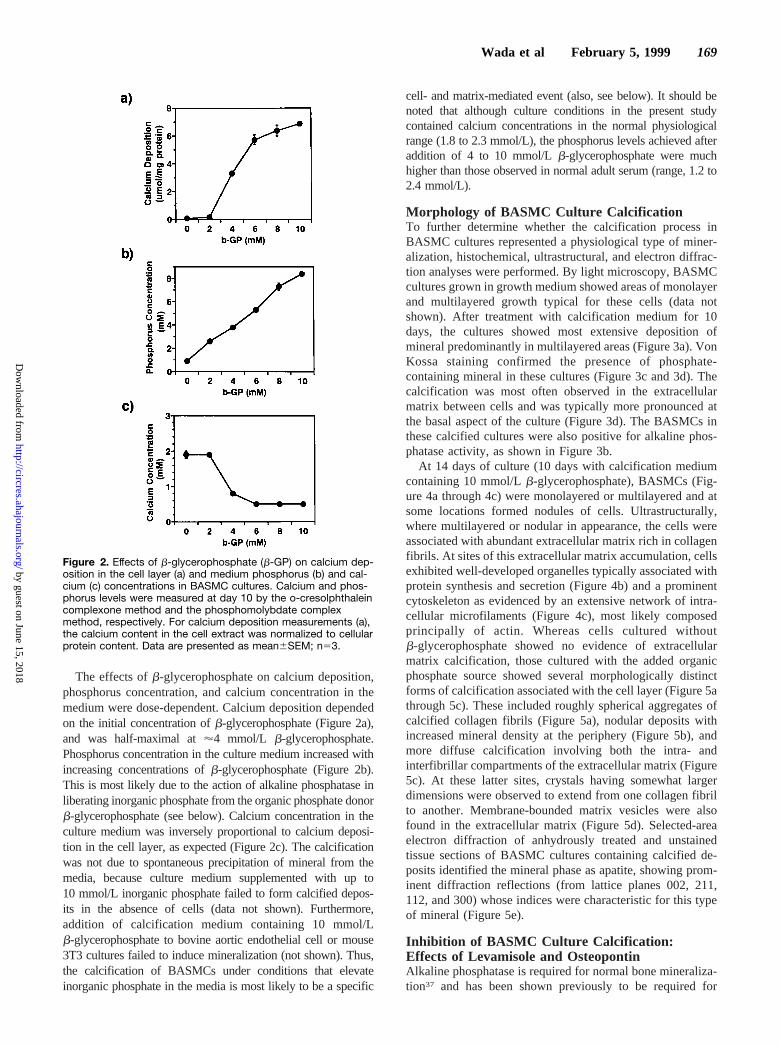
The effects ofb-glycerophosphate on calcium deposition,phosphorus concentration, and calcium concentration in themedium were dose-dependent. Calcium deposition dependedon the initial concentration ofb-glycerophosphate (Figure 2a),and was half-maximal at'4 mmol/L b-glycerophosphate.Phosphorus concentration in the culture medium increased withincreasing concentrations ofb-glycerophosphate (Figure 2b).This is most likely due to the action of alkaline phosphatase inliberating inorganic phosphate from the organic phosphate donorb-glycerophosphate (see below). Calcium concentration in theculture medium was inversely proportional to calcium deposi-tion in the cell layer, as expected (Figure 2c). The calcificationwas not due to spontaneous precipitation of mineral from themedia, because culture medium supplemented with up to10 mmol/L inorganic phosphate failed to form calcified depos-its in the absence of cells (data not shown). Furthermore,addition of calcification medium containing 10 mmol/Lb-glycerophosphate to bovine aortic endothelial cell or mouse3T3 cultures failed to induce mineralization (not shown). Thus,the calcification of BASMCs under conditions that elevateinorganic phosphate in the media is most likely to be a specific
cell- and matrix-mediated event (also, see below). It should benoted that although culture conditions in the present studycontained calcium concentrations in the normal physiologicalrange (1.8 to 2.3 mmol/L), the phosphorus levels achieved afteraddition of 4 to 10 mmol/Lb-glycerophosphate were muchhigher than those observed in normal adult serum (range, 1.2 to2.4 mmol/L).
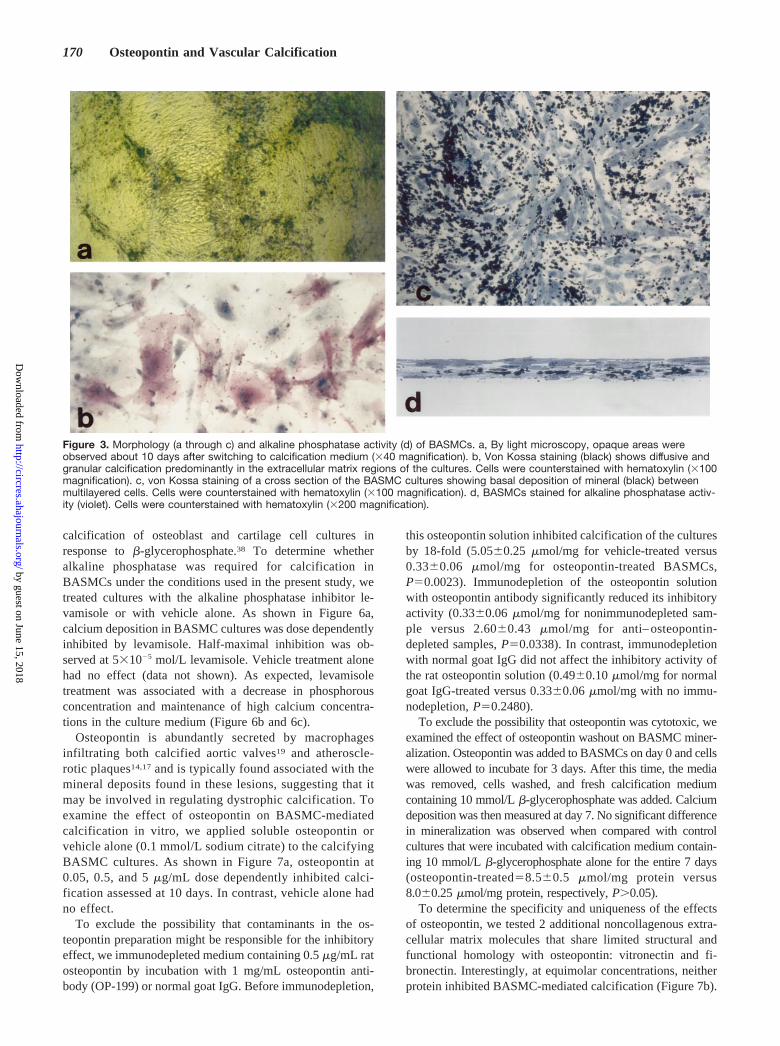
Morphology of BASMC Culture CalcificationTo further determine whether the calcification process inBASMC cultures represented a physiological type of miner-alization, histochemical, ultrastructural, and electron diffrac-tion analyses were performed. By light microscopy, BASMCcultures grown in growth medium showed areas of monolayerand multilayered growth typical for these cells (data notshown). After treatment with calcification medium for 10days, the cultures showed most extensive deposition ofmineral predominantly in multilayered areas (Figure 3a). VonKossa staining confirmed the presence of phosphate-containing mineral in these cultures (Figure 3c and 3d). Thecalcification was most often observed in the extracellularmatrix between cells and was typically more pronounced atthe basal aspect of the culture (Figure 3d). The BASMCs inthese calcified cultures were also positive for alkaline phos-phatase activity, as shown in Figure 3b.
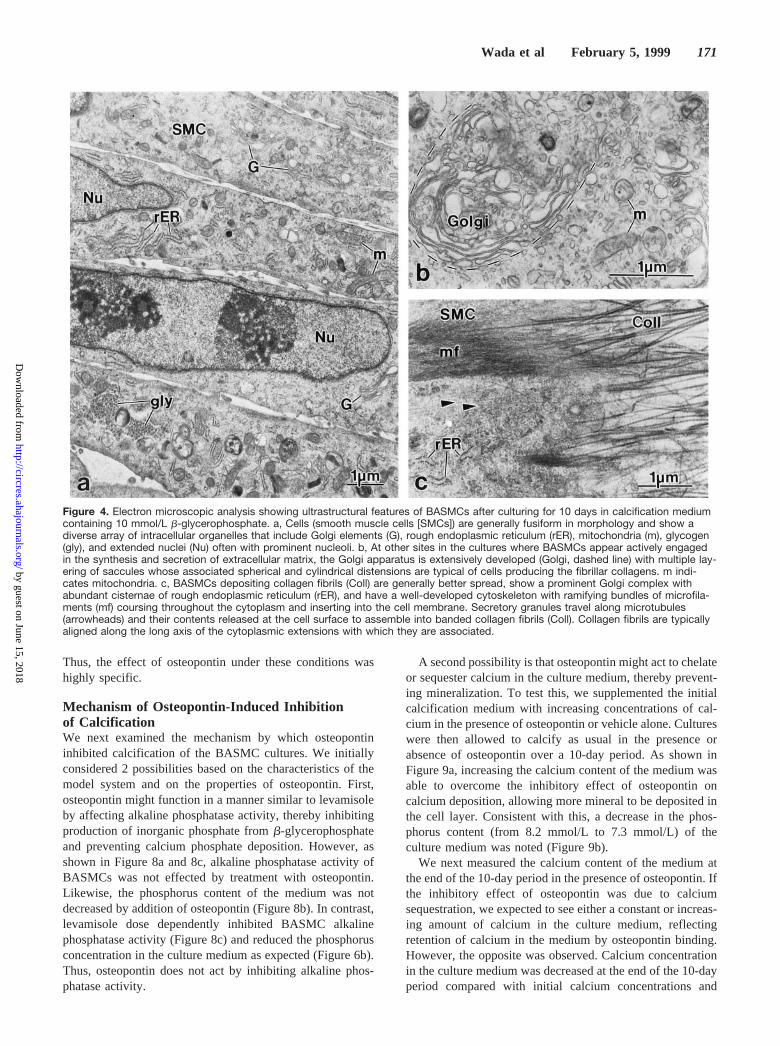
At 14 days of culture (10 days with calcification mediumcontaining 10 mmol/Lb-glycerophosphate), BASMCs (Fig-ure 4a through 4c) were monolayered or multilayered and atsome locations formed nodules of cells. Ultrastructurally,where multilayered or nodular in appearance, the cells wereassociated with abundant extracellular matrix rich in collagenfibrils. At sites of this extracellular matrix accumulation, cellsexhibited well-developed organelles typically associated withprotein synthesis and secretion (Figure 4b) and a prominentcytoskeleton as evidenced by an extensive network of intra-cellular microfilaments (Figure 4c), most likely composedprincipally of actin. Whereas cells cultured withoutb-glycerophosphate showed no evidence of extracellularmatrix calcification, those cultured with the added organicphosphate source showed several morphologically distinctforms of calcification associated with the cell layer (Figure 5athrough 5c). These included roughly spherical aggregates ofcalcified collagen fibrils (Figure 5a), nodular deposits withincreased mineral density at the periphery (Figure 5b), andmore diffuse calcification involving both the intra- andinterfibrillar compartments of the extracellular matrix (Figure5c). At these latter sites, crystals having somewhat largerdimensions were observed to extend from one collagen fibrilto another. Membrane-bounded matrix vesicles were alsofound in the extracellular matrix (Figure 5d). Selected-areaelectron diffraction of anhydrously treated and unstainedtissue sections of BASMC cultures containing calcified de-posits identified the mineral phase as apatite, showing prom-inent diffraction reflections (from lattice planes 002, 211,112, and 300) whose indices were characteristic for this typeof mineral (Figure 5e).
Inhibition of BASMC Culture Calcification:Effects of Levamisole and OsteopontinAlkaline phosphatase is required for normal bone mineraliza-tion37 and has been shown previously to be required for
Figure 2. Effects of b-glycerophosphate (b-GP) on calcium dep-osition in the cell layer (a) and medium phosphorus (b) and cal-cium (c) concentrations in BASMC cultures. Calcium and phos-phorus levels were measured at day 10 by the o-cresolphthaleincomplexone method and the phosphomolybdate complexmethod, respectively. For calcium deposition measurements (a),the calcium content in the cell extract was normalized to cellularprotein content. Data are presented as mean6SEM; n53.
Wada et al February 5, 1999 169
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
calcification of osteoblast and cartilage cell cultures inresponse tob-glycerophosphate.38 To determine whetheralkaline phosphatase was required for calcification inBASMCs under the conditions used in the present study, wetreated cultures with the alkaline phosphatase inhibitor le-vamisole or with vehicle alone. As shown in Figure 6a,calcium deposition in BASMC cultures was dose dependentlyinhibited by levamisole. Half-maximal inhibition was ob-served at 531025 mol/L levamisole. Vehicle treatment alonehad no effect (data not shown). As expected, levamisoletreatment was associated with a decrease in phosphorousconcentration and maintenance of high calcium concentra-tions in the culture medium (Figure 6b and 6c).
Osteopontin is abundantly secreted by macrophagesinfiltrating both calcified aortic valves19 and atheroscle-rotic plaques14,17and is typically found associated with themineral deposits found in these lesions, suggesting that itmay be involved in regulating dystrophic calcification. Toexamine the effect of osteopontin on BASMC-mediatedcalcification in vitro, we applied soluble osteopontin orvehicle alone (0.1 mmol/L sodium citrate) to the calcifyingBASMC cultures. As shown in Figure 7a, osteopontin at0.05, 0.5, and 5mg/mL dose dependently inhibited calci-fication assessed at 10 days. In contrast, vehicle alone hadno effect.
To exclude the possibility that contaminants in the os-teopontin preparation might be responsible for the inhibitoryeffect, we immunodepleted medium containing 0.5mg/mL ratosteopontin by incubation with 1 mg/mL osteopontin anti-body (OP-199) or normal goat IgG. Before immunodepletion,
this osteopontin solution inhibited calcification of the culturesby 18-fold (5.0560.25mmol/mg for vehicle-treated versus0.3360.06 mmol/mg for osteopontin-treated BASMCs,P50.0023). Immunodepletion of the osteopontin solutionwith osteopontin antibody significantly reduced its inhibitoryactivity (0.3360.06mmol/mg for nonimmunodepleted sam-ple versus 2.6060.43 mmol/mg for anti– osteopontin-depleted samples,P50.0338). In contrast, immunodepletionwith normal goat IgG did not affect the inhibitory activity ofthe rat osteopontin solution (0.4960.10mmol/mg for normalgoat IgG-treated versus 0.3360.06mmol/mg with no immu-nodepletion,P50.2480).
To exclude the possibility that osteopontin was cytotoxic, weexamined the effect of osteopontin washout on BASMC miner-alization. Osteopontin was added to BASMCs on day 0 and cellswere allowed to incubate for 3 days. After this time, the mediawas removed, cells washed, and fresh calcification mediumcontaining 10 mmol/Lb-glycerophosphate was added. Calciumdeposition was then measured at day 7. No significant differencein mineralization was observed when compared with controlcultures that were incubated with calcification medium contain-ing 10 mmol/Lb-glycerophosphate alone for the entire 7 days(osteopontin-treated58.560.5 mmol/mg protein versus8.060.25mmol/mg protein, respectively,P.0.05).
To determine the specificity and uniqueness of the effectsof osteopontin, we tested 2 additional noncollagenous extra-cellular matrix molecules that share limited structural andfunctional homology with osteopontin: vitronectin and fi-bronectin. Interestingly, at equimolar concentrations, neitherprotein inhibited BASMC-mediated calcification (Figure 7b).
Figure 3. Morphology (a through c) and alkaline phosphatase activity (d) of BASMCs. a, By light microscopy, opaque areas wereobserved about 10 days after switching to calcification medium (340 magnification). b, Von Kossa staining (black) shows diffusive andgranular calcification predominantly in the extracellular matrix regions of the cultures. Cells were counterstained with hematoxylin (3100magnification). c, von Kossa staining of a cross section of the BASMC cultures showing basal deposition of mineral (black) betweenmultilayered cells. Cells were counterstained with hematoxylin (3100 magnification). d, BASMCs stained for alkaline phosphatase activ-ity (violet). Cells were counterstained with hematoxylin (3200 magnification).
170 Osteopontin and Vascular Calcification
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
Thus, the effect of osteopontin under these conditions washighly specific.
Mechanism of Osteopontin-Induced Inhibitionof CalcificationWe next examined the mechanism by which osteopontininhibited calcification of the BASMC cultures. We initiallyconsidered 2 possibilities based on the characteristics of themodel system and on the properties of osteopontin. First,osteopontin might function in a manner similar to levamisoleby affecting alkaline phosphatase activity, thereby inhibitingproduction of inorganic phosphate fromb-glycerophosphateand preventing calcium phosphate deposition. However, asshown in Figure 8a and 8c, alkaline phosphatase activity ofBASMCs was not effected by treatment with osteopontin.Likewise, the phosphorus content of the medium was notdecreased by addition of osteopontin (Figure 8b). In contrast,levamisole dose dependently inhibited BASMC alkalinephosphatase activity (Figure 8c) and reduced the phosphorusconcentration in the culture medium as expected (Figure 6b).Thus, osteopontin does not act by inhibiting alkaline phos-phatase activity.
A second possibility is that osteopontin might act to chelateor sequester calcium in the culture medium, thereby prevent-ing mineralization. To test this, we supplemented the initialcalcification medium with increasing concentrations of cal-cium in the presence of osteopontin or vehicle alone. Cultureswere then allowed to calcify as usual in the presence orabsence of osteopontin over a 10-day period. As shown inFigure 9a, increasing the calcium content of the medium wasable to overcome the inhibitory effect of osteopontin oncalcium deposition, allowing more mineral to be deposited inthe cell layer. Consistent with this, a decrease in the phos-phorus content (from 8.2 mmol/L to 7.3 mmol/L) of theculture medium was noted (Figure 9b).
We next measured the calcium content of the medium atthe end of the 10-day period in the presence of osteopontin. Ifthe inhibitory effect of osteopontin was due to calciumsequestration, we expected to see either a constant or increas-ing amount of calcium in the culture medium, reflectingretention of calcium in the medium by osteopontin binding.However, the opposite was observed. Calcium concentrationin the culture medium was decreased at the end of the 10-dayperiod compared with initial calcium concentrations and
Figure 4. Electron microscopic analysis showing ultrastructural features of BASMCs after culturing for 10 days in calcification mediumcontaining 10 mmol/L b-glycerophosphate. a, Cells (smooth muscle cells [SMCs]) are generally fusiform in morphology and show adiverse array of intracellular organelles that include Golgi elements (G), rough endoplasmic reticulum (rER), mitochondria (m), glycogen(gly), and extended nuclei (Nu) often with prominent nucleoli. b, At other sites in the cultures where BASMCs appear actively engagedin the synthesis and secretion of extracellular matrix, the Golgi apparatus is extensively developed (Golgi, dashed line) with multiple lay-ering of saccules whose associated spherical and cylindrical distensions are typical of cells producing the fibrillar collagens. m indi-cates mitochondria. c, BASMCs depositing collagen fibrils (Coll) are generally better spread, show a prominent Golgi complex withabundant cisternae of rough endoplasmic reticulum (rER), and have a well-developed cytoskeleton with ramifying bundles of microfila-ments (mf) coursing throughout the cytoplasm and inserting into the cell membrane. Secretory granules travel along microtubules(arrowheads) and their contents released at the cell surface to assemble into banded collagen fibrils (Coll). Collagen fibrils are typicallyaligned along the long axis of the cytoplasmic extensions with which they are associated.
Wada et al February 5, 1999 171
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

Figure 5. Electron micrographs selected to show various calcification (Cal) events associated with the collagenous (Coll) extracellularmatrix produced by BASMCs (SMCs) after 10 days in calcification medium. a, Electron-dense calcified deposits frequently appear asdiscrete, micrometer-sized masses within the collagen-rich extracellular matrix surrounding the cells. Closer examination of the calcifi-cation pattern reveals that at the margins of these sites, a finely textured mineral commonly aligns with fibrillar collagen and progressesalong individual collagen fibrils (arrowheads, inset). Additional mineral deposition appears to occur within the interior of these calcifica-tion foci, extending into the interfibrillar volume as well, thus rendering them more electron dense and homogeneous. Nu indicatesnucleus; m, mitochondria. b, Another calcification pattern typically seen in the extracellular matrix of the BASMC cultures consists ofsmall nodular accretions of mineral (asterisks) with an increased density at their periphery (open arrows). c, Frequently observed at themargins of calcification sites are relatively large crystals (small arrows) spanning the interfibrillar compartment and passing from onecollagen fibril to another. This mineral texture differs from that seen in panel a (inset). d, Membrane-bounded matrix vesicles (mv) areoccasionally encountered within the extracellular matrix. e, Selected-area electron diffraction for mineral characterization from anunstained LR White section of an anhydrously prepared sample identifies predominantly apatite within the calcified deposits, with dif-fraction maxima typical for apatite as indicated.
172 Osteopontin and Vascular Calcification
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

correlated inversely with calcium deposition (compare Figure9a and 9c). Thus, although the inhibitory effect of osteopontinon mineralization was calcium-dependent (ie, decreased byincreasing calcium concentrations), it does not appear to besimply attributable to chelation of the calcium available in themedium.
Finally, we examined the ultrastructural localization ofendogenous and exogenous osteopontin in the BASMC cul-tures by immunogold labeling. For these experiments, os-teopontin was omitted (vehicle alone) or added on day 7 afterinitiation of mineralization with calcification medium con-taining 10 mmol/Lb-glycerophosphate. Under these condi-tions, exogenously applied osteopontin (0.5mg/mL) was stillable to inhibit BASMC culture calcification by 50% at day 10(data not shown). As shown in Figure 10a, a low level ofendogenous osteopontin was found in untreated, mineralizingcultures, typically in a diffuse pattern in the mineralizedareas. In contrast, in osteopontin-treated cultures, gold parti-cles were abundant at sites of calcification, typically accu-mulating at the margins of small calcified masses (Figure10b) or associating with individual crystal profiles (Figure
10c). No gold particles were observed when preimmuneserum and protein A–gold complex alone were used ascontrols (data not shown). These data suggest that a directinteraction of osteopontin with the growing apatite crystals isrequired for its inhibitory function.
DiscussionCalcification of vascular tissue is a common finding inatherosclerosis, diabetes, renal failure, aging, aortic stenosis,and prosthetic valve replacements. Osteopontin, an acidicnoncollagenous protein, is invariably found at sites of dys-trophic calcification.13,14,17–22To investigate the mechanismsof vascular calcification and osteopontin function in thiscontext, we have examined the mineralization of vascularsmooth muscle cell cultures in vitro. Our findings indicatethat calcification of the matrix deposited by BASMCs resem-bles the mineralization observed in atherosclerotic plaquesand other types of dystrophic calcification with regard to(1) mineral type (apatite), (2) the association of mineral withextracellular collagen fibrils, and (3) the presence of matrixvesicles. Furthermore, osteopontin profoundly inhibits themineralization of these cultures by a mechanism distinct fromcalcium chelation or inhibition of alkaline phosphatase activ-ity but most likely involves direct adsorption and inhibition ofapatite crystal growth.
Our studies support and extend previous findings of Shioiet al28 using BASMCs treated withb-glycerophosphate as a
Figure 6. Effects of levamisole on calcium deposition (a) andmedium phosphorus (b) and calcium (c) concentrations inBASMCs. Calcium and phosphorus levels were measured atday 10 by the o-cresolphthalein complexone method and thephosphomolybdate complex method, respectively. In mineraldeposition experiments (a), the calcium content in cell layerextracts was normalized to cellular protein content. Data arepresented as mean6SEM; n53.
Figure 7. Effects of osteopontin (a) and vitronectin andfibronectin (b) on calcification of BASMC cultures. Cells werecultured in growth medium (Grow M.) or calcification medium(Calc. M.) containing 10 mmol/L b-glycerophosphate for 10days in the absence (2) or presence of the indicated concentra-tions of osteopontin (OPN), vitronectin (VN), and fibronectin (FN).The calcium levels were measured by the o-cresolphthaleincomplexone method and were normalized to cellular proteincontent. In panel a, calcium deposition is represented as a per-centage of calcified control (5.760.6 mmol/mg protein) and rep-resents the average of 2 separate experiments, n53 to 6 foreach treatment group per experiment. In panel b, calcium depo-sition is expressed as mean6SEM; n53.
Wada et al February 5, 1999 173
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

model for vascular calcification. In agreement with that work,we found that alkaline phosphatase activity was required forBASMC-mediated calcification in the presence ofb-glycerophosphate. However, in the previous reports, thetype of mineral formed, the presence of extracellular matrix,and the ultrastructure of the calcified deposits and theirmatrix relationships were not determined, leading to ques-tions regarding the physiological relevance of the observedcalcification. In the present study, we show that apatite is thepredominant mineral formed in BASMC cultures under theseconditions and that the mineral deposits share considerablemorphological similarity to deposits found in calcifyingatherosclerotic plaques as well as bone, cartilage, and teeth.8,9
First, mineralization of the BASMC cultures occurred pre-dominantly extracellularly in association with collagen fibrilsand rarely intracellularly or associated with necrotic debris.Second, nodular calcifications suggesting spherulitic crystal
growth, which are commonly observed in calcified athero-sclerotic plaques and valves,8 were also present in calcifyingBASMC cultures. Finally, we identified matrix vesicles inassociation with calcifying extracellular matrix in theBASMC cultures. These vesicular structures have been re-ported in calcified atherosclerotic plaques in association withelevated alkaline phosphatase activity,8 suggesting that theymay play a role in vascular dystrophic calcification aspostulated for bone, calcified cartilage, and teeth.39 More-over, matrix vesicles were also observed in the calcifiedarterial media of the MGP-null mice.16 Thus, the BASMCsused in the present studies are able to create an extracellularmilieu capable of mineralization that, in many ways, ismorphologically similar to that observed in calcified vasculartissues in vivo.
Apatite formation has also been demonstrated in peri-cytes derived from retinal microvessels40 as well as clonedbovine aortic medial cells, termed calcifying vascularcells,10 when grown in culture. In the former cells, the
Figure 8. Effects of added osteopontin on alkaline phosphataseactivity of BASMCs (a) and phosphorus concentration in themedium (b) and the direct effects of levamisole and osteopontin(OPN) on alkaline phosphatase (ALP) activity (c). a and b, Cellswere cultured in growth medium (Grow M.) or calcificationmedium (Calc. M.) for 10 days in the absence (2) or presence ofthe indicated concentrations of osteopontin (OPN). Alkalinephosphatase (ALP) activities and phosphorus content in themedium were measured 10 days after switching to 10 mmol/Lb-glycerophosphate containing calcification medium. ALP activi-ties were normalized to cellular protein content and are present-ed as mean6SEM; n53.
Figure 9. Effects of osteopontin (OPN) on calcium deposition (a)and medium phosphorus (b) and calcium (c) concentrations inBASMC cultures at various initial calcium concentrations. a, Cal-cium level in cell layer extracts was measured at day 10 by theo-cresolphthalein complexone method and normalized to cellu-lar protein content. b and c, Calcium and phosphorus levels inthe medium were measured at day 0 (pre) and day 10 (vehicleand OPN) by the o-cresolphthalein complexone method andphosphomolybdate complex method, respectively, and are pre-sented as mean6SEM; n53.
174 Osteopontin and Vascular Calcification
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

ultrastructure of mineral deposition was quite similar tothat observed in theb-glycerophosphate–treated BASMCsused in the present study, including presence of calcifyingcollagen fibrils, matrix vesicles, and agglomerated calci-fications. In contrast to the BASMCs used in the presentstudy, however, pericytes and calcifying vascular cellswere able to mineralize their matrices spontaneously in theabsence of an exogenous phosphate donor. Interestingly,addition ofb-glycerophosphate or culturing on a collagensubstrate substantially accelerated calcification of thesecultures.40 The ability of pericytes to induce calcificationin the absence of an exogenous phosphate donor may berelated to their ability to form multicellular nodules andthereby create calcium- and phosphate-rich microenviron-ments. However, we have been unsuccessful in eliminatingthe b-glycerophosphate requirement of BASMCs by cul-turing on a glass substrate, which enhances nodule forma-tion in these cultures substantially (Wada and Giachelli,unpublished observation, 1996). Alternatively, the require-ment for exogenousb-glycerophosphate may be related toas yet unknown mineralization promoting factors absent(or conversely, mineralization-inhibitory factors present)in the various types of culture medium or secreted by thecells themselves. In support of this, lipid oxidation prod-ucts, b-estradiol, transforming growth factor-b, andthrombospondin-1 have all been shown to modulate min-
eralization of pericyte cultures,41– 44 and parathyroid hor-mone–related protein dramatically inhibits calcification ofb-glycerophosphate–treated BASMCs.33 It is also notablethat we have been able to observe spontaneous calcifica-tion in cultures of human aorta– derived smooth musclecell under conditions of physiological calcium and phos-phorus levels (Jono and Giachelli, unpublished observa-tion, 1997). Clearly, a deeper understanding of the require-ments for calcification by particular vascular cells isneeded to address these issues.
Noncollagenous bone proteins including secreted, acidicphosphoproteins and Gla-containing proteins have been sug-gested to regulate physiological mineralization in general andhave recently been associated with dystrophic calcification aswell. Transgenic mice lacking MGP have recently beengenerated and show extensive dystrophic calcification ofmuscular and elastic arteries (in addition to premature andexcessive cartilage calcification), suggesting that this proteinmay normally serve to inhibit the calcification process in thevasculature.16 In addition, osteocalcin, osteopontin, and os-teonectin have also been found in calcified natural andprosthetic aortic valves, as well as calcified atheroscleroticlesions, suggesting a role for these proteins at these dystro-phic calcification sites as well.19,45–47Although studied mostextensively in bone,24,48,49 the precise functions of theseproteins are still not understood.
Figure 10. Localization of endogenous and exogenous osteopontin in mineralizing BASMC cultures. a, After immunocytochemicallabeling for endogenous osteopontin, gold particles are diffusely located over calcified areas (CAL) of the extracellular matrix. Collagenfibrils at the periphery of such areas appear calcified (arrowheads). b, After addition of osteopontin to the cultures, calcified masses aresomewhat smaller and exhibit an immunolabeling pattern distinct from that of endogenous osteopontin alone in that intense immunola-beling is observed at the margins of the calcified deposits (asterisks, CAL) and in association with peripherally located, calcified colla-gen fibrils (arrowheads). Coll indicates uncalcified collagen fibrils. c, At other calcification sites (CAL) within the cultures, osteopontin isintimately associated with relatively large crystal profiles (arrows) found among the collagen fibrils.
Wada et al February 5, 1999 175
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

We have been particularly interested in elucidating thefunction of osteopontin because of its consistent associationwith calcified vascular tissues in vivo.17–22,45–47Although therole of osteopontin in physiological mineralization has beencontroversial, in bone, osteopontin has been proposed to actas a regulator of bone formation by inhibiting calcification.Osteopontin is present in the osteoid matrix at small calcifi-cation foci, while levels of osteopontin increase dramaticallyat the mineralization front deeper in the bone. In otherstudies, osteopontin has been shown to facilitate the adhesionof osteoblast-lineage cells, particularly osteocytes and bone-lining cells,48 both of which suggest a role in bone-formativeprocesses. However, osteopontin also promotes osteoclastresorption of bone,27 thus alternatively suggesting a degrada-tive role.
In the present study, osteopontin was found to profoundlyinhibit BASMC matrix mineralization at very low concentra-tions. To the best of our knowledge, this is the first study totest the effect of exogenously applied osteopontin on extra-cellular matrix mineralization in a calcifying cell culturesystem. The mechanism of this inhibition appears to bedistinct from that of levamisole, an alkaline phosphataseinhibitor that inhibits the release of phosphate and alsoblocked calcification in this model. Thus, osteopontin treat-ment of BASMC cultures did not reduce alkaline phosphataseactivity or phosphorus levels in the culture medium. We alsoruled out simple calcium chelation/sequestration by os-teopontin, because we did not observe retention of calcium inthe medium by osteopontin at any time. Thus, although theinhibitory effect of osteopontin on mineralization wascalcium-dependent (ie, decreasing with increasing initialcalcium concentrations), it did not appear to be simply due tochelation of free calcium available in the medium. This is alsoconsistent with the calcium-binding properties of osteopontin,which have shown that'50 molecules of calcium can bebound by osteopontin at physiological calcium concentra-tions.50 Hence, it would require roughly 40mmol/L osteopon-tin (2.7 mg/mL) to chelate 2 mmol/L calcium, which is morethan 5000 times the amount of osteopontin used (0.5mg/mL)and demonstrated to be effective in inhibiting vascularcalcification in our assay. The ability of calcium to overcomethe effect of osteopontin may be related to a calcium-sensitiveconformational change in osteopontin, which has been pre-viously described.50,51
Our studies strongly suggest that osteopontin inhibition ofmineralization occurs via direct binding of this protein tocrystal surfaces. In osteopontin-treated cultures, osteopontinwas found to be intimately associated with apatite crystalsand not with unmineralized matrix or cells. Consistent withthis hypothesis, osteopontin was able to inhibit mineralizationeven when added after nucleation was allowed to proceed,suggesting that it can bind to growing crystal surfaces andinhibit any further propagation. Likewise, the accumulationof osteopontin (as laminae limitantes) on bone surfaces36,48aswell as in the concentric organic lamellae of kidney stones,where growth and calcification have been arrested in vivo,supports this hypothesis. Our findings are also consistent within vitro studies of urinary osteopontin, which was found toinhibit calcium oxalate precipitation,52 and cell-free nucle-
ation studies in which osteopontin failed to nucleate hydroxy-apatite crystals but potently inhibited crystal growth in bothgelatin and agar gels.25,26 Taken together, these studiessupport the concept that an important function for osteopontinin mineralizing tissues is as an inhibitor of crystal growthduring calcification.
Osteopontin may play multiple roles in the vascular re-sponse to injury. We and others have observed that osteopon-tin can facilitate adhesion and migration of vascular smoothmuscle and endothelial cells and increase the survival ofendothelial cells.30,53–55 These processes are integrin-mediated and require the arginine-glycine-aspartate (RGD)domain of osteopontin. These functions of osteopontin maybe particularly important for early phases of tissue remodel-ing such as in arterial neointimal formation and angiogenesis,in which osteopontin and its receptors are expressed simul-taneously by migrating vascular cells and might facilitateautocrine adhesion, migration, and/or survival of these celltypes. In fact, inhibiting osteopontin with neutralizing anti-bodies was found to block arterial intimal formation afterballoon catheter injury in the rat.56
The ability of osteopontin to regulate vascular matrixmineralization may be more important at later phases oftissue remodeling in response to injury. In atheroscleroticplaques and calcifying aortic valves, osteopontin is intimatelyassociated with calcified deposits. In both pathologies, mac-rophages associated with the chronic inflammation seen inthese diseases appear to be the major source of osteopon-tin.13,14,17–22 Our findings suggest that the high levels ofosteopontin secreted by macrophages may be a protectivemechanism aimed at preventing pervasive vascular calcifica-tion. Although there is no evidence that osteoclast-likeresorption of mineral deposits occurs in calcified vasculartissues, the finding that osteopontin might act as an opsoninfor apatite-containing particulate phagocytosis by macro-phages during wound healing57 is also consistent with ananti-calcific role of osteopontin in vascular calcification.
The capacity of osteopontin to modulate mineralizationmay be unrelated to its cell RGD-dependent adhesive func-tions. Calcium-binding domains in osteopontin have beenidentified and include an aspartate-rich domain58 and phos-phorylated residues.25,26 Our finding that vitronectin andfibronectin, both RGD-containing peptides, failed to inhibitmineralization emphasizes that non-RGD sites in osteopontinare important in regulating mineralization. In support of this,preliminary data indicate that RGD peptides do not affectmineralization nor do they block the inhibitory effect ofosteopontin on calcification of BASMC cultures (Wada andGiachelli, unpublished observation, 1996). Thus, osteopontinappears to exist as a multifunctional protein whose diversesequence motifs operate not only at the cellular level but inthe extracellular matrix as well. Importantly, the ability ofthis protein to inhibit vascular smooth muscle cell–mediatedcalcification in vitro suggests that osteopontin might be usedin therapies aimed at preventing dystrophic calcification.
AcknowledgmentsThis work was supported by NIH grants HL40079-6A2 andHL18645 (to C.M.G.), NSF grant EEC9520161 (to C.M.G.), and by
176 Osteopontin and Vascular Calcification
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

the MRC of Canada (to M.D.M.). Dr Giachelli is an EstablishedInvestigator of the American Heart Association, and Dr McKee is aScholar of the FRSQ of Quebec. We gratefully acknowledgeHsueh-Ying Yang and Manuela Almeida for purification of ratosteopontin. M.D.M. also thanks the members of the Laboratory forthe Study of Calcified Tissues and Biomaterials for their collectivesupport of this study.
References1. Cotran RS, Kumar V, Robbins SL. Cellular injury and cellular death. In:
Robbins SL, ed.Pathological Basis of Disease. 5th ed. Philadelphia, Pa:W.B. Saunders Co; 1994:1–35.
2. Schoen FJ, Levy RJ, Piehler HR. Pathological considerations inreplacement cardiac valves.Cardiovasc Pathol. 1992;1:29–52.
3. Beadenkopf WG, Daoud AS, Love BM. Calcification in the coronaryarteries and its relationship to arteriosclerosis and myocardial infarction.AJR Am J Roentgenol. 1964;92:865–871.
4. Niskanen LK, Suhonen M, Siitonen O, Lehtinen JM, Uusitupa MI. Aorticand lower limb artery calcification in type II (non-insulin-dependent)diabetic patients and non-diabetic control subjects. A five year follow-upstudy.Atherosclerosis. 1990;84:61–71.
5. Fitzgerald PJ, Ports TA, Yock PG. Contribution of localized calciumdeposits to dissection after angioplasty: an observational study usingintravascular ultrasound.Circulation. 1992;86:64–70.
6. Locker TH, Schwartz RS, Cotta CW, Hickman JR. Fluoroscopic coronaryartery calcification and associated coronary disease in asymptomaticyoung men.J Am Coll Cardiol. 1992;19:1167–1172.
7. Puentes G, Detrano R, Tang W, Wong N, French W, Narahara K,Brundage B, Baksheshi H. Estimation of coronary calcium mass usingelectron beam computed tomography: a promising approach for pre-dicting coronary events?Circulation. 1995;92(suppl 1):I-313. Abstract.
8. Kim KM. Calcification of matrix vesicles in human aortic valve andaortic media.Fed Proc. 1976;35:156–162.
9. Tanimura A, McGregor DH, Anderson HC. Calcification in atheroscle-rosis, I: human studies.J Exp Pathol. 1986;2:261–272.
10. Bostrom K, Watson KE, Horn S, Wortham C, Herman IM, Demer LL.Bone morphogenetic protein expression in human atherosclerotic lesions.J Clin Invest. 1993;91:1800–1809.
11. Keeley FW. The extraction and partial characterization of proteinsreleased by decalcification from calcified human aortic plaques.BiochimBiophys Acta. 1977;494:384–394.
12. Levy RJ, Howard SL, Oshry LJ. Carboxyglutamic acid (Gla) containingproteins of human calcified atherosclerotic plaque solubilized by EDTA.Atherosclerosis. 1986;59:155–160.
13. Shanahan CM, Weissberg PL, Metcalfe JC. Isolation of gene markers ofdifferentiated and proliferating vascular smooth muscle cells.Circ Res.1993;73:193–204.
14. Shanahan CM, Cary NRB, Metcalfe JC, Weissberg PL. High expressionof genes for calcification-regulating proteins in human atheroscleroticplaques.J Clin Invest. 1994;93:2393–2402.
15. Fleet JC, Hock JM. Identification of osteocalcin mRNA in nonosteroidtissue of rats and humans by reverse transcription-polymerase chainreaction.J Bone Miner Res. 1994;9:1565–1573.
16. Luo G, Ducy P, McKee MD, Pinero GJ, Loyer E, Behringer RR, KarsentyG. Spontaneous calcification of arteries and cartilage in mice lackingmatrix GLA protein.Nature. 1997;386:78–81.
17. Giachelli CM, Bae N, Almeida M, Denhardt DT, Alpers CE, SchwartzSM. Osteopontin is elevated during neointima formation in rat arteriesand is a novel component of human atherosclerotic plaques.J Clin Invest.1993;92:1686–1696.
18. O’Brien ER, Garvin MR, Stewart DK, Hinohara T, Simpson JB, SchwartzSM, Giachelli CM. Osteopontin is synthesized by macrophages, smoothmuscle, and endothelial cells in primary and restenotic human coronaryatherosclerotic plaques.Arterioscler Thromb. 1994;14:1648–1656.
19. O’Brien KD, Kuusisto J, Reichenbach DD, Ferguson M, Giachelli CM,Alpers CE, Otto CM. Osteopontin is expressed in human aortic valvularlesions: evidence that aortic valve calcification may be actively regulated.Circulation. 1995;92:2163–2168.
20. Ikeda T, Shirasawa T, Esaki Y, Yoshiki S, Hirokawa K. OsteopontinmRNA is expressed by smooth muscle-derived foam cells in humanatherosclerotic lesions of the aorta.J Clin Invest. 1993;92:2814–2820.
21. Hirota S, Imakita M, Kohri K, Ito A, Morii E, Adachi S, Kim H-M,Kitamura Y, Yutani C, Nomura S. Expression of osteopontin messenger
RNA by macrophages in atherosclerotic plaques: a possible associationwith calcification.Am J Pathol. 1993;143:1003–1008.
22. Fitzpatrick LA, Severson A, Edwards WD, Ingram RT. Diffuse calcifi-cation in human coronary arteries: association of osteopontin with ath-erosclerosis.J Clin Invest. 1994;94:1597–1604.
23. Giachelli CM, Schwartz SM, Liaw L. Molecular and cellular biology ofosteopontin: potential role in cardiovascular disease.Trends CardiovascMed. 1995;5:88–95.
24. Butler WT, Ridall AL, McKee MD. Osteopontin. In: Bilezikian JP, RaiszLG, Rodan GA, eds.Principles of Bone Biology. San Diego, Calif:Academic Press; 1996:167–181.
25. Boskey AL, Maresca M, Ullrich W, Doty SB, Butler WT, Prince CW.Osteopontin-hydroxyapatite interactions in vitro: inhibition of hydroxy-apatite formation and growth in a gelatin-gel.Bone Miner. 1993;22:147–159.
26. Hunter GK, Kyle CL, Goldberg HA. Modulation of crystal formation bybone phosphoproteins: structural specificity of the osteopontin mediatedinhibition of hydroxyapatite formation.Biochem J. 1994;300:723–728.
27. Ross FP, Chappel J, Alvarez JI, Sander D, Butler WT, Farach-CarsonMC, Mintz KA, Robey PG, Teitelbaum SL, Cheresh DA. Interactionsbetween the bone matrix proteins osteopontin and bone sialoprotein andthe osteoclast integrinavb3 potentiate bone resorption.J Biol Chem.1993;268:9901–9907.
28. Shioi A, Nishizawa Y, Jono S, Koyama H, Hosoi M, Morii H.B-glycerophosphate accelerates calcification in cultured bovine vascularsmooth muscle cells.Arterioscler Thromb Vasc Biol. 1995;15:2003–2009.
29. Ross R. The smooth muscle cell, II: growth of smooth muscle in cultureand formation of elastic fibers.J Cell Biol. 1971;50:172–186.
30. Liaw L, Almeida M, Hart CE, Schwartz SM, Giachelli CM. Osteopontinpromotes vascular cell adhesion and spreading and is chemotactic forsmooth muscle cells in vitro.Circ Res. 1994;74:214–224.
31. Frid MG, Shekhonin BV, Koteliansky VE, Glukhova MA. Phenotypicchanges of human smooth muscle cells during development: lateexpression of heavy caldesmon and calponin.Dev Biol. 1992;153:185–193.
32. Mallory FB. Pathological Techniques. 2nd ed. Philadelphia, Pa: W.B.Saunders Co; 1942.
33. Jono S, Nishizawa Y, Shioi A, Morii H. Parathyroid hormone-relatedpeptide as a local regulator of vascular calcification: its inhibitory actionon in vitro calcification by bovine vascular smooth muscle cells.Arte-rioscler Thromb Vasc Biol. 1997;17:1135–1142.
34. Bessey OA, Lowry OH, Brock MJ. Method for rapid determination ofalkaline phosphatase with 5 cubic millimeters of serum.J Biol Chem.1946;164:321–329.
35. Landis WJ, Glimcher MJ. Electron diffraction and electron probe micro-analysis of the mineral phase of bone tissue prepared by anhydroustechniques.J Ultrastruct Res. 1978;63:188–223.
36. McKee MD, Nanci A. Osteopontin at mineralized tissue interfaces inbone, teeth and osseointegrated implants: ultrastructural distribution andimplications for mineralized tissue formation, turnover, and repair.Microsc Res Tech. 1996;33:141–164.
37. Whyte MP. Hypophosphatasia and the role of alkaline phosphatase inskeletal mineralization.Endocr Rev. 1994;15:439–461.
38. Tenenbaum HC. Levamisole and inorganic phosphate inhibitb-glycerophosphate induced mineralization of bone formed in vitro.BoneMiner. 1987;2:13–26.
39. Anderson HC. Matrix vesicle calcification: review and update. In: PeckW, ed. Bone and Mineral Research. New York, NY: Elsevier SciencePublishers; 1985:109–149.
40. Schor AM, Canfield AE, Sutton AB, Arciniegas E, Allen TD. Pericytedifferentiation.Clin Orthop. 1995;313:81–91.
41. Watson KE, Bostrom K, Ravindranath R, Lam T, Norton B, Demer LL.TGF-b1 and 25-hydroxycholesterol stimulate osteoblast-like vascularcells to calcify.J Clin Invest. 1994;93:2106–2113.
42. Balica M, Bostrom K, Shin V, Tillisch K, Demer LL. Calcifying sub-population of bovine aortic smooth muscle cells is responsive to 17b-estradiol.Circulation. 1997;95:1954–1960.
43. Parahami F, Morrow AD, Balucan J, Leitinger N, Watson AD, Tintut Y,Berlinger JA, Demer LL. Lipid oxidation products have opposite effectson calcifying vascular cell and bone. A possible explanation for theparadox of arterial calcification in osteoporotic patients.ArteriosclerThromb Vasc Biol. 1997;17:680–687.
Wada et al February 5, 1999 177
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

44. Canfield AE, Sutton AB, Hoyland JA, Schor AM. Association ofthrombospondin-1 with osteogenic differentiation of retinal pericytes invitro. J Cell Sci. 1996;109:343–353.
45. Shen M, Marie P, Farge D, Carpentier S, De Pollak C, Hott M, Chen L,Martinet B, Carpentier A. Osteopontin is associated with bioprostheticheart valve calcification in humans.C R Acad Sci III. 1997;320:49–57.
46. Gura TA, Wright KL, Veis A, Webb CL. Identification of specificcalcium-binding noncollagenous proteins associated with glutaraldehyde-preserved bovine pericardium in the rat subdermal model.J BiomedMater Res. 1997;35:483–495.
47. Srivatsa SS, Harrity PJ, Maerchlein PB, Kleppe L, Veinot J, EdwardsWD, Johnson CM, Fitzpatrick LA. Increased cellular expression of matrixproteins that regulate mineralization is associated with calcification ofnative human and porcine xenograft bioprosthetic heart valves.J ClinInvest. 1997;99:996–1009.
48. McKee MD, Nanci A. Osteopontin: an interfacial extracellular matrixprotein in mineralized tissues.Connect Tissue Res. 1996;35:197–205.
49. Boskey AL. Matrix proteins and mineralization: an overview.ConnectTissue Res. 1996;35:357–363.
50. Chen Y, Bal BS, Gorski JP. Calcium and collagen binding properties ofosteopontin, bone sialoprotein, and bone acidic glycoprotein-75 frombone.J Biol Chem. 1992;267:24871–24878.
51. Gorski JP, Kremer E, Ruiz-Perez J, Wise GE, Artigues A. Conforma-tional analysis on soluble and surface bound osteopontin.Ann N Y AcadSci. 1995;760:12–23.
52. Shiraga H, Min W, VanDusen WJ, Clayman MD, Miner D, Terrell CH,Sherbotie JR, Foreman JW, Przysiecki C, Neilson EG, Hoyer JR. Inhi-
bition of calcium oxalate crystal growth in vitro by uropontin: anothermember of the aspartic acid-rich protein superfamily.Proc Natl Acad SciU S A. 1992;89:426–430.
53. Liaw L, Lindner V, Schwartz SM, Chambers AF, Giachelli CM.Osteopontin andb3 integrin are coordinately expressed in regeneratingendothelium in vivo and stimulate RGD-dependent endothelial migrationin vitro. Circ Res. 1995;77:665–672.
54. Liaw L, Skinner MP, Raines EW, Ross R, Cheresh DA, SchwartzSM, Giachelli CM. The adhesive and migratory effects ofosteopontin are mediated via distinct cell surface integrins: role ofavb3 in smooth muscle migration to osteopontin.J Clin Invest.1994;95:713–724.
55. Giachelli CM, Scatena M, Wada T. Osteopontin: potential roles invascular function and dystrophic calcification.J Bone Miner Metab.1997;15:179–183.
56. Liaw L, Lombardi D, Almeida MM, Schwartz SM, deBlois D, GiachelliCM. Neutralizing antibodies directed against osteopontin inhibit ratcarotid neointimal thickening following endothelial denudation.Arte-rioscler Thromb Vasc Biol. 1997;17:188–193.
57. McKee MD, Nanci A. Secretion of osteopontin by macrophages and itsaccumulation at tissue surfaces during wound healing in mineralizedtissues: a potential requirement for macrophage adhesion and phago-cytosis.Anat Rec. 1996;245:394–409.
58. Singh K, Deonarine D, Shanmugam V, Senger DR. Calcium-bindingproperties of osteopontin derived from non-osteogenic sources.J Biochem (Tokyo). 1993;114:702–707.
178 Osteopontin and Vascular Calcification
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

Takeo Wada, Marc D. McKee, Susie Steitz and Cecilia M. GiachelliCalcification of Vascular Smooth Muscle Cell Cultures: Inhibition by Osteopontin
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1999 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.84.2.1661999;84:166-178Circ Res.
http://circres.ahajournals.org/content/84/2/166World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from