auditory hair cells in human fetuses: synaptogenesis and ciliogenesis
TRANSCRIPT

JOURNAL OF ELECTRON MICROSCOPY TECHNIQUE 15:115-122 (1990)
Auditory Hair Cells in Human Fetuses: Synaptogenesis and Ciliogenesis MIREILLE LAVIGNE-REBILLARD AND REMY PUJOL INSERM U. 254 and Uniuerstt6 de Montpe l lw II Laboratoire de Neurobiologte de l’Audition, H6prtal St Charles, 34059 Montpellier Cedex (France)
KEY WORDS Development, Human fetus, Cochlea, Hair cells, Synaptogenesis, Ciliogenesis
ABSTRACT This paper brings together the most recent findings concerning the development of human fetus cochlear hair cells, as observed using transmission and scanning electron micro- scopy (TEM and SEMI. Specific attention is drawn to the formation of synapses and the growth of stereocilia. In both types of hair cells, synaptogenesis begins before ciliogenesis (week 10 of gesta- tion versus week 12). In the inner hair cell (IHC), an adultlike stage is seen around week 15 for synapses, but not before week 22 for stereocilia. In the outer hair cell (OHC), both synapses and stereocilia are not yet fully mature a t week 22. Classic gradients of maturation are found: a base-to-apex gradient, and an IHC-to-OHC gradient. By comparing these results with the anatom- ical and functional data on cochlear development in laboratory mammals, the onset of cochlear function in the human fetus can be estimated to occur around week 18. The completion of cochlear maturation based upon the same anatomical criteria should occur about 10 weeks later.
INTRODUCTION The gross anatomical development of the human fe-
tal cochlea is relatively well documented (see Retzius, 1884; Streeter, 1917; Bast and Anson, 1949). Bredberg (1968) added to these descriptions a good qualitative and quantitative analysis using mainly a surface prep- aration technique. By contrast, until recently, the fine morphology has been the matter of only few descrip- tions (Tanaka et al., 1979; Igarashi, 1980; Sanchez- Fernandez et al., 1983; Hoshino and Nakamura, 1985). A few years ago, we began to investigate fetal cochleas a t different stages of its development using both scan- ning and transmission electron microscopy (SEM and TEM) (Pujol and Lavigne-Rebillard, 1985; Lavigne-Re- billard and Pujol, 1986, 1987, 1988). The aim of the present paper is to bring together these results focus- ing on two different aspects of the cochlear develop- ment, i.e., the formation of synapses, and the growing of stereocilia and the tectorial membrane. Comparison is made with what is well known in other mammals (see Pujol and Uziel, 1988, for a recent review), and, finally, some physiological correlations will be dis- cussed.
MATERIALS AND METHODS Thirty-four cochleas from fetuses ranging in age
from 9 to 22 gestational weeks were investigated. The fetuses were obtained from legal abortions (for the ear- lier stages) or from therapeutic abortions. The fetal ages were determined by the menstrual phases and confirmed by echography for most of the cases; they were expressed in weeks from fecundation. Generally, when it was possible, one cochlea was used for trans- mission electron microscopic (TEM) observations, and the other belonging to the same fetus, for scanning electron microscopic (SEMI observations.
The fetal debris or the temporal bone, depending on
the age and the abortion technique, were immersed in the fixative (2.5% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4) for 2 to 24 hr. The cochleas were dis- sected out in the same fixative and rinsed in buffer. The specimens were postosmicated (1% osmium tetrox- ide in 0.1 M cacodylate buffer) for 1 hr and dehydrated in a graded series of ethanol.
The specimens (19) investigated for the synaptogen- esis studies were then classically processed for TEM observations. The observations were performed with a Philips 300 microscope. Concerning the specimens (15) used for the auditory epithelial surface observations, the cochleas were dissected in 70% ethanol to remove the otic capsule; the Reissner’s membrane and, when it was possible, the tectorial membrane were also re- moved from the oldest stages. After the last immersion in 100% ethanol, the dehydration was completed in a critical point drier using C02 as the transition fluid. The specimens were then coated with gold and viewed a t 15 kV in a Jeol JSM 255 microscope.
RESULTS In all specimens observed, the two general gradients
of cochlear maturation were confirmed, whatever the type of investigation performed (TEM or SEM). The maturation of the basal turn of the cochlea led that of the apex by 1 or 2 weeks. At a given level of the CO- chlea, the maturation of the inner hair cells (IHCs) preceded that of the outer hair cells (OHCs).
Received December 21, 1988; accepted in revised form January 18, 1989. Address reprint requests to Dr. M. Lavigne-Rebillard, INSERM U. 254, Lab-
oratoire de Neurobiologie de I’Audition, CHR HBpital St. Charles, 34059 Mont- pellier Cedex (France).
0 1990 WILEY-LISS. INC

116 M. LAVIGNE-REBILLARD AND R. PUJOL
9 gestational weeks The cochlea was completely coiled (2.5 turns); the
cochlear duct was well formed but no perilymphatic space was noted. The great and small epithelial ridges of the Corti’s primordium were already noticeable on the medial wall. SEM observations did not show any signs of ciliary differentiation at the apical surface of the cochlear epithelium. Low-magnification TEM ob- servations confirmed these findings.
From the spiral ganglion neurons, fibers could be followed up to the undifferentiated epithelium, run- ning below the basilar membrane and entering the ep- ithelium, probably a t the level of the future habenula perforata. These fibers spread between the lower layers of the epithelial cells.
10 gestational weeks SEM observations showed a still undifferentiated
sensory epithelium. But the tectorial membrane, seen for the first time in a 9Yz-week fetal cochlea, was now identifiable throughout the cochlea. At the apical level, only an amorphous substance could be seen on the in- ternal side, with some fibrils on its top. But basal- wards, a network of slanted fibers covering a large part of the cochlear epithelium was already present (Fig. 1). This nascent tectorial membrane was so firmly at- tached to the epithelial surface which is very difficult to remove from the specimens up to the 12th gesta- tional week.
As far as the Corti’s epithelium was concerned, TEM observations did not show noticeable differences from the previous stage. Nevertheless, the nerve endings be- came more abundant in the deep and intermediate lay- ers of the sensory epithelium (Fig. 2). In most of the sections, some of these fibers even reach the apical pole of the epithelium. The base of some cells could be found surrounded by nerve endings (see Figs. 11 and 12 in Pujol and Lavigne-Rebillard, 1985).
11-12 gestational weeks The apical level of the cochlea showed no signs of
epithelial differentiation. At the medial and basal turns of a 12-week cochlea, the first signs of ciliary differentiation could be observed (Fig. 3). Both inner (IHCs) and outer (OHCs) hair cells first showed round bundles of cilia arising among numerous microvilli (Fig. 3A). The ciliary bundles belonging to the IHCs were taller than those belonging to the OHCs. For both types of hair cells, the stereocilia developed a three- dimensional V-like arrangement (Fig. 3B); i.e., the longest cilia were located near the kinocilium, while the length of the cilia decreased both laterally and to- ward the cochlear internal side (Fig. 3C). During this development of the stereocilia arrangement, glabrous portions of the hair cell surface appeared both on the internal and external sides.
By TEM observations, the sensory cells appeared now well cytodifferentiated from the surrounding epithelial cells. In the basal turn, one IHC and three (often four) OHCs could be clearly recognized in each section. The base of both types of hair cells was surrounded by nerve endings, but those below the IHC were more numerous.
Fig. 1. Scanning electron micrograph of a surface view of the Cor- ti’s epithelium from a 10-week fetal cochlea. The epithelium is undif- ferentiated, all cells being covered with microvilli. The arrows point to the area of the future organ of Corti. On the modiolar side, the fibrillar material of the nascent tectorial membrane (TM) is seen. Scale bar: 100 pm.
Fig. 2. Transmission electron micrograph of a transverse section of the Corti’s epithelium from a 10-week fetal cochlea. At the level of the spiral vessel (sv) nerve fibers (arrows) are seen within the stria- tum of epithelial cells. Scale bar: 10 Wm.
Synaptic specializations such as asymmetric thickening or presynaptic body and vesicles were found at the junc- tions between afferent dendrites, and IHCs or OHCs. Below the IHCs some densely and regularly vesiculated endings could be found.

DEVELOPMENT OF HUMAN AUDITORY HAIR CELLS 117
because of the gradient of maturation from the apical level, which was poorly differentiated, to the basal level, which was much more advanced, one could ob- serve different developmental stages of IHCs and OHCs along the same cochlea (see Figs. 1 to 12 in Lav- igne-Rebillard and Pujol, 1986). At the basal end of a 131h-week cochlea, some IHCs had begun to exhibit a more rectilinear arrangement of stereocilia. One to 2 weeks later, an adultlike ciliary arrangement could be observed on most of the basal IHCs. Supernumerary IHCs could be seen a t this stage. On the top of the OHCs, the three-dimensional V-like arrangements of stereocilia became striking. The shorter cilia disap- peared, making the apical glabrous surfaces of the OHCs more extended, especially on the internal side. Often, a fourth row of OHCs could be seen. During this period, the surface of the different supporting cells be- came clearly recognizable. The surface of Deiters’ cells were well distinguishable from that of Hensen’s cells and Claudius’ cells.
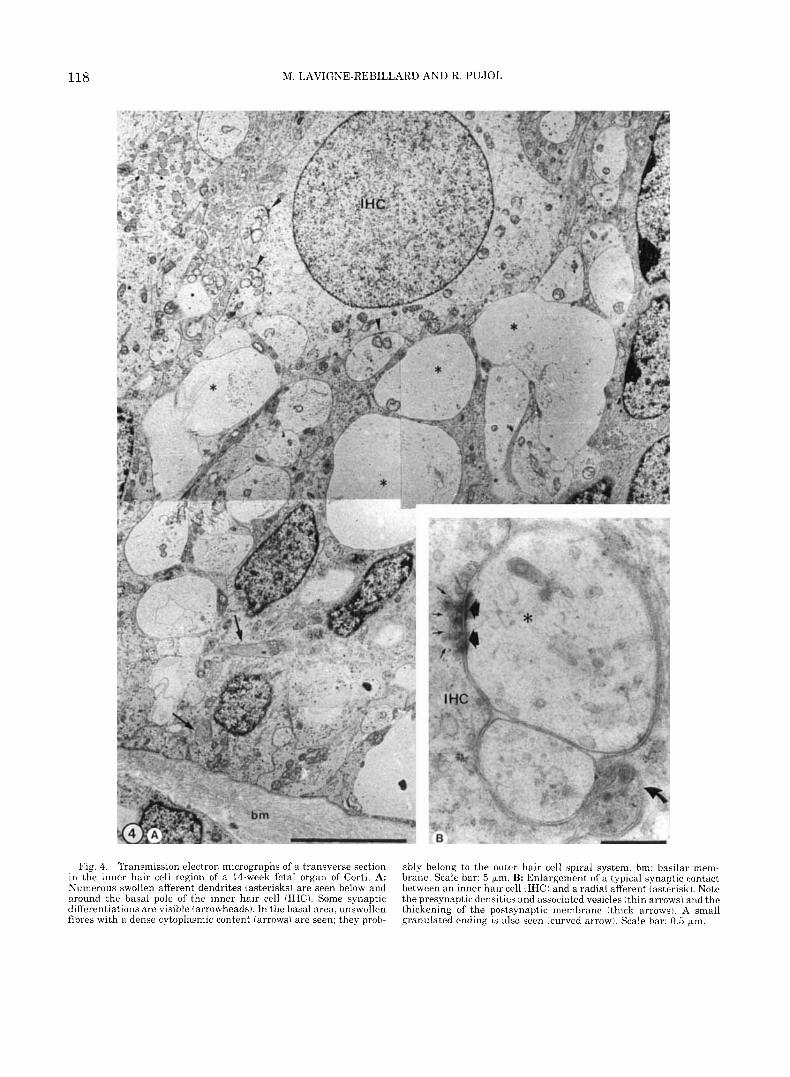
Concerning the innervation, the afferent nerve end- ings were very abundant a t the base of both types of hair cells (Figs. 4A, 5A), and numerous well defined synaptic contacts could be seen (Figs. 4B, 5B). At the base of the IHCs, most of the afferent endings appeared swollen, with very poor cytoplasmic content (Fig. 4). Below the IHCs, some varicosities were detected in the inner spiral bundle, and a few axodendritic synapses were also observed (see Fig. 3 in Lavigne-Rebillard and Pujol, 1988). Below the OHCs, together with the same kind of “swollen” afferents, smaller fibers could be no- ticed spiralling between Deiters cells (Fig. 5). These fibers had a dense cytoplasm containing a regular ar- ray of well-preserved microtubules.
16-18 gestational weeks Few data can be added to our description, as we could
obtain only few well-preserved specimens from this stage, especially for TEM observations. Moreover, a layer of amorphous substance below the fibrous tecto- rial membrane increased the difficulty in obtaining a clean surface of the sensory epithelium. Large pieces of this amorphous substance remained on the top and be- tween the hair cells. In addition, strands from the fibrillar tectorial membrane were still firmly anchored to the microvilli of the supporting cells, especially the Hensen’s cells. The surface of the head of the inner
Fig. 3. Scanning electron micrographs of the surface views of the organ of Corti from a 12-week fetal cochlea, at three different levels: upper medial turn (A), lower medial turn (B), and basal turn (C). The two main gradients of maturation of stereocilia are clearly seen in these pictures; i.e., a basal-to-apical gradient, and an inner (IHCb to-outer hair cells (OHC) gradient. When they first appear, the ciliary tufts are round (both types of hair cells in A, and OHCs in B) . Then, a tridimensional V-like arrangement is noticed (IHCs in B, and both types of hair cells in C) . All supporting cells are covered with mi- crovilli. Note the four rows of OHCs in C. Scale bars: 5 Fm.
13-15 gestational weeks The sensory epithelium exhibited advanced signs of
differentation throughout the cochlea. Nevertheless,
pillar cells could be seen, for the first time, in the basal turn of a 18-week fetal cochlea (see Fig. 2 in Lavigne- Rebillard and Pujol, 1987).
20-22 gestational weeks As shown in Figure 6, the surface of the developing
inner pillar cells were now identifiable throughout the cochlea; i t looked fully mature in the basal turn of a 22-week-old cochlea. IHCs exhibited an adult rectilin- ear ciliary arrangement, but still with the presence of the kinocilium. OHCs showed a W-shaped pattern of the stereocilia only at the basal turn, even in 22-week- old fetuses. The kinocilium was still present on the OHC surface but now at some distance from the stere- ocilia bundle. The OHCs in the first row showed a reg- ular alignment as well as a constant orientation of
118 M. LAVIGNE-REBILLARD AND R. PUJOL
Fig. 4. Transmission electron micrographs of a transverse section in the inner hair cell region of a 14-week fetal organ of Corti. A: Numerous swollen afferent dendrites (asterisks) are seen below and around the basal pole of the inner hair cell (IHC). Some synaptic differentiations are visible (arrowheads). In the basal area, unswollen fibres with a dense cytoplasmic content (arrows) are seen; they prob-
ably belong to the outer hair cell spiral system. bm: basilar mem- brane. Scale bar: 5 bm. B: Enlargement of a typical synaptic contact between an inner hair cell (IHC) and a radial afferent (asterisk). Note the presynaptic densities and associated vesicles (thin arrows) and the thickening of the postsynaptic membrane ithick arrows). A small granulated ending is also seen (curved arrow). Scale bar: 0.5 bm.

DEVELOPMENT OF HUMAN AUDITORY HAIR CELLS 119
Fig. 5. Transmission electron micrographs of a transverse section in the outer hair cell region of a 14-week fetal organ of Corti (same section as in Fig. 4). A: The base of an outer hair cell (OHC) is con- tacted by two kinds of afferents: three swollen and clear endings (as- terisks) are seen contacting the synaptic pole of the hair cell. They present the same characteristics as radial afferents in contact with the inner hair cell (compare with Fig. 4). Other nerve profiles (arrows)
are smaller and present a dense arrangement of microtubules: most of them are cut on their spiraling course between two Deiters’ cells (D). bm: basilar membrane. Scale bar: 5 p,m. B: Enlargement of a synapse between an outer hair cell (OHC) and an afferent ending (asterisk). Typically, a t this early stage of the outer hair cell development, the presynaptic specialization is characterized by multiple presynaptic bodies (arrows) surrounded by microvesicles. Scale bar: 0.5 p,m.

120 M. LAVIGNE-REBILLARD AND R. PUJOL
their stereocilia, while OHCs in the 2nd, and even more in the 3rd row exhibited numerous irregularities. Supernumerary IHCs and OHCs were still commonly found.
The organ of Corti, observed by TEM, looked mature, with a well-open tunnel of Corti and adult-shaped OHCs separated by the spaces of Nuel (Fig. 7A). As far as the innervation was concerned, the only noticeable changes to be pointed out occurred a t the base of the OHCs. Afferent dendrites now seemed to belong to only one type and no swollen dendrites were seen. The num- ber of the presynaptic specializations within the OHCs had been greatly reduced. Beside these afferents, effer- ent endings were seen, forming typical axosomatic syn- apses a t the base of OHCs (Fig. 7B). Nevertheless, most of the postsynaptic cisterns were still incompletely formed, even a t the basal level of our oldest specimens.
DISCUSSION The formation of synapses and the development of
stereocilia confirm that the human cochlea follows the same general gradients of maturation as other mam- malian cochleas. Moreover, the human cochlea, with its slow development, appears to be a good model for describing and timing some stages precisely that are difficult to interpret in small laboratory animals.
Base-to-apex gradient This general base-to-apex gradient of the cochlear
maturation has been previously discussed (see Rubel, 1978; Romand, 1983; Pujol and Uziel, 1988 for review). A possibility still exists that the development starts a t a certain distance from the basal end and progresses toward both extremities (Bredberg, 1968). The difficul- ties in macro- and microdissecting the hook region are obvious, and a few if any reliable findings have been reported concerning the extreme base of the cochlea. The base-to-apex gradient is particularly clear in SEM during the formation of the tectorial membrane (see Figs. 4 to 7 in Lavigne-Rebillard and Pujol, 1987) and the growth of the stereocilia. The difference of about 2 weeks exists in their development between the basal and apical turns. The most apical tip should be consid- ered separately: for the arrangement of hair cells in regular rows, the regular patterning of stereocilia, and the complete formation of tectorial membrane to an adultlike (or basal-like) stage never seems to be reached (Lavigne-Rebillard and Pujol, 1986; see also Lenoir et al., 1980, 1987 in the rat).
IHC-to-OHC gradient IHCs mature before OHCs. This is also generally ac-
cepted (see Romand, 1983; Pujol and Uziel, 1988, for review). A good example of such a gradient is given by the overall shape of hair cells. The IHC gets its adult- like shape well before OHCs, which markedly change during the last stages. Similarly, while IHC synapto- genesis seems to be achieved quite early (week 15 in human fetus), a t the OHC synaptic pole, changes occur much later, efferent synapses are not distinguishable before week 22. In human cochleas, the IHC-to-OHC gradient is particularly clear when observing the grow- ing stereocilia. At a given level of the cochlea, the time
interval by which the development of the IHC stereo- cilia leads those of OHCs is about a few days. This difference is less expressed during the latter develop- mental stages. At week 22, for example, a kinocilium (last sign of immaturity) is still present on both IHCs and OHCs.
Comparison between synaptogenesis and ciliogenesis
At both IHC and OHC levels synaptogenesis begins before ciliogenesis. At least, when stereocilia are first seen growing on the top of an hair cell (week 12/13), this hair cell is always synaptically contacted by nerve fibers. Conversely, nerve endings have been observed contacting cells still only covered by microvilli (Pujol and Lavigne-Rebillard, 1985). Sans and Dechesne (1985) investigating the vestibular epithelium from our specimens have reported identical findings. This point has been extensively discussed elsewhere (Pujol and Sans, 19861, and such an arrival of fibers before the development of stereocilia does not necessarily mean that nerve fibers trigger the hair cell ciliogenesis. Syn- aptogenesis also ends before ciliogenesis, a t least a t the IHC level, where an adult stage is reached first for synapses (week 15) then for stereocilia (after week 22). This is not so obvious however, for OHCs because of the prolonged maturation of the medial efferent synapses: a t week 22, neither synaptogenesis nor ciliogenesis is achieved at this level.
Duality of OHC afferent innervation During the first steps of development a typical pat-
tern of immature innervation is observed a t OHC level. The OHC at this stage is only innervated by nonvesic- ulated endings which could well belong to the two dif- ferent types of afferents (from type I and I1 spiral gan- glion neurons). This feature is particularly clear a t stage 13-15 weeks, when small and swollen afferent endings are observed. Small afferents with dense cyto- plasm and regular microtubules are seen spiraling be- tween Deiters cells. These fibers most probably belong to type I1 neurons and form the classical afferent con- tacts with OHCs. The swollen afferent endings, with clear or floculescent cytosplasm, look completely dif- ferent. They actually show similar characteristics as the radial afferents connected with the IHCs (cf. Figs. 4 and 5). Their swollen appearance could be explained by a similar sensitivity to anoxic conditions. The pos- sibility for some radial afferents to temporarily branch and innervate OHCs has already been suggested in the rat pup cochlea. In this species, during the first week of postnatal life, at a time when no vesiculated endings from the medial efferent could be detected, the OHC is clearly connected by endings reacting to kainic acid. Such a reaction, consisting mainly of a drastic swell- ing, is characteristic (at all developmental stages as well as in adults) of radial afferent connected to IHC (Pujol, 1986; Pujol et al., 1985).
Physiological correlations It would be difficult from structural human data
alone to draw any physiological conclusions. Actually, the first recorded auditory responses in premature ba-

DEVELOPMENT OF HUMAN AUDITORY HAIR CELLS 121
Fig. 6. Scanning electron micrograph of a surface view of the or- gan of Corti from the medial turn of a 22-week-old fetal cochlea. The stereocilia are fully developed on both inner (IHC) and outer (OHC) hair cells; they are linearly organized on IHC and display a V-like arrangement on OHCs. The surface of the head of inner pillar (IP) is well developed. A supernumerary IHC (arrow) and an incomplete fourth row of OHCs (arrowheads) are seen. Scale bar: 10 pm.
Fig. 7. Transmission electron micrographs of a transverse section in the outer hair cell region of a 22-week fetal cochlea. A Four outer hair cells (1,2,3,4) are seen in this section. They have an elongated shape and a mature cuticular plate; they are separated by Nuel's spaces. Scale bar: 10 pm. B: Base of an outer hair cell (OHC) in contact with a vesiculated efferent nerve ending. The postsynaptic cistern (arrowheads) is visible on the opposite cell side. Bar: 1 pm.

122 M. LAVIGNE-REBILLARD AND R. PUJOL
bies were reported a t week 25 (Starr et al., 1977), and behavioral evidence of hearing in utero comes from about the same stage (week 24, Birnholz and Benacer- raf, 1983). However, the anatomical development of the cochlea is remarkably similar in all mammals, thus allowing quite good extrapolations. A specific stage in ultrastructural development of human fetal cochlea is first compared with what is well known in an animal model (e.g., the rat) which offers precise correlations between anatomy and physiology (Lenoir et al., 1980, 1987; Puel and Uziel, 1987); then, an estimation of the physiological status can be made in humans. The onset of function presumably should take place by week 18 (Pujol and Uziel, 1988; Pujol et al., 1990). One month later, the OHCs and tectorial membrane have reached a stage where some tuning would be expected in the cochlear responses. The lack of good specimens after week 22 of gestation does not allow for precise timing concerning the end of the cochlear maturation. Never- theless, it can be estimated by most of the morpholog- ical and probably physiological criteria that the human cochlea achieves its maturation by the 7th to 8th month of pregnancy.
ACKNOWLEDGMENTS Authors are indebted to the Service of Neonatalogie
(Professor H. Bonnet) et d'obstetrique (Professor F. Laffargue) du CHR de Montpellier for their efficient collaboration. Thanks are due to P. Sibleyras, who printed the photographs, and to A. Bara, who edited the manuscript.
REFERENCES Bast, T.H., and Anson, B.J. (1949) The Temporal Bone and Ear.
Charles C Thomas, Springfield, Illinois. Birnholz, J .C. and Benacerraf, B.R. (1983) Development of human
fetal hearing. Science, 222:516-518. Bredberg, G. (1968) Cellular pattern and nerve supply of the human
organ of Corti. Acta Otolaryngol. (Stockh.1, Suppl. 236. Hoshino, T., and Nakamura, K. (1985) Nerve fibers in the fetal organ
of Corti. Scanning electron microscopic study. Ann. Otol. Rhinol. Laryngol.. 94:304-308.
Igarashi. Y. (19801 Cochlea of the human fetus: a scanning electron microscope study. Arch. Histol. Jpn. , 43:195-209.
Lavigne-Rebillard, M., and Pujol, R. (19861 Development of auditory hair cell surface in human fetuses: A scanning electron microscopy study. Anat. Embryol., 174:369-377.
Lavigne-Rebillard, M., and Pujol, R. (19871 Surface aspects of the
developing human organ of Corti. Acta Otolaryngol. (Stockh. ), 436 (suppl:):43-50.
Lavigne-Rebillard, M., and Pujol, R. (1988) Hair cell innervation in the fetal human cochlea. Acta Otolarvnpol. (Stockh.), 105:398-402.
Lenoir, M., Shnerson, A,, and Pujol, R, (79803 Cochlear receptor de- velopment in the ra t with emphasis on synaptogenesis. Anat. Em- bryol., 160:253-262.
Lenoir, M., Puel, J.-L., and Pujol, R. (1987) SEM study of the rat cochlear development with emphasis on stereocilia and tectorial membrane. Anat. Embryol., 175:477-487.
Pujol, R. (1986) Synaptic plasticity in the developing cochlea. In: The Biology of Change in Otolaryngology. R.W. Ruben, T.R. Van de Water, and E.W. Rubel, eds. Elsevier Science Publishers B.V. (Bio- medical Division), New York, pp. 47-54.
Pujol, R., and Lavigne-Rebillard, M. (1985) Early stages of innerva- tion and sensory cell differentiation in the human organ of Corti. Acta Otolaryngol. (Stockh.!, 423 (suppl.):43-50.
Pujol, R., and Sans, A. (1986) Synaptogenesis in the mammalian in- ner ear. In: Advances in Neural and Behavioral Development. R. A s h , ed. Ablex Press, Norwood, New York, pp. 1-18.
Puel, J.-L., and Uziel, A. (1987) Correlative development of cochlear action potential sensitivity, latency, and frequency selectivity. Dev. Brain Res., 37:179-188.
PUJO~, R., and Uziel, A (1988) Auditory development: peripheral as- pects. In: Handbook of Human Growth and Developmental Biology. Vol I: Neural, Sensory, Motor, and Integrative Development. Part B: Sensory, Motor, and Integrative development. E. Meisami and P.S. Timiras, eds. CRC Press, Inc., Boca Katon, Florida, pp. 109- 130.
PUJO~, R., Lenoir, M., Robertson, D., Eybalin, M., and Johnstone, B.M. ( 1985) Kainic acid selectively alters auditory dendrites connected with cochlear inner hair cells. Hearing Res., 18:145-151.
Pujol, R., Lavigne-Rehillard, M., and Uziel, A. (1990) Development of human cochlea. Acta Otolaryngol. (Stockh.). in press.
Retzius, G. (1884) Gehororgan des Wirbeltiere. In: Das Gehororgan der Reptilien, der Vogel, und der Saugetiere. Vol. 2. Samsom and Wallin. Stockholm.
Romand, R. (19831 Development of Auditory and Vestibular Systems. Academic Press, New York.
Rubel, E.W. (1978) Ontogeny of structure and function in the verte- brate auditory system. In: Handbook of Sensory Physiology. Val 9. H. Autrum, R. Jung, W.R. Loewenstein, D.M. MacKay, and H.L. Teuber, eds. Springer-Verlag, Berlin, pp. 135-237.
Sanchez-Fernandez. J.M., Rivera, J.M.. and Macias, J.A. (1983) Early aspects of human cochlea development and tectorial membrane his- togenesis. Acta Otolaryngol. (Stockh.), 95:460-469.
Sans, A., and Dechesne, C. (19853 Early development of vestibular receptors in human embryos. Acta Otolaryngol. (Stockh. 1, 423 (suppl. 15-58,
Starr, A., Amlie, R.N., Martin, W.H., and Sanders, S. (19771 Devel- opment of auditory function in newborn infants revealed by a d - tory brainstem potentials. Pediatrics, 60:831-839.
Streeter, G.L. (1917) The develoument of the scala tvmuani. scala " . vestibuli and perioticular cistern in the human embryo. Am. J. Anat.. 21:299-320.
Tanaka, K., Sakai, N., and Terayama, Y. (1979) Organ of Cori in the human fetus. Scanning and transmission electronmicroscope stud- ies. Ann. Otol. Rhinol. Laryngol., 88:749-758.