avian schistosomes and outbreaks of cercarial …horses, in the southern regions of north america...
TRANSCRIPT

Avian Schistosomes and Outbreaks of Cercarial Dermatitis
Petr Horaacuteka Libor Mikeša Lucie Lichtenbergovaacutea Vladimiacuter Skaacutelaa Miroslava Soldaacutenovaacuteb Sara Vanessa Brantc
Department of Parasitology Faculty of Science Charles University in Prague Prague Czech Republica Institute of Parasitology Biology Centre of the Academy of Sciencesof the Czech Republic Ceskeacute Budejovice Czech Republicb Museum Southwestern Biology Department of Biology University of New Mexico Albuquerque New MexicoUSAc
SUMMARY 165INTRODUCTION 165DIVERSITY OF SCHISTOSOMES CAUSING DERMATITIS 166MOLLUSCAN AND AVIAN HOST SPECIFICITY 168INTRAMOLLUSCAN DEVELOPMENT OF AVIAN SCHISTOSOMES 169VERTEBRATE HOST FINDING AND PENETRATION 170PATHOGENICITY OF AND IMMUNE REACTIONS AGAINST AVIAN SCHISTOSOMES 173
Survival and Migration in Avian Hosts 173Pathology Caused by Visceral Species in Birds and Mammals 173Pathology Caused by Nasal Species in Birds and Mice 174Skin Immune Response and Cercarial Dermatitis 176
DETECTION AND IDENTIFICATION OF AVIAN SCHISTOSOMES 176CLINICAL FEATURES DIAGNOSIS TREATMENT AND PROPHYLAXIS OF HUMAN INFECTIONS 177ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OF AVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS 179
Global Warming and Eutrophication 179Recreational Activities and Cercarial Dermatitis 179Control Measures Related to the Ecology of Avian Schistosomes 180
PERSPECTIVES 180ACKNOWLEDGMENTS 181REFERENCES 181AUTHOR BIOS 189
SUMMARY
Cercarial dermatitis (swimmerrsquos itch) is a condition caused byinfective larvae (cercariae) of a species-rich group of mammalianand avian schistosomes Over the last decade it has been reportedin areas that previously had few or no cases of dermatitis and isthus considered an emerging disease It is obvious that avian schis-tosomes are responsible for the majority of reported dermatitisoutbreaks around the world and thus they are the primary focusof this review Although they infect humans they do not matureand usually die in the skin Experimental infections of avian schis-tosomes in mice show that in previously exposed hosts there is astrong skin immune reaction that kills the schistosome Howeverpenetration of larvae into naive mice can result in temporary mi-gration from the skin This is of particular interest because theworms are able to migrate to different organs for example thelungs in the case of visceral schistosomes and the central nervoussystem in the case of nasal schistosomes The risk of such migra-tion and accompanying disorders needs to be clarified for humansand animals of interest (eg dogs) Herein we compiled the mostcomprehensive review of the diversity immunology and epide-miology of avian schistosomes causing cercarial dermatitis
INTRODUCTION
Cercarial dermatitis is a condition caused by both mammalianand avian schistosomes (Trematoda Schistosomatidae)
Which of those species is more prevalent in a dermatitis outbreakdepends on where you are in the world and how humans andbirdsmammals (and by association snails) come into contactwith a particular type of aquatic environment The name ldquocer-
carial dermatitisrdquo is derived from the term ldquocercariardquo the last lar-val stage developing in an aquatic snail Cercaria is the infectivestage that after leaving the snail searches for and invades a warm-blooded vertebrate host via skin penetration Besides the officialname ldquocercarial dermatitisrdquo many local terms are used (ldquosawahitchrdquo ldquokoganbyordquo etc) with the most widely used name beingldquoswimmerrsquos itchrdquo
Schistosome cercariae were disclosed as the causative agent ofcercarial dermatitis in the United States in 1928 (1) Since thattime numerous reports of cercarial dermatitis have been docu-mented from different parts of the world Global economic lossesdue to outbreaks of cercarial dermatitis are not known as there isno systematic method of reporting either the number of cases orincurred economic losses in terms of recreation or person workhours Furthermore what data do exist that estimate local costsare usually not available to the public domain but it is acceptedthat outbreaks can have considerable impacts on local tourism-based economies in the areas of recreational lakes (2) For exam-ple in the recreational area of Naroch Lake (Belarus) 4737 casesof cercarial dermatitis were recorded between 1995 and 2006 (3)In addition cercarial dermatitis may represent a debilitating oc-
Citation Horaacutek P Mikeš L Lichtenbergovaacute L Skaacutela V Soldaacutenovaacute M Brant SV 2015Avian schistosomes and outbreaks of cercarial dermatitis Clin Microbiol Rev28165ndash190 doi101128CMR00043-14
Address correspondence to Petr Horaacutek petrhorakpetrhorakeu
Copyright copy 2015 American Society for Microbiology All Rights Reserved
doi101128CMR00043-14
January 2015 Volume 28 Number 1 cmrasmorg 165Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cupational disease among rice farmers (4) and may incur costs interms of lost person work hours Older reports refer to 75 ormore of the population experiencing the characteristic symptomsof ldquokoganbyordquo in the areas of Japan where the disease is mosthighly endemic (5) Recent reviews (6ndash9) agree that in some re-gions cercarial dermatitis has appeared as a new problem eitherbecause the dermatitis was previously unknown (eg the USSouthwest and Chile) or because the number of reports of out-breaks increased (8 10 11) Consequently cercarial dermatitis isnow regarded as an emerging disease Besides human schisto-somes (Schistosoma spp) no animal (eg avian) schistosomeshave any other presently known pathogenic effects on humansThus the use of animal models to study the potential risk of ani-mal (avian) schistosomes to human health is invaluable
The last decade has revealed diverse avian schistosome speciesand biology as well as the snails that host them These discoverieshave outpaced the equally essential host-parasite biological im-munological pathological and epidemiological studies of speciesdiversity in terms of incorporating the results of such studies intothe current known diversity of schistosomes Such studies are dif-ficult and time-consuming and consequently only a few specieshave been adapted to experimental conditions Nevertheless suchstudies are crucial to understanding the current and future rolesthat these species might play in the frequency and distribution ofcercarial dermatitis as well as understanding how to break the lifecycle to prevent outbreaks What has been documented and isdetailed in the following sections points to an understanding ofthe avian schistosome-host relationships and thus offers the foun-dation on which future studies will be modeled
DIVERSITY OF SCHISTOSOMES CAUSING DERMATITIS
Considering only the named species in the literature there are 4schistosome genera from mammals and 10 from birds with about30 described species from mammals and about 67 from birds (12)The total is close to 100 species with 70 of them being avianschistosomes distributed around the world that may initiate cer-carial dermatitis The role of some of the species of avian schisto-somes as dermatitis agents has not been studied sufficiently asthey are not often found in areas where people most commonlyare in contact with water and snails To discuss the distributionand diversity of schistosomes causing dermatitis within the phy-logenetic framework of the family Schistosomatidae we refer toFig 1 (12ndash14)
The basal clade of the family tree (Fig 1 clade A) comprises theexclusively marine avian schistosomes Austrobilharzia (4 species)and Ornithobilharzia (2 species) the species shown in the tree arethose for which there are genetic data Species of these two generaare associated with outbreaks of dermatitis in shallow marine en-vironments (15 16) Infection often occurs in people who areswimming playing in tidal pools or working for example col-lecting tidal invertebrates in the sand (15 17ndash27) Both of thesegenera have robust large worms as adults and are common schis-tosomes of marine birds particularly gulls Species of Austrobil-harzia are more often implicated as a cause of dermatitis out-breaks (25)
The next main clade includes the remaining schistosomes (Fig1 clades B C and D) Clades B and C are exclusively freshwatermammalian schistosomes The largest clade of mammalian schis-tosomes includes the genus Schistosoma with 25 species (cladeB) In particular three of these species (Schistosoma mansoni
Schistosoma haematobium and Schistosoma japonicum) cause oneof the most devastating helminth diseases in humans schistoso-miasis affecting about 220 million people mainly in the tropicaland subtropical latitudes around the world (WHO) All but onespecies (S mansoni) occur exclusively in the Eastern HemisphereThese species are not typically implicated in dermatitis outbreaksyet there is a mild eruption of dermatitis following penetration byall schistosomes (28) Most reported cases of dermatitis caused bythe genus Schistosoma are from parasites that infect domesticatedwork animals such as cattle and buffalo mainly in Asia For ex-ample in countries such as India and Nepal the species Schisto-soma turkestanicum Schistosoma nasale Schistosoma indicum andSchistosoma spindale are often implicated in outbreaks of derma-titis (29ndash38) This relationship may not be a surprise as bovids arethe definitive host and the people in these areas depend uponthese animals for their livelihood in farming Additionally thesnail host for the major species causing dermatitis (S nasale Sindicum and S spindale) is Indoplanorbis exustus a widespreadand abundant snail that is found mainly in Nepal and India to theexclusion of Biomphalaria and Bulinus snail hosts for a majorityof the African transmitted species of Schistosoma
The genus Bivitellobilharzia is considered a schistosome of el-ephants but it has also been reported from wild rhinoceroses inNepal (38ndash41) There are no known reports of cercarial dermatitisin humans from areas inhabited by African elephants (with theBivitellobilharzia loxodontae schistosome) but in areas where do-mesticated Asian elephants are used there have been cases of der-matitis in the mahouts or elephant handlers when the elephantsare taken for bathing (eg in Sri Lanka [40]) In Nepal Bivitello-bilharzia nairi has thus far been found in wild not domesticatedelephants (38) The snail host remains unknown but is likely apulmonate snail (42) At least two species of Schistosoma fromBiomphalaria snails infect the African hippopotamus but thesespecies have not been implicated directly in dermatitis outbreaksdespite the presence of humans working on lakeshores wherethere are hippopotamuses (43ndash45) Given the prevalence of hu-man schistosomiasis in these areas however dermatitis caused byhippopotamus schistosomes may easily go undetected
The small clade C (Fig 1) has two species of mammalian schis-tosomes that as far as we know are found only in North Americaand are not frequently associated with dermatitis outbreaksthough they both produce a skin reaction (46ndash50) These twospecies are parasites of lymnaeid snails (often Stagnicola elodes)usually with raccoons and muskrats as mammalian hosts Schisto-somatium douthitti adults inhabit aquatic and semiaquatic ro-dents in more northern latitudes or at high elevations (48 51 52)Heterobilharzia americana has been reported from a wide range ofmammalian hosts (rivaling Schistosoma japonicum) includinghorses in the southern regions of North America (49 53 54)
Perhaps the most remarkable clade of schistosomes responsi-ble for dermatitis is clade D a large clade of avian schistosomeswhose adults are long and threadlike (except Dendritobilharziaand Bilharziella) and that includes both freshwater and marinespecies In particular the genus Trichobilharzia has achieved no-toriety as the primary etiological agent for dermatitis outbreaksaround the world The diversity of aquatic environments hostuse morphology definitive host habitat and cercarial behavior isunparalleled in any other group of schistosomes and probablymost other groups of trematodes (12 14) Figure 1 includes amolecular phylogeny of all the known genera of schistosomes ex-
Horaacutek et al
166 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cept one Jilinobilharzia has not been reported since the originalpaper reporting it from the duck Anas crecca in northeasternChina its snail host remains unknown (55) Morphological char-acteristics and host use suggest that Jilinobilharzia belongs in thelarge clade of avian schistosomes (Fig 1 clade D) perhaps even toTrichobilharzia
At the base of clade D is an unresolved group of avian schisto-somes most of which have been implicated in dermatitis out-breaks and comprise the most diverse range of both bird and snail(9 families) host use (13 40 56ndash66) Current results based on all ofthe available sequence data in GenBank for the internal tran-scribed spacer (ITS) region indicate that there are about nine dis-tinct lineages only two of which are described Gigantobilharziaand Dendritobilharzia (40 64ndash69) Most of the lineages in this part
of clade D have one to a few species and have been seen in only afew cases many related to dermatitis outbreaks (12) (Table 1)Thus far the literature suggests that species in this clade causedermatitis in more local areas whereas Trichobilharzia causes cer-carial dermatitis globally For example in the San Francisco Bayarea (California) one beach in particular has annual cases of der-matitis (64) The prevalence of dermatitis caused by schistosomesfrom Valvata or Melanoides snails (63 65 66) depends on howoften people use areas where these snails release cercariae
Species of Trichobilharzia (Fig 1 clade D) are globally distrib-uted and cause the majority of recreational and occupational re-ports of dermatitis found in the literature especially in the tem-perate latitudes In North America and Europe where most of theresearch has been focused outbreaks occur in recreational ponds
FIG 1 Phylogenetic tree showing generic and species positions based on Bayesian analysis of the nuclear ribosomal DNA 28S region (1200 bp) of Schistoso-matidae Panels A to D refer to the clades discussed in the text This tree is based on genetic data not morphological data and as such there are more species thathave been described morphologically than genetically Asterisks denote significant posterior probabilities (095)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 167Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and reservoirs These outbreaks have been reviewed extensively (78 70 71) Species of Trichobilharzia have been reported to causedermatitis from other areas as well such as Rwanda-Burundi (72)South Africa (73 74) New Zealand and Australia (75ndash79) Malay-siaIndonesia (80ndash82) Iran (65 83ndash86) United Arab Emirates(66) Thailand (87) and China (88 89) in the Eastern Hemisphereand Argentina (57 90 91) Chile (11) and El Salvador (92) in theWestern Hemisphere We are just beginning to better under-stand the significant disease components for dermatitis as aglobal problem
Cercarial dermatitis is also recognized as an occupational haz-ard in many areas of the world especially in areas where rice isgrown (82 93) Rice fields are areas where snails domestic andwild ducks cattle and humans seasonally use the water so the lifecycle is maintained consistently (5 84 94ndash104) Species of Tricho-bilharzia are identified most often though not exclusively (5)Rice fields are plowed by water buffalo and cattle in areas whereIndoplanorbis exustus occurs and hence dermatitis may be causedby one of the species of Schistosoma as noted above NonethelessTrichobilharzia is still by far the most common etiological agent
MOLLUSCAN AND AVIAN HOST SPECIFICITY
Schistosomes have colonized many families of snails as first inter-mediate hosts (12 62) Mammalian schistosomes use 3 families ofsnails compared to 15 families used by avian schistosomes assummarized in Table 1 The majority of schistosome species aretransmitted by the pulmonate snail families Physidae Lymnaei-dae and Planorbidae (10 70 105) Interestingly two snail familiescontain species (eg Biomphalaria and Indoplanorbis in the Plan-orbidae family and Stagnicola in the Lymnaeidae family) that canhost both avian and mammalian schistosomes that cause derma-titis (40 51 70 104 106)
Previous papers have reviewed the details of host specificity inmammalian schistosomes (Fig 1 clades B and C) (107 108) Be-cause avian schistosomes are the major group of schistosomescausing dermatitis our own discussion focuses on their avian andsnail hosts For two reasons these avian schistosome species com-prise most of the dermatitis reports first many avian hosts sea-sonally migrate consequently disseminating avian schistosomesas they fly (domestic ducks can serve as definitive hosts particu-
TABLE 1 Summary of general host use of known genera of schistosomes reflecting current knowledge habitat in the definitive host and broadgeographic locality
Genus Snail host Mammalianavian host
Definitivehosthabitat Locality
Aquatichabitat
Major areas foroutbreaks
Austrobilharzia Nassariidae BatillaridaeLittoriniidaePotamididae
Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Ornithobilharzia Batillaridae Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Macrobilharzia Unknown Suliformes (Anhinga) Visceral NorthAmericaAfrica
Unknown Unknown if causesdermatitis
Bivitellobilharzia Unknown Elephantidae Rhinocerotidae Visceral Africa Asia Freshwater Probably freshwaterrivers
Schistosoma PlanorbidaeLymnaeidaePomatiopsidae
Mammalia VisceralNasal
Eurasia AfricaSouthAmerica
Freshwater Mostly eutrophic ponds
Heterobilharzia Lymnaeidae Mammalia Visceral North America Freshwater Marshy areasSchistosomatium Lymnaeidae Rodentia Visceral North America Freshwater Marshy areasBilharziella Planorbidae Anseriformes Gruiformes
CiconiformesPodicipediformes
Visceral Europe Freshwater Eutrophic ponds
Species isolatedfromHaminoea
Haminoeidae CharadriiformesPelicaniformes
Visceral North America Marine Shallow marine areastidal pools
Gigantobilharziaa Physidae Passeriformes Visceral North America Freshwater Marshy areas usuallywith cattails
Dendritobilharzia Planorbidae Anseriformes GruiformesPelicaniformesGaviiformes
Visceral Global Freshwater Unknown if reports ofdermatitis
Jilinobilharzia Unknown Anseriformes (Anatidae) Visceral China Unknown Unknown if reports ofdermatitis
Allobilharzia Unknown Anseriformes (swans) Visceral NorthernHemisphere
Unknown Unknown if causesdermatitis
Anserobilharzia Planorbidae Anseriformes (geese) Visceral NorthernHemisphere
Freshwater Eutrophic pondsreservoirs
Trichobilharzia Lymnaeidae Physidae Anseriformes (Anatidae) Visceralnasal
Global Freshwater Eutrophic ponds glaciallakes reservoirs
a Since Gigantobilharzia is not a monophyletic genus the information listed here is for G huronensis only
Horaacutek et al
168 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
larly for Trichobilharzia) and second some snail hosts are habitatgeneralists (eg Physa [syn Physella] acuta and Lymnaea stagna-lis) that are now globally distributed As a result the opportunitiesfor birds and snails to come into contact across time and space arevast For example P acuta (host to Gigantobilharzia huronensisTrichobilharzia physellae and Trichobilharzia querquedulae)thrives in both natural and altered environments with a widetolerance for water temperature and chemistry including the con-ditions found in ponds drainage ditches rivers marshes andephemeral water (109ndash111) In Europe the lymnaeid snail L stag-nalis (host of Trichobilharzia szidati) has also been linked to casesof dermatitis in people who acquired it while working withaquaria (112 113) providing evidence of the snailrsquos ability to per-sist in addition to its local global presence
From an evolutionary perspective the vagility and habitatspecificity of most bird hosts in concert with the availability ofsnails in aquatic habitats are likely mechanisms for widespreadhost switching in snails and thus for diversification of avian schis-tosomes (12 114) Our knowledge of the current schistosome-snail associations indicates that once a schistosome is hosted by aparticular species of snail it possesses little ability to utilize morethan a few species within that genus (eg Radix Stagnicola andPhysa) (115) Two exceptions are Dendritobilharzia pulverulentawhich uses Anisus vortex in Europe (116) and Gyraulus parvus inNorth America (117) both of which are small related planorbidsnails and Trichobilharzia regenti which employs lymnaeid snailsof the genus Radix in Europe and Austropeplea tomentosa in NewZealand (118) There is little evidence of schistosome speciescrossing snail families naturally or experimentally (for an excep-tion see references 119 and 120) Trichobilharzia franki was re-ported to be widespread across Europe but detailed molecularstudies are showing that it may represent several species related tosnail host use (70 121 122)
Cercarial dermatitis is caused not only by species of schisto-somes from indigenous snails but also by those from invasive orintroduced snails For example Haminoea japonica (originallyfrom Japan but now off the California coast) and Ilyanassa obso-leta (originally from the east coast but now on the west coast ofNorth America) are responsible for annual dermatitis outbreaks atmarine swimming beaches (25 64) In freshwater L stagnalis isfound commonly in the northern Eastern hemisphere yet in thenorthern Western hemisphere it is only locally common When Lstagnalis is found to be infected the infecting species is related to acommon European species Trichobilharzia szidati (70 123) In-terestingly P acuta is one of the most invasive pulmonate snailsand can host at least four species of avian schistosomes in NorthAmerica (10 70) yet there are no reports of this snail hostingschistosomes in their invasive range (outside North America) It isalso noteworthy that most of the schistosome species transmittedby physid snails have thus far been found only in North America(at least based on genetic comparisons) (eg G huronensis Tphysellae and one undescribed lineage of schistosome [10]) Thelymnaeid snail genera Stagnicola (found in North America) andRadix (found in the Eastern Hemisphere) are the main snail hostsfor most species of Trichobilharzia in fact thus far Radix main-tains most of the reported species diversity of Trichobilharzia(122) Trichobilharzia regenti has been recognized to cause derma-titis in Lake Wanaka in New Zealand and it may have been intro-duced from Europe in wild duck breeds (Anas platyrhynchos) usedfor hunting It is now found commonly in the endemic nonmigra-
tory scaup Aythya novaeseelandiae and the snail Austropeplea to-mentosa (118) Schistosomes seem to be specific to particular snailhosts at the species or genus level but not as much to their avian ormammalian hosts though loose specificity of definitive host useexists at higher taxonomic levels (Table 1) (eg Schistosomahaematobium in humans Bivitellobilharzia in elephants and Allo-bilharzia in swans)
The diversity of avian schistosomes found around the world isin no small part due to the ability of thousands of migratory birdsto carry their parasites across several latitudes and longitudes ex-posing commonly encountered snails This propensity to migratelarge distances distinguishes avian schistosomes from mammalianschistosomes in terms of distribution diversification and hostuse perhaps with the exception of Schistosoma mansoni (in termsof long-distance migration only) (124) Yet the schistosome spe-cies found in a wide range of avian host orders (eg Bilharziellaand Dendritobilharzia) (Table 1) are not the ones recurrently re-sponsible for outbreaks and are also species that are not geneticallydiverse compared to other species (69 125 126) Currently Dpulverulenta might be the most widespread single species of avianschistosome crossing both the Northern and Southern Hemi-spheres (117 125 127 128)
The most derived clade or most recently evolved clade in cladeD has three genera Allobilharzia Anserobilharzia and Trichobil-harzia (Fig 1) Allobilharzia from swans and Anserobilharziafrom geese both have a circumpolar distribution (69 70 105 129130) Anserobilharzia brantae which is common in North Amer-ica (in the Canada goose [Branta canadensis] and the snow goose[Chen caerulescens]) but also found in Europe (in the greylaggoose [Anser anser]) has been identified in at least one outbreak ofdermatitis in the United States (10) The area was a eutrophicmunicipal lakepond with a dense population of Gyraulus parvussnails and Canada geese The third genus Trichobilharzia themost species-rich genus in the family is found almost exclusivelyin ducks It should be noted that there are several species of Tricho-bilharzia reported from other avian families (Trichobilharzia corvifrom passeriforms along with other species [131 132]) but basedon the morphology of adults and eggs and snail host use (whenknown) these probably represent new genera or these avian hostsare not the primary (competent) hosts or might be aberrant cases(70 78) Within the genus Trichobilharzia several clades are spe-cific to certain groups of ducks for example Trichobilharzia stag-nicolae and Trichobilharzia mergi are found in mergansers (Mer-ginae) (70 133) T querquedulae in the ldquoblue-winged duckrdquo clade(Anas clypeata Anas discors and Anas cyanoptera) (70 134) Tphysellae in an ecological group of diving ducks that includesducks of the Aythinae and Merginae and a common undescribedspecies of Trichobilharzia species A in Anas americana (70) It isnot yet clear which duck groups (phylogenetic or ecological) aremore specific for T franki or T szidati Interestingly T regenti hasprobably been reported from the most diverse duck species (135)and does not appear to have a preferred host within the AnatidaeIn North America T stagnicolae and T physellae are most oftenidentified in dermatitis outbreaks (70)
INTRAMOLLUSCAN DEVELOPMENT OF AVIANSCHISTOSOMES
As noted above Trichobilharzia is the most diverse schistosomegenus and has most often been implicated in outbreaks of cercarialdermatitis As a result studies on the avian schistosome-snail in-
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 169Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
termediate host relationship have focused primarily on species ofthe genus Trichobilharzia Additionally long-term laboratorymaintenance of T szidati and L stagnalis enabled the experimentsthat uncovered the intimate molecular interactions of avian schis-tosomes and their snail hosts (The Trichobilharzia ocellata organ-ism used as an experimental model in European laboratories forthe last few decades is identical to T szidati and the latter name ispreferred and is used here [123] If T ocellata is used in the textbody then it refers to the non-European isolates of the parasite)
After hatching from eggs in an aquatic environment schisto-some miracidia search for and invade an appropriate snail hostspecies This behavior must be accomplished quickly as miracidiahave a temperature-dependent limited life span of around 20 h asreported for eg T stagnicolae (136) Studies on the miracidialbehavior of T szidati have shown a progression of steps from hostfinding to penetration and migration Miracidia respond to envi-ronmental stimuli such as light or gravity that direct them to themicrohabitat occupied by the host snails (137) Snails release var-ious chemical compounds that form an ldquoactive spacerdquo aroundthem and serve as chemoattractants for the miracidia Miracidiarecognize macromolecular glycoconjugates termed miracidium-attracting glycoproteins (MAGs) or miraxones that consist of aprotein core and carbohydrate chains linked O-glycosidically viaN-acetyl-D-galactosamine and serinethreonine (138 139) Theattractant for miracidia is encoded in these carbohydrate moietiesUpon entering the ldquoactive spacerdquo of the snail miracidia modifytheir movement by increased random turns within the increasingattractant gradient and by a turn-back form of swimming withinthe decreasing attractant gradient (140) This mode of orientation(chemokinesis) observed eg in T szidati or T franki (115 138)results in the first contact of the miracidia with the snail which isthen followed by repeated investigationprobing of the snail sur-face and finally miracidial attachment (140) Penetration of thesnail surface follows although the factors contributing to this pro-cess such as the components of miracidial penetration glandsremain largely unknown
After penetrating the surface epithelium of the snail miracidiaof avian schistosomes transform into mother sporocysts that giverise to daughter sporocysts which migrate to the snail hepatopan-creas where production of the final larval stage the cercariaetakes place (13) The prepatent period lasts about 3 to 10 weeksdepending on several factors such as the miracidial dose or tem-perature (13 141) Infection by avian schistosomes may lead toalterations of the snail internal defense system (IDS) metabolismand endocrine functions Such alterations have been studiedwidely in L stagnalis the intermediate host of T szidati (142ndash145)
The snail IDS is based solely on innate immune mechanismscomposed of humoral and cellular limbs Lectins are essential hu-moral components whereas hemocytes represent the main effec-tor cells (146 147) Both limbs of the L stagnalis IDS appear to beactivated and then suppressed during early and late stages respec-tively of a T szidati infection (143) In vitro hemocytes failed toencapsulate and destroy T szidati sporocysts (148) It has beensuggested that parasite excretory-secretory products participate inthe modulation of hemocyte activities (149) Disruption of hemo-cyte signaling pathways such as protein kinase C (PKC) and ex-tracellular signal-regulated kinase (ERK) pathways may alsoinfluence hemocyte activities (150ndash152) For example carbohy-drates known to be present on larval surfaces of T szidati and Tregenti such as D-galactose and L-fucose (153ndash156) affected he-
mocyte PKC and ERK signaling in L stagnalis which suggests animmunosuppressive role (157 158) However experiments inves-tigating the direct effect of Trichobilharzia larvae on hemocytesignaling have not been performed Alterations of humoral de-fense components also occur during infections by avian schisto-somes and at least two molecules molluscan defense molecule(MDM) and granularin have been investigated in this respect(159 160) Both molecules produced in L stagnalis by granularcells of connective tissue are related to phagocytic activity of he-mocytes MDM enhances phagocytosis of hemocytes and the ex-pression of a corresponding gene for MDM is downregulated in Lstagnalis infected with T szidati (159 161) In contrast treatmentof hemocytes with granularin decreases phagocytic activity andthe encoding gene is upregulated in parasitized snails (160 161)
In the snail host avian schistosomes also interfere with metab-olism such as causing abnormal body growth and endocrinefunctions such as causing a reduction of egg laying (161) In Lstagnalis these processes are regulated by neuroendocrine cells inthe central nervous system (CNS) (162 163) Trichobilharzia szi-dati releases an undescribed substance that induces the snail hostto produce schistosomin (a peptide of 87 kDa consisting of 79amino acids) from its connective tissue and hemocytes (145)Schistosomin acts as a neuropeptide that interferes with somehormones such as calfluxin a neuropeptide that stimulates theinflux of Ca2 into the mitochondria of albumen gland cells (164165) As a consequence ovulation and egg laying are inhibited inthe snail Excitability of neuroendocrine cells (light green cells[LGCs]) responsible for growth (162) increases in response toschistosomin (166) As a result the body size of infected snailsmay become considerably larger than that of uninfected snails(142 161) Other neuropeptides (FMRFamide-related peptides)are also upregulated during infection of L stagnalis by T szidatiand these peptides via inhibition of neuroendocrine cells may beresponsible for the suppression of snail metabolism and repro-duction (167) All these changes in infected snails may provideenergy resources and space that can be exploited by the schisto-somes for development (145 161)
The survival rate of infected snails releasing schistosome cer-cariae as the agent of cercarial dermatitis varies among species Alimited number of studies have focused on survival rates of avianschistosome-infected versus uninfected snails As an example90 of L stagnalis snails infected experimentally with a singlemiracidium of T ocellata (North American isolate) were alive at28 weeks of age (three infected snails were alive for 19 months)whereas all uninfected snails were dead (168) In contrast L stag-nalis or Planorbarius corneus snails naturally infected with T szi-dati or Bilharziella polonica respectively lived a shorter time onaverage than the corresponding uninfected individuals (169170)
VERTEBRATE HOST FINDING AND PENETRATION
Cercariae emerging from the snail intermediate host are the infec-tive stage to the definitive host and are also the stage responsiblefor causing cercarial dermatitis A cercaria is a multicellular larvacomprised of an oblong body and a slender tail that is bifurcated(furcocercous) at the posterior end (Fig 2) Cercariae of schisto-somes leave their snail hosts actively For this purpose they em-ploy a pair of specialized unicellular escape glands that are obviousin mature cercariae within sporocysts Once the cercariae haveemerged only their ducts lined by microtubules are visible sug-
Horaacutek et al
170 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

cupational disease among rice farmers (4) and may incur costs interms of lost person work hours Older reports refer to 75 ormore of the population experiencing the characteristic symptomsof ldquokoganbyordquo in the areas of Japan where the disease is mosthighly endemic (5) Recent reviews (6ndash9) agree that in some re-gions cercarial dermatitis has appeared as a new problem eitherbecause the dermatitis was previously unknown (eg the USSouthwest and Chile) or because the number of reports of out-breaks increased (8 10 11) Consequently cercarial dermatitis isnow regarded as an emerging disease Besides human schisto-somes (Schistosoma spp) no animal (eg avian) schistosomeshave any other presently known pathogenic effects on humansThus the use of animal models to study the potential risk of ani-mal (avian) schistosomes to human health is invaluable
The last decade has revealed diverse avian schistosome speciesand biology as well as the snails that host them These discoverieshave outpaced the equally essential host-parasite biological im-munological pathological and epidemiological studies of speciesdiversity in terms of incorporating the results of such studies intothe current known diversity of schistosomes Such studies are dif-ficult and time-consuming and consequently only a few specieshave been adapted to experimental conditions Nevertheless suchstudies are crucial to understanding the current and future rolesthat these species might play in the frequency and distribution ofcercarial dermatitis as well as understanding how to break the lifecycle to prevent outbreaks What has been documented and isdetailed in the following sections points to an understanding ofthe avian schistosome-host relationships and thus offers the foun-dation on which future studies will be modeled
DIVERSITY OF SCHISTOSOMES CAUSING DERMATITIS
Considering only the named species in the literature there are 4schistosome genera from mammals and 10 from birds with about30 described species from mammals and about 67 from birds (12)The total is close to 100 species with 70 of them being avianschistosomes distributed around the world that may initiate cer-carial dermatitis The role of some of the species of avian schisto-somes as dermatitis agents has not been studied sufficiently asthey are not often found in areas where people most commonlyare in contact with water and snails To discuss the distributionand diversity of schistosomes causing dermatitis within the phy-logenetic framework of the family Schistosomatidae we refer toFig 1 (12ndash14)
The basal clade of the family tree (Fig 1 clade A) comprises theexclusively marine avian schistosomes Austrobilharzia (4 species)and Ornithobilharzia (2 species) the species shown in the tree arethose for which there are genetic data Species of these two generaare associated with outbreaks of dermatitis in shallow marine en-vironments (15 16) Infection often occurs in people who areswimming playing in tidal pools or working for example col-lecting tidal invertebrates in the sand (15 17ndash27) Both of thesegenera have robust large worms as adults and are common schis-tosomes of marine birds particularly gulls Species of Austrobil-harzia are more often implicated as a cause of dermatitis out-breaks (25)
The next main clade includes the remaining schistosomes (Fig1 clades B C and D) Clades B and C are exclusively freshwatermammalian schistosomes The largest clade of mammalian schis-tosomes includes the genus Schistosoma with 25 species (cladeB) In particular three of these species (Schistosoma mansoni
Schistosoma haematobium and Schistosoma japonicum) cause oneof the most devastating helminth diseases in humans schistoso-miasis affecting about 220 million people mainly in the tropicaland subtropical latitudes around the world (WHO) All but onespecies (S mansoni) occur exclusively in the Eastern HemisphereThese species are not typically implicated in dermatitis outbreaksyet there is a mild eruption of dermatitis following penetration byall schistosomes (28) Most reported cases of dermatitis caused bythe genus Schistosoma are from parasites that infect domesticatedwork animals such as cattle and buffalo mainly in Asia For ex-ample in countries such as India and Nepal the species Schisto-soma turkestanicum Schistosoma nasale Schistosoma indicum andSchistosoma spindale are often implicated in outbreaks of derma-titis (29ndash38) This relationship may not be a surprise as bovids arethe definitive host and the people in these areas depend uponthese animals for their livelihood in farming Additionally thesnail host for the major species causing dermatitis (S nasale Sindicum and S spindale) is Indoplanorbis exustus a widespreadand abundant snail that is found mainly in Nepal and India to theexclusion of Biomphalaria and Bulinus snail hosts for a majorityof the African transmitted species of Schistosoma
The genus Bivitellobilharzia is considered a schistosome of el-ephants but it has also been reported from wild rhinoceroses inNepal (38ndash41) There are no known reports of cercarial dermatitisin humans from areas inhabited by African elephants (with theBivitellobilharzia loxodontae schistosome) but in areas where do-mesticated Asian elephants are used there have been cases of der-matitis in the mahouts or elephant handlers when the elephantsare taken for bathing (eg in Sri Lanka [40]) In Nepal Bivitello-bilharzia nairi has thus far been found in wild not domesticatedelephants (38) The snail host remains unknown but is likely apulmonate snail (42) At least two species of Schistosoma fromBiomphalaria snails infect the African hippopotamus but thesespecies have not been implicated directly in dermatitis outbreaksdespite the presence of humans working on lakeshores wherethere are hippopotamuses (43ndash45) Given the prevalence of hu-man schistosomiasis in these areas however dermatitis caused byhippopotamus schistosomes may easily go undetected
The small clade C (Fig 1) has two species of mammalian schis-tosomes that as far as we know are found only in North Americaand are not frequently associated with dermatitis outbreaksthough they both produce a skin reaction (46ndash50) These twospecies are parasites of lymnaeid snails (often Stagnicola elodes)usually with raccoons and muskrats as mammalian hosts Schisto-somatium douthitti adults inhabit aquatic and semiaquatic ro-dents in more northern latitudes or at high elevations (48 51 52)Heterobilharzia americana has been reported from a wide range ofmammalian hosts (rivaling Schistosoma japonicum) includinghorses in the southern regions of North America (49 53 54)
Perhaps the most remarkable clade of schistosomes responsi-ble for dermatitis is clade D a large clade of avian schistosomeswhose adults are long and threadlike (except Dendritobilharziaand Bilharziella) and that includes both freshwater and marinespecies In particular the genus Trichobilharzia has achieved no-toriety as the primary etiological agent for dermatitis outbreaksaround the world The diversity of aquatic environments hostuse morphology definitive host habitat and cercarial behavior isunparalleled in any other group of schistosomes and probablymost other groups of trematodes (12 14) Figure 1 includes amolecular phylogeny of all the known genera of schistosomes ex-
Horaacutek et al
166 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cept one Jilinobilharzia has not been reported since the originalpaper reporting it from the duck Anas crecca in northeasternChina its snail host remains unknown (55) Morphological char-acteristics and host use suggest that Jilinobilharzia belongs in thelarge clade of avian schistosomes (Fig 1 clade D) perhaps even toTrichobilharzia
At the base of clade D is an unresolved group of avian schisto-somes most of which have been implicated in dermatitis out-breaks and comprise the most diverse range of both bird and snail(9 families) host use (13 40 56ndash66) Current results based on all ofthe available sequence data in GenBank for the internal tran-scribed spacer (ITS) region indicate that there are about nine dis-tinct lineages only two of which are described Gigantobilharziaand Dendritobilharzia (40 64ndash69) Most of the lineages in this part
of clade D have one to a few species and have been seen in only afew cases many related to dermatitis outbreaks (12) (Table 1)Thus far the literature suggests that species in this clade causedermatitis in more local areas whereas Trichobilharzia causes cer-carial dermatitis globally For example in the San Francisco Bayarea (California) one beach in particular has annual cases of der-matitis (64) The prevalence of dermatitis caused by schistosomesfrom Valvata or Melanoides snails (63 65 66) depends on howoften people use areas where these snails release cercariae
Species of Trichobilharzia (Fig 1 clade D) are globally distrib-uted and cause the majority of recreational and occupational re-ports of dermatitis found in the literature especially in the tem-perate latitudes In North America and Europe where most of theresearch has been focused outbreaks occur in recreational ponds
FIG 1 Phylogenetic tree showing generic and species positions based on Bayesian analysis of the nuclear ribosomal DNA 28S region (1200 bp) of Schistoso-matidae Panels A to D refer to the clades discussed in the text This tree is based on genetic data not morphological data and as such there are more species thathave been described morphologically than genetically Asterisks denote significant posterior probabilities (095)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 167Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and reservoirs These outbreaks have been reviewed extensively (78 70 71) Species of Trichobilharzia have been reported to causedermatitis from other areas as well such as Rwanda-Burundi (72)South Africa (73 74) New Zealand and Australia (75ndash79) Malay-siaIndonesia (80ndash82) Iran (65 83ndash86) United Arab Emirates(66) Thailand (87) and China (88 89) in the Eastern Hemisphereand Argentina (57 90 91) Chile (11) and El Salvador (92) in theWestern Hemisphere We are just beginning to better under-stand the significant disease components for dermatitis as aglobal problem
Cercarial dermatitis is also recognized as an occupational haz-ard in many areas of the world especially in areas where rice isgrown (82 93) Rice fields are areas where snails domestic andwild ducks cattle and humans seasonally use the water so the lifecycle is maintained consistently (5 84 94ndash104) Species of Tricho-bilharzia are identified most often though not exclusively (5)Rice fields are plowed by water buffalo and cattle in areas whereIndoplanorbis exustus occurs and hence dermatitis may be causedby one of the species of Schistosoma as noted above NonethelessTrichobilharzia is still by far the most common etiological agent
MOLLUSCAN AND AVIAN HOST SPECIFICITY
Schistosomes have colonized many families of snails as first inter-mediate hosts (12 62) Mammalian schistosomes use 3 families ofsnails compared to 15 families used by avian schistosomes assummarized in Table 1 The majority of schistosome species aretransmitted by the pulmonate snail families Physidae Lymnaei-dae and Planorbidae (10 70 105) Interestingly two snail familiescontain species (eg Biomphalaria and Indoplanorbis in the Plan-orbidae family and Stagnicola in the Lymnaeidae family) that canhost both avian and mammalian schistosomes that cause derma-titis (40 51 70 104 106)
Previous papers have reviewed the details of host specificity inmammalian schistosomes (Fig 1 clades B and C) (107 108) Be-cause avian schistosomes are the major group of schistosomescausing dermatitis our own discussion focuses on their avian andsnail hosts For two reasons these avian schistosome species com-prise most of the dermatitis reports first many avian hosts sea-sonally migrate consequently disseminating avian schistosomesas they fly (domestic ducks can serve as definitive hosts particu-
TABLE 1 Summary of general host use of known genera of schistosomes reflecting current knowledge habitat in the definitive host and broadgeographic locality
Genus Snail host Mammalianavian host
Definitivehosthabitat Locality
Aquatichabitat
Major areas foroutbreaks
Austrobilharzia Nassariidae BatillaridaeLittoriniidaePotamididae
Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Ornithobilharzia Batillaridae Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Macrobilharzia Unknown Suliformes (Anhinga) Visceral NorthAmericaAfrica
Unknown Unknown if causesdermatitis
Bivitellobilharzia Unknown Elephantidae Rhinocerotidae Visceral Africa Asia Freshwater Probably freshwaterrivers
Schistosoma PlanorbidaeLymnaeidaePomatiopsidae
Mammalia VisceralNasal
Eurasia AfricaSouthAmerica
Freshwater Mostly eutrophic ponds
Heterobilharzia Lymnaeidae Mammalia Visceral North America Freshwater Marshy areasSchistosomatium Lymnaeidae Rodentia Visceral North America Freshwater Marshy areasBilharziella Planorbidae Anseriformes Gruiformes
CiconiformesPodicipediformes
Visceral Europe Freshwater Eutrophic ponds
Species isolatedfromHaminoea
Haminoeidae CharadriiformesPelicaniformes
Visceral North America Marine Shallow marine areastidal pools
Gigantobilharziaa Physidae Passeriformes Visceral North America Freshwater Marshy areas usuallywith cattails
Dendritobilharzia Planorbidae Anseriformes GruiformesPelicaniformesGaviiformes
Visceral Global Freshwater Unknown if reports ofdermatitis
Jilinobilharzia Unknown Anseriformes (Anatidae) Visceral China Unknown Unknown if reports ofdermatitis
Allobilharzia Unknown Anseriformes (swans) Visceral NorthernHemisphere
Unknown Unknown if causesdermatitis
Anserobilharzia Planorbidae Anseriformes (geese) Visceral NorthernHemisphere
Freshwater Eutrophic pondsreservoirs
Trichobilharzia Lymnaeidae Physidae Anseriformes (Anatidae) Visceralnasal
Global Freshwater Eutrophic ponds glaciallakes reservoirs
a Since Gigantobilharzia is not a monophyletic genus the information listed here is for G huronensis only
Horaacutek et al
168 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
larly for Trichobilharzia) and second some snail hosts are habitatgeneralists (eg Physa [syn Physella] acuta and Lymnaea stagna-lis) that are now globally distributed As a result the opportunitiesfor birds and snails to come into contact across time and space arevast For example P acuta (host to Gigantobilharzia huronensisTrichobilharzia physellae and Trichobilharzia querquedulae)thrives in both natural and altered environments with a widetolerance for water temperature and chemistry including the con-ditions found in ponds drainage ditches rivers marshes andephemeral water (109ndash111) In Europe the lymnaeid snail L stag-nalis (host of Trichobilharzia szidati) has also been linked to casesof dermatitis in people who acquired it while working withaquaria (112 113) providing evidence of the snailrsquos ability to per-sist in addition to its local global presence
From an evolutionary perspective the vagility and habitatspecificity of most bird hosts in concert with the availability ofsnails in aquatic habitats are likely mechanisms for widespreadhost switching in snails and thus for diversification of avian schis-tosomes (12 114) Our knowledge of the current schistosome-snail associations indicates that once a schistosome is hosted by aparticular species of snail it possesses little ability to utilize morethan a few species within that genus (eg Radix Stagnicola andPhysa) (115) Two exceptions are Dendritobilharzia pulverulentawhich uses Anisus vortex in Europe (116) and Gyraulus parvus inNorth America (117) both of which are small related planorbidsnails and Trichobilharzia regenti which employs lymnaeid snailsof the genus Radix in Europe and Austropeplea tomentosa in NewZealand (118) There is little evidence of schistosome speciescrossing snail families naturally or experimentally (for an excep-tion see references 119 and 120) Trichobilharzia franki was re-ported to be widespread across Europe but detailed molecularstudies are showing that it may represent several species related tosnail host use (70 121 122)
Cercarial dermatitis is caused not only by species of schisto-somes from indigenous snails but also by those from invasive orintroduced snails For example Haminoea japonica (originallyfrom Japan but now off the California coast) and Ilyanassa obso-leta (originally from the east coast but now on the west coast ofNorth America) are responsible for annual dermatitis outbreaks atmarine swimming beaches (25 64) In freshwater L stagnalis isfound commonly in the northern Eastern hemisphere yet in thenorthern Western hemisphere it is only locally common When Lstagnalis is found to be infected the infecting species is related to acommon European species Trichobilharzia szidati (70 123) In-terestingly P acuta is one of the most invasive pulmonate snailsand can host at least four species of avian schistosomes in NorthAmerica (10 70) yet there are no reports of this snail hostingschistosomes in their invasive range (outside North America) It isalso noteworthy that most of the schistosome species transmittedby physid snails have thus far been found only in North America(at least based on genetic comparisons) (eg G huronensis Tphysellae and one undescribed lineage of schistosome [10]) Thelymnaeid snail genera Stagnicola (found in North America) andRadix (found in the Eastern Hemisphere) are the main snail hostsfor most species of Trichobilharzia in fact thus far Radix main-tains most of the reported species diversity of Trichobilharzia(122) Trichobilharzia regenti has been recognized to cause derma-titis in Lake Wanaka in New Zealand and it may have been intro-duced from Europe in wild duck breeds (Anas platyrhynchos) usedfor hunting It is now found commonly in the endemic nonmigra-
tory scaup Aythya novaeseelandiae and the snail Austropeplea to-mentosa (118) Schistosomes seem to be specific to particular snailhosts at the species or genus level but not as much to their avian ormammalian hosts though loose specificity of definitive host useexists at higher taxonomic levels (Table 1) (eg Schistosomahaematobium in humans Bivitellobilharzia in elephants and Allo-bilharzia in swans)
The diversity of avian schistosomes found around the world isin no small part due to the ability of thousands of migratory birdsto carry their parasites across several latitudes and longitudes ex-posing commonly encountered snails This propensity to migratelarge distances distinguishes avian schistosomes from mammalianschistosomes in terms of distribution diversification and hostuse perhaps with the exception of Schistosoma mansoni (in termsof long-distance migration only) (124) Yet the schistosome spe-cies found in a wide range of avian host orders (eg Bilharziellaand Dendritobilharzia) (Table 1) are not the ones recurrently re-sponsible for outbreaks and are also species that are not geneticallydiverse compared to other species (69 125 126) Currently Dpulverulenta might be the most widespread single species of avianschistosome crossing both the Northern and Southern Hemi-spheres (117 125 127 128)
The most derived clade or most recently evolved clade in cladeD has three genera Allobilharzia Anserobilharzia and Trichobil-harzia (Fig 1) Allobilharzia from swans and Anserobilharziafrom geese both have a circumpolar distribution (69 70 105 129130) Anserobilharzia brantae which is common in North Amer-ica (in the Canada goose [Branta canadensis] and the snow goose[Chen caerulescens]) but also found in Europe (in the greylaggoose [Anser anser]) has been identified in at least one outbreak ofdermatitis in the United States (10) The area was a eutrophicmunicipal lakepond with a dense population of Gyraulus parvussnails and Canada geese The third genus Trichobilharzia themost species-rich genus in the family is found almost exclusivelyin ducks It should be noted that there are several species of Tricho-bilharzia reported from other avian families (Trichobilharzia corvifrom passeriforms along with other species [131 132]) but basedon the morphology of adults and eggs and snail host use (whenknown) these probably represent new genera or these avian hostsare not the primary (competent) hosts or might be aberrant cases(70 78) Within the genus Trichobilharzia several clades are spe-cific to certain groups of ducks for example Trichobilharzia stag-nicolae and Trichobilharzia mergi are found in mergansers (Mer-ginae) (70 133) T querquedulae in the ldquoblue-winged duckrdquo clade(Anas clypeata Anas discors and Anas cyanoptera) (70 134) Tphysellae in an ecological group of diving ducks that includesducks of the Aythinae and Merginae and a common undescribedspecies of Trichobilharzia species A in Anas americana (70) It isnot yet clear which duck groups (phylogenetic or ecological) aremore specific for T franki or T szidati Interestingly T regenti hasprobably been reported from the most diverse duck species (135)and does not appear to have a preferred host within the AnatidaeIn North America T stagnicolae and T physellae are most oftenidentified in dermatitis outbreaks (70)
INTRAMOLLUSCAN DEVELOPMENT OF AVIANSCHISTOSOMES
As noted above Trichobilharzia is the most diverse schistosomegenus and has most often been implicated in outbreaks of cercarialdermatitis As a result studies on the avian schistosome-snail in-
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 169Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
termediate host relationship have focused primarily on species ofthe genus Trichobilharzia Additionally long-term laboratorymaintenance of T szidati and L stagnalis enabled the experimentsthat uncovered the intimate molecular interactions of avian schis-tosomes and their snail hosts (The Trichobilharzia ocellata organ-ism used as an experimental model in European laboratories forthe last few decades is identical to T szidati and the latter name ispreferred and is used here [123] If T ocellata is used in the textbody then it refers to the non-European isolates of the parasite)
After hatching from eggs in an aquatic environment schisto-some miracidia search for and invade an appropriate snail hostspecies This behavior must be accomplished quickly as miracidiahave a temperature-dependent limited life span of around 20 h asreported for eg T stagnicolae (136) Studies on the miracidialbehavior of T szidati have shown a progression of steps from hostfinding to penetration and migration Miracidia respond to envi-ronmental stimuli such as light or gravity that direct them to themicrohabitat occupied by the host snails (137) Snails release var-ious chemical compounds that form an ldquoactive spacerdquo aroundthem and serve as chemoattractants for the miracidia Miracidiarecognize macromolecular glycoconjugates termed miracidium-attracting glycoproteins (MAGs) or miraxones that consist of aprotein core and carbohydrate chains linked O-glycosidically viaN-acetyl-D-galactosamine and serinethreonine (138 139) Theattractant for miracidia is encoded in these carbohydrate moietiesUpon entering the ldquoactive spacerdquo of the snail miracidia modifytheir movement by increased random turns within the increasingattractant gradient and by a turn-back form of swimming withinthe decreasing attractant gradient (140) This mode of orientation(chemokinesis) observed eg in T szidati or T franki (115 138)results in the first contact of the miracidia with the snail which isthen followed by repeated investigationprobing of the snail sur-face and finally miracidial attachment (140) Penetration of thesnail surface follows although the factors contributing to this pro-cess such as the components of miracidial penetration glandsremain largely unknown
After penetrating the surface epithelium of the snail miracidiaof avian schistosomes transform into mother sporocysts that giverise to daughter sporocysts which migrate to the snail hepatopan-creas where production of the final larval stage the cercariaetakes place (13) The prepatent period lasts about 3 to 10 weeksdepending on several factors such as the miracidial dose or tem-perature (13 141) Infection by avian schistosomes may lead toalterations of the snail internal defense system (IDS) metabolismand endocrine functions Such alterations have been studiedwidely in L stagnalis the intermediate host of T szidati (142ndash145)
The snail IDS is based solely on innate immune mechanismscomposed of humoral and cellular limbs Lectins are essential hu-moral components whereas hemocytes represent the main effec-tor cells (146 147) Both limbs of the L stagnalis IDS appear to beactivated and then suppressed during early and late stages respec-tively of a T szidati infection (143) In vitro hemocytes failed toencapsulate and destroy T szidati sporocysts (148) It has beensuggested that parasite excretory-secretory products participate inthe modulation of hemocyte activities (149) Disruption of hemo-cyte signaling pathways such as protein kinase C (PKC) and ex-tracellular signal-regulated kinase (ERK) pathways may alsoinfluence hemocyte activities (150ndash152) For example carbohy-drates known to be present on larval surfaces of T szidati and Tregenti such as D-galactose and L-fucose (153ndash156) affected he-
mocyte PKC and ERK signaling in L stagnalis which suggests animmunosuppressive role (157 158) However experiments inves-tigating the direct effect of Trichobilharzia larvae on hemocytesignaling have not been performed Alterations of humoral de-fense components also occur during infections by avian schisto-somes and at least two molecules molluscan defense molecule(MDM) and granularin have been investigated in this respect(159 160) Both molecules produced in L stagnalis by granularcells of connective tissue are related to phagocytic activity of he-mocytes MDM enhances phagocytosis of hemocytes and the ex-pression of a corresponding gene for MDM is downregulated in Lstagnalis infected with T szidati (159 161) In contrast treatmentof hemocytes with granularin decreases phagocytic activity andthe encoding gene is upregulated in parasitized snails (160 161)
In the snail host avian schistosomes also interfere with metab-olism such as causing abnormal body growth and endocrinefunctions such as causing a reduction of egg laying (161) In Lstagnalis these processes are regulated by neuroendocrine cells inthe central nervous system (CNS) (162 163) Trichobilharzia szi-dati releases an undescribed substance that induces the snail hostto produce schistosomin (a peptide of 87 kDa consisting of 79amino acids) from its connective tissue and hemocytes (145)Schistosomin acts as a neuropeptide that interferes with somehormones such as calfluxin a neuropeptide that stimulates theinflux of Ca2 into the mitochondria of albumen gland cells (164165) As a consequence ovulation and egg laying are inhibited inthe snail Excitability of neuroendocrine cells (light green cells[LGCs]) responsible for growth (162) increases in response toschistosomin (166) As a result the body size of infected snailsmay become considerably larger than that of uninfected snails(142 161) Other neuropeptides (FMRFamide-related peptides)are also upregulated during infection of L stagnalis by T szidatiand these peptides via inhibition of neuroendocrine cells may beresponsible for the suppression of snail metabolism and repro-duction (167) All these changes in infected snails may provideenergy resources and space that can be exploited by the schisto-somes for development (145 161)
The survival rate of infected snails releasing schistosome cer-cariae as the agent of cercarial dermatitis varies among species Alimited number of studies have focused on survival rates of avianschistosome-infected versus uninfected snails As an example90 of L stagnalis snails infected experimentally with a singlemiracidium of T ocellata (North American isolate) were alive at28 weeks of age (three infected snails were alive for 19 months)whereas all uninfected snails were dead (168) In contrast L stag-nalis or Planorbarius corneus snails naturally infected with T szi-dati or Bilharziella polonica respectively lived a shorter time onaverage than the corresponding uninfected individuals (169170)
VERTEBRATE HOST FINDING AND PENETRATION
Cercariae emerging from the snail intermediate host are the infec-tive stage to the definitive host and are also the stage responsiblefor causing cercarial dermatitis A cercaria is a multicellular larvacomprised of an oblong body and a slender tail that is bifurcated(furcocercous) at the posterior end (Fig 2) Cercariae of schisto-somes leave their snail hosts actively For this purpose they em-ploy a pair of specialized unicellular escape glands that are obviousin mature cercariae within sporocysts Once the cercariae haveemerged only their ducts lined by microtubules are visible sug-
Horaacutek et al
170 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

cept one Jilinobilharzia has not been reported since the originalpaper reporting it from the duck Anas crecca in northeasternChina its snail host remains unknown (55) Morphological char-acteristics and host use suggest that Jilinobilharzia belongs in thelarge clade of avian schistosomes (Fig 1 clade D) perhaps even toTrichobilharzia
At the base of clade D is an unresolved group of avian schisto-somes most of which have been implicated in dermatitis out-breaks and comprise the most diverse range of both bird and snail(9 families) host use (13 40 56ndash66) Current results based on all ofthe available sequence data in GenBank for the internal tran-scribed spacer (ITS) region indicate that there are about nine dis-tinct lineages only two of which are described Gigantobilharziaand Dendritobilharzia (40 64ndash69) Most of the lineages in this part
of clade D have one to a few species and have been seen in only afew cases many related to dermatitis outbreaks (12) (Table 1)Thus far the literature suggests that species in this clade causedermatitis in more local areas whereas Trichobilharzia causes cer-carial dermatitis globally For example in the San Francisco Bayarea (California) one beach in particular has annual cases of der-matitis (64) The prevalence of dermatitis caused by schistosomesfrom Valvata or Melanoides snails (63 65 66) depends on howoften people use areas where these snails release cercariae
Species of Trichobilharzia (Fig 1 clade D) are globally distrib-uted and cause the majority of recreational and occupational re-ports of dermatitis found in the literature especially in the tem-perate latitudes In North America and Europe where most of theresearch has been focused outbreaks occur in recreational ponds
FIG 1 Phylogenetic tree showing generic and species positions based on Bayesian analysis of the nuclear ribosomal DNA 28S region (1200 bp) of Schistoso-matidae Panels A to D refer to the clades discussed in the text This tree is based on genetic data not morphological data and as such there are more species thathave been described morphologically than genetically Asterisks denote significant posterior probabilities (095)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 167Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and reservoirs These outbreaks have been reviewed extensively (78 70 71) Species of Trichobilharzia have been reported to causedermatitis from other areas as well such as Rwanda-Burundi (72)South Africa (73 74) New Zealand and Australia (75ndash79) Malay-siaIndonesia (80ndash82) Iran (65 83ndash86) United Arab Emirates(66) Thailand (87) and China (88 89) in the Eastern Hemisphereand Argentina (57 90 91) Chile (11) and El Salvador (92) in theWestern Hemisphere We are just beginning to better under-stand the significant disease components for dermatitis as aglobal problem
Cercarial dermatitis is also recognized as an occupational haz-ard in many areas of the world especially in areas where rice isgrown (82 93) Rice fields are areas where snails domestic andwild ducks cattle and humans seasonally use the water so the lifecycle is maintained consistently (5 84 94ndash104) Species of Tricho-bilharzia are identified most often though not exclusively (5)Rice fields are plowed by water buffalo and cattle in areas whereIndoplanorbis exustus occurs and hence dermatitis may be causedby one of the species of Schistosoma as noted above NonethelessTrichobilharzia is still by far the most common etiological agent
MOLLUSCAN AND AVIAN HOST SPECIFICITY
Schistosomes have colonized many families of snails as first inter-mediate hosts (12 62) Mammalian schistosomes use 3 families ofsnails compared to 15 families used by avian schistosomes assummarized in Table 1 The majority of schistosome species aretransmitted by the pulmonate snail families Physidae Lymnaei-dae and Planorbidae (10 70 105) Interestingly two snail familiescontain species (eg Biomphalaria and Indoplanorbis in the Plan-orbidae family and Stagnicola in the Lymnaeidae family) that canhost both avian and mammalian schistosomes that cause derma-titis (40 51 70 104 106)
Previous papers have reviewed the details of host specificity inmammalian schistosomes (Fig 1 clades B and C) (107 108) Be-cause avian schistosomes are the major group of schistosomescausing dermatitis our own discussion focuses on their avian andsnail hosts For two reasons these avian schistosome species com-prise most of the dermatitis reports first many avian hosts sea-sonally migrate consequently disseminating avian schistosomesas they fly (domestic ducks can serve as definitive hosts particu-
TABLE 1 Summary of general host use of known genera of schistosomes reflecting current knowledge habitat in the definitive host and broadgeographic locality
Genus Snail host Mammalianavian host
Definitivehosthabitat Locality
Aquatichabitat
Major areas foroutbreaks
Austrobilharzia Nassariidae BatillaridaeLittoriniidaePotamididae
Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Ornithobilharzia Batillaridae Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Macrobilharzia Unknown Suliformes (Anhinga) Visceral NorthAmericaAfrica
Unknown Unknown if causesdermatitis
Bivitellobilharzia Unknown Elephantidae Rhinocerotidae Visceral Africa Asia Freshwater Probably freshwaterrivers
Schistosoma PlanorbidaeLymnaeidaePomatiopsidae
Mammalia VisceralNasal
Eurasia AfricaSouthAmerica
Freshwater Mostly eutrophic ponds
Heterobilharzia Lymnaeidae Mammalia Visceral North America Freshwater Marshy areasSchistosomatium Lymnaeidae Rodentia Visceral North America Freshwater Marshy areasBilharziella Planorbidae Anseriformes Gruiformes
CiconiformesPodicipediformes
Visceral Europe Freshwater Eutrophic ponds
Species isolatedfromHaminoea
Haminoeidae CharadriiformesPelicaniformes
Visceral North America Marine Shallow marine areastidal pools
Gigantobilharziaa Physidae Passeriformes Visceral North America Freshwater Marshy areas usuallywith cattails
Dendritobilharzia Planorbidae Anseriformes GruiformesPelicaniformesGaviiformes
Visceral Global Freshwater Unknown if reports ofdermatitis
Jilinobilharzia Unknown Anseriformes (Anatidae) Visceral China Unknown Unknown if reports ofdermatitis
Allobilharzia Unknown Anseriformes (swans) Visceral NorthernHemisphere
Unknown Unknown if causesdermatitis
Anserobilharzia Planorbidae Anseriformes (geese) Visceral NorthernHemisphere
Freshwater Eutrophic pondsreservoirs
Trichobilharzia Lymnaeidae Physidae Anseriformes (Anatidae) Visceralnasal
Global Freshwater Eutrophic ponds glaciallakes reservoirs
a Since Gigantobilharzia is not a monophyletic genus the information listed here is for G huronensis only
Horaacutek et al
168 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
larly for Trichobilharzia) and second some snail hosts are habitatgeneralists (eg Physa [syn Physella] acuta and Lymnaea stagna-lis) that are now globally distributed As a result the opportunitiesfor birds and snails to come into contact across time and space arevast For example P acuta (host to Gigantobilharzia huronensisTrichobilharzia physellae and Trichobilharzia querquedulae)thrives in both natural and altered environments with a widetolerance for water temperature and chemistry including the con-ditions found in ponds drainage ditches rivers marshes andephemeral water (109ndash111) In Europe the lymnaeid snail L stag-nalis (host of Trichobilharzia szidati) has also been linked to casesof dermatitis in people who acquired it while working withaquaria (112 113) providing evidence of the snailrsquos ability to per-sist in addition to its local global presence
From an evolutionary perspective the vagility and habitatspecificity of most bird hosts in concert with the availability ofsnails in aquatic habitats are likely mechanisms for widespreadhost switching in snails and thus for diversification of avian schis-tosomes (12 114) Our knowledge of the current schistosome-snail associations indicates that once a schistosome is hosted by aparticular species of snail it possesses little ability to utilize morethan a few species within that genus (eg Radix Stagnicola andPhysa) (115) Two exceptions are Dendritobilharzia pulverulentawhich uses Anisus vortex in Europe (116) and Gyraulus parvus inNorth America (117) both of which are small related planorbidsnails and Trichobilharzia regenti which employs lymnaeid snailsof the genus Radix in Europe and Austropeplea tomentosa in NewZealand (118) There is little evidence of schistosome speciescrossing snail families naturally or experimentally (for an excep-tion see references 119 and 120) Trichobilharzia franki was re-ported to be widespread across Europe but detailed molecularstudies are showing that it may represent several species related tosnail host use (70 121 122)
Cercarial dermatitis is caused not only by species of schisto-somes from indigenous snails but also by those from invasive orintroduced snails For example Haminoea japonica (originallyfrom Japan but now off the California coast) and Ilyanassa obso-leta (originally from the east coast but now on the west coast ofNorth America) are responsible for annual dermatitis outbreaks atmarine swimming beaches (25 64) In freshwater L stagnalis isfound commonly in the northern Eastern hemisphere yet in thenorthern Western hemisphere it is only locally common When Lstagnalis is found to be infected the infecting species is related to acommon European species Trichobilharzia szidati (70 123) In-terestingly P acuta is one of the most invasive pulmonate snailsand can host at least four species of avian schistosomes in NorthAmerica (10 70) yet there are no reports of this snail hostingschistosomes in their invasive range (outside North America) It isalso noteworthy that most of the schistosome species transmittedby physid snails have thus far been found only in North America(at least based on genetic comparisons) (eg G huronensis Tphysellae and one undescribed lineage of schistosome [10]) Thelymnaeid snail genera Stagnicola (found in North America) andRadix (found in the Eastern Hemisphere) are the main snail hostsfor most species of Trichobilharzia in fact thus far Radix main-tains most of the reported species diversity of Trichobilharzia(122) Trichobilharzia regenti has been recognized to cause derma-titis in Lake Wanaka in New Zealand and it may have been intro-duced from Europe in wild duck breeds (Anas platyrhynchos) usedfor hunting It is now found commonly in the endemic nonmigra-
tory scaup Aythya novaeseelandiae and the snail Austropeplea to-mentosa (118) Schistosomes seem to be specific to particular snailhosts at the species or genus level but not as much to their avian ormammalian hosts though loose specificity of definitive host useexists at higher taxonomic levels (Table 1) (eg Schistosomahaematobium in humans Bivitellobilharzia in elephants and Allo-bilharzia in swans)
The diversity of avian schistosomes found around the world isin no small part due to the ability of thousands of migratory birdsto carry their parasites across several latitudes and longitudes ex-posing commonly encountered snails This propensity to migratelarge distances distinguishes avian schistosomes from mammalianschistosomes in terms of distribution diversification and hostuse perhaps with the exception of Schistosoma mansoni (in termsof long-distance migration only) (124) Yet the schistosome spe-cies found in a wide range of avian host orders (eg Bilharziellaand Dendritobilharzia) (Table 1) are not the ones recurrently re-sponsible for outbreaks and are also species that are not geneticallydiverse compared to other species (69 125 126) Currently Dpulverulenta might be the most widespread single species of avianschistosome crossing both the Northern and Southern Hemi-spheres (117 125 127 128)
The most derived clade or most recently evolved clade in cladeD has three genera Allobilharzia Anserobilharzia and Trichobil-harzia (Fig 1) Allobilharzia from swans and Anserobilharziafrom geese both have a circumpolar distribution (69 70 105 129130) Anserobilharzia brantae which is common in North Amer-ica (in the Canada goose [Branta canadensis] and the snow goose[Chen caerulescens]) but also found in Europe (in the greylaggoose [Anser anser]) has been identified in at least one outbreak ofdermatitis in the United States (10) The area was a eutrophicmunicipal lakepond with a dense population of Gyraulus parvussnails and Canada geese The third genus Trichobilharzia themost species-rich genus in the family is found almost exclusivelyin ducks It should be noted that there are several species of Tricho-bilharzia reported from other avian families (Trichobilharzia corvifrom passeriforms along with other species [131 132]) but basedon the morphology of adults and eggs and snail host use (whenknown) these probably represent new genera or these avian hostsare not the primary (competent) hosts or might be aberrant cases(70 78) Within the genus Trichobilharzia several clades are spe-cific to certain groups of ducks for example Trichobilharzia stag-nicolae and Trichobilharzia mergi are found in mergansers (Mer-ginae) (70 133) T querquedulae in the ldquoblue-winged duckrdquo clade(Anas clypeata Anas discors and Anas cyanoptera) (70 134) Tphysellae in an ecological group of diving ducks that includesducks of the Aythinae and Merginae and a common undescribedspecies of Trichobilharzia species A in Anas americana (70) It isnot yet clear which duck groups (phylogenetic or ecological) aremore specific for T franki or T szidati Interestingly T regenti hasprobably been reported from the most diverse duck species (135)and does not appear to have a preferred host within the AnatidaeIn North America T stagnicolae and T physellae are most oftenidentified in dermatitis outbreaks (70)
INTRAMOLLUSCAN DEVELOPMENT OF AVIANSCHISTOSOMES
As noted above Trichobilharzia is the most diverse schistosomegenus and has most often been implicated in outbreaks of cercarialdermatitis As a result studies on the avian schistosome-snail in-
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 169Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
termediate host relationship have focused primarily on species ofthe genus Trichobilharzia Additionally long-term laboratorymaintenance of T szidati and L stagnalis enabled the experimentsthat uncovered the intimate molecular interactions of avian schis-tosomes and their snail hosts (The Trichobilharzia ocellata organ-ism used as an experimental model in European laboratories forthe last few decades is identical to T szidati and the latter name ispreferred and is used here [123] If T ocellata is used in the textbody then it refers to the non-European isolates of the parasite)
After hatching from eggs in an aquatic environment schisto-some miracidia search for and invade an appropriate snail hostspecies This behavior must be accomplished quickly as miracidiahave a temperature-dependent limited life span of around 20 h asreported for eg T stagnicolae (136) Studies on the miracidialbehavior of T szidati have shown a progression of steps from hostfinding to penetration and migration Miracidia respond to envi-ronmental stimuli such as light or gravity that direct them to themicrohabitat occupied by the host snails (137) Snails release var-ious chemical compounds that form an ldquoactive spacerdquo aroundthem and serve as chemoattractants for the miracidia Miracidiarecognize macromolecular glycoconjugates termed miracidium-attracting glycoproteins (MAGs) or miraxones that consist of aprotein core and carbohydrate chains linked O-glycosidically viaN-acetyl-D-galactosamine and serinethreonine (138 139) Theattractant for miracidia is encoded in these carbohydrate moietiesUpon entering the ldquoactive spacerdquo of the snail miracidia modifytheir movement by increased random turns within the increasingattractant gradient and by a turn-back form of swimming withinthe decreasing attractant gradient (140) This mode of orientation(chemokinesis) observed eg in T szidati or T franki (115 138)results in the first contact of the miracidia with the snail which isthen followed by repeated investigationprobing of the snail sur-face and finally miracidial attachment (140) Penetration of thesnail surface follows although the factors contributing to this pro-cess such as the components of miracidial penetration glandsremain largely unknown
After penetrating the surface epithelium of the snail miracidiaof avian schistosomes transform into mother sporocysts that giverise to daughter sporocysts which migrate to the snail hepatopan-creas where production of the final larval stage the cercariaetakes place (13) The prepatent period lasts about 3 to 10 weeksdepending on several factors such as the miracidial dose or tem-perature (13 141) Infection by avian schistosomes may lead toalterations of the snail internal defense system (IDS) metabolismand endocrine functions Such alterations have been studiedwidely in L stagnalis the intermediate host of T szidati (142ndash145)
The snail IDS is based solely on innate immune mechanismscomposed of humoral and cellular limbs Lectins are essential hu-moral components whereas hemocytes represent the main effec-tor cells (146 147) Both limbs of the L stagnalis IDS appear to beactivated and then suppressed during early and late stages respec-tively of a T szidati infection (143) In vitro hemocytes failed toencapsulate and destroy T szidati sporocysts (148) It has beensuggested that parasite excretory-secretory products participate inthe modulation of hemocyte activities (149) Disruption of hemo-cyte signaling pathways such as protein kinase C (PKC) and ex-tracellular signal-regulated kinase (ERK) pathways may alsoinfluence hemocyte activities (150ndash152) For example carbohy-drates known to be present on larval surfaces of T szidati and Tregenti such as D-galactose and L-fucose (153ndash156) affected he-
mocyte PKC and ERK signaling in L stagnalis which suggests animmunosuppressive role (157 158) However experiments inves-tigating the direct effect of Trichobilharzia larvae on hemocytesignaling have not been performed Alterations of humoral de-fense components also occur during infections by avian schisto-somes and at least two molecules molluscan defense molecule(MDM) and granularin have been investigated in this respect(159 160) Both molecules produced in L stagnalis by granularcells of connective tissue are related to phagocytic activity of he-mocytes MDM enhances phagocytosis of hemocytes and the ex-pression of a corresponding gene for MDM is downregulated in Lstagnalis infected with T szidati (159 161) In contrast treatmentof hemocytes with granularin decreases phagocytic activity andthe encoding gene is upregulated in parasitized snails (160 161)
In the snail host avian schistosomes also interfere with metab-olism such as causing abnormal body growth and endocrinefunctions such as causing a reduction of egg laying (161) In Lstagnalis these processes are regulated by neuroendocrine cells inthe central nervous system (CNS) (162 163) Trichobilharzia szi-dati releases an undescribed substance that induces the snail hostto produce schistosomin (a peptide of 87 kDa consisting of 79amino acids) from its connective tissue and hemocytes (145)Schistosomin acts as a neuropeptide that interferes with somehormones such as calfluxin a neuropeptide that stimulates theinflux of Ca2 into the mitochondria of albumen gland cells (164165) As a consequence ovulation and egg laying are inhibited inthe snail Excitability of neuroendocrine cells (light green cells[LGCs]) responsible for growth (162) increases in response toschistosomin (166) As a result the body size of infected snailsmay become considerably larger than that of uninfected snails(142 161) Other neuropeptides (FMRFamide-related peptides)are also upregulated during infection of L stagnalis by T szidatiand these peptides via inhibition of neuroendocrine cells may beresponsible for the suppression of snail metabolism and repro-duction (167) All these changes in infected snails may provideenergy resources and space that can be exploited by the schisto-somes for development (145 161)
The survival rate of infected snails releasing schistosome cer-cariae as the agent of cercarial dermatitis varies among species Alimited number of studies have focused on survival rates of avianschistosome-infected versus uninfected snails As an example90 of L stagnalis snails infected experimentally with a singlemiracidium of T ocellata (North American isolate) were alive at28 weeks of age (three infected snails were alive for 19 months)whereas all uninfected snails were dead (168) In contrast L stag-nalis or Planorbarius corneus snails naturally infected with T szi-dati or Bilharziella polonica respectively lived a shorter time onaverage than the corresponding uninfected individuals (169170)
VERTEBRATE HOST FINDING AND PENETRATION
Cercariae emerging from the snail intermediate host are the infec-tive stage to the definitive host and are also the stage responsiblefor causing cercarial dermatitis A cercaria is a multicellular larvacomprised of an oblong body and a slender tail that is bifurcated(furcocercous) at the posterior end (Fig 2) Cercariae of schisto-somes leave their snail hosts actively For this purpose they em-ploy a pair of specialized unicellular escape glands that are obviousin mature cercariae within sporocysts Once the cercariae haveemerged only their ducts lined by microtubules are visible sug-
Horaacutek et al
170 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-
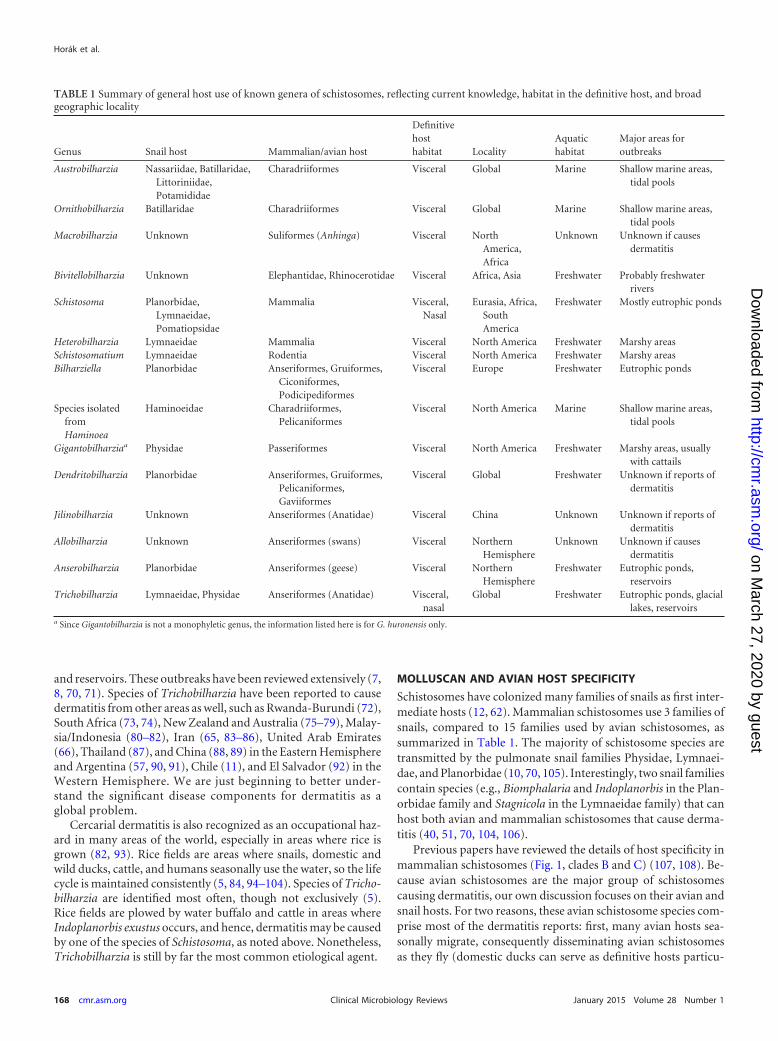
and reservoirs These outbreaks have been reviewed extensively (78 70 71) Species of Trichobilharzia have been reported to causedermatitis from other areas as well such as Rwanda-Burundi (72)South Africa (73 74) New Zealand and Australia (75ndash79) Malay-siaIndonesia (80ndash82) Iran (65 83ndash86) United Arab Emirates(66) Thailand (87) and China (88 89) in the Eastern Hemisphereand Argentina (57 90 91) Chile (11) and El Salvador (92) in theWestern Hemisphere We are just beginning to better under-stand the significant disease components for dermatitis as aglobal problem
Cercarial dermatitis is also recognized as an occupational haz-ard in many areas of the world especially in areas where rice isgrown (82 93) Rice fields are areas where snails domestic andwild ducks cattle and humans seasonally use the water so the lifecycle is maintained consistently (5 84 94ndash104) Species of Tricho-bilharzia are identified most often though not exclusively (5)Rice fields are plowed by water buffalo and cattle in areas whereIndoplanorbis exustus occurs and hence dermatitis may be causedby one of the species of Schistosoma as noted above NonethelessTrichobilharzia is still by far the most common etiological agent
MOLLUSCAN AND AVIAN HOST SPECIFICITY
Schistosomes have colonized many families of snails as first inter-mediate hosts (12 62) Mammalian schistosomes use 3 families ofsnails compared to 15 families used by avian schistosomes assummarized in Table 1 The majority of schistosome species aretransmitted by the pulmonate snail families Physidae Lymnaei-dae and Planorbidae (10 70 105) Interestingly two snail familiescontain species (eg Biomphalaria and Indoplanorbis in the Plan-orbidae family and Stagnicola in the Lymnaeidae family) that canhost both avian and mammalian schistosomes that cause derma-titis (40 51 70 104 106)
Previous papers have reviewed the details of host specificity inmammalian schistosomes (Fig 1 clades B and C) (107 108) Be-cause avian schistosomes are the major group of schistosomescausing dermatitis our own discussion focuses on their avian andsnail hosts For two reasons these avian schistosome species com-prise most of the dermatitis reports first many avian hosts sea-sonally migrate consequently disseminating avian schistosomesas they fly (domestic ducks can serve as definitive hosts particu-
TABLE 1 Summary of general host use of known genera of schistosomes reflecting current knowledge habitat in the definitive host and broadgeographic locality
Genus Snail host Mammalianavian host
Definitivehosthabitat Locality
Aquatichabitat
Major areas foroutbreaks
Austrobilharzia Nassariidae BatillaridaeLittoriniidaePotamididae
Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Ornithobilharzia Batillaridae Charadriiformes Visceral Global Marine Shallow marine areastidal pools
Macrobilharzia Unknown Suliformes (Anhinga) Visceral NorthAmericaAfrica
Unknown Unknown if causesdermatitis
Bivitellobilharzia Unknown Elephantidae Rhinocerotidae Visceral Africa Asia Freshwater Probably freshwaterrivers
Schistosoma PlanorbidaeLymnaeidaePomatiopsidae
Mammalia VisceralNasal
Eurasia AfricaSouthAmerica
Freshwater Mostly eutrophic ponds
Heterobilharzia Lymnaeidae Mammalia Visceral North America Freshwater Marshy areasSchistosomatium Lymnaeidae Rodentia Visceral North America Freshwater Marshy areasBilharziella Planorbidae Anseriformes Gruiformes
CiconiformesPodicipediformes
Visceral Europe Freshwater Eutrophic ponds
Species isolatedfromHaminoea
Haminoeidae CharadriiformesPelicaniformes
Visceral North America Marine Shallow marine areastidal pools
Gigantobilharziaa Physidae Passeriformes Visceral North America Freshwater Marshy areas usuallywith cattails
Dendritobilharzia Planorbidae Anseriformes GruiformesPelicaniformesGaviiformes
Visceral Global Freshwater Unknown if reports ofdermatitis
Jilinobilharzia Unknown Anseriformes (Anatidae) Visceral China Unknown Unknown if reports ofdermatitis
Allobilharzia Unknown Anseriformes (swans) Visceral NorthernHemisphere
Unknown Unknown if causesdermatitis
Anserobilharzia Planorbidae Anseriformes (geese) Visceral NorthernHemisphere
Freshwater Eutrophic pondsreservoirs
Trichobilharzia Lymnaeidae Physidae Anseriformes (Anatidae) Visceralnasal
Global Freshwater Eutrophic ponds glaciallakes reservoirs
a Since Gigantobilharzia is not a monophyletic genus the information listed here is for G huronensis only
Horaacutek et al
168 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
larly for Trichobilharzia) and second some snail hosts are habitatgeneralists (eg Physa [syn Physella] acuta and Lymnaea stagna-lis) that are now globally distributed As a result the opportunitiesfor birds and snails to come into contact across time and space arevast For example P acuta (host to Gigantobilharzia huronensisTrichobilharzia physellae and Trichobilharzia querquedulae)thrives in both natural and altered environments with a widetolerance for water temperature and chemistry including the con-ditions found in ponds drainage ditches rivers marshes andephemeral water (109ndash111) In Europe the lymnaeid snail L stag-nalis (host of Trichobilharzia szidati) has also been linked to casesof dermatitis in people who acquired it while working withaquaria (112 113) providing evidence of the snailrsquos ability to per-sist in addition to its local global presence
From an evolutionary perspective the vagility and habitatspecificity of most bird hosts in concert with the availability ofsnails in aquatic habitats are likely mechanisms for widespreadhost switching in snails and thus for diversification of avian schis-tosomes (12 114) Our knowledge of the current schistosome-snail associations indicates that once a schistosome is hosted by aparticular species of snail it possesses little ability to utilize morethan a few species within that genus (eg Radix Stagnicola andPhysa) (115) Two exceptions are Dendritobilharzia pulverulentawhich uses Anisus vortex in Europe (116) and Gyraulus parvus inNorth America (117) both of which are small related planorbidsnails and Trichobilharzia regenti which employs lymnaeid snailsof the genus Radix in Europe and Austropeplea tomentosa in NewZealand (118) There is little evidence of schistosome speciescrossing snail families naturally or experimentally (for an excep-tion see references 119 and 120) Trichobilharzia franki was re-ported to be widespread across Europe but detailed molecularstudies are showing that it may represent several species related tosnail host use (70 121 122)
Cercarial dermatitis is caused not only by species of schisto-somes from indigenous snails but also by those from invasive orintroduced snails For example Haminoea japonica (originallyfrom Japan but now off the California coast) and Ilyanassa obso-leta (originally from the east coast but now on the west coast ofNorth America) are responsible for annual dermatitis outbreaks atmarine swimming beaches (25 64) In freshwater L stagnalis isfound commonly in the northern Eastern hemisphere yet in thenorthern Western hemisphere it is only locally common When Lstagnalis is found to be infected the infecting species is related to acommon European species Trichobilharzia szidati (70 123) In-terestingly P acuta is one of the most invasive pulmonate snailsand can host at least four species of avian schistosomes in NorthAmerica (10 70) yet there are no reports of this snail hostingschistosomes in their invasive range (outside North America) It isalso noteworthy that most of the schistosome species transmittedby physid snails have thus far been found only in North America(at least based on genetic comparisons) (eg G huronensis Tphysellae and one undescribed lineage of schistosome [10]) Thelymnaeid snail genera Stagnicola (found in North America) andRadix (found in the Eastern Hemisphere) are the main snail hostsfor most species of Trichobilharzia in fact thus far Radix main-tains most of the reported species diversity of Trichobilharzia(122) Trichobilharzia regenti has been recognized to cause derma-titis in Lake Wanaka in New Zealand and it may have been intro-duced from Europe in wild duck breeds (Anas platyrhynchos) usedfor hunting It is now found commonly in the endemic nonmigra-
tory scaup Aythya novaeseelandiae and the snail Austropeplea to-mentosa (118) Schistosomes seem to be specific to particular snailhosts at the species or genus level but not as much to their avian ormammalian hosts though loose specificity of definitive host useexists at higher taxonomic levels (Table 1) (eg Schistosomahaematobium in humans Bivitellobilharzia in elephants and Allo-bilharzia in swans)
The diversity of avian schistosomes found around the world isin no small part due to the ability of thousands of migratory birdsto carry their parasites across several latitudes and longitudes ex-posing commonly encountered snails This propensity to migratelarge distances distinguishes avian schistosomes from mammalianschistosomes in terms of distribution diversification and hostuse perhaps with the exception of Schistosoma mansoni (in termsof long-distance migration only) (124) Yet the schistosome spe-cies found in a wide range of avian host orders (eg Bilharziellaand Dendritobilharzia) (Table 1) are not the ones recurrently re-sponsible for outbreaks and are also species that are not geneticallydiverse compared to other species (69 125 126) Currently Dpulverulenta might be the most widespread single species of avianschistosome crossing both the Northern and Southern Hemi-spheres (117 125 127 128)
The most derived clade or most recently evolved clade in cladeD has three genera Allobilharzia Anserobilharzia and Trichobil-harzia (Fig 1) Allobilharzia from swans and Anserobilharziafrom geese both have a circumpolar distribution (69 70 105 129130) Anserobilharzia brantae which is common in North Amer-ica (in the Canada goose [Branta canadensis] and the snow goose[Chen caerulescens]) but also found in Europe (in the greylaggoose [Anser anser]) has been identified in at least one outbreak ofdermatitis in the United States (10) The area was a eutrophicmunicipal lakepond with a dense population of Gyraulus parvussnails and Canada geese The third genus Trichobilharzia themost species-rich genus in the family is found almost exclusivelyin ducks It should be noted that there are several species of Tricho-bilharzia reported from other avian families (Trichobilharzia corvifrom passeriforms along with other species [131 132]) but basedon the morphology of adults and eggs and snail host use (whenknown) these probably represent new genera or these avian hostsare not the primary (competent) hosts or might be aberrant cases(70 78) Within the genus Trichobilharzia several clades are spe-cific to certain groups of ducks for example Trichobilharzia stag-nicolae and Trichobilharzia mergi are found in mergansers (Mer-ginae) (70 133) T querquedulae in the ldquoblue-winged duckrdquo clade(Anas clypeata Anas discors and Anas cyanoptera) (70 134) Tphysellae in an ecological group of diving ducks that includesducks of the Aythinae and Merginae and a common undescribedspecies of Trichobilharzia species A in Anas americana (70) It isnot yet clear which duck groups (phylogenetic or ecological) aremore specific for T franki or T szidati Interestingly T regenti hasprobably been reported from the most diverse duck species (135)and does not appear to have a preferred host within the AnatidaeIn North America T stagnicolae and T physellae are most oftenidentified in dermatitis outbreaks (70)
INTRAMOLLUSCAN DEVELOPMENT OF AVIANSCHISTOSOMES
As noted above Trichobilharzia is the most diverse schistosomegenus and has most often been implicated in outbreaks of cercarialdermatitis As a result studies on the avian schistosome-snail in-
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 169Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
termediate host relationship have focused primarily on species ofthe genus Trichobilharzia Additionally long-term laboratorymaintenance of T szidati and L stagnalis enabled the experimentsthat uncovered the intimate molecular interactions of avian schis-tosomes and their snail hosts (The Trichobilharzia ocellata organ-ism used as an experimental model in European laboratories forthe last few decades is identical to T szidati and the latter name ispreferred and is used here [123] If T ocellata is used in the textbody then it refers to the non-European isolates of the parasite)
After hatching from eggs in an aquatic environment schisto-some miracidia search for and invade an appropriate snail hostspecies This behavior must be accomplished quickly as miracidiahave a temperature-dependent limited life span of around 20 h asreported for eg T stagnicolae (136) Studies on the miracidialbehavior of T szidati have shown a progression of steps from hostfinding to penetration and migration Miracidia respond to envi-ronmental stimuli such as light or gravity that direct them to themicrohabitat occupied by the host snails (137) Snails release var-ious chemical compounds that form an ldquoactive spacerdquo aroundthem and serve as chemoattractants for the miracidia Miracidiarecognize macromolecular glycoconjugates termed miracidium-attracting glycoproteins (MAGs) or miraxones that consist of aprotein core and carbohydrate chains linked O-glycosidically viaN-acetyl-D-galactosamine and serinethreonine (138 139) Theattractant for miracidia is encoded in these carbohydrate moietiesUpon entering the ldquoactive spacerdquo of the snail miracidia modifytheir movement by increased random turns within the increasingattractant gradient and by a turn-back form of swimming withinthe decreasing attractant gradient (140) This mode of orientation(chemokinesis) observed eg in T szidati or T franki (115 138)results in the first contact of the miracidia with the snail which isthen followed by repeated investigationprobing of the snail sur-face and finally miracidial attachment (140) Penetration of thesnail surface follows although the factors contributing to this pro-cess such as the components of miracidial penetration glandsremain largely unknown
After penetrating the surface epithelium of the snail miracidiaof avian schistosomes transform into mother sporocysts that giverise to daughter sporocysts which migrate to the snail hepatopan-creas where production of the final larval stage the cercariaetakes place (13) The prepatent period lasts about 3 to 10 weeksdepending on several factors such as the miracidial dose or tem-perature (13 141) Infection by avian schistosomes may lead toalterations of the snail internal defense system (IDS) metabolismand endocrine functions Such alterations have been studiedwidely in L stagnalis the intermediate host of T szidati (142ndash145)
The snail IDS is based solely on innate immune mechanismscomposed of humoral and cellular limbs Lectins are essential hu-moral components whereas hemocytes represent the main effec-tor cells (146 147) Both limbs of the L stagnalis IDS appear to beactivated and then suppressed during early and late stages respec-tively of a T szidati infection (143) In vitro hemocytes failed toencapsulate and destroy T szidati sporocysts (148) It has beensuggested that parasite excretory-secretory products participate inthe modulation of hemocyte activities (149) Disruption of hemo-cyte signaling pathways such as protein kinase C (PKC) and ex-tracellular signal-regulated kinase (ERK) pathways may alsoinfluence hemocyte activities (150ndash152) For example carbohy-drates known to be present on larval surfaces of T szidati and Tregenti such as D-galactose and L-fucose (153ndash156) affected he-
mocyte PKC and ERK signaling in L stagnalis which suggests animmunosuppressive role (157 158) However experiments inves-tigating the direct effect of Trichobilharzia larvae on hemocytesignaling have not been performed Alterations of humoral de-fense components also occur during infections by avian schisto-somes and at least two molecules molluscan defense molecule(MDM) and granularin have been investigated in this respect(159 160) Both molecules produced in L stagnalis by granularcells of connective tissue are related to phagocytic activity of he-mocytes MDM enhances phagocytosis of hemocytes and the ex-pression of a corresponding gene for MDM is downregulated in Lstagnalis infected with T szidati (159 161) In contrast treatmentof hemocytes with granularin decreases phagocytic activity andthe encoding gene is upregulated in parasitized snails (160 161)
In the snail host avian schistosomes also interfere with metab-olism such as causing abnormal body growth and endocrinefunctions such as causing a reduction of egg laying (161) In Lstagnalis these processes are regulated by neuroendocrine cells inthe central nervous system (CNS) (162 163) Trichobilharzia szi-dati releases an undescribed substance that induces the snail hostto produce schistosomin (a peptide of 87 kDa consisting of 79amino acids) from its connective tissue and hemocytes (145)Schistosomin acts as a neuropeptide that interferes with somehormones such as calfluxin a neuropeptide that stimulates theinflux of Ca2 into the mitochondria of albumen gland cells (164165) As a consequence ovulation and egg laying are inhibited inthe snail Excitability of neuroendocrine cells (light green cells[LGCs]) responsible for growth (162) increases in response toschistosomin (166) As a result the body size of infected snailsmay become considerably larger than that of uninfected snails(142 161) Other neuropeptides (FMRFamide-related peptides)are also upregulated during infection of L stagnalis by T szidatiand these peptides via inhibition of neuroendocrine cells may beresponsible for the suppression of snail metabolism and repro-duction (167) All these changes in infected snails may provideenergy resources and space that can be exploited by the schisto-somes for development (145 161)
The survival rate of infected snails releasing schistosome cer-cariae as the agent of cercarial dermatitis varies among species Alimited number of studies have focused on survival rates of avianschistosome-infected versus uninfected snails As an example90 of L stagnalis snails infected experimentally with a singlemiracidium of T ocellata (North American isolate) were alive at28 weeks of age (three infected snails were alive for 19 months)whereas all uninfected snails were dead (168) In contrast L stag-nalis or Planorbarius corneus snails naturally infected with T szi-dati or Bilharziella polonica respectively lived a shorter time onaverage than the corresponding uninfected individuals (169170)
VERTEBRATE HOST FINDING AND PENETRATION
Cercariae emerging from the snail intermediate host are the infec-tive stage to the definitive host and are also the stage responsiblefor causing cercarial dermatitis A cercaria is a multicellular larvacomprised of an oblong body and a slender tail that is bifurcated(furcocercous) at the posterior end (Fig 2) Cercariae of schisto-somes leave their snail hosts actively For this purpose they em-ploy a pair of specialized unicellular escape glands that are obviousin mature cercariae within sporocysts Once the cercariae haveemerged only their ducts lined by microtubules are visible sug-
Horaacutek et al
170 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

larly for Trichobilharzia) and second some snail hosts are habitatgeneralists (eg Physa [syn Physella] acuta and Lymnaea stagna-lis) that are now globally distributed As a result the opportunitiesfor birds and snails to come into contact across time and space arevast For example P acuta (host to Gigantobilharzia huronensisTrichobilharzia physellae and Trichobilharzia querquedulae)thrives in both natural and altered environments with a widetolerance for water temperature and chemistry including the con-ditions found in ponds drainage ditches rivers marshes andephemeral water (109ndash111) In Europe the lymnaeid snail L stag-nalis (host of Trichobilharzia szidati) has also been linked to casesof dermatitis in people who acquired it while working withaquaria (112 113) providing evidence of the snailrsquos ability to per-sist in addition to its local global presence
From an evolutionary perspective the vagility and habitatspecificity of most bird hosts in concert with the availability ofsnails in aquatic habitats are likely mechanisms for widespreadhost switching in snails and thus for diversification of avian schis-tosomes (12 114) Our knowledge of the current schistosome-snail associations indicates that once a schistosome is hosted by aparticular species of snail it possesses little ability to utilize morethan a few species within that genus (eg Radix Stagnicola andPhysa) (115) Two exceptions are Dendritobilharzia pulverulentawhich uses Anisus vortex in Europe (116) and Gyraulus parvus inNorth America (117) both of which are small related planorbidsnails and Trichobilharzia regenti which employs lymnaeid snailsof the genus Radix in Europe and Austropeplea tomentosa in NewZealand (118) There is little evidence of schistosome speciescrossing snail families naturally or experimentally (for an excep-tion see references 119 and 120) Trichobilharzia franki was re-ported to be widespread across Europe but detailed molecularstudies are showing that it may represent several species related tosnail host use (70 121 122)
Cercarial dermatitis is caused not only by species of schisto-somes from indigenous snails but also by those from invasive orintroduced snails For example Haminoea japonica (originallyfrom Japan but now off the California coast) and Ilyanassa obso-leta (originally from the east coast but now on the west coast ofNorth America) are responsible for annual dermatitis outbreaks atmarine swimming beaches (25 64) In freshwater L stagnalis isfound commonly in the northern Eastern hemisphere yet in thenorthern Western hemisphere it is only locally common When Lstagnalis is found to be infected the infecting species is related to acommon European species Trichobilharzia szidati (70 123) In-terestingly P acuta is one of the most invasive pulmonate snailsand can host at least four species of avian schistosomes in NorthAmerica (10 70) yet there are no reports of this snail hostingschistosomes in their invasive range (outside North America) It isalso noteworthy that most of the schistosome species transmittedby physid snails have thus far been found only in North America(at least based on genetic comparisons) (eg G huronensis Tphysellae and one undescribed lineage of schistosome [10]) Thelymnaeid snail genera Stagnicola (found in North America) andRadix (found in the Eastern Hemisphere) are the main snail hostsfor most species of Trichobilharzia in fact thus far Radix main-tains most of the reported species diversity of Trichobilharzia(122) Trichobilharzia regenti has been recognized to cause derma-titis in Lake Wanaka in New Zealand and it may have been intro-duced from Europe in wild duck breeds (Anas platyrhynchos) usedfor hunting It is now found commonly in the endemic nonmigra-
tory scaup Aythya novaeseelandiae and the snail Austropeplea to-mentosa (118) Schistosomes seem to be specific to particular snailhosts at the species or genus level but not as much to their avian ormammalian hosts though loose specificity of definitive host useexists at higher taxonomic levels (Table 1) (eg Schistosomahaematobium in humans Bivitellobilharzia in elephants and Allo-bilharzia in swans)
The diversity of avian schistosomes found around the world isin no small part due to the ability of thousands of migratory birdsto carry their parasites across several latitudes and longitudes ex-posing commonly encountered snails This propensity to migratelarge distances distinguishes avian schistosomes from mammalianschistosomes in terms of distribution diversification and hostuse perhaps with the exception of Schistosoma mansoni (in termsof long-distance migration only) (124) Yet the schistosome spe-cies found in a wide range of avian host orders (eg Bilharziellaand Dendritobilharzia) (Table 1) are not the ones recurrently re-sponsible for outbreaks and are also species that are not geneticallydiverse compared to other species (69 125 126) Currently Dpulverulenta might be the most widespread single species of avianschistosome crossing both the Northern and Southern Hemi-spheres (117 125 127 128)
The most derived clade or most recently evolved clade in cladeD has three genera Allobilharzia Anserobilharzia and Trichobil-harzia (Fig 1) Allobilharzia from swans and Anserobilharziafrom geese both have a circumpolar distribution (69 70 105 129130) Anserobilharzia brantae which is common in North Amer-ica (in the Canada goose [Branta canadensis] and the snow goose[Chen caerulescens]) but also found in Europe (in the greylaggoose [Anser anser]) has been identified in at least one outbreak ofdermatitis in the United States (10) The area was a eutrophicmunicipal lakepond with a dense population of Gyraulus parvussnails and Canada geese The third genus Trichobilharzia themost species-rich genus in the family is found almost exclusivelyin ducks It should be noted that there are several species of Tricho-bilharzia reported from other avian families (Trichobilharzia corvifrom passeriforms along with other species [131 132]) but basedon the morphology of adults and eggs and snail host use (whenknown) these probably represent new genera or these avian hostsare not the primary (competent) hosts or might be aberrant cases(70 78) Within the genus Trichobilharzia several clades are spe-cific to certain groups of ducks for example Trichobilharzia stag-nicolae and Trichobilharzia mergi are found in mergansers (Mer-ginae) (70 133) T querquedulae in the ldquoblue-winged duckrdquo clade(Anas clypeata Anas discors and Anas cyanoptera) (70 134) Tphysellae in an ecological group of diving ducks that includesducks of the Aythinae and Merginae and a common undescribedspecies of Trichobilharzia species A in Anas americana (70) It isnot yet clear which duck groups (phylogenetic or ecological) aremore specific for T franki or T szidati Interestingly T regenti hasprobably been reported from the most diverse duck species (135)and does not appear to have a preferred host within the AnatidaeIn North America T stagnicolae and T physellae are most oftenidentified in dermatitis outbreaks (70)
INTRAMOLLUSCAN DEVELOPMENT OF AVIANSCHISTOSOMES
As noted above Trichobilharzia is the most diverse schistosomegenus and has most often been implicated in outbreaks of cercarialdermatitis As a result studies on the avian schistosome-snail in-
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 169Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
termediate host relationship have focused primarily on species ofthe genus Trichobilharzia Additionally long-term laboratorymaintenance of T szidati and L stagnalis enabled the experimentsthat uncovered the intimate molecular interactions of avian schis-tosomes and their snail hosts (The Trichobilharzia ocellata organ-ism used as an experimental model in European laboratories forthe last few decades is identical to T szidati and the latter name ispreferred and is used here [123] If T ocellata is used in the textbody then it refers to the non-European isolates of the parasite)
After hatching from eggs in an aquatic environment schisto-some miracidia search for and invade an appropriate snail hostspecies This behavior must be accomplished quickly as miracidiahave a temperature-dependent limited life span of around 20 h asreported for eg T stagnicolae (136) Studies on the miracidialbehavior of T szidati have shown a progression of steps from hostfinding to penetration and migration Miracidia respond to envi-ronmental stimuli such as light or gravity that direct them to themicrohabitat occupied by the host snails (137) Snails release var-ious chemical compounds that form an ldquoactive spacerdquo aroundthem and serve as chemoattractants for the miracidia Miracidiarecognize macromolecular glycoconjugates termed miracidium-attracting glycoproteins (MAGs) or miraxones that consist of aprotein core and carbohydrate chains linked O-glycosidically viaN-acetyl-D-galactosamine and serinethreonine (138 139) Theattractant for miracidia is encoded in these carbohydrate moietiesUpon entering the ldquoactive spacerdquo of the snail miracidia modifytheir movement by increased random turns within the increasingattractant gradient and by a turn-back form of swimming withinthe decreasing attractant gradient (140) This mode of orientation(chemokinesis) observed eg in T szidati or T franki (115 138)results in the first contact of the miracidia with the snail which isthen followed by repeated investigationprobing of the snail sur-face and finally miracidial attachment (140) Penetration of thesnail surface follows although the factors contributing to this pro-cess such as the components of miracidial penetration glandsremain largely unknown
After penetrating the surface epithelium of the snail miracidiaof avian schistosomes transform into mother sporocysts that giverise to daughter sporocysts which migrate to the snail hepatopan-creas where production of the final larval stage the cercariaetakes place (13) The prepatent period lasts about 3 to 10 weeksdepending on several factors such as the miracidial dose or tem-perature (13 141) Infection by avian schistosomes may lead toalterations of the snail internal defense system (IDS) metabolismand endocrine functions Such alterations have been studiedwidely in L stagnalis the intermediate host of T szidati (142ndash145)
The snail IDS is based solely on innate immune mechanismscomposed of humoral and cellular limbs Lectins are essential hu-moral components whereas hemocytes represent the main effec-tor cells (146 147) Both limbs of the L stagnalis IDS appear to beactivated and then suppressed during early and late stages respec-tively of a T szidati infection (143) In vitro hemocytes failed toencapsulate and destroy T szidati sporocysts (148) It has beensuggested that parasite excretory-secretory products participate inthe modulation of hemocyte activities (149) Disruption of hemo-cyte signaling pathways such as protein kinase C (PKC) and ex-tracellular signal-regulated kinase (ERK) pathways may alsoinfluence hemocyte activities (150ndash152) For example carbohy-drates known to be present on larval surfaces of T szidati and Tregenti such as D-galactose and L-fucose (153ndash156) affected he-
mocyte PKC and ERK signaling in L stagnalis which suggests animmunosuppressive role (157 158) However experiments inves-tigating the direct effect of Trichobilharzia larvae on hemocytesignaling have not been performed Alterations of humoral de-fense components also occur during infections by avian schisto-somes and at least two molecules molluscan defense molecule(MDM) and granularin have been investigated in this respect(159 160) Both molecules produced in L stagnalis by granularcells of connective tissue are related to phagocytic activity of he-mocytes MDM enhances phagocytosis of hemocytes and the ex-pression of a corresponding gene for MDM is downregulated in Lstagnalis infected with T szidati (159 161) In contrast treatmentof hemocytes with granularin decreases phagocytic activity andthe encoding gene is upregulated in parasitized snails (160 161)
In the snail host avian schistosomes also interfere with metab-olism such as causing abnormal body growth and endocrinefunctions such as causing a reduction of egg laying (161) In Lstagnalis these processes are regulated by neuroendocrine cells inthe central nervous system (CNS) (162 163) Trichobilharzia szi-dati releases an undescribed substance that induces the snail hostto produce schistosomin (a peptide of 87 kDa consisting of 79amino acids) from its connective tissue and hemocytes (145)Schistosomin acts as a neuropeptide that interferes with somehormones such as calfluxin a neuropeptide that stimulates theinflux of Ca2 into the mitochondria of albumen gland cells (164165) As a consequence ovulation and egg laying are inhibited inthe snail Excitability of neuroendocrine cells (light green cells[LGCs]) responsible for growth (162) increases in response toschistosomin (166) As a result the body size of infected snailsmay become considerably larger than that of uninfected snails(142 161) Other neuropeptides (FMRFamide-related peptides)are also upregulated during infection of L stagnalis by T szidatiand these peptides via inhibition of neuroendocrine cells may beresponsible for the suppression of snail metabolism and repro-duction (167) All these changes in infected snails may provideenergy resources and space that can be exploited by the schisto-somes for development (145 161)
The survival rate of infected snails releasing schistosome cer-cariae as the agent of cercarial dermatitis varies among species Alimited number of studies have focused on survival rates of avianschistosome-infected versus uninfected snails As an example90 of L stagnalis snails infected experimentally with a singlemiracidium of T ocellata (North American isolate) were alive at28 weeks of age (three infected snails were alive for 19 months)whereas all uninfected snails were dead (168) In contrast L stag-nalis or Planorbarius corneus snails naturally infected with T szi-dati or Bilharziella polonica respectively lived a shorter time onaverage than the corresponding uninfected individuals (169170)
VERTEBRATE HOST FINDING AND PENETRATION
Cercariae emerging from the snail intermediate host are the infec-tive stage to the definitive host and are also the stage responsiblefor causing cercarial dermatitis A cercaria is a multicellular larvacomprised of an oblong body and a slender tail that is bifurcated(furcocercous) at the posterior end (Fig 2) Cercariae of schisto-somes leave their snail hosts actively For this purpose they em-ploy a pair of specialized unicellular escape glands that are obviousin mature cercariae within sporocysts Once the cercariae haveemerged only their ducts lined by microtubules are visible sug-
Horaacutek et al
170 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

termediate host relationship have focused primarily on species ofthe genus Trichobilharzia Additionally long-term laboratorymaintenance of T szidati and L stagnalis enabled the experimentsthat uncovered the intimate molecular interactions of avian schis-tosomes and their snail hosts (The Trichobilharzia ocellata organ-ism used as an experimental model in European laboratories forthe last few decades is identical to T szidati and the latter name ispreferred and is used here [123] If T ocellata is used in the textbody then it refers to the non-European isolates of the parasite)
After hatching from eggs in an aquatic environment schisto-some miracidia search for and invade an appropriate snail hostspecies This behavior must be accomplished quickly as miracidiahave a temperature-dependent limited life span of around 20 h asreported for eg T stagnicolae (136) Studies on the miracidialbehavior of T szidati have shown a progression of steps from hostfinding to penetration and migration Miracidia respond to envi-ronmental stimuli such as light or gravity that direct them to themicrohabitat occupied by the host snails (137) Snails release var-ious chemical compounds that form an ldquoactive spacerdquo aroundthem and serve as chemoattractants for the miracidia Miracidiarecognize macromolecular glycoconjugates termed miracidium-attracting glycoproteins (MAGs) or miraxones that consist of aprotein core and carbohydrate chains linked O-glycosidically viaN-acetyl-D-galactosamine and serinethreonine (138 139) Theattractant for miracidia is encoded in these carbohydrate moietiesUpon entering the ldquoactive spacerdquo of the snail miracidia modifytheir movement by increased random turns within the increasingattractant gradient and by a turn-back form of swimming withinthe decreasing attractant gradient (140) This mode of orientation(chemokinesis) observed eg in T szidati or T franki (115 138)results in the first contact of the miracidia with the snail which isthen followed by repeated investigationprobing of the snail sur-face and finally miracidial attachment (140) Penetration of thesnail surface follows although the factors contributing to this pro-cess such as the components of miracidial penetration glandsremain largely unknown
After penetrating the surface epithelium of the snail miracidiaof avian schistosomes transform into mother sporocysts that giverise to daughter sporocysts which migrate to the snail hepatopan-creas where production of the final larval stage the cercariaetakes place (13) The prepatent period lasts about 3 to 10 weeksdepending on several factors such as the miracidial dose or tem-perature (13 141) Infection by avian schistosomes may lead toalterations of the snail internal defense system (IDS) metabolismand endocrine functions Such alterations have been studiedwidely in L stagnalis the intermediate host of T szidati (142ndash145)
The snail IDS is based solely on innate immune mechanismscomposed of humoral and cellular limbs Lectins are essential hu-moral components whereas hemocytes represent the main effec-tor cells (146 147) Both limbs of the L stagnalis IDS appear to beactivated and then suppressed during early and late stages respec-tively of a T szidati infection (143) In vitro hemocytes failed toencapsulate and destroy T szidati sporocysts (148) It has beensuggested that parasite excretory-secretory products participate inthe modulation of hemocyte activities (149) Disruption of hemo-cyte signaling pathways such as protein kinase C (PKC) and ex-tracellular signal-regulated kinase (ERK) pathways may alsoinfluence hemocyte activities (150ndash152) For example carbohy-drates known to be present on larval surfaces of T szidati and Tregenti such as D-galactose and L-fucose (153ndash156) affected he-
mocyte PKC and ERK signaling in L stagnalis which suggests animmunosuppressive role (157 158) However experiments inves-tigating the direct effect of Trichobilharzia larvae on hemocytesignaling have not been performed Alterations of humoral de-fense components also occur during infections by avian schisto-somes and at least two molecules molluscan defense molecule(MDM) and granularin have been investigated in this respect(159 160) Both molecules produced in L stagnalis by granularcells of connective tissue are related to phagocytic activity of he-mocytes MDM enhances phagocytosis of hemocytes and the ex-pression of a corresponding gene for MDM is downregulated in Lstagnalis infected with T szidati (159 161) In contrast treatmentof hemocytes with granularin decreases phagocytic activity andthe encoding gene is upregulated in parasitized snails (160 161)
In the snail host avian schistosomes also interfere with metab-olism such as causing abnormal body growth and endocrinefunctions such as causing a reduction of egg laying (161) In Lstagnalis these processes are regulated by neuroendocrine cells inthe central nervous system (CNS) (162 163) Trichobilharzia szi-dati releases an undescribed substance that induces the snail hostto produce schistosomin (a peptide of 87 kDa consisting of 79amino acids) from its connective tissue and hemocytes (145)Schistosomin acts as a neuropeptide that interferes with somehormones such as calfluxin a neuropeptide that stimulates theinflux of Ca2 into the mitochondria of albumen gland cells (164165) As a consequence ovulation and egg laying are inhibited inthe snail Excitability of neuroendocrine cells (light green cells[LGCs]) responsible for growth (162) increases in response toschistosomin (166) As a result the body size of infected snailsmay become considerably larger than that of uninfected snails(142 161) Other neuropeptides (FMRFamide-related peptides)are also upregulated during infection of L stagnalis by T szidatiand these peptides via inhibition of neuroendocrine cells may beresponsible for the suppression of snail metabolism and repro-duction (167) All these changes in infected snails may provideenergy resources and space that can be exploited by the schisto-somes for development (145 161)
The survival rate of infected snails releasing schistosome cer-cariae as the agent of cercarial dermatitis varies among species Alimited number of studies have focused on survival rates of avianschistosome-infected versus uninfected snails As an example90 of L stagnalis snails infected experimentally with a singlemiracidium of T ocellata (North American isolate) were alive at28 weeks of age (three infected snails were alive for 19 months)whereas all uninfected snails were dead (168) In contrast L stag-nalis or Planorbarius corneus snails naturally infected with T szi-dati or Bilharziella polonica respectively lived a shorter time onaverage than the corresponding uninfected individuals (169170)
VERTEBRATE HOST FINDING AND PENETRATION
Cercariae emerging from the snail intermediate host are the infec-tive stage to the definitive host and are also the stage responsiblefor causing cercarial dermatitis A cercaria is a multicellular larvacomprised of an oblong body and a slender tail that is bifurcated(furcocercous) at the posterior end (Fig 2) Cercariae of schisto-somes leave their snail hosts actively For this purpose they em-ploy a pair of specialized unicellular escape glands that are obviousin mature cercariae within sporocysts Once the cercariae haveemerged only their ducts lined by microtubules are visible sug-
Horaacutek et al
170 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

gesting a release of granular gland content likely containing histo-lytic enzymes during their migration through the snail tissue(171)
Once in the water schistosome cercariae express a complexpattern of behaviors that is composed of movement cycles that arerepeated in defined frequencies (172) In general they expressnegative geotaxy and positive phototaxy which result in the con-centration of cercariae just beneath the water surface where ap-propriate definitive hosts may occur (173) Surface tension of thewater enables clinging of cercariae via their ventral sucker (acetab-ulum) (173) This resting phase is interrupted by phases of activeswimming (173) The effect of light on the behavior of swimmingcercariae was studied in detail for Trichobilharzia szidati Cercar-iae are able to react to a moving shadow stimulus (produced by apotential host) by a burst of forward swimming (body first) awayfrom the source of light to deeper levels where they can encoun-ter the feet of duck definitive hosts A shadow stimulus applied inthe active phase inhibits swimming and prolongs the followingpassive phase (173) Two types of photoreceptors located near thedorsal surface of the body were described for this species a pair oflens-covered pigment cup ocelli and a special type of unpig-mented rhabdomeric photoreceptors composed of three cells ar-ranged in a three-dimensional (3-D) configuration The lens-cov-
ered pigment cup ocelli probably serve to detect the direction ofincoming light and to control the direction of swimming in rela-tion to a light source while the unpigmented photoreceptors serveas monitors for light intensity (174) The pigmented ocelli are alsopresent in other genera of avian (Bilharziella DendritobilharziaGigantobilharzia and Austrobilharzia) and mammalian (Schisto-somatium and Heterobilharzia) schistosomes but are absent inSchistosoma
Moving shadows also trigger a readiness for cercarial attach-ment to substrates further stimulated by thermal and chemicalhost cues (172) As for the latter signals compounds in the hostskin (ceramides and cholesterol) stimulate enduring contact ofcercariae of T szidati with the host skin (173 175) There is vari-ability among schistosome species in responses to light shadowphysical and chemical cues such that for different species someof the signals may not work or may include some additional onessuch as touch water turbulence andor additional chemical com-pounds (172 176) The variation among the different avian schis-tosome species reflects the diversity in biology and ecology ofschistosomes and their adaptations to the spectra of avian hosts(172 176)
Invasion of the bird or mammal skin is initiated by the cercar-iae receiving the proper signal Surprisingly there were few differ-ences between the avian T szidati and human S mansoni organ-isms in their pattern of invasive behavior toward living humanskin most cercariae did not penetrate the skin immediately afterattachment but performed a leech-like creeping which lasted 0 to80 s for T szidati and 15 s to 558 min for S mansoni (177) Suchbehavior guided the cercariae to skin wrinkles or hair follicleswhere most penetration sites were located (178) (Fig 3) Penetra-tion behavior and production of secretions stimulate neighboringcercariae to use the same entry site on the skin (177 178) Invasionof the skin is facilitated by secretions of cercarial penetrationglands released from openings at the apex of the muscular headorgan by spasmodic contractions of cercarial body musculature(179) The head organ performs concurrent thrust movementsagainst the skin surface while the cercaria is firmly attached by theventral sucker (176 178 180) Signals for skin invasion seem to beuniversal for schistosomesmdashfatty acids especially polyunsatu-rated fatty acids containing 18 carbons and two or three cis doublebonds (linoleic and linolenic acids) which are bound in cell mem-branes and occur as free molecules on the surface of human andbird skin (176)
For T szidati the tail is shed within 0 to 105 s after the onset ofpenetration sometimes during creeping This shedding seems tobe generated at least partially by contractions of a muscular collarat the body-tail junction which plays a role in the closure of thecercarial hind body after tail shedding Cercarial penetration oc-curs in a nearly surface-parallel direction while the spined ventralsucker supports squeezing of the cercarial body into the openingin the skin caused by histolytic gland secretions Full penetrationof living human skin was achieved within a mean of 4 min (83 s to133 min) which was significantly faster than the case for thehuman parasite S mansoni (658 min on average) (177) Fasterpenetration of avian schistosome cercariae might be a conse-quence of these parasitesrsquo adaptation to lower concentrations offatty acids in duck skin therefore reaction to higher concentra-tions in human skin may induce faster invasion (177 178) An-other explanation may be that different histolytic enzymes areused for penetration (see below) Skin penetration success rates
FIG 2 Furcocercaria of Trichobilharzia regenti with protruded acetabulum(arrow) lateral view Bar 200 m (Courtesy of J Bulantovaacute reproduced withpermission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 171Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

may vary greatly In experiments with T szidati and human vol-unteers the highest penetration success rate was 49 underneaththe forearm (181)
The ultrastructure and chemical composition of avian schisto-some cercarial penetration glands and their secretions have beenpoorly studied relative to the case for human schistosomes (Schis-tosoma) In fact most of our knowledge is based on only twospecies of Trichobilharzia There are five pairs of unicellular pen-etration glands three pairs are located behind the ventral sucker(postacetabular glands) and two pairs are located around the ven-tral sucker (circumacetabular or preacetabular glands) Theseglands are filled with secretory vesicles that are released throughthe gland processes at the surface of the head organ A 3-D modelof T regenti acetabular glands shows that they occupy more thanone-fourth of the cercarial body volume (postacetabular glandsca 15 and circumacetabular glands ca 12) Differences wereobserved in the appearance of granular materialsecretory vesiclescontained in the glands pH value and the ability to bind variousdyes and fluorescent markers (180 182)
In T szidati proteolytic activity was detected in cercarial glandsecretions induced by linoleic acid This activity was linked withan orthologue of a chymotrypsin-like serine peptidase namedcercarial elastase and characterized from S mansoni and S haema-tobium cercariae A protein on blots of T szidati cercarial secre-tions as well as in histological sections of penetration glands im-munologically cross-reacted with antibodies against elastases of Smansoni and S haematobium (183 184) However in anotherstudy the reaction of antibodies raised against elastase of S man-soni was observed neither with the penetration glands of T szidatinor with the cercarial secretions on Western blots (180) In con-trast high activities of cysteine peptidases were noticed in inducedcercarial secretions and homogenates for T szidati and T regenti(180 185) In addition the presence of cercarial elastase in thelatter species was not confirmed by screening of a cDNA library(186) On the other hand a papain-like cysteine peptidase termedcathepsin B2 was found in the postacetabular penetration glandsof T regenti This enzyme was shown to cleave proteins of the hostskin similar to the case with S japonicum (187 188) Its expres-sion was even higher in intravertebrate stages (schistosomula andadults) suggesting that there are multiple roles of this enzymeduring the life cycle (189) It seems that the use of particular pep-tidase families for skin penetration and tissue invasion is diverseamong schistosomes (190) and may confer host specificity
In the postacetabular penetration glands of T szidati and Tregenti cercariae a lectin(s) specific for -13- and -14-linkedsaccharide chains and their sulfated derivatives is present thoughits biological function is still unknown (180 191) It is interestingthat there are high concentrations of calcium in the circumac-etabular glands of both species (180) Several hypotheses suggestthat the role(s) of calcium in the glands (including those of Smansoni) may be to regulate gland peptidase activity stimulateglycocalyx removal interact with connective tissue proteoglycansregulate host blood coagulation or polymerize the adhesive sub-stance from postacetabular glands However none of these hy-potheses (except for a regulation of peptidase activity) has beenconfirmed adequately (180 192 193) Following contact with thehost skin or a linoleic acid-coated surface (L Mikeš unpublisheddata) cercariae start to expel small amounts of gland content dur-ing the creeping movement which is ldquoprintedrdquo as the cercariaetouch the surfacemdashthese ldquokissing marksrdquo are made of a stickysubstance In S mansoni this substance is a product of the posta-cetabular glands and is composed of neutral and acidic mucosub-stances (194) This product might serve adhesive or enzyme-di-rective functions Similar material is produced by cercariae ofTrichobilharzia spp yet differs in chemical composition (180)Finally the production of three types of eicosanoids by T szidaticercariae is stimulated by linoleate (195) Eicosanoids may have arole in host invasion (vasodilatation) and their involvement inimmune evasion was proven by the inhibition of superoxide pro-duction by human neutrophils (195)
During cercarial penetration dramatic changes of surfacestructures and metabolism lead to the transformation of the cer-caria to a schistosomulum The thick glycocalyx that served as aprotective layer for the free-living cercaria is shed as its carbohy-drate-rich composition is a target of the host complement cascadeIn the schistosome species studied so far the glycocalyx is mark-edly rich in fucose residues (154 196ndash198) There is an obviousloss of saccharide moieties at the surfaces of transformed schisto-somula of T szidati and T regenti leading to reduced immunore-activity and attractiveness for fucose-specific lectins (154 155199 200) Also similar to the case for human schistosomes andmembers of other families of blood flukes the trilaminar surfacemembrane of the outer cercarial tegument gradually changes tothe doubled heptalaminar membrane of the schistosomulumwhich has a protective function against the host immune system(199) In T regenti stimulated by linoleate in vitro shedding of the
FIG 3 Scanning electron micrographs of cercariae of Trichobilharzia regenti penetrating the skin of a duck leg (A) An individual larva entering the skin the tailis still preserved (B) Tails of three cercariae penetrating the skin in a group Bars 100 m (Courtesy of J Bulantovaacute reproduced with permission)
Horaacutek et al
172 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

glycocalyx starts at the anterior of the cercaria surrounding theopenings of penetration glands (J Chaloupeckaacute and L Mikešunpublished data) Sticky products of the glands adhere to thesurface of the body and as the cercaria crawls forward the glyco-calyx with the gland products is shed in a sleeve-like manneruntil it is detached at the end of the hind body (Fig 4) Trans-formed cercariaeschistosomula then lose their osmotic resistancetoward water and become dependent on isosmotic conditions ofthe host (Chaloupeckaacute and Mikeš unpublished data) Whetherany compounds of gland secretions take part directly (eg enzy-matically) in the process of glycocalyx shedding is still unclear
It should be mentioned that cercariae of avian schistosomesreadily penetrate other (soft) tissues and peroral infections ofbirds (definitive hosts) and mice (accidental hosts) with cercariaeof T regenti and T szidati have been confirmed (L Kolaacuterovaacute KBlažovaacute V Pech and P Horaacutek unpublished data) This phenom-enon is also known for mammalian schistosomes of the genusSchistosoma (201ndash203) It is not clear however how much theseperoral infections contribute to the transmission of schistosomesunder natural conditions Due to the features of the esophagealmucosa the penetration of Trichobilharzia cercariae does not re-quire the penetration glands to be emptied completely and theirtails may be preserved for some time The stimuli triggering thispenetration behavior remain unknown
PATHOGENICITY OF AND IMMUNE REACTIONS AGAINSTAVIAN SCHISTOSOMES
Survival and Migration in Avian Hosts
Once the cercariae have transformed into schistosomula andreached their final location it is not clear how long avian schisto-somes live in their definitive hosts For example in experimentalinfections of ducks adults of Trichobilharzia parocellata werefound at 86 days postinfection (pi) (204) Whereas experimentalinfections with T szidati and T regenti last for about 3 to 5 weeks(13 205) the adult worms of a Canadian isolate of T ocellata werefound in the liver at 370 days pi (206) Data describing the schis-tosome life span and length of time for egg release in a bird hostwill be necessary in considering the epidemiology of dermatitisNonetheless avian schistosomes have a preference for two major
habitats within the avian host the visceral venous system (mesen-teric renal cloacal and portal vessels) and the nasal passages (ex-cept for Dendritobilharzia which is found in the arterial system)(13)
Migration and localization have been characterized for a fewvisceral schistosomes and only one (T regenti) of the eight nasalschistosomes (205 207) Visceral schistosomes in birds have amigration pattern similar to those of Schistosoma spp in mam-mals After skin penetration schistosomula of T szidati navigatetoward deeper skin layers by following dark and higher concen-trations of D-glucose and L-arginine (208 209) Once a blood cap-illary is found the worms penetrate it and migrate to the heart andlungs In the lungs the worms enter free air space and then reenterthe blood system (206 210) Finally visceral blood vessels (usuallyportal and mesenteric veins) are the preferred habitat (13) How-ever there are at least two exceptions as follows (i) the adults of TszidatiT ocellata leave the blood system and enter the layers of thehost intestinal wall and mucosa (206 211) and (ii) Dendritobilhar-zia pulverulenta prefers the arterial system of its hosts where it isfound in the lower dorsal aorta and the femoral arteries (212) Forthe nasal schistosome T regenti migration is dramatically differ-ent Schistosomula leave the skin and then seek and penetrateperipheral nerves (Fig 5) to migrate to the spinal cord and brain oftheir host From there the adult worms appear intra- and ex-travascularly in the nasal mucosa (207)
Pathology Caused by Visceral Species in Birds andMammals
Most pathological studies of avian schistosomes in the avian hosthave been detailed for experimental birds though there are a fewreports from wild birds As for accidental mammalian hosts onlyexperimental infections have been evaluated The schistosomulamigrate through the heart and lungs of birds and mammals andonly in the avian host do they reach their final destination Migra-tion through host lungs has been shown to cause damage (16 210213) Infections of duck and mouse lungs by T szidati are accom-panied by hemorrhages in the periphery of the lungs (210) Mi-gration of schistosomula in the lungs of ducks leads to formationof lymphocytic lesions and an influx of macrophages heterophils
FIG 4 Living Trichobilharzia regenti cercaria in vitro shedding its glycocalyx upon stimulation by linoleate The sticky products of the penetration glands stainedwith lithium carmine adhere to the surface of the body and as the cercaria crawls forward by periodical constrictions (A) and extensions (B) the glycocalyx andbound secretions are removed from the surface in a sleeve-like manner Arrows indicate detached sleeve-like remnants of glycocalyx and gland products Bar 100m (Courtesy of J Chaloupeckaacute reproduced with permission)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 173Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

and eosinophils into the afflicted tissue (210) However in mouselungs schistosomula do not evoke a specific inflammatory reac-tion only alveolar congestion and edema are observed (210)Damage to the lung tissue in general and formation of alveolarcongestion in particular might be caused by schistosomula thatleave the blood system and localize extravascularly in the alveolarwalls Their inability to reenter the blood system might be linkedto their relative size and loss of orientation in a noncompatiblemouse host (210) Similarly migrations to mammalian lungs andaccompanied hemorrhages have been observed in hamstersguinea pigs rabbits and rhesus monkeys experimentally exposedto three species of Trichobilharzia (213) Pulmonary infections ofchickens and pigeons with the marine schistosome Ornithobilhar-zia canaliculata led to the development of lesions in the arterialand venous vascular systems (lymphocytic endarteritis periarteri-tis and segmental proliferation of the vascular endothelium) hy-perplasia of smooth muscles in tertiary bronchi and thickening ofalveolar septa Cellular infiltrates consisted mainly of histiocytesheterophils and lymphocytes (16) In contrast to the case withvisceral schistosomes infections of duck and mouse lungs with thenasal species T regenti probably represent an ectopic localizationof schistosomula (207 214 215)
As for patent infections (exclusively in birds) major pathologyis caused by granulomas around eggs and only partly by adult
worms Obliterative endophlebitis caused by adult schistosomes(probably Trichobilharzia filiformis) in the intestinal veins of muteswans (Cygnus olor) has been recorded (216) The intestinal sur-face showed various stages of villous atrophy with intestinal mu-cosal lesions associated with infiltration of the lamina propria ofthe jejunum and ileum by lymphocytes and plasma cells and by asmaller number of heterophils and eosinophils Eggs were ob-served multifocally in the lamina propria of the small and largeintestines and their presence triggered mild to severe granuloma-tous reactions (216) Eggs laid by the adult worms of Austrobilhar-zia variglandis in experimentally infected chickens caused edemacellular infiltration and hyperplasia of smooth muscle of the mus-cularis externa of the intestine Around the eggs mononuclearcells formed early granulomas with a few eosinophils and hetero-phils followed by accumulation of giant and epithelioid cells(217)
Similarly examination of Atlantic brant geese (Branta berniclahrota) infected with Trichobilharzia sp (probably Anserobilharziabrantae based on current taxonomy) revealed the development ofgranulomas around eggs located in the colon (218) Granulomaswere also observed in the duodenum and small intestine of pi-geons infected with Ornithobilharzia canaliculata (16) Three spe-cies of ducks infected with T physellae showed no associated tissuereaction in the vicinity of adults located in mesenteric veins but agranulomatous reaction around the eggs was detected occasion-ally in the mucosa and submucosa of the intestine (219) On theother hand the most serious lesions and fibroplasia of the portaltriads and adjacent parenchyma were observed in the livers ofthose ducks and were attributed to mature T physellae (219) Inthe case of pigeons infected with O canaliculata granulomatouslesions surrounding collapsed eggs were observed in the liver pa-renchyma (16) Intestinal pathology may be accompanied by poornutritional conditions as noticed in pigeons infected with Ocanaliculata (16) Exceptionally a more serious manifestation ofthe infection in naturally infected wild ducks was reported wherethe adults and eggs of T physellae caused partial to complete pa-ralysis of the cervical wing and leg muscles foul-smelling diar-rhea and half-closed pasted eyelids (219)
Pathology Caused by Nasal Species in Birds and Mice
Histological examination of the nasal tissue of birds showed theextra- and intravascular locations of adult worms of T regenti(220) with their first appearance on day 13 pi (205) Immatureeggs appeared from day 15 pi and at day 19 pi eggs were fullydeveloped and observed extravascularly in the nasal mucosa withthe maximum number of eggs seen on day 22 pi (221) The areasurrounding the eggs was infiltrated by numerous eosinophilsheterophils histiocytes and multinucleated giant cells and a fewplasma cells and mononucleated cells (220) The formation ofgranulomas around the eggs was noted from day 22 pi (221)Miracidia that hatched directly in the host tissue were surroundedby lymphocytes eosinophils and heterophils without granulomaformation (221) While an infected bird is drinkingfeeding onlythe miracidia leave the tissue to enter the water which representsan exceptional mode of transmission among schistosomes (205)The presence of adults only did not initiate an influx of immunecells to their vicinity but the presence of large worms and eggscaused the development of focal hemorrhages throughout the na-sal mucosa (207 221) Probably the more devastating aspect of thepathology of this species is the effect on the CNS of hosts (birds
FIG 5 Cercaria of Trichobilharzia regenti in vitro penetrating a peripheralnerve isolated from a duck The tail is already detached and the head organburrows into the nerve Bar 200 m (Courtesy of J Bulantovaacute reproducedwith permission)
Horaacutek et al
174 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

and experimental mammals) As stated above cercariae penetratethe skin and migrate to the nasal passages via the CNS rather thanthe circulatory system
CNS infections of ducks and mice by T regenti may lead to thedevelopment of various transient or permanent neuromotorsymptoms such as weak to severe leg paralysis and balanceorien-tation disorders (207 214 215) In avian hosts schistosomula inthe CNS initiated an accumulation of inflammatory cells such aseosinophils and heterophils that represented the most abundantcell infiltrates yet minimal damage to nervous system cells wasdetected (220) In a few cases however damage to the CNS wasobserved in birds with visceral schistosomes For example gran-ulomatous encephalitis in mute swans caused by Dendritobilhar-zia sp has been described (222 223) Schistosome eggs found inthe cerebrums and cerebellums of naturally infected swans weresurrounded by giant cells macrophages lymphocytes and to alesser extent heterophils and fibroblasts (223) The presence ofDendritobilharzia eggs in the CNS represents an ectopic localiza-tion
Because of the pathology caused by T regenti and thus theimplications for human health most experimental work has beendone in mice rather than birds For up to 3 days pi migration ofschistosomula through the murine nervous tissue did not evokeinflammation or tissue damage and all detected parasites wereintact (215 224) A host reaction to the infection was visible ondays 6 and 7 pi The presence of parasites led to the accumulationof immune cells predominantly microglia macrophages andneutrophils and to a lesser extent CD3 lymphocytes (215 224)
Proliferating astrocytes formed ldquoglial scarsrdquo at the sites of previ-ously migrating schistosomula (215) Ongoing infection was as-sociated with a more intense inflammatory reaction in white andgray matter of the spinal cord Microglia macrophages neutro-phils eosinophils and CD3 lymphocytes participated in the for-mation of inflammatory lesions surrounding the disintegratingschistosomula and damage to the axons was detected (215) (Fig6) The localization of schistosomula outside the solid tissue inthe subarachnoidal space of the spinal cord and the brain and inthe cavity of the 4th ventricle of the brain led neither to damagenor to inflammation of the adjacent nervous tissue (215) It seemsthat schistosomula located in the cavities of CNS were able todelay destruction by the immune cells Nevertheless most of theworms were eliminated by 21 days pi (215) Challenge infectionstriggered a strong immune response which efficiently and rapidlyeliminated the schistosomes (215 224)
In immunodeficient SCID mice primary infections as well asreinfections did not evoke a significant skin reaction and theschistosomula often escaped from the skin to the CNS (224)where migrating schistosomula caused axonal damage and an in-flux of immune cells (215) In comparison to the case with immu-nocompetent mice the schistosomula survived longer in the CNSprobably due to the absence of T and B lymphocytes (215 224)cells that may represent important effectors in destruction ofschistosomula Larger numbers of schistosomula in the CNS andtheir extended time of migration via nervous tissue resulted in ahigher rate of occurrence of paralysis of immunodeficient SCIDmice (215)
FIG 6 Destruction of a schistosomulum (arrows) in the thoracic part of the spinal cord of a BALBc mouse at 21 days pi (longitudinal sections) (A and B)Inflammatory lesion consisting of CD3 lymphocytes (dark spots) (A) and microglia cells (brown-stained ramified cells) (B) that were detected by use ofanti-mouse CD3 and anti-mouse Iba-1 antibodies respectively Nuclei of other cells were stained blue by hematoxylin (C and D) Tissue around theschistosomulum contains damaged axons Axonal damage was accompanied by formation of spheroids (asterisks) in the site of axonal disruption and wasvisualized immunohistochemically by use of anti-mouse nonphosphorylated and phosphorylated neurofilament antibodies (SMI-311 and SMI-312 respec-tively) (C) and anti-mouse -amyloid precursor protein antibodies (D) Bars 100 m (A and B) and 50 m (C and D) (The figure was created by LLichtenbergovaacute)
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 175Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

The above-mentioned damage to the nervous tissues of birdsand mice demonstrates that not only the eggs but also the otherstages of schistosomes are responsible for major pathology In thisparticular case just the schistosomula (migrating juveniles) of Tregenti can be regarded as the most pathogenic stage of the para-site
Skin Immune Response and Cercarial Dermatitis
Skin immune reactions of birds to the penetration of avian schis-tosome cercariae are insufficiently described In any case birds dorespond to penetrating cercariae as shown in a histological obser-vation of chicken skin infected by Ornithobilharzia canaliculataSevere infiltrations of the dermis by histiocytes and heterophilsand aggregation of lymphocytes around dilated capillaries in thedermis were recorded (16) Dead and destructed schistosomulasurrounded by heterophils and histiocytes were found in the epi-dermises of chickens at 12 h pi At 24 h pi lymphocytes histio-cytes and heterophils still persisted in the dermis but the numberof immune cells decreased (16)
Immunohistopathology of cercarial dermatitis in humans rec-ognized three phases of cellular responses (leucocytic lympho-cytic and histiocytic) against Trichobilharzia larvae (225) How-ever because studies on humans are rarely performed (112 225226) a mouse model was established to provide more-detailedstudies on immunohistopathology Primary infection of micewith T regenti causes an acute inflammatory reaction with edemaand vasodilatation (227) Parasites located in the dermis are sur-rounded by large inflammatory cellular foci that are formed byneutrophils macrophages mast cells major histocompatibilitycomplex class II (MHC II) antigen-presenting cells and a smallnumber of CD4 lymphocytes (227) Repeated infections causeperivasculitis folliculitis and substantially more influx of thesame cell types that are noted after primary infection (227) Ex-tensive skin inflammation leads to the formation of large abscessesand subsequently to dermal and epidermal necrosis Sites of pre-vious cercarial penetration are characterized by intraepidermalpustulae and parakeratosis (227)
In vitro culture of skin biopsy specimens from primary mouseinfections by T regenti revealed a release of the acute-phase cyto-kines interleukin-1 (IL-1) and IL-6 and an increased produc-tion of IL-12 (227) Larger amounts of IL-12 correlated with ele-vated production of gamma interferon (IFN-) in the cell culturesupernatant (antigen-stimulated lymphocytes) from the skindraining lymph nodes (227) IFN- and IL-12 are associated withTh1 cell differentiation and IL-1 and IL-6 are important in Th17polarization (228) Although the role of Th17 in host tissue im-munopathology has been described for some infections by hel-minths (eg human schistosomes) (228) participation of Th17cells in the processes associated with skin penetration by avianschistosomes needs to be clarified
Reinfections of mice with T regenti were accompanied byedema that developed as a consequence of local vascular permea-bility caused by histamine released from activated mast cells (227)Histamine has a regulatory function in the Th1 and Th2 polariza-tion of the immune response (229) Mast cells also rapidly releaseda large amount of IL-4 which has been detected in the superna-tants of skin biopsy specimens from T regenti-reinfected mice(227) Like mast cells basophils degranulate and release IL-4 as aresponse to the presence of T regenti antigens (230) Elevation oftotal serum IgE levels implies that histamine and IL-4 production
by mast cells and basophils occurs in an IgE-dependent manner(227 230) Dominance of the Th2 response was also supported byan elevation of antigen-specific IgG1 antibodies and a decrease ofIgG2b antibodies (Th1 associated) in the sera of mice reinfectedwith T regenti (230) Cercarial dermatitis in reinfected mice istherefore Th2 polarized with a response comprised of an earlytype I hypersensitivity reaction and late-phase skin inflammation(227)
It has been shown in several cases that mammals (includinghumans) are unsuitable hosts for avian schistosomes such thatthe worms cannot mature and reproduce (except for Austrobilhar-zia variglandis which is able to reach sexual maturity in the lungsof Meriones unguiculatus gerbils [231]) Nevertheless cercarialdermatitis is probably not the only interaction of avian schisto-somes and mammals Dermatitis develops as an immune (aller-gic) reaction of the already sensitized person it represents a pow-erful protection of the body against worms in the skin Howeverin a naive (nonsensitized) or immunodeficient experimental hostat least some worms survive leave the skin and migrate through-out the body (213 232) Mild to severe consequences of suchmigration may appear (see above) most importantly T regenti isneurotropic and can cause damage to the central nervous system(155 207 214 215 220 224) To date no information aboutmigration of avian schistosomes in human bodies is available Ittherefore seems that laboratory animals (mice rats etc) are in-dispensable for assessing all risks associated with infections ofmammals (humans) by avian schistosomes
DETECTION AND IDENTIFICATION OF AVIANSCHISTOSOMES
Prior to or during the dermatitis season especially following adermatitis outbreak a standard protocol for the detection ofschistosome cercariae involves the collection of and screening forcercarial emergence in snails Usually in a laboratory individualsnails are placed in a beakersmall wells with clean water and ex-posed to a lamp to stimulate shedding of cercariae Subsequentlycercariae are collected and identified under a light microscope Ifno cercariae emerge from the snails there are three options thesnails can be dissected to find the schistosomes the snail tissue canbe pooled and molecular techniques applied to detect a schisto-some infection andor additional snails can be collected fromaffected and surrounding areas (233 234) Detection of microor-ganisms directly from water samples is developing rapidly butamong schistosomes these techniques have been optimized forhuman schistosomes (235 236) Methods thus far to detect avianschistosomes Trichobilharzia in particular (237 238) involveconcentrating the cercariae from a water sample and using a PCRassay to detect a single cercaria in plankton (05 g) and snail tissues(025 g) (239) Using definitive host-seeking behavior cercariaecan be lured to a trap that contains linoleic acid a known stimulusfor cercarial penetration (240) Irrespective of a dermatitis out-break identifying which adult worms can be found in the habitatcan be completed by examination of feces for eggs by using sedi-mentationflotation (241) or Kato-Katz fecal smear methods(242) andor postmortem examination of the arterialvenous sys-tem nasals liver or kidney of the host For nasal schistosomespecies lavage of the nasal cavity of birds represents a method ofchoice After rinsing out the nasal cavity with saline or water freelymoving miracidia (and partly the liberated eggs) can be detectedin the wash fluid (13) The above-described examination of snails
Horaacutek et al
176 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

and birds for avian schistosomes is thoroughly summarized inreference 243
Examination of water or the snail or bird host is important forspecies delineation Not only is there a highly diverse populationof schistosomes especially avian schistosomes that are responsi-ble for dermatitis but these species differ markedly in their mor-phology and pathogenicity in birds and experimental animals (1213) Identification of schistosomes particularly avian schisto-somes is challenging Traditional methods using morphology(adults mostly but also eggs and cercariae) are usually not suffi-cient to separate species particularly within a genus as these char-acteristics can be missing or minute and their recognition oftendepends on the experience of the observer However some char-acteristics have been found to be informative for rough speciesidentification such as the position of internal organs and tegu-ment spination of adults (13) morphology of eggs (244) or dis-tribution of sensory papillae on cercariae (68 198)
In addition to the morphological features there are some non-morphological criteria that can be used to differentiate species forexample the behavior of cercariae (245) compatibility with thesnail hosts and organtissue affinity of adults in birds (135) If eggsand adults from a host are examined it is not always clear whichadultseggs are conspecific or which match cercariae from snailsunless experimental trials are conducted Experimental infectionswith cercariae from snails to obtain adult worms for morpholog-ical assessment are ideal but these are time-consuming and oftenyield low prevalences of infection Because of the reported speciesmorphological and pathogenic diversity of avian schistosomes(12 13) from an epidemiological perspective on dermatitis thereis a need for species delineation and host use and molecular tech-niques have proven a necessary and excellent tool for more rapididentifications (63 70 135) Such techniques have allowed us togenetically connect larval stages from snails to adult stages frombirds greatly advancing the epizootology and epidemiology ofavian schistosomes in particular (10 40 123)
Molecular identification of an avian schistosome providesclues to the host source and biology and possible targets for con-trol There have been steady efforts and testing of markers thathave revealed some consistency and validity for species identifica-tion of schistosomes (123 246 247) Molecular phylogenetics hashad a major impact on the taxonomy and discovery of new lin-eages of avian schistosomes particularly the major etiologicalagents of cercarial dermatitis ie species of Trichobilharzia (1040 63 67 69 70 121ndash123 125 129 135 248ndash251) Most of theeffort has been conducted using three gene regions To date thenuclear ribosomal DNA D1-D2 regions of 28S the internal tran-scribed spacer (ITS) region and the mitochondrial cox1 gene havebeen tested as molecular markers for systematics (40 63 67 6970 105 123 248 249 251) epidemiology (252) and diagnostics(121 253) Sequencing of gene regions such as the nuclear ITSregion and the mitochondrial cox1 gene not only has linked lifecycle stages but also has suggested that our recognition of thediversity of avian schistosomes continues to grow (10 12 40 65ndash67 69 70 105 129 133 135)
While the molecular identification of avian schistosomes is stillin the early stages these markers have thus far proved successful inattributing most samples to a known species At the level of (infr-a)populations genetic diversity of T szidati cercariae from 7 snailscollected at 3 localities in Russia was recently shown by use ofrandomly amplified polymorphic DNA (RAPD) (254) At this
time neither microsatellite markers nor next-generation sequenc-ing protocols have been developed that would allow detection ofspecific populations or host strains The development of genome-wide sequencing protocols for population genetic analyses wouldgreatly aid in identifying the epidemiological determinants of cer-carial dermatitis outbreaks Although genetic identifications haveprovided a framework for circumscribing species and for rapiddetection caution must be exercised since genetic identificationalone is not sufficient in the absence of data on disease dynamicsand morphology (255 256) A species designation should ideallyreflect all data available ie host species location and morpho-logical characteristics of adult worms and eggs In addition com-parative molecular analyses must be performed to obtain reliableand convincing results of species identification
Most importantly specimens and other data eg genetic datashould be archived in a permanent museum collection or an ar-chivable Web-based database for data resulting from a specimensuch as the sequence archive GenBank Voucher samples of anylife cycle stage of these schistosomes (or any parasite and host)should be preserved and deposited in a permanent museum col-lection (257ndash259) This is imperative for several reasons the mostimportant is for the question that has not yet been asked In theevent that the parasite sample does not match known species orhas odd features further work may be necessary Access to imagesand measurements plus an additional sample(s) will be used infurther sequencing Moreover documentation of any species ofschistosome coming through a clinic is an important record thatcan contribute to epidemiological studies especially if it is associ-ated with people affected by dermatitis
CLINICAL FEATURES DIAGNOSIS TREATMENT ANDPROPHYLAXIS OF HUMAN INFECTIONS
As for the clinical symptoms and signs penetration of avian schis-tosome cercariae into mammalian (human) skin may initiate animmediate prickling sensation that persists for approximately 1 h(4 260) The development and intensity of the subsequent allergicreaction depend on the number and duration of previous cercarialcontacts as well as individual susceptibility (4 7 260) The pri-mary contact with cercariae may lead to either an imperceptible(7 261) or mild skin reaction with the development of small andtransient macules maculopapules or inconspicuous papules ofabout 1 to 2 mm after 05 to 2 days pi A delayed reaction in theform of small papules can be observed in some persons as late as 8days pi (7 238)
Repeated infections cause a more pronounced cutaneous reac-tion followed by diffuse edema and development of erythematouspapules or papulovesicles (4 261) More specifically the first tran-sitory macules (up to 10 mm) and primary itching can appear assoon as 4 to 20 min after exposure Thereafter (1 to 15 h pi)macules are replaced by papules (about 3 to 8 mm) and an intenseitching (secondary itching) is experienced In addition erythemaand edema may occur in the afflicted area for a few days Vesiclesof about 1 to 8 mm may form on papules at 2 to 3 days pi and mayrupture as a result of scratching As a consequence bacterial su-perinfection may result in formation of pustules Papules usuallyregress and disappear at 4 to 10 days pi leaving a pigmented spot(about 1 to 4 mm) on the skin for weeks (7 225 238) however insome cases the symptoms may persist for about 20 days pi (7238 262 263) Every maculapapula is a reaction against the pen-etrating cercaria(e) and thus represents the part of the body in
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 177Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

direct contact with cercariae (Fig 7) An attack by many cercariaemay be accompanied by generalized reactions such as limb andlymph node swelling nausea diarrhea and fever (7 238)
Diagnosis is rather problematic Skin reactions to cercariaefrom freshwater or marine environments may resemble insectbites bacterial dermatitis contact dermatitis or skin reactionsagainst nematocysts of larval cnidarians (sea anemones thimblejellyfish etc) (7) Anamnestic data (suggesting recent contactwith water reservoirs) and maculopapular skin eruption on thebody parts that were in contact with water are important indica-tors Direct proof of schistosome infection may be shown by skinbiopsy of the papulae no later than 48 h pi Individual papulaeshould be excisedshaved off under local anesthesia put in Bouinrsquosfixative cut into 10-m sections and stained with hematoxylin-eosin (225 226) As for basic laboratory tests increased eosinophilcounts and elevated levels of total IgE may indicate an attack byavian schistosomes (7 230 238) Specific immunologicalserolog-ical assays (skin test ldquoCercarienhuumlllenreaktionrdquo [a precipitationreaction surrounding the cercarial body in the presence of a spe-cific antibody] indirect fluorescent-antibody test [IFAT] en-zyme-linked immunosorbent assay [ELISA] and complementfixation test) to detect penetration by avian schistosomes are notsufficiently specific or sensitive (7 238) Exceptionally some re-action has been obtained with sera of dermatitis patients and aheterologous antigenmdashSchistosoma mansoni cercariaemdash used inCercarienhuumlllenreaktion and IFAT (263 264) Selection of a reli-able (recombinant) antigen or primer for serological or DNA-based tests respectively is in progress (our unpublished data)
Therapy for afflicted areas includes only symptomatic (notcausal) treatment of the condition in the form of soothing agentsFor example water chestnut planters in India use mustard oil to
relieve the itching and rash in mild cases of dermatitis (265) Inserious cases application of systemic antihistamines (tablets orgels eg hydroxyzine) or mild corticosteroids (eg 01 triam-cinolon cream or 1 hydrocortisone ointment) may be consid-ered (7 226 262 266)
There are several recommendations to protect individualsWearing rubber waders or gloves or neoprene diving suits is 100reliable although not always appropriate or realistic Upon con-tact with cercariae in water the number of penetrating larvae canbe reduced if action is taken within seconds to a few minutes (seethe part on vertebrate host penetration above) For example thor-ough toweling and exposure of skin to the sun are recommendedfor bathers immediately after leaving water In the case of an acci-dental exposure in the laboratory skin can be washed with 70ethanol or warm water (as much as one can tolerate) and soap(our personal experience) Several chemicals have been tested asbarriers to cercarial penetration For example LipoDEET a long-acting liposome formulation of DEET (NN-diethyl-m-tolua-mide) a common safe and available insect repellent has beenused successfully to prevent penetration of S mansoni (267)However a cream formulation of DEET was poorly effectiveagainst T szidati penetrating living human skin (181) Two otherformulations were effective (i) SafeSea lotion against jellyfishstings where the effective compound may be H1-antihistaminediphenhydramine and (ii) niclosamide in a dosage as low as 01in water-resistant sunscreens We hypothesize that sunscreensbased on plant oils (nonmedicated) will trigger cercarial penetra-tion due to a high content of unsaturated fatty acids In this regarda negative effect in terms of human protection was observed fordimethicones (polydimethylsiloxanes and silicone oils) which arecommon ingredients in many skin care products (181)
FIG 7 Development of cercarial dermatitis on the dorsal (1) and ventral (2) parts of the left hand of a sensitized volunteer infected experimentally byTrichobilharzia szidati (the whole hand was immersed into a beaker containing water with cercariae) Images were taken at 1 (A) 2 (B) 3 (C) and 4 (D) dayspostexposure Formation of macules (A) and papules with vesicles (B to D) can be seen No penetrationreaction was recorded for the palm Noticeable swellingof the hand is shown mainly in panel B1 (The figure was created by H Kuliacutekovaacute and P Horaacutek)
Horaacutek et al
178 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

ECOLOGICAL FACTORS INFLUENCING THE OCCURRENCE OFAVIAN SCHISTOSOMES AND CERCARIAL DERMATITIS
Due to climate changes and land alterations the seasonal windowfor parasite transmission may become longer in addition tochanges in the behavioral and physiological patterns of parasitehosts For example the accelerated growth of both snail and trem-atode larval populations (71) or changes in phenology of aquaticmigratory birds to sedentary (268ndash270) can increase chances foroutbreaks of cercarial dermatitis in both space and time
Global Warming and Eutrophication
At least in temperate climates schistosome cercariae occur seasonallyonce the ambient temperature is high enough for snail activity andthus cercarial emergence (271) Also snails infected during late sum-mer can survive through the winter season and serve as a source ofcercariae in spring (13 272 273) Global warming is predicted to havea positive effect on trematode intramolluscan stages because theirdevelopment is strongly temperature dependent leading to increasedcercarial emission rates For example a shorter prepatent period inrelation to higher temperatures was demonstrated in experimentalinfections with T szidati (141 274) A slight rise in temperature willincrease developmental rates and transmission success (number ofcercariae available to find the definitive host) For example a 5-foldincrease in cercarial emergence with a 10degC increase in temperaturewas recorded for Trichobilharzia sp from Radix peregra (275) Inaddition a link has been suggested between the rise of 08degC withinthe last century and the increased frequency of cercarial dermatitisand Trichobilharzia prevalence at higher latitudes in Europe (9 276)What does not change is the life span of cercariae which is limited tothe restricted source of reserve glycogen (cercariae do not take up anyfood) In this regard temperature strongly affects cercarial survivalFor example the half-life of T szidati was 16 h at 16degC but only 78h at 25degC (169) and the life span of Trichobilharzia arcuata was 72h at 4degC but only 48 h at 26degC (277) Infected snails change theirthermal microhabitat selection eg Lymnaea stagnalis infected withT szidati prefers a colder water temperature than that preferred byuninfected snails (1997degC versus 25degC) This may be an adaptationfor either the snail or the schistosome because at these temperatureslarval development is slower and the rate of cercarial emission islower leading to less tissue damage in the snail (169 170)
Eutrophication promotes excessive plant growth and decaycausing alterations in the dynamics of freshwater communitiesand thus leading to an increased risk of acquiring cercarial derma-titis (11 278ndash280) Greater biomass of primary producers is asso-ciated with faster development and growth of snails due to theincrease in food availability (280ndash282) Dense snail populationsmay increase the probability that a miracidium will find a snailand lead to a higher prevalence of infection In addition aquaticbirds are attracted to such nutrient-rich environments also con-tributing to an increase in parasite transmission (283) Using datafrom a mark-recapture study of L stagnalis in eutrophic fish-ponds rapid trematode recruitment into the snail populationswas demonstrated (284) Maximum annual rates of colonizationrecruitment for T szidati were shown to reach up to 300 suchthat the odds of trematode establishment in an individual snailwere 3 times per year versus once in 10 years in other mainlymarine trematode-snail systems (284 285)
Large snail populations with a high prevalence of avian schisto-somes have been reported for several localities in Europe (71
286ndash288) North America (289 290) and Australia (291) Usu-ally natural preserves support diverse and abundant populationsof potential hosts of schistosomes so these areas may serve as hotspots for outbreaks of cercarial dermatitis Although eutrophica-tion contributes to infection risk cases of cercarial dermatitis orfindings of avian schistosomes have also been reported from oli-gotrophic or mesotrophic systems (8) In addition other abioticand biotic factors and human-induced habitat alterations mayinfluence the occurrence of schistosomes and cercarial dermatitissuch as altered hydrology conditions with water-level fluctuationice cover acidification or dam constructions (268 292 293) an-thropogenic pollutants (268 294ndash297) biodiversity change interms of introducing nonindigenous species that may affect en-demic parasites (298) host susceptibility or resistance (9) preda-tion upon trematode free-swimming larval stages by fish andother aquatic animals (295 299 300) or interspecific competitionof parasites within the same snail host (301ndash303)
Recreational Activities and Cercarial Dermatitis
The data in the literature suggest that lakes represent high-riskareas for cercarial dermatitis as they are attractive for a large num-ber of people usually for recreational purposes (8) Cercarial der-matitis develops as a result of sensitization of the human immunesystem (232 260) and repeated exposures to cercariae influencethe occurrence and intensity of subsequent infections For exam-ple longer exposure in water increases cercarial penetration viafrequent water visits and more time spent in shallow water (4304ndash306) While gender does not influence the risk of infection (4304 305 307) some studies found a higher risk among children ofless than 15 years of age since they tend to spend more time inshallow water (280 308 309) There are rare cases of nonsensitiveindividuals that may be due to host desensitization (310) or indi-vidual nonsusceptibilitynonattractivity to cercariae (4 311)
Locally the highest risk of infection usually occurs in shallowwarm and vegetation-rich shore areas where the snails accumu-late and release cercariae (308 309) Cercariae of avian schisto-somes are concentrated just beneath the water surface (see theinformation on host finding) so swimming in deeper water mayreduce the infection probability (280) However snail preferencesfor habitat differ Lymnaea stagnalis is found mostly in patches ofaquatic vegetation feeding on periphyton Radix spp accumulatein vegetation-free areas on stones or muddy sediments and phy-sid snails prefer detritus as a source of food (312 313) In additioncercariae of avian schistosomes can be transported by wind andwater currents for several kilometers (2 304 305) Therefore tak-ing the local situation into account is essential for risk assessmentbecause conditions unsuitable for one snail species may providean ideal habitat for another
Season and time of day were shown to have considerable effectsMost cases of cercarial dermatitis correlate with high air and watertemperatures during the summer months when schistosome devel-opment in snails is amplified and emission rates of cercariae are stim-ulated by sunlight (4 11 279 280 304 305 309 314) Productioncan reach several thousands of cercariaesnailday (169 300 315)and these emission rates may even counterbalance the loss of cercarialcapability to survive and infect hosts at high temperatures (seeabove) Irrespective of the season outbreaks of cercarial dermatitishave been reported in geothermally heated lakes and ponds in Iceland(71) Time of day proved to be another risk factor Production ofschistosome cercariae is usually discontinuousmdashparticular species
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 179Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

may differ in circadian rhythms of cercarial release This is usuallyattributed to the peak activity of the preferred definitive hosts ThusTrichobilharzia and some other genera infecting aquatic birds leavethe snail host during the light period of the day mainly in the morn-ing hours (2 4 280 305) Increased illumination temperature andsnail locomotory activity trigger cercarial emergence (169 211 316317) Patterns of cercarial emergence can be changed almost imme-diately by reversing the dark-light regimen suggesting that light has adecisive influence on the direction of the process (317 318) Thenumber of cercariae released from snails may vary greatly dependingon the parasitehost species and the phase (age) of infection largespecies of snails represent a higher risk as more cercariae are pro-duced per snail and released into the environment (10 275 315) Thismay also explain why some species of avian schistosomes that usesmaller species of snails are not often detected during an outbreak
Control Measures Related to the Ecology of AvianSchistosomes
In order to reduce the risk of cercarial dermatitis either water useduring peak cercarial emergence should be avoided or the lifecycle of avian schistosomes should be interrupted Interruption ofthe schistosome life cycle has been tried on a number of occasionsOne option is to reduce the prevalence of adult avian schistosomesin birds by eliminating ducks from high-risk areas However pub-lic acceptance of regular hunting or repelling of birds (ducks) atrecreational lakes during summer months is uncertain Anotheroption is to treat birds with the antihelminthic drug praziquantelBirds are captured in the field by use of modified dive traps andsubsequently treated with praziquantel and then released Recap-ture results showed a significant reduction of infections in mal-lards (319ndash321) Furthermore in the subsequent year there was amarked decline in prevalence of avian schistosomes in snails(320) However treatment of the entire duck population is time-consuming and labor-intensive
Reduction or elimination of snail populations is another strat-egy of cercarial dermatitis control by interrupting the life cycle Inthe past application of commercial molluscicides such as niclos-amide or copper sulfate was common (319) However repeatedand unlimited treatments had a deleterious effect on animal com-munities In addition after repeated exposures some snails maybecome resistant to copper sulfate or avoid the treatment by bur-rowing into the mud (322 323) Manual collection of snails (309324) or destruction of their habitats by local removal of littoralvegetation (2) represents a less harmful approach to snail controlLarge-scale destruction of snail populations by use of heavy ma-chines along the shores of Annecy Lake (France) and Cultus Lake(Canada) led to substantial reductions of cercarial dermatitis (2)As for the biological control of snail populations use of mollus-civorous fish or prawns as natural snail predators is promising forlong-term control (325ndash328) although it represents a risk for na-tive flora and fauna if nonindigenous organisms are introduced
Free-swimming miracidia and cercariae may represent a prom-ising target for life cycle interruption Snail finding and recogni-tion by miracidia are based markedly on recognition of host mol-ecules called miraxones (see the information on intramolluscandevelopment) which led to a proposal to use miracidial trapscontaining miraxones (329) Unfortunately due to the high levelof diversity of avian schistosomes and presumed variability ofsnail recognition (139) this proposal proved unrealistic Simi-larly cercarial traps based on species-specific host-finding behav-
ior (see the information on vertebrate host finding) might providelocal protection (330) For example fatty acid-stimulated trans-formation to the schistosomulum stage could be initiated in thesetraps leading to the loss of resistance against a hypo-osmotic wa-ter environment (178 331 332) As far as trophic interactions oforganisms are concerned predation by small aquatic animals mayrepresent a promising means of biological control Schistosomemiracidia and cercariae may serve as prey for larval aquatic insectscrustaceans oligochaetes shrimps and fish (271 333ndash338) Inparticular the annelid Chaetogaster limnaei sensu lato living com-mensally or parasitically on the shell surface or in the mantle andpulmonary cavities of freshwater snails may act as an efficientpredator and prevent penetration of miracidia (339 340) or feedon cercariae as they are released from the snail (341 342)
Finally competitive interactions among trematode larval com-munities within an individual snail can substantially influence es-tablishment and survival of schistosome intramolluscan stages Interms of interspecific interactions particular species of trema-todes can be dominant usually by producing competitively ag-gressive rediae or subordinate because they have less aggressivesporocysts and no redial stage Schistosomes belong to the lattergroup and their sporocysts are eliminated via the predatory inter-actions of redia-producing trematodes eg echinostomes (301343) Yet schistosomes may exert a dominant effect in some casesThe avian schistosome Trichobilharzia brevis persists in coinfec-tions with two dominant echinostomes Echinostoma audyi andHypoderaeum dingeri (343) and can cause developmental sup-pression of the latter species The nonpredatory exclusion of Hdingeri by T brevis has also been shown experimentally (344)Intertrematode competition was recently identified as an impor-tant determinant in transmission of avian schistosomes For ex-ample competitive exclusion was estimated to result in an 180reduction of Trichobilharzia szidati in Lymnaea stagnalis (345)Interestingly T szidati may represent a subordinate species yet itfrequently cooccurs in snails with other trematodes Therefore Tszidati may be either an obligate secondary invader of snails with acompromised immune system or a schistosome that can coexist indouble infections as was demonstrated for other species ofTrichobilharzia and Austrobilharzia (302 343 346)
PERSPECTIVES
The more we know about avian schistosome diversity and distri-bution the better we are positioned to understand their evolu-tionary history and potential for the future dynamics of dermatitisoutbreaks These days we have practical means for their identifi-cation and can better gauge the likelihood that some of these spe-cies may emerge in new contexts to cause unexpected problemsAn effort to describe the avian schistosome species diversity intheir hosts in the circumpolar regions where most migratorybirds spend the summer (and young birds become infected)seems to be a critical component to understanding the epidemi-ology of cercarial dermatitis In addition the condition itself andthe host-parasite interaction require further characterization atthe molecular level At least three examples of future applicationscan be mentioned (i) A more detailed knowledge of cercarialpenetration mechanisms may help in the prevention of dermatitisby introducing new formulations containing inhibitory mole-cules (ii) Characterization of biologically active secretions pro-duced by avian schistosomes may help us to understand the mo-lecular basis of tissue pathology in avian and mammalian hosts
Horaacutek et al
180 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

(iii) Newly designed primers or carefully selected antigens may bedeveloped for reliable antibody- or DNA-based diagnostic toolsSuch tools may be used to screen people or animals with healthproblems following recent water contact to assess the risk associ-ated with exposure to cercariae of avian schistosomes
ACKNOWLEDGMENTS
The University of New Mexico partially supported this study through aNational Institutes of Health grant to Eric S Loker (R01 AI101438) and aNational Science Foundation grant to SVB (DEB 1021427) MS isgrateful for financial support from the Institute of Parasitology (RVOgrant 60077344) Biology Centre of the Academy of Sciences of the CzechRepublic The scientific activities of PH LM LL and VS were finan-cially supported by Charles University in Prague (research program P41PrFUK UNCE grant 204017 GAUK grant 435911 and grant SVV2600742014) and the Czech Science Foundation (grant 13-29577S)
REFERENCES1 Cort WW 1928 Schistosome dermatitis in the United States (Michi-
gan) JAMA 901027ndash1029 httpdxdoiorg101001jama192802690400023010
2 Leighton BJ Nervos S Webster JM 2000 Ecological factors in schis-tosome transmission and an environmentally benign method for con-trolling snails in a recreational lake with a record of schistosome derma-titis Parasitol Int 499 ndash17 httpdxdoiorg101016S1383-5769(99)00034-3
3 Nesterenko SV Malko NJ 2007 Dynamics of human allergic derma-tites caused by schistosomes in the Mjadel district (1995ndash2006) p 202ndash207 In Problems of cercarial dermatitis in the Naroch district Scien-tific seminar Medisont Minsk Belarus
4 Chamot E Toscani L Rougemont A 1998 Public health importanceand risk factors for cercarial dermatitis associated with swimming inLake Leman at Geneva Switzerland Epidemiol Infect 120305ndash314 httpdxdoiorg101017S0950268898008826
5 Hunter GW Ritchie LS Tanabe H 1951 The epidemiology of schis-tosome dermatitis (lsquokoganbyorsquo) in Japan Trans R Soc Trop Med Hyg45103ndash112 httpdxdoiorg101016S0035-9203(51)90604-9
6 Beer SA Voronin MV 2007 Cercarioses in urbanized ecosystemsNauka Moscow Russia
7 Kolaacuterovaacute L Horaacutek P Skiacuternisson K Mareckovaacute H Doenhoff M 2013Cercarial dermatitis a neglected allergic disease Clin Rev Allergy Immu-nol 4563ndash74 httpdxdoiorg101007s12016-012-8334-y
8 Soldaacutenovaacute M Selbach C Kalbe M Kostadinova A Sures B 2013Swimmerrsquos itch etiology impact and risk factors in Europe TrendsParasitol 2965ndash74 httpdxdoiorg101016jpt201212002
9 Horaacutek P Kolaacuterovaacute L 2011 Snails waterfowl and cercarial dermatitisFreshwater Biol 56779 ndash790 httpdxdoiorg101111j1365-2427201002545x
10 Brant SV Loker ES 2009 Schistosomes in the southwest United Statesand their potential for causing cercarial dermatitis or lsquoswimmerrsquos itchrsquo JHelminthol 83191ndash198 httpdxdoiorg101017S0022149X09308020
11 Valdovinos C Balboa C 2008 Cercarial dermatitis and lake eutrophi-cation in south-central Chile Epidemiol Infect 136391ndash394 httpdxdoiorg101017S0950268807008734
12 Brant SV Loker ES 2013 Discovery-based studies of schistosome di-versity stimulate new hypotheses about parasite biology Trends Parasi-tol 29449 ndash 459 httpdxdoiorg101016jpt201306004
13 Horaacutek P Adema CM Kolaacuterovaacute L 2002 Biology of the schistosomegenus Trichobilharzia Adv Parasitol 52155ndash233 httpdxdoiorg101016S0065-308X(02)52012-1
14 Loker ES Brant SV 2006 Diversification dioecy and dimorphism inschistosomes Trends Parasitol 22521ndash528 httpdxdoiorg101016jpt200609001
15 Grodhaus G Keh B 1958 The marine dermatitis-producing cercariaeof Austrobilharzia variglandis in California (Trematoda Schistosoma-tidae) J Parasitol 44633ndash 638 httpdxdoiorg1023073274549
16 Morales GA Helmboldt CF Penner LR 1971 Pathology of experimen-tally induced schistosome dermatitis in chickens the role of Ornithobil-
harzia canaliculata (Rudolphi 1819) Odhner 1912 (Trematoda Schisto-somatidae) Avian Dis 15262ndash276 httpdxdoiorg1023071588696
17 Penner LR 1950 Cercaria littorinalinae sp nov a dermatitis-producingschistosome larvae from the marine snail Littorina planaxis Philippi JParasitol 36466 ndash 472 httpdxdoiorg1023073273174
18 Orris L Combes FC 1952 Clam diggerrsquos dermatitis schistosome der-matitis from sea water Arch Derm Syphilol 66367ndash370 httpdxdoiorg101001archderm195201530280071011
19 Sindermann CJ Gibbs RF 1953 A dermatitis producing schistosomewhich causes ldquoclam diggers itchrdquo along the central Maine coast MaineDept Sea Shore Fish Res Bull 12143ndash162
20 Bearup AJ 1955 A schistosome larva from the marine snail Pyrazusaustralis as a cause of cercarial dermatitis in man Med J Aust 1955ndash960
21 Sindermann CJ 1960 Ecological studies of marine dermatitis-producing schistosome larvae in northern New England Ecology 41678 ndash 684 httpdxdoiorg1023071931801
22 Short RB Holliman RB 1961 Austrobilharzia penneri a new schisto-some from marine snails J Parasitol 47447ndash 452 httpdxdoiorg1023073275378
23 Rohde K 1977 The bird schistosome Austrobilharzia terrigalensis fromthe Great Barrier Reef Australia Z Parasitenkd 5239 ndash51 httpdxdoiorg101007BF00380557
24 Appleton CC 1989 Translocation of an estuarine whelk and its trema-todes parasites in Australia Environ Conserv 16172ndash182 httpdxdoiorg101017S0376892900008973
25 Leighton BJ Ratzlaff D McDougall D Stewart G Nadan A GustafsonL 2004 Schistosome dermatitis at Crescent Beach Preliminary reportEnviron Health Rev 485ndash13
26 Walker JC 2005 Medical importance marine schistosome dermatitis p439 ndash 442 In Rhodes K (ed) Marine parasitology CSIRO PublishingCollingwood Victoria Australia
27 Al-Kandari WY Al-Bustan SA Isaac AM George BA Chandy BS2012 Molecular identification of Austrobilharzia species parasitizingCerithidia cingulata (Gastropoda Potamididae) from Kuwait Bay J Hel-minthol 86470 ndash 478 httpdxdoiorg101017S0022149X11000733
28 Boros DL 1989 Immunopathology of Schistosoma mansoni infectionClin Microbiol Rev 2250 ndash269
29 Buckley JJC 1938 On a dermatitis in Malays caused by the cercariae ofSchistosoma spindale Montgomery 1906 J Helminthol 16117ndash120 httpdxdoiorg101017S0022149X00018368
30 Anantaraman M 1958 On schistosome dermatitis I Dermatitis in In-dia caused by cercariae of Schistosoma spindale Montgomery 1906 In-dian J Helminthol 1046 ndash52
31 Islam KS Awal MA 1991 Schistosome dermatitis in Bangladesh In-dian J Parasitol 15165ndash169
32 Narain K Mahanta J Dutta R Dutta P 1994 Paddy field dermatitis inAssam a cercarial dermatitis J Commun Dis 2626 ndash30
33 Narain K Rajguru SK Mahanta J 1998 Incrimination of Schistosomaspindale as a causative agent of farmerrsquos dermatitis in Assam with a noteon liver pathology in mice J Commun Dis 301ndash 6
34 Agrawal MC Rao VG Vohra S Bhondeley MK Ukey MJ AnvikarAR Yadav R 2007 Is active human schistosomiasis present in IndiaCurr Sci 92889
35 Rao VG Dash AP Agrawal MC Yadav RS Anvikar AR Vohra SBhondeley MK Ukey MJ Das SK Minocha RK Tiwari BK 2007Cercarial dermatitis in central India an emerging health problem amongtribal communities Ann Trop Med Parasitol 101409 ndash 413 httpdxdoiorg101179136485907X176463
36 Wang CR Chen J Zhao JP Chen AH Zhai YQ Li L Zhu XQ 2009Orientobilharzia species neglected parasitic zoonotic agents Acta Trop109171ndash175 httpdxdoiorg101016jactatropica200811009
37 Agrawal MC 2012 Schistosomes and schistosomiasis in South AsiaSpringer New Delhi India
38 Devkota R Brant SV Thapa A Loker ES 2014 Sharing schistosomesthe elephant schistosome Bivitellobilharzia nairi also infects the greaterone-horned rhinoceros (Rhinoceros unicornis) in Chitwan National ParkNepal J Helminthol 8832ndash 40 httpdxdoiorg101017S0022149X12000697
39 Mudaliar SV Ramanujachary G 1945 Schistosoma nairi n sp from anelephant Indian Vet J 121ndash5
40 Brant SV Morgan JAT Mkoji GM Snyder SD Rajapakse JRPV LokerES 2006 An approach to revealing blood fluke life cycles taxonomy anddiversity provision of key reference data including DNA sequence from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 181Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

single life cycle stages J Parasitol 9277ndash 88 httpdxdoiorg101645GE-35151
41 Brant SV Pomajbiacutekovaacute K Modryacute D Petrželkovaacute KJ Loker ES 2013Molecular phylogenetics of the elephant schistosome Bivitellobilharzialoxodontae (Trematoda Schistosomatidae) from Central African Repub-lic J Helminthol 87102ndash107 httpdxdoiorg101017S0022149X1200003X
42 Vogel H Minning W 1940 Bilharziose bei Elefanten Arch Schiffs TropHyg 42562ndash574
43 Thurston JP 1963 Schistosomes from Hippopotamus amphibious L IThe morphology of Schistosoma hippopotami sp nov Parasitology 5349 ndash54
44 Thurston JP 1964 Schistosomes from Hippopotamus amphibious L IIThe morphology of Schistosoma edwardiense sp nov Parasitology 5467ndash72
45 Thurston JP 1971 Further studies on Schistosoma hippopotami andSchistosoma edwardiense in Uganda Rev Zool Bot Afr 84145ndash152
46 Cort WW 1914 Larval trematodes from North American fresh-watersnails J Parasitol 165ndash 84 httpdxdoiorg1023073271183
47 Cort WW 1936 Studies on schistosome dermatitis IV Further infor-mation on distribution in Canada and the United States Am J Hyg(Lond) 24318 ndash333
48 Penner LR 1942 Studies on dermatitis producing schistosomes in east-ern Massachusetts with emphasis on the status of Schistosomatium path-locopticum Tanabe 1923 J Parasitol 28103ndash112 httpdxdoiorg1023073272720
49 Malek EA Armstrong JC 1967 Infection with Heterobilharzia ameri-cana in primates Am J Trop Med Hyg 16708 ndash714
50 Wall RC 1968 ldquoSwimmerrsquos itchrdquo in the Great Lakes region Wards Bull71ndash7
51 Price HF 1931 Life history of Schistosomatium douthitti (Cort) Am JHyg (Lond) 13685ndash727
52 Malek EA 1977 Geographical distribution hosts and biology of Schis-tosomatium douthitti (Cort 1914) Price 1931 Can J Zool 55661ndash 671httpdxdoiorg101139z77-087
53 McKown RD Veatch JK Fox LB 1991 New locality record for Het-erobilharzia americana J Wildl Dis 27156 ndash160 httpdxdoiorg1075890090-3558-271156
54 Corapi WV Snowden KF Rodrigues A Porter FB Buote MA BirchSM Jackson ND Eden KB Whitley DB Mansell J Edwards JF HardyJ Chaffin MK 2012 Natural Heterobilharzia americana infection inhorses in Texas Vet Pathol 49552ndash556 httpdxdoiorg1011770300985811432346
55 Liu Z Bai G 1976 On bird schistosomes from Jilin Province Jilinobil-harzia crecci gen nov sp nov (Schistosomatidae Bilharziellinae) with adiscussion on the taxonomy of the subfamily Bilharziellinae Acta ZoolSin 22385ndash392
56 Leigh WH 1955 The morphology of Gigantobilharzia huttoni (Leigh1953) an avian schistosome with marine dermatitis producing larvae JParasitol 41262ndash269 httpdxdoiorg1023073274202
57 Szidat L 1958 Investigaciones sobre Cercaria chascomusi n sp agentecausal de una nueva enfermedad humana en la Argentina lsquola dermatitisde los banistas de la laguna Chascomusrsquo Bol Museo Argent Cienc Nat181ndash13
58 Ito J 1960 Studies on the morphology and life cycle of Pseudobilharziellacorvi Yamaguti 1941 (Trematoda Schistosomatidae) Jpn J Med Sci Biol1353ndash58 httpdxdoiorg107883yoken19521353
59 Ewers WH 1961 A new intermediate host of schistosome trematodesfrom New South Wales Nature 190283ndash284
60 Leedom WS Short RB 1981 Cercaria pomaceae sp n a dermatitisproducing schistosome cercaria from Pomacea paludosa the Florida ap-ple snail J Parasitol 67257ndash261 httpdxdoiorg1023073280646
61 Martorelli SR 1984 Sobre una cercaria de la familia Schistosomatidae(Digenea) parasita de Chilina gibbosa Sowerby 1841 en el Lago Pel-legrini Provincia de Rio Negro Republica Argentina Neotropica 3097ndash106
62 Blair D Davis GM Wu B 2001 Evolutionary relationships between trem-atodes and snails emphasizing schistosomes and paragonimids Parasitology123S229ndashS243 httpdxdoiorg101017S003118200100837X
63 Aldhoun JA Faltyacutenkovaacute A Karvonen A Horaacutek P 2009 Schistosomesin the North a unique finding from a prosobranch snail using moleculartools Parasitol Int 58314 ndash317 httpdxdoiorg101016jparint200903007
64 Brant SV Cohen AN James D Hui L Hom A Loker ES 2010Cercarial dermatitis transmitted by an exotic marine snail Emerg InfectDis 161357ndash1365 httpdxdoiorg103201eid1609091664
65 Karamian M Aldhoun JA Maraghi S Hatam G Farhangmehr BSadjjadi SM 2011 Parasitological and molecular study of the furcocer-cariae from Melanoides tuberculata as a probable agent of cercarial der-matitis Parasitol Res 108955ndash962 httpdxdoiorg101007s00436-010-2138-x
66 Schuster RK Aldhoun JA OrsquoDonovan D 2014 Gigantobilharzia mela-noidis n sp (Trematoda Schistosomatidae) from Melanoides tuberculata(Gastropoda Thiaridae) in the United Arab Emirates Parasitol Res 113959 ndash972 httpdxdoiorg101007s00436-013-3728-1
67 Aldhoun JA Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2009 Bird schisto-some diversity in Iceland molecular evidence J Helminthol 83173ndash180httpdxdoiorg101017S0022149X09289371
68 Aldhoun JA Podhorskyacute M Holickaacute M Horaacutek P 2012 Bird schisto-somes in planorbid snails in the Czech Republic Parasitol Int 61250 ndash259 httpdxdoiorg101016jparint201110006
69 Jouet D Ferteacute H Hologne C Kaltenbach ML Depaquit J 2009 Avianschistosomes in French aquatic birds a molecular approach J Helmint-hol 83181ndash189 httpdxdoiorg101017S0022149X09311712
70 Brant SV Loker ES 2009 Molecular systematics of the avian schisto-some genus Trichobilharzia (Trematoda Schistosomatidae) in NorthAmerica J Parasitol 95941ndash963 httpdxdoiorg101645GE-18701
71 Skiacuternisson K Aldhoun JA Kolaacuterovaacute L 2009 A review on swimmerrsquositch and the occurrence of bird schistosomes in Iceland J Helminthol83165ndash171 httpdxdoiorg101017S0022149X09336408
72 Fain A 1955 Sur une furcocercaire du groupe Ocellata produisant ex-perimentalement la dermatitie des nageurs a Astrida (Ruanda-Urundi)Ann Soc Belg Med Trop 35701ndash707
73 De Meillon B Stoffberg N 1954 Swimmerrsquos itch in South Africa S AfrMed J 281062ndash1064
74 Appleton CC Brock K 1986 The penetration of mammalian skin bycercariae of Trichobilharzia sp (Trematoda Schistosomatidae) fromSouth Africa Onderstepoort J Vet Res 53209 ndash211
75 Johnston TH 1941 Batherrsquos itch (schistosome dermatitis) in the Mur-ray swamps South Australia Trans R Soc S Aust 65276 ndash284
76 Macfarlane WV 1944 Schistosome dermatitis in the southern lakes aninvestigation of swimmersrsquo itch N Z Med J 43136 ndash140
77 Macfarlane WV 1952 Schistosome dermatitis in Australia Med J Aust39669 ndash 672
78 Blair D Islam KS 1983 The life cycle and morphology of Trichobilhar-zia australis n sp (Digenea Schistosomatidae) from the nasal bloodvessels of the black duck (Anas superciliosa) in Australia with a review ofthe genus Trichobilharzia Syst Parasitol 589 ndash117 httpdxdoiorg101007BF00049237
79 Featherston DW Weeks PJ Featherston N 1988 Schistosome derma-titis in Lake Wanaka Cercaria longicauda prevalence in Lymnaea tomen-tosa 1978 ndash 83 N Z J Zool 15381ndash386 httpdxdoiorg10108003014223198810422963
80 Sandosham AA 1953 Malaysian parasites XII Cercarial dermatitiscaused by a member of the ldquoelvaerdquo group Malaya 26195ndash198
81 Margono SS 1968 Cercaria of Trichobilharzia brevis Basch 1966 inDjakarta as a possible cause of schistosome dermatitis Med J Malaya22306 ndash312
82 Wooi CE Hong SLL Ambu S 2007 Cercarial dermatitis in KelantanMalaysia an occupation related health problem Int eJ Sci Med Educ169 ndash73
83 Farahnak A Essalat M 2003 A study on cercarial dermatitis in Khuz-estan province southwestern Iran BMC Public Health 335ndash38 httpdxdoiorg1011861471-2458-3-35
84 Athari A Gohar-Dehi S Rostami-Jalilian M 2006 Determination ofdefinitive and intermediate hosts of cercarial dermatitis-producingagents in northern Iran Arch Iran Med 911ndash15
85 Maleki SH Athari A Haghighi A Taghipour N Gohardehi SHSeyyed Tabaei S 2012 Species identification of birds nasal Trichobilhar-zia in Sari North of Iran Iran J Parasitol 782ndash 85
86 Gohardehi S Fakhar M Madjidaei M 2013 Avian schistosomes andhuman cercarial dermatitis in a wildlife refuge in Mazandaran provincenorthern Iran Zoonoses Public Health 60442ndash 447 httpdxdoiorg101111zph12020
87 Kullavanijaya P Wongwaisayawan H 1993 Outbreak of cercarial der-
Horaacutek et al
182 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

matitis in Thailand Int J Dermatol 32113ndash115 httpdxdoiorg101111j1365-43621993tb01448x
88 Liu Z Minsheng C Hui K Lin L 1980 A survey of the etiological agentof paddy field dermatitis with studies on the life cycle of Trichobilharziajianensis Liu et al 1977 Acta Zool Sin 26153ndash159
89 Li C Ma C Gu J Lu W 1998 An epidemiological survey of schisto-some cercarial dermatitis among the residents living along the banks ofthe Huaihe River Chin J Parasitol Parasit Dis 16384 ndash387
90 Szidat L 1951 Cercarias schistosomicas y dermatitis schistosomica hu-mana en la Republica Argentina Commun Inst Nac Invest Cienc NatMuseo Argent Cienc Nat Bernardino Rivadavia 2129 ndash150
91 Bosq P Szidat L Soria MF 1955 Dermatitis schistosomica por Cercariachascomusi Prensa Med Argent 423500 ndash3504
92 Reyes E 1944 Dermatitis sistosomiasica en El Salvador Arch HospRosales 313ndash 8
93 Joshi SK Phil M 2002 Rice field work and the occupational hazardsOccup Med 4111ndash114
94 Tanaka M 1960 Studies on Trichobilharzia physellae in Oki Islands 2Four kinds of schistosome cercariae parasitic in Lymnaea japonica in OkiIslands Jpn J Parasitol 9604 ndash 609
95 Yang K Tchou T Trsquoang CC Ho T Luo H 1965 A study on dermatitisin rice farmers Chin Med J 84143ndash159
96 Basch PF 1966 The life cycle of Trichobilharzia brevis n sp an avianschistosome from Malaya Z Parasitenkd 27242ndash251
97 Bearup AJ Langsford WA 1966 Schistosome dermatitis in associationwith rice growing in the northern territory of Australia Med J Aust1521ndash525
98 Kumada N Ohya S Makiya K 1970 Paddy field dermatitis in AichiPrefecture II Results of the survey of fresh-water snails and parasiticcercariae in the western region of the prefecture Rep Aichi Inst PublicHealth 2065ndash73
99 Kumada N Ohya S Makiya K 1971 Paddy field dermatitis in AichiPrefecture V Eggs and adults of the genus Trichobilharzia recoveredfrom resident waterfowls in the endemic area Rep Aichi Inst PublicHealth 2139 ndash 46
100 Oda T 1973 Schistosome dermatitis in Japan Prog Med Parasitol Jpn55ndash 63
101 Suzuki N Kawanaka M 1980 Trichobilharzia brevis Basch 1966 as acause of an outbreak of cercarial dermatitis in Japan Jpn J Parasitol291ndash11
102 Makiya K Ishiguro T 1982 Population studies on Austropeplea ollula(Gould) the snail intermediate host of dermatitis-producing avian schis-tosomes Nagoya J Med Sci 4447ndash55
103 Hoque MA Skerratt LF Rahman MA Alim MA Grace D BummowB Rabiul Alam Beg ABM Debnath NC 2011 Monitoring the healthand production of household Jinding ducks on Hatia Island of Bangla-desh Trop Anim Health Prod 43431ndash 440 httpdxdoiorg101007s11250-010-9710-3
104 Fernandez MV Hamann MI Nafiez MO 2013 Larval trematodes ofBiomphalaria straminea (Mollusca Planorbidae) in ricefield in Corrien-tes Province Argentina Rev Mex Biodivers 84756 ndash764 httpdxdoiorg107550rmb33748
105 Brant SV Jouet D Ferteacute H Loker ES 2013 Anserobilharzia gen n(Digenea Schistosomatidae) and redescription of A brantae (Farr ampBlankemeyer 1956) comb n (syn Trichobilharzia brantae) a parasite ofgeese (Anseriformes) Zootaxa 3670193ndash206 httpdxdoiorg1011646zootaxa367025
106 Devkota R Brant SV Thapa A Loker ES 2014 Two avian schistosomecercariae from Nepal including a Macrobilharzia-like species from Indo-planorbis exustus Parasitol Int 63374 ndash380 httpdxdoiorg101016jparint201312009
107 Loker ES 1983 A comparative study of the life-histories of mammalianschistosomes Parasitology 87343ndash369 httpdxdoiorg101017S0031182000052689
108 Rollinson D Southgate VR 1987 The genus Schistosoma a taxonomicappraisal p 1ndash 49 In Rollinson D Simpson AJG (ed) The biology ofschistosomes from genes to latrines Academic Press United Kingdom
109 Paraense WL Pointier JP 2003 Physa acuta Draparnaud 1805 (Gas-tropoda Physidae) a study of topotypic specimens Mem Inst OswaldoCruz 98513ndash517 httpdxdoiorg101590S0074-02762003000400016
110 Taylor DW 2003 Introduction to Physidae (Gastropoda Hygrophila)
biogeography classification morphology Rev Biol Trop 51(Suppl 1)1ndash263 265ndash287
111 Kock KN Wolmarans CT 2007 Distribution and habitats of the alieninvader freshwater snail Physa acuta in South Africa Water SA (Pretoria)33717ndash722
112 Bastert J Sing A Wollenberg A Korting HC 1998 Aquarium derma-titis cercarial dermatitis in an aquarist Dermatology 19784 ndash 86 httpdxdoiorg101159000017965
113 Folster-Holst R Disko R Rowert J Bockeler W Kreiselmaier IChristophers E 2001 Cercarial dermatitis contracted via contact withan aquarium case report and review Br J Dermatol 145638 ndash 664 httpdxdoiorg101046j1365-2133200104414x
114 Brant SV Loker ES 2005 Can specialized pathogens colonize distantlyrelated hosts Schistosome evolution as a case study PLoS Pathog 1e38httpdxdoiorg101371journalppat0010038
115 Kock S 2001 Investigations of intermediate host specificity help toelucidate the taxonomic status of Trichobilharzia ocellata (DigeneaSchistosomatidae) Parasitology 12367ndash70 httpdxdoiorg101017S0031182001008101
116 Khalifa R 1976 Studies on Schistosomatidae Looss 1899 (Trematoda)of aquatic birds of Poland III Notes on the morphology and life cycle ofDendritobilharzia pulverulenta (Braun 1901) Acta Parasitol Pol 241ndash9
117 Brant SV Bochte CA Loker ES 2011 New intermediate host recordsfor the avian schistosomes Dendritobilharzia pulverulenta Gigantobilhar-zia huronensis and Trichobilharzia querquedulae from North America JParasitol 97946 ndash949 httpdxdoiorg101645GE-27431
118 Davis NE 2011 Avian schistosome research accomplished in New Zea-land from 1991 through the present Where to from here poster 17986th Meet Am Soc Parasitol Anchorage AK
119 Leite ACR Costa HMA Costa JO 1978 Trichobilharzia jequitibaensissp n (Trematoda Schistosomatidae) in Cairina moschata domestica(Anatidae) Rev Braz Biol 38843ndash 846
120 Leite ACR Costa HMA Costa JO 1979 Life cycle of Trichobilharziajequitibaensis Leite Costa and Costa 1978 (Trematoda Schistosoma-tidae) Rev Braz Biol 39341ndash345
121 Picard D Jousson O 2001 Genetic variability among cercariae of theSchistosomatidae (Trematoda Digenea) causing swimmerrsquos itch inEurope Parasite 8237ndash242 httpdxdoiorg101051parasite2001083237
122 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Molecular diversityof Trichobilharzia franki in two intermediate hosts (Radix auricularia andRadix peregra) a complex of species Infect Genet Evol 101218 ndash1227httpdxdoiorg101016jmeegid201008001
123 Rudolfovaacute J Hampl V Bayssade-Dufour C Lockyer AE LittlewoodDTJ Horaacutek P 2005 Validity reassessment of Trichobilharzia speciesusing Lymnaea stagnalis as the intermediate host Parasitol Res 9579 ndash 89httpdxdoiorg101007s00436-004-1262-x
124 Morgan JA Dejong RJ Adeoye GO Ansa ED Barbosa CS BreacutemondP Cesari IM Charbonnel N Correcirca LR Coulibaly G DrsquoAndrea PSDe Souza CP Doenhoff MJ File S Idris MA Incani RN Jarne PKaranja DM Kazibwe F Kpikpi J Lwambo NJ Mabaye A MagalhatildeesLA Makundi A Moneacute H Mouahid G Muchemi GM Mungai BNSeacutene M Southgate V Tchuenteacute LA Theacuteron A Yousif F Zanotti-Magalhatildees EM Mkoji GM Loker ES 2005 Origin and diversificationof the human parasite Schistosoma mansoni Mol Ecol 143889 ndash3902httpdxdoiorg101111j1365-294X200502709x
125 Bayssade-Dufour C Jouet D Rudolfovaacute J Horaacutek P Ferteacute H 2006Seasonal morphological variations in bird schistosomes Parasite 13205ndash214 httpdxdoiorg101051parasite2006133205
126 Chrisanfova GG Lopatkin AA Shestak AG Mishchenkov VA Zhu-kova TV Akimova LN Semyenova SK 2011 Polymorphism of thecox1 mtDNA gene from cercarial isolates of the avian schistosome Bil-harziella polonica (Trematoda Schistosomatidae) from Belarussianlakes Russ J Genet 47603ndash 609 httpdxdoiorg101134S1022795411050097
127 Rind S 1989 Dendritobilharzia pulverulenta (Trematoda Schistosoma-tidae) in New Zealand N Z J Zool 16215ndash220 httpdxdoiorg10108003014223198910422571
128 Birmani NA Dharejo AM Khan MM Shaikh AM 2013 New recordof Dendritobilharzia pulverulenta (Trematoda Schistosomatidae) fromPakistan J Anim Plant Sci 231215ndash1218
129 Kolaacuterovaacute L Rudolfovaacute J Hampl V Skirnisson K 2006 Allobilharziavisceralis gen nov sp nov (Schistosomatidae-Trematoda) from Cygnus
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 183Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

cygnus (L) (Anatidae) Parasitol Int 55179 ndash186 httpdxdoiorg101016jparint200510009
130 Brant SV 2007 The occurrence of the avian schistosome Allobilharziavisceralis Kolaacuterovaacute Rudolfovaacute Hampl et Skiacuternisson 2006 (Schistosoma-tidae) in the tundra swan Cygnus columbianus (Anatidae) from NorthAmerica Folia Parasitol 5499 ndash104 httpdxdoiorg1014411fp2007013
131 Fain A 1956 Les schistosomes drsquooiseaux du genre TrichobilharziaSkrjabin et Zakharow 1920 au Ruanda-Urundi Rev Zool Bot Afr 54147ndash178
132 Rudolfovaacute J Sitko J Horaacutek P 2006 Unusual finding of Trichobilharziasp in Motacilla alba in the Czech Republic J Helminthol 8083ndash 85 httpdxdoiorg101079JOH2005321
133 Kolaacuterovaacute L Skiacuternisson K Ferteacute H Jouet D 2013 Trichobilharzia mergisp nov (Trematoda Digenea Schistosomatidae) a visceral schistosomeof Mergus serrator (L) (Aves Anatidae) Parasitol Int 62300 ndash308 httpdxdoiorg101016jparint201303002
134 Johnson KJ Sorenson MD 1999 Phylogeny and biogeography of thedabbling ducks (genus Anas) a comparison of molecular and morpho-logical evidence Auk 116792ndash 805 httpdxdoiorg1023074089339
135 Jouet D Skiacuternisson K Kolaacuterovaacute L Ferteacute H 2010 Final hosts andvariability of Trichobilharzia regenti under natural conditions ParasitolRes 107923ndash930 httpdxdoiorg101007s00436-010-1953-4
136 Murray JA 2002 Effect of temperature on the longevity and infectivityof Trichobilharzia stagnicolae miracidia in Stagnicola emarginata p 137th Int Cong Med Appl Malacol Los Banos Laguna Philippines
137 Hertel J Holweg A Haberl B Kalbe M Haas W 2006 Snail odour-clouds spreading and contribution to the transmission success of Tricho-bilharzia ocellata (Trematoda Digenea) miracidia Oecologia 147173ndash180 httpdxdoiorg101007s00442-005-0239-5
138 Kalbe M Haberl B Haas W 1997 Miracidial host-finding in Fasciolahepatica and Trichobilharzia ocellata is stimulated by species-specific gly-coconjugates released from the host snails Parasitol Res 83806 ndash 812httpdxdoiorg101007s004360050344
139 Kalbe M Haberl B Haas W 2000 Snail host finding by Fasciola hepat-ica and Trichobilharzia ocellata compounds analysis of ldquomiracidia-attracting glycoproteinsrdquo Exp Parasitol 96231ndash242 httpdxdoiorg101006expr20004579
140 Haas W Haberl B Kalbe M Koumlrner M 1995 Snail-host-finding bymiracidia and cercariae chemical host cues Parasitology 11468 ndash 472
141 Sluiters JE Brussaard-Wust CM Meuleman EA 1980 The relationshipbetween miracidial dose production of cercariae and reproductive ac-tivity of the host in the combination Trichobilharzia ocellata and Lym-naea stagnalis Z Parasitenkd 6313ndash26 httpdxdoiorg101007BF00927722
142 Joosse J Van Elk R 1986 Trichobilharzia ocellata physiological char-acterization of giant growth glycogen depletion and absence of repro-ductive activity in the intermediate snail host Lymnaea stagnalis ExpParasitol 621ndash13 httpdxdoiorg1010160014-4894(86)90002-0
143 Amen RI Baggen JMC Bezemer PD de Jong-Brink M 1992 Modu-lation of the activity of the internal defence system of the pond snailLymnaea stagnalis by the avian schistosome Trichobilharzia ocellata Par-asitology 10433ndash 40 httpdxdoiorg101017S0031182000060777
144 Nuntildeez PE Adema CM de Jong-Brink M 1994 Modulation of thebacterial clearance activity of haemocytes from the freshwater molluscLymnaea stagnalis by the avian schistosome Trichobilharzia ocellataParasitology 109299 ndash310 httpdxdoiorg101017S0031182000078331
145 de Jong-Brink M 1995 How schistosomes profit from the stress re-sponses they elicit in their hosts Adv Parasitol 35177ndash256 httpdxdoiorg101016S0065-308X(08)60072-X
146 Horaacutek P van der Knaap WPW 1997 Lectins in snail-trematode im-mune interactions a review Folia Parasitol 44161ndash172
147 van der Knaap WPW Adema CM Sminia T 1993 Invertebrate bloodcells morphological and functional aspects of the haemocytes in thepond snail Lymnaea stagnalis Comp Haematol Int 320 ndash26 httpdxdoiorg101007BF00394923
148 Adema CM van Deutekom-Mulder EC van der Knaap WPW SminiaT 1994 Schistosomicidal activities of Lymnaea stagnalis haemocytes therole of oxygen radicals Parasitology 109479 ndash 485 httpdxdoiorg101017S0031182000080732
149 Nuntildeez PE de Jong-Brink M 1997 The suppressive excretory-secretoryproduct of Trichobilharzia ocellata a possible factor for determining
compatibility in parasite-host interactions Parasitology 115193ndash203httpdxdoiorg101017S0031182097001169
150 Plows LD Cook RT Davies AJ Walker AJ 2004 Activation of extra-cellular-signal regulated kinase is required for phagocytosis by Lymnaeastagnalis haemocytes Biochim Biophys Acta 169225ndash33 httpdxdoiorg101016S0167-4889(04)00042-4
151 Lacchini AH Davies AJ Mackintosh D Walker AJ 2006 Beta-13-glucan modulates PKC signalling in Lymnaea stagnalis defence cells arole for PKC in H2O2 production and downstream ERK activation J ExpBiol 2094829 ndash 4840 httpdxdoiorg101242jeb02561
152 Walker AJ Lacchini AH Sealey KL Mackintosh D Davies AJ 2010Spreading by snail (Lymnaea stagnalis) defence cells is regulated throughintegrated PKC FAK and Src signaling Cell Tissue Res 341131ndash145httpdxdoiorg101007s00441-010-0986-4
153 Gerhardus MJT Baggen JMC van der Knaap WPW Sminia T 1991Analysis of surface carbohydrates of Trichobilharzia ocellata miracidiaand sporocysts using lectin binding techniques Parasitology 10351ndash59httpdxdoiorg101017S003118200005928X
154 Horaacutek P 1995 Developmentally regulated expression of surface carbo-hydrate residues on larval stages of the avian schistosome Trichobilharziaszidati Folia Parasitol 42255ndash265
155 Blažovaacute K Horaacutek P 2005 Trichobilharzia regenti the developmentaldifferences in natural and abnormal hosts Parasitol Int 54167ndash172 httpdxdoiorg101016jparint200503003
156 Chanovaacute M Bulantovaacute J Maacuteslo P Horaacutek P 2009 In vitro cultivationof early schistosomula of nasal and visceral bird schistosomes (Trichobil-harzia spp Schistosomatidae) Parasitol Res 1041445ndash1452 httpdxdoiorg101007s00436-009-1343-y
157 Plows LD Cook RT Davies AJ Walker AJ 2005 Carbohydrates thatmimic schistosome surface coat components affect ERK and PKC signal-ling in Lymnaea stagnalis haemocytes Int J Parasitol 35293ndash302 httpdxdoiorg101016jijpara200411012
158 Walker AJ 2006 Do trematode parasites disrupt defence cell signallingin their snail host Trends Parasitol 22154 ndash159 httpdxdoiorg101016jpt200602003
159 Hoek RM Smit AB Frings H Vink JM de Jong-Brink M GeraertsWPM 1996 A new Ig-superfamily member molluscan defence mole-cule (MDM) from Lymnaea stagnalis is down-regulated during parasi-tosis Eur J Immunol 26939 ndash944 httpdxdoiorg101002eji1830260433
160 Smit AB de Jong-Brink M Li KW Sassen MM Spijker S Van Elk RBuijs S Van Minnen J Van Kesteren RE 2004 Granularin a novelmolluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation FASEB J 18845ndash 847 httpdxdoiorg101096fj03-0590fje
161 de Jong-Brink M Bergamin-Sassen M Solis Soto M 2001 Multiplestrategies of schistosomes to meet their requirements in the intermediatesnail host Parasitology 123S129 ndashS141
162 Geraerts WP 1992 Neurohormonal control of growth and carbohy-drate metabolism by the light green cells in Lymnaea stagnalis GenComp Endocrinol 86433ndash 444 httpdxdoiorg1010160016-6480(92)90068-U
163 Hermann PM de Lange RP Pieneman AW ter Maat A Jansen RF1997 Role of neuropeptides encoded on CDCH-1 gene in the organiza-tion of egg-laying behavior in the pond snail Lymnaea stagnalis J Neu-rophysiol 782859 ndash2869
164 de Jong-Brink M Elsaadany M Soto M 1991 The occurrence ofschistosomin an antagonist of female gonadotropic hormones is a gen-eral phenomenon in haemolymph of schistosome-infected freshwatersnails Parasitology 103371ndash378 httpdxdoiorg101017S0031182000059886
165 Schallig HDFH Sassen MJM Hordijk PL de Jong-Brink M 1991Trichobilharzia ocellata influence of infection on the fecundity of itsintermediate snail host Lymnaea stagnalis and cercarial induction of therelease of schistosomin a snail neuropeptide antagonizing female gonad-otropic hormones Parasitology 10285ndash91 httpdxdoiorg101017S0031182000060376
166 Hordijk PL de Jong-Brink M ter Maat A Pieneman AW Lodder JCKits KS 1992 The neuropeptide schistosomin and haemolymph fromparasitized snails induce similar changes in excitability in neuroendo-crine cells controlling reproduction and growth in a fresh-water snailNeurosci Lett 136193ndash197 httpdxdoiorg1010160304-3940(92)90047-B
Horaacutek et al
184 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

167 Hoek RM Li KW van Minnen J Lodder JC de Jong-Brink M SmitAB van Kesteren RE 2005 LFRFamides a novel family of parasitation-induced -RFamide neuropeptides that inhibit the activity of neuroendo-crine cells in Lymnaea stagnalis J Neurochem 921073ndash1080 httpdxdoiorg101111j1471-4159200402927x
168 McClelland G Bourns TKR 1969 Effects of Trichobilharzia ocellata ongrowth reproduction and survival of Lymnaea stagnalis Exp Parasitol24137ndash146 httpdxdoiorg1010160014-4894(69)90150-7
169 Zbikowska E 2005 Do larvae of Trichobilharzia szidati and Echinostomarevolutum generate behavioral fever in Lymnaea stagnalis individualsParasitol Res 9768 ndash72 httpdxdoiorg101007s00436-005-1394-7
170 Zbikowska E Cichy A 2012 Symptoms of behavioural anapyrexiamdashreverse fever as a defence response of snails to fluke invasion J InvertebrPathol 109269 ndash273 httpdxdoiorg101016jjip201112006
171 Stirewalt MA 1974 Schistosoma mansoni cercaria to schistosomuleAdv Parasitol 12115ndash182 httpdxdoiorg101016S0065-308X(08)60388-7
172 Haas W 1992 Physiological analysis of cercarial behavior J Parasitol78243ndash255 httpdxdoiorg1023073283471
173 Feiler W Haas W 1988 Host-finding in Trichobilharzia ocellata cercar-iaemdashswimming and attachment to the host Parasitology 96493ndash505httpdxdoiorg101017S0031182000080136
174 Sopott-Ehlers B Haas W Ehlers U 2003 Ultrastructure of pigmentedand unpigmented photoreceptors in cercariae of Trichobilharzia ocellata(Plathelminthes Trematoda Schistosomatidae) evidence for the evolu-tion of parasitism in Neodermata Parasitol Res 91109 ndash116 httpdxdoiorg101007s00436-003-0933-3
175 Feiler W Haas W 1988 Trichobilharzia ocellatamdash chemical stimuli ofduck skin for cercarial attachment Parasitology 96507ndash517 httpdxdoiorg101017S0031182000080148
176 Haas W 2003 Parasitic worms strategies of host finding recognitionand invasion Zoology 106349 ndash364 httpdxdoiorg1010780944-2006-00125
177 Haas W Haumlberlein S 2009 Penetration of cercariae into the livinghuman skin Schistosoma mansoni vs Trichobilharzia szidati ParasitolRes 1051061ndash1066 httpdxdoiorg101007s00436-009-1516-8
178 Haas W van de Roemer A 1998 Invasion of the vertebrate skin bycercariae of Trichobilharzia ocellata penetration processes and stimulat-ing host signals Parasitol Res 84787ndash795 httpdxdoiorg101007s004360050489
179 Bulantovaacute J Chanovaacute M Houžvickovaacute L Horaacutek P 2011 Trichobil-harzia regenti (Digenea Schistosomatidae) changes of body wall muscu-lature during the development from miracidium to adult worm Micron4247ndash54 httpdxdoiorg101016jmicron201008003
180 Mikeš L Ziacutedkovaacute L Kašnyacute M Dvoraacutek J Horaacutek P 2005 In vitrostimulation of penetration gland emptying by Trichobilharzia szidati andT regenti (Schistosomatidae) cercariae Quantitative collection and par-tial characterization of the products Parasitol Res 96230 ndash241 httpdxdoiorg101007s00436-005-1347-1
181 Wulff C Haumlberlein S Haas W 2007 Cream formulations protectingagainst cercarial dermatitis by Trichobilharzia Parasitol Res 10191ndash97httpdxdoiorg101007s00436-006-0431-5
182 Ligasovaacute A Bulantovaacute J Šebesta O Kašnyacute M Koberna K Mikeš L2011 Secretory glands in cercaria of the neuropathogenic schistosomeTrichobilharzia regentimdashultrastructural characterization 3-D model-ling volume and pH estimations Parasit Vectors 4162 httpdxdoiorg1011861756-3305-4-162
183 Bahgat M Francklow K Doenhoff MJ Li YL Ramzy RM Kirsten CRuppel A 2001 Infection induces antibodies against the cercarial secre-tions but not against the cercarial elastases of Schistosoma mansoniSchistosoma haematobium Schistosoma japonicum and Trichobilharziaocellata Parasite Immunol 23557ndash565 httpdxdoiorg101046j1365-3024200100417x
184 Bahgat M Ruppel A 2002 Biochemical comparison of the serine pro-tease (elastase) activities in cercarial secretions from Trichobilharzia ocel-lata and Schistosoma mansoni Parasitol Res 88495ndash500 httpdxdoiorg101007s00436-002-0597-4
185 Kašnyacute M Mikeš L Dalton JP Mountford AP Horaacutek P 2007 Com-parison of cysteine peptidase activities in Trichobilharzia regenti andSchistosoma mansoni cercariae Parasitology 1341599 ndash1609 httpdxdoiorg101017S0031182007002910
186 Doleckovaacute K Kašnyacute M Mikeš L Mutapi F Stack C Mountford APHoraacutek P 2007 Peptidases of Trichobilharzia regenti (Schistosomatidae)
and its molluscan host Radix peregra s lat (Lymnaeidae) constructionand screening of cDNA library from intramolluscan stages of the para-site Folia Parasitol 5494 ndash98 httpdxdoiorg1014411fp2007012
187 Doleckovaacute K Kašnyacute M Mikeš L Cartwright J Jedelskyacute P SchneiderEL Dvoraacutek J Mountford AP Craik CS Horaacutek P 2009 The functionalexpression and characterisation of a cysteine peptidase from the invasivestage of the neuropathogenic schistosome Trichobilharzia regenti Int JParasitol 39201ndash211 httpdxdoiorg101016jijpara200806010
188 Dvoraacutek J Mashiyama ST Braschi S Sajid M Knudsen GM Hansell ELim KC Hsieh I Bahgat M Mackenzie B Medzihradszky KF BabbittPC Caffrey CR McKerrow JH 2008 Differential use of protease fam-ilies for invasion by schistosome cercariae Biochimie 90345ndash358 httpdxdoiorg101016jbiochi200708013
189 Doleckovaacute K Albrecht T Mikeš L Horaacutek P 2010 Cathepsins B1 and B2in the neuropathogenic schistosome Trichobilharzia regenti distinct geneexpression profiles and presumptive roles throughout the life cycle Para-sitol Res 107751ndash755 httpdxdoiorg101007s00436-010-1943-6
190 Pleass RJ Aboobaker AA Schmitt AO 2008 Piggybacking schistosomeinvasion similarities are only skin deep Trends Parasitol 24153ndash156httpdxdoiorg101016jpt200801003
191 Horaacutek P Grubhoffer L Mikeš L Tichaacute M 1997 Lectins of Trichobil-harzia szidati cercariae Parasite 427ndash35 httpdxdoiorg101051parasite1997041027
192 McKerrow JH Pinoheiss S Lindquist R Werb Z 1985 Purificationand characterization of an elastinolytic proteinase secreted by cercariaeof Schistosoma mansoni J Biol Chem 2603703ndash3707
193 Modha J Redman CA Thornhill JA Kusel JR 1998 Schistosomesunanswered questions on the basic biology of the host-parasite relation-ship Parasitol Today 14396 ndash 401 httpdxdoiorg101016S0169-4758(98)01321-0
194 Linder E 1986 Fluorochrome-labelled lectins reveal secreted glycocon-jugates of schistosome larvae Parasitol Today 2219 ndash221 httpdxdoiorg1010160169-4758(86)90085-2
195 Nevhutalu PA Salafsky B Haas W Conway T 1993 Schistosomamansoni and Trichobilharzia ocellatamdash comparison of secreted cercarialeikosanoids J Parasitol 79130 ndash133
196 Horaacutek P Mikeš L 1995 Cercarial surface saccharides of six trematodespecies from the pond snail Lymnaea stagnalis Parasite 2419 ndash 421
197 Hokke CH Neeleman AP Koeleman CAM van den Eijnden DH1998 Identification of an alpha 3-fucosyltransferase and a novel alpha2-fucosyltransferase activity in cercariae of the schistosome Trichobilhar-zia ocellata biosynthesis of the Fuc alpha 1 iexcl 2Fuc alpha 1 iexcl 3[Gal-(NAc)beta 1 iexcl 4]GlcNAc sequence Glycobiology 8393ndash 406 httpdxdoiorg101093glycob84393
198 Podhorskyacute M Huzovaacute Z Mikeš L Horaacutek P 2009 Cercarial dimen-sions and surface structures as a tool for species determination of Tricho-bilharzia spp Acta Parasitol 5428 ndash36 httpdxdoiorg102478s11686-009-0011-9
199 Horaacutek P Kovaacuter L Kolaacuterovaacute L Nebesaacuterovaacute J 1998 Cercaria-schistosomulum surface transformation of Trichobilharzia szidati and itsputative immunological impact Parasitology 116139 ndash147 httpdxdoiorg101017S0031182097002059
200 Chanovaacute M Lichtenbergovaacute L Bulantovaacute J Mikeš L Horaacutek P 2012Trichobilharzia regenti antigenic structures of intravertebrate stagesCent Eur J Biol 783ndash90 httpdxdoiorg102478s11535-011-0074-0
201 Boulanger D Schneider D Sidikou F Capron A Chippaux J-P SellinB 1999 The oral route as a potential way of transmission of Schistosomabovis in goats J Parasitol 85464 ndash 467 httpdxdoiorg1023073285779
202 Oleaga A Ramajo V 2004 Efficiency of the oral intramuscular andsubcutaneous routes for the experimental infection of hamster and sheepwith Schistosoma bovis Vet Parasitol 12443ndash53 httpdxdoiorg101016jvetpar200406022
203 Giver H Johansen MV Christensen NO Boslashgh H Nansen P 1999Peroral infection of pigs with Schistosoma japonicum cercariae Vet Para-sitol 83161ndash165 httpdxdoiorg101016S0304-4017(99)00048-5
204 Islam KS Copeman DB 1986 The morphology and life cycle of Tricho-bilharzia parocellata (Johnston amp Simpson 1939) Islam amp Copeman1980 from the visceral blood vessels of Australian anatids Syst Parasitol839 ndash 49 httpdxdoiorg101007BF00010308
205 Horaacutek P Kolaacuterovaacute L Dvoraacutek J 1998 Trichobilharzia regenti n sp(Schistosomatidae Bilharziellinae) a new nasal schistosome from
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 185Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

Europe Parasite 5349 ndash357 httpdxdoiorg101051parasite1998054349
206 Bourns TKR Ellis JC Rau ME 1973 Migration and development ofTrichobilharzia ocellata (Trematoda Schistosomatidae) in its duck hostsCan J Zool 511021ndash1030 httpdxdoiorg101139z73-148
207 Horaacutek P Dvoraacutek J Kolaacuterovaacute L Trefil L 1999 Trichobilharzia regentia pathogen of the avian and mammalian central nervous system Parasi-tology 119577ndash581 httpdxdoiorg101017S0031182099005132
208 Grabe K Haas W 2004 Navigation within host tissues Schistosomamansoni and Trichobilharzia ocellata schistosomula respond to chemicalgradients Int J Parasitol 34927ndash934 httpdxdoiorg101016jijpara200404012
209 Grabe K Haas W 2004 Navigation within host tissues cercariae ori-entate towards dark after penetration Parasitol Res 93111ndash113 httpdxdoiorg101007s00436-003-1045-9
210 Chanovaacute M Vuong S Horaacutek P 2007 Trichobilharzia szidati the lungphase of migration within avian and mammalian hosts Parasitol Res1001243ndash1247 httpdxdoiorg101007s00436-006-0398-2
211 Neuhaus W 1952 Biologie und Entwicklung von Trichobilharzia szidatin sp (Trematoda Schistosomatidae) einem Erreger von Dermatitis beiMenschen Z Parasitenkd 15203ndash266
212 Vusse FJV 1979 Host-parasite relations of Dendritobilharzia pulveru-lenta (Trematoda Schistosomatidae) and anatids J Parasitol 65894 ndash897 httpdxdoiorg1023073280244
213 Olivier L 1953 Observations on the migrations of avian schistosomes inmammals previously unexposed to cercariae J Parasitol 39237ndash243httpdxdoiorg1023073273943
214 Hraacutedkovaacute K Horaacutek P 2002 Neurotropic behaviour of Trichobilharziaregenti in ducks and mice J Helminthol 76137ndash141 httpdxdoiorg101079JOH2002113
215 Lichtenbergovaacute L Lassmann H Jones MK Kolaacuterovaacute L Horaacutek P 2011Trichobilharzia regenti host immune response in the pathogenesis ofneuroinfection in mice Exp Parasitol 128328 ndash335 httpdxdoiorg101016jexppara201104006
216 van Bolhuis GH Rijks JM Dorrestein GM Rudolfovaacute J van Dijk MKuiken T 2004 Obliterative endophlebitis in mute swans (Cygnus olor)caused by Trichobilharzia sp (Digenea Schistosomatidae) infection VetPathol 41658 ndash 665 httpdxdoiorg101354vp41-6-658
217 Wood LM Bacha WJ Jr 1983 Distribution of eggs and the host re-sponse in chickens infected with Austrobilharzia variglandis (Trema-toda) J Parasitol 69682ndash 688 httpdxdoiorg1023073281141
218 Wojcinski ZW Barker IK Hunter DB Lumsden H 1987 An outbreakof schistosomiasis in Atlantic brant geese Branta bernicla hrota J WildlDis 23248 ndash255 httpdxdoiorg1075890090-3558-232248
219 Pence DB Rhodes MJ 1982 Trichobilharzia physellae (Digenea Schis-tosomatidae) from endemic waterfowl on the high plains of Texas JWildl Dis 1869 ndash74 httpdxdoiorg1075890090-3558-18169
220 Kolaacuterovaacute L Horaacutek P Cada F 2001 Histopathology of CNS and nasalinfections caused by Trichobilharzia regenti in vertebrates Parasitol Res87644 ndash 650 httpdxdoiorg101007s004360100431
221 Chanovaacute M Horaacutek P 2007 Terminal phase of bird schistosomiasiscaused by Trichobilharzia regenti (Schistosomatidae) in ducks (Anasplatyrhynchos f domestica) Folia Parasitol 54105ndash107 httpdxdoiorg1014411fp2007014
222 Levine ND Clark DT Hanson LE 1956 Encephalitis in a swan due toDendritobilharzia sp (Trematoda Schistosomatidae) J Parasitol 42496 ndash500 httpdxdoiorg1023073274443
223 Wilson RB New JC Scholtens RG 1982 Granulomatous encephalitiscaused by schistosomiasis in swans J Am Vet Med Assoc 1811386 ndash1387
224 Kourilovaacute P Syrucek M Kolaacuterovaacute L 2004 The severity of mousepathologies caused by the bird schistosome Trichobilharzia regenti inrelation to host immune status Parasitol Res 938 ndash16 httpdxdoiorg101007s00436-004-1079-7
225 Haemmerli U 1953 Schistosomen-Dermatitis am Zuumlrichsee Dermato-logica 107301ndash341
226 Tremaine AM Whittemore DE Gewirtzman AJ Bartlett BL MendozaN Rapini RP Tyring SK 2009 An unusual case of swimmerrsquos itch J AmAcad Dermatol 60174 ndash176 httpdxdoiorg101016jjaad200807060
227 Kourilovaacute P Hogg KG Kolaacuterovaacute L Mountford AP 2004 Cercarialdermatitis caused by bird schistosomes comprises both immediate andlate phase cutaneous hypersensitivity reactions J Immunol 1723766 ndash3774 httpdxdoiorg104049jimmunol17263766
228 Larkin BM Smith PM Ponichtera HE Shainheit MG Rutitzky LIStadecker MJ 2012 Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis Semin Immunopathol 34873ndash 888 httpdxdoiorg101007s00281-012-0341-9
229 Jutel M Watanabe T Klunker S Akdis M Thomet OA Malolepszy JZak-Nejmark T Koga R Kobayashi T Blaser K Akdis CA 2001Histamine regulates T-cell and antibody responses by differential expres-sion of H1 and H2 receptors Nature 413420 ndash 425 httpdxdoiorg10103835096564
230 Lichtenbergovaacute L Kolbekovaacute P Kourilovaacute P Kašnyacute M Mikeš L HaasH Schramm G Horaacutek P Kolaacuterovaacute L Mountford AP 2008 Antibodyresponses induced by Trichobilharzia regenti antigens in murine and hu-man hosts exhibiting cercarial dermatitis Parasite Immunol 30585ndash595httpdxdoiorg101111j1365-3024200801059x
231 Bacha W Jr Roush R Jr Icardi S 1982 Infection of the gerbil by theavian schistosome Austrobilharzia variglandis (Miller and Northup 1926Pener 1953) J Parasitol 68505ndash507 httpdxdoiorg1023073280970
232 Horaacutek P Kolaacuterovaacute L 2001 Bird schistosomesmdash do they die in mam-malian skin Trends Parasitol 1766 ndash 69 httpdxdoiorg101016S1471-4922(00)01770-0
233 Melo FL Gomes ALD Barbosa CS Werkhauser RP Abath FGC 2006Development of molecular approaches for the identification of transmis-sion sites of schistosomiasis Trans R Soc Trop Med Hyg 1001049 ndash1055httpdxdoiorg101016jtrstmh200512008
234 Kane RA Stothard JR Rollinson D Leclipteux T Evraerts J StandleyCJ Allan F Betson M Kaba R Mertens P Laurent T 2013 Detectionand quantification of schistosome DNA in freshwater snails using eitherfluorescent probes in real-time PCR or oligochromatographic dipstickassays targeting the ribosomal intergenic spacer Acta Trop 128241ndash249httpdxdoiorg101016jactatropica201110019
235 Worrell C Xiao N Vidal JE Chen L Zhong B Remais J 2011 Fielddetection of Schistosoma japonicum cercariae in environmental watersample by quantitative PCR Appl Environ Microbiol 772192ndash2195httpdxdoiorg101128AEM01561-10
236 Onichandran S Kumar T Lim YAL Sawangjaroen N Andiappan HSalibay CC Chye TT Ithoi I Dungca JZ Sulaiman WYW Ling LYNissapatorn V 2013 Waterborne parasites and physico-chemical as-sessment of selected lakes in Malaysia Parasitol Res 1124185ndash 4191httpdxdoiorg101007s00436-013-3610-1
237 Schets FM Lodder WJ Husman AMD 2010 Confirmation of thepresence of Trichobilharzia by examination of water samples and snailsfollowing reports of cases of cercarial dermatitis Parasitology 13777ndash 83httpdxdoiorg101017S0031182009990849
238 Horaacutek P Schets FM Kolaacuterovaacute L Brant SV 2013 Trichobilharzia p455ndash 465 In Liu D (ed) Molecular detection of human parasitic patho-gens Taylor and Francis CRC Press Boca Raton FL
239 Hertel J Hamburger J Haberl B Haas W 2002 Detection of birdschistosomes in lakes by PCR and filter-hybridization Exp Parasitol 10157ndash 63 httpdxdoiorg101016S0014-4894(02)00036-X
240 Graczyk TK Shiff CJ 2000 Recovery of avian schistosome cercariaefrom water using penetration stimulant matrix with an unsaturated fattyacid Am J Trop Med Hyg 63174 ndash177
241 Cringoli G Rinaldi L Utzinger J 2010 FLOTAC new multivalenttechniques for qualitative and quantitative copromicroscope diagnosisof parasites in animals and humans Nat Protoc 5503ndash515 httpdxdoiorg101038nprot2009235
242 Katz N Chavez A Pellegrino J 1972 A simple device for quantitativestool thicksmear technique in schistosomiasis mansoni Rev Inst MedTrop Sao Paulo 14397ndash 400
243 Kolaacuterovaacute L Horaacutek P Skiacuternisson K 2010 Methodical approaches in theidentification of areas with potential risk of infection by bird schisto-somes causing cercarial dermatitis J Helminthol 84327ndash335 httpdxdoiorg101017S0022149X09990721
244 Skiacuternisson K Kolaacuterovaacute L 2008 Diversity of bird schistosomes in an-seriform birds in Iceland based on egg measurements and egg morphol-ogy Parasitol Res 10343ndash50 httpdxdoiorg101007s00436-008-0925-4
245 Lu DB Wang TP Rudge JW Donnelly CA Fang GR Webster JP2009 Evolution in a multi-host parasite chronobiological circadianrhythm and population genetics of Schistosoma japonicum cercariae in-dicates contrasting definitive host reservoirs by habitat Int J Parasitol391581ndash1588 httpdxdoiorg101016jijpara200906003
246 Nolan MJ Cribb TJ 2005 The use and implications of ribosomal DNA
Horaacutek et al
186 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

sequencing for the discrimination of digenean species Adv Parasitol 60101ndash163 httpdxdoiorg101016S0065-308X(05)60002-4
247 Vilas R Criscione C Blouin M 2005 A comparison between mito-chondrial DNA and the ribosomal internal transcribed regions in pros-pecting for cryptic species of platyhelminth parasites Parasitology 131839 ndash 846 httpdxdoiorg101017S0031182005008437
248 Dvoraacutek J Vanaacutecovaacute S Hampl V Flegr J Horaacutek P 2002 Comparisonof European Trichobilharzia species based on ITS1 and ITS2 sequencesParasitology 124307ndash313 httpdxdoiorg101017S0031182001001238
249 Chrisanfova GG Lopatkin AA Mishchenkov VA Kheidorova EEDorozhenkova TE Zhukova TV Ryskov AP Semyenova SK 2009Genetic variability of bird schistosomes (class Trematoda family Schis-tosomatidae) of Naroch Lake identification of a new species in theTrichobilharzia ocellata group Dokl Biochem Biophys 428268 ndash272httpdxdoiorg101134S1607672909050123
250 Jouet D Ferteacute H Depaquit J Rudolfovaacute J Latour P Zanella DKaltenbach ML Leacuteger N 2008 Trichobilharzia spp in natural condi-tions in Annecy Lake France Parasitol Res 10351ndash58 httpdxdoiorg101007s00436-008-0926-3
251 Rizevsky SV Cherviakovsky EM Kurchenko VP 2011 Moleculartaxonomic identification of Schistosomatidae from Naroch Lake andPolonevichi Lake in Belarus Biochem Syst Ecol 3914 ndash21 httpdxdoiorg101016jbse201012014
252 Ferteacute H Depaquit J Carreacute S Villena I Leacuteger N 2005 Presence ofTrichobilharzia szidati in Lymnaea stagnalis and T franki in Radix auric-ularia in northeastern France molecular evidence Parasitol Res 95150 ndash154 httpdxdoiorg101007s00436-004-1273-7
253 Semyenova SK Chrisanfova GG Filippova EK Beeacuter SA Voronin MVRyskov AP 2005 Individual and population variation in cercariae ofbird schistosomes of the Trichobilharzia ocellata species group as revealedwith the polymerase chain reaction Genetika 4112ndash16
254 Korsunenko A Chrisanfova G Lopatkin A Beer SA Voronin MRyskov AP Semyenova SK 2012 Genetic differentiation of cercariaeinfrapopulations of the avian schistosome Trichobilharzia szidati basedon RAPD markers and mitochondrial cox1 gene Parasitol Res 110833ndash841 httpdxdoiorg101007s00436-011-2562-6
255 Besansky NJ Severson DW Ferdig MT 2003 DNA barcoding ofparasites and invertebrate disease vectors what you donrsquot know can hurtyou Trends Parasitol 19545ndash546 httpdxdoiorg101016jpt200309015
256 Hansen H Bakke TA Bachmann L 2007 DNA taxonomy and barcod-ing of monogenean parasites lessons from Gyrodactylus Trends Parasi-tol 23363ndash367 httpdxdoiorg101016jpt200706007
257 Peterson AT Moyle RG Nyaacuteri AS Robbins MB Brumfield RTRemsen JV Jr 2007 The need for proper vouchering in phylogeneticstudies of birds Mol Phylogenet Evol 451042ndash 4044 httpdxdoiorg101016jympev200708019
258 Olsson U Sundberg P Alstroumlm P Gelang M Ericson PGP 2008 Whatis the proper vouchering in phylogenetic studies of birdsmdasha reply toPeterson et al (2007) Mol Phylogenet Evol 48383ndash385 httpdxdoiorg101016jympev200803031
259 Hoberg EP Pilitt PA Galbreath KE 2009 Why museums matter a taleof pinworms (Oxyuroidea Heteroxynematidae) among pikas (Ochotonaprinceps and O collaris) in the American West J Parasitol 95490 ndash501httpdxdoiorg101645GE-18231
260 Olivier L 1949 Schistosome dermatitis a sensitization phenomenonAm J Hyg (Lond) 49290 ndash302
261 Augustine DL Weller TH 1949 Experimental studies on the specificityof skin tests for the diagnosis of schistosomiasis J Parasitol 35461ndash 466httpdxdoiorg1023073273648
262 Ulrich H Landthaler M Vogt T 2008 Aquatic dermatoses J DtschDermatol Ges 6133ndash146 httpdxdoiorg101111j1610-0387200706590x
263 Kimmig P Meier M 1985 Parasitologische Untersuchungen Diagnoseund Klinik der Zerkariendermatitis-hygienische Bedeutung fuumlr Ba-degewaumlsser gemaumlssigter Zonen Zentralbl Bakteriol Hyg I Abt Orig B181390 ndash 408
264 Kourilovaacute P Kolaacuterovaacute L 2002 Variations in immunofluorescent anti-body response against Trichobilharzia and Schistosoma antigens in com-patible and incompatible hosts Parasitol Res 88513ndash521 httpdxdoiorg101007s00436-002-0607-6
265 Agrawal MC Gupta S George J 2000 Cercarial dermatitis in IndiaBull World Health Organ 78278
266 Fraser SJ Allan SJR Roworth M Smith HV Holme SA 2009 Cer-carial dermatitis in the UK Clin Exp Dermatol 34344 ndash346 httpdxdoiorg101111j1365-2230200802903x
267 Salafsky B Ramaswamy K He YX Li J Shibuya T 1999 Developmentand evaluation of LIPODEET a new long-acting formulation of NN-diethyl-m-toluamide (DEET) for the prevention of schistosomiasis AmJ Trop Med Hyg 61743ndash750
268 Marcogliese DJ 2001 Implications of climate change for parasitism ofanimals in the aquatic environment Can J Zool 791331ndash1352 httpdxdoiorg101139z01-067
269 Cotton PA 2003 Avian migration phenology and global climate changeProc Natl Acad Sci U S A 10012219 ndash12222 httpdxdoiorg101073pnas1930548100
270 Suter W 1994 Overwintering waterfowl on Swiss lakes how are abun-dance and species richness influenced by trophic status and lake mor-phology Hydrobiologia 2792801ndash14
271 Morley NJ 2012 Cercariae (Platyhelminthes Trematoda) as neglectedcomponents of zooplankton communities in freshwater habitats Hy-drobiologia 17ndash19 httpdxdoiorg101007s10750-012-1029-9
272 Barber KE Caira JN 1995 Investigation of the life cycle and adultmorphology of the avian blood fluke Austrobilharzia variglandis (Trem-atoda Schistosomatidae) from Connecticut J Parasitol 81584 ndash592httpdxdoiorg1023073283857
273 Davis NE 1998 Population dynamics of and larval trematode interac-tions with Lymnaea tomentosa and the potential for biological control ofschistosome dermatitis in Bremner Bay Lake Wanaka New ZealandJ Helminthol 72319 ndash324 httpdxdoiorg101017S0022149X00016679
274 Sluiters JE 1981 Development of Trichobilharzia ocellata in Lymnaeastagnalis and the effects of infection on the reproductive system of thehost Z Parasitenkd 64303ndash319
275 Poulin R 2006 Global warming and temperature-mediated increases incercarial emergence in trematode parasites Parasitology 132143ndash151httpdxdoiorg101017S0031182005008693
276 Larsen AH Bresciani J Buchmann K 2004 Increasing frequency ofcercarial dermatitis at higher latitudes Acta Parasitol 49217ndash221
277 Islam KS 1986 The morphology and life-cycle of Trichobilharzia arcu-ata n sp (Schistosomatidae Bilharziellinae) a nasal schistosome of waterwhistle ducks (Dendrocygna arcuata) in Australia Syst Parasitol 8117ndash128 httpdxdoiorg101007BF00009868
278 Allgoumlwer R 1995 Zerkariendermatitismdash ein Warnsignal fuumlr zuneh-mende Gewaumlssereutrophierung Limnol Aktuell 7193ndash203
279 de Gentile L Picot H Bourdeau P Bardet R Kerjan A Piriou M LeGuennic A Bayssade-Dufour C Chabasse D Mott KE 1996 Ledermatite cercarienne en Europe un probleme de la santeacute publique nou-veau Bull World Health Organ 74159 ndash163
280 Lindblade KA 1998 The epidemiology of cercarial dermatitis and itsassociation with limnological characteristics of a Northern MichiganLake J Parasitol 8419 ndash23 httpdxdoiorg1023073284521
281 Chase JM 2003 Strong and weak trophic cascades along a productivitygradient Oikos 101187ndash195 httpdxdoiorg101034j1600-0706200312062x
282 McKenzie VJ Townsend AR 2007 Parasitic and infectious diseaseresponses to changing global nutrient cycles EcoHealth 4384 ndash396 httpdxdoiorg101007s10393-007-0131-3
283 Marcogliese DJ 2008 The impact of climate change on the parasites andinfectious diseases of aquatic animals Rev Sci Tech 27467ndash 484
284 Soldaacutenovaacute M Kostadinova A 2011 Rapid colonisation of Lymnaeastagnalis by larval trematodes in eutrophic ponds in Central Europe IntJ Parasitol 41981ndash990 httpdxdoiorg101016jijpara201105005
285 Curtis LA 2002 Ecology of larval trematodes in three marine gastro-pods Parasitology 124S43ndashS56 httpdxdoiorg101017S0031182002001452
286 Muumlller V Kimmig E 1994 Trichobilharzia franki n spmdash die Ursachefuumlr Badedermatitiden in suumldwestdeutschen Baggerseen Appl Parasitol3512ndash31
287 Vaumlyrynen T Siddall R Valtonen ET Taskinen J 2000 Patterns oftrematode parasitism in lymnaeid snails from northern and central Fin-land Ann Zool Fenn 37189 ndash199
288 Soldaacutenovaacute M Faltyacutenkovaacute A Scholz T Kostadinova A 2011 Parasitesin a man-made landscape contrasting patterns of trematode flow in a
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 187Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

fishpond area in Central Europe Parasitology 138789 ndash 807 httpdxdoiorg101017S0031182011000291
289 Graham AL 2003 Effects of snail size and age on the prevalence andintensity of avian schistosome infection relating laboratory to fieldstudies J Parasitol 89458 ndash 463 httpdxdoiorg1016450022-3395(2003)089[0458EOSSAA]20CO2
290 Coady NR Muzzall PM Burton TM Snider RJ Saxton J Sergeant MSommers A 2006 Ubiquitous variability in the prevalence of Trichobil-harzia stagnicolae (Schistosomatidae) infecting Stagnicola emarginata inthree northern Michigan lakes J Parasitol 9210 ndash15 httpdxdoiorg101645GE-33361
291 Hurley M Hearnden MN Blair D Kay BH 1994 Larval trematodes infreshwater snails at the Ross River reservoir northern Australia withemphasis on Trichobilharzia sp(p) causative agents of swimmerrsquos itchAust J Mar Freshw Res 45563ndash567 httpdxdoiorg101071MF9940563
292 Patz JA Graczyk TK Gelle N Vittor AY 2000 Effect of environmentalchange on emerging parasitic diseases Int J Parasitol 301395ndash1405 httpdxdoiorg101016S0020-7519(00)00141-7
293 Mas-Coma S Valero AV Bargues MD 2009 Climate change effects ontrematodiases with emphasis on zoonotic fascioliasis and schistosomia-sis Vet Parasitol 163264 ndash280 httpdxdoiorg101016jvetpar200903024
294 Lafferty KD 1997 Environmental parasitology what can parasites tellus about human impacts on the environment Parasitol Today 13251ndash255 httpdxdoiorg101016S0169-4758(97)01072-7
295 Pietrock M Marcogliese DJ 2003 Free-living endohelminth stages atthe mercy of environmental conditions Trends Parasitol 19293ndash299httpdxdoiorg101016S1471-4922(03)00117-X
296 Sures B 2008 Environmental parasitology interactions between para-sites and pollutants in the aquatic environment Parasite 15434 ndash 438httpdxdoiorg101051parasite2008153434
297 Vidal-Martiacutenez VM Pech D Sures B 2010 Can parasites really revealenvironmental impact Trends Parasitol 2644 ndash51 httpdxdoiorg101016jpt200911001
298 Poulin R Paterson RA Townsend C Tompkins DM Kelly DW 2011Biological invasions and the dynamics of endemic diseases in freshwaterecosystems Freshw Biol 56676 ndash 688 httpdxdoiorg101111j1365-2427201002425x
299 Johnson PTJ Dobson A Lafferty KD Marcogliese DJ Memmott JOrlofske SA Poulin R Thieltges DW 2010 When parasites becomeprey ecological and epidemiological significance of eating parasitesTrends Ecol Evol 35362ndash371 httpdxdoiorg101016jtree201001005
300 Thieltges DW Jensen KT Poulin R 2008 The role of biotic factors inthe transmission of free-living endohelminth stages Parasitology 135407ndash 426 httpdxdoiorg101017S0031182007000248
301 Lim HK Heyneman D 1972 Intramolluscan inter-trematode antago-nism a review of factors influencing the host-parasite system and itspossible role in biological control Adv Parasitol 10191ndash268 httpdxdoiorg101016S0065-308X(08)60175-X
302 Kuris AM 1990 Guild structure of larval trematodes in molluscan hostsprevalence dominance and significance of competition p 69 ndash100 InEsch GW Bush AO Aho JM (ed) Parasite communities patterns andprocesses Chapman amp Hall London United Kingdom
303 Sousa WP 1992 Interspecific interactions among larval trematode par-asites of freshwater and marine snails Am Zool 32583ndash592
304 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Pro-spective study of swimmers itch incidence and severity J Parasitol 90697ndash704 httpdxdoiorg101645GE-237R
305 Verbrugge LM Rainey JJ Reimink RL Blankespoor HD 2004 Swim-merrsquos itch incidence and risk factors Am J Public Health 94738 ndash741httpdxdoiorg102105AJPH945738
306 Schets FM Lodder WJ van Duynhoven YTHP Husman AMD 2008Cercarial dermatitis in the Netherlands caused by Trichobilharzia spp JWater Health 6187ndash195 httpdxdoiorg102166wh2008028
307 Houmlrweg C Sattman H Auer H 2006 Cercarial dermatitis in Austriaquestionnaires as useful tools to estimate risk factors Middle Eur J Med11877ndash 80 httpdxdoiorg101007s00508-006-0674-2
308 Appleton CC Lethbridge RC 1979 Schistosome dermatitis in the swanestuary western Australia Med J Aust 131141ndash144
309 Leacutevesque B Giovenazzo P Guerrier P Laverdieacutere D PrudrsquoHomme H
2002 Investigation of an outbreak of cercarial dermatitis Epidemiol In-fect 129379 ndash386 httpdxdoiorg101017S0950268802007379
310 Berg K Reiter HEH 1960 Observations on schistosome dermatitis inDenmark Acta Derm Venereol 40369 ndash380
311 Kolaacuterovaacute L Horaacutek P Sitko J 1997 Cercarial dermatitis in focusschistosomes in the Czech Republic Helminthologia 34127ndash139
312 Thorp JH Covich AP 2001 Ecology and classification of NorthAmerican freshwater invertebrates 2nd ed p 1056 Academic PressNew York NY
313 Dillon RT 2004 The ecology of freshwater molluscs p 509 CambridgeUniversity Press Cambridge United Kingdom
314 Allgoumlwer R Effelsberg W 1991 Swimmerrsquos dermatitis in an excavationpoolmdashan incentive for the status analysis of the water and for the prep-aration of an ecology-friendly utilization concept Offentl Gesund-heitswes 53138 ndash143
315 Kagan IG Short RB Nez MM 1954 Maintenance of Schistosomatiumdouthitti (Cort 1914) in the laboratory (Trematoda Schistosomatidae)J Parasitol 40424 ndash 439 httpdxdoiorg1023073273890
316 Macy RW Moore DJ Price WS Jr 1955 Studies on dermatitis-producing schistosomes in the Pacific Northwest with special referenceto Trichobilharzia oregonensis Trans Am Microsc Soc 74235ndash251 httpdxdoiorg1023073224097
317 Anderson PA Nowosielsky LW Croll NA 1976 Emergence of cercar-iae of Trichobilharzia ocellata and its relationship to activity of its snailhost Lymnaea stagnalis Can J Zool 541481ndash1487 httpdxdoiorg101139z76-171
318 Olivier L 1951 The influence of light on the emergence of Schistosoma-tium douthitti cercariae from their snail host J Parasitol 37201ndash204httpdxdoiorg1023073273454
319 Blankespoor HD Reimink RL 1991 The control of swimmerrsquos itch inMichigan past present and future Mich Acad 247ndash23
320 Reimink RL DeGoede JA Blankespoor HD 1995 Efficacy of praziqu-antel in natural populations of mallards infected with avian schisto-somes J Parasitol 811027ndash1029 httpdxdoiorg1023073284067
321 Blankespoor CL Reimink RL Blankespoor HD 2001 Efficacy of prazi-quantel in treating natural schistosome infections in common mergan-sers J Parasitol 87424 ndash 426 httpdxdoiorg1016450022-3395(2001)087[0424EOPITN]20CO2
322 Blankespoor HD Cameron SC Cairns J Jr 1985 Resistance of pul-monate snail populations to repeated treatments of copper sulfate En-viron Manage 9455ndash 458 httpdxdoiorg101007BF01866345
323 Mitchell AJ 2002 A copper-sulfate-citric acid pond shoreline treatmentto control the rams-horn snail Planorbella trivolvis N Am J Aquacult64182ndash187 httpdxdoiorg1015771548-8454(2002)0640182ACSCAP20CO2
324 Graefe G Aspoumlck H Picher O 1973 Auftreten von Bade-Dermatitis inOsterreich und Moumlglichkeiten ihrer Bekaumlmpfung Zentralbl BakteriolHyg 225398 ndash 405
325 Ledford JJ Kelly AM 2006 A comparison of black carp Mylopharyn-godon piceus redear sunfish Lepomis microlophus and blue catfish Icta-lurus furcatus as biological controls of snail populations N Am J Aquac-ult 68339 ndash347 httpdxdoiorg101577A05-0621
326 Venable DL Gaude AP Klerks PL 2000 Control of the trematodeBolbophorus confusus in channel catfish Ictalurus punctatus ponds usingsalinity manipulation and polyculture with black carp Mylopharyngodonpiceus J World Aquacult Soc 31158 ndash166 httpdxdoiorg101111j1749-73452000tb00349x
327 Wui YS Engle CR 2007 The economic impact of restricting use of blackcarp for snail control on hybrid striped bass farms N Am J Aquacult69127ndash138 httpdxdoiorg101577A05-0881
328 Sokolow SH Lafferty KD Kuris AM 2014 Regulation of laboratorypopulations of snails (Biomphalaria and Bulinus spp) by river prawnsMacrobrachium spp (Decapoda Palaemonidae) implications for con-trol of schistosomiasis Acta Trop 13264 ndash74 httpdxdoiorg101016jactatropica201312013
329 Haas W Haberl B Kalbe M Stoll K 1998 Traps for schistosomemiracidiacercariae p 359 ndash363 In Tada I (ed) IXth International Con-gress of Parasitology Monduzzi Editore Bologna Italy
330 Ahmed AA Babiker A Eltash LA Shiff C 2002 Development of amodified baited trap for detection of schistosome cercariae using naturaloils rich in polyunsaturated fatty acids in Sudan Acta Trop 82363ndash368httpdxdoiorg101016S0001-706X(02)00039-6
331 Haas W 1984 Schistosoma mansoni cercaricidal effect of 2-tetrade-
Horaacutek et al
188 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

cenoic acid a penetration stimulant Exp Parasitol 58215ndash222 httpdxdoiorg1010160014-4894(84)90037-7
332 Naples JM Shiff C Halden RU 2005 Reduction of infectivity of schis-tosome cercariae by application of cercaricidal oil to water Am J TropMed Hyg 73956 ndash961
333 Bunnag T Freitas JR De Scott HG 1977 The predatory activity ofLebistes reticulatus (Peter 1859) on Schistosoma mansoni miracidia inlaboratory experiments Trop Geogr Med 29411ndash 414
334 Knight WB Ritchie LS Liard F Chiriboga J 1970 Cercariophagicactivity of guppy fish (Lebistes reticulatus) detected by cercariae labelledwith radioselenium (75Se) Am J Trop Med Hyg 19620 ndash 625
335 Holliman RB Mecham J 1971 Macrostomum gigas (Turbellaria Rhab-docoela) a predator of Schistosoma mansoni cercariae J Parasitol 57680 ndash 681 httpdxdoiorg1023073277941
336 Christensen NO 1979 Schistosoma mansoni interference with cercarialhost-finding by various aquatic organisms J Helminthol 537ndash14 httpdxdoiorg101017S0022149X00005678
337 Christensen NO Frandsen F Nansen P 1980 The interaction of someenvironmental factors influencing Schistosoma mansoni cercarial host-finding J Helminthol 54203ndash205 httpdxdoiorg101017S0022149X0000660X
338 Thieltges DW de Montaudouin X Fredensborg B Jensen KT Ko-privnikar J Poulin R 2008 Production of marine trematode cercariaea potentially overlooked path of energy flow in benthic systems Mar EcolProg Ser 372147ndash155 httpdxdoiorg103354meps07703
339 Ibrahim MM 2007 Population dynamics of Chaetogaster limnaei (Oli-
gochaeta Naididae) in the field population of freshwater snails and itsimplications as a potential regulator of trematode larvae communityParasitol Res 10125ndash33 httpdxdoiorg101007s00436-006-0436-0
340 Michelson EH 1964 The protective action of Chaetogaster limnaei onsnails exposed to Schistosoma mansoni J Parasitol 50441ndash 444 httpdxdoiorg1023073275851
341 Wajdi N 1964 The predation of Schistosoma mansoni by the oligochaeteannelid Chaetogaster J Helminthol 38391ndash392 httpdxdoiorg101017S0022149X00033939
342 Hopkins SR Wyderko JA Sheehy RR Belden LK Wojdak JM 2013Parasite predators exhibit a rapid numerical response to increased para-site abundance and reduce transmission to hosts Ecol Evol 34427ndash 4438httpdxdoiorg101002ece3634
343 Lie KJ 1973 Larval trematode antagonism principles and possible ap-plication as a control method Exp Parasitol 33343ndash349 httpdxdoiorg1010160014-4894(73)90038-6
344 Ow-Yang CK Lie KJ 1972 Interaction between Hypoderaeum dingeriand Trichobilharzia brevis in the Lymnaea intermediate host ProcTwelfth Pac Sci Congr 1203
345 Soldaacutenovaacute M Kuris AM Scholz T Lafferty KD 2012 The role ofspatial and temporal heterogeneity and competition in structuring trem-atode communities in the great pond snail Lymnaea stagnalis (L) J Para-sitol 98460 ndash 471 httpdxdoiorg101645GE-29641
346 Walker JC 1979 Austrobilharzia terrigalensis a schistosome dominantin interspecific interactions with the molluscan host Int J Parasitol9137ndash140 httpdxdoiorg1010160020-7519(79)90104-8
Petr Horaacutek MSc PhD is a Professor of Par-asitology and Head of the Department of Para-sitology at the Faculty of Science Charles Uni-versity in Prague Czech Republic He obtainedhis parasitologybiology degree from theCharles University in Prague The President ofthe Czech Republic appointed Petr Horaacutek as aProfessor of Parasitology in 2005 ProfessorHoraacutek has presented and published on manytopics dealing with parasitic helminths in gen-eral and cercarial dermatitisavian schistosomesin particular His teaching activities lie in general parasitology and the biol-ogy of helminths His research is focused on functional morphology of hel-minths trematodes and avian schistosomes (systematics life cycles andhost-parasite interactions) Details of his personal data can be found athttpwwwpetrhorakeu
Libor Mikeš MSc PhD is an Assistant Pro-fessor of Parasitology at the Department of Par-asitology Faculty of Science Charles Universityin Prague from which he received his masterrsquosand doctoral degrees His international experi-ence includes eg stays at the Keele UniversityUnited Kingdom and the Institute for the Bio-technology of Infectious Diseases UTS SydneyAustralia Dr Mikeš is currently the Presidentof the Czech Society for Parasitology His basicacademic interests lie in the general biology ofhelminths (with a special focus on the trematode superfamilies Schistoso-matoidea and Diplostomoidea) He is also involved in teaching molecularand cellular interactions between parasites and hosts and biochemical para-sitology and he leads several field courses in parasitology His research andpublication activities are focused mainly on the biochemistry of helminths(proteomics peptidases of trematodes and monogeneans and glycobiologyof larval trematodes) and immune reactions of hosts against avianschistosomes
Lucie Lichtenbergovaacute MSc PhD is a Post-doctoral Fellow at the Department of Parasitol-ogy Faculty of Science Charles University inPrague She finished her masterrsquos study at theMasaryk University in Brno Czech Republicand obtained her PhD from the Charles Uni-versity in Prague Czech Republic Since herdoctoral study she has focused on the researchof interactions of avian schistosomes and theirnatural avian and accidental mammalian hostsImmunology and histopathology are her cen-ters of interest
Vladimiacuter Skaacutela MSc is a PhD student at theDepartment of Parasitology Faculty of ScienceCharles University in Prague Czech RepublicHis masterrsquos degree was obtained at the sameinstitution and his masterrsquos thesis focused onthe morphology and development of the mono-stome fluke Notocotylus attenuatus (DigeneaNotocotylidae) At present he is investigatingthe defensive activity of hemocytes of two lym-naeid snails transmitting avian schistosomesand the immunomodulation caused by trema-todes in their snail hosts His research is carried out in collaboration withAnthony J Walker from Kingston University London United KingdomVladimiacuter has presented the results of his research at several national andinternational scientific meetings
Continued next page
Cercarial Dermatitis
January 2015 Volume 28 Number 1 cmrasmorg 189Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-

Miroslava Soldaacutenovaacute MSc PhD received aPhD in parasitology at the Faculty of ScienceUniversity of South Bohemia CeskeacuteBudejovice Czech Republic in 2011 Simulta-neously she started as a Research Assistant atthe Institute of Parasitology Biology CentreAcademy of Sciences of the Czech Republicwhere she currently continues as a PostdoctoralFellow in the research team focused on trema-tode ecology and systematics Her 1-year post-doctoral fellowship (2012ndash2013) at the Univer-sity of Duisburg-Essen (Faculty of Biology Aquatic Ecology Germany)focused on the impact of eutrophication on the composition of trematodecommunities in large lake ecosystems in the Ruhr River screening of fresh-water snails in relation to trematode infection with avian schistosomes andassessment of potential risks of cercarial dermatitis for humans Her fields ofinterest comprise parasite ecology (trematode community structure and or-ganization in freshwater molluscs) environmental parasitology (use of par-asite communities in molluscs as bioindicators) and trematode taxonomy
Sara Vanessa Brant PhD is an Assistant Re-search Professor and Senior Collections Man-ager of the Museum of Southwestern BiologyDivision of Parasites at the University of NewMexico She obtained her parasitologybiologyMS and PhD from the University of Nebras-ka-Lincoln Her main research focus is the evo-lutionary and ecological diversity of trema-todes in particular those in the blood flukefamilies (Schistosomatidae Spirorchiidae San-guinicolidae) Her recent efforts have beenfunded to address global avian schistosome diversity and their host interac-tions She is also involved in efforts to motivate and promote the importanceof placing both parasite and host vouchers in permanent museum collec-tions for future studies
Horaacutek et al
190 cmrasmorg January 2015 Volume 28 Number 1Clinical Microbiology Reviews
on March 27 2020 by guest
httpcmrasm
orgD
ownloaded from
- Avian Schistosomes and Outbreaks of Cercarial Dermatitis
-