author's personal copy - cpb-us-w2.wpmucdn.com · author's personal copy the human...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The human amygdala plays a stimulus specific role in the detection of novelty
Nicholas L. Balderston a, Doug H. Schultz a, Fred J. Helmstetter a,b,⁎a Department of Psychology, University of Wisconsin-Milwaukee, Milwaukee, WI 53211, USAb Department of Neurology, Medical College of Wisconsin, Milwaukee, WI 53226, USA
a b s t r a c ta r t i c l e i n f o
Article history:Received 26 September 2010Revised 7 January 2011Accepted 12 January 2011Available online 20 January 2011
Keywords:HumanAmygdalaNoveltyEmotionFacesfMRI
The primary focus of research on the amygdala has been on the detection of and response to emotion but theamygdala also sometimes responds to new or unexpected stimuli without specific emotional content. Verylittle is currently known about why the amygdala responds to some new stimuli but not to others. Here weinvestigated the conditions that are necessary and sufficient for the expression of novelty specific amygdalaresponses by presenting novel and repeated images to human participants and varying the content of theseimages while measuring blood-oxygenation level dependent (BOLD) responses. In Experiment 1 wepresented novel and repeated emotional and neutral images. Both emotional and neutral images of humansevoked more amygdala activity when novel than when repeated. In Experiment 2 we presented novel andrepeated images of humans and scenes. Images of humans but not scenes evoked more amygdala activitywhen novel than when repeated. Our results suggest that the amygdala plays a stimulus-specific role in thebrain's novelty detection network. Surprisingly, emotion was not necessary for amygdalar novelty responses,but the presence of a human representation was important. Amygdala responses evoked by novel faces mayreflect our need to use others' faces as clues for important events in the environment.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Activity in the brain's novelty detection network is thought torepresent an early stage in memory encoding, focusing attention onunexpected stimuli or events (Tulving et al., 1996). The amygdalaplays an important role in the formation of new memories foremotional events (Canli et al., 2000). Although often overlooked innovelty detection studies (Daselaar et al., 2006; Menon et al., 2000;Tulving et al., 1996; Yamaguchi et al., 2004), the amygdala sometimesresponds to novel stimuli much like other regions (i.e. hippocampus,parahippocampal gyrus). For example, Schwartz and colleaguespresented blocks of faces and found that the amygdala respondedmaximally when the faces in a given block were presented only once(Schwartz et al., 2003). In contrast, Yamaguchi and colleagues showedthat the hippocampus but not the amygdala was activated by novelpresentations of animals, buildings and landscapes, suggesting thatnovelty per se does not drive amygdala responses (Yamaguchi et al.,2004).
Although larger blood-oxygenation level dependent (BOLD)responses have been observed in the human amygdala for stimulithat are novel to the observer, we still lack a basic understanding ofthe conditions that are necessary and sufficient for such responses.We hypothesized that novelty-specific amygdala responses are notevoked by all stimuli, but are dependent on characteristics of thenovel stimulus.
We used functional magnetic resonance imaging (fMRI) todetermine which stimulus characteristics are most important fornovelty-specific amygdala responses. We presented novel andrepeated images and systematically varied the content of theseimages based on the previously established framework of amygdalafunctions. In Experiment 1 we sought to determine if emotionalcontent played an important role in amygdalar novelty responding. InExperiment 2 we sought to determine if human representations werenecessary for novelty-specific amygdala responses. Finally, weinvestigated the temporal properties of novelty-specific amygdalarresponses by comparing the responses evoked by novel and repeatedstimuli across trials.
Materials and methods
Participants
Fifty-three neurologically healthy undergraduate students (Age:M=20.78, SD=2.90) at the University of Wisconsin-Milwaukeeparticipated in this experiment and received $20 for participation, as
NeuroImage 55 (2011) 1889–1898
Abbreviations: NE, novel emotional image; NN, novel neutral image; RE, repeatedemotional image; RN, repeated neutral image; NH, novel human image; NS, novel sceneimage; RH, repeated human image; RS, repeated scene image; IAPS, InternationalAffective Picture System; ITI, intertrial interval; SCL, skin conductance level; SCR, skinconductance response; ROI, region of interest; FFA, fusiform face area; PPA,parahippocampal place area.⁎ Corresponding author at: 2441 E. Hartford Ave., Garland Hall 224, Milwaukee, WI
53212, USA. Fax: +1 414 229 5219.E-mail address: [email protected] (F.J. Helmstetter).
1053-8119/$ – see front matter © 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.neuroimage.2011.01.034
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r.com/ locate /yn img

Author's personal copy
well as extra credit in their psychology classes and a picture of theirbrain. Thirty-three were female. Three participants were excludedfrom the study because of computer/recording failures. Ten partici-pants were excluded because of excessive head motion. Of theremaining participants, 20 individuals (14 female) participated inExperiment 1, and 20 individuals (12 female) participated inExperiment 2. All participants gave informed consent, and theprotocol was approved by the Institutional Review Boards forhuman subject research at the University of Wisconsin-Milwaukeeand the Medical College of Wisconsin.
Procedure
Stimuli were presented using the software package Presentation(Neurobehavioral Systems, Inc., Albany, CA), using aDell laptop (model:Inspiron 9300, Dell Inc., Red Rock, TX). All participants saw a series of20 eight second presentations of novel and repeated images whilepositioned in the fMRI scanner. Participants viewed the stimuli via aback projection system with prism glasses mounted to the head coil.
In the novel conditions, we presented a series of five differentimages, where each image was presented once. In the repeatedconditions, we presented a single image six times. For the repeatedconditions we included only trials where the image had been repeated.Therefore, the initial presentation of these stimuli was counted in therespective novel categories. In total, there were five trials of eachstimulus type, corresponding to the experimental categories for eachexperiment. Repeated stimuli were counterbalanced across subjects.
In Experiment 1, we manipulated the emotional content of thestimuli by presenting emotional (novel = NE; repeated = RE) andneutral (novel = NN; repeated = RN) images of humans from theInternational Affective Picture System (IAPS) database (Lang et al.,2005). See Fig. 1 for design summary. Emotional images depictedmutilated human bodies. Neutral images depicted healthy individualswith their gaze directed toward the camera. Normative ratings of theimages confirm that the emotional images were rated as morearousing and more emotionally negative than the neutral images (seeSupplementary Table 3 for picture codes and normative ratings). Each
picture was presented centrally, and presentations were separated bya 20 second average variable intertrial interval (ITI; ±4 s).
In Experiment 2, we presented images of humans (novel = NH;repeated= RH) and scenes (novel = NS; repeated= RS), all of whichwere emotionally neutral (Fig. 2). This experiment was a partialreplication of Experiment 1 because our images of humans were thesame images used for the neutral images above. The scenes werechosen from the same picture database (Lang et al., 2005) andequated for arousal and valence ratings. As in Experiment 1, eachpicture was presented centrally, and presentations were separated bya 20 second average variable intertrial interval (ITI; ±4 s).
Prior to the experiment participants underwent a procedurewhere they rated the intensity of an electrical stimulus as part of aseparate study. In order to ensure that the participants were attendingto the stimuli, they were asked to continuously rate their expectancyof receiving this stimulation by manipulating a dial (SupplementaryFig. 1). However at no time during the actual experiment did theindividuals receive the stimulation. We also measured skin conduc-tance responses throughout the experiments (see SupplementaryTables 4 and 5 and Supplementary Fig. 2).
MRI
We conducted whole brain imaging using a 3T short bore GE SignaExcite MRI system. Functional images were acquired using a T2*weighted gradient-echo, echoplanar pulse sequence. Contiguous fourmillimeter sagittal slices (TR=2 s; TE=25 ms; field of view=24 cm;flip angle=90°) were collected during the experiment. Two hundredand ninety whole brain images were collected. High resolutionspoiled gradient recalled (SPGR) acquisition images were collectedto serve as a three-dimensional anatomical map for the functionalimages.
MRI segmentation
Subcortical segmentation was performed using the Freesurfersoftware package, which is freely available online and has beendescribed previously (Fischl et al., 2002, 2004). Freesurfer generated
Fig. 1. In Experiment 1 we presented emotional and neutral images while measuringBOLD activity. (a) Images were presented sequentially in an event related design. (b,c)The images are representative of the emotional (b) and neutral (c) images shown toparticipants. All participants saw 5 presentations of novel emotional (NE) and 5presentations of novel neutral (NN) images indicated by the orange and light blueoutlines, respectively. In addition all saw 5 repetitions of one emotional (RE) and oneneutral (RN) image, shown in red and blue respectively. +The initial presentation ofthe repeated stimuli was counted as novel.
Fig. 2. In Experiment 2 we presented images of humans and scenes while measuringBOLD activity. (a) As in Experiment 1, images were presented sequentially in an eventrelated design. (b,c) The images are representative of the human images (b) and sceneimages (c) shown to participants. All participants saw 5 presentations of novel human(NH) and 5 presentations of novel scene (NS) images, which are indicated by the lightblue and light green outlines, respectively. In addition all saw 5 repetitions of onehuman (RH) and one scene (RS) image, shown with dark blue and dark green outlines,respectively. +The initial presentation of the repeated stimuli was counted as novel.
1890 N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
volumes were then realigned to native space using the Analysis ofFunctional NeuroImages software package (AFNI). These realignedvolumes were then manually edited to conform to previouslydescribed standards (Morey et al., 2009).
Functional imaging data acquisition
Functional imaging data were reconstructed and processed usingAFNI (Cox, 1996). fMRI data were passed through motion correctionand edge detection algorithms, then registered to the fifth image inthe timeseries. Raw fMRI data were manually inspected for large headmovements. Images that contained discrete head movements werecensored, and participants showing excessive movement (greaterthan 2 mm displacement or more than 5 instances of discrete headmovements) were excluded from further analyses. Head motion anddial movement regressors were included in the analysis as regressorsof no interest. Timeseries data were deconvolved with stimuluscanonicals using a least squares procedure, to yield average impulseresponse functions (IRFs).
Functional imaging data analysis
For whole brain analyses SPGR images were manually warped intoTalairach space using anatomical markers. Images one through five ofthe IRFs were used to calculate percent area under the curve (%AUC).The %AUC maps were then registered to Talairach space andresampled to 1 mm isotropic voxels using linear interpolation. Imageswere then blurred using a 4 mm full-width at half-maximumGaussian kernel. The resulting maps were used in the group levelanalyses. We used cluster thresholding (Forman et al., 1995) tocorrect for multiple comparisons across the voxels in the whole brainvolume (p=0.005; rmm=2; xyz=1; volume=228 μL; correctedα=0.05).
For the ROI analyses, image three from the IRF was registered tothe unwarped SPGR data on a subject-by-subject basis and resampledto 1 mm isotropic voxels using linear interpolation. The images usedfor the ROI analyses were not warped or blurred, in order to forego thedistortion caused by these procedures. Because voxelwise data werenot used in the group-level analyses, these steps were unnecessary.We chose an alpha level of 0.05 for all analyses.
Skin conductance responses
Skin conductance level (SCL) was recorded via two surface cupelectrodes (silver/silver chloride, 8 mm diameter, Biopac modelEL258-RT, Goleta, CA) filled with electrolyte gel (Signa Gel, Parkerlaboratories Fairfield, NJ) attached to the bottom of the participant'sleft foot approximately 2 cm apart, and sampled at 200 Hz throughoutthe experiment. We sampled SCL during the 8 second stimulus periodand the preceding two second baseline period. Raw values for eachtrial were normalized to that trial's average baseline SCL andexpressed as a percent change from that baseline value. SCRtimecourse data were obtained by averaging the percent changevalues across participants and across trials at each timepoint duringthe stimulus period. Statistical tests were computed at each timepointwith an alpha level of 0.05. Monte Carlo simulations were used tocorrect for multiple comparisons across timepoints (see Supplemen-tary Table 5 and Supplementary Fig. 2).
SCR Monte Carlo simulations
Monte Carlo simulations were conducted to correct for multiplecomparisons carried out on SCRs, in a manner similar to clusterthresholding of fMRI data. We generated 200,000 strings of randomp-values thatwere the same length as the SCR samples (2000 values),using a random number generator. We then counted the number of
sequential significant p-values (Tp=timepoint p-value), using a time-point α (Tα) of 0.05, and determined the likelihood that a given stretchof sequential significant p-values could have arisen due to chance alone.We found that less than 0.002% of the simulations yielded samplelengths (length of sequential significant p-values) greater than or equalto 7 timepoints in length, which is equivalent to 35 ms in duration.Therefore to correct for multiple comparisons, we thresholded our datausing a combination of individual timepoint p-value (Tα=0.05) andsample length (λpN7; see Supplementary Table 4).
Electrical stimulation
As part of another experiment, participants were given presenta-tions of an electrical stimulation before the experiment began.Electrical stimulation was administered via an AC (60 Hz) source(Contact Precision Instruments, Model SHK1, Boston, MA) throughtwo surface cup electrodes (silver/silver chloride, 8 mm diameter,Biopac model EL258-RT, Goleta, CA) filled with electrolyte gel (SignaGel, Parker laboratories Fairfield, NJ). The electrodes were placed onthe skin over the subject's right tibial nerve over the right medialmalleolus. Participants were given several half-second presentationsof the shock. They rated the shock on a scale from zero (no sensation)to ten (painful but tolerable). Intensity was increased gradually in mAuntil participants rated the sensation as a ten.
Shock expectancy
In order to ensure that the participants attended to the stimuli,they continuously rated their expectancy of receiving the electricalstimulation. To do so, participants controlled a visual analog scale onthe computer screen using dial. The analog scale was anchored with 0and 100. Participants were instructed to move the cursor to 0 if theywere absolutely sure that they would not receive an electricalstimulation, to move the cursor to 100 if they were absolutely surethat they would receive a stimulation, and to keep the cursor near 50if they felt like there was an equal probability of receiving or notreceiving a stimulation. Responses were recorded throughout theexperiment and sampled at 40 Hz (see Supplementary Fig. 1). Analpha level of 0.05 was used for all analyses.
Results
Novelty-specific BOLD responses in the amygdala and hippocampus arenot dependent on emotion
We began by investigating emotion as a potential mediating factorfor two reasons. First, the amygdala is important for the perception ofand response to emotional stimuli (Adolphs et al., 1994; Cheng et al.,2003; Williams et al., 2001). Second, amygdala activity at encodingcorrelates with subsequent memory for emotional scenes, suggestingthat amygdala activity may help facilitate memory for emotionalevents (Canli et al., 2000). Amygdalar novelty responses may facili-tate the encoding of emotional stimuli, which would provide a linkbetween amygdala activity and memory performance. To test thiswe presented novel and repeated emotional and neutral imageswhile measuring BOLD activity.
Based on the novelty/encoding hypothesis and the link betweenamygdala activity and facilitated recognition for emotional scenes, wepredicted that there would be larger amygdala responses to novelemotional images than repeated emotional images, but similarmagnitude amygdala responses to novel and repeated neutral images.To determine the effects of novelty and emotion on brain activity, weperformed a mixed effects ANOVA on BOLD intensity using noveltyand emotion as fixed factors and subject as a random factor. The moststriking finding of the whole brain BOLD analysis was a robust maineffect for novelty bilaterally in the amygdala and hippocampus, but no
1891N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
main effect for emotion and no novelty by emotion interaction inthese structures (Figs. 3a–b, see Supplementary Table 1 for full list ofactivations).
To characterize these effects, we sampled the stimulus evokedBOLD responses within the amygdala and hippocampus. To indepen-dently sample activity within these two structures, we createdanatomical regions of interest (ROIs; see Figs. 3c–f) for the amygdalaand hippocampus on a subject-by-subject basis, using an automatedsubcortical segmentation algorithm (Fischl et al., 2002, 2004). Theseautomatically generated anatomical ROIs were then manually editedto conform to previously described standards (Morey et al., 2009). Wesampled the BOLD data from the final 2 s of the stimulus period(seconds 7–8), which corresponded to the overall peak of the impulseresponse functions (see Figs. 4a and c). We performed a novelty byemotion by laterality ANOVA on these values, with left/righthemisphere as a repeated measure. Consistent with the whole brainanalysis, novel stimuli evoked a robust BOLD response in theamygdala and hippocampus whether they are emotional or neutral(Novelty main effect: Amygdala, F(1,76)=19.71, p=3.02E−5;Hippocampus, F(1,76)=15.32, p=1.97E−4; see Fig. 4). In addition,emotional stimuli evoked more activity than neutral stimuli in bothstructures (Emotion main effect: Amygdala, F(1,76)=5.18, p=0.025;Hippocampus, F(1,76)=9.61, p=0.003). However, the novelty effectsare similar for emotional and neutral stimuli (Novelty×Emotioninteraction: Amygdala, F(1,76)=0.19, p=0.667; Hippocampus, F(1,76)b0.01, p=0.986). Also, this pattern of responses is consistentacross hemispheres (Laterality main effect, Amygdala, F(1,76)=1.24,p=0.269; Hippocampus, F(1,76)=0.01, p=0.92).
Novel faces but not novel scenes activate the amygdala
In Experiment 1, we showed that the amygdala responds to novelstimuli whether they are emotional or neutral. In contrast, severalprevious novelty studies have failed to observe significant effects for theamygdala (Daselaar et al., 2006; Menon et al., 2000; Tulving et al., 1996;Yamaguchi et al., 2004). This lack of effect may be due to theheterogeneous collection of stimuli used in these previous studies (e.g.,
words, animals, landscapes, etc.). All of our stimuli depicted humans.Similarly, other studies showing novelty-evoked BOLD responses in theamygdala used images of humans as well (Schwartz et al., 2003;Wrightet al., 2003). Thus, the presenceof humans/faces in the stimulusmaybe akey stimulus attribute, necessary for the expressionof amygdalarnoveltyresponses, although for a recent counterpoint see Blackford et al. (2010).To test this we presented novel and repeated images of humans andscenes while measuring BOLD activity.
To determine the effects of novelty and picture content on brainactivity, we performed a mixed effects ANOVA on BOLD intensityusing novelty and picture content as fixed factors and subject as arandom factor (see Supplementary Table 2 for full list of activations).In this whole brain BOLD analysis we did not observe a significantnovelty by picture content interaction in the amygdala or hippocam-pus. But because we had a priori hypotheses about the amygdala andhippocampus and because we used an anatomical ROI approach inExperiment 1, we continued that approach here.
We created anatomical ROIs for the amygdala and hippocampuson a subject-by-subject basis, using the same process as the firstexperiment. We then sampled the BOLD data within the amygdalaand hippocampus using these ROIs (seconds 7–8; see Figs. 5a and c).Next we performed a novelty by picture content by laterality ANOVAon these values, with left/right hemisphere as a repeated measure.Fig. 5 shows that novel humans evoke larger magnitude BOLDresponses in the amygdala than repeated humans but novel scenesevoke similar magnitude BOLD responses as repeated scenes(Novelty×Picture content interaction: F(1,76)=5.19, p=0.026;post hoc Bonferroni-corrected t-tests: Novel vs. Repeated humans,t(38)=2.99, p=0.005; Novel vs. Repeated scenes, t(38)=0.04,p=0.970). In contrast, novel humans and novel scenes both evokelarger magnitude hippocampal responses than repeated humansand scenes (Novelty main effect: F(1,76)=11.75, p=0.001),although the effect size may be larger for humans (Novelty×Picturecontent interaction: F(1,76)=3.71, p=0.058). This pattern ofresponses was consistent across hemispheres for both structures(Laterality main effect: Amygdala, F(1,76)=1.52, p=0.222; Hip-pocampus, F(1,76)=0.01, p=0.940).
Fig. 3. Whole brain BOLD analysis reveals significant main effect for novelty bilaterally in the amygdala and hippocampus. (a,b) Axial slices showing main effect for novelty inamygdala and hippocampus (data from Experiment 1). Colors indicate size of F-statistic depicted on brain slice and correspond to the colors on the scale to the left. Arrows indicateareas where clusters overlap with the amygdala (pink) and the hippocampus (green). (c–f) We used automated subcortical segmentation to identify the amygdala (pink) andhippocampus (green) on a subject-by-subject bases. (c–f) Representation of a sample subject's segmentation in coronal (c), axial (d), sagittal (e), and 3-D views (f).
1892 N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
Neural activity in the amygdala and hippocampus decreases after asingle stimulus presentation
The results from these two experiments demonstrate novelty-specific responses in the amygdala and hippocampus, consistent withseveral previous studies (Schwartz et al., 2003; Wright et al., 2003).Furthermore, they strongly suggest that amygdalar novelty responsesare specific to stimuli that contain biologically relevant information,such as images of conspecifics. However, because responses wereaveraged across several stimulus repetitions, it is difficult to determinehow the BOLD responses decrease across repeated stimulus presenta-tions. If the results are driven by novelty, then the magnitude of theBOLD response should not depend on the number of times a givenstimulus is repeated. That is, we would expect to see a large initialresponse followed by an immediate decrease to a stable baseline. If theresults are driven by a more gradual process like habituation, then themagnitude of the BOLD response should dependon the number of timesa given stimulus is repeated. That is, we would expect to see a largeinitial response, followed by a gradual decay toward baseline (Grovesand Thompson, 1970). Habituation is commonly evoked to explaindecreases in the magnitude of amygdala responses, but is rarely testedspecifically (Britton et al., 2008; Buchel et al., 1998; Fischer et al., 2003).To distinguish between effects driven by novelty and effects driven byhabituation,we reexamined the novelty effect from Experiment 1whileexplicitly accounting for the number of stimulus repetitions.
To understand the temporal characteristics of the novelty evokedresponses in the amygdala and hippocampus, we reanalyzed the data
from the Experiment 1 to determine the BOLD response on each of thetrials. Because we did not observe a novelty by emotion interaction inthe previous analyses, we collapsed across emotion. Thus we wereable to deconvolve the single subject BOLD data with separatecanonicals for each trial, using the same least squares procedure asbefore. This allowed us to determine the effects of trial number on theBOLD response in the amygdala and hippocampus. To do this weperformed an ANOVA on the BOLD activity (seconds 7–8) within theanatomical amygdala and hippocampal ROIs. The most importantthing to understand about this analysis is that the initial presentationof the to-be-repeated image is counted as novel. This is importantbecause the initial data point for the repeated condition is actually thesecond presentation (first repetition) of the repeated image. Again, ifour effects are driven by a rapid process, we should see a main effectfor novelty but no novelty by trial interaction. If our effects are drivenby a more gradual habituation process, we should see a smalldifference on the beginning trials followed by a larger difference onsubsequent trials, which would yield a Novelty×Trial interaction.Novel stimuli evoke a larger magnitude BOLD response in theamygdala (Novelty main effect: F(1,190)=29.58, p=1.63E−7; seeFig. 6) and the hippocampus (Novelty main effect: F(1,190)=26.33,p=7.08E−7). In both structures the novelty effect is consistent acrosshemispheres (Laterality main effect: Amygdala, F(1,190)=1.85,p=0.176; Hippocampus, F(1,190)=0.02, p=0.883) and presenteven on early trials. In the amygdala this response seems to beconsistent across trials (Trial main effect: F(4,190)=1.04, p=0.390;Novelty×Trial interaction: F(4,190)=0.47, p=0.790). However
Fig. 4. Novel emotional and novel neutral stimuli drive BOLD activity in the amygdala and hippocampus. (a,c) Line graphs represent BOLD timecourse in the amygdala (a) andhippocampus (c) during Experiment 1. (b,d) Bar graphs represent the percent signal change in the amygdala (b) and hippocampus (d) during the last 2 s of the stimulus period. Alldata points represent mean±SEM. (NE = novel emotional, RE = repeated emotional, NN = novel neutral, RN = repeated neutral).
1893N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
there may be a trend towards habituation to the novel stimuli in thehippocampus (Trial main effect: F(4,190)=1.03, p=0.393; Noveltyby Trial interaction: F(4,190)=2.27, p=0.063).
Humans and scenes evoke BOLD responses in distinct cortical areas
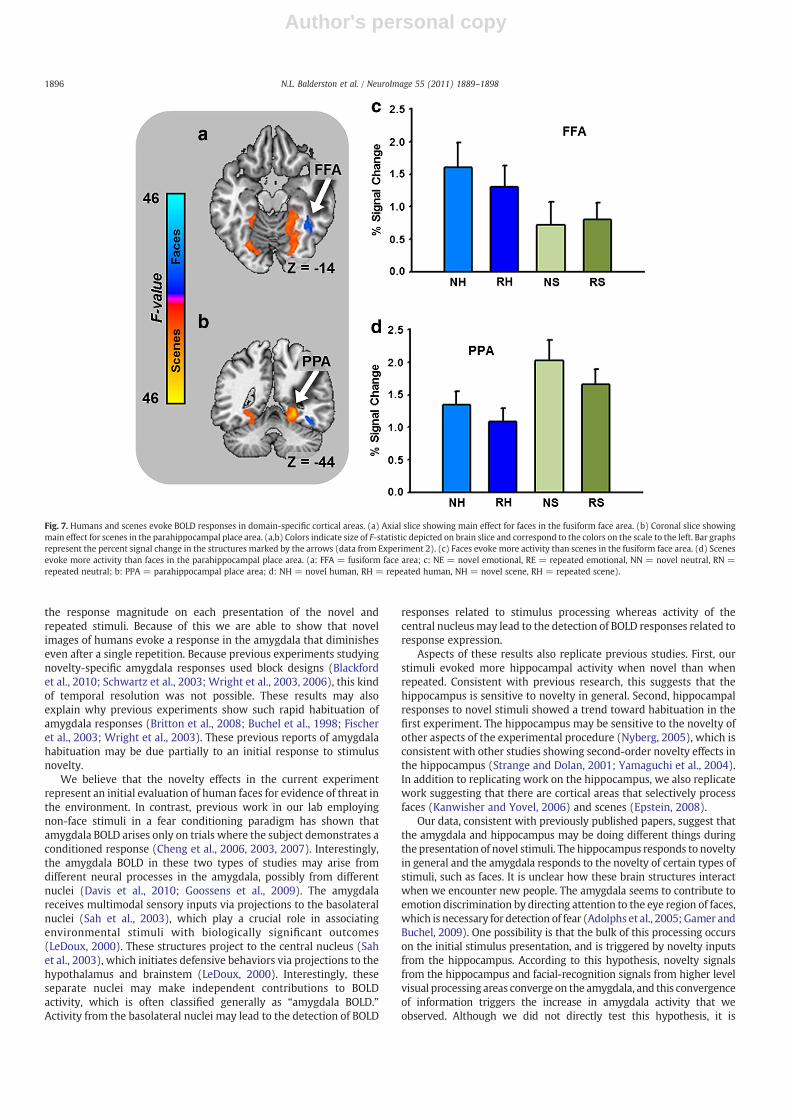
In Experiment 2, we presented images of humans and images ofscenes. Based on previous work (Epstein, 2008; Kanwisher and Yovel,2006), we expected to find activations in areas of the cortex thatcorresponded to the domains of the different image types. Specifically,we expected to observe larger responses to faces in the fusiform facearea (FFA) and larger responses to scenes in the parahippocampalplace area (PPA). As a manipulation check we performed anexploratory whole brain mixed effects ANOVA on BOLD intensitywith novelty and picture content as fixed factors and subject as arandom factor, using the methods from the first experiment. Asexpected we observed significant activations in FFA and PPA. Thepattern of means in these areas is consistent with previousobservations. That is, faces evoke larger magnitude BOLD responsesthan scenes in FFA and scenes evoke larger magnitude BOLDresponses than faces in PPA (see Fig. 7 and Supplementary Table 2).
Behavioral results
Shock expectancyAs part of a separate experiment, participants received presenta-
tions of an electrical stimulation prior to the data that were collected.
To ensure that participants were attending to the stimuli, we toldthem to continuously rate their expectancy of receiving an electricalstimulation using an onscreen visual analog scale.
In the first experiment we performed a novelty by emotion by trialANOVA on these values, with trial as a repeated measure. Participantsindicated that the stimulation was more likely to occur duringpresentations of the emotional stimuli than during the neutral stimuli(Emotion main effect: F(1,76)=13.31; p=0.0005; see SupplementaryFig. 1). Participants showed a decrease in expectancy across trials (Trialmain effect: F(4,76)=2.64; p=0.034), which seemed to be smaller foremotional stimuli (Emotion by Trial interaction: F(4,76)=2.53;p=0.041). Results suggest that participants are more likely to expectaversive outcomes when presented with emotional pictures.
In the secondexperimentweperformed anovelty bypicture contentby trial ANOVA on these values, with trial as a repeated measure.Participants indicated that the stimulation was more likely to occurduring presentations of the novel stimuli than during the repeatedstimuli (Noveltymain effect: F(1,76)=21.72; p=1.3E−5). Participantsshowed a decrease in expectancy across trials (F(4,76)=3.75;p=0.005), which seemed to be similar for both stimulus types(F(4,76)=1.065; p=0.374).
Skin conductance responsesIn the first experiment we performed ANOVAs at each timepoint of
the stimulus period using novelty and emotion as factors. Weobserved a significant novelty by emotion interaction during thesecond half of the stimulus period (see Supplementary Table 5 and
Fig. 5. Novel faces but not novel scenes drive amygdala BOLD. (a,c) Line graphs represent BOLD timecourse in the amygdala (a) and hippocampus (c) during Experiment 2. (b,d) Bargraphs represent the percent signal change in the amygdala (b) and hippocampus (d) during the last 2 s of the stimulus period. All data points represent mean±SEM. (NH= novelhuman, RH = repeated human, NH = novel scene, RH = repeated scene).
1894 N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
Supplementary Fig. 2a). Results suggest that autonomic responsesevoked by emotional stimuli habituate with repeated stimuluspresentations.
In the second experiment we performed ANOVAs at eachtimepoint of the stimulus period using novelty and picture contentas factors.We observed a significant main effect for novelty during thesecond half of the experiment (see Supplementary Table 5 andSupplementary Fig. 2b). Results suggest that neutral stimuli generallyevoke autonomic responses that habituate with repeated stimuluspresentations. However, results from the first experiment suggest thatneutral stimuli fail to evoke autonomic responses when presented inthe same context as emotional stimuli.
Discussion — the amygdala plays a stimulus specific role in thedetection of novel stimuli
We show that the amygdala is sensitive to stimulus novelty, butonly when certain types of stimuli are used. Surprisingly, thesefindings do not depend on the emotional content of the images. Novelemotional and neutral images of humans each evoke robust amygdalaresponses. Our findings are original because we show that neutralscenes do not evoke a novelty response in the amygdala. Remarkably,amygdala activity does not seem to gradually habituate with repeated
stimulus presentations. Instead, activity diminishes after a singlepresentation and this difference remains consistent across subsequenttrials.
In Experiment 1, we measured the effects of novelty and emotionon amygdala activity and found a clear superiority for novelty. Itshould be noted that our lack of emotion effect could be due to a lackof power, given the limited number of trials (see below for a moredetailed discussion). Even so, the fact that we observe such a robusteffect for novelty is surprising because many think that the primaryfunction of the amygdala is to detect and respond to emotional stimuli(Öhman and Mineka, 2001). For example, patients with amygdalalesions fail to recognize fearful facial expressions (Adolphs et al.,1994). These patients also show impairments in social learning andemotional empathy tasks (Hurlemann et al., 2010). Fear-relevantstimuli are also more easily associated with an aversive stimulus(Öhman and Soares, 1993) and capture attention in complex visualarrays (Öhman et al., 2001). In addition to fearful stimuli, theamygdala is also sensitive to other emotional facial expressions(Adolphs, 2008; Britton et al., 2006) such as anger (Whalen et al.,2001) and surprise (Kim et al., 2003). Furthermore, amygdalaresponses can be elicited by either emotional faces or scenes (Brittonet al., 2006; Canli et al., 2000; Hariri et al., 2002; Irwin et al., 1996;Stark et al., 2004) although expressions tend to evoke more activitythan scenes (Britton et al., 2006; Hariri et al., 2002). In Experiment 2,we observed an amygdalar novelty effect for images of humans butnot scenes, which is consistent with observations that the amygdalareceives highly processed visual information (McDonald, 1998) and issensitive to images of faces (Adolphs, 2008) and body gestures(Hadjikhani and de Gelder, 2003). Also, the amygdala and fusiformgyrus are often coactivated during the presentation of faces (Brittonet al., 2008), and lesions of the amygdala decrease face evokedfusiform gyrus activity (Vuilleumier et al., 2004). Lesions of theamygdala impair the ability to recognize fear from a fearfulexpression, which may stem from an impairment in the ability toorient one's gaze towards the eyes of a fearful expression (Adolphset al., 2005). Likewise, amygdala responses in normal individualspredict gaze shifts toward fearful eyes (Gamer and Buchel, 2009), andare larger when individuals view subjects with large pupils (Demoset al., 2008). Amygdala responses are smaller for familiar faces thanfor unfamiliar faces (Gobbini et al., 2004; Leibenluft et al., 2004).Taken together, these results suggest that the amygdala responds tonovel instances of particular classes of stimuli, such as human faces.These results may also help to explain why some studies havereported novelty effects (Daselaar et al., 2006; Grunwald et al., 1998;Knight, 1996) while others failed to report novelty effects in theamygdala (Schwartz et al., 2003; Wright et al., 2003).
Our results however differ from those of a recent study by Blackfordet al. (2010). These two studies differ because blackford and colleaguesused stimuli that did not contain a representation of a human. Even so,they show that novel common and uncommon stimuli each activatethe amygdala. In addition, they show that novel uncommon stimuliactivate the amygdala more than novel common stimuli. In light ofthese recent findings, it is difficult to conclude that amygdalar noveltyresponses are exclusive stimuli depicting humans, especially giventhat to do so would require testing an infinite number of differentstimulus types.Whatwe can say from our data is that there are certaintypes of stimuli that do not evoke such a response, and that moreresearch is needed to determine the key feature or features thatdistinguish between stimuli that do and stimuli that do not evoke anamygdalar novelty response. One possibility is that Blackford andcolleagues used stimuli with strong foreground objects, whereas weused scenes where the focus was on the background content. Futurestudies should investigate the difference between amygdala noveltyresponses evoked by foreground and background content.
Anotherway that our study differs from previous novelty studies isthat we used an event related design, which allowed us to determine
Fig. 6. BOLD responses in the amygdala and hippocampus decrease after single stimuluspresentation. (a,b) Graphs depict BOLD response in the amygdala (a) and hippocampus(b) across trials during Experiment 1. Data points reflect the mean±SEM BOLDresponse evoked by the novel (dark red) and the repeated (light red) images duringeach trial, +Note that for the repeated images, Trial 1 is actually the second imagepresentation, because on the first presentation the image is novel.
1895N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898
Author's personal copy
the response magnitude on each presentation of the novel andrepeated stimuli. Because of this we are able to show that novelimages of humans evoke a response in the amygdala that diminisheseven after a single repetition. Because previous experiments studyingnovelty-specific amygdala responses used block designs (Blackfordet al., 2010; Schwartz et al., 2003; Wright et al., 2003, 2006), this kindof temporal resolution was not possible. These results may alsoexplain why previous experiments show such rapid habituation ofamygdala responses (Britton et al., 2008; Buchel et al., 1998; Fischeret al., 2003; Wright et al., 2003). These previous reports of amygdalahabituation may be due partially to an initial response to stimulusnovelty.
We believe that the novelty effects in the current experimentrepresent an initial evaluation of human faces for evidence of threat inthe environment. In contrast, previous work in our lab employingnon-face stimuli in a fear conditioning paradigm has shown thatamygdala BOLD arises only on trials where the subject demonstrates aconditioned response (Cheng et al., 2006, 2003, 2007). Interestingly,the amygdala BOLD in these two types of studies may arise fromdifferent neural processes in the amygdala, possibly from differentnuclei (Davis et al., 2010; Goossens et al., 2009). The amygdalareceives multimodal sensory inputs via projections to the basolateralnuclei (Sah et al., 2003), which play a crucial role in associatingenvironmental stimuli with biologically significant outcomes(LeDoux, 2000). These structures project to the central nucleus (Sahet al., 2003), which initiates defensive behaviors via projections to thehypothalamus and brainstem (LeDoux, 2000). Interestingly, theseseparate nuclei may make independent contributions to BOLDactivity, which is often classified generally as “amygdala BOLD.”Activity from the basolateral nuclei may lead to the detection of BOLD
responses related to stimulus processing whereas activity of thecentral nucleusmay lead to the detection of BOLD responses related toresponse expression.
Aspects of these results also replicate previous studies. First, ourstimuli evoked more hippocampal activity when novel than whenrepeated. Consistent with previous research, this suggests that thehippocampus is sensitive to novelty in general. Second, hippocampalresponses to novel stimuli showed a trend toward habituation in thefirst experiment. The hippocampus may be sensitive to the novelty ofother aspects of the experimental procedure (Nyberg, 2005), which isconsistent with other studies showing second-order novelty effects inthe hippocampus (Strange and Dolan, 2001; Yamaguchi et al., 2004).In addition to replicating work on the hippocampus, we also replicatework suggesting that there are cortical areas that selectively processfaces (Kanwisher and Yovel, 2006) and scenes (Epstein, 2008).
Our data, consistent with previously published papers, suggest thatthe amygdala and hippocampus may be doing different things duringthe presentation of novel stimuli. The hippocampus responds to noveltyin general and the amygdala responds to the novelty of certain types ofstimuli, such as faces. It is unclear how these brain structures interactwhen we encounter new people. The amygdala seems to contribute toemotion discrimination by directing attention to the eye region of faces,which is necessary for detection of fear (Adolphs et al., 2005; Gamer andBuchel, 2009). One possibility is that the bulk of this processing occurson the initial stimulus presentation, and is triggered by novelty inputsfrom the hippocampus. According to this hypothesis, novelty signalsfrom the hippocampus and facial-recognition signals from higher levelvisual processing areas converge on the amygdala, and this convergenceof information triggers the increase in amygdala activity that weobserved. Although we did not directly test this hypothesis, it is
Fig. 7. Humans and scenes evoke BOLD responses in domain-specific cortical areas. (a) Axial slice showing main effect for faces in the fusiform face area. (b) Coronal slice showingmain effect for scenes in the parahippocampal place area. (a,b) Colors indicate size of F-statistic depicted on brain slice and correspond to the colors on the scale to the left. Bar graphsrepresent the percent signal change in the structures marked by the arrows (data from Experiment 2). (c) Faces evoke more activity than scenes in the fusiform face area. (d) Scenesevoke more activity than faces in the parahippocampal place area. (a: FFA = fusiform face area; c: NE = novel emotional, RE = repeated emotional, NN = novel neutral, RN =repeated neutral; b: PPA = parahippocampal place area; d: NH = novel human, RH = repeated human, NH = novel scene, RH = repeated scene).
1896 N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
supported by our data and can be tested by measuring the functionalconnectivity between the amygdala andhippocampus during novel andrepeated presentations of faces.
The presence of a human in a novel image is sufficient to evoke anovelty-specific amygdala response, and emotional content is notnecessary for these responses. However, it is currently unclearwhether emotional content is sufficient to evoke such responseswhen the novel image does not depict a human subject. A recent studyby Larson and colleagues found that novel spider images evoke largermagnitude amygdala responses than novel neutral images (Larsonet al., 2006), however without a condition where the images arerepeated, it is difficult to tell whether this effect is driven by novelty.In addition, a study by Weierich et al. (2010) suggests that novelpositive and negative images are able to evoke amygdalar noveltyresponses; however, without explicitly controlling for presence of ahuman representation, it is difficult to determine whether emotion ingeneral is sufficient to evoke an amygdalar novelty response.
Again, our results suggest that novel neutral faces but not scenesactivate the amygdala, but we are far from understanding what isspecial about faces. According to some, the amygdala has evolved torapidly and automatically detect stimuli that have historicallysignaled threat in the environment (Isbell, 2006; Öhman and Mineka,2001). Like faces, images of threatening animals like snakes andspiders also meet this criterion. Research suggesting that theamygdala responds to novel snakes or spiders would support thisevolutionary hypothesis.
It's also unclear which aspects of human representations arenecessary and sufficient to evoke novelty responses in the amygdala.The amygdala is sensitive to emotional facial expressions (Adolphs,2008; Britton et al., 2006; Kim et al., 2003; Whalen et al., 2001) andlesions to the amygdala impair patients' ability to direct their gazetoward the eye regions of photographs (Adolphs et al., 2005).Therefore, images of just faces or more specifically images of justthe eye region of the face may be sufficient to evoke an amygdalarnovelty response. However, emotional body gestures also evokeamygdala activity (Hadjikhani and de Gelder, 2003), suggesting thatimages of bodies may be sufficient to evoke an amygdala noveltyresponse. Finally, our results suggest that the amygdala responds tothe novelty of faces independent of emotional content; however ourstudy did not include happy faces. Given that neutral faces can beperceived as more ambiguous and evoke more amygdala activity thanhappy faces (Kukolja et al., 2008), it is possible that happy faces maynot evoke an amygdala novelty response.
Amygdalar novelty responses are larger in individualswith inhibitedtemperament (Schwartz et al., 2003). However, it is currently unclearhow they relate to other behaviors previously associatedwith amygdalaactivity. For instance, there is often a high correspondence between skinconductance responses and amygdala activity (Cheng et al., 2003;Williams et al., 2001). In our current experiments we measured skinconductance responses, but the patterns did not correspond to theamygdala data (see Supplementary Fig. 2).
One limitation to this study is that the original experiment waspart of another separate fear conditioning experiment. As a result,subjects were exposed to an aversive electrical stimulation prior totheir fMRI scan. Because of this exposure, it is difficult to say whetherthese results are generalizable to contexts where the aversivestimulation is not present. This is an empirical question that needsto be answered with additional studies. However, there are severalreasons why we believe that these results are not dependent uponprevious exposure to an aversive electrical stimulation. First, the basicnovelty effect has been demonstrated in studies where an aversiveelectrical stimulation was never presented (Schwartz et al., 2003;Wright et al., 2003, 2006). Our study adds to this literature by showingthat the amygdalar novelty response is not evoked by all stimulustypes. Given that uncertainty about aversive outcomes has beenshown to upregulate amygdala activity (Sarinopoulos et al., 2010), it
is unlikely that the exposure to electrical stimulation prior to theexperiment accounts for the specificity of amygdalar noveltyresponses that we show here. In addition, the pattern of shockexpectancy in these experiments did not correspond to the pattern ofamygdala BOLD, suggesting that differences in shock expectancyacross stimulus types are not sufficient to explain our findings. First,individuals in Experiment 1 generally believed that the shock wasmore likely during emotional stimuli; however these individualsshowed larger BOLD responses to novel emotional and neutral stimuli.Second, individuals believed that the shock was more likely on earliertrials, but our novelty effect in the amygdala was consistent acrosstrials. Finally, subjects in Experiment 2 expected the shock moreduring novel stimuli than repeated stimuli generally, but showedlarger amygdala responses only to the novel faces.
Another limitation to our experiment is that we included only 5trials for each stimulus types. Therefore, our marginal emotion effectin Experiment 1, and our lack of a novelty effect for scenes inExperiment 2 may have been due to type II error. With regard toemotion, this is a valid concern. We originally expected an effect foremotion in Experiment 1, and the pattern of amygdala activity showsmarginally more activity for emotional stimuli. There is no doubt thatamygdala activity contributes emotional processing, and that ourdesign is not completely sensitive to this activity. However, given thatwe see such a striking effect for novelty at this level of sensitivity, ourresults suggest that novelty is a powerful driver of amygdala activity.With regard to scenes, it is unlikely that our lack of a novelty effect isdue to a lack of sensitivity. If this were the case, there should havebeen marginally larger responses to novel scenes coupled with a largeamount of error. Instead amygdala responses are roughly equivalentfor novel and repeated scenes, and significantly smaller thanamygdala responses to novel humans.
The novelty/encoding hypothesis suggests that novelty processingmodulates encoding by prioritizing the to-be-encoded information(Tulving et al., 1996). It is unclear whether amygdala noveltyresponses facilitate encoding. In Experiment 2 we attempted toaddress this question by measuring subsequent memory for the teststimuli, but the small number of stimuli made recognition too easyand participants performed at ceiling levels. However, by identifyingstimulus sets that both evoke and fail to evoke amygdala noveltyresponses, our results provide a tool for subsequent researchers toaddress this question.
Our difficulty finding behavioral outputs that correspond toamygdalar novelty responses further suggest that these responsesrepresent a sensory evaluation of the stimuli. We believe that theprimary function of the amygdala is to allow the individual to rapidlyinitiate defensive behaviors indangerous situations, but in order todosothe amygdala must keep the organism vigilant by allocating attentionalresources to stimuli that potentially signal biologically significantoutcomes (Davis and Whalen, 2001; Holland and Gallagher, 1999).
Faces offer a snapshot of the outside world as viewed through theeyes of another, which can alert an individual to biologicallysignificant events that may have otherwise gone unperceived(Whalen, 2007). We believe that all novel faces, independent ofemotional content, engage the basolateral nuclei of the amygdala,which evaluate the face and engage the central nucleus if and only ifthe face signals a biologically significant outcome.
Acknowledgments
This study was supported by the National Institute of MentalHealth (MH060668 and MH069558).
Appendix A. Supplementary data
Supplementary data to this article can be found online atdoi:10.1016/j.neuroimage.2011.01.034.
1897N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898

Author's personal copy
References
Adolphs, R., 2008. Fear, faces, and the human amygdala. Curr. Opin. Neurobiol. 18,166–172.
Adolphs, R., Tranel, D., Damasio, H., Damasio, A., 1994. Impaired recognition of emotionin facial expressions following bilateral damage to the human amygdala. Nature372, 669–672.
Adolphs, R., Gosselin, F., Buchanan, T.W., Tranel, D., Schyns, P., Damasio, A.R., 2005. Amechanism for impaired fear recognition after amygdala damage. Nature 433,68–72.
Blackford, J.U., Buckholtz, J.W., Avery, S.N., Zald, D.H., 2010. A unique role for the humanamygdala in novelty detection. Neuroimage 50, 1188–1193.
Britton, J.C., Taylor, S.F., Sudheimer, K.D., Liberzon, I., 2006. Facial expressions and complexIAPS pictures: common and differential networks. Neuroimage 31, 906–919.
Britton, J.C., Shin, L.M., Barrett, L.F., Rauch, S.L., Wright, C.I., 2008. Amygdala andfusiform gyrus temporal dynamics: responses to negative facial expressions. BMCNeurosci. 9, 44.
Buchel, C., Morris, J., Dolan, R.J., Friston, K.J., 1998. Brain systems mediating aversiveconditioning: an event-related fMRI study. Neuron 20, 947–957.
Canli, T., Zhao, Z., Brewer, J., Gabrieli, J.D., Cahill, L., 2000. Event-related activation in thehuman amygdala associates with later memory for individual emotional experience.J. Neurosci. 20 RC99..
Cheng, D.T., Knight, D.C., Smith, C.N., Stein, E.A., Helmstetter, F.J., 2003. Functional MRIof human amygdala activity during Pavlovian fear conditioning: stimulusprocessing versus response expression. Behav. Neurosci. 117, 3–10.
Cheng, D.T., Knight, D.C., Smith, C.N., Helmstetter, F.J., 2006. Human amygdala activityduring the expression of fear responses. Behav. Neurosci. 120, 1187–1195.
Cheng, D.T., Richards, J., Helmstetter, F.J., 2007. Activity in the human amygdalacorresponds to early, rather than late period autonomic responses to a signal forshock. Learn. Mem. 14, 485–490.
Cox, R.W., 1996. AFNI: software for analysis and visualization of functional magneticresonance neuroimages. Comput. Biomed. Res. 29, 162–173.
Daselaar, S.M., Fleck, M.S., Cabeza, R., 2006. Triple dissociation in the medial temporallobes: recollection, familiarity, and novelty. J. Neurophysiol. 96, 1902–1911.
Davis, M., Whalen, P.J., 2001. The amygdala: vigilance and emotion. Mol. Psychiatry 6,13–34.
Davis, F.C., Johnstone, T., Mazzulla, E.C., Oler, J.A., Whalen, P.J., 2010. Regional responsedifferences across the human amygdaloid complex during social conditioning.Cereb. Cortex 20, 612–621.
Demos, K.E., Kelley, W.M., Ryan, S.L., Davis, F.C., Whalen, P.J., 2008. Human amygdalasensitivity to the pupil size of others. Cereb. Cortex 18, 2729–2734.
Epstein, R.A., 2008. Parahippocampal and retrosplenial contributions to human spatialnavigation. Trends Cogn. Sci. 12, 388–396.
Fischer, H., Wright, C.I., Whalen, P.J., McInerney, S.C., Shin, L.M., Rauch, S.L., 2003. Brainhabituation during repeated exposure to fearful and neutral faces: a functional MRIstudy. Brain Res. Bull. 59, 387–392.
Fischl, B., Salat, D.H., Busa, E., Albert, M., Dieterich, M., Haselgrove, C., van der Kouwe, A.,Killiany, R., Kennedy, D., Klaveness, S., Montillo, A., Makris, N., Rosen, B., Dale, A.M.,2002. Whole brain segmentation: automated labeling of neuroanatomicalstructures in the human brain. Neuron 33, 341–355.
Fischl, B., Salat, D.H., van der Kouwe, A.J., Makris, N., Segonne, F., Quinn, B.T., Dale, A.M.,2004. Sequence-independent segmentation of magnetic resonance images. Neuro-image 23 (Suppl 1), S69–S84.
Forman, S.D., Cohen, J.D., Fitzgerald, M., Eddy, W.F., Mintun, M.A., Noll, D.C., 1995.Improved assessment of significant activation in functional magnetic resonanceimaging (fMRI): use of a cluster-size threshold. Magn. Reson. Med. 33, 636–647.
Gamer, M., Buchel, C., 2009. Amygdala activation predicts gaze toward fearful eyes.J. Neurosci. 29, 9123–9126.
Gobbini, M.I., Leibenluft, E., Santiago, N., Haxby, J.V., 2004. Social and emotionalattachment in the neural representation of faces. Neuroimage 22, 1628–1635.
Goossens, L., Kukolja, J., Onur, O.A., Fink, G.R., Maier, W., Griez, E., Schruers, K.,Hurlemann, R., 2009. Selective processing of social stimuli in the superficialamygdala. Hum. Brain Mapp. 30, 3332–3338.
Groves, P.M., Thompson, R.F., 1970. Habituation: a dual-process theory. Psychol. Rev.77, 419–450.
Grunwald, T., Lehnertz, K., Heinze, H.J., Helmstaedter, C., Elger, C.E., 1998. Verbalnovelty detection within the human hippocampus proper. Proc. Natl Acad. Sci. USA95, 3193–3197.
Hadjikhani, N., de Gelder, B., 2003. Seeing fearful body expressions activates thefusiform cortex and amygdala. Curr. Biol. 13, 2201–2205.
Hariri, A.R., Tessitore, A., Mattay, V.S., Fera, F., Weinberger, D.R., 2002. The amygdalaresponse to emotional stimuli: a comparison of faces and scenes. Neuroimage 17,317–323.
Holland, P.C., Gallagher, M., 1999. Amygdala circuitry in attentional and representa-tional processes. Trends Cogn. Sci. 3, 65–73.
Hurlemann, R., Patin, A., Onur, O.A., Cohen, M.X., Baumgartner, T., Metzler, S., Dziobek,I., Gallinat, J., Wagner, M., Maier, W., Kendrick, K.M., 2010. Oxytocin enhancesamygdala-dependent, socially reinforced learning and emotional empathy inhumans. J. Neurosci. 30, 4999–5007.
Irwin, W., Davidson, R.J., Lowe, M.J., Mock, B.J., Sorenson, J.A., Turski, P.A., 1996. Humanamygdala activation detected with echo-planar functional magnetic resonanceimaging. NeuroReport 7, 1765–1769.
Isbell, L.A., 2006. Snakes as agents of evolutionary change in primate brains. J. Hum.Evol. 51, 1–35.
Kanwisher, N., Yovel, G., 2006. The fusiform face area: a cortical region specialized forthe perception of faces. Philos. Trans. R. Soc. Lond. B Biol. Sci. 361, 2109–2128.
Kim, H., Somerville, L.H., Johnstone, T., Alexander, A.L., Whalen, P.J., 2003. Inverseamygdala and medial prefrontal cortex responses to surprised faces. NeuroReport14, 2317–2322.
Knight, R., 1996. Contribution of human hippocampal region to novelty detection.Nature 383, 256–259.
Kukolja, J., Schlapfer, T.E., Keysers, C., Klingmuller, D., Maier, W., Fink, G.R., Hurlemann,R., 2008. Modeling a negative response bias in the human amygdala bynoradrenergic-glucocorticoid interactions. J. Neurosci. 28, 12868–12876.
Lang, P.J., Bradley, M.M., Cuthbert, B.N., 2005. International Affective Picture System(IAPS): Digitized Photographs, InstructionManual and Affective Ratings. Universityof Florida, Gainesville, FL.
Larson, C.L., Schaefer, H.S., Siegle, G.J., Jackson, C.A.B., Anderle, M.J., Davidson, R.J., 2006.Fear is fast in phobic individuals: amygdala activation in response to fear-relevantstimuli. Biol. Psychiatry 60, 410–417.
LeDoux, J.E., 2000. Emotion circuits in the brain. Annu. Rev. Neurosci. 23, 155–184.Leibenluft, E., Gobbini, M.I., Harrison, T., Haxby, J.V., 2004. Mothers' neural activation in
response to pictures of their children and other children. Biol. Psychiatry 56,225–232.
McDonald, A.J., 1998. Cortical pathways to the mammalian amygdala. Prog. Neurobiol.55, 257–332.
Menon, V., White, C.D., Eliez, S., Glover, G.H., Reiss, A.L., 2000. Analysis of a distributedneural system involved in spatial information, novelty, and memory processing.Hum. Brain Mapp. 11, 117–129.
Morey, R.A., Petty, C.M., Xu, Y., Pannu Hayes, J., Wagner Ii, H.R., Lewis, D.V., LaBar, K.S.,Styner, M., McCarthy, G., 2009. A comparison of automated segmentation andmanual tracing for quantifying hippocampal and amygdala volumes. Neuroimage45, 855–866.
Nyberg, L., 2005. Any novelty in hippocampal formation and memory? Curr. Opin.Neurol. 18, 424–428.
Öhman, A., Mineka, S., 2001. Fears, phobias, and preparedness: toward an evolvedmodule of fear and fear learning. Psychol. Rev. 108, 483–522.
Öhman, A., Soares, J.J., 1993. On the automatic nature of phobic fear: conditionedelectrodermal responses to masked fear-relevant stimuli. J. Abnorm. Psychol. 102,121–132.
Öhman, A., Flykt, A., Esteves, F., 2001. Emotion drives attention: detecting the snake inthe grass. J. Exp. Psychol. Gen. 130, 466–478.
Sah, P., Faber, E.S., Lopez De Armentia, M., Power, J., 2003. The amygdaloid complex:anatomy and physiology. Physiol. Rev. 83, 803–834.
Sarinopoulos, I., Grupe, D.W., Mackiewicz, K.L., Herrington, J.D., Lor, M., Steege, E.E.,Nitschke, J.B., 2010. Uncertainty during anticipation modulates neural responses toaversion in human insula and amygdala. Cereb. Cortex 20, 929–940.
Schwartz, C.E., Wright, C.I., Shin, L.M., Kagan, J., Rauch, S.L., 2003. Inhibited anduninhibited infants “grown up”: adult amygdalar response to novelty. Science 300,1952–1953.
Stark, R., Schienle, A., Walter, B., Kirsch, P., Blecker, C., Ott, U., Schafer, A., Sammer, G.,Zimmermann, M., Vaitl, D., 2004. Hemodynamic effects of negative emotionalpictures — a test–retest analysis. Neuropsychobiology 50, 108–118.
Strange, B.A., Dolan, R.J., 2001. Adaptive anterior hippocampal responses to oddballstimuli. Hippocampus 11, 690–698.
Tulving, E., Markowitsch, H.J., Craik, F.E., Habib, R., Houle, S., 1996. Novelty andfamiliarity activations in PET studies of memory encoding and retrieval. Cereb.Cortex 6, 71–79.
Vuilleumier, P., Richardson, M.P., Armony, J.L., Driver, J., Dolan, R.J., 2004. Distantinfluences of amygdala lesion on visual cortical activation during emotional faceprocessing. Nat. Neurosci. 7, 1271–1278.
Weierich, M.R., Wright, C.I., Negreira, A., Dickerson, B.C., Barrett, L.F., 2010. Novelty as adimension in the affective brain. Neuroimage 49, 2871–2878.
Whalen, P.J., 2007. The uncertainty of it all. Trends Cogn. Sci. 11, 499–500.Whalen, P.J., Shin, L.M., McInerney, S.C., Fischer, H., Wright, C.I., Rauch, S.L., 2001. A
functional MRI study of human amygdala responses to facial expressions of fearversus anger. Emotion 1, 70–83.
Williams, L.M., Phillips, M.L., Brammer, M.J., Skerrett, D., Lagopoulos, J., Rennie, C.,Bahramali, H., Olivieri, G., David, A.S., Peduto, A., Gordon, E., 2001. Arousaldissociates amygdala and hippocampal fear responses: evidence from simulta-neous fMRI and skin conductance recording. Neuroimage 14, 1070–1079.
Wright, C.I., Martis, B., Schwartz, C.E., Shin, L.M., Fischer, H., McMullin, K., Rauch, S.L.,2003. Novelty responses and differential effects of order in the amygdala,substantia innominata, and inferior temporal cortex. Neuroimage 18, 660–669.
Wright, C.I., Wedig, M.M., Williams, D., Rauch, S.L., Albert, M.S., 2006. Novel fearful facesactivate the amygdala in healthy young and elderly adults. Neurobiol. Aging 27,361–374.
Yamaguchi, S., Hale, L.A., D'Esposito, M., Knight, R.T., 2004. Rapid prefrontal-hippocampal habituation to novel events. J. Neurosci. 24, 5356–5363.
1898 N.L. Balderston et al. / NeuroImage 55 (2011) 1889–1898