biochemical characterization of tgal - a plant - t-space
TRANSCRIPT

Biochemical Characterization of TGal - a Plant Heterotrimeric G-protein a Subunit Homologue
Gilad Shevah Aharon
A thesis subrnitted in confonnity with the requirements for the degree of Master of Science
Graduate Department of Botany University of Toronto
Copflght by Gilad Shevah Aharon

National Library 1+m OfCamda Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliog raphic Services services bibliographiques
395 Wellington Street 395. nie Wellington OttawaON K 1 A W OtWwaON K 1 A W canada Canada
The author has granted a non- exclusive licence allowing the National Library of Canada to reproduce, loan, distriiute or seLl copies of this thesis in microform, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fîom it may be printed or otherwise reproduced without the author's permission.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/fh, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation,

ABSTRACT
A cDNA encoding an a subunit homologue of a heterotrimeric G-protein in
tomato (TGAI) was overexpressed in Escherichia coi1 as a GST fusion protein and
purified by glutat hione affinity . The molecular weight of the purified protein (TGa 1 ),
afier thrombin proteolysis to release the GST fragment, was 45 kDa Assays using
[ y - 3 2 ~ ] - ~ ~ ~ determined the K.,, value of GTP hydrolysis (0.075-0.125 min-') while
no hydrolysis was detected when [y-32~]-~TP was used as an alternative nucleotide.
A conserved glutamine residue in position 223 was show to regulate GTP hydrolysis
by the enzyme. A point mutation in Q223-+L resulted in a GTPase-deficient
enzyme, capable of binding GTP but with an extremely slow rate of hydrolysis.
Polyclonal antibodies were raised using TGal as an antigen and used for its
subcellular localization. Immunoblot analysis using the affinity-purified anti-TGa l
antibody detected a 45 kDa protein most abundantly in the plasma membrane and
endoplasmic reticulurn. The stimulation of the plasma membrane K-ATPase in
response to mastoparan was inhibited by the use of synthetic peptides corresponding
to the C-terminus of TGu 1. This suggests the coupling of TGa 1 to a seven-
transmembrane s p d n g receptor in plants with its C-terminus as a receptor - G-
protein contact. An immunologically-based interactive cloning screen identified an
interaction between TGal and a Dnal-like protein. This interaction may represent a
role for TGul in regulation of polypeptide translocation into the ER a mechanism
for its processing and targeting; or a mechanism for its stabilization under stress
responses.

AKNOWLEDGEMENTS
The completion of this thesis was facilitated by numerous people which I
wouid like to thank and acknowledge for their continuous help and support.
1 am forever indebted to Prof. Eduardo Blumwald for having the courage to
allow a timid young man the opporauiity to engage in scientific research under his
guidance. His drive, patience, enthusiasrn and hands-on approach enabled me to
surpass the pit-fdls often associated with research work Moreover, his fnendship and
warm personality dong with his open-mindedness always allowed for mie dialogue
and exchange of ideas. 1 am gratetiil for his past and present belief in me and hope
that this thesis is a smail demonstration of that.
I would also like to thank Dr. Michael Mayne, Dr. Roumiana Alexandrova
and Dr. John Marshall for sharing with me their expertise in molecular biology and
biochemistry. A special acknowledgernent goes out to Dr. Wayne Snedden with
whom 1 had the fortune to work as a wlleague. His inexhaustible enthusiasm dong
with his unwavering standards for science were a true driving force for excellence. in
addition, special thanks go out to Dr. Angie Gelli for spending her valuable time
patch-clamping, helping to put my work into context. For the past and present
members of the Blumwald research group, 1 would like to th& you al1 for making
that second home of ours an enjoyable place of work.
1 am grateful to rny loving partner and fiiend Margarita, for reminding me of
the existence of a wondemil world of experiences beyond the lab. Her patience,
devotion and love for life have often helped in settùig my priorities straight.

I am thankfbl to my parents and my grandmothers for supporting me
throughout both doicult and happy times. Their love and encouragement have
always allowed me to comfortably f i s my energy towards my goals. 1 am grateful
to my brother Michael for being a mie role mode1 for me throughout my life both
academically and persondly, I wish him well.

TABLE OF CONTENTS
ABSTRACC
ACKNOWEDGEMENTS
TABLE OF CONTENTS
LIST OF FIGURES
LIST OF ABBREVlATiONS
INTRODUCTION
1 .1 Biologicai Signai Transduction
1.2 The G-protein Superfamily
1.3 The Small (Monomeric) G-Proteins
1 -4 The Heîerotrirneric G-proteins
1.5 Biochemical Tools in Studying Receptor-Coupled G-Protein Systems
1.6 Biochemical Evidence for Membrane Associated G-proteins in Plant Signalhg
1.7 Cloning and Characterization of Plant Genes Homologous to the Heterotruneric G-Rotein Family
MATERIALS AM) METHODS
2.1 Cloning and mutagenesis of the TGA 1 cDN A
2.2 Overe.vpressio11 and -cation of Wild Type and Midant TGul
2.3 SDS-PAGE and Western hunoblotting
2.1 GTP HydroIysk and Binding Assays
2.5 Production of Pofyclonal anti-TGar 1 a n t i i e s
2.6 f lant Material
2.7 Isolation of Plant Membranes and Cytosol
2.8 Detennjnaîion of Protein Concentration
Page
1
I l
iv
vi
.-. VUt
1
1
3
6
7
9
14
19
2 1
22
22
25
28
29
31
32
3 2
33

2.9 Measurement of P h m a Membrane AïPase Activities
2.10 Synthetic Peptides
2.1 1 Immunologi~-Based Interactive Cloning
RESULTS
3.1 Overexpression and Purification of Recombinant TGu 1 Proteins
3.2 GTP Hydrolysis and Binding by TGal
3.3 Immunological Detection and Subcellular Localization of TGa 1
3.4 Cbaracterization of a Putative G-Protein-Coupled Receptor Contact
3 -5 Screening for TGa 1 Interacting Proteins
DISCUSSION
4.1 Purification of Recombinant TGa 1
4.3 Characterization of the TGa I -Q223L Mutant
4.5 Possiile d e s for TGa 1 in the ER
4.6 Fundonal Intpiicatious of TGa 1 - interacting with a DnaJ-üke Protein
4.7 Possible roles for TGa 1 in the Plasma Membrane
1.8 Mechanisms of Heterotrimeric G-protein Involvement in Host-Pathogen interactions
REFERENCES

LIST OF FIGURES
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Figure 6
Figure 7
Figure 8
Figure 9
Figure 10
Figure 11
Figure 12
Figure 13
The GTPase cycle
Mechanisms of heterotrimeric G-protein action
Scheme for PCR-based sitedirecteci mutagenesis
Cloning strategy of the TGA 1 constxucts into the pGEX-2TK expression vector
Strategy for an immunologically-based interactive cloning screen
Coomassieblue stained 1 OYO SSDS-PAGE of total proteins fkom bacteria carrying the pGEX- TGA I expression vector
A western immunoblot ushg an anti-GST antibody to identifjr the GST-TGal fiision protein in total proteins &om bacteria carrying the pGEX- TGAI expression vector
Purification of recombinant GST-TGa l (WTjQ223L) and TGu 1 (WT/Q223 L) by glutathione affinity
GTP hydrolysis assays using [ y - ) 2 ~ ] ~ ~ ~ to monitor [ 3 2 ~ i ] released by recombinant TGa 1 -WT and TGaLQ223L enzymes
GTP binding and hydrolysis assays on enzymes transferred to nitrocelluloce membranes
A western immunoblot using an ffiity-purified anti-TGa l polyclonal antibody
Identification of a potential receptor contact region in the C-terminal sequence of TGa 1
Inhibition of mastoparan-stimulateci activation of the plasma membrane K-ATPase by synthetic peptides, correspondhg to the TGul C-terminus sequence
Page
4
11
23
26
37
40
43
46
48
52
55
58

Figure 14 Sequence alignment of the predicted amino acids (fkom the p d a l 5 ' DNA sequences) of the four proteins identifid as interacting with TGal - GIP4- 1, 6-2,8- 1 and 9-1 65
Figure 15 Sequence alignment of the compileci 205 amino acids (from ail four TGa 1 -interacting proteins) with known DnaJ-like proteins
Figure 16 A proposed mode1 for the regulation of a DnaJ-like protein, by TGa 1, in polypeptide translocation in the ER
Figure 17 Regulation of a plasma membrane ca2- channel by TGal
Figure 18 Putative mechanism for the involvement of membrane-associated G-proteins in host-pat hogen interactions

LIST OF ABBREVLATIONS
A - adenine ADP - adenosine diphosphate ATP - adenosine triphosphate BCIP - 5-brorno-4-chloro-3 -indolyl-p hosphate bp - base pair BSA - bovine serum albumin C - cytosine Ca - calcium CAMP - adenosine 3 *, 5 kyclic mono phosphate cGMP - guanosine 3 *,5 O-cyclic monophosphate Cl - chloride CTX - Choiera toxin Da - dalton O C - degrees centigrade DNA - deoxyribonucleic acid D'M' - 1,4-dithiothreitol EDTA - ethylenediaminetetra-acetic acid EGTA - ethyleneglycol-bi~~aminoethyIether)wN,N,N,N,- tetraacetic acid ER - Endoplasmic reticulum Fas - antigen-binding hgment g - gram x g - times g force G - guanine Gprotein - GTP-binding protein GAP - GTPase - activating proteins GDP - guanosine diphosphate GDP(f3)S - guanosine-5 '-O-(2-thiodip hosp hate) [~PP]GTP - guanoçined '-[a-32~]-triphosphate [FP] GTP - guanosine-5 '-[y-32~] -trip hosp hate [ 3 S ~ ] ~ T P y ~ - Guanosine 5 '[y-35~]-thiotriphosphate GEF - guanine nucleotide exchange factors C;I - inhibitory G-protein GNRP - guanine nucleotide reiease proteins Ga- olfact ory G-prot ein Gpp(NB[lp - guanosine 5 '-@,y-imid0)triphosphate Gs - stimulatory G-protein GST - Glutathione-S-ansf ferase Gt - transducin G-protein GTP - guanosine triphosphate GTPyS - guanosine 5 '-[y-thioltrip hosphate GUS - B - glucuronidase

- hydrogen peroxide Hepes - 4-(2-hydroxyethyl)- l -piperazineethanesulfonic acid ER - hypersensifve response IAA - indole-3-acetic acid 4 - inositol- 1 -4.5-triphosphate rm*rG - isopropyl f3-D-galactopyranoside k - kilo K - potassium LB - Luria-Bertani media p - micro pCi - microCurie(s) m - mili M - molar mas7 - synthetic mastoparan mwCP - mastoparan, control peptide min - minute(s) Na - sodium NAD - nicotinarnide adenine dinucleotide NBT - nitn>bIue tetrazolium ORF - open reading frarne p - pic0 PAGE - polyacrylamide gel electophoresis PBSm - phosphate b e e r dindween-20 PCR - polymerase chah reaction Pi - inorganic phosphate Pm - phosphatidylinositol-4,5-diphosphate PMSF - pheny lmethylsulp hony 1 fluoride PO4 - phosphate PR protein - pathogenesis-related protein PTX - pemissis toxin R gene - Resistance gene RGS - regdators pf G-protein signalling SA - salicylic acid SDS - sodium dodecyl sulfate 7TMS - seven-transmembrane spanning S 0 4 - sulfate T - thymine TBS/T - Tris buffer sdine/Tween-20 v - volts v/v - volume per volume whr - weight per vohune

INTRODUCTION
1.1 Biological Signal Transduction
Signaling in multicellular organisms requires a complex network of
communication which relies both on extracellular as well as intracellular components.
This signaling apparatus enables the perception of signals. their propagation and
translation into a subsequent set of responses. This cascade cm happen within or
between tissues allowing for the CO-ordination of signals on which the organism relies
for normal growth and development.
The concept of signai transduction implies the linkage of a number of
components that convert information from one form to another. The study of signal
transduction pathways therefore involves the elucidation of the particular components
in the biochemical reaction cascades - the triggering signai, mechanisms of its
intemalization, propagation and direction to produce appropriate short- and long-tem
responses.
Despite the variety of signds, responses and the complexity of multicellular
organisms, some cornmon themes appear to emerge from the different mode1 systems
investigated to date. This cm be exemplified by the following signaling paradigm in
which the signai (e.g. a hormone) is perceived at the plasma membrane. leading
directly or indirectiy to the release of a secondary messenger inside the cell.
Modulation of the concentration of secondary rnessengers may affect enzyme
activities through changes in the phosphorylation and dephosphorylation status of the

ce11 through the action of protein kinases and phosphatases as a pst-translational
modification (Barford, 1991). Within the cell. the cascade can be further transmitted
to other compartmentalized organelles such as the nucleus. Within the nucleus.
nucleic acid processing may be regulated to modulate the levels of particular
transcripts in accordance with their requirement. by transcriptional factors.
Like al1 organisrns, plants have to face a wide variety of environmental
conditions. These include light, gravity, wind, temperature, predators, pathogens. and
moisture. Unlike animals, plants are sessile and have to cope with environmental
stresses through a specialized anatomy and physiology. In addition. at the cellular and
subcellular levels, more active responses have to be employed in dealing with
environmental conditions imposed on the plant. Since many aspects of signal
transduction in animals are understood in considerable detail. this a priori knowledge
is often used in many cases as a reference mode1 when investigaring plant signaling.
Evolutionary relationships and hindamental biochemical and molecular similarities
between ail organisms give credence to this assumption. However, phenornena that
are of focus in animals such as muscle contraction. immune responses and oncology
do not appear to have much in comrnon with phenornena in plants (Verhey and
Lomax, 1993). Therefore, although there may be a biochemical similarity between
plants and anirnals, the context of these components is still unclear and remains to be
uncovered.

1.2 The G-protein Superfamily
The G-protein superfamily is an example of important signaling molecules in
anirnals that have been only recently demonstrated in plants. It is a large family of
guanine-nucleotide-binding proteins that have ken classified. on the basis of subunit
composition and size, into heterotrimenc (consisting of a, and y subunits) and
small (monomeric) G-proteins (Ma 1994). Regulation of G-protein signaling is
dependent on the binding and hydrolysis of GTP, known as the GTPase cycle. This
cycle includes three conformational States of the protein: (i) A GDP-bound 'inactive'
state, which in the case of the heterotrimeric G-proteins results in the association of
the different subunits; (ii) A transient, 'empty' state resulting from the release of
b a n d GDP replaced by GTP. Since cells contain -100 rnM GTP and -10 mM GDP
(Boume et al.. 199 1). it appean that the substitution of nucleotides from GDP to GTP
is a random event rather than enzyme mediated. The higher concentration ratio of
GTP to GDP in the cytosol would preferentially promote GTP to enter the empty
guanine nucleotide binding site; and (iii) Upon binding of GTP. the protein assumes
an 'active' conformation which allows its regulation of downstream elements (Fig. 1)
(Bourne et al., 1990).
The length of activation of downstream elements by G-proteins is determined
by the relative rate of two reactions: the rate of dissociation of GDP from the GDP-
bound form (kuseGDP) and the hydrolysis of bound GTP (kc;itm). These two reaction
rates determine the fraction of protein molecules in the 'active' conformation.

Figure 1. The GTPase cycle.
This cycle includes three conformational States: (i) A GDP-bound, 'inactive*
state. (ii) A transient, 'empty* state. resulting from the release of bound GDP to be
replaced by GTP. (iii) A GTP-bound, 'active' state.
The length of activation of a G-protein is determined by the relative rate of two
reactions. Transition from the 'inactive* state to the 'empty' state is controlled by the
rate of dissociation of GDP (KdbsGDP) which can be catalyzed by GNRPs (guanine
nucleotide release proteins). Transition from the 'active' state to the 'inactive' state
depends on the rate of GTP hydrolysis (KCa,-), which can be increased by GAPs
(GTPase - activating proteins).

E mpty S tde
GDP 9
Inactive S tate Active S tate

6
The ratio between the GTP-bound and GDP-bound forms is represented by the
following equation:
This equation depends on the assumption that the concentration of GTP is not
limiting, and thus, cm rapidly bind to an empty site on the G-protein.
According to this equation, increasing the proportion of 'active' G-proteins
depends either on the acceleration of kdkGDP or the reduction of kt-. In many G-
proteins the intrinsic rate constants of GDP release and GTP hydrolysis are quite low
(4.03 min-') but were found to be regulated by either of two classes of regulatory
proteins: (i) Proteins that catalyze the release of bound GDP, promoting exchange by
GTP, known as the guanine nucleotide releasdexchange proteins/factors (GNRP's 1
GEF's), and (ii) GTPase - activating proteins (GAP'S), that increase the rate of GTP
hydrolysis (Fig. 1) (Bourne et al., 199 1).
1.3 The Small (Monomeric) G-Proteins
The small G-proteins (20-30 kDa) have a consensus sequence for GTP-
binding, which is related to that found in the a subunit of the heteroaimeric G
proteins. On the basis of amino acid homologies, the small G proteins have been
grouped into several subfamilies. These include the ras, rab/ypt and rho/rac. The best
studied subfarnily is the ras family which encodes several proto-oncogenes
homologous in invertebrate animals and in yeast. Regulation of the Ras protein is

directed both by GEF's and GAP'S. In addition. evidence suggests a regdation of Ras
activity by extracellular signals, mediated through a tyrosine kinase membrane
receptor. Downstream of Ras. a conserved MAP kinase cascade has been uncovered
(Kaziro et al., 199 1). Data from Drosophila and C-elegans suggests a role for Ras in
signal transduction during development (Boguski and McCormick. 1993). The Rab
proteins appear to be involved in vesicular transport in the secretory pathway.
Evidence suggests their association with membrane vesicles which shuttle between
donor and acceptor membrane structures (Nuoffer and Balch, 1994). Finally, the rho
subfamily is known to function in celi polarity and cytoskeletal functions (Hall.
1 9%)).
1.4 The Heterotrimeric G-proteins
The heterotrimeric G-proteins were first identified in mammalian
transmembrane signaling. Initial evidence indicated the requirernent for GTP in the
hormonal activation of adenylyl cyclase (ATP pyrophosphate-lyase cyclizing enzyme)
which is responsible for the production of the CAMP secondq messenger (Rodbell et
al., 1971a). In addition to this observation, the presence of GTP was reported to
decrease binding of the hormone glucagon to recepton. which regulate adenylyl
cyclase activity (Rodbell et al., 197 lb). This effect was discovered to be specific for
agonists, which affinity to cognate receptors decreased, in the presence of guanine
nucleotides (Maguire et al., 1976). The fmt GTPase assays were done in turkey
erythrocyte membranes in response to catecholamine. These experiments

demonstrated the presence of G-protein - linked systems which were activated upon
the binding of GTP and deactivated by its hydrolysis. Dissociation of GDP was
determined to be the rate lirniting step, controlied by a receptor (Cassel et al., 1977).
Ce11 hision experiments, in which the components of the adenylyl cyclase were mixed
and exchanged (Orly et al., 1976), allowed their subsequent reconstitution in vitro.
Purification of the signaling components, i.e. the p-adrenergic receptor (Shorr et al,
1981). G-proteins (Nonhup et al.. 1989) and the adenylyl cyclase itself (Pfeuffer et
al., 1982) were faciiitated both by the assays developed as well as by
chromatographic techniques (Gilman 1987).
Since their initial discovery, other mamrndian G a proteins that are
structuraily and functionally related to Gs have k e n identified. The activation of the
adenylyl cyclase by Gs (stimulatory) was found to be subsequently inhibited by a
different G-protein named Gi (inhibitory) (Gilman, 1984). The G, (transducins) were
found to regulate visual excitation by stimulating the retinal cyclic GMP
phosphodiesterase in retinal rod outer segments in response to light (Fung et
al.,l981). The Goif - for olfactory response - were observed to regulate taste sensation
(Jones and Reed, 1987). Furthemore, G-proteins have k e n implicated in regulating
the phosphodiesteratic cleavage of phosphatidylinositol-4,s-diphosphate (PB) to
inositol-l,4,5-triphosphate (Pd. This phosphoinositide hydrolysis, mediated by the
enzyme phospholipase C, is known to result in the release of intracellular ca2+ stores
in response to many hormones (Stemweis and Srnrcka., 1992). In addition, there is
growing evidence suggesting direct regdation of ion channels by G-proteins. These

include neuronal, inwardly rectifjmg K? channel, cardiac ~ a ' + channels and others
(Clapham, 1994). Although the known heterotrimeric G-proteins are mostly localized
to the ce11 plasma membrane, there is evidence supporting their presence in other
intracellular membranes. As an example, Gm has been localized to Golgi membranes
in several cell types. Its role appears to be in vesicular transport, but its precise
function still remains unknown (Vries et al., 1995). Unlike plasma membrane G-
proteins, very little is known about possible receptors and effectors coupled to
heterotrimeric G proteins associated with intracellular membranes.
It is worth noting that activation of downstream elements by heterotrimenc G-
proteins is not exclusive to any of the protein subunits. Both the a and py complex
are involved in signal mediation, and were observed to exert both stimulatory and
inhibitory effects (Clapharn 1993).
1.5 Biochemical Twls in Shidying Receptor-CoupIed GProtein Systems
A number of diagnostic and biochemical techniques are available to examine
the role of G-proteins in a particular system. The basic criteria to be considered when
examining a system for regdation by G-proteins include: (i) Both a ligand for the
receptor of interest and GTP are required to initiate the response in question; (ii) The
use of non-hydrolyzable analogues of GTP (GïF@ or Gpp[NII]p) can mimic the
response in the absence of the appropriate Ligand; (iii) Cholem toxin ((JTX) andior
pertussis toxin (PTX) have characteristic eEects on the function of known G-proteins.

These bacterid exotoxins are able to catalyze the covalent modification of sorne G-
proteins. CTX isolated fiom cultures of Vibrio cholerae. is able to catalyze the ADP-
ribosylation of Ga, using NAD as a substrate. The effect of this modification on Gsa
is to reduce its rate of GTP hydrolysis. G,a is therefore maintained in its active
conformation (Cassel, and Zelinger, 1 977; Moss and Vaughan, 1977). S imilarly,
PTX, isolated from cultures of Bordetella pertussis. cataiyzes the ADP-ribosylation of
the 'Gi-like' G-proteins on a cysteine residue in the C-terminus of the protein. The
functional effect of this modification is the uncoupiing of the receptor - G-protein
contact (Katada and Ui, 1982). As a result, Gi will maintain its inactive aey
conformation even in the presence of an agonist. In animai systems the in vivo
consequences of both these toxins is the elevation of CAMP levels by the constitutive
activation of G,a by CTX, or inactivation of the inhibitory Gia by PTX (Fig.2)
(Simon et al.. 1991); (iv) Immunological detection by antibodies with different
reactivities for individual G-proteins; and (v) the reconstitution of the individual
components of a particular pathway - such as in the case of the adenylyl cyclase
system. (Gilman 1987).
Mastoparan is the major component of wasp venom and its application was
observed to cause the degranulation of mast cells. It is a 14 amino acid peptide with a
molecular weight of 1480 D a Circular dichroism studies of mastoparan have
determined that its Iargely unordered structure in aqueous solution is convened to an
a-helical tetramer upon binding with a phospholipid membrane. This conversion is

Figure 2. Mechanisms of heterotrimeric G-protein action.
A number of biochemical compounds are available to study G-protein
activation or inhibition. Activation of heterotrimeric G-proteins can be achieved by
the use of GTPyS, a non-hydrolyzable GTP-analogue, or CTX (Choiera toxin) which
covaiently modifies the a subunit to inhibit its GTP hydrolysis. Both result in the
constitutive activation of heterotrimenc G-proteins. The PTX (Pertussis toxin)
covaiently modifies the a subunit to uncouple its receptor contact. preventing
activation of the G-protein in response to an agonist
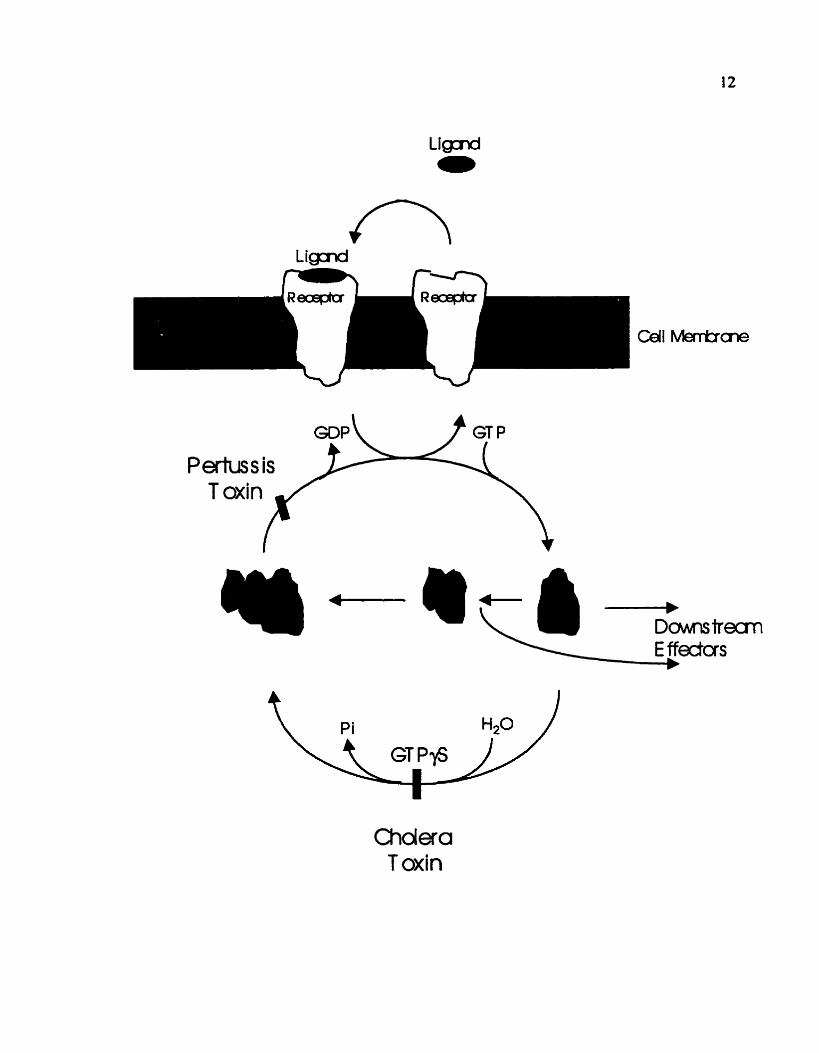
Choiera T oxin

thought to occur as a result of the interaction of the hydrophobic moiety of
mastoparan with the hydrophobic interior of the phospholipid membrane
(Higashijima et al., 1984). hirified G proteios reconstituted into phospholipid
vesicles had an increased GTPase activity and GTP-binding in the presence of
mastoparan (Higashijima et al., 1988). These results suggested a direct interaction
between mastoparan and G a which was Iater confmed by Wenigarten et al. (1990).
Polyclonal antibodies raised against the C-terminal peptide of Gi biocked mastoparan-
stimulated GTPase activity while mastoparan antagonized the ability of the antibody
to detect Gi. Interestingly, biochemical studies of the structure of G a have suggested
that its C-terminal region is involved in receptor contact. Experiments using synthetic
oligopeptides corresponding to the C-terminal sequences as well as monoclonal
antibodies raised against this portion of Gta were used to block its interaction with
the photoreceptor rhodopsin (Kaziro, 1992). The above observations suggested that
mastoparan interacts with G proteins at a conserved receptor-binding domain in a
marner smicturally as well as functionally similar to that of hormone receptos. It has
k e n suggested that the stnictured a helix, formed upon binding of mastoparan to
phospholipid membranes, is reminiscent of the third intracellular Ioop of the seven-
transmembrane spanning (7TMS) receptors. These fonn the largest family of G
protein-coupled receptors. and their cytoplasmic domain (mimicked by mastoparan) is
thought to be important in determinhg G protein interactions (Higashijima et al..
1988). Mastoparan and its synthetic analogue mas7 are therefore commonly used as

14
diagnostics for the existence of G protein coupled TïMS receptors in a variety of
systems.
1.6 Biochemical Evidence for Membrane Associated G-proteins in Plant
Signaling
In recent years there has been steady accumulation of biochemical evidence
suggesting a role for membrane associated G-proteins in plant signaling. Initial
studies included demonstrations of high-affinity binding of the GTP analogue
[ 3 s ~ ] ~ T P @ to membrane fractions frorn various plant species such as Arabidopsis
thaliana (Blum et al., 1988), Lemna paucicustata (Hasunuma et al., 1987) zucchini
(Jacobs et al., 1988) as well as tobacco and maize (Wise et al., 1991). The specificity
of GTP binding to membrane fractions was confmed in these experiments by
nucleotide cornpetition assays.
In addition to these preliminary experiments, immunological studies have
been carried out using antiserum raised against animal G a or synthetic peptides
derived from animal G a sequences. ~uno log ica l ly related proteins of expected
sizes were detected in the plasma membrane of Arabidopsis (Blum et al., 1988;
Clarkson et al., 1991,), zucchini (Jacobs et ai., 1988). bean (Blum et al., 1988). pea
(Warpeha et al., 1991), soybean (Legendre et al., 1992) and tornato (Xng et al.,
1997). Aso, studies using the cholera andfor pertussis toxins demonstrated the ADP-
ribosylation of plasma membrane proteins of simüar size to cornmon G-proteins in

Lemna paucicostatu, pea, soybean. and tomato. (Hasunuma et al., 1987; Warpeha et
ai.. 199 1; Legendre et al., 1992; Xing et ai.. 1997).
A number of studies have implicated GTP-binding proteins in light-stimulated
signaling. Romero et al. (1991). observed 2 1% stirnulated binding of [ 3 5 ~ ] ~ T P y S in
etiolated Avenu seedluigs in response to a 5 minute red-light irradiation. The red-light
induced stimulation was abolished in response to far-red light. Neuhaus et al. (1993)
used the GTP analogues GTPyS (30-100 pM) and Gpp(NH)p (50-100 pM)
intracellularly. to mimic the effects of the light receptor phytochrome A (PhyA) on
light-dependent gene expression in mutant tomato ceik deficient in PhyA. These
include the synthesis of anthocyanins, and the expression of the GUS reporter gene
fused to the light regulated cab promoter in tomato seedlings. The use of caZ' and
calmodulin, mirnicked the same gene expression patterns as observed in response to
the GTP analogues, suggesting a role for G-proteins in plant caZ+ signaling. These
results, as weil as the red light/far-red-light responses. strongly suggested
phytochrome-mediated G-protein activation. In addition to red Light, Warpeha et al.
(1991) described blue iight stimulation of GTPase activity, as weil as GTP-binding
activity in the plasma membrane of etiolated pea seedlings. A 40 D a protein,
irnmunologically related to an a subunit, was ADP-ribosylated by PTX only in the
absence of GTP or blue light. These results were explained by the fact that PTX ADP-
ribosylation is more efficient in GDP-bound. inactive a subunits.

A possible role for G-proteins was described in response to the plant hormone
auxin, indole-3-acetic acid (IAA). Its application to membrane vesicles, denved fiom
rice coleoptiles, was observed to stimulate binding of GTPyS. Furthemore, pn-
incubation of vesicles with GTPyS caused a reduction in auxin binding (Zaina et al.,
1990). These results rnay be explained in one of two ways: firstly, the auxin receptor
was desensitized by the activation of the G-protein by GTPyS. or secondly. the auxin
receptor required the G-protein to be in its inactive. GDP-bound fom, to interact with
the auxin ligand (Ma, 1994).
Some lines of evidence have suggested the presence of a system in plants
similar to that of the receptor-coupled G-protein in animals. Much of it is due to the
use of mastoparan to investigate the role of G proteins and possibly 7TMS receptors
in plant signaling. Wise et al. (1993) demonstrated a two-fold increase in the binding
affinity of [3'~]~TPyS to the plasma membrane of both pea and maize upon treatment
with a mastoparan analogue masi. These observations indicated activation of plant G
proteins in an analogous manner to that of animals. In addition, Armstrong et al.
(1995) reported the inhibition of inward K* currents by mas7 in intact Vicia foba
guard cells, as well as by GïPyS. The control peptide rnasCP, which is similar in
structure to mas7 but lacks the ability to activate G proteins. had no effect. The
authors' conclusion was the existence of a receptor - G-protein coupling system in
guard cells.

Legendre et al. (1992) used mastoparan in investigating the defense response
of rapid oxidative burst in cultured soybean cells. This oxidative burst, reached
maximum intensity within 1-5 minutes upon addition of relatively pure pathogenic
elicitors. Mastoparan was shown to mimic the elicitor's effect in a concentration-
dependent manner. Additional experiments included the introduction of the antigen-
binding fragment (F*) of an ânti-Ga~o-n, (raised against a consensus sequence of
animal G proteins recognizing an irnmunologically related plant protein), into
soybean ceils, using a biotin-rnediated delivery technique. Interestingly, the rapid
oxidative burst in response to elicitoa was enhanced in those cells up to 10-fold.
Aside fkom demonstrating a functional role for G proteins in plants, these results
suggested that more than one GTP-binding protein may participate in the regdation of
the elicitor-stimulated oxidative burst. This possibility stems from the fact that the
antibody used, rather than inhibiting H20t production, led to a more intense. longer
lasting burst, once the reaction has k e n started by an elicitor.
There is additional growing evidence for the involvement of G-proteins in the
signaling cascades involved in plant-pathogen interactions. Beffa et al. (1995)
conducted experiments using the cholera toxin (CTX) which is used in activating
signaling pathways dependent on heterotrimeric G-proteins. The researchen
transformed tobacco plants with a chimenc gene encoding the A l subunit of CTX
regulated by a Light-inducible promoter Cab-1. Reduced susceptibility to the bacterial
pathogen Pseudomonas tabaci was observed in tissues of transgenic plants. Detailed
molecular analysis of the transformed tissues demonstrated the accumulation of high

levels of salicylic acid (SA) and the constitutive expression of pathogenesis-related
(PR) protein genes encoding PR- 1, the class II isoforms of PR-2 (p- 1 f -glucanase)
and PR-3 (chitinase). This subset of PR proteins, as well as the accumulation of SAT
are a characteristic of the systemic acquired resistance (SM) response in plants.
Genes encoding the class 1 PR-2 and PR-3 isoforms are induced in tobacco by
ethylene or by other stress, and locally as part of the HR, but they are not induced
systemically in SAR. These genes were not induced in the CTX transformed tissues
and showed normal regulation. Beffa et ai. (1995) suggested that CTX expression
does not trigger non-specific stress reactions. Furthemore. microinjection
experiments showed that CTX induces expression of PRI-GUS but not that of the
GU-GUS transgene (containing the promoter region of the GLB gene encoding a
class 1 isoform of the PR-2). These results seem to suggest a role for CTX-sensitive
G-proteins in specific defense responses such as SAR.
Geili et al., (1997) observed the activation of a plasma membrane ca2'-
permeable channel, in response to Cladosponum fulvum race-specific eliciton, in
tomato suspension culture cells. The stimulation of a ca2+ influx in response to
fungal elicitors, was mirnicked by the use of GTPyS or mastoparan. Furthermore, pre-
incubation with GDPPS, a GDP analogue that locks heterotrimeric G-proteins into
their inactivated state, abolished the channel activation induced by the hingai elicitors.
These results not only implicated G-proteins directly in host-pathogen interactions.

19
they also demonstrated a role for G-proteins in modulating an important plant
secondary messenger which may have a role in the plant defense response.
1.7 Cloning and Characterization of Plant Genes Homologous to the
Heterotrirneric G-Protein Family
In spite of the overwhelming evidence for the role of membrane-associated G-
proteins in plant signal transduction, none were identifed or purified to date.
However, a number of cDNA's, showing homology to subunits of the heterotrimenc
G-proteins were cloned (Ma, 1994). The fmt to be cloned, by degenerate PCR, was
GPAl from Arabidopsis thaliana (Ma et al., 1990). Its putative open-reading-frame is
36% identicai and 73% similar (with conservative changes) to the mamrnalian Gi and
transducins. Northem blot anaiysis, using GPAI cDNA as a probe, suggested that
GPAI mRNA was most abundant in vegetative tissues, including leaves and/or roots.
less in floral stems, and least in floral buds and floral meristem. Southern blot
analysis, at low- stringency hybridization with the GPAI cDNA, uncovered additional
bands, suggesting the presence of other homologous cDNA's (Ma et al., 1990).
Nevertheless, PCR and low-stringency hybndization screenings have not yet
uncovered additional homologues for GPAl in Arabidopsis thdiana (Ma., 1994).
Instead, GPAI homologues from other plants were cloned using low-stringency
hybridization with GPAl as a probe. These include the tomato TGAI. which is 84%
identical to the GPAl protein sequence (Ma et al., 199 1). and others from Lotus
japonicus (Poulsen et al., 1994), two from soybean (Kim et al, 1995; Gotor et al.,

1996), and two from rice (Seo et al., 1995; Iwasaki et al., 1997). Also, genes encoding
proteins showing sequence similarities to the animal f! subunit of the heterotrimeric
G-proteins were isolated from Arabidopsis and maize (Weiss et al., 1994), tobacco
(Ishida et al., 1993) and from rice (Ishikawa et al, 1996).
Initial work, airning at the identification of their functions, has been carried
out with a number of these clones. Weiss et al., (1993) used a specific antibody raised
against a peptide from the C-terminal region of GPal - the GPAI gene product - to
conduct extensive irnmunological detection and localization throughout
development. The authors found higher levels of the GPAI gene product in immature
organs than in mature organs. In mature organs GPal was present primarily in the
vascular tissue and mesophyll cells. In developing organs, GPal was present at high
levels in the mot meristem and elongation zone, in the shoot and floral menstems, in
the leaf and floral organ primordia, in developing embryos, and in growing pollen
tubes and nectaries (Weiss et al, 1993). The authors suggested that the complex
localization patterns of GPal were indicative of its role in different signaling
pathways, depending on the plant's developmental stage. SubceIlular localization of
GPal in plant membrane fraction, by irnmunological detection, indicated its presence
both in the plasma membrane and the ER (Weiss et al., 1997).
Wise et al. (1997) have overexpressed GPal in E-coli and demonstrated the
GTP-binding capacity of the recombinant enzyme. The rice RGAI product,
overexpressed in E. d i , was shown to be ADP-ribosylated by PT'X in vitro (Seo et

al.. 1995). In addition. both rice gene products - RGa1 and rGricea were subcellularly
localized to the plasma membrane by western blot andysis, and shown to have GTP
binding and hydrolysis activities (Seo et al., 1997; Iwasaki et ai., 1997).
OBJECTllVES
The aim of this study is to biochemically characterize the gene product of
TGAl. the only tomato heterotrimeric G-protein a subunit homologue cloned to date
(Ma et ai., 1991).
The particular objectives are: (i) To overexpress the TGAl gene product in a
heterologous system (such as E.colî) to allow for its purification as a recombinant
enzyme. (ii) To use the purified protein as an antigen for the production of specific
polyclonal antibodies to be used for its subcellular localization. (iii) Identifiy
important domains and amino acid residues involved in regulathg its activity. (iv)
Isolate proteins that interact with the TGAI gene product, in an attempt to elucidate its
potential functions.

MATERIALS AND METHODS
2.1 Cloning and mutagenesis of the TGAI cDNA
Subcloning was carried out by PCR using Vent DNA polymerase (NEB)
and TGAl (provided by Dr. Ma, H. Cold Spring Harbor, NY, Ma et al.. 1991) cDNA
as a template. Two oligonucleotides:
Barn-tga 1 - 5'-TATGGATCCATGGGCTTCGTTGTGC-3 ' and
Eco-tgal - 5 '-mGAATTCTCATAGTAAACCTGC-3 ' were designed for
amplification of the coding sequence of TGAl with BamHYEcoRI restriction sites for
in-frame cloning with GST into the BamHUEcoRI sites in the pGEX-2TK vector
(Pharmacia).
PCR-based point mutagenesis was canied out as described by Zhao et al.
(1993) with some modifications. An oligonucleotide - Q223LtgaI - containing a
base substitution 5 =GïTGGAGGTCnAGAAATGAG-3 ' was used in conjunction
with primer Eco-tgal to ampli@ a 506 bp Eragrnent corresponding to the 3' region of
the TGAl template. The PCR product was purifed frorn an agarose gel using the
geneclean iII kit (BiolOl) and used as a primer in a second PCR reaction dong with
primer Barn-tgal to generate the full-length coding sequence of TGAI carrying the
one base-pair substitution (Fig.3). For all PCR reactions, a maximum of 21
amplification cycles were used, to minimize nucleotide misincorporation. Since Vent
DNA polymerase produces blunt-end products, both wild type and rnutagenized PCR

Figure 3. Scheme for PCR-based site-directed mutagenesis.

A ï E A 1 cDNA template 5'
Use the 506bp product as a primer dong with Barn-tgal using TGA 1 as a cDNA template
Amplify a 506bp fragment w ith Q223L-tga I and Eco-tgal primers
5' 3' Eco- tga 1 h
506bp Mutagenized PCR product
BamHI 5' 3'
EcoRI Site
3' Site
A full length TGAl - Q223L cDNA Mutant Ready for Subcloning in Designated Vector
506 bp mutagenized PCR product

25
product were bluntend ligated into pGEM-7Zf(+) digested with SmaI. The inserts
were subsequentiy digested using BamHIIEcoRI and Ligated into pGEX-Zn< that was
cut with those same enzymes (Fig.4). AU constnicts were sequenced to confim
mutagenesis and fidelity.
2.2 Overexpression and Purikation of Wild Type and Mutant TGal
Wild type and mutagenized constructs cloned in pGEX-2TK were used to
transfom E.coli BU1 (pLysS). An overnight culture grown in LI3 medium
containing 50 pg/d ampicillin was diluted 1: 100 into 800 mi of LB medium. Culture
was grown at 30% for 2 hours at which point isopropyl P-D-gaIactopyranoside
(IPTG) was added to a final concentration of 0.05 mM to induce expression.
Incubation continued for another 3 houn at 3 0 ' ~ . After centrihigation. bacterial
pellets were resuspended in a 50 mM Na-Hepes pH 8.0, 200 mM NaCI. 0.1 rnM
PMSF. Following lysis of the bacterial pellet (by freeze-thaw) and DNase I treatment
to remove nucleic acids, the fusion protein was purified from the bacterial extract
using giutathione afini ty chromatography as per manufacturer's instructions
(Pharmacia). Elution of fusion protein fiom Glutathione-Agarose column (Sigma)
was performed by a glutathione elution buffer consisting of 10 rnM reduced
glutathione in 50 mM Na-Hepes pH 8.0. To isolate cleaved TGal, thrombin (50 uni&
per mg protein. Pharmacia) was used to proteolytically release TGal from GST
directly on the affinity column as per manufacturer's instructions (Pharmacia).

26
Figure 4. Cloning strategy of the TGAI coostnicts into the pGEX-2TK
expression vector.

Blunt-end TGAI - WTIQ223L PCR produet (-13 Kbp)
-1-1- 'Blunt-endy ligation BamHI into a vector
digested with Smal + - pGEM-TGA I
Vector digested with EcoIWBamHI to release the TGAl
DNA fragment
Biunrn EcoRI Wicky-endY iigation into the expression vector digested with
EcoRUBamEU 1-l- - +
BamHI EcoRI
@EX-2TK The TGAI- Open-reading-frame (- 4.9Kbp)
is cloned in-frame with GST.

2.3 SDS-PAGE and Western Immunoblotting
Proteins were prepared for electrophoresis by resuspension in a 2% SDS
Laemrnli sample buffer ( L a e d i 1970) and boiled for 2 min. Samples were loaded
ont0 a 10% (wlv) linear acrylamide gels. Electrophoresis was carried out at a constant
voltage of 200V for approximately one hour. Molecular weight markers included,
Trypsin inhibitor (2 1,500), Carbonic Anhyârase (3 1 ,O), Ovalburnin (45,000),
Semm Alburnin (66,200), Phosphorylase B (97,400), P-galactosidase (1 16,250) and
Myosin - H chah (200,000). Mer eIectrophoresis, gels were stained with Coomassie
brilliant blue R250 (0.25% in 50% methanol, 7% acetic acid) for 30 min. Overnight
destaining was carried out in 1: 1:8 (v/v/v) methanol/acetic acid/distilled water. For
western immunobloning, unstained SDS-PAGE-separated proteins were
electrophoretically transferred onto nitrocellulose membranes using the Multiphor II
semi-dry blotter (Pharamacia). Eighty mA of current for 45 min was passed through a
stacked 'sandwich' that was placed from the anode to the cathode as follows (i) a
Whatmann 3MM paper soaked in 0.3 M Tris, 20% (vlv) methanol; (ii) a Whatmann
3MM p a p a and the nitrocellulose membrane soaked in 25 rnM Tris, 20% methanol
(vlv) ont0 which the gel was laid; (iii) 2 Whatmann 3MM papers soaked in 25 m .
Tris, 20% rnethanol (v/v), supplemented with 40 mM E - amino -n- caproic acid. To
assess the efficiency of transfer, the nitrocellulose membrane was stained with 0.2%
ponceau S for 2-4 min and rinsed with distilled water. The position of molecular
weight marken was marked with indelible ink.

For western blot analysis, the nitrocellulose membrane was blocked in
Phosphate-Buffered Saline (lxPBST) containing 140 mM NaCl, 2.7 mM K I ,
10 m M Na2HP04, 1.8 rnM KH2P04, 0.1% (vfv) Tween 20 (pH 7.3). and 0.05%
sodium azide, supplemented with 5% fat-free milk (Carnation) for 2 hours at room
temperature. Membranes were then incubated with a primary antibody (anti-GST. at
1: 1000 dilution - Pharmcia / anti-TGal - refer to section 2.5) for 2 hours at room
temperature. The membranes were then washed three t h e s in IxPBST. The
membranes were then incubated with the appropriate allcaline phosphatase-conjugated
secondary antibody (anti -goat/rabbit IgG - Sigma) at room temperature for 1 hour.
The membranes were then washed three tirnes a s described above, and the immune
complexes were detected by adding a mixture containing 0.17 pg per ml nitroblue
tetrazolium (NBT) and 0.33 pg per ml 5-bromo4chloro-3-indolyl-phosphate (BCIP)
in a substrate buffer containing 10 rnM Tris-HCI pH 9.5, 100 rnM NaCl and 10 mM
MgC12, to the nitrocellulose membrane.
2.4 GTP Hydrolysis and Binding Assays
GTPase activity was detemiined as descnbed by Graziano and GiIman (1989).
The reaction mixture included 1 pM [Y-~*P]-GTP (5000 Cilmm01 Amersham) in a
50 rnM Na-Hepes pH 8.0,2 rnM MgS04. 1 mM DïT buffer. Reactions were initiated
by the addition of 20 pmoi (-1 pg) of TGal to a total volume of 500 pl reaction

mixture. Determination of ["pi] released was carried out as descnbed by Brandt et
al. (1983). At designated t h e points, 50 pi aliquots were withdrawn and added to
750 pl of c W e d 5% Norit (wfv) in 50 mM NaH2P04 buffer. Samples were
centrifbged at 2000 rpm in a bench-top centrifuge (Biofuge A - Canlab) for 10 min.
400 pl aiiquots were used for determination of ] in a liquid scintillator counter
(Beckrnan LS600 K).
For GTP binding and hydrolysis assays on nitrocellulose membranes, 2 pg of
purified GST, TGa l wild type and mutant 42231. fusion proteins were separated by
10% SDS-PAGE and transferred to a nitrocellulose membrane. The membrane was
used for a GTP binding and hydrolysis assay as described by Yang et ai. (1993).
Nitrocellulose membranes were washed twice for 10 min in a GTP-binding buffer
containing 20 mM Tris-HC1 pH 8.0, L rnM MgS04. 5 mM DTT, and 0.3% (v/v)
Tween 20. Membranes were then incubated at room temperature for 30 min in the
same buffer as above containing 0.3% (w/v) BSA and 1.0 pCi [$?PI/ [a-32~] -GTP
(5000 Ci/mrnol Arnersham) per ml. Membranes were subsequently washed twice with
the GTP-binding buffer, and exposed to X-Omat autoradiography film (Eastman
Kodak).

2 5 Production of Polyclonal anti-TGal antibodies
Approximately 100 pg of a recombinant TGal fusion protein was emulsified
in Freund's complete adjuvant by repetitive pipetting. The mixture was then injected
into a 3 rnonth old New Zedand White rabbit. Booster injections of the antigen
(100 pg protein) in Freund's incomplete adjuvant were given every two weeks. Fifty
ml blwd sarnples were collected, following the second booster injection. in a period
of 5 weeks. The collected blwd was stored at room temperature for 4 hours and then
centrifuged at 2500 rpm (Beckman GPR) to collect the blood plasma Cross-reactivity
with the recombinant protein was confmed by western blot analysis. Antibodies
were affinity purified using immunoblots as described by Harlow and Lane (1988).
Approximately 250 pg of recombinant TGal was transferred to a nitrocellulose
membrane. The membrane was blocked with 5% miik (wh) in PBST for 2 hours.
10 ml of sera, diluted to IxPBST, was incubated with the blot for 12 hours at 4 ' ~ .
The blot was subsequently washed 5 times with IxPBST, with a final wash in
0.lxPBST. Antibodies were eluted fiom the blot using a 0.2 M glycine-HC1 pH 2.5
buffer, for 15 min at 4 ' ~ . The solution was subsequently neutralized using 2 M Tris
base, to a pH of 6& The titer of the antibody was determined by western blot
analysis. Antisera was aliquoted. and stored at - 8 0 ' ~ .

2.6 Plant Material
Cell suspensions derived from a Line of tomato (Lycopersicon esculentum L.)
cv Moneyrnaker were grown in 500 mi Erlenmeyer flasks containing 120 ml of
Murashige and Skoog medium in the dark at 2 5 ' ~ on a rotary shaker at 120 rpm.
Cells were subcultured into fresh medium weekly. Cells used for al1 experiments were
3 to 4 days old.
2.7 Isolation of Plant Membranes and Cytosol
Microsomal membranes were isolated according to the method of Blumwald
and Poole (1987). Three to four day-old tomato cells (100- 150 g fresh weight) were
collected by vacuum filtration of the cell suspensions ont0 Whatman No.1 filters
using a Buchner funnel. Cells were then homogenized in a Bead-Beater Ce11
Homogenizer (Biospec Products) containing 80 ml of 0.5 mm g l a s beads pre-soaked
in 200 ml of ice-cold homogenization buffer. Homogenization was carried out at 4 ' ~
with 5 pulses of 45 seconds each and 30 seconds rest period in between pulses. The
homogenization buffer consisted of 10% (w/v) glycerol. 0.5% (w/v) BSA. 0.25 mM
dibucaine, 0.5 mM butylated hydroxytoluene, 1 mM PMSF, 5% (wlv) PVP- IO, 5 mM
EGTA, 5 rnM MgS04, 0.25 M mannitol, 2 mM DTT, 26 mM potassium metabisulfate
(&&O5) and 30 mM Tris (adjusted to pH 8.0 with H2S04). The homogenate was
filtered through four layers of cheesecloth and centrifuged at 10.000 x g for 20 min to
remove ce11 debris and mitochondria. The supematant was then centrifuged at
100,000 x g for 45 min in a Beckman type 35 rotor. The cytosol (supematant) was

collected and concentrated in an Amicon ultraFütration unit using a 3 kDa cut-off
füter (YM or low protein-binding membrane) under 10 psi N2. The microsomal pellet
was resuspended in a suspension buffer of 6 rnM Tris-Mes (pH 8.0), 10% (wlv)
giycerol, 250 rnM mannit01 and 2 mM DTT. The microsomal pellet was then layered
ont0 discontinuous sucrose gradients of 9 ml of 16% (wlv) 1 9 ml 34%(w/v) 1 9 ml
38%(w/v) sucrose in suspension buffer. After centrifugation at 100,OO x g for 2 houn
in a Beckman SW 28 rotor, tonoplast, ER and plasma membranes were retrieved from
the sampIe/l6%, 16%/34% and 34%/38% interfaces, respectively. The membranes
were diluted in suspension buffer and pelleted ai 100,00 x g for 45 min in a Beckrnan
Ti 60 rotor.
2.8 Determination of Protein Concentration
Protein content was determined using a dye reagent concentrate (Bio-Rad,
Canada) as described by Bradford (1976). Membrane proteins were first solubilized
by 0.5% (wlw) Triton X-100 for 5 min, before the addition of the dye. BSA was used
as the protein standard for absorbance measurements at 595 nm.
2.9 Measurement of Plasma Membrane ATPase Activities
Plasma membrane ATPase activities were monitored by the release of
inorganic phosphate as described by Ames (1966). Plasma membrane vesicles (25 pg
protein) were incubated in 0.5 ml buffer 30 mM Tris-Mes (pH 6.5) of 3 mM Tris-

ATP (pH 6.5),3 rnM MgS04. 50 mM KCI, containing LOO pM sodium molybdate - to
inhibit non-specific phosphatase contamination fiom other membrane fractions,
10 mM sodium azide - to inhibit rnitochondrial p ~ T P a s e and 5 pM gramiciciin-D -
to ensure the breakdown of the proton gradient that could stall the FI+-pump activity.
Measurernents of ATPase activities were carried out in response to mastoparan (1-
1Op.M) or GïFyS (100 pM). Okadaic acid, when used, was dissolved in DMSO and
used at 5 pM. The reaction medium was incubated at room temperature for 30 min.
The reaction was stopped by adding the Ames solution (1 voi 10% ascorbic acid and 6
vol 0.42% ammonium molybdate in H2S04), and the absorbante was measured at 820
nm.
2.10 Synthetic Peptides
Imrnunological-grade custom peptides were synthesized by Genemed
S ynthesis Inc. (San Francisco, CA). Two peptides were used in the experiments - (i) a
peptide corresponding to the last 11 amino acids of the TGal C-terminus -
N - RRRNLFEAGLL - C; and (ii) a control peptide, consisting of the same amino
acid composition with a randomized sequence - N - NAEUGRFLREL - C. For the
measurement of plasma membrane ATPase activities, in response to mastoparan (2
IrM) in the presence of the peptides, the membranes were pre-incubated for 10 min at
room temperature with 10 ph4 of the peptides, before adding the rest of the reaction
mixture. Detection of Pi released was perfonned as described above (section 2.9).

2.1 1 Immunologicall y-Based Interactive Cloning
cDNA clones for TGal-interacting proteins were isolated by a method similar
to the one described by Chapline et al. (1993). A ZAP Express cDNA library
(Stratagene) was used. constmcted from reverse-iranscribed mRNA, isolated from 4
day-old tomato suspension culture cells carrying the Cf5 resistance gene, infected for
1.5 hours with intracellu1 ar fiuids of leaf tissue infected with Clodosporium filvum
funys race 4. IPTG-induced proteins were immobilized on nitrocellulose lifts. The
nitrocellulose lifts were blocked with 5% non-fat dry milk, and then incubated with
10 p g h l recombinant TGal. Incubation was for 6 hours at room temperature in Tris-
buffer saline (1 x TBS), 30 mM Tris-HCI (pH 7.5), 150 rnM NaCI. After 3 washes in
1 x TBS, bound TGal was cross-linked by incubating the filters in 1 % fonnaldehyde.
Excess fomaldehyde was inactivated with a 2% glycine wash. After brief washing
with 1 x PBST, the ni~ocellulose lifts were incubated with anti-TGal polyclonal
antibody ( 1 : 1000 dilution) in 1 x PBST supplemented with 1 Q non-fat dry rnik.
Positive clones were identified after incubation with an anû-rabbit IgG-aikaline
phosphatase-conjugated secondary antibody and after colour developmeot (as
described in section 2.5). in the secondary screen, putative positives were examined to
determine whether they directly interacied with the anti-TGal antibody. Those that
did not, were carried through to a tertiary interaction screen, as desclibed above
(Fig.5). Positive clones were plaque-purified and in vivo excised, and the excised
phagemid was infected into the E-coli XLOLR strain as per manufacturer's

36
instructions (Stratagene). Phagemid DNA @BK-CMV) was purified using the Wizard
Plus Miniprep DNA purification system (Promega). Insert clones were sequenced
from their 5' - end using a T3 primer by cycle sequencing. with dye-labelled dideoxy
c hain terminators, at the York University core rnolecular biology facili ty.

Figure S. Strategy for an immunologically-based interactive cloning screen.

Plate tomato cDNA library Overlay PTG-soaked NC filter to induce expression
incubation PRIlMARY s c m
Detect with anti- TGal antibody
Lift NC tilters and use for screening. Add TGa 1 -WT/Q223L (6 hours)
@y3 4-1 Wash and cross-link
SECONDARY SCREEN
F a k ImmunologicaU y Putative positive related interaction
+ = incubated with TGal; - = without TGal

RESULTS
3.1 Overexpression and Purifkation of Recombinant TGal Proteins
The putative open-reading-he of the TGAl gene product cloned into the
pGEX-2TK expression vector was used to transfonn E.coli BL2I pLysS ceus. To
facilitate an optimal yield of purification, the conditions for IPTG - induction of
bacterial expression, were assessed. Special consideration was given to the
temperature of bacteriai growth. and to the PTG concentration used. Total protein
proNes from bactena induced under the different experimental conditions, carrying
the pGEX-TGAI construct versus a conaol condition using E.coli carrying the pGEX
vector alone, were compared by SDS-PAGE (Fig.6). A -66 kDa product, visible by
Coomassie blue staining, was present in bacteria carrying the pGEX-TGAI construct
but not in the controt. To optimize the expression of the 66 kDa product, varying
concentrations of IPTG, ranging from 10 pM to LOO pM. were used to induce bacteria
entenng log phase growth. In addition, different incubation times foilowing IPTG
induction of protein synthesis (2-3 hours) were assessed. The temperature was kept at
3 0 ' ~ to allow longer induction times. Optimal expression was observed upon
induction, of bactena entering log phase, with 50 pM F ï G for 3 hours (Fig.6).
Similar conditions were used for the expression of TGAl-Q223L.
The pGEX-TGAI constmct was designed to express TGal as a GST
(Glutathione-S-transferase)-fusion protein with GST at its N-terminal end. The

40
Figure 6. Coomassie-blue stained 10% SDS-PAGE of total proteins fiom
bacteria canying the pGEX-TGAI expression vector under different expression
conditions. Lane 1 is the negative control lane with total proteins from bacteria
carrying the pGEX-2TK vector alone. Expression conditions were as follows: Lanes
2-5 - induction with IPTG for 2 hours; lanes 6-9 - induction for 3 hours. Lanes 2 and
6 - 10 p M IPTG; lanes4
and 8 - 50 pM IPTG; lanes 5 and 9 - 100 pM IPTG. The arrow points at the 66 kDa
band that appeared with varying intensities under the different expression conditions,
suggesting it to be the expected fusion protein.
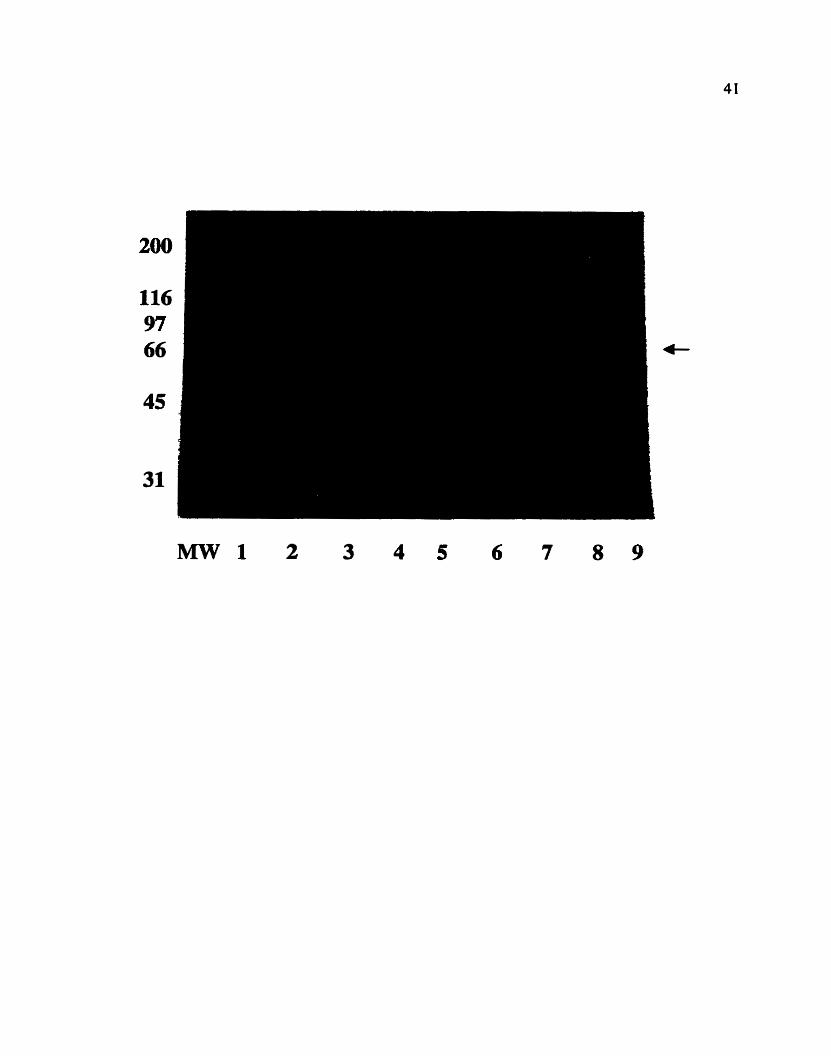

expected open-reading-frame of the TGAI gene product has a predicted molecular
weight of 45 kDa. GST is a 29kDa protein. As the product detected on a 10% SDS-
PAGE was 66 kDa and the expected GST-fusion protein size is 45+29 = 74 kDa, it
was necessaiy to determine whether the 66 kDa product was indeed that fusion
protein. A western blot using an anti-GST antibody on lysates frorn induced bactena
was perforrned. As expected, the antibody detected a 29 kDa product in IPTG-induced
bactena canying the pGEX vector, coding for GST. In addition, the 66 kDa product
was detected by the antibody, confirrning it to be the GST-TGal fusion protein
(Fig.7). The discrepancy in expected size is not of concem because the 10% SDS-
PAGE is not optimized for detection of proteins at this range of molecular weights.
Since GST was included in the gel as a positive control, lower percentage gels were
not used.
Having a protein fused to GST, dows for its purification by affinity
chromatography on a glutathione column. Furthermore, the pGEX-2TK vector was
designed to contain a thrombin-specific cleavage site between GST and the desired
product. This allows for fast and simple purification of the fusion protein from
bacterial lysates as weli as the subsequent release of the desired product from GST.
Purification of wild type and mutant TGAI gene products, transformed with the
pGEX-TGAI constructs, was optimized, resulting in yields of 1-1.5 mg of GST-
fusion protein per litre of bacterial culture. Thrombin cleavage directly on the column,

Figure 7. (a) A western immunoblot using a . anti-GST antibody to identify
the GST-TGal fusion protein in total proteins from bacteria carrying the pGEX-
TGAl expression vector. Lane 1 is the positive control lane with bactena carrying the
pGEX-2TK vector alone. Lane 2 - bacteria carrying the @EX-TGAI -WT vector.
Lane 3 - bacteria carrying the pGEX-TGAIQ223L vector. Induction of expression
was carried out in the same manner for dl bacteria using 50 pM IPTG for 3 hours.
(b) The corresponding Coomassie-blue stained 10% SDS-PAGE of total bacterial
proteins as described in (a). The arrow points at the 66 kDa protein that is present
oniy in lanes 2 and 3 and corresponds to the hision protein detected by the anti-GST
antibody.


generating the recombinant protein lacking the GST fragment, resulted in yields of
0.5- 1 mg per litre (Fig.8a).
Figure 8(b), is the sequence alignmeat of TGal, compared to other known G a
subunits, within a conserved region in which mutations were observed to have
significant effects on GTP binding and hydrolysis (Ma et al., 1991). Specifically, the
substitution of the conserved glutamine residue (4223 in TGal) to a leocine, was
observed to significantly reduce the Kcat of GTP hydrolysis in other G-proteins
(Graziano and Gilman; 1989;Masters et al., 1989). The effect of this mutation is the
constitutive activation of the G-protein a subunit. Since G-proteins are naturally
found in an inactive (GDP-bound) state, to facilitate the biochernical characterization
of TGal, the same mutation was introduced into its sequence and its effects were
studied.
3 3 GTP Hydrolysis and Binding by TGal
The predicted TGal sequence showed high homology to known a subunits of
the heterotrimeric G-proteins. GTP binding proteins have an exclusive high &nity
and specificity to guanine nucleotides. To asses whether TGal is indeed a G protein,
as suggested by its primary sequence, both its GTP hydrolytic activity and GTP-
binding capacity were examined.
Using labeled the enzyme-dependent GTP hydrolysis was
measured by monitoring [32~i] released in the reaction mixture. As seen in figure 9(a),

Figure 8. Purification of recombinant GST-TGal (WTIQ223L) and
TGal (WTlQ223L) by glutathione afflnity. (a) A Coornassie-blue stained 10% SDS-
PAGE showing the purification of the different gene products fkom total bacterial
proteins as follows: GST - 1 and 4; GST-TGal -WT - 2 and 5; GST-TGaLQ223L -
3 and 6. Lanes 7 and 8 are the released TGal-WT and TGabQ223L products
respectively, following thrombin proteolysis to remove GST from the fusion proteins.
(b) Sequence alignment of the deduced TGal amino acid sequence, with other known
a subunits. The conserved Gln residue (223) was mutated to Leu in TGotl. Identical
residues are coloured in blue.

220 & 230 240
KSGEVYRLFD VGGQRNERRK WIHLFEGVTA KSGEWRLFD VGGURNERRK WIHLFEGVTA FKELTFKMVD VGGQRSERKK WIHCFEGVTA VDKVNFHNFD VGGQRDERRK - WlQCFNDVTA

Figure 9. GTP hydrolysis assays using [ y 3 2 ~ 1 ~ ~ ~ to monitor [-'*pi]
released by recombinant TGal -WT and TGaLQ223L enzymes. (a) Enzyme-
dependent hydrolysis was monitored by increasing the concentration of TGal-WT
(I) in the reaction mixture from lû-100 pmol in a total volume of 0.5 ml. GST (e)
was used as a negative control. [)'pi] released was measured (as described in
materials and methods) at t= 30 min (b) Time-course experiments were carried out
using 10 pmol of either TGal-WT (*) or TGaLQ223L (m. Time points were t a ,
5, 15. 30 min. Al1 values are representative of three independent experiments carried
out in triplkates (n=3).

Enzyme-Dependent GTP Hydrolysis
O 50 100
pmole Enzyme 1
Determining Kcat of Hydrolysis
I
Q223L Mutant
O 10 20 30
T lme (min)

increasing concentrations of TGal ( 10- 100 prnoles) resulted in increased [ 3 2 ~ i ]
release. Purified GST was used as a negative control indicating that no contaminating
bacterial G-proteins were purified by glutathione affinity from bacterial extracts.
Hydrolysis was completely dependent on the presence of MC ions. When 5 mM
EDTA was included, and no M ~ Z C was present in the reaction mixture, no hydrolysis
was observed. In addition. no significant hydrolysis was detected when using [y-
3 2 ~ ] ~ ~ ~ as an alternative nucleotide. indicating a guanine nucleotide specificity. as
expected for a G-protein.
The intrinsic rate of GTP hydrolysis by TGal was measured by monitoring
['*pi] release in a time course experiment (Fig.9b.). The rate of hydrolysis in these
experiments was 0.075-0.125 mol Pi mol min-' ( 1.5-2.5 prnoles Pi pg
protein'' min-') The rate of hydrolysis of GTP for the TGal-Q223L mutant was
negiigible within the time course of the expenment (Fig.9b.).
Lack of hydrolysis by the Q223L mutant could either be due to its inability to
hydrolyze GTP or to a significantly reduced affinity for GTP. In order to characterize
the mutation. GTP binding and hydrolysis assays were carried out on nitrocellulose
membranes to which the proteins were transferred. The membranes were incubated
with GTP labeled either at the a or y phosphate. Although the proteins were subjected
to denaturing conditions by SDS-PAGE, a fraction of the G-protein may renature,
and specifically bind andlor hydrolyze GTP. The resulting autoradiogram of the
membrane allowed for the visualkation of the assay.

51
Approximately equal amounts (2 pg) of GST, TGal-WT and TGal-Q223L
GST fusions, were used in the experiments, a s seen in figure 1 O a showing the
Coomassie blue - stained gel after SDS-PAGE. In the hydrolysis of GTP. it is the y
phosphate that is released while the a phosphate remains in the resulting GDP
molecule. Figure lob. is an autoradiogram of a nitrocellulose membrane blotted with
the three GST-fusion proteins, incubated with, [ a - 3 2 ~ ] ~ ~ ~ . in this assay, binding of
GTP was observed, irrespective of the rate of hydrolysis by the G-protein. The GST
lane served as a negative control, ensuring that no contaminating bacterial G-proteins
were present after glutathione purification. A 66 kDa labeled-protein appeared both in
the TGal-WT and TGal-Q223L lanes, corresponding to the size of the purified
fusion proteins. The intensity of both labeled proteins was comparable to the
Coomassie-stained proteins in fig.lO(a), suggesting sirnilar affhity to GTP by both
proteins. Figure lO(c). is an autoradiogram of a nitrocellulose membrane incubated
with, [ y - 3 2 ~ ] ~ ~ ~ . The TGaI-WT lane showed a 66 kDa of a faint-labeled protein
whiie the TGaLQ223L lane showed a labeled-protein appearing with greater
intensity, indicating that GTP was stiil bound to the enzyme. As expected, no labeled-
proteins appeared in the GST lane in both experiments.

Figure 10. GTP binding and hydrolysis assays on enzymes transfemd to
nitrocellulose membranes. (a) A Coornassie-blue stained lO%SDS-PAGE run with
2 pg of I - GST; 2 - TGal-WT; 3 - TGaLQ223L. (b) Proteins (as in a.) were
transferred to a nitrocellulose membrane which was incubated with [a-"P]GTP for 30
min (as described in materials and methods). (c) As in (b), but with [Y-~~P]GTP.
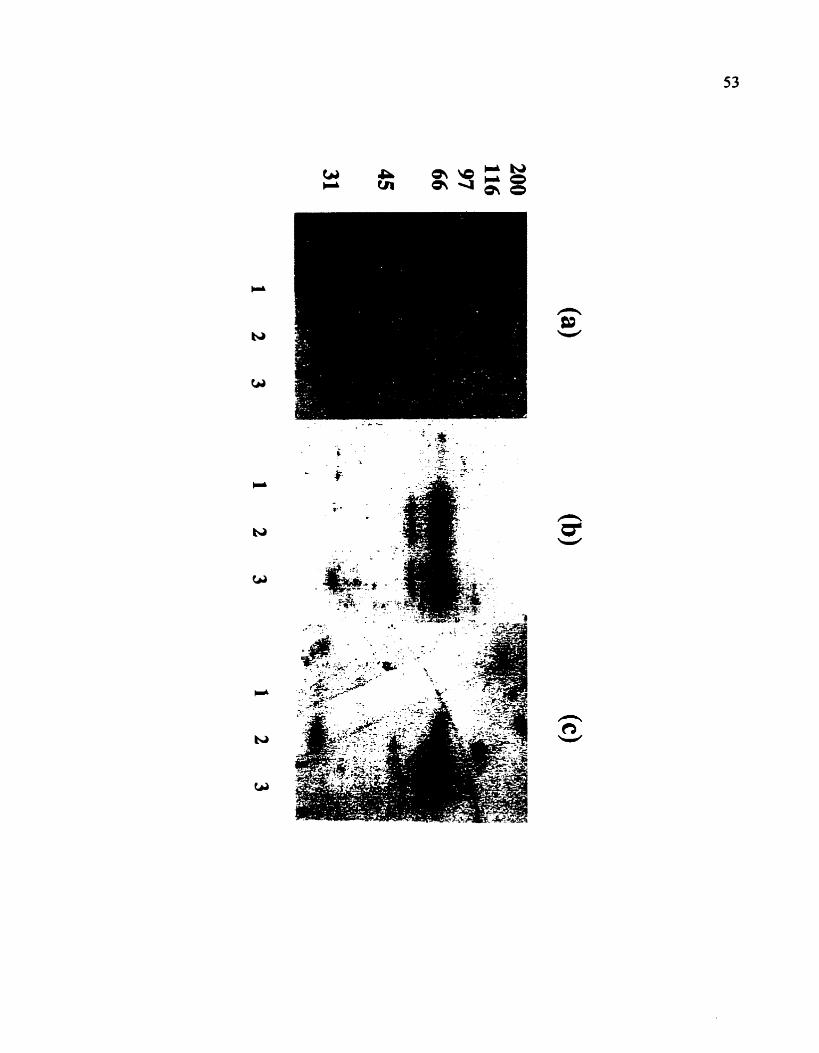

3 3 Immunological Detection and Subcellular Localization of TGal
Polyclonal antibodies were afin@-purified from the serum of a rabbit
injected with recombinant TGal. To determine the subceiiular location of the protein.
Immunoblot analysis, using different subcellular fractions from tomato suspension
cells, was performed. Fig. 1 1 shows a western blot. using the anti-TGal antibody. of
various enriched ce11 membranes as well as cytosol (30 pg eacch), separated by SDS-
PAGE. As seen, a protein of a molecular mass of 45 kDa, corresponding to the size of
the recombinant TGal, was strongly detected in the microsornal membranes, plasma
membrane and endoplasrnic reticulum fractions. A faint irnrnuno-reactive protein was
visible in the tonoplast lane, possibly due to contamination from other membrane
fractions during the purification procedure. No irnmuno-reactive proteins were
detected in the cytosol (Fig. 1 1).
3.4 Characterization of a Putative G-Protein-Coupled Receptor Contact
In mammalian systerns, heterotrimenc G-proteins are known to be coupled to
membrane receptors of the seven-transmembrane-spanning family. These receptors
are involved in regulating the activation of G-proteins through protein-protein
interactions. Genetic, biochemical (Conklin et al., 1993) and X-ray crystallography
studies (Lambright et ai., 1996) have implicated the C-terminal of the G-protein a
subunits as a site for receptor - G-protein contact. The C-terminal is characterized by
a short stretch (-10-15) of hydrophobie amino acids that tend to form an a helical

Figure 11. A western Immunoblot using an affinity-purified anti-TGal
polyclond antibody. (a) A Cwmassie-bIue stained 10% SDS-PAGE of membrane
proteins purified by sucrose gradient and concentrateci cytosol. 30 pg of each fraction
was loaded. -100 ng of pure TGal was used as a positive controi (b) Immunoiogical
detection of TGal in the different subcellular hctions.


secondary structure (Sullivan et ai. 1987). To examine the possibility that the TGal
C-terminus is involved in receptor contact, a number of sequence analyses were
carried out. Figure 12(a) is the hydropathy profile (Kyte Doolittle) of the deduced
amino acid sequence of TGal. The enzyme is predicted to predorninately be
hydrophilic. This prediction is supported by its presence in the soluble fraction of
bacterial lysates. upon its expression as a recombinant enzyme (Fig.6). The last
stretch of 11 amino acids at the C-terminus of TGal showed an overall stronger
hydrophobic character. Figure 12@), is a secondaq structure prediction (Chou
Fasman) based on primary sequence analysis. Although only suggestive, it predicts
the secondary structure with the highest probability for a stretch of amino acids. based
on known protein structures. This analysis predicts an a helical structure at the C-
terminus of TGal. These two analyses have identified an l l amino acid stretch at the
C-terminus as having a hydrophobic character with a tendency to form an a-helix.
Fig. 12(c), is an alignment of the C-terminal sequences from different a subunits
including two plant homologues. The 11 amino acid C-terminal sequence of TGal
and GPal (Arabidopsis) are 91% identicai while Gs and G, are only 36% identicai to
the plant homologues. All sequences contain hydrophobic residues, with a high
probability to form a-helicai secondary structures.
Synthetic peptides, correspondhg to the C-terminal sequence of TGal, were
used to uncouple, in competitive inhibition assays, the putative receptor - G protein
contact. Uncoupling means the inhibition of signal mediatioa hom the receptor to the

Figure 12. Identification of a potential receptor contact region in the C-
terminal sequence of TGal. (a) The hycûopathy profile of the predicted TGal
sequence (Kyte Doolittie). (b) The prediaed secondary structure of the TGal
sequence (Chou Fasman). ( c ) Sequence a l i m e n t of the C-terminus of TGal
compared to other known a subunits. Identical residues were put in bold.

Residue
TGal LVKKTFKLVD E RRRNLFE AGLL GPal LVKKTFKLVD E RRRNLLE AGLL
Gt NIQFVFDAVT IQNNLKY IGLC
Putative Receptor Contact

G-protein molecule. As a result. G-protein activation does not occur in response to an
agonist, and downstrearn elements in the signal transduction pathway do not respond.
As there is no direct evidence to the role of G-proteins in the action of any plant
agonists, the mastoparan peptide was used instead. As descnbed before, mastoparan
in the presence of a hydrophobic environment forms a structure rnirnicking the
activated third intracellular loop of 7TMS receptors. In addition, mastoparan was
shown to directly interact with the C-terminal of some a subunits. In the plasma
membrane of tomato cells, there is a G-protein regulated w - ~ T P a s e pump (Vera
Estrella et al.. 1994; Xing et al., 1997). In addition. these studies have shown that the
activation of the proton pump is dependent on a dephosphorylation event, mediated
by a membrane associated phosphatase. Figure 13(a), shows the measurement of
plasma membrane ATPase activity in response to increasing concentrations of
mastoparan (0-10 pM). In the absence of rnastoparan, the basai ATPase activity in
the plasma membrane was 11 m o l Pi mg-' x h-'. Maximal ATPase activation of 32
jmol Pi mg-' x h-' (2 10% of control) was observed when 10 jM mastoparan was
used. Figure 13(a) includes the ATPase activities in response to mastoparan, pre-
incubated with 5 pM of the phosphatase inhibitor - okadaic acid. Inhibition of activity
was almost complete when lower concentrations of mastoparan were used (1-2.5
pM). At 5 p M mastoparan inhibition by okadaic acid was 79% while at 10 jM
mastoparan only 57% inhibition was seen. We have therefore chosen 2.5 p M
mastoparan in ai l subsequent experiments - a concentration that is within the linear

Figure 13. Inhibition of mastoparan-stimulated activation of the plasma
membrane H+-ATPase by synthetic peptides, corresponding to the TGal C-terminus
sequence. (a) Concentration-dependent activation of the plasma-membrane
H'-AT~ase by mastoparan (+) using concentrations of 1-10 pM, and its inhibition
by 5 pM okadaic acid (m). Pi was determined (as described in materials and
methods) at t=30 min. (b) Effect of pre-incubation of plasma membrane with 10
of the synthetic peptides, corresponding io the C-terminus of TGal, in response to 2.5
pM mastoparan. GTPyS was used at 100 pM.


activation of the ATPase activity and is almost completely inhibited by 5 pM okadaic
acid.
Figure 13(b), is a graph showing the effects of pre-incubation of plasma
membrane with synthetic peptides corresponding to the C-terminus of TGal prior to
the addition of mastoparan. Both the addition of 2.5 p M mastoparan and
100 pM GTPyS resulted in -7û-75% stimulation of the plasma membrane ATPase
activity. Pn-incubation with 10 p M of Ci i-TGal (The TGal - C-terniinus 1 1 amino
acids) resulted in 87% inhibition of the rnastoparan-stimulated activity. Pre-
incubation with 10 pM Cil-Con (the control peptide with the same arnino acid
composition in a randomized order) did not result in a significant inhibition of plasma
membrane stimulated ATPase activity.
3.5 Screening for TGal Interacting Proteins
Approximately 2~1d ph, of a tomato cDNA expression library, were used to
screen for TGal interacting proteins. Immunological detection, using the anti-TGal
antibody, after incubation with recombinant TGal, identified 11 potential positives
which were carried thmugh to a secondary screen. Three of the positives cross-reacted
with the antibody alone, and therefore, do not represent binding proteins. Four did not
reappear, and therefore were considered false positives. Four positives GIP4- 1, GIP6-
2, GIP8- 1 and GIP 9-1, did not cross-reacting with the antibody alone, yet did cross-
react if they were pre-incubated with recombinant TGal. The plaques, corresponding

to these putative interacting proteins, were isolated, excised and partially sequenced
h m their 5 ' end. Ail DNA sequences were <ranslated to identiS, the possible ORF's
of the cDNA's. Cornparison of the predicted amino acid sequence of dl four proteins
GP4- 1, 6-2, 8- 1 and 9- 1 reveaied them al1 to code for the same sequence. Figure 14.
is the sequence alignment of the predicted amino acid sequence of the four cDNA1s
isolated. Both GIP4-1 and 6-2 appeared to code for amino acid residues downstream
of the coding region of GIP8-1 and 9-1, indicating that they lacked a 5' end DNA
sequence present in the other cDNA's. As a result, both GIP4-1 and 6-2 provided an
extended 3' DNA sequence which allowed the identification of additional
downstream amino acid residues. This compiled sequence of al1 four proteins has
therefore allowed for the identification of 205 amino acids in totd, that were
homologous in at l es t two of the reported sequences. This combined sequence was
subsequently used in searching the GeneBank (NCBI) databases for sequence
sirnilarities. A 'BLAST' search revealed high homology, of the deduced amino acid
sequence, to a family of proteins known as the Dnd-like proteins . Figure 15 is the
homology alignment of the compiled 205 amino acids of the TGal-interacting protein
(designated LEJI for Lycopersicon esculentwn L.) with other known Dnd-like
sequences. As the four interacting proteins were only partially sequenced from their
5' end, the homology to the known proteins was only within their C-terminal
sequences as follows: 56% identical and 7 1% similar (with conserved changes) to
RDJZ (Rattus norvegicus); 54% identical and 70% similar to STJ3 (Solarium
tuberosum); and 52% identical, 70%

Figure 14. Sequence alignrnent of the predicted amino acids (from the partial
5' DNA sequences) of the four proteins identified as interacting with TGaI - GIP4- 1.
6-2,8- 1 and 9- 1 . Biack boxes indicate identical residues,


67
similar to ATJ3 (Arabidopsis thliana) (Fig. 15). Because of the extensive homology
at the C-terminal sequences, it is possible that the methionine residue at position 5 in
the compiled sequence represents the start codon of the DNA sequence. If that is the
case, both GP8-1 and 9-1 represent the full-length clone of that Dnd-like protein in
tomato.

Figure 15. Sequence alignment of the compiled 205 arnino acids (from al1
four TGal-interacting proteins) with known Dnd-Wre proteins. The tomato clone
(designated LUI) was : 56% identical and 7 1% sirnilar (with conserved changes) to
RDJ2 (Raiîus norvegicus); 54% identicai and 70% similar to STJ3 (Solarium
tuberosum); and 52% identical. 70% similar to ATJ3 (Arabidopsis thalinna) within
the C-terminal regions. Black boxes indicate identical residues, while grey boxes
indicate homology with conserved changes.

U U W W U W 'OP-

DISCUSSION
4.1 Purification of Recombinant T k l
The tornato TGAl cDNA has a predicted amino acid sequence showing
significant homology to a G-protein a subunit. The DNA sequence, coding for the
open-reading-frame, was cloned into an expression vector to facilitate over-
expression and purification of the recombinant protein from bacterid cells.
Purification of the recombinant enzyme was achieved at high yields (0.5- 1.5 mg / L of
bacterial culture), allowing for its biochernical characterization, as well as use as an
antigen for the generation of antibodies. The recombinant product, after thrombin
proteolysis to release the GST fragment from the TGal fusion protein, resulted in a
45 kDa protein (Fig.8a). This f d s within the 35-45 kDa size range of known a
subunits from animais (Ma, 1994), and corresponds to the approximate size of
immunologically-related polypeptides detected in plant membranes (Blum et al.,
1988; Jacobs et al.. 1988; Legendre et al., 1992; Weiss et ai., 1997; Xing et ai., 1997).
4.2 GTP Hydrolysis by TGal
To examine whether TGal is indeed a GTP - binding protein, radiolabelled
GTP was used to detect enzyme-dependent GTPase activity. While increasing
concentrations of TGal (10-100 pmol) resulted in increased release of L~'P~] in the

7 1
presence of [I '32~]~TP (Fig-9a), none was released when [ y - 3 2 ~ ] ~ ~ ~ was used as an
alternative nucleotide. This guanine-nucleotide specificity is characteristic of GTP-
binding proteins and generally, alternative nucleotides cannot compte for the GTP
binding site, even at high concentrations (Yang et alJ993). GTP hydrolysis was
completely dependent on the presence of magnesium ions in the reaction mixture. The
role of magnesium ions in the activity of G-protein a subunits was correlated with a
number of events. These include; the activation of hoio- G-protein, Le. dissociation of
the a& heterotrimer, as well as GTP bindllig and hydrolysis. GTP binding
experiments using [ 3 S ~ ] ~ T P y S demonstrated the formation of an extremely stable
GTPyS -magnesium complex which did not dissociate even if kee magnesium was
removed by the addition of EDTA (Higashijima et al., 1987; Slepak et al., 1993).
Tirnecourse experiments, for the determination of K,, of GTP hydrolysis for
TGal, provided an estimate of 0.0754.125 moles Pi mol protein-' min-' (Fig.9b).
This rate of hydrolysis is lower than the rate described for most other mammalian a
subunits. The K, reported for a recombinant Gsa was 3.5 min-' (Graziano and
Gilman, 1989); 2.2 min-' for Goa (linder et al., 1990) and 2.4 min' for Gia (Linder
et al., 1990). Within the K, range of TGal is Gza at 0.05 min-' (Casey et al., 1990).
Interestingly, the two recombinant rice a subunits were also reported to have slower
rates of hydrolysis than mammalian G-proteins. Seo et al. (1997) reported the Kcat of
hydrolysis for the recombinant product of RGAI to be 0.0075 min-'. Iwasaki et al.
(1997) characterized rGricea, and detennined its Kcat tu be 0.44 min*'. The different

enzymatic characteristics arnong the mammalian a subunits in their GTPase
activities, seems to be due to differences in their signal transduction roles. The value
of l/K,, represents the time that an a subunit remains active in a steady-state
reaction. The recombinant TGal would bind GTP in a scde of minutes, while most
mammalian a subunits would remain active for 10-20 seconds. There is no direct
evidence to conclude that the in vitro biochemical characteristics of the recombinant
TGal reflect those of the natural a subunit in higher plants. In the case of the
mammalian a subunits, the interaction of guanine nucleotides with the recombinant
products was essentially the sarne as that of their natural counterparts. It would
therefore not be surpnsing if the same principle held in plants. Nonetheless, one has
to consider that under in vivo physiological conditions some of these characteristics
rnay be altered. A recombinant ras- srna11 G-protein. has a reported Kcul of 0.02 min-'.
Yet, in the presence of a GTPase activating protein (GAP) its K,, increases to values
of >1 min" (Masten et al., 1989). Attenuation of the GTPase activity, is not
exclusive to the small (monomeric) G-proteins. Protein regulators of heterotrimeric
G-protein signaling, refemd to as RGS's, were shown to be negative regulators of
signal transduction. In vitro experiments with purified RGS proteins established their
capacity to accelerate the GTPase activity of certain G-protein a subunits (Dohlman
and Thomer, 1997; Huang et al., 1997; Wang et al., 1997). In addition. there is
growing evidence supporting the role of the G-protein eflector molecules in
stimulating GTPase activity, such as the case of the cGMP-phosphodiesterase for Gt

a in the visual system (reviewed by Fields and Casey, 1997). Similar attenuation
mechanisms may well be present in plants to regulate heterotrimeric G-pmtein
activation.
As reported by Weiss et al. (1993), the locdization pattern of the Arabidopsis
GPal homologue is quite compiex in various tissues and amounts throughout
development. It was found to be most abundant in immature tissue. Its abundance in
rapidly dividing, actively differentiating tissue would indicate its requirement, and
therefore a need to keep GPal active for longer periods. This requirement could
correlate with the slower rates of GTP hydrolysis observed for TGal. Interestingly. a
G-protein subfamily, designated G12, which was observed to stimulate cell growth,
was reported to have K, values at the 0.1-0.2 min-' range (Offermanns and Schultz.
1994; Kozasa and Gilrnan, 1995)- comparable to that of TGal. It is therefore
plausible that prolonged activation of G-proteins, by having an intrinsically low Kea,,
is a developmental requirement in actively dividing tissues. In more mature organs,
where GPal was observed to be less abundant, it may serve different functions.
Thus, its GTPase activity. if required, may be attenuated by RGS molecules or by the
presence of effector molecules specific to that tissue.

4.3 Characterization of the TGal-Q223L Mutant
No significant release of rJ2pi] could be detected in the GTP hydrolysis assay
for the TGal-Q223L mutant (Fig.9b). Its Kcar of GTP hydrolysis was therefore not
estimated. GTP binding and hydrol ysis assays on nitrocellulose membranes using
GTP radiolabelled at the a or y phosphate were carried out in order to characterize the
effect of the arnino acid substitution on substrate binding. Binding was assessed by
incubating both wild-type and mutant TGal with [CC-~~P]G'TP. which would retain its
3 2 ~ isotope even upon GTP hydrolysis to GDP. For the hydrolysis assay, [Y-~*P]GTP
was used. From these assays, the effect of the Q223L substitution on TGal is
apparent. While binding of GTP was equivaient for both enzymes fig.lOb),
hydrolysis of GTP by the wild type protein was much more significant than by the
mutant (Fig. lûc). These results demonstrate that TGabQ223L is a GTPase-defcient
enzyme but it is not deficient in its GTP binding capacity. This mutation would
therefore render the enzyme, once bound by GTP. constitutively in the active state.
The same substitution in the cognate regions of other G-proteins was observed
to result in a >LOO fold reduction in the K, for GTP hydrolysis (Graziano and
Gilman, 1989; Masters et al., 1989). Drawing h m the crystal structure of p21".
Masters et ai. (1989) proposed that the Q-tL substitution in Gsa affects a 'switch'
mechanism that mediates the conformational transition between the GTP- and GDP-
bound forms of the enzyme. The location of the glutamine residue in p21rm is not
involved in direct contact with the phosphoryls of GDP. The authon therefore

hypothesized that GTP hydrolysis is slowed because the Q+L substitution slows or
prevents transition h m the GTP- to the GDP-bound conformation, but not because
the mutation directly alters the rate of GTP hydrolysis. The recent crystallization of
G p in a heterovimeric complex (Lambnght et al. 1996) demonstrated the
conformational changes in Gta upon its activation. The conserved glycine residue,
which is adjacent to the glutamine (Gly222 versus Gln223 in TGal, Fig.8b), was seen
to interact with the y phosphate of GTP, triggering the 'active' conformational
change. Thus, the role of the glutamine residue is in stabitizing the transition state to a
GTP hydrolysis mode. These observations seem to confirm the proposed mode1 by
Masters et al. (1989).
4.4 Subcellular Localization of TGal
Western blot analysis of difierent subcellular fractions from plant suspension
cultures indicates that TGal is predominantly associated with the plasma membrane
and ER (Fig. 11). The appearance of a faint immuno-reactive product in the tonoplast
is most probably due to contarninating membranes, but the presence of TGal in the
tonoplast fraction, in lower abundance. cannot be ruled out by these experiments. The
ER fraction likely consists of both the rough and smooth ER. as well as of significant
Golgi membranes. Therefore, the precise location of TGal in these intracellular
structures cannot be determined from these expenrnents. No signal was observed in
the cytosol, indicating the TGal is exclusively membrane associated, as would be

expected h m a heterotrimeric G-protein. These results are consistent with the
observations of Weiss et al. ( 1997) for the Arabidopsis GPal homologue. Subceliular
localization of GPal in meristematic cells of ArabUiopsis and cauliflower iadicated
its presence in both the plasma membrane and ER. The ER fraction was M e r
separated into the rough and smwth components and examined by western blot
analysis. GPal was found to be associated with both ER membrane fractions-
Western blot analysis of isolated Golgi apparatus indicatd that GPal may associate
with the Golgi membranes, but to much lower level than with the plasma and ER
membranes. The antibody detection of TGal. and the results presented by Weiss et al.
( 1997), would seem to suggest a role for TGal (and its GPal homologue) at the
plasma membrane and the ER.
4.5 Possible roles for TGal in the ER
In the past few years, animal heterotrimeric G-proteins have been found to be
associated not only with the plasma membrane, but also with endomembranes such
as the ER (Audigier et al., 1988) and the Golgi complex (Vries et ai, 1995). However,
the role of the heterotrimeric G-proteins in the endomembranes is still largely
unknown. The data available is mostly derived from the use of basic biochemical
tools such as GïF analogues, FTX and known dmgs, suggesting a role in the
secretory pathway (Stow and de Almeida, 1993; Yamaguchi et ai., 1997). More direct
evidence was produced by the observation that overexpression of G ~ L , (which

localizes to the Golgi membranes) in kidney cells inhibited constitutive secretion.
These experiments suggested that G w plays an inhibitory role in the secretory
pathway (Stow et ai., 1991). In contrat, srnall G-proteins, and specifically the Rab
family, have been widely implicated in regulating the transport of proteins fiom the
ER to the Golgi complex, the formation of vesicles within the Golgi complex. the
formation of endocytic and transcytotic vesicles, as weil as endosorne fusion
(reviewed by Nuoffer and Balch, 1994).
By analogy with animal G-proteins, it is conceivable that TGal is involved in
regulating vesicle transport. It may well be involved in regulating other processes of
the ER such as protein synthesis or protein modification or possibly membrane
trafficking between both the plasma membrane and the ER. The relatively low Km of
GTP hydrolysis reported here for TGai, indicates that it has the capacity to be active
for longer tirnes. That, and the abundance of GPal in immature, developing tissues,
could support the notion of TGal k i n g involved in long distance cellular transport of
vesicles (Weiss et al., 1993). These type of cellular processes would be at their
highest peak in the early developmental stages of organs, coinciding with the high
levels of GPal .
Weiss et al. ( 1997) suggested a second possibility for the presence of GPal in
the ER membranes of plants. The authors cited the fact that several plant hormones
are known to be membrane penneable. In addition, the sequence of an auxin binding
protein suggests that it may be localized to the ER. Furthemore, they pointed out that

both types of light receptors, phytochromes for red light, and the HY4 protein for blue
iight, appear to be cytoplasmic. Since G-proteias have k e n impiicated in both auxin
and Light signaling, the authors hypothesized that G-protein signaling in plants does
not have to be across a membrane. In that case, receptors that are coupled to the ER-
located G-proteins do not need to be trammembrane recepton as is the case in
animals. However, this hypothesis fails to account for the observed stimulation of
GTP binding to the plasma membrane of plants in response to light and auxin
(Romero et al. 1991; Warpeha et al. 1991; Zaina et al., 1990).
4.6 Functional Implications of TGal - Interacüng with a DnaJ-like Protein
To help elucidate potential functions of TGal , an interactive cloning
technique was used to identiQ proteins that interact with TGal . Four cDNA's were
isolated, and their partial sequencing revealed them to be identicai. The protein
sequence of the translated DNA shows high homology to a protein family designated
Dnd. Aithough the interaction was found in vitro, and was not fully characterized by
other methods and most importantiy has not k e n shown NI vivo, it is worthwhile to
consider the possible implications of that interaction.
The DnaJ family of proteins was originally found in Eschenchia coli, and its
role was identified in regulating the chaperone activity of another bacterial protein
DnaK (Hsp7O - a 70 kDa heat shock protein). The E . d i DnaJ interacts directly with
DnaK thereby stimulating its ATPase activity, and thus acts as a chaperone in

conjunction with DnaK. These activities enable DnaJ to assist DnaK in protein
folding and in the assembly and disassembly of protein complexes. Genes that encode
proteins homologous to the E.coli DnaJ were identified in eukaryotic organisms
ranging from yeast to plants and humans (reviewed by Silver and Way, 1993; Cyr et
al. 1994). In eukaryotic cells, as in bactenal cells, DnaJ proteins were observed, in
conjunction with Hsp70, to mediate a wide variety of cellular events that involve
modulation of protein structure. The Hsp70 famiiy have been identified in al1 free
living organisms so far examined. They were originally identified because of their
inducible expression by heat shock but are now known to be constitutively expressed
as well, with specific farnily members targeted to different cellular organelles. They
function as molecular chaperones by binding to other polypeptides (Caplan et al..
1993). Current evidence indicates that both Hsp70 and Dnd-Wre proteins act as
molecular chaperones and bind polypeptides, but with different substrate specificities.
Hsp7O proteins bind unfolded proteins, recognizing shon stretches of amino acids that
assume an extended conformation. By contrast, Dnd-like proteins bind protein
substrates exhibiting secondary and tertiary structure. These differences in substrate
specificity may allow DnaJ-like proteins to facilitate the interaction of nascent
proteins with Hsp70, which would not nomally be bound (Cyr et al., 1994).
The best studied eukaryotic DnaJ-like proteins are SEC63 and YDJl in yeast.
They have been shown to function in two separate transport pathways in targeting and
sorting of polypeptides foilowing their synthesis. Mutants in SEC63 were found to
affect transport of polypeptides across the ER membrane and into the nucleus. YDJl

functions in the cytosol to transport polypeptides targeted to both the mitochondria
and the ER. Both SEC63 and YDJl appear to function with Hsp7O proteins whose
role is in maintainhg polypeptides in a transport-competent conformation before their
import into mitochondria or translocation across the ER membrane (Lyman and
Schekman. 1996). Data would seem to suggest that YDJl stimulates the Hsp7O
ATPase resulting in the dissociation of the Hsp7O:polypeptide complexes and thus,
k i n g the polypeptide for its interaction with a putative receptor or other component
of the translocation apparatus. SEC63 in turn, appears to be an integral component of
the membrane-bound translocation machinery facilitating the polypeptide
translocation into the ER (Caplan et al., 1993).
Within the context of the observed roles of the DnaJ-like protein family, a
number of functional roles can be speculated for the possible interaction of TGal
with a Dnd-like plant homologue. The Dnd-like proteins appear to regulate the
ATP-dependent Hsp7O:polypeptide dissociation, although experimentai evidence
supporting the regdation of the DnaJ-like protein is lacking. in keeping with the
known regulatoiy role of G-proteins in signal transduction. and the presence of TGal
in the plant ER, one may hypothesize a regulatory role for TGal in polypeptide
release. Upon arriva1 of a Hsp7O:polypeptide complex to the ER membrane, TGal in
association with a Dnd-like protein may become activated thereby resulting in an
interaction between the Hsp70:polypeptide and TGa1:Dna.J-like protein. This
interaction may result in the hydrolysis of the ATP-bound to Hsp70, which in turn

prompts the dissociation of the transported polypeptides to be translocated into the
ER. Inactivation of TGal , by the hydrolysis of its bound GTP, leads to the
dissociation of the TGa1:DnaJ:HspiO complex, resulting in the release of Hsp70 to
reassume its chaperone function (Fig. 16).
Another possible explmation for the observed interaction lies in the fact that
the protein used in the interaction screen was a recombinant enzyme expressed in a
heterologous system. Since the recombinant TGal was observed to be a soluble
enzyme, but its natural counterpart was found to be associated with plant membranes,
it is probable that it lacks structurai modifications to allow its incorporation into
phospholipid mcieties. Heterotrimeric G-proteins are known to be modified by acyl
groups allowing their incorporation into membranes, and the sequence of TGal
shows potential residues for these modifications (Ma et al., 1991). It is possible that
the interaction between TGal and the Dnd-like protein represents an intermediate
complex destined for pst-translationai modifications. In this case, the Dnd-like
protein itself serves as a chaperone directing the covalent modifications of TGal by
other proteins, and is possibly involved in targeting TGal to either the ER or plasma
membrane.
An additional potential role for this putative interaction may concern the
suggested roles of Dnd-like proteins in stress responses. The cDNA expression
library screened was constmcted from cells exposed to race specific fungal eliciton.
Although not confmed by Northem analysis, the isolation of four identical cDNA's

Figure 16. A proposed mode1 for the regulation of a DnaJ-like protein. by
TGal, in polypeptide translocation in the ER. (a) TGal is associated with the DnaJ
protein in the ER. (b) Upon arriva of the Hsp7O:polypeptide complex to the ER
membrane. an unknown signal activates TGal. (c) The activated TGal allows for the
interaction of DnaJ with Hsp70. inducing ATP hydrolysis by Hsp7O. (d) The
hydrolysis of the ATP by Hsp70 results in the release of the polypeptide. (e) GTP
hydroIysis by TGal results in the dissociation of the DnakHsp70 complex. (f) The
high concentration of ATP in the cytosol allows for the binding of ATP by Hsp70. (f)
Hsp7O bound by ATP is free to associate with other polypeptides. The polypeptide
delivered by Hsp70 is free to be processed by the ER translocation apparatus.

ATP

from the iimîted number of plaques screened could suggest upregulation of that
hwscript under fungal attack. The role of Dnd-like proteins as molecular chaperones
independent of Hsp7O have been already suggested. In plants, Zhu et al. (1993)
observed transcriptional up-regulation of ANJ 1 (a D n d homologue) upon salinity
stress in ArBplex numrnularia tissue culture, while transcription levels of Hsp7O were
unchanged. Since both ANJl and Hsp70 are upregdated in a CO-ordinated manner
upon heat stress, this suggests a separate function for ANJl when osmolarity changes.
Under extreme physiological changes induced by stress, molecular chaperones may
interact to stabilize enzymes and maintain their activity. Under fungal attack, the
DnaJ chaperones may interact with TGal either at the ER or plasma membrane to
proteci ifs enzymatic activity. ensuring its continuous function. In yeast, raising the
incubation temperatures to 37'~ for 1 hour led to the redistribution of YDJl from
20% to 50% membrane bound, indicating a possible stabilization function for these
DnaJ-like proteins under stress (Caplan et al., 1993).
4.7 Possible roles for TGal in the Plasma Membrane
The presence of TGal in the plasma membrane fits the classical mode1 for the
involvement of heterotrimeric G-proteins in signal perception and propagation of
extracellular stimuli. It still remains to be uncovered what plant agonist(s) is
responsible for the activation of TGal. Since G-protein a subunits are known to be
coupled to 7TMS receptors through contact at their C-terminal end, a putative TGal

receptor contact was examined. Since no 7TMS receptor - G-protein system is known
in plants, mastoparan which is known to interact with the C-terminal of some animai
Ga's, and have measurable effects in plants, was used instead. The activation of the
plasma membrane w-ATPase by mastoparan was used as an assay system.
Experiments using synthetic peptides corresponding to the C-terminus of TGal
demonstrated specific, sequencedependent inhibition (Fig. 13b). These results would
suggest direct interaction of the TGal C-tenninus with mastoparan. Pre-incubation of
membrane vesicles wi th the anti-TGa l antibody did not inhibit mastoparan-
dependent activation of the w-~TPase. Furthennore, addition of recombinant TGal
to membrane vesicles did not result in increased ATPase activity. Although these
results would suggest that TGal is not the G-protein regulating the H+-ATP~s~
activity through a membrane-associated phosphatase (Vera-Estrelia et al., 1994; Xing
et al., 1997). it is still likely that the observed inhibition represents direct interaction
of the C-terminus of TGal with mastoparan. When different punfied animal Ga's
were reconstituted into phospholipid vesicles, mastoparan was observed to stimulate
their GTPase activity to different extents (Higashijima et al., 1988). These results may
indicate different aEnities of mastoparan to the C-terminai of different mammalian
Ga's and the situation may be similar in plants. Unfortunately, experiments using
TGal, reconstituted into phospholipid vesicles. did not result in an increased K, of
GTP hydrolysis. That may be due to the lack of an acyl group modification on TGal.
which cannot be achieved in prokaryotic expression systems. Without an acyl

modification, TGal would not associate with the lipid moiety and properly interact
with mastoparan, which has to be embedded in the membrane to assume its a-helical
conformation. To examine this possibility, these experiments would need to be
repeated using TGal , either purified from plant celIs or expressed in a eukaryotic
system capable of the required pst-translational modifications. The inhibition of the
effects of mastoparan by the C-terminus of TGal may suggest the presence of 7 T M S
receptors in plants and their coupling to G-proteins. The recent cloning of a putative
G-protein-coupled ï T M S receptor in Arabidopsis would seem to confirm these
observations (Josefsson and Rask, 1997).
Other experiments using recombinant wild-type and 42231. - mutant TGal ,
demonstrated its regdation of a previously characterized plasma membrane ca2+
channel in plants (Fig.17) (Aharon et al., manuscript in preparation). These results
suggest an important regulatory role for TGal in ce11 signaling, The role of ca2+ in
plants as a secondary messenger is well documented. Many cellular processes are
regulated by changes in cytosolic ca2+ levels (Bush 1993). The discovery of the
CDPK (ca2+ dependent protein kinases) family, unique to plants, having a
caimodulin-like domain fused to the kinase domain, demonstrates the unique
importance of ~ a " in plant signaling (Roberts and Hannon, 1992). Interestingly. the
TGal-regulated ca2+ channel was previously shown to be regulated by mastoparan in
addition to Cladosporium fitlvwn fungal elicitors (Gelli et al., 1997). Blocking
heterotrimeric G-proteins by using GDPPS, in the presence of fungd elicitors,

Figure 17. Regulation of a plasma membrane ca2+ channel by TGal .
Protoplast were kept in an g las bathing chamber containing 50 rnM CaC12, O. 1 mM
potassium glutamate, 1 rnMMgS04, 5 rnM Tris-Mes pH 6.1 and sorbitol io give a
final osmolarity of 450 mOsmol. Glass pipettes pulled with a vertical puller from
borosilicate glas capillaries coated with silicone and fire polished, had a tip
resistance of 10 MR when filled with 100 mM potassium glutamate, 1 rnM MgS04, 3
mM Mg-ATP, 0.05 rnM CaClz + 0.1 mM BAPTA to give a final free [ ~ a " ] = 100
nM, 5 mM Tris-Mes (pH 7.3) and sorbitol with a final osmolarity of 475 mOsmol.
GTP (20 IrM) and recombinant proteins (1 pmollml) were added where indicated.
Outside-out patches of plasma membranes were obtained after the whole-celi
configuration mode (Gelli et al., 1997) by quickly pulling the pipette away fiom the
protoplasts. Single channel currents were measured at 23 OC in voltage-clamp mode
using a Dagan 3900 amplifier (Dagan Corporation) digitized on line (TL4 DMA
interface), stored on a 386 based 33 MHz cornputer and acquired and analyzed with
pClarnp 6.0.2 software (Axon Instruments). Data were filtered with a four-pole Bessel
filter at 200 Hz, digitized at 2 KHz and stored on disk. Open channei probabilities
(Po) were caiculated with amplitude histograms obtained from single channel
recordings as described by Gelli et al., (1997).
The figure shows some original single-channel recordings from isolated patches of
membranes and the carresponding amplitude histograms. Under control conditions,
channel activity was detected at membrane potential ciifferences that corresponded to

the movement of ca2+ into the cytosol. Amplitude histograms of the single channels
revealed at Least one channel in the patch of membrane +.th an amplitude of -1pA. In
the presence of 20 pM. the channel events became more fiequent an a two-fold
increase in the mean Po was observed. The presence of recombinant a-subunits in the
pipette resulted in a significant increase in channel activity as shown by the increase
in channel events and by the increase in the mean P, from 0.19 to 0.30 and 0.53 in
the presence of TGal-WT and TGaLQ223L. respectively. The single channel
conductance (1 1 pS) did not significantiy change in the presence of the recombinant
a-subunits. thus single channel current levels remaineci unchanged.

CONTROL
- P,=0.10fO.M

abolished the observed channel activation, demonstrating a direct role for G-proteins
in mediating elicitor signaling (Gelli et al., 1997). These results rnay suggest a
possible involvement of TGal in the Cladosporiumfulvum-tomato interaction
4.8 Mechanisms of Heterotrimeric G-proteln Involvement in Host-Pathogen
Interactions
The possible involvement of membrane-associated plant G-proteins in
response to pathogens is well documented. (Legendre et a1.,1992; Vera Estrella et al.,
1994; Beffa et al. 1995; Gelli et al., 1997; Xing et al., 1997). Whether TGal is
involved in mediating the signal, or any other membrane G-protein, their mechanism
of involvement is puvling in light of recent knowledge. It is therefore worth
considering the possible implications of G-proteins mediating responses in host-
pathogen interactions
The recent cloning of plant resistance (R) genes, from a number of host-
pathogen model systerns, represents a leap in the molecular understanding of host
resistance (reviewed by Bent 1996). Disease resistance in many plant-pathogen
interactions requires the complementary expression of two genes - a dominant or
semi-dominant plant R gene and a corresponding dominant pathogen avimlence (avr)
gene. The proposed model suggests that the direct or indirect interaction of the AVR
product with the R gene product triggen resistance (Dangl, 1995). Based on sequence
analysis of known R genes, it is hypothesized that the R gene product codes for a
membrane receptor capable of binding the AVR product as a ligand (Bent et al., 1994;

Jones et al., 1994; Song et al., 1995). Interestingly, although a possible role for G-
proteins has k e n described for a number of host-pathogen interactions, including the
Cladosporiwn fulvum-tomato interaction (Vera Estrella et al., L 994; Gelli et al., 1997;
Xing et al., 1997). none of the R gene products described to date in general. and
specifically in the CIadosporiwt fulvum-tomato system, show any homology to
known G-protein -coupled receptors (Jones et al.. 1994; Dixon et al., 1996). The
same is true for other stimuli which were observed to involve membrane G-proteins
in plants for which their receptors are known, such as light (Neuhaus et al. 1993) and
the plant hormone auxin (Zaina et al., 1990). The question which arises is therefore:
what are the possible mechanisms in plants goveniing membrane associated G-protein
activation? By addressing the issue specifically in host-pathogen interactions, some
insight may be gained as to the general mechanisms of G-protein regulation in plants.
Of the known plant R genes, the Cf gene family in tomato is the most
intriguing. Specific Cf genes were found to be responsible for resistance of tomato to
different races of the Cladosporiwn fulvum fungus (Jones et al.. 1994; Dixon et al..
1996). The predicted protein sequences of aimost all known R genes shows that they
are compnsed of leucine-rich-repeats (LRR) dong with some other structural motifs
(Bent, 1996). But in the case of the Cf family, they appear to consist almost entirely of
these Lm ' s (Jones et al., 1994; Dixon et al., 1996; Dangl and Holub, 1997). The
role of Lm's in mediating protein-protein interactions, peptide-ligand binding, and
proteincarbohydrate interactions has been weil estabiished ( reviewed by Kobe and
Deisenho fer, 1994).

Sequence cornparisons of the four known Cf resistance gene products.
revealed a putative membrane anchor and a small cytoplasmic domain ai their C -
terminus. Nearly ail arnino acid differences between the Cf proteins were found in
-30% of the amino-terminal LRR domains, suggesting that they determine specificity
to different fungal AVR products (Dangl and Holub. 1997). The carboxy-terminal
LRR domains are therefore fairiy conserved (while the cytosolic C-terminai is not),
suggesting their functional importance in al1 Cf proteins.
Since the carboxy-terminal LRR's are predicted to be extracellular, there is no
indication as to how the signals are intemalized by the Cf proteins. A possible model
would implicate the conserved LRR's in extracellular protein interaction with other
membrane associated receptor molecules. These in turn, may intemalize the signal
and propagate it. In light of the evidence supporting the role of G-proteins in
mediating hingai elicitor responses, an interesting hypothesis would be that the Cf
proteins interact extracellularly with G-protein - coupled 7TMS recepton (Fig. 18).
A combinatorid receptor model, in the Cladosporium filvum - tomato
interaction, was already suggested by Kooman-Gersmann et al. (1996) when
exarnining the of the AVR9 elicitor to the plasma membranes of different
plants. The AVR9 is a race-specific peptide elicitor, and its corresponding R gene
(Cf9) was cloned (Jones et al.. 1994). Interestingly, the authors observed high affinity
binding sites for the AVRP peptide in both resistant tomato Iines (cmying the C'
gene), and susceptible lines (Cfl - canying no resistance gene). The presence of other
receptors, capable of binding the Am9 elicitor, was therefore hypothesized.

Figure 18. Putative mechanism for the involvement of membrane-associated
G-proteins in host-pathogen interactions. Binding of the AVR peptide to the Cf
protein results in the activation of a G-protein - coupled 7TMS receptor via
extracellular protein-protein interaction. The conserved carboxy LRR's (coloured in
blue) may be the ones mediating that interaction.

suggesting a CO-ordinated role for the perception of the signal. Such examples are
known in plants, such as the interaction between the SLG and SRK protein in the
Brassica self-incompatibility system (Nasrailah., 1997).
It is possible that membrane signaling in plants involves a complex network of
receptor interactions. A mechanism could therefore be postulated where various
signals are perceived by their respective receptor molecule. which in tum activate G-
protein - coupled receptors through protein-protein interactions. The recent cloning of
a 7TMS receptor-like protein in Arabidopsis (Josefsson and Rask, 1997). suggests
that receptor-mediated G-protein activation is similar to that descnbed in animais.
The questions which still remain to be solved are what are the plant agonists? and
what are the precise mechanisms by which they are perceived at the plasma
membrane to allow their signal to be internalized by membrane-associated G-
proteins.
FZlTURE WORK
Future goals in the characterization of TGal have to focus on in vivo studies
in light of the results generated in vitro by this project. The generation of the GTPase-
deficient TGal, resulting in its constitutive activation, as weli as the identification of
its putative receptor contact region, present an attractive method for studying G-
protein function in transgenic plants. These could be transformed with the Q223L
mutation, resulting in over-activation of G-protein mediated pathways. h addition.
transformation of plants with a truncated TGal. in its C-terminus, codd inhibit those

same pathways by uncoupling receptor - G-protein contacts. The association of G-
proteins in a heterotrimeric cornplex, facilitates the generation of dominant-negative
mutations as descnbed above. Furthermore, purification of plasma membranes fkom
transgenic plants, could allow the direct assessrnent of the role of different plant
agonists in the activation of TGal by simpie GTPase assays. An observed increase in
GTP hydrolysis for a given agonist in a wild type plant, while not observed in a
mutant plant, could point to the role of that agonist in TGal activation.
The interaction of TGal with DnaJ should as well be examined in vivo by
techniques such as CO-immunopercipitation. Specificaily. the interaction should be
subcellulaily localized to the membrane in which it occurs i.e. plasma membrane or
ER, In addition, the interaction should be examined under stress conditions such as
fungal elicitors, heat-shock, sait stress etc. Only when the interaction is observed in
vivo in a specific location or under a specific response, the different hypotheses could
be further examined. Furthermore, additional screens, by the use of the same
technique for detecting protein-protein interactions, should be camied out to identiS
other targets of interaction. This should include the use of the Q223L mutant, which
may have higher affinities, than the wild type product, to some effector molecules.

Aharon, G., Gelli, A., Snedden, W.A. and Blumwald, E. (1998). Regulation of a plant plasma membrane ca2+channel by TGal, a heterotrimenc G-protein a subunit homologue. (manuscript in preparation).
Ames, B.N. ( 1966). Assay of inorganic phosphate. total phosphate and phosp hatases. Methods Enzymol. 8: 1 15- 1 18
Armstrong, F. and Blatt, M.R (1995). Evidence for K+ channel control in Vicia guard cells coupled by G-proteins to a 7TMS receptor mimetic. Plant J. 8: 187- 198
Audigier, Y., Nigam, S.K. and Blobel, Go (1988). Identification of a G Protein in Rough Endoplasmic Reticulum of Canine Pancreas. J. Biol. Chem. 263: 16352- 16357
Barford, D. (1991). Molecular mechanisms for the control of enzymatic activity by protein phosphorylation. Biochim. Biophys. Acta 1133: 55-62
Beffa, R., Szell, M., Meuwly, P., Pay, A., Vogeli-Lange, R, Mebaux, J-P., Neuhaus, Go, Meins, Fm and Nagy, Fm (1995). Cholem toxin elevates pathogen resistance and induces pathogenesis-related gene expression in tobacco. EMBO J. 14: 5753-5761
Bent, A.F. (1996). Plant Disease Resistance Genes: Function Meets Structure. Plant Ce11 8: 1757-177 1
Bent, A.F., Kunkel, B.N., Dahlbeck, D., Brown, K.L., Schmidt, R., Giraudat, J., Leung, J. and Staskawicz, BJ. (1994). RPS2 of Arabidopsis thaliana: a leucine-rich repeat class of plant disease resistance genes. Science 265: 1856- 1860
Blum, W., Hinsch, K.D., Schultz, G. and Weiier, E.W. (1988). Identification of GTP-Binding Proteins in the Plasma Membrane of Higher Plants. Biochem Biophys. Res. Commun. 156: 954-959
Blumwald, E., and Poole, RJ. (1987). Salt tolerance in suspension cultures of sugar beet. Plant Physiol. 83: 884-887

Boguski, MS. and McCormick, Fm (1993). Proteins regulating ras and its relatives. Nature 366: 643-654
Bourne, HmRm (1988). Do GTPases Direct Membrane Traffic in Secretion? Ce11 53: 669-67 1
Boume, H.R., Sanders, DA. and McCormick, F. (1990). The GTPase superfamily: a conserved switch for diverse ceU functions. Nature 348: 125- 132
Bourne, H.R., Sanders, DA. and McCormiek, F. (199 1). The GTPase superfamily: consewed s tmcture and molecular mechanism. Nature 349: 1 17- 127
Bradford, M.M. (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248-254
Brandt, D.R., Asano, T., Pedersen, S.E. and Ross, E.M. (1983). Reconstitution of catecholamine-stimulated guanosinetriphosphatase activity. Biochernistry 22: 4357-4362
Bush, DS. (1993). Regulation of cytosolic calcium in plants. Plant Physiol. 103: 7- 13
Caplan, AJ., Cyr. D.M. and Douglas, M.G. (1993). Eukaryotic Homologues of Escherichia coli dnd : A Diverse Protein Family That Functions with HSWO Stress Proteins. Mol. Biol. Ce11 4: 555-563
Casey, PJ., Fong, HmKWm, Simon, M.1. and Gilrnan, A.G. (1990). Gz, a guanine nucleotide-binding protein with unique biochemical properties. J. Biol. Chem. 265: 2383-2390
Cassel, D. and Selinger, 2. (1977). Mechanism of adenylate cyclase activation by cholera toxin: Inhibition of GTP hydrolysis at the regulatory site. Proc. NatL Ac& Sci. USA 74: 3307-33 1 1
Chapline, C., Ramsay, K., Klauck, T., and Jaken, S. (1993). Interaction Cloning of Protein Kinase C Substrates. J. Biol. Chem. 268: 6858-6861
Clapham, D., E. and Neer, E J. (1993). New roles for G-protein By-dimers in transmernebrane signaling. Nature 365: 403-406

Clapham, D., E. (1994). Direct G Protein Activation of Ion Channels?. Annu. Rev. Neurosci. 17: 44 1 -464
Clarkson, J., White, I.R. and Milner, PA. ( 199 1). Specific immune detection and partial purification of G-proteins h m Arabidopsis thaliana. . Biochem. Soc. Tram. 20: 9s
Conklin, B.R., Farfel, Z., Lustig, K.D., Juiius, D. and Bourne, H.R. (1993). Substitution of three amino acids switches receptor specificity of G,a to that of G i a Nature 363: 274-28 1
Cyr, D.M., Langer, T. and Douglas, M.G. (1994). DnaJ-like proteins: molecular chaperones and specific regulators of Hsp70. Trends Biochem. Sci. 19: 176- 18 1
Dangl, J.L. (1995). Pièce de Resistance: novel classes of plant disease resistance genes. Cell80: 363-366
Dan& J. and Holub, E. (1997). La Dolce Vita: A Molecular Feast in Plant-Pathogen Interactions. Cell 91: 17-24
Dixon, M.S., Jones, D.A., Keddie, JS., Thomas, CM., Hamson, K. and Jones, J.D.G. (1996). The tomato Cf-2 disease resistance locus comprises two functional genes encoding leucine-nch repeat proteins. Cell 84: 45 1-459
Dohlman, H.G. and Thorner, J. (1997). RGS Proteins and Signaling by Heterotrimeric G Proteins. J. Biol. Chem. 272: 387 1 -3874
Fields, TA. and Casey, P J. (1 997). Signaling functions and biochernical properties of pertussis toxin-resistant G-proteins. Biochem. J. 321: 56 1-57 1
Fung, B.K.IC, Hurley, J.B., and Stryer, L. (198 1). Flow of information in the Iight- tnggered cyclic nucleotide cascade of vision. Proc. Natl. Acad Sci. USA 78: 152- 156
Gelli, A., Higgins, V J. and Blumwald, E. ( 1997). Activation of Plant Plasma Membrane ca2+-permeable Charnels by Race-Specific Fungal Elicitors. Piant Physiol. 113: 269-279
Gilman, A.G. (1984). G Proteins and Dual Control of Adenylate Cyclase. Ce11 36: 577-579

Gilman, A.G. (1987). G Proteins: Transducers of Recepetor-Generated Signais. Ann. Rev. Biochem. 56: 6 15-649
Gotor, C., Lam, E., Cejudo, F J. and Romero, L.C. (1996). Isolation and analysis of the soybean SGA2 gene (cDNA), encoding a new member of the plant G- protein farnily of signal transducers. Plant Mol. Biol. 32: 1227- 1234
Graziano, M.P. and Giiman, AmGe ( 1989). Synthesis in Escherichia coli of GTPase- deficient Mutants of Gs,. J. Biol. Chem. 264: 15475- 15482 guanylnucleotides on binding of 1251-glucagon. J. Biol. Chem. 246: 1872- 1876
Hall, A. (1990). The cellular functions of small GTP-binding proteins. Science 249: 634-640
Harlow, E. and Lane, D. (1988). Antibodies - a laboratory manuai. Cold Spring Laboratoty p.498
Hasunuma, K. and Funadera, K. (1987). GTP-Binding Protein(s) in Green Plant, Lemna paucicostata. Biochem. Biophys. Res. Commun. 143: 908-9 12
Higashijima, T, Ferguson, KM., Sternweis, P.C., Smigel, M.D. and Gilman, A.G. (1987). Effects of Mg2+ and the beta gamma-subunit complex on the interactions of guanine nucleotides with G proteins. J. Biol. Chem. 262: 762-766
Higashijima, T., Uzu, S., Nakajima, T. and Ross, EM. (1988). Mastoparan, a Peptide Toxin from Wasp Venom, Mimics Receptors by Activating GTP- binding Regdatory Proteins (G Proteins). J. Biol. Chem. 263: 649 1-6494
Higashijima, T., Wakamatru, K., Saito, K., Fujino, M., Nakajima, T. and Miyazawa, T. (1984). Molecular Aggregation and Conformationai Change of Wasp Venom Mastoparan as Induced by Salt in Aqueous Solution. Biochim. Biophys. Acta 802: 157- 16 1
Huang, C., Hepler, J.R., Gilman, A.G. and Mumby, S.M. ( 1997). Attenuation of Gi- and Gq-mediated signaling by expression of RGS4 or GAP in marnmalian cells. Proc. Natl. Acad. Sci. USA 94: 6 159-6 163
Ishida, B., Takahashi, Y. and Nagata, T. (1993). Isolation of cDNA of an auxin- regulated gene encoding a G protein P subunit-like protein from tobacco BY-2 cefis. Proc. Natl. Acad. Sci. USA 90: 1 1 152-1 1156

Ishikawa, A., Iwasaki, Y. and Asahi, T. (1996). Molecular cloning and charactenzation of a cDNA for the beta subunit of a G protein from rice. Plant Celt Physiol. 37: 223-228
Iwasaki, Y., Kato, Tm, Kaidoh, T., Ishikawa, Ao and Asahi. Tm (1997). Characterization of the putative a subunit of a heterotrheric G protein in rice. Plant Mol. Biol. 34: 563-572
Jacobs, M., Thelen, MOP., Famdale, ReWo, Astle, MX. and Rubery, P.H. (1988). Specific Guanine Nucleotide Binding By Membranes from Cucurbita pepo Seedlings. Biochem Biophys. Res. Commun. 155: 1478- 1484
Jones, D A , Thomas, CMo, Hamrnond-Kosack, KE.9 Balint-Kurti, P JO and Jones, JD-G. (1994). Isolation of the tomato Cf-9 gene for resistance to Cladosporiwnjùlvwn by transposon tagging. Science 266: 789-793
Jones, DOT and Reed, R.R. (1987). Molecular Cloning of Five GTP-binding Protein cDNA Species from Rat Olfactory Neuroepithelium. J. Biol. Chem. 262: 1424 1 - 14249
Josefsson, L.-G. and Rask, L. (1997). Cloning of a putative G-protein-coupled receptor fiom Arabidopsis thaliana. Eur. J. Biochem. 249: 4 15-420
Katada, T. and UI, M o (1982). Direct modification of the membrane adenylate cyclase system by islet-activating protein due to ADP-ribosylation of a membrane protein. Proc. Nat1 Acad Sci. USA 79: 3 129-3 133
m i r o , Y. ( 1992). Signal Transducing G-proteins: a Subunits. In: Brann, M.R., ed. Molecular Biology of G-Protein-Coupled Receptors. Birkhauser, 234-269.
Kaziro, Y., Itoh, Ho, Kozasa, T., Nakafuku, M. and Satoh, T. (1991). Structure and hinction of signai-transducing GTP-binding proteins. Annu. Rev. Biochem. 60: 349-400
Kim, W.Y., Cheong, NoEq Lee, D.C., Je, Deyo, Bahk, J.D., Cho, MJ. and Lee, S.Y. (1995). Cloning and Sequencing Analysis of a Full-Length cDNA Encoding a G Prctein a Subunit, SGAI, from Soybean. Plant Physiol. 108: 1315-1316
Kobe, B. and Deisenhofer, Jo (1994). The leucine-rich repeat: a versatile binding motif. Trends Biochem Sci 19: 415-421

Kobe, B. and Deisenhofer, J. (1994). The leucine-rich repeat: a versatile binding motif. Trends Biochem. Sci. 19: 4 15-42 1
Kooman-Gersmann, M., Honée, G., Bonnerna, G. and De Wit, P J.G.M. (1996). A high-affinity binding site for the AVR9 peptide elicitor of Clodosporiwn filvum is present on plasma membranes of tomato and other solanaceous plants. Plant Ce11 8: 929-938
Kooman-Getsmann, M., Honée, G., Bonnema, G. and De Wit, P J.G.M. (1996). A high-aKhity binding site for the AVR9 peptide elicitor of Cladosporium fulvum is present on plasma membranes of tomato and other solanaceous plants. Plant Ce11 8: 929-938
Kozasa, T. and Gilman, A.G. (1995). Purification of recombinant G proteins from Sf9 cells by hexahistidine tagging of associated subunits. Characterization of a12 and inhibition of adenylyl cyclase by a=. J. Biol. Chem. 270: 1734- 174 1
Laemmli, U.K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685
Lambright, D.G., Sondek, J., Bohm, A., Skiba, N.P., Hamm, H.E. and Sigler, PB. (1996). The 2 A crystal structure of a heterotrimeric G protein. Nature 379: 31 1-319
Legendre, L., Heinstein, P.F. and Low, P.S. (1992). Evidence for participation of GTP-binding proteins in elicitation of the rapid oxidative bunt in cultured soybean cells. J. Biol. Chem. 267: 20 140-20147
Linder, M.E., Ewald, DA., Miller, R J. and alman, A.G. (1990). Purification and characterization of Goa and three types of Gia after expression in Escherichia coli. J. Biol. Chem. 265: 8243-825 1
Lyman, S.K. and Schekman, R. (1996). Polypeptide translocation rnachinery of the yeast endoplasmic reticulum. Experientia 52: 1042- 1049
Ma, He 1994. GTP-binding proteins in plants: new members of an old family. Plant Mol. Biol. 26: 16 1 1- 1636
Ma, H., Yanofsky, M.F. and Hai, HA. (1991). Isolation and sequence analysis of TGAI cDNAs encoding a tomato G-protein a subunit. Gene 107: 189- 195

Ma, H., Yanofsky, MaFe and Meyerowk, E.M. (1990). Molecular cloning and characterization of GPAI, a G protein a subunit gene from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 87: 3 82 1 -3825
Maguire, M.&, Van Arsdaie, P.,Mm and Gflmsn, Ao,Go (1976). An agooist-specific effect of guanine nucleotides on binding to the beta adrenergic receptor. Mol. Phannacol. 12: 335-339
Masters, S.B., Miller, RrnT., Chi, M-H., Chang, F-Hm, Beiderrnan, B., Lopez, N.G. and Bourne, H.R. (1989). Mutations in the GTP-binding Site of Gsa Alter S tirnulation of Aden y1 y1 Cyclase. J. Biol. Chem. 264: 1 5467- 1 5474
Moss, J. and Vaughan, M. (1977). Mechanism of Action of Choleragen- Enidence for ADP-ribosyltransferase activity with arganine aç an acceptor. J. Biol. Chem 252: 2455-2457
Nasrallah, JJ. (1997). Evolution of the Brassica self-incompatibility locus: A look into S-locus gene polyrnorphisrns. Proc. Natl. Ac&. Sci. USA 94: 95 16-95 19
Neuhaus, Ge, Bowler, Cm, Kern, R and Chua, N. (1993). Calciurn/Calmodulin- Dependent and -Independent Phytochrome Signal Transduction Pathways. Cell. 73: 937-952.
Northup, J.K, Sternweis, P.C., Smigel, MODO, Schleifer, L.S., Ross, E.M. and GiIman, A.G. (1980). hirification of the regulatory component of adenylate cyclase. Proc. Natl. Acad. Sci. USA 77: 65 16-6520
Nuoffer, C. and Balch, W.E. (1994). GTPases: multifunctional molecular switches regulating vesicular transport. Annu. Rev. Biochem. 63: 949-990
Offermanns, S. and Schultz, G. (1994). What are the functions of the pertussis toxin-insensitive G proteins Gt z, G13 and Gz? Mol. Cell. Endocrinol. 100: 7 1-74
Orly, J. and Schramrn, M. (1976). Coupling of catecholamine receptor frorn one ce11 with adenylate cyclase from another celi by ceii fusion. Proc. Natl. Acad Sci. USA 73: 4410-4414
Pfeuffer, Ee, Mollner, S., Lancet, D. and Pfeuffer, T. (1989). Olfactory adenyiyl cyclase. Identification and purification of a novel enzyme form. J-Bioi. Chem 264 : 18803- 18807
Poulsen, C , Mai, X.M. and Borg, S. (1994). A L o u japonicur cDNA Encoding an a Subunit of a Heterotrirneric G-Protein. Plart Physiol. 105: 1453- 1454

Roberts, D.M. and Hannon, A.C. (1992). Calcium-modulated proteins: Targets of intracellular calcium signals in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43: 375-414
Rodbell, M., Bimbaumer, L., Phol, S.L. and Krans, MJ. (1971a). The Glucagon- sensitive Adenyl Cyclase System in Plasma Membranes of Rat Liver. J. Biol. Chem. 246: 1 877- 1882
Rodbell, M., Krans, H.M., Pohl, S.L. and Bimbaumer, L. (197lb). The glucagon sensitive adenyl cyclase system in plasma membranes of rat Iiver. TV. Effects of
Romero, L.C., Sommer, D., Gotor, C. and Song, P.S. (1991). G-proteins in etiolated Avena seedlings. Possible phytochrome replation. FEBS Len. 282: 34 1-346
Seo, AS., Kim, H.Y., Jeong, J.Y., Lee, S.Y., Cho, MJ. and Bahk, J.D. (1995). Molecular cloning and characterization of RGAl encoding a G protein a subunit from rice (Oryza sativa L. IR-36). Plant Mol. Biol. 27: 1 1 19- L 13 1
Seo, AS., Choi, CH., Lee, S.Y., Cho, M J. and Bahk, J.D. (1997). Biochernical characteristics of a nce (Oryra sativa L.. IR36) G-protein a-subunit expressed in Escherichia coli. Biochem. J. 324: 273-28 1
Shorr, R.G., Lefiowitz, RJ., and Caron, M.G. (198 1). Purification of the 8- Adrenergic Receptor - Identification of the hormone binding subunit. J. Biol. Chem. 256: 5820-5826
Süver, PA. and Way, J-C. (1993). Eukaryotic DnaJ Homologs and the Specificity of Hsp70 Activity. Cell74: 5-6
Simon, ML, Strathrnann, M.P. and Gautam, N. (199 1). Divenity of G Proteins in Signal Transduction. Science 252: 802-808
Song, W.Y., Wang, G.L., Chen, LL., Kim, HS., Pi, L.Y., Holsten, T., Gardner, J., Wang, B., Zhai, W.X., Zhu, L.H., Fauquet, C. and Ronald, P. (1995). A receptor kinase-like protein encoded by the nce disease resistance gene, Xa2 1. Science 270: 1804- 1806
Sternweis, P.C. and Smrcka, A.V. (1992). Regulation of phospholipase C by G proteins. Trends Biochem. Sci. 17: 502-506

Stow, J.L., de Almeida, J.B. (1993). Distribution and role of the heterotrimeric G proteins in the secretory pathways of polarized epithelial cells. J. Ceil Sci. Suppl. 17: 33-39
Stow, J.L., de Almeida, J.B, Narula, N, Holtzman, EJ., Ercolani, L. and AusielIo, D.A. (1991). A heterotrimeric G protein, G*, on Golgi membranes regulates the secretion of heparan suifate proteoglycan in LLC-PKI epitheiial cells. J. Cell Biol. 114: 1 1 13- 1 124
Sullivan, KA, Miller, RT, Mastem, SB., Beiderman, B., Heidernan, W. and Bourne, H.R. (1987). Identification of receptor contact site involved in receptor-G protein coupLing. h r e 330: 758-760
Vera-Estreila, R., Barkla, B J., H i m 7 V J. and Blumwald, E. (1994). Plant Defense Response to Fungal Pathogens. Plmt PhysioL 104: 209-2 15
Verhey, S.D. and Lomax, T.L. 1993. Signai Transduction in Vascular Plants. J Plant Regul. 12: 179- 195
Vries, L.D., Mousü, M., Wurmser, A. and Farquhar, G. (1995). GAP, a protein that specifically interacts with the trimeric G protein Gw, is a member of a protein family with a highiy conserved core domain. Proc. Natl. Acad. Sci. USA 92: 11916-11920
Wang, J., Tu, Y., Woodson, JO, Song, X. and Ross, E.M. (1997). A GTPase- activating Protein for the G Protein Ga, J. BioL Chem 272: 5732-5740
Warpeha, W., HX., Rasenick, M.M. and Kaufman, LS. (1991). A blue-iight-activated GTP-binding protein in the plasma membranes of etiolated peas. Proc. Natl. Acad. Sci. USA 88: 8925-8929
Weiss, CA., Garoaat, C.W., Mukai, K., Hu, Y. and Ma, H . (1994). Isolation of cDNAs encoding guanine nucleotide-binding protein P-subunit homologues h m maize (ZGB1) and Arabidopsis (AGB1). Proc. Nad Acad Sci. USA 91: 9554-9558
Weiss, CA, Huang, H. and Ma, E (1993). Immunolocaiization of the G Protein a Subunit Encoded by the GPAI Gene in Arabidopsis. P lmt Cell5: 15 13- 1528
Weiss, C.A., White, E, Huang, H. and Ma, H. (1997). The G protein a subunit (GPal) is associated with the ER and the plasma membrane in meristematic ceils of Arabidopsis and cauliflower. FEBS Le#. 407: 36 1-367

Wenigarten, R, Ransnas, Le, MueUer, H., Sklar, LA. and Bokoch G.M. (1990). Mastoparan Interacts with the Carboxyl Terminus of the a Subunit of Gi. J. Biol. Chem, 265: 1 1044- 1 1049
Wise, A. and Millner, P.A. (199 1). Evidence for the presence of GTP-binding proteins in tobacco leaf and maize hypocotyl plasmalemma. Biochem. Soc. Tram. 20: 7s
Wise, A., Thomas, P.G., Cam, T.H., Murphy, GA. and Milner, P.A. (1997). Expression of the Arabidopsis G-protein GPal: purification and characterisation of the recombinant protein. Plant Moi. Biol. 33: 723-728
Wise, A., White, I.R. and m e r P.A. (1993). Stimulation of guanosine 5'-0- (3-thio) triphosphate binding to higher plant plasma membranes by the mastoparan-analogue, Mas-7. Biochem Soc. T m . 21: 228s
Xing, T, Biggins VJ. and Blumwald 33. 1997. Identification of G proteins mediating fungai elicitor-induced dephosphrylation of host plasma membrane r - ~ T P a s e . J. Erp. Bot. 48: 229-237
Yang, 2. and Watson, J.C. (1993). Molecular cloning and characterization of rho. a ras-related smaii GTP-binding protein from the garden pea. Proc. Notl. Acad. Sci. USA 90: 8732-8736
Zaina, S., Reggiani, Re and Bertani, A. (1990). Preliminary evidence for involvement of GTP-binding protein(s) in auxin signal transduction in rice (Oryzn sativa L.). J. Plant Physwl. 136: 653-658
Zhao, LJ. Zhang, Q.X. and Padmanabhan, R (1993). Polymerase Chain Reaction-Based Point Mutagenesis Protocol. Methods E n m o l . 217: 2 18-227
Zhu, J.K., Shi, J., Bressan, RA. and Hasegawa, P.M. (1993). Expression of an Arriplex numrnulnria gene encoding a protein homologous to the bacteriai molecular chaperone dnd. Plant Ce11 5: 34 1-349
IMAGE EVALUATION TEST TARGET (QA-3)
APPLIED 2 IMAGE, lnc t 653 East Main Street - -- - - Rochester. NY 14609 USA -- --= Phone: 71 W482-0300 -- - Fa>c 71 61288-5989