auxin-mediated ribosomal biogenesis regulates vacuolar
TRANSCRIPT

Auxin-Mediated Ribosomal Biogenesis Regulates VacuolarTrafficking in Arabidopsis W
AbelRosado,a Eun JuSohn,a,1 GeorgiaDrakakaki,a Songqin Pan,a Alexandra Swidergal,a YuqingXiong,b Byung-HoKang,b Ray A. Bressan,c and Natasha V. Raikhela,2
a Department of Botany and Plant Sciences and Center for Plant Cell Biology, University of California, Riverside, California 92521b Electron Microscopy and Bioimaging Lab, Interdisciplinary Center for Biotechnology Research, University of Florida,
Gainesville, Florida 32611c Department of Horticulture and Landscape Architecture, Purdue University, West Lafayette, Indiana 47907
In plants, the mechanisms that regulate the transit of vacuolar soluble proteins containing C-terminal and N-terminal
vacuolar sorting determinants (VSDs) to the vacuole are largely unknown. In a screen for Arabidopsis thaliana mutants
affected in the trafficking of C-terminal VSD containing proteins, we isolated the ribosomal biogenesis mutant rpl4a
characterized by its partial secretion of vacuolar targeted proteins and a plethora of developmental phenotypes derived
from its aberrant auxin responses. In this study, we show that ribosomal biogenesis can be directly regulated by auxins and
that the exogenous application of auxins to wild-type plants results in vacuolar trafficking defects similar to those observed
in rpl4a mutants. We propose that the influence of auxin on ribosomal biogenesis acts as a regulatory mechanism for auxin-
mediated developmental processes, and we demonstrate the involvement of this regulatory mechanism in the sorting of
vacuolar targeted proteins in Arabidopsis.
INTRODUCTION
A functional vacuole and intact protein trafficking system are
necessary for plant cell viability and function. Perturbations of the
trafficking machinery often affect vital cellular processes, such
as plant hormone responses, cytokinesis, and the development
of tissue specificity (Surpin and Raikhel, 2004). In the classical
view, many soluble plant vacuolar proteins are sorted away from
proteins destined for secretion at the trans-Golgi network, a
process that requires the presence of positive sorting signals in
the primary sequence of vacuolar proteins (Matsuoka and
Nakamura, 1999; Vitale and Raikhel, 1999; Robinson et al.,
2005). Two of these sorting signals, an N-terminal propeptide
(NTPP) and a C-terminal propeptide (CTPP), are directed to the
vacuole by distinct pathways that converge at the prevacuolar
compartment (PVC) (Miao et al., 2008). The NTPP pathway is
believed to be common to plants and yeast, and several com-
ponents of the machinery involved in the sorting of NTPP-type
cargoes have been characterized (Zheng et al., 1999; Ahmed
et al., 2000; Bassham and Raikhel, 2000). The CTPP pathway is
believed to be unique to plants, and different genetic approaches
have been used to identify components that are specific for that
pathway (Sanmartın et al., 2007; Sohn et al., 2007). To isolate
newcomponents of the plant-specificCTPP sortingmachinery in
Arabidopsis thaliana, we used a T-DNA–mutagenized population
in the Vac2 background (Vac2 T-DNA). The Vac2 line contains a
genetically engineered CLAVATA3 (CLV3) fused to the barley
(Hordeum vulgare) lectin C-terminal vacuolar sorting signal
(CLV3:CTPPBL) in the clv3-2 mutant background (Figure 1A).
Previous studies using genetic crosses and ethyl methanesulfo-
nate mutagenesis of the Vac2 line have been successfully used
for the identification of components involved in the specific
sorting of CTPP proteins (Sanmartın et al., 2007; Sohn et al.,
2007). In this report, the use of the Vac2 T-DNA screen allowed
us to identify a previously unknown component of the auxin-
mediated vacuolar sorting machinery, the cytosolic ribosomal
protein (r-protein) L4/L1 (RPL4A).
Although protein trafficking defects of ribosomal mutants have
not been evaluated, the association of ribosomal mutations with
deficient auxin perception and distribution has been widely
reported in the literature. The pointed first leaf 1 (pfl1)/r-protein
S18 (rps18a), pfl2/rps13b, and the semidominant Arabidopsis
minute-like1 (aml1)/rps5a mutants display auxin-related devel-
opmental defects, including growth retardation, narrow leaves
with reductions in the palisade mesophyll layer, and cotyledon
vascular pattern defects (Van Lijsebettens et al., 1994; Ito et al.,
2000; Weijers et al., 2001). Mutations in the short valve1 (stv1)/
r-protein L24 (rpl24b) result in an apical-basal patterning defect
of the gynoecium by influencing the translation of the auxin
response factors ARF3 and ARF5 (Nishimura et al., 2005). STV1
together with the proteins RPL28A and RPL5A has been shown
to have important roles in specifying leaf adaxial identity (Yao
et al., 2008), andRPL5A, RPL10A, andRPL9 have been shown to
modulate leaf patterning mechanisms via the ribosome-mediated
translational regulation of genes in theHD-ZIPIII-KANADI pathway
1Current address: Division of Molecular and Life Sciences, PohangUniversity of Science and Technology, Pohang 790-784, Korea.2 Address correspondence to [email protected] author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantcell.org) is: Natasha V. Raikhel([email protected]).WOnline version contains Web-only data.www.plantcell.org/cgi/doi/10.1105/tpc.109.068320
The Plant Cell, Vol. 22: 143–158, January 2010, www.plantcell.org ã 2010 American Society of Plant Biologists
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022
(Pinon et al., 2008). Finally, mutations in the nucleolar protein
PARL1, involved in ribosomal biogenesis, cause similar auxin
related developmental defects, suggesting that auxin regulation
depends on protein turnover and ribosome biogenesis in areas of
growth (Petricka and Nelson, 2007).
In bacteria, the RPL4 protein has been shown to be crucial for
the maintenance of ribosomal translational efficiency and fidelity
(O’Connor et al., 2004), but aside from structural roles within the
ribosome, no other specific function has yet been assigned in
plants. In this study, we analyze the implications of themutations
in theArabidopsisRPL4 family for protein trafficking aswell as for
hormonal regulation leading to the altered sorting of vacuolar
targeted proteins.
The Arabidopsis cytosolic r-protein RPL4 family is composed
of two transcriptionally active genes (RPL4A and RPL4D) and
two nonexpressed pseudogenes (RPL4B and RPL4C) (Barakat
et al., 2001). Our analysis of the two transcriptionally active
members of the RPL4 family in Arabidopsis suggests that the
RPL4A and RPL4D proteins have equivalent functions and are
coexpressed. Mutations in either RPL4 gene cause a similar
auxin-related developmental defect, and both proteins are in-
volved in the delivery of vacuolar targeted proteins to the
vacuole. Moreover, our results suggest that the sorting defects
in rpl4mutants are due to problems in protein turnover and auxin
perception in metabolically active tissues. We propose that
ribosomal biogenesis influenced by auxins is a high level regu-
latory mechanism in metabolically active tissues that regulates
the vacuolar delivery of not only CTPP, but also NTPP and
recycled proteins.
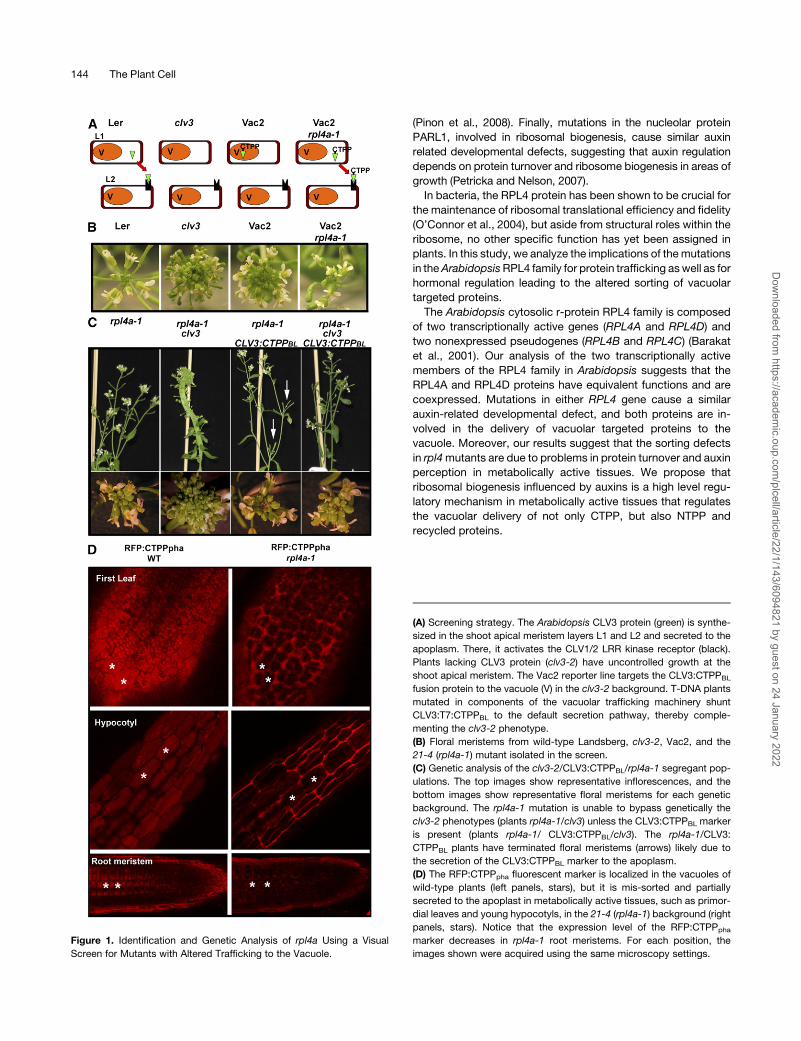
Figure 1. Identification and Genetic Analysis of rpl4a Using a Visual
Screen for Mutants with Altered Trafficking to the Vacuole.
(A) Screening strategy. The Arabidopsis CLV3 protein (green) is synthe-
sized in the shoot apical meristem layers L1 and L2 and secreted to the
apoplasm. There, it activates the CLV1/2 LRR kinase receptor (black).
Plants lacking CLV3 protein (clv3-2) have uncontrolled growth at the
shoot apical meristem. The Vac2 reporter line targets the CLV3:CTPPBL
fusion protein to the vacuole (V) in the clv3-2 background. T-DNA plants
mutated in components of the vacuolar trafficking machinery shunt
CLV3:T7:CTPPBL to the default secretion pathway, thereby comple-
menting the clv3-2 phenotype.
(B) Floral meristems from wild-type Landsberg, clv3-2, Vac2, and the
21-4 (rpl4a-1) mutant isolated in the screen.
(C) Genetic analysis of the clv3-2/CLV3:CTPPBL/rpl4a-1 segregant pop-
ulations. The top images show representative inflorescences, and the
bottom images show representative floral meristems for each genetic
background. The rpl4a-1 mutation is unable to bypass genetically the
clv3-2 phenotypes (plants rpl4a-1/clv3) unless the CLV3:CTPPBL marker
is present (plants rpl4a-1/ CLV3:CTPPBL/clv3). The rpl4a-1/CLV3:
CTPPBL plants have terminated floral meristems (arrows) likely due to
the secretion of the CLV3:CTPPBL marker to the apoplasm.
(D) The RFP:CTPPpha fluorescent marker is localized in the vacuoles of
wild-type plants (left panels, stars), but it is mis-sorted and partially
secreted to the apoplast in metabolically active tissues, such as primor-
dial leaves and young hypocotyls, in the 21-4 (rpl4a-1) background (right
panels, stars). Notice that the expression level of the RFP:CTPPpha
marker decreases in rpl4a-1 root meristems. For each position, the
images shown were acquired using the same microscopy settings.
144 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

RESULTS
rpl4a-1 Has Altered Protein Sorting to the Vacuole
rpl4a-1 is a mutant characterized from a T-DNA–mutagenized
population in the Arabidopsis Vac2 background (Rojo et al.,
2002). The Vac2 line was transformed with the pSKI015 plasmid,
and ;8000 lines were divided into 144 independent pools
(Koiwa et al., 2006). The pools were screened for complemen-
tation of the clv3-2 shoot meristem phenotype (Sanmartın et al.,
2007; Sohn et al., 2007; Zouhar et al.,2009) (Figure 1A). By this
approach, we identified amutant designated 21-4 that displayed
reduced floral meristem size (Figure 1B). Genetic analyses
showed that the 21-4 line harbored one functional T-DNA inser-
tion that caused a recessive mutation in a single nuclear locus
(see Supplemental Table 1 online). Thermal asymmetric inter-
laced PCR was used to determine the T-DNA insertion position
and the genomic sequence flanking the T-DNA left border. As a
result, a T-DNA insertion located 283 bp downstream of the ATG
translation start site in the first exon of the At3g09630 locus was
identified. The At3g09630 gene was annotated as the cytosolic
L4/L1 protein of the large 60S ribosomal complex subunit
(RPL4A). Genetic linkage analysis confirmed that the reduced
meristems, the T-DNA insertion in RPL4A, and the developmen-
tal phenotypes observed in different rpl4a alleles (Figure 2) were
linked within a genetic distance of 0.5 centimorgans (see Sup-
plemental Table 1 online). Upon identification of the mutation in
RPL4A, we performed phenotypic and genetic analyses to test
whether the mutation acted as a bypass suppressor of clv3-2.
The analysis of populations in which rpl4a-1, clv3-2, and CLV3:
CTPPBL were segregating indicated that rpl4a-1 mutation was
unable to genetically bypass the clv3 phenotypes (plants rpl4a-1/
clv3) unless CLV3:CTPPBL was present (plants rpl4a-1/clv3/
CLV3:CTPPBL) (Figure 1C). Interestingly, the combination rpl4a-
1/CLV3:CTPPBL caused terminated meristems that mimicked
the phenotypes observed in CLV3-overexpressing lines (Rojo
et al., 2002) (Figure 1C). Based on these results, we concluded
that the reduced floral meristem phenotype in 21-4 (rpl4a-1/clv3/
CLV3:CTPPBL) was due to the secretion of the CLV3:CTPPBL
fusion protein to the apoplasm and that rpl4a-1was not a bypass
mutation of clv3.
To confirm further that rpl4a-1 causes the secretion of vac-
uolar targeted proteins at the cellular level, we crossed the
rpl4a-1 mutant line with two lines expressing fluorescent vac-
uolar markers. The first marker contained the vacuolar sorting
signal of proricin appended to the N terminus of the monomeric
red fluorescent protein RFP1 (NTPPpro:RFP) (Hunter et al.,
2007). The secondmarker contained the vacuolar sorting signal
of phaseolin fused to the C terminus of RFP1 (RFP:CTPPpha)
(Hunter et al., 2007). Both markers included the signal peptide
from sporamine fused to the N-terminal end of RFP1 that
caused the reporter to be directed into the default secretion
pathway when the vacuolar sorting signals were not recognized
(Craddock et al., 2008). Homozygous lines from the F2 gener-
ation were obtained, and the localization of the RFP fusion
proteins in different tissues and developmental stages using
scanning fluorescent confocal microscopy was determined. As
shown in Figure 1D, wild-type plants displayed vacuolar local-
ization of the RFP:CTPPpha in all tissues, whereas secretion in
the first pair of true leaves and lower hypocotyls and decreased
expression and partial secretion in root meristems were ob-
served in the rpl4a-1 line. Similar analyses were performed with
the NTPPpro:RFP marker, which presented similar vacuolar
localization in wild-type plants, but the secretion of NTPPpro:
RFP in different tissues from rpl4a-1 mutants was not as
dramatic as in the case of RFP:CTPPpha (see Supplemental
Figure 1 online). The differential results obtained with the
different vacuolar markers might be partially explained by the
lower expression of NTPPpro:RFP in both wild-type and rpl4a
backgrounds that, in turn, decreased the marker secretion
signal (see Supplemental Figure 1 online). The results obtained
from the vacuolar fluorescent markers analyses validate the
genetic screen for vacuolar trafficking mutants and suggest
that RPL4A is required for the proper sorting of vacuolar-
targeted proteins.
rpl4a Displays Aberrant Auxin-Related
Developmental Phenotypes
Early in its development, the rpl4a-1 mutant displays very
characteristic narrow pointed first leaves (pfl) and retarded
growth (Figure 2A). The pfl phenotype in rpl4a-1 mutants re-
sembled the phenotypes of the r-protein knockout mutants
rps13b and rps18a (Van Lijsebettens et al., 1994; Ito et al., 2000)
and was also similar to the phenotypes observed in RPL23-
silenced lines (Degenhardt and Bonham-Smith, 2008). To con-
firmwhethermutations in theRPL4A genewere the cause of the
pfl phenotypes, two additional T-DNA alleles in the Columbia-0
(Col-0) genetic background, rpl4a-2 (SALK_130595) and rpl4a-3
(SALK_063782), were isolated. Homozygous rpl4a-2 and rpl4a-3
seedlings exhibited similar developmental phenotypes to those
of rpl4a in a Landsberg background (Figure 2A). In early stages
of development, it was also observed that rpl4a-1 displayed
altered cotyledon architecture associated with aberrant distri-
butions of auxin maxima sites (Figure 2B). The vascular struc-
ture in the first leaves of rpl4a-1 deviated from the closed and
reticulate venation of controls. rpl4a-1 presented substantially
reduced venation, few or no tertiary or quaternary veins, and
aberrant anastamosis close to the hypocotyl-petiole junction
(Figure 2C). Other physiological and developmental conse-
quences of the rpl4a-1 mutation included reduced root elon-
gation, altered root gravitropic responses, delayed transition to
reproductive phase, and decreased hypocotyl elongation in
etiolated seedlings (Figures 2D to 2F). Impaired auxin homeo-
stasis may explain these phenotypic traits, which were remi-
niscent of mutants with altered ribosome biogenesis and auxin
responses (for references, see Supplemental Table 2 online).
However, the severely altered auxin responses observed in
rpl4a-1 did not influence the subcellular localization, polarity,
and tissue expression of auxin transporters, such as AUX1,
PIN2, and PIN7, in light-incubated rpl4a seedlings (see Sup-
plemental Figure 2 online).
Because rpl4a-1 shared common features with a plethora of
previously reported ribosomal mutants, we investigated whether
other common phenotypes found in different r-protein mutants
were present in rpl4a-1 (see Supplemental Table 2 online). Leaf
Ribosomal Regulation of Vacuolar Sorting 145
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

patterning organization and adaxial-abaxial polarity were dis-
rupted at the cellular level in rpl4a-1. Epidermal cells of the
adaxial lamina surface of the first leaf of rpl4a-1were altered and
presented angular lobes as opposed to the jigsawpatterning and
smooth lobes in the wild type. Abaxial surfaces in rpl4a-1 were
also different and displayed a mosaic pattern of normal abaxial
patches mixed with outgrowths and heterogeneous cell sizes
(Figure 2G). The stained transverse sections of first leaf cells in
rpl4a-1 presentedmany enlarged cells and intercellular spaces in
the adaxial palisade region, and the number of subepidermal
palisade cells was fewer than that of the wild type (Figure 2H).
Finally, the rpl4a-1 root meristem cells displayed more disor-
ganized nucleolar structures with an abundance of large nu-
cleolar vacuoles (71% of rpl4a nuclei n = 44) compared with
controls (10% of Vac2 nuclei n = 39), suggesting that ribosomal
biogenesis could be affected in that tissue (Figure 2I). The
extensive phenotypic similarities of rpl4a-1 with previously
reported r-protein mutants involved in ribosomal biogenesis,
together with the aberrant nucleolar structures in rpl4a-1,
support the hypothesis that the ribosome per se, and not
r-proteins acting independently, regulates auxin-mediated de-
velopmental processes.
Figure 2. The rpl4 Family Mutants Display Aberrant Auxin-Related Developmental Defects.
(A) Independent rpl4a alleles display narrow pointed first leaves. The original rpl4a-1mutant was compared with the wild-type Landsberg, whereas the
rpl4a-2 (SALK_130595) and rpl4a-3 (SALK_063782) alleles were compared with the wild-type Col-0.
(B) rpl4a-1 cotyledon structures and auxin maxima localizations. In the far right panel, GUS staining of rpl4a-1 cotyledons expressing the auxin reporter
DR5pro:GUS. The right cotyledon shows aberrant auxin maxima localizations (black stars), and the left cotyledon shows the expected apical auxin
maxima localization (white star).
(C) to (F) Auxin-related phenotypes in rpl4a-1 include incomplete vascular development (C), reduced root elongation and altered root gravitropic
responses (D), delayed transition to reproductive phase (E), and reduced hypocotyl elongation in etiolated seedlings (F).
(G) Scanning electron micrographs of the adaxial and abaxial surfaces of late rosette leaves from 28-d-old plants of the wild type (top) and rpl4a-1
(bottom).
(H) Stained transverse sections of first leaf cells in rpl4a-1 presented many enlarged cells and intercellular spaces in the adaxial palisade region and
fewer subepidermal palisade cells than did the wild type.
(I) Transmission electron microscopy of root apical meristem cells shows aberrant nucleolar structures in the rpl4a-1 mutant. NUC, nucleolus; NP,
nucleoplasma; NV, nuclear vacuole.
146 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

Arabidopsis Has Heterogeneous RPL4-Containing
Ribosomes in Specific Tissues
The RPL4 family in Arabidopsis comprises two nearly identical
expressed members, RPL4A and RPL4D (Barakat et al., 2001;
Carroll et al., 2008) (Figure 3A). RPL4A is a basic protein (pI =
11.04) of 406 amino acids that shares an overall 95.1% amino
acid identity with RPL4D. The T-DNA mutants in the RPL4A
(rpl4a-2 and rpl4a-3) and the RPL4D (rpl4d-1 and rpl4d-2) genes
in the Col-0 background were isolated from the SALK collection
and their mRNA expressions analyzed by RT-PCR. As shown in
Figure 3B, the rpl4a-2 mutant (hereafter referred to as rpl4a)
showed reducedRPL4A expression, whereas the rpl4a-3, rpl4d-1
(hereafter referred to as rpl4d), and rpl4d-2 mutants showed no
discernible transcripts after 35 PCR cycles.
When the rpl4a and rpl4d mutants were grown side by side,
they were indistinguishable and displayed similar auxin-related
phenotypes (see Supplemental Figure 3 online). The high degree
of amino acid identity between RPL4A and RPL4D, together with
the similar phenotypes observed in the rpl4a and rpl4dmutants,
suggested that both proteins have similar functions. To evaluate
further their spatial and temporal equivalence, their respective
promoter:b-glucuronidase (GUS) expression patterns were an-
alyzed, and no significant differences between RPL4Apro:GUS
and RPL4Dpro:GUS expression patterns in our experimental
conditions were observed (Figures 6A to 6F; see Supplemental
Figure 4D online). Because a given ribosomal complex contains
only one RPL4 family member at a time (Ban et al., 2000), these
results suggested that distinct ribosomal populations (RPL4A
and RPL4D ribosomal complexes) simultaneously exist in wild-
type plants. This conclusion is supported by previous proteomic
studies that demonstrated the presence of ribosomal heteroge-
neity in Arabidopsis (Chang et al., 2005; Giavalisco et al., 2005;
Carroll et al., 2008).
The RPL4 Family Requires at Least Two Functional Gene
Copies for Plant Viability
We attempted to enhance the phenotypes observed in rpl4
mutants by eliminating both RPL4 proteins or, in case of lethality,
to determine the minimum RPL4 gene dosage required for plant
viability. For that purpose, we successively eliminated RPL4
gene function by performing reciprocal crosses between the
Figure 3. Genetic Analysis of the F2 Population from the Cross rpl4a 3 rpl4d.
(A) Exon-intron organization and schematic representation of the locations of the T-DNA insertions in the RPL4A and RPL4D genes. Orange, exon;
purple, intron; red, untranslated regions.
(B) RPL4 transcripts detected in 2-week-old seedlings in wild-type (Col-0) and rpl4a and rpl4d mutant alleles using RT-PCR. The positions of the
oligonucleotides used for the RT-PCR (F and R) are indicated with arrows in (A).
(C) Genotypes and segregation analysis of the F2 individuals from the cross rpl4a-23 rpl4d-1 (rpl4a3 rpl4d). The possible genotypic F2 combinations
derived from the rpl4a3 rpl4d cross are represented with green and red bars. Green bars indicate RPL4 genomic copies without T-DNA insertions. Red
bars with black triangles indicate RPL4 genomic copies containing T-DNA insertions.
Ribosomal Regulation of Vacuolar Sorting 147
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

rpl4a and rpl4dmutants. Surprisingly, F1 plants from the rpl4a3rpl4d cross that were heterozygous for both recessive mutations
displayed similar morphological defects as the single mutants,
with one exception: cleared siliques of either of the single rpl4
mutants were normal, but the double heterozygous plants
showed empty spaces in the siliques, indicating embryo lethality
of;30% of expected seeds (see Supplemental Figure 5 online).
To confirm this result, phenotypic and genotypic analyses of 144
segregant F2 plants from the rpl4a 3 rpl4d cross without pfl
phenotype and 144 segregant F2 plants with pfl phenotype were
completed (Figure 3C). The analysis of the plants without phe-
notype resulted in 36 wild-type plants and 108 plants with a
T-DNA insertion in one of the four RPL4 copies, providing the
expected segregation ratio. However, all 144 plants that dis-
played the pfl phenotype had T-DNA mutations in two RPL4
copies and significant segregation distortions (x2 >16.75, P <
0.005) were observed. In this random population, we did not
identify plants with insertions inmore than twoRPL4 copies. This
result suggests that two active copies of RPL4, independent of
their identities, define the minimum threshold for plant viability.
The rpl4Mutants Have Altered Ribosomal Functions
In Escherichia coli, multiple defects in translation associatedwith
altered ribosomal protein L4 activity (the homolog of the Arabi-
dopsis RPL4s) have been shown (O’Connor et al., 2004). In
Arabidopsis, r-protein mutants, such as stv1/rpl24b (Nishimura
et al., 2005), rpl5a, and rpl9 (Yao et al., 2008), and the three
piggybackmutants (Pinon et al., 2008) have translational defects
due to their aberrant ribosomal function. To test whether the
Arabidopsis rpl4mutations compromised ribosomal function in a
similar way to their bacterial counterpart and other Arabidopsis
r-proteins, we used an in vivo protoplast transient expression
assay. In this assay, various green fluorescent protein (GFP)
fusion proteins under the control of the 35S cauliflower mosaic
virus promoter were expressed in protoplasts isolated fromwild-
type and rpl4 seedlings, and the levels of accumulation of
different GFP-fused proteins were evaluated temporally using
anti-GFP antibodies.
As shown in Figure 4A, the levels of GFP accumulation using a
cytosolic 35S:GFP construct were reduced in rpl4a and rpl4d
when compared with the wild-type protoplast 6 h after transfor-
mation. Those differences were reduced when longer incubation
times were analyzed (9 and 12 h), until near-wild-type levels of
accumulationwere observed after 18 h in rpl4a. In our conditions,
rpl4d showed a slightly lower GFP accumulation than rpl4a in all
time points likely due to the residual RPL4A expression of the
rpl4a mutant allele (Figure 2B).
Because we determined that the cytosolic GFP levels after 6 h
were different in wild-type and rpl4 protoplasts, we tested
whether this reduction was a general effect or specific for
cytosolic proteins. As shown in Figure 4B, the levels of accumu-
lation of different GFP fusion proteins, which included proteins
destined for different compartments such as the ER, cytoplasm,
chloroplast, and vacuole, were reduced in both rpl4 mutants
compared with the control. These results suggest that indepen-
dently of their subcellular localization, the rate of protein synthe-
sis was reduced in rpl4 protoplasts; hence, it takes longer to
reach the same protein levels in rpl4 mutants relative to the wild
type. However, we cannot exclude the possibility that the differ-
ential protein accumulation observed after 6 h was due to
transcriptional differences or enhanced degradation of newly
synthesized polypeptides in rpl4 backgrounds.
Once we determined that rpl4 mutations cause protein syn-
thesis defects, we evaluated whether the altered rpl4 ribosomal
function was due to either a structural defect in fully assembled
ribosomes or to the lack of ribosomes. For that purpose, we
analyzed rpl4 responses against an array of antibiotics with
known ribosomal targeting locations in prokaryotes (see Sup-
plemental Table 3 online). The rpl4 mutants did not display
increased sensitivity to any tested compounds, including the
eukaryotic protein synthesis inhibitor cycloheximide, which
binds the eukaryotic ribosomal complexes in a stoichiometic
manner (Oleinick, 1977). Moreover, rpl4 mutants displayed mild
resistance to chloramphenicol and erythromycin in shoots (Fig-
ure 4C) and streptomycin in roots and isolated protoplasts
(Figures 4D and 4E). The lack of general resistance or hypersen-
sitivity to antibiotics or cycloheximide in rpl4 suggested that the
total number of ribosomeswas similar to that in thewild type, and
the resistance for specific antibiotics suggested that the protein
synthesis problems in the rpl4 background were due to the
presence of an aberrant population of ribosomes unable to bind
properly to specific antibiotics. Interestingly, the presence of
aberrant ribosomes in rpl4 did not trigger an enhanced unfolded
protein response as demonstrated by the similar expression of
ER resident chaperones at the transcriptional and translational
levels (see Supplemental Figure 6 online) and the similar sensi-
tivity to tunicamycin (see Supplemental Table 3 online) of the rpl4
mutants compared with wild-type plants.
The rpl4Mutants Activate Compensatory Mechanisms at
the Protein Level
Because the RPL4 family is composed of two coexpressed
members, we hypothesized that the general protein accumula-
tion defects in rpl4 might be partially buffered by compensatory
mechanisms at the protein level. To test this hypothesis, we
checked the levels of total RPL4 protein in both rpl4a and rpl4d
single mutant backgrounds compared with the wild type. In this
assay, the RPL4 proteins were indistinguishable due to high
homology at the amino acid sequence level, so the observed
signals using human antiRPL4-specific antibodies accounted for
the contributions of both members. As shown in Figure 5A,
upregulation of the combined RPL4 proteins was observed in
both rpl4a and rpl4d mutants compared with the wild type.
Because our mutant lines were knockdowns, it was possible that
the lack of one member increased the expression of the other
family member to compensate functionally. To test this hypoth-
esis, liquid chromatography/tandem mass spectrometry (LC/
MS/MS) analysis was performed using protein extracts from
rpl4a and rpl4d 2-week-old seedlings. As shown in Figures 5B
and 5C, specific peptides for RPL4A and RPL4D were identified
by LC/MS/MS, and the ion chromatograms for the correspond-
ing peptides were quantified. Figure 5D shows the quantification
of the RPL4A- and RPL4D-specific peptides, which indicated
that the rpl4a mutant expressed more RPL4D protein and vice
148 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022
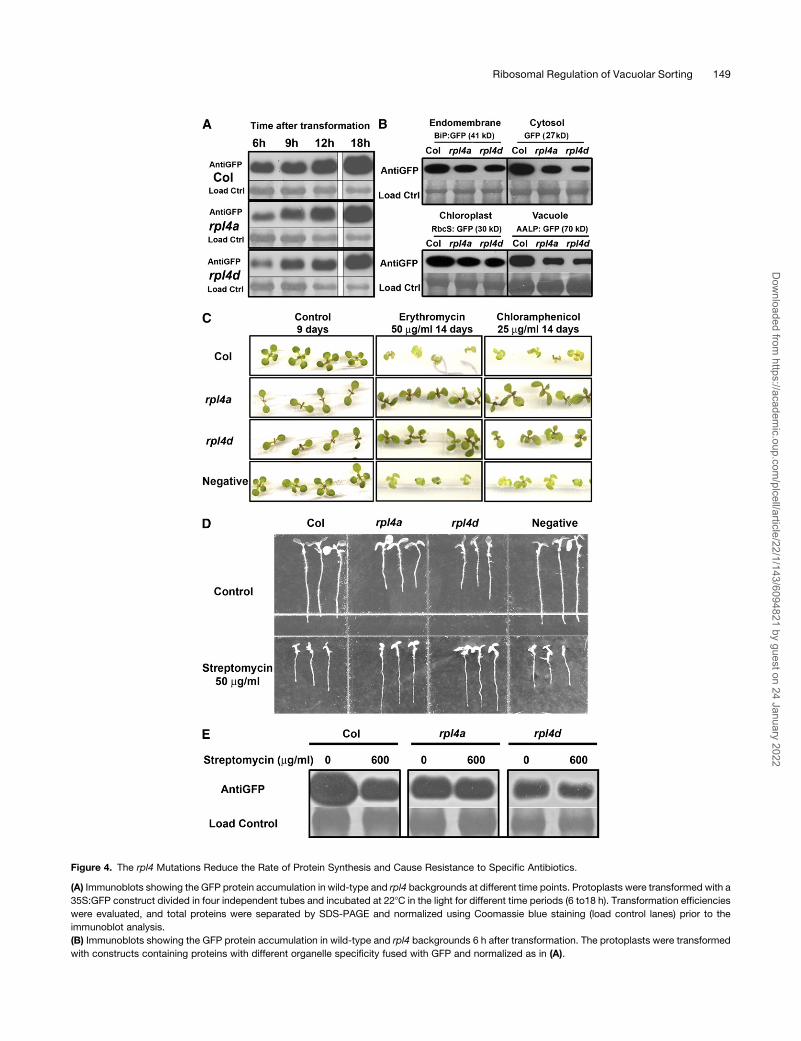
Figure 4. The rpl4 Mutations Reduce the Rate of Protein Synthesis and Cause Resistance to Specific Antibiotics.
(A) Immunoblots showing the GFP protein accumulation in wild-type and rpl4 backgrounds at different time points. Protoplasts were transformed with a
35S:GFP construct divided in four independent tubes and incubated at 228C in the light for different time periods (6 to18 h). Transformation efficiencies
were evaluated, and total proteins were separated by SDS-PAGE and normalized using Coomassie blue staining (load control lanes) prior to the
immunoblot analysis.
(B) Immunoblots showing the GFP protein accumulation in wild-type and rpl4 backgrounds 6 h after transformation. The protoplasts were transformed
with constructs containing proteins with different organelle specificity fused with GFP and normalized as in (A).
Ribosomal Regulation of Vacuolar Sorting 149
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

versa. This compensation at the protein level in rpl4 backgrounds
correlated with an increased transcriptional activity of the alter-
native RPL4 protein and an increased transcriptional activity of
the r-protein S6 (see Supplemental Figure 6 online), which is
an important regulator of ribosome biogenesis in eukaryotes
(Volarevic et al., 2000).
Although the compensatory mechanisms in planta cannot fully
fulfill the requirements of RPL4protein dosage in specific tissues,
this mechanism could explain several observations derived from
our study: the relatively mild phenotypes observed in the adult
rpl4 plants (Figure 2E), the specificity of the secretion patterns for
certain tissues (Figure 1D), and the lack of differences between
the wild type and rpl4a when the protein levels of the vacuolar
trafficking machinery in planta were analyzed (see Supplemental
Figure 6 online).
RPL4A Transcription Is Regulated by Auxins
A model including a general protein accumulation defect in
metabolically active tissues is inadequate to explain the speci-
ficity of the auxin phenotypes in rpl4a. To link ribosomal function
and specific auxin responses, we analyzed whether auxins
mediate transcriptional regulation of theRPL4 promoters. Based
on the phenotypes observed in Figure 2, we expected that the
RPL4 promoter activity would be correlated with patterns of free
auxin distribution. To confirm our hypothesis, we characterized
theGUS staining patterns ofArabidopsis plants transformedwith
the RPL4Apro:GUS and RPL4Dpro:GUS reporter gene con-
structs. As shown in Figure 6A and Supplemental Figures 4A to
4D online, plants transformed with the RPL4Apro:GUS and
RPL4Dpro:GUS constructs showed very similar staining distri-
bution that strongly resembled the auxin distribution patterns
throughout the Arabidopsis development described by Teale
et al. (2006). Briefly, RPL4Apro:GUS expression was strong
during early stages of primordia development, and leaf apical
dominance was evident in the elongating tip (Figure 6A, primor-
dial leaf 1 and leaf 2). In later stages of leaf development, GUS
expression progressed basipetally along the margins (Figure 6A,
leaf 3), until its final appearance in the central regions of the
lamina (Figure 6A, leaf 4), and in a more diffuse fashion in mature
leaf mesophyll cells (Figure 6A, leaf 5). In main and lateral roots,
RPL4Apro:GUS expression also correlated with auxin distribu-
tion patterns. In themain root,RPL4Apro:GUS stainingwasmore
intense toward the root tip and more diffuse in the epidermis
(Figure 6B). In lateral roots, theGUSactivity started from the stele
to the new root tip and then continued through the epidermis
(Figure 6C). Finally, GUS staining was also observed in second-
ary sites of free auxin production, such as the seed endosperm
cap, stigma, stamen, and pollen (Figure 6D to 6F). These results
suggested that ribosome biogenesis and auxin biosynthesis and
transport are tightly linked processes. To confirm further that this
linkage exists, we tested whether exogenously applied auxins
alter the expression of the RPL4Apro:GUS lines. As shown in
Supplemental Figure 4E online, exogenous applications of
auxins at physiological levels (10 mM concentrations for 24 h)
caused a general decrease in the RPL4A promoter activity in the
root meristem. Although this result suggested that auxins regu-
late RPL4A expression, no strong conclusions about the relative
sensitivity of the RPL4A promoter to different auxins could be
reached. To determine whether the different auxin treatments
were equivalent, exogenous applications of higher hormonal
concentrations (50 mM concentrations for 4 h) were used. As
shown in Figure 6G, the application of the natural auxin indole-3-
acetic acid (IAA), as well as the same concentration of the auxin
transport inhibitor 1-naphthylphthalamic acid (NPA), completely
abolished the RPL4A promoter activity in the root meristem and
root elongation zone without modifying the expression patterns
in the vasculature of fully developed roots. The same concentra-
tions and incubation times using 1-naphthaleneacetic acid (NAA),
2,4-D, and the auxin transport inhibitor 2,3,5-triiodobenzoic acid
caused a moderate decrease in GUS activity in root meristems,
suggesting that the promoter response to those treatments was
weaker (Figure 6G). As a positive control of induction, we evalu-
ated the effect of the similar treatments using the auxin inducible
DR5pro:GUS transgenic line (Ulmasov et al., 1997). As shown in
Figure 6H, DR5pro:GUS expression was induced by auxins;
however, no differences in induction were observed among the
NAA, IAA or 2,4-D treatments. In our conditions, no repression of
DR5pro:GUS was observed in the 2,3,5-triiodobenzoic acid and
NPA treatments compared with nontreated plants. Based on
these results, we concluded that auxins regulate ribosomal bio-
genesis in root meristems and that the RPL4 promoter is more
sensitive to IAA and NPA treatments.
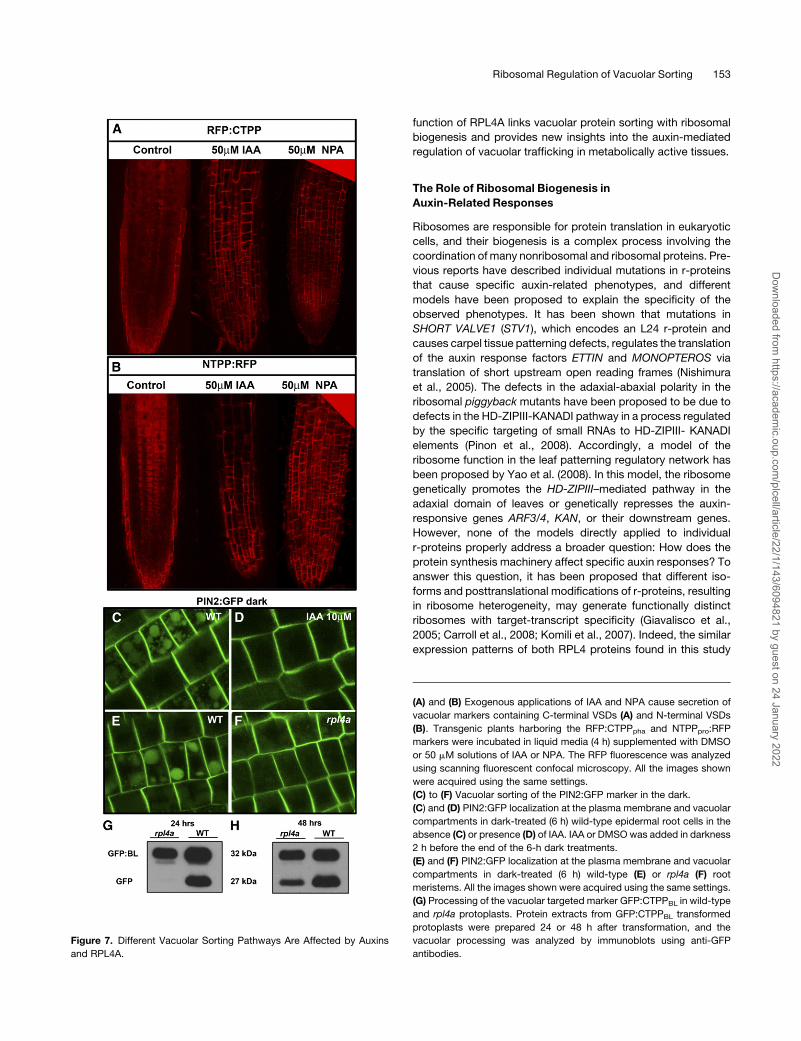
Different Vacuolar Sorting Pathways Are Affected byAuxins
and rpl4a
Since IAA and NPA treatments (50 mM concentrations for 4 h)
were able to completely abolish the RPL4A promoter activity in
root meristems, we hypothesized that those treatments applied
to wild-type plants might mimic the vacuolar sorting defects
caused by the rpl4amutation. As shown in Figures 7A and 7B, the
IAA and NPA treatments caused large amounts of secretion of
both RFP:CTPPpha and NTPPpro:RFP fluorescent markers
Figure 4. (continued).
(C) rpl4mutants shoots are resistant to erythromycin and chloramphenicol. Wild-type and rpl4 seeds were surface sterilized and directly germinated on
plates with or without antibiotic. Plates were incubated horizontally in a growth chamber under long-day conditions. An independent kanamycin-
resistant line SALK_069239 was used as a negative control to evaluate antibiotic cross-resistance.
(D) rpl4mutants roots are resistant to streptomycin. The assay was performed as in (C), but the plates were incubated vertically and pictures were taken
7 d after sowing.
(E) Transient expression assay in streptomycin-treated protoplasts. The GFP protein was expressed in protoplasts for 6 h, followed by a treatment with
0 and 600 mg/mL of antibiotic for 12 additional hours.
150 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022
restricted to the root meristems and the elongation zone of the
root. Since the hormonal treatments were not specific for either
RFP:CTPPpha or NTPPpro:RFP markers, we hypothesized that
RPL4A acts in a regulatory mechanism that affects vacuolar
trafficking generally, not just markers containing C- or N-terminal
propeptides. To test that hypothesis, we analyzed the vacuolar
transport of the auxin-efflux carrier PIN2, a protein without
vacuolar sorting signals that is targeted for vacuolar degradation
after darkness treatments (Kleine-Vehn et al., 2008). As shown in
Figure 7C, dark-incubated wild-type PIN2:GFP plants displayed
fluorescent signals in the plasma membrane, endosomes, and
vacuole; however, the combined treatment of darkness and IAA
in wild-type plants restricted the PIN2:GFP fluorescent signal to
the plasma membrane (Figure 7D). This result is in accordance
with previous studies that demonstrated that PIN2 endocytosis,
and, therefore, the delivery of the PIN2:GFP marker to the
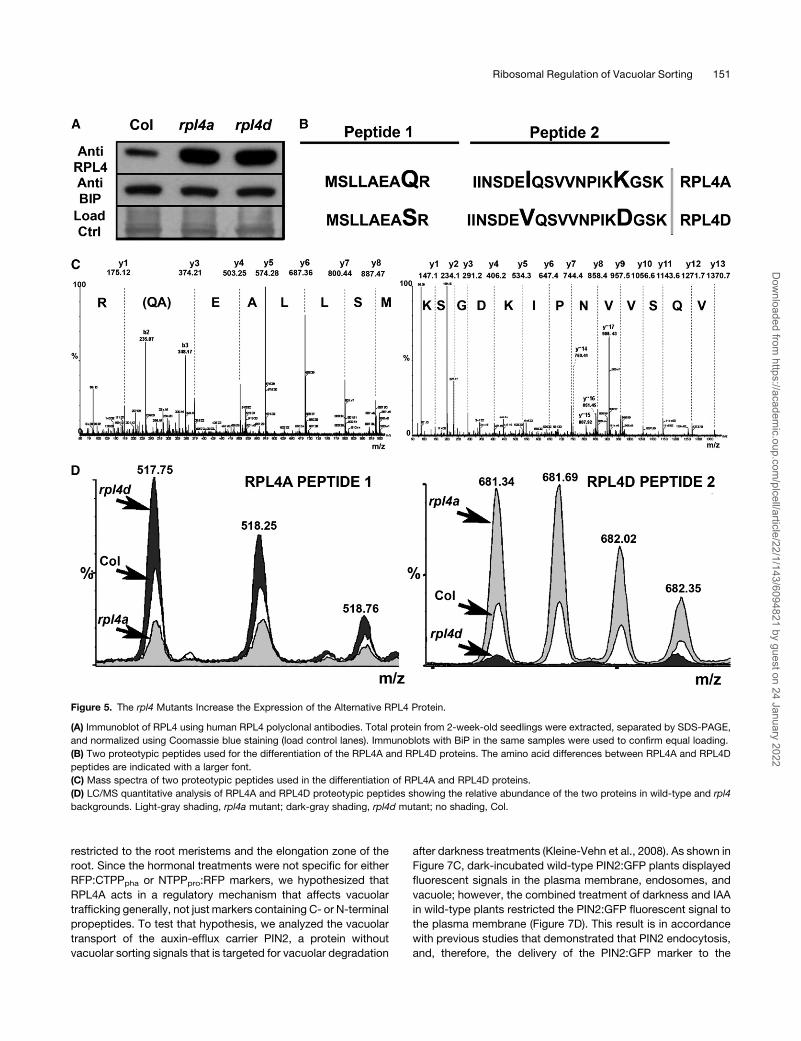
Figure 5. The rpl4 Mutants Increase the Expression of the Alternative RPL4 Protein.
(A) Immunoblot of RPL4 using human RPL4 polyclonal antibodies. Total protein from 2-week-old seedlings were extracted, separated by SDS-PAGE,
and normalized using Coomassie blue staining (load control lanes). Immunoblots with BiP in the same samples were used to confirm equal loading.
(B) Two proteotypic peptides used for the differentiation of the RPL4A and RPL4D proteins. The amino acid differences between RPL4A and RPL4D
peptides are indicated with a larger font.
(C) Mass spectra of two proteotypic peptides used in the differentiation of RPL4A and RPL4D proteins.
(D) LC/MS quantitative analysis of RPL4A and RPL4D proteotypic peptides showing the relative abundance of the two proteins in wild-type and rpl4
backgrounds. Light-gray shading, rpl4a mutant; dark-gray shading, rpl4d mutant; no shading, Col.
Ribosomal Regulation of Vacuolar Sorting 151
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022
vacuole, was inhibited by treatments with NAA (Paciorek et al.,
2005). Once we confirmed that IAA was also able to inhibit the
release of the PIN2:GFP marker to the vacuole, we crossed the
PIN2:GFP line with rpl4a and analyzed the vacuolar transport
of PIN2:GFP in rpl4a backgrounds and isogenic wild-type lines.
As shown in Figure 7E, the PIN2:GFP fluorescent signal of
dark-treated wild-type plants grown in solid media appeared in
plasma membrane, endosomes, and vacuole. However, the
PIN2:GFP signal of isogenic rpl4a plants was similar at the
plasma membrane but showed a marked decrease in the vac-
uole (Figure 7F). This result suggested that rpl4a mimics the
effect of exogenously applied IAA by partially inhibiting the
vacuolar delivery of PIN2:GFP. The partial inhibition of the PIN2:
GFP vacuolar delivery in rpl4a correlates with the partial secre-
tion of the RFP:CTPPpha and NTPPpro:RFP markers and sug-
gests that the trafficking of vacuolar targeted proteins is delayed
but not totally abolished in rpl4a. Finally, to analyze directly the
kinetics of trafficking to the vacuole, we analyzed the vacuolar
processing of a marker containing the Barley lectin vacuolar
propeptide fused with GFP (GFP:CTPPBL) in transformed wild-
type and rpl4a protoplasts. As shown in Figure 7G, 24 h after
transformation, a shift in the molecular weight of the GFP:
CTPPBL marker was observed in wild-type but not rpl4a proto-
plasts. This shift is due to the vacuolar processing of the BL
propeptide in wild-type plants. Forty-eight hours after transfor-
mation, rpl4a protoplasts started to process the marker; how-
ever, the ratio of nonprocessed versus processed GFP:CTPPBL
was higher in rpl4a than in wild-type protoplasts (Figure 7H). This
result confirms that trafficking toward the vacuole was strongly
delayed in rpl4a.
DISCUSSION
The results provided by the characterization of the ribosomal
rpl4a mutant isolated in this screen demonstrate that the Vac2
T-DNA collection is an excellent tool to describe mechanisms of
transport toward the plant vacuole. RPL4A belongs to a totally
different group of proteins compared with the ones identified
using ethyl methanesulfonate mutagenesis or genetic crosses of
the Vac2 line (Sanmartın et al., 2007; Sohn et al., 2007), demon-
strating that the Vac2T-DNA collection can identify unknown com-
ponents of plant vacuolar trafficking pathways. The described
Figure 6. The RPL4 Expression Correlates with Patterns of Free Auxin
Distribution, and It Is Regulated by Auxins.
(A) to (F) GUS activity assay in transgenic plants harboring the RPL4A
promoter-GUS fusion.
(A) Whole plant showing GUS staining in leaves at different develop-
mental stages (1 to 5).
(B) Root tip and root vasculature.
(C) Water-embedded seeds.
(D) Lateral root.
(E) Anthers and pollen grains.
(F) Stigma and petals.
(G) GUS activity assay in the presence of auxin transport inhibitors and
auxins in RPL4Apro:GUS transgenic plants. TIBA, 2,3,5-triiodobenzoic
acid.
(H) The auxin-inducible DR5pro:GUS transgenic lines were used as a
control.
152 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022
function of RPL4A links vacuolar protein sorting with ribosomal
biogenesis and provides new insights into the auxin-mediated
regulation of vacuolar trafficking in metabolically active tissues.
The Role of Ribosomal Biogenesis in
Auxin-Related Responses
Ribosomes are responsible for protein translation in eukaryotic
cells, and their biogenesis is a complex process involving the
coordination of many nonribosomal and ribosomal proteins. Pre-
vious reports have described individual mutations in r-proteins
that cause specific auxin-related phenotypes, and different
models have been proposed to explain the specificity of the
observed phenotypes. It has been shown that mutations in
SHORT VALVE1 (STV1), which encodes an L24 r-protein and
causes carpel tissue patterning defects, regulates the translation
of the auxin response factors ETTIN and MONOPTEROS via
translation of short upstream open reading frames (Nishimura
et al., 2005). The defects in the adaxial-abaxial polarity in the
ribosomal piggyback mutants have been proposed to be due to
defects in the HD-ZIPIII-KANADI pathway in a process regulated
by the specific targeting of small RNAs to HD-ZIPIII- KANADI
elements (Pinon et al., 2008). Accordingly, a model of the
ribosome function in the leaf patterning regulatory network has
been proposed by Yao et al. (2008). In this model, the ribosome
genetically promotes the HD-ZIPIII–mediated pathway in the
adaxial domain of leaves or genetically represses the auxin-
responsive genes ARF3/4, KAN, or their downstream genes.
However, none of the models directly applied to individual
r-proteins properly address a broader question: How does the
protein synthesis machinery affect specific auxin responses? To
answer this question, it has been proposed that different iso-
forms and posttranslational modifications of r-proteins, resulting
in ribosome heterogeneity, may generate functionally distinct
ribosomes with target-transcript specificity (Giavalisco et al.,
2005; Carroll et al., 2008; Komili et al., 2007). Indeed, the similar
expression patterns of both RPL4 proteins found in this study
Figure 7. Different Vacuolar Sorting Pathways Are Affected by Auxins
and RPL4A.
(A) and (B) Exogenous applications of IAA and NPA cause secretion of
vacuolar markers containing C-terminal VSDs (A) and N-terminal VSDs
(B). Transgenic plants harboring the RFP:CTPPpha and NTPPpro:RFP
markers were incubated in liquid media (4 h) supplemented with DMSO
or 50 mM solutions of IAA or NPA. The RFP fluorescence was analyzed
using scanning fluorescent confocal microscopy. All the images shown
were acquired using the same settings.
(C) to (F) Vacuolar sorting of the PIN2:GFP marker in the dark.
(C) and (D) PIN2:GFP localization at the plasma membrane and vacuolar
compartments in dark-treated (6 h) wild-type epidermal root cells in the
absence (C) or presence (D) of IAA. IAA or DMSOwas added in darkness
2 h before the end of the 6-h dark treatments.
(E) and (F) PIN2:GFP localization at the plasma membrane and vacuolar
compartments in dark-treated (6 h) wild-type (E) or rpl4a (F) root
meristems. All the images shown were acquired using the same settings.
(G) Processing of the vacuolar targeted marker GFP:CTPPBL in wild-type
and rpl4a protoplasts. Protein extracts from GFP:CTPPBL transformed
protoplasts were prepared 24 or 48 h after transformation, and the
vacuolar processing was analyzed by immunoblots using anti-GFP
antibodies.
Ribosomal Regulation of Vacuolar Sorting 153
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

support that heterogeneous ribosomal populations are present
in the same tissues, but also, the similarity of both rpl4 mutant
phenotypes suggest that RPL4A and RPL4D-containing ribo-
somes are equivalent and do not account for the target transcript
specificity. In accordance with the equivalence of RPL4 ribo-
somal types,RPL4 gene dosage, and not the identity of the RPL4
proteins, is the key element forArabidopsis survival. In our study,
50% RPL4 gene dosage reductions caused similar phenotypes,
independent of RPL4 identity, and 75% RPL4 gene dosage
reductions caused lethality. Furthermore, the activation of com-
pensatory mechanisms that increased the protein levels of the
alternate RPL4 protein in mutant backgrounds suggest that
RPL4 ribosomal heterogeneity does not confer auxin specificity,
but acts as a backup mechanism to maintain basic cellular
functions when RPL4 availability is limited.
To explain the auxin specificity of the r-protein mutant re-
sponses, our study uses a global analysis of the phenotypes
derived from ribosomal mutations (see Supplemental Table 2
online). Since most of the r-protein mutant phenotypes are
similar, we propose that those phenotypes are downstream
effects of a failure in a common regulatory mechanism for all
r-proteins, the auxin-mediated ribosomal biogenesis. At least
four independent lines of evidence were provided to support this
model. First, as indicated above, except for variations in the
severity of the phenotypes, and rare specific phenotypes that
could be associated with extraribosomal functions (Wool, 1996),
several independent r-protein mutants described in the literature
display very similar auxin-related phenotypes (see Supplemental
Table 2 online). Second, parl1, a mutant in a nucleolin that
globally affects ribosomal biogenesis (Petricka and Nelson,
2007) displays auxin-related phenotypes similar to those of rpl4
mutants. Third, the transcriptional levels of RPL4, and presum-
ably other coexpressed r-proteins, can be modified by direct
application of auxins and auxin transport inhibitors in certain
tissues. Finally, our general model predicts that alterations in the
auxin-mediated ribosomal biogenesis might regulate auxin-
related processes not previously reported, for example, the
sorting of vacuolar proteins (see below).
Role of Ribosomal Biogenesis in the Vacuolar Sorting
of Proteins
rpl4a was initially isolated in a screen for mutants with altered
protein sorting to the vacuole, and we hypothesize that the
mis-sorting is caused by an imbalance in the regulation of the
auxin-mediated ribosomal biogenesis. To illustrate the role of
ribosomal biogenesis in vacuolar trafficking in Arabidopsis, we
will use a process known to be regulated by auxins and vacuolar
trafficking elements, leaf vascular development. Leaf veins are
known to form by differentiation of vascular cells from ground
meristem cells in a process regulated by the polar flow of auxins
(Galweiler et al., 1998). In this context, the PVC and vacuoles
have recently been reported to function in the regulation of the
polarized transport and recycling of the auxin efflux carriers PIN1
and PIN2 (Kleine-Vehn et al., 2008; Laxmi et al., 2008; Shirakawa
et al., 2009; Spitzer et al., 2009). Severalmutants belonging to the
PVC-to-vacuole pathway, including vti11 vti12/+, vps9a, and
vam3, have been reported to have defects in protein trafficking
and vascular network formation (Shirakawa et al., 2009). VPS9A
is the homolog of yeast VPS9, which is involved in vacuolar
sorting, the vti11 vti12/+mutant has defects in the sorting of seed
storage protein (Goh et al., 2007, Sanmartın et al., 2007), and
vam3belongs to the vacuolar SNARE (solubleN-ethylmaleimide-
sensitive factor attachment protein receptor) complex and is
reported to be localized to the PVC and the vacuolar membrane
(Sato et al., 1997; Sanderfoot et al., 1999, 2001). Since we have
Figure 8. Model: Ribosomal Biogenesis Affects the Sorting of Vacuolar
Proteins in a Process Regulated by Auxins.
Local auxin accumulations cause the transcriptional repression of ribo-
somal proteins (1), followed by the translational repression of auxin-
responsive factors (such as HD-ZIPIII, KANADI, and ARF3/ARF4) (2).
r-protein mutations mimic the effect of exogenous auxin application by
decreasing the auxin-responsive factor synthesis rate (2). Whether the
decreased protein synthesis rate acts as a feedback mechanism that
represses auxin biosynthetic genes remain unclear (3). The translational
repression of auxin-responsive factors slows down the PVC-to-vacuole
delivery of vacuolar sorted markers, such as CTPP:RFPpha, NTPPpro:
RFP, NTPPBL:GFP, and PIN2:GFP, either directly or through PVC ele-
ments, such as VAM3, VPS9A, or VTI11 (4). The delayed delivery of the
CTPP:RFPpha and NTPPpro:RFP markers to the vacuole leads to their
partial secretion (5), whereas the delayed processing of PIN2:GFP (6)
likely modifies the auxin polar flows and causes the array of auxin-related
developmental phenotypes observed in rpl4 mutants (7). Black arrows:
normal delivery of CTPP:RFPpha, NTPPpro:RFP, and PIN2 to the vacuole
in wild-type Arabidopsis. Dashed arrows: effects of local auxin accumu-
lations or rpl4a backgrounds in vacuolar trafficking.
154 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

shown that the delivery of vacuolar proteins is delayed in rpl4a, it
is not surprising that the vascular defects observed in mutants
with altered PVC-to-vacuole trafficking resemble the pheno-
types observed in many r-protein mutants and particularly rpl4.
Interestingly, the explanatory models are highly complementary.
From the vacuolar trafficking perspective, the defects in protein
trafficking caused by the PVC-to-vacuole mutants modify the
distribution of auxin. This is achieved by regulating the localiza-
tion of auxin carriers (PINs), which in turn, determines where
procambium cells are located in the leaf primordium (Shirakawa
et al., 2009). In our model (Figure 8), auxin-dependent ribosomal
biogenesis affects the sorting of vacuolar targeted proteins, as
demonstrated by the delayed processing of the vacuolar tar-
geted protein GFP:CTPPBL, the secretion of the RFP:CTPPpha
and NTPPpro:RFP markers, and the delayed processing of PIN2
in the rpl4 backgrounds. Thus, in specific tissues, auxin accu-
mulations cause the transcriptional repression of r-proteins,
followed by the translational repression of auxin-responsive
factors, such as HD-ZIPIII, KANADI, and ARF3/ARF4, regulated
by the r-proteins (Nishimura et al., 2005; Pinon et al., 2008; Yao
et al., 2008). As a consequence, downstream elements affecting
the sorting of vacuolar targeted proteins are repressed, and the
vacuolar delivery of the CTPP:RFPpha, NTPPpro:RFP, NTPPBL:
GFP, and PIN2:GFP markers is delayed. The delayed delivery of
theCTPP:RFPpha andNTPPpro:RFPmarkers to the vacuole leads
to their partial secretion; meanwhile, the delayed processing of
PIN2 likely modifies the auxin polar flow required for the forma-
tion of the vascular network. In this way, the delayed vacuolar
degradation of PIN2, or other PIN proteins, might explain the
vascular defects and other auxin-related developmental pheno-
types observed in the rpl4 mutants.
In conclusion, we speculate that auxin-dependent ribosomal
biogenesis is an active part of a previously unknown mechanism
regulating the proper delivery of vacuolar targeted proteins (such
as PIN2) to the vacuole. Using this mechanism, ribosomal
biogenesis might regulate auxin fluxes and influence auxin-
related developmental processes. Whether this regulation oc-
curs through general mechanisms dependent on protein and
lipid synthesis, for example, of membrane sterols (Pan et al.,
2009), or through the inhibition of specific PVC elements, such as
VPS9A, VAM3, or VTI11, as suggested by the similarities in the
vascular defects when compared with rpl4 mutants is still un-
clear, and it is the focus of our current research.
METHODS
Plant Materials, Insertion Identification, and Growth Conditions
The Arabidopsis thaliana Vac2 T-DNA insertion lines in the Landsberg
background used in the screen were a gift from Ray A. Bressan (Koiwa
et al., 2006). The mutant screen was performed as described (Sohn et al.,
2007), and the DNA flanking the left border of the inserted T-DNA was
isolated by thermal asymmetric interlaced-PCR (Liu et al., 1995; Koiwa
et al., 2002). The NTPPpro:RFP and RFP:CTPPpha vacuolar markers
(Hunter et al., 2007) were a gift from Lorenzo Frigerio. For histochemical
analysis of the RPL4pro:GUS expression, the DR5pro:GUS transgenic
line (Ulmasov et al., 1997) was used as a control. The Arabidopsis Col-0
T-DNA insertion mutants SALK_130595 (rpl4a-2), SALK_063782 (rpl4a-3),
SALK_029203 (rpl4d-1), and SALK_065625 (rpl4d-2) and the control line
SALK_069239 were obtained from the ABRC stock center. The amplified
fragments in the 21-4 mutant were sequenced, and genetic cosegrega-
tion analysis was performed using an F2 population created by back-
crossing 21-4 to Vac2. Primer sets used are shown in Supplemental Table
4 online. For growth assays, surface-sterilized and cold-stratified Arabi-
dopsis seeds were sown onto half-strength Murashige and Skoog
phytoagar medium (0.53 Murashige and Skoog salts, 10 g/L sucrose,
and 7 g/L phytoagar, pH 5.7). Phytoagar plates containing sterilized seed
were incubated in an environmental chamber that was set for long-day
lighting conditions (16 h light/8 h dark) and a temperature of 228C. For
antibiotic treatments, the appropriate amounts of filter sterilized antibiotic
stock solutions were added to cooled autoclaved growth media. The
seeds were directly germinated on the antibiotic-containing plates, and
the plates were incubated horizontally for shoot analysis or vertically for
root elongation assays for up to 21 d.
RNA Isolation and Transcript Analyses
Total RNAwas prepared from2-week-old Col-0, rpl4a-2, rpl4a-3, rpl4d-1,
and rpl4d-2 Arabidopsis plantlets grown in sterile conditions using the
RNeasy plant mini kit (Qiagen), and cDNA was synthesized with the
Superscript III cDNA synthesis kit (Invitrogen) applying anchored-oligo
(dT)18 primer with equalized amounts of template RNA. PCR (30, 35, and
40 cycles) were performed with the cDNAs and ExTaq polymerase
(TaKaRa), and products were visualized with ethidium bromide staining.
For quantitative PCRs, SYBRGreen Supermix (Bio-Rad) was used in
reaction volume of 25 mL. Reactions were run on an iQ5 thermocycler
(Bio-Rad). For relative quantification, ratios were calculated according to
the method of Pfaffl (2001). All Ct values were the average of three
replicates, and all treatments were subject to three biological replicates.
Primer sets used are shown in Supplemental Table 4 online.
RPL4pro:GUS Constructs and Histochemical Analysis
To obtain theRPL4A andRPL4Dpromoter:GUS-fusions, 1.7 and 1.5 kb of
the genomic sequence upstream of the RPL4A and RPL4D translation
start sites were cloned using the Gateway recombination cloning technol-
ogy (Invitrogen) into the binary vector PMDC162 (Curtis and Grossniklaus,
2003) and transformed into Agrobacterium tumefaciens strain GV3101.
The primer sets used are shown in Supplemental Table 4 online. Wild-
type Col-0 plants were transformed using floral dipping, and primary
transformants were isolated using hygromycin as a selective marker.
Eight independent homozygous lines were isolated for each construct.
For hormonal treatments, the transgenic RPL4Apro:GUS and DR5pro:
GUS lines were incubated in liquid media supplemented with 50 mM
solutions of each compound for 4 h, andGUS activity was detected in situ
as described (Jefferson et al., 1987).
Microscopy
For scanning electron microscopy of adaxial and abaxial leaf surfaces,
fresh leaves were mounted on scanning electron microscopy stubs, flash
frozen in liquid air, and imaged in a Hitachi TM-1000 scanning electron
microscope at an accelerating voltage of 5 kV. Confocal images of water-
mounted seedlings were collected using a Leica TCS SP2/UV fitted with
320 or 363 water immersion objectives. A 543-nm line from a He-Ne
laser was used to excite RFP, and fluorescence was detected in the 560-
to 640-nm range. A 488-nm line from an argon laser was used to excite
GFP, and fluorescence was detected in the 500- to 530-nm range. For
analysis of leaf vasculature, leaf 5 of 28-d-old plants were fixed in 3%
glutaraldehyde, dehydrated through an ethanol series to 100% ethanol,
and embedded in JB4 resin (Agar Scientific). Embedded tissue was
sectioned at 3 mm and subsequently stained with 0.02% Toluidine Blue.
Ribosomal Regulation of Vacuolar Sorting 155
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

Images were obtained using a Nikon E800 microscope. For transmission
electron microscopy, 5-d-old Vac2 and rpl4a-1 root tips were dissected
out, transferred to a B-type aluminum planchette (Technotrade Interna-
tional) containing a 150 mM sucrose solution, and frozen with a HPM 100
high-pressure freezer (Leica). The frozen root tipswere freeze-substituted
in acetone containing 2% OsO4 at 2808C for 4 d. After substitution, the
sample temperaturewas slowly warmed up to2208Cover 24 h, from220
to 48C over 12 h, and from 48C to room temperature over 4 h. The root tips
werewashed three timeswith anhydrous acetone and embedded in Epon
resin (Ted Pella). After polymerization, samples were sliced into 80-nm
sections and stained with uranylacetate solution (2% w/v) and subse-
quently with lead citrate solution (26 g/L lead nitrate and 35g/L sodium
citrate). Electron micrographs were captured with a Hitachi TEM H-7000
operated at 80 kV.
Proteomic Analysis
Total protein extracts purified from 2-week-old Arabidopsis seedlings
were separated by SDS-PAGE. The gels were blotted onto nitrocellulose,
and immunoblot analysis was performed using human RPL4-specific
antibodies (Proteintech Group). The protein amounts were normalized
using Coomassie Brilliant Blue staining and AntiBiP antibodies (Rose
Biotechnology). Protein bands corresponding to the immunoblot signal
were excised and prepared as previously described (Carter et al., 2004).
Proteotypic peptides ions for RPL4A and RPL4D were selected based on
the criteria described by Baerenfaller et al. (2008) for the inclusion list to
perform targeted data-dependent acquisition analysis using a nanoLC/
MS/MS system with combination of the nano-Acquity UPLC and Q-TOF
Premier (Waters). After peptide sequenceswere confirmed by the tandem
mass spectra, the relative abundance of RPL4A and RPL4D in different
backgrounds was evaluated using LC/MS quantitative analysis as pre-
viously described (Rojo et al., 2004).
Transient Expression Analysis and in Vivo Translational Assays
in Protoplasts
Protoplasts were isolated from 2-week-old seedlings grown on B5 agar
media, and the preparations were transformed with 10 mg of the BiP:GFP
(Kim et al., 2001), RbcS:GFP (Lee et al., 2002), AALP:GFP (Sohn et al.,
2003), and GFP constructs using a polyethylene glycol–mediated proce-
dure previously described (Jin et al., 2001). Equal transformation effi-
ciencies were ensured measuring the ratio transformed:nontransformed
protoplast using confocal microscopy. For the streptomycin-treated
protoplasts, the GFP protein was expressed in protoplasts for 6 h,
followed by a treatment with 0 and 600 mg/mL of antibiotic for 12
additional hours. Protein extracts from protoplasts derived from the
different assays were prepared at different time points after transforma-
tion as described (Sohn et al., 2003). Briefly, protoplast incubation media
was collected after centrifugation at 6000g for 5 min. One milligram of
BSA was added to the media, which was then precipitated with 10%
trichloroacetic acid and followed by centrifugation at 10,000g for 5 min.
Precipitated protein aggregates were dissolved in 0.1 M NaOH. Prior to
the immunoblot analysis, the protoplast preparations were normalized
measuring total protein contents. Immunoblots were developed using a
Living Colors GFPMonoclonal antibody (Clontech) and an ECL detection
kit (Pierce Biotechnology).
Accession Numbers
Sequence data from this article can be found in the Arabidopsis Genome
Initiative or GenBank/EMBL databases under the following accession
numbers: RPL4A, locus At3g09630 cDNA, NM_001035586; protein,
NP_001030663; RPL4D, locus At5g02870 cDNA NM_001125687; pro-
tein, NP_001119159. T-DNA insertion lines SALK_130595 (rpl4a-2),
SALK_063782 (rpl4a-3), SALK_029203 (rpl4d-1), SALK_065625 (rpl4d-
2), and SALK_069239 (negative control for the antibiotic assays).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. The Secretion of the Vacuolar Markers in
rpl4a-1 Depends on Their Expression Levels and Developmental
Stage.
Supplemental Figure 2. Cellular Localization of Auxin Transporters in
rpl4a-1.
Supplemental Figure 3. The rpl4a and rpl4d Mutants Display Similar
Developmental Phenotypes.
Supplemental Figure 4. Histochemical Analysis of RPL4pro:GUS
Transgenic Lines.
Supplemental Figure 5. Double Heterozygous rpl4a-rpl4d Plants
Have Similar Phenotypes as the Single Mutants.
Supplemental Figure 6. The rpl4a Mutation Does Not Cause En-
hanced Unfolded Protein Response or Translational Defects in Vac-
uolar Trafficking Machinery.
Supplemental Table 1. Genetic Linkage Analysis Indicates That the
21-4 Phenotypes and the Protein Trafficking Defects Are Linked to the
Mutation in RPL4A.
Supplemental Table 2. Auxin-Related Phenotypes in Arabidopsis
Mutants with Altered Ribosome Biogenesis.
Supplemental Table 3. Specificity of the Antibiotic Response in rpl4
Mutants.
Supplemental Table 4. Primers Used in This Study.
ACKNOWLEDGMENTS
We thank Glenn Hicks, Marci Surpin, Julia Bailey-Serres, and Patty
Springer (University of California, Riverside) for their critical reading of
the manuscript. We also thank Mien Van De Ven, April Agee, Latasha
Johnson, and Alexander Michkov for technical assistance, L. Frigerio
(University of Warwick, UK) for the gift of the NTPPpro:RFP and the RFP:
CTPPpha marker lines, and I. Hwang (Pohang University, Korea) for the
gift of the BiP:GFP, GFP, RbcS:GFP, and AALP:GFP constructs. We
thank the anonymous reviewers for their useful comments and sugges-
tions. This work was funded by Department of Energy, Division of
Energy Biosciences, Grant DE-FG03-02ER15295/A000 (to N.V.R.) and
by the Spanish Fulbright postdoctoral fellowship MEC-FU-0248-2006
(to A.R.).
Received April 29, 2009; revised December 7, 2009; accepted December
19, 2009; published January 8, 2010.
REFERENCES
Ahmed, S., Rojo, E., Kovaleva, V., Venkataraman, S., Dombrowski,
J., Matsuoka, K., and Raikhel, N. (2000). The plant vacuolar sorting
receptor AtELP is involved in transport of NH(2)-terminal propeptide-
containing vacuolar proteins in Arabidopsis thaliana. J. Cell Biol. 149:
1335–1344.
Baerenfaller, K., Grossmann, J., Grobei, M., Hull, R., Hirsch-Hoffmann,
M., Yalovsky, S., Zimmermann, P., Grossniklaus, U., Gruissem,
156 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

W., and Baginsky, S. (2008). Genome-scale proteomics reveals
Arabidopsis thaliana gene models and proteome dynamics. Science
320: 938–941.
Ban, N., Nissen, P., Hansen, J., Moore, P., and Steitz, T. (2000). The
complete atomic structure of the large ribosomal subunit at 2.4 A
resolution. Science 289: 905–920.
Barakat, A., Szick-Miranda, K., Chang, I., Guyot, R., Blanc, G.,
Cooke, R., Delseny, M., and Bailey-Serres, J. (2001). The organi-
zation of cytoplasmic ribosomal protein genes in the Arabidopsis
genome. Plant Physiol. 127: 398–415.
Bassham, D., and Raikhel, N. (2000). Unique features of the plant
vacuolar sorting machinery. Curr. Opin. Cell Biol. 12: 491–495.
Carroll, A., Heazlewood, J., Ito, J., and Millar, A. (2008). Analysis of
the Arabidopsis cytosolic ribosome proteome provides detailed in-
sights into its components and their post-translational modification.
Mol. Cell. Proteomics 7: 347–369.
Carter, C., Pan, S., Zouhar, J., Avila, E., Girke, T., and Raikhel, N.
(2004). The vegetative vacuole proteome of Arabidopsis thaliana
reveals predicted and unexpected proteins. Plant Cell 16: 3285–3303.
Chang, I., Szick-Miranda, K., Pan, S., and Bailey-Serres, J. (2005).
Proteomic characterization of evolutionarily conserved and vari-
able proteins of Arabidopsis cytosolic ribosomes. Plant Physiol.
137: 848–862.
Craddock, C., Hunter, P., Szakacs, E., Hinz, G., Robinson, D., and
Frigerio, L. (2008). Lack of a vacuolar sorting receptor leads to non-
specific missorting of soluble vacuolar proteins in Arabidopsis seeds.
Traffic 9: 408–416.
Curtis, M., and Grossniklaus, U. (2003). A gateway cloning vector set
for high-throughput functional analysis of genes in planta. Plant
Physiol. 133: 462–469.
Degenhardt, R., and Bonham-Smith, P. (2008). Arabidopsis ribosomal
proteins RPL23aA and RPL23aB are differentially targeted to the
nucleolus and are desperately required for normal development. Plant
Physiol. 147: 128–142.
Galweiler, L., Guan, C., Muller, A., Wisman, E., Mendgen, K.,
Yephremov, A., and Palme, K. (1998). Regulation of polar auxin
transport by AtPIN1 in Arabidopsis vascular tissue. Science 282:
2226–2230.
Giavalisco, P., Wilson, D., Kreitler, T., Lehrach, H., Klose, J., Gobom,
J., and Fucini, P. (2005). High heterogeneity within the ribosomal
proteins of the Arabidopsis thaliana 80S ribosome. Plant Mol. Biol. 57:
577–591.
Goh, T., Uchida, W., Arakawa, S., Ito, E., Dainobu, T., Ebine, K.,
Takeuchi, M., Sato, K., Ueda, T., and Nakano, A. (2007). VPS9a, the
common activator for two distinct types of Rab5 GTPases, is essential
for the development of Arabidopsis thaliana. Plant Cell 19: 3504–3515.
Hunter, P., Craddock, C., Di Benedetto, S., Roberts, L., and Frigerio,
L. (2007). Fluorescent reporter proteins for the tonoplast and the
vacuolar lumen identify a single vacuolar compartment in Arabidopsis
cells. Plant Physiol. 145: 1371–1382.
Ito, T., Kim, G., and Shinozaki, K. (2000). Disruption of an Arabidopsis
cytoplasmic ribosomal protein S13-homologous gene by transposon-
mediated mutagenesis causes aberrant growth and development.
Plant J. 22: 257–264.
Jefferson, R., Kavanagh, T., and Bevan, M. (1987). GUS fusions: Beta-
glucuronidase as a sensitive and versatile gene fusion marker in
higher plants. EMBO J. 6: 3901–3907.
Jin, J., Kim, Y., Kim, S., Lee, S., Kim, D., Cheong, G., and Hwang, I.
(2001). A new dynamin-like protein, ADL6, is involved in trafficking
from the trans-Golgi network to the central vacuole in Arabidopsis.
Plant Cell 13: 1511–1526.
Kleine-Vehn, J., Leitner, J., Zwiewka, M., Sauer, M., Abas, L.,
Luschnig, C., and Friml, J. (2008). Differential degradation of PIN2
auxin efflux carrier by retromer-dependent vacuolar targeting. Proc.
Natl. Acad. Sci. USA 105: 17812–17817.
Kim, D., Eu, Y., Yoo, C., Kim, Y., Pih, K., Jin, J., Kim, S., Stenmark, H.,
and Hwang, I. (2001). Trafficking of phosphatidylinositol 3-phosphate
from the trans-Golgi network to the lumen of the central vacuole in
plant cells. Plant Cell 13: 287–301.
Koiwa, H., et al. (2002). C-terminal domain phosphatase-like family
members (AtCPLs) differentially regulate Arabidopsis thaliana abiotic
stress signaling, growth, and development. Proc. Natl. Acad. Sci. USA
99: 10893–10898.
Koiwa, H., Bressan, R., and Hasegawa, P. (2006). Identification of
plant stress-responsive determinants in Arabidopsis by large-scale
forward genetic screens. J. Exp. Bot. 57: 1119–1128.
Komili, S., Farny, N., Roth, F., and Silver, P. (2007). Functional
specificity among ribosomal proteins regulates gene expression.
Cell 131: 557–571.
Laxmi, A., Pan, J., Morsy, M., and Chen, R. (2008). Light plays an
essential role in intracellular distribution of auxin efflux carrier PIN2 in
Arabidopsis thaliana. PLoS One 3: e1510.
Liu, Y., Mitsukawa, N., Oosumi, T., and Whittier, R. (1995). Efficient
isolation and mapping of Arabidopsis thaliana T-DNA insert junctions
by thermal asymmetric interlaced PCR. Plant J. 8: 457–463.
Lee, M., Min, M., Lee, Y., Jin, J., Shin, D., Kim, D., Lee, K., and
Hwang, I. (2002). ADP-ribosylation factor 1 of Arabidopsis plays a
critical role in intracellular trafficking and maintenance of endoplasmic
reticulum morphology in Arabidopsis. Plant Physiol. 129: 1507–1520.
Matsuoka, K., and Nakamura, K. (1999). Large alkyl side-chains of
isoleucine and leucine in the NPIRL region constitute the core of the
vacuolar sorting determinant of sporamin precursor. Plant Mol. Biol.
41: 825–835.
Miao, Y., Li, K., Li, H., Yao, X., and Jiang, L. (2008). The vacuolar
transport of aleurain-GFP and 2S albumin-GFP fusions is mediated by
the same pre-vacuolar compartments in tobacco BY-2 and Arabi-
dopsis suspension cultured cells. Plant J. 56: 824–839.
Nishimura, T., Wada, T., Yamamoto, K., and Okada, K. (2005). The
Arabidopsis STV1 protein, responsible for translation reinitiation, is
required for auxin-mediated gynoecium patterning. Plant Cell 17:
2940–2953.
O’Connor, M., Gregory, S., and Dahlberg, A. (2004). Multiple defects
in translation associated with altered ribosomal protein L4. Nucleic
Acids Res. 32: 5750–5756.
Oleinick, N. (1977). Initiation and elongation of protein synthesis in
growing cells: Differential inhibition by cycloheximide and emetine.
Arch. Biochem. Biophys. 182: 171–180.
Paciorek, T., Zazımalova, E., Ruthardt, N., Petrasek, J., Stierhof, Y.,
Kleine-Vehn, J., Morris, D., Emans, N., Jurgens, G., Geldner, N.,
and Friml, J. (2005). Auxin inhibits endocytosis and promotes its own
efflux from cells. Nature 435: 1251–1256.
Pan, J., Fujioka, S., Peng, J., Chen, J., Li, G., and Chen, R. (2009). The
E3 ubiquitin ligase SCFTIR1/AFB and membrane sterols play key
roles in auxin regulation of endocytosis, recycling, and plasma mem-
brane accumulation of the auxin efflux transporter PIN2 in Arabidopsis
thaliana. Plant Cell 21: 568–580.
Petricka, J., and Nelson, T. (2007). Arabidopsis nucleolin affects plant
development and patterning. Plant Physiol. 144: 173–186.
Pfaffl, M. (2001). A new mathematical model for relative quantification in
real-time RT-PCR. Nucleic Acids Res. 29: e45.
Pinon, V., Etchells, J., Rossignol, P., Collier, S., Arroyo, J.,
Martienssen, R., and Byrne, M. (2008). Three PIGGYBACK genes
that specifically influence leaf patterning encode ribosomal proteins.
Development 135: 1315–1324.
Robinson, D., Oliviusson, P., and Hinz, G. (2005). Protein sorting to the
storage vacuoles of plants: A critical appraisal. Traffic 6: 615–625.
Ribosomal Regulation of Vacuolar Sorting 157
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022

Rojo, E., Martın, R., Carter, C., Zouhar, J., Pan, S., Plotnikova, J.,
Jin, H., Paneque, M., Sanchez-Serrano, J., Baker, B., Ausubel, F.,
and Raikhel, N. (2004). VPEgamma exhibits a caspase-like activity
that contributes to defense against pathogens. Curr. Biol. 14: 1897–
1906.
Rojo, E., Sharma, V., Kovaleva, V., Raikhel, N., and Fletcher, J.
(2002). CLV3 is localized to the extracellular space, where it activates
the Arabidopsis CLAVATA stem cell signaling pathway. Plant Cell 14:
969–977.
Sanderfoot, A., Kovaleva, V., Bassham, D., and Raikhel, N. (2001).
Interactions between syntaxins identify at least five SNARE com-
plexes within the Golgi/prevacuolar system of the Arabidopsis cell.
Mol. Biol. Cell 12: 3733–3743.
Sanderfoot, A., Kovaleva, V., Zheng, H., and Raikhel, N. (1999). The
t-SNARE AtVAM3p resides on the prevacuolar compartment in
Arabidopsis root cells. Plant Physiol. 121: 929–938.
Sanmartın, M., Ordonez, A., Sohn, E., Robert, S., Sanchez-Serrano,
J., Surpin, M., Raikhel, N., and Rojo, E. (2007). Divergent functions
of VTI12 and VTI11 in trafficking to storage and lytic vacuoles in
Arabidopsis. Proc. Natl. Acad. Sci. USA 104: 3645–3650.
Sato, M., Nakamura, N., Ohsumi, Y., Kouchi, H., Kondo, M., Hara-
Nishimura, I., Nishimura, M., and Wada, Y. (1997). The AtVAM3
encodes a syntaxin-related molecule implicated in the vacuolar as-
sembly in Arabidopsis thaliana. J. Biol. Chem. 272: 24530–24535.
Shirakawa, M., Ueda, H., Shimada, T., Nishiyama, C., and Hara-
Nishimura, I. (2009). Vacuolar SNAREs function in the formation of
the leaf vascular network by regulating auxin distribution. Plant Cell
Physiol. 50: 1319–1328.
Sohn, E., Kim, E., Zhao, M., Kim, S., Kim, H., Kim, Y., Lee, Y., Hillmer,
S., Sohn, U., Jiang, L., and Hwang, I. (2003). Rha1, an Arabidopsis
Rab5 homolog, plays a critical role in the vacuolar trafficking of
soluble cargo proteins. Plant Cell 15: 1057–1070.
Sohn, E., Rojas-Pierce, M., Pan, S., Carter, C., Serrano-Mislata, A.,
Madueno, F., Rojo, E., Surpin, M., and Raikhel, N. (2007). The shoot
meristem identity gene TFL1 is involved in flower development and
trafficking to the protein storage vacuole. Proc. Natl. Acad. Sci. USA
104: 18801–18806.
Spitzer, C., Reyes, F., Buono, R., Sliwinski, M., Haas, T., and Otegui,
M. (2009). The ESCRT-related CHMP1A and B proteins mediate
multivesicular body sorting of auxin carriers in Arabidopsis and are
required for plant development. Plant Cell 21: 749–766.
Surpin, M., and Raikhel, N. (2004). Traffic jams affect plant develop-
ment and signal transduction. Nat. Rev. Mol. Cell Biol. 5: 100–109.
Teale, W., Paponov, I., and Palme, K. (2006). Auxin in action: Signal-
ling, transport and the control of plant growth and development. Nat.
Rev. Mol. Cell Biol. 7: 847–859.
Ulmasov, T., Murfett, J., Hagen, G., and Guilfoyle, T. (1997). Aux/IAA
proteins repress expression of reporter genes containing natural and
highly active synthetic auxin response elements. Plant Cell 9: 1963–
1971.
Van Lijsebettens, M., Vanderhaeghen, R., De Block, M., Bauw, G.,
Villarroel, R., and Van Montagu, M. (1994). An S18 ribosomal
protein gene copy at the Arabidopsis PFL locus affects plant
development by its specific expression in meristems. EMBO J. 13:
3378–3388.
Vitale, A., and Raikhel, N. (1999). What do proteins need to reach
different vacuoles? Trends Plant Sci. 4: 149–155.
Volarevic, S., Stewart, M., Ledermann, B., Zilberman, F., Terracciano,
L., Montini, E., Grompe, M., Kozma, S., and Thomas, G. (2000).
Proliferation, but not growth, blocked by conditional deletion of 40S
ribosomal protein S6. Science 288: 2045–2047.
Weijers, D., Franke-van Dijk, M., Vencken, R., Quint, A., Hooykaas,
P., and Offringa, R. (2001). An Arabidopsis Minute-like phenotype
caused by a semi-dominant mutation in a RIBOSOMAL PROTEIN S5
gene. Development 128: 4289–4299.
Wool, I. (1996). Extraribosomal functions of ribosomal proteins. Trends
Biochem. Sci. 21: 164–165.
Yao, Y., Ling, Q., Wang, H., and Huang, H. (2008). Ribosomal proteins
promote leaf adaxial identity. Development 135: 1325–1334.
Zheng, H., von Mollard, G., Kovaleva, V., Stevens, T., and Raikhel, N.
(1999). The plant vesicle-associated SNARE AtVTI1a likely mediates
vesicle transport from the trans-Golgi network to the prevacuolar
compartment. Mol. Biol. Cell 10: 2251–2264.
Zouhar, J., Rojo, E., and Bassham, D. (2009). AtVPS45 is a positive
regulator of the SYP41/SYP61/VTI12 SNARE complex involved in
trafficking of vacuolar cargo. Plant Physiol. 149: 1668–1678.
158 The Plant Cell
Dow
nloaded from https://academ
ic.oup.com/plcell/article/22/1/143/6094821 by guest on 24 January 2022