asymmetric distribution of phosphatidylethanolamine in …repository.ias.ac.in/15935/1/318.pdf ·...
TRANSCRIPT

Asymmetric Distribution of Phosphatidylethanolaminein C. albicans: Possible Mediation by CDRl, aMultidrug Transporter Belonging to ATP BindingCassette (ABC) SuperfamilySMRITI DOGRA1, SHANKARLING KRISHNAMURTHY', VINITA GUPTAl, BHARAT L. DIXIT2,C. M. GUPTA2, DOMINIQUE SANGLARD3 AND RAJENDRA PRASAD'*
'Membrane Biology Laboratory, School of Life Sciences, Jawaharlal Nehru University, New Delhi-l10067, India2Division of Membrane Biology, Central Drug Research Institute, Lucknow, India3Institut de Microbiologie, Centre Hospitalier Universitaire Vaudois, 1011 Lausanne, Switzerland
By using two molecular probes, we demonstrate that only 4% of total phosphatidylethanolamine (PtdEtn) in theplasma membrane (PM) of a human pathogenic yeast, Candida albicans, is present in its external half. Evidence ispresented to show that the availability of PtdEtn could be related to the expression of a multidrug transporter CDRlof C. albicans, and the process is energy-dependent. A homozygous CDRI disruptant strain of C. albicans showsalmost 23% reduction in the external labelling of PtdEtn. This report shows that, similar to human MDRs, yeastmultidrug transporter could also be involvedin aminophospholipidtranslocation. ' -= - ~ - ",
KEYWORDS- membrane asymmetry; multidrug resistance; aminophospholipid translocation; ABC transporter;CDRl; C. albicans
INTRODUCTION
..
Membrane phospholipids are asymmetricallydistributed across the plasma membrane (PM) ofa variety of cells (Diaz and Schroit, 1996;Williamson and Schlegel, 1994). Most of the phos-phatidylethanolamine (PtdEtn) and phosphatidyl-serine (PtdSer) are located on the inner leafletof the lipid bilayer, while phosphatidylcholine(PtdCho) and other lipids are predominantlylocalized on the outer half of the bilayer (Schroitand Zwaal, 1991). The asymmetrical distributionof membrane lipids is very specific and the loss ofthis asymmetry has been linked to various physio-logical processes (Kean et al., 1997; Diaz andSchroit, 1996;Tang et al., 1996; Utsugi et al., 1991;Herrmann and Devaux, 1990).
Overexpression of a 170 kDa glycoprotein (P-glycoprotein) is involved in multidrug resistance(MDR). Such MDR proteins belong to a group of*Correspondence to: Prof. R. Prasad, Membrane BiologyLaboratory, School of Life Sciences, Jawaharlal NehruUniversity, New Delhi-ll0067, India. Fax: 618-7338,616-5886; e-mail: [email protected]
.f
membrane transporters known as ABC (ATPbinding cassette) transporters, which are widelydistributed throughout the evolutionary scale(Prasad et al., 1996; Hyde et al., 1990; Higgins,1992; Lewis, 1994). The completion of the S.eerevisiae genome project has revealed that thereare 29 putative ABC proteins, which could behomologues of MDRs (Decottignies and Goffeau,1997). The multidrug transporters in yeast notonly include members of the ABC family but alsoa family of membrane facilitators (MFS), whichare probably anti porters (Goffeau et al., 1997;Balzi and Goffeau, 1994). In addition, regulatorynetworks of PDR (pleiotropic drug resistance)genes, backed by transcription regulators, are alsowell characterized (Balzi and Goffeau, 1994, 1995).
The studies involving MDR in yeast haveattracted much attention, since they are goodmodel systems and some of them are also patho-genic. Candida albieans is a dimorphic and oppor-tunistic human pathogen (Gow et al., 1994;Prasad, 1991), which is naturally more resistant toseveral drugs than S. eerevisiae (Prasad et al.,
Yeast 15,111-121(1999)

1996). In recent years, the incidence of C. albieanscells acquiring resistance to azoles and polyeneshas increased alarmingly (Prasad et al., 1996;Van den Bossche, 1995). We had earlier clonedand sequenced a multidrug transporter, CDRl(Candida drug resistance) of C. albieans, which is ahomologue of S. eerevisiae multidrug efflux pumpPDR5 (Prasad et al., 1995b; Balzi and Goffeau,1994, 1995). A few more homologues of CDRlhave been identified recently in C. albieans (Walshet al., 1997; Balan et al., 1997; Sanglard et al.,1997) and many genes specific to fluconazole andbenomyl resistance have also been characterized(Alarco et al., 1997; Prasad et al., 1995; Prasadet al., 1996).
Earlier, human Mdrlp and Mdr3p andmouse mdr2p have been shown to have flippaseactivity, which mediates phospholipid translo-cation directed towards the exoplasmic leaflet ofthe lipid bilayer (Ruetz and Gros, 1994; Boschet al., 1997; Van Helvoort et al., 1996). Recently,Kean et al. (1997) have shown that translocationof PtdEtn in S. eerevisiae is controlled by PDRland PDR3 and identified two new alleles, tpel-l(dominant) and tpe2-1 (semidominant) for PDRIand PDR3, respectively. Pdrlp and Pdr3p are tran-scription regulators which control the expressionof several genes including ABC transporterslike PDR5 and SNQ2 (Balzi and Goffeau, 1994,1995).
Asymmetrical distribution of phospholipids isnot restricted to mammalian cells. The PM of S.eerevisiae cells is also asymmetric with regard toaminophospholipids and is controlled by anenergy-dependent amino phospholipid translocase(Kean et al., 1997; Tang et al., 1996). This com-munication, for the first time, by using twomolecular probes, shows that C. albieans PM isasymmetric, where PtdEtn is predominantly con-fined to the cytoplasmic leaflet of PM. We furthershow that the expression of CDRl, a multidrugresistance ABC transporter gene, affects theavailability of PtdEtn in the outer half of the PMin an energy-dependent process, and presentevidence that Cdrl p could be a phospholipidtranslocator.
MATERIALS AND METHODS
Materials
Yeast media were obtained from Difcolauoratories (USA). All other chemicals were
purchased from Sigma Chemicals Co. (USA).Precoated silica gel G-60 F TLC plates wereobtained from E. Merck.
Strains and culture conditions
Candida albieans ATCC 10261 was grown inYEPD medium (2% bactopeptone, 1% bactoyeastextract and 2% glucose) (Jethwaney et al., 1997).C. albieans strains CAF2-1 (Llura3::imm434/URA3), DSY449 (Lledrl::hisG/L1edrl::hisG),DSY1025 (Lledrl::hisG/L1edrl::hisG, Lledr2::hisG/Lledr2::hisG) were also maintained and grown inYEPD medium, as described earlier (Sanglardet al., 1997). S. eerevisiae JG436 (Mat a,PDR5::Tn5, leu2, met5, ura3-52, mak71, KRBl)was a kind gift from Dr J. Golin, CatholicUniversity of America, Washington, DC (Prasadet al., 1995b). S. eerevisiae AD 1234568 (Mat a,pdrl-3, hisl, ura3, pdr5Ll, snq2Ll, pdrlOLl, pdrllLl,pdrl5Ll, yorlL1, yeflL1), was a kind gift from A.Decottignies, E. Balzi and A. Goffeau, UniversiteCatholique de Louvain, Belgium. Both the strains,which were hypersensitive to cycloheximide, weretransformed with the CDRI gene by a methoddescribed earlier (Prasad et al., 1995b). The result-ing transformants S-12 and AD-CDRI werehyper-resistant to cycloheximide and to manyother unrelated drugs and were expressing CDRl,as was evident from their Northern analysis (datanot shown).
Trinitrobenzenesulphonie acid (TN BS) labelling
TNBS labelling was done essentially by themethod described earlier (Balasubramanian andGupta, 1996). C. albieans cells were harvested inmid-log phase by centrifuging at 3000 rpm for5 min at 4°C and washed two or three times withbuffer A (100 mM potassium phosphate, 5 mMEDT A, pH 7'5). The harvested cells (0'6 g wetweight) were resuspended in 5 ml of buffer B(100 mM potassium phosphate and 600 mM KCI,pH 8'2) and kept at 4°C with gentle swirling. Anequal volume of indicated concentration of TNBS(freshly prepared and chilled) in buffer B wasadded to the cell suspension. After indicated time,the reaction was stopped by adding equal volumeof buffer C (200 mM potassium acetate, 600 mMKCI, pH 4'5). The suspension was centrifuged andwashed two or three times with buffer C untilthe colour of free TNBS disappeared from thesupernatant.
Yeast 15,111-121(1999)
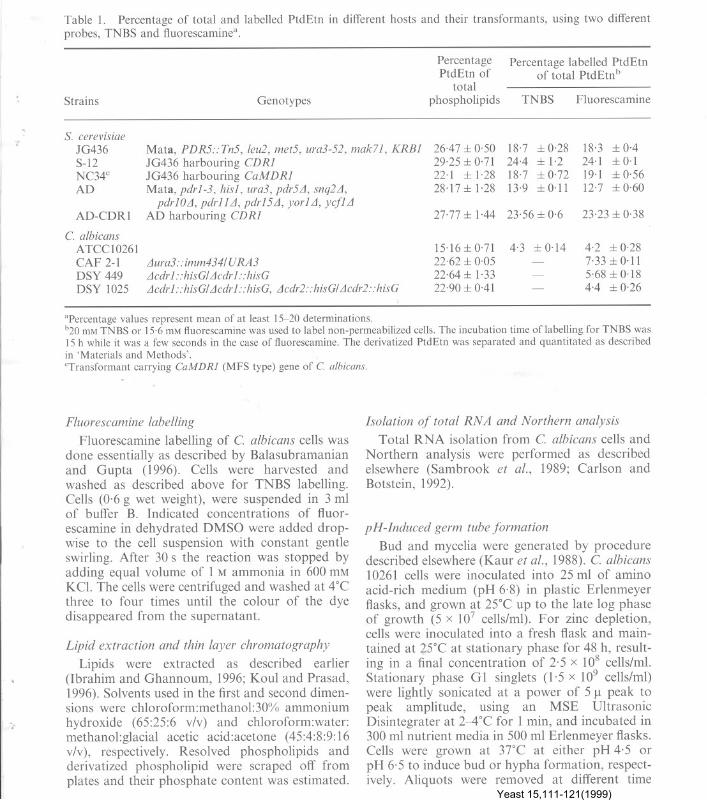
Table I. Percentage of total and labelled PtdEtn in different hosts and their transformants, using two differentprobes, TNBS and fluorescaminea.
Strains Genotypes
PercentagePtdEtn of
totalphospholipids
Percentage labelled PtdEtnof total PtdEtnb
TNBS Fluorescamine
"Percentage values represent mean of at least 15-20 determinations.b20 mM TNBS or 15.6 mM ftuorescamine was used to label non-permeabilized cells. The incubation time of labelling for TNBS was15 h while it was a few seconds in the case of ftuorescamine. The derivatized PtdEtn was separated and quantitated as describedin 'Materials and Methods'.
cTransformant carrying CaMDRl (MFS type) gene of C. albicans.
Fluorescamine labelling
Fluorescamine labelling of C. albicans cells wasdone essentially as described by Balasubramanianand Gupta (1996). Cells were harvested andwashed as described above for TNBS labelling.Cells (0'6 g wet weight), were suspended in 3 mlof buffer B. Indicated concentrations of fluor-escamine in dehydrated DMSO were added drop-wise to the cell suspension with constant gentleswirling. After 30 s the reaction was stopped byadding equal volume of 1M ammonia in 600 mMKC!. The cells were centrifuged and washed at 4°Cthree to four times until the colour of the dyedisappeared from the supernatant.
Lipid extraction and thin layer chromatography
Lipids were extracted as described earlier(Ibrahim and Ghannoum, 1996; Koul and Prasad,1996). Solvents used in the first and second dimen-sions were chloroform:methanol:30% ammoniumhydroxide (65:25:6 v/v) and chloroform:water:methanol:glacial acetic acid:acetone (45:4:8:9:16v/v), respectively. Resolved phospholipids andderivatized phospholipid were scraped off fromplates and their phosphate content was estimated.
Isolation of total RNA and Northern analysisTotal RNA isolation from C. albicans cells and
Northern analysis were performed as describedelsewhere (Sambrook et al., 1989; Carlson andBotstein, 1992).
pH-Induced germ tubeformation
Bud and mycelia were generated by proceduredescribed elsewhere (Kaur et al., 1988). C. albicans10261 cells were inoculated into 25 ml of aminoacid-rich medium (pH 6'8) in plastic Erlenmeyerflasks, and grown at 25°C up to the late log phaseof growth (5 x 107 cells/ml). For zinc depletion,cells were inoculated into a fresh flask and main-tained at 2YC at stationary phase for 48 h, result-ing in a final concentration of 2.5 x 108 cells/m!.Stationary phase G 1 singlets (I' 5 x 109 cells/ml)were lightly sonicated at a power of 5 /l peak topeak amplitude, using an MSE UltrasonicDisintegrater at 2--4°Cfor 1min, and incubated in300 ml nutrient media in 500 ml Erlenmeyer flasks.Cells were grown at 37°C at either pH 4,5 orpH 6,5 to induce bud or hypha formation, respect-ively. Aliquots were removed at different time
S. cerevisiaeJG436 Mata, PDR5::Tn5, leu2, met5, ura3-52, mak71, KRB1 26.47::1: 0,50 18.7 ::I:0.28 18.3 ::I:0,4
S-12 JG436 harbouring CDR1 29.25::1: 0.71 24.4 ::I:1.2 24.1 ::I:0.1NC34c JG436 harbouring CaMDR1 22.1 ::I: 1.28 18.7 ::I: 0,72 19.1 ::1:0.56AD Mata, pdr1-3, his1, ura3, pdr5£l, snq2£l, 28.17::1: 1.28 13-9 ::I:0.11 12.7 ::I:0,60
pdrlO£l, pdrll£l, pdr15£l, yorl£l, ycf1£lAD-CDRI AD harbouring CDR1 27.77 ::I: 1.44 23,56::1: 0,6 23.23::1: 0,38
C. albicansATCC I0261 15.16::1: 0,71 4,3 ::I:0.14 4.2 ::I:0.28
CAF 2-1 £lura3::imm434IURA3 22.62 ::I: 0,05 - 7.33::1:0.11DSY 449 £lcdr1::hisGI £lcdr1: :hisG 22.64::1: 1.33 - 5.68::1: 0.18DSY 1025 £lcdr1::hisGI £lcdr1::hisG, £lcdr2::hisGI £lcdr2::hisG 22.90::1: 0-41 - 4.4 ::I:0.26
Yeast 15,111-121(1999)
(a)I
4.5
4
3.5
i 3 6
'3 i t 5o 2.5...
.....
J 40
2"0 3....-.rf21.5-
'*- <f. 1O I I I
0.5 +i I v 5 10 15 20 25
Time (he)II I
0
0 5 10 15 20 25 30
Concentration of TNBS (mM)I
(b)
I4.5
4
3.5
i 3
2.5
.....0
2....
1.5-'*-
1
0.5
00 5 10 15 20 25
Concentration of fluorescamine (mM)
Yeast 15,111-121(1999)

intervals, and cell divergence was microscopicallyexamined.
RESULTS
Labelling of plasma membrane PtdEtn of intactC. albicans cells
TNBS has been widely used in determiningaminophospholipid distribution in membranes ofdifferent cell types. Recently, Balasubramanianand Gupta (1996) have shown the suitability ofTNBS and fluorescamine for labelling the externalPtdEtn of yeast PM. It is evident from Table I thatboth the probes were equally efficient in labellingthe exposed PtdEtn of different yeast strains. Thesuitability of these probes was further establishedwhen we checked their impermeability under ourlabelling conditions. Under experimental con-ditions, the probes were impermeable to yeast cellsand could only label the PtdEtn of the externalleaflet. If cells were permeabilized, the samereagents could label up to 80% of the aminophos-pholipids, including PtdSer (it was not labelled innormal cells by either of the reagents, since mostof PtdSer tS localized in the cytoplasmic leaflet).Under the microscope, permeabilized cells labelledwith fluorescamine showed dispersed fluorescence,while the latter was localized to the exteriors ofnon-permeabilized cells (data not shown). Theresults of the present study thus confirm thatTNBS and fluorescamine can be used to labelPtdEtn of C. albicans and S. cerevisiae cells.
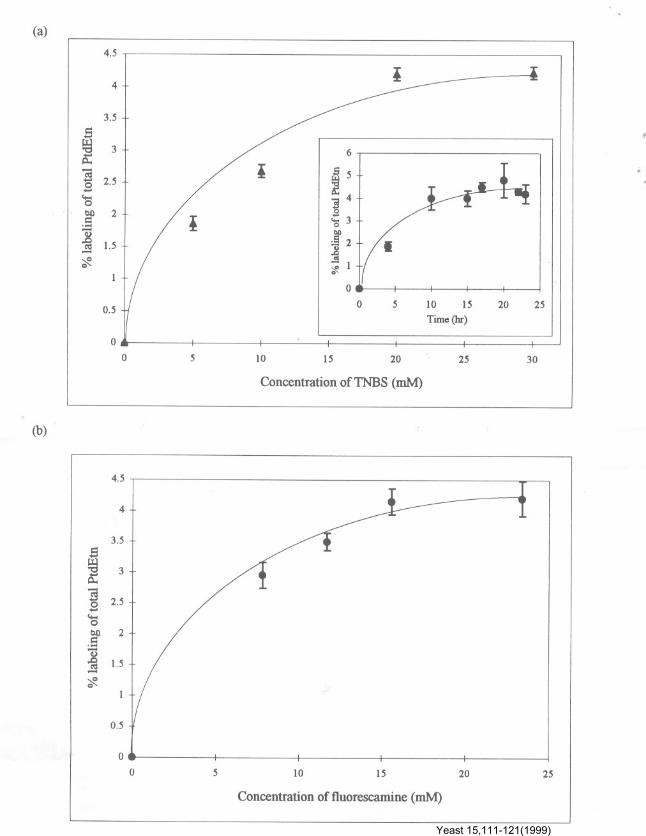
As can be seen from Figure la, b, PtdEtn couldbe labelled by both the probes in a concentration-dependent manner. In purified PM of C. albicans,of the total phospholipids present, '" 15% wasPtdEtn (data not shown) and only ",4% of it waslabelled with both the aminophospholipid labellingreagents in intact cells. This would mean that, outof total PtdEtn in non-permeabilized C. albicanscells, only '" 4% is available for external labellingand the rest ( '" 96%) is localized in the inner leafletof PM. It must be pointed out that about 10-15 hwere required for TNBS (20 mM)labelling (Figurela, inset), while fluorescamine (15'6 mM) couldlabel the amino phospholipid in 30 s (data notshown). Since the labelling with fluorescamine isvery rapid, it is expected to cause much less
perturbation in the lipid organization, andtherefore was routinely used in the followingexperiments (Rawyler et al., 1984).
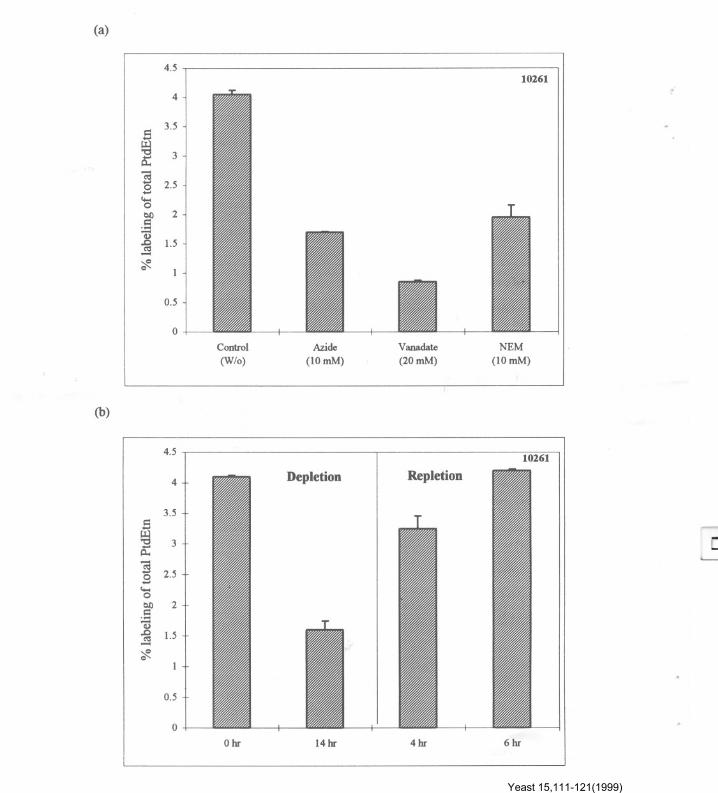
PtdEtn translocation from inner to outer leaflet isenergy dependent and protein-mediated
The movement of PtdEtn across PM ofC. albicans could be prevented if cells werepreincubated with energy inhibitors like sodiumazide and sodium orthovanadate. Figure 2a,depicts labelling of PtdEtn in the presence of theseinhibitors. Sodium azide, which is a mitochondrialATP synthesis inhibitor, and sodium ortho-vanadate, which is a typical inhibitor ofPM-ATPase, if added prior to labelling of PtdEtnwith fluorescamine, resulted in reduced labelling.Almost 58-80% labelling of PtdEtn was inhibitedby 10 mM of azide and 20 mM of vanadate. NEM(10 mM),which blocks free thiol groups of proteinsand thus prevents protein-mediated processes,when added before labelling, was also able toblock PtdEtn labelling with fluorescamine byabout 52% (Figure 2a). If cells were starved ofglucose for 14 h for the depletion of cellular ATP,the labelling was reduced by 60%. However, if cellswere repleted with glucose, the total labelling ofPtdEtn was again restored within 6 h. As shownearlier, for S. cerevisiae cells (Balasubramanianand Gupta, 1996) there may be an energy depen-dent amino phospholipid translocase in C. albicanswhich controls the movement of PtdEtn across thetwo mono layers (Figure 2b). It must be mentionedthat the total phospholipid composition of C.albicans, determined under various experimentalconditions, did not vary significantly (data notshown).
Expression of CDRI is linked to PtdEtntranslocation
In order to ascertain whether CDRI expressioncould contribute in PtdEtn translocation, S.cerevisiae transformant S-12, which was expressingCDRl, was used to examine the PtdEtn distri-bution (Prasad et al., 1995b). Interestingly, in S-12strain out of total PtdEtn, almost 24% of it waslabelled by both fluorescamine and TNBS, as
Figure I. Percentage labelling of PtdEtn of plasma membrane of Candida albicans ATCC 10261 with (a) TNBS and (b)fluorescamine. The inset (a) shows the optimum time required for a 20 mMTNBS labelling. The labelling was done as described inMethods. Derivatized PtdEtn and other phospholipids were extracted and quantified as mentioned in 'Materials and Methods'.
Yeast 15,111-121(1999)
(a)
I4.5
I
10261-4
S 3.5
3
'3o 2.5-'C)
.s 2
J 1.5
*"1
0.5
0
Control Azide Vanadate NEM
(W/o) (10mM) (20mM) (10 mM)
I(b)
I4.5 ,
10261
4-!- Depletion I Repletion
3.5
3+ I II C
'3o 2.5-'C)
2....-v
1.5-*"
I
0.5
0
Ohr 14hr 4hr 6hr
Yeast 15,111-121(1999)

,,,nmvrnV.3rnULll'lU lKAN:SLUCATION IS MEDIATED BY MULTIDRUG TRANSPORTER II7
compared to its host strain, JG436 ('" 19%) lackingCDRI and also having disrupted PDR5, which isa homologue of CDRI (Table I). That CDRIcould be involved in PtdEtn translocation wasfurther confirmed when another transformant ofS. cerevisiae (NC34), which was expressing a non-ABC transporter benomyl resistance (CaMDRI)gene, a membrane facilitator (Prasad et al., per-sonal communication) was checked for PtdEtnlocalization. It was observed that", 19% of PtdEtnwas labelled in NC34 as in its host strain JG436(Table 1). It must be pointed out that only 4%of total PtdEtn could be labelled in C. albicans10 261 cells, while 18-24% of it was labelled inS. cerevisiae strains.
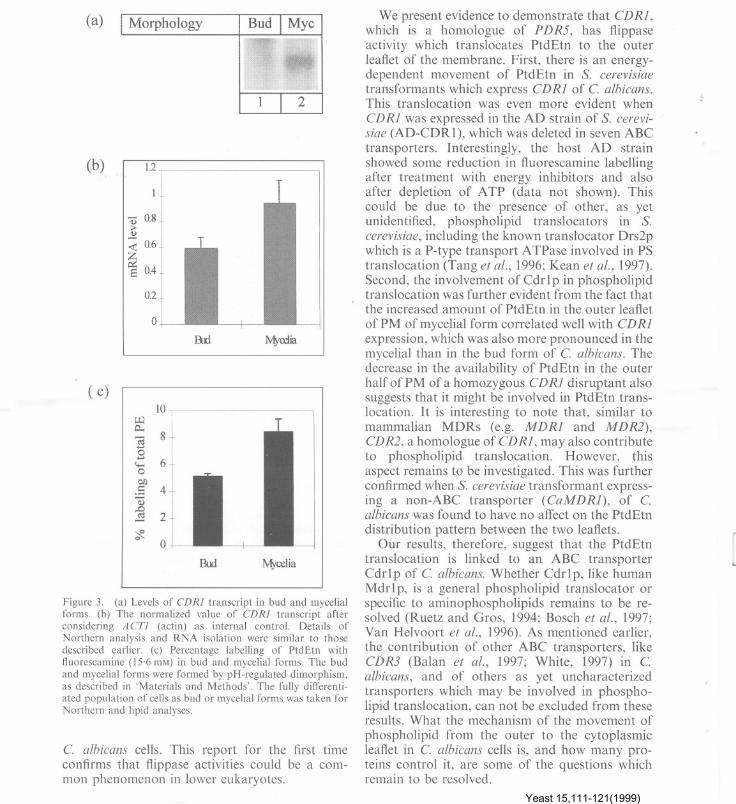
The expression of CDRI is morphologicallyregulated. As can be seen from Figure 3a thatCDRI transcript is higher in mycelial form than inthe bud form of C. albicans (Figure 3a and 3b).Interestingly, PtdEtn, in the outer leaflet ofmembrane, was also found to be much higher inthe mycelial form as compared to bud form(Figure 3c). It is pertinent to mention that therewas no significant change in phospholipid com-position including that of PtdEtn between bud ormycelial forms of C. albicans cells (data notshown). Thus, the expression of CDRI appears tobe linked to PtdEtn translocation.
S. cerevisiae strain S-12 having disrupted PDR5,is known to harbour several other ABC trans-porters, which might mask the function of Cdrl p(Decottignies and Goffeau, 1997). We, therefore,transformed S. cerevisiae (ADI234568) which was(called as AD hereafter, in this paper) constructedby A. Decottignies (in A. Goffeau's group), where-in seven ABC transporter genes, e.g. PDR5 (pleio-tropic drug resistance gene), PDRJO, PDRll,PDRI5, SNQ2 (Saccharomyces nitroquinoline-N-oxide resistance gene), ycn (yeast cadmium fac-tor gene) and YORI (yeast oligomycin resistancegene) were disrupted (Decottignies et al., 1998).AD strain was transformed with the CDRI gene(Prasad et al., 1995b) and the resulting transform-ant was designated as AD-COR I. Both the strainswere subjected to fluorescamine labelling. Thelabelling of PtdEtn was about 24% in AD-COR I,as compared to its host strain AD where only
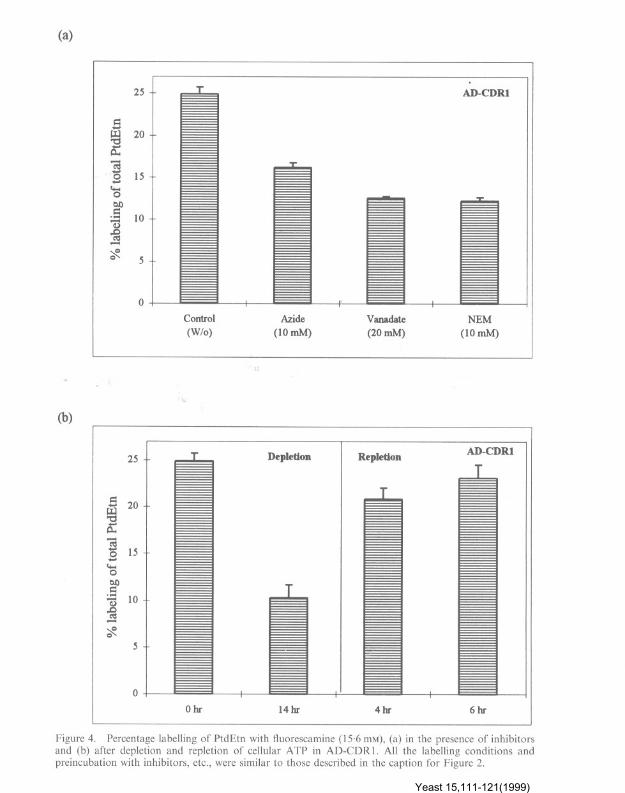
13-14% of PtdEtn could be labelled with fluor-escamine (Table I). Similar to C. albicans cells(Figure 2a, b), the translocation of PtdEtnmediated by CDRI was energy dependent as thelabelling with fluorescamine was inhibited by azideand vanadate. NEM could also prevent the trans-location of this aminophospholipid in AD-COR 1strain (Figure 4a). ATP depletion of cells clearlyprevented the translocation of PtdEtn which couldbe restored upon its repletion (Figure 4b). Ahomozygous disrupted CDRI strain of C. albicansDSY449 was used to check if the non-availabilityof functional Cdr Ip could affect the availability ofexternal PtdEtn (Sanglard et al., 1997). It is evi-dent from Table 1 that DSY449 has about 23%less of external PtdEtn than its parent strainCAF2-1. Interestingly, a double disrupt ant ofCDRI and CDR2 (a homologue of CDRl)DSYI025, showed even less (",40% less as com-pared to parent strain) of PtdEtn in the externalhalf of PM (Table 1) suggesting the involvementof both the ABC transporters in PtdEtn trans-location. It should be mentioned that the differ-ence in external labelling of PtdEtn in strain 10261and CAF2-1 could be due to their genotypicdifferences.
DISCUSSION
We have shown that PM of C. albicans cells isasymmetric with regard to PtdEtn distribution.Most of the PtdEtn was localized in inner leafletand only 4% of it was exposed to external leaflet.The distribution of PtdEtn was affected if cellswere preincubated with energy inhibitors prior tothe labelling with fluorescamine. It was apparentfrom our results that availability of PtdEtn inouter half was mediated by an energy-dependentprocess. In this regard it is pertinent to mentionabout recent reports where aminophospholipidtranslocase activities in various systems includingyeast cells have been shown to control the amino-phospholipidmovement(Decottignieset al., 1998;Kean et al., 1997; Balasubramanian and Gupta,1996; Tang et al., 1996). However, the presenceof such translocase was not demonstrated in
Figure 2. Effect of (a) different inhibitors, (b) depletion and repletion of cellular ATP on PtdEtn of the plasma membrane ofCandida albicans ATCC 10261 with fluorescamine (15'6 mM). Cells were incubated with the indicated concentrations of sodiumazide (2 h), sodium orthovanadate (30 min) and NEM (10 min). For ATP depletion, cells were starved for 14 h and were repletedwith the addition of 2% glucose for 4-6 h. All the treated cells were subjected to fluorescamine labelling, as described in 'Materialsand Methods'.
Yeast 15,111-121(1999)
(a) I Morphology
fv¥.elia
Figure 3. (a) Levels of CDRI transcript in bud and mycelialforms. (b) The normalized value of CDRI transcript afterconsidering A cn (actin) as internal control. Details ofNorthern analysis and RNA isolation were similar to thosedescribed earlier. (c) Percentage labelling of PtdEtn withfluorescamine (15.6 mM) in bud and mycelial forms. The budand mycelial forms were formed by pH-regulated dimorphism,as described in 'Materials and Methods'. The fully differenti-ated population of cells as bud or mycelial forms was taken forNorthern and lipid analyses.
C. albieans cells. This report for the first timeconfirms that flippase activities could be a com-mon phenomenon in lower eukaryotes.
We present evidence to demonstrate that CDRI,which is a homologue of PDR5, has flippaseactivity which translocates PtdEtn to the outerleaflet of the membrane. First, there is an energy-dependent movement of PtdEtn in S. eerevisiaetransformants which express CDRI of C. albieans.This translocation was even more evident whenCDRI was expressed in the AD strain of S. eerevi-siae (AD-CDR1), which was deleted in seven ABCtransporters. Interestingly, the host AD strainshowed some reduction in fluorescamine labellingafter treatment with energy inhibitors and alsoafter depletion of ATP (data not shown). Thiscould be due to the presence of other, as yetunidentified, phospholipid translocators in S.eerevisiae, including the known translocator Drs2pwhich is a P-type transport ATPase involved in PStranslocation (Tang et al., 1996; Kean et al., 1997).Second, the involvement of Cdrl p in phospholipidtranslocation was further evident from the fact thatthe increased amount of PtdEtn in the outer leafletof PM of mycelial form correlated well with CDRIexpression, which was also more pronounced in themycelial than in the bud form of C. albieans. Thedecrease in the availability of PtdEtn in the outerhalf of PM of a homozygous CDRI disruptant alsosuggests that it might be involved in PtdEtn trans-location. It is interesting to note that, similar tomammalian MDRs (e.g. MDRI and MDR2),CDR2, a homologue of CDRI, may also contributeto phospholipid translocation. However, thisaspect remains to be investigated. This was furtherconfirmed when S. eerevisiae transformant express-ing a non-ABC transporter (CaMDRI), of C.albieans was found to have no affect on the PtdEtndistribution pattern between the two leaflets.
Our results, therefore, suggest that the PtdEtntranslocation is linked to an ABC transporterCdrl p of C. albieans. Whether Cdrl p, like humanMdrlp, is a general phospholipid translocator orspecific to amino phospholipids remains to be re-solved (Ruetz and Gros, 1994; Bosch et al., 1997;Van Helvoort et al., 1996). As mentioned earlier,the contribution of other ABC transporters, likeCDR3 (Balan et al., 1997; White, 1997) in C.albieans, and of others as yet uncharacterizedtransporters which may be involved in phospho-lipid translocation, can not be excluded from theseresults. What the mechanism of the movement ofphospholipid from the outer to the cytoplasmicleaflet in C. albieans cells is, and how many pro-teins control it, are some of the questions whichremain to be resolved.
...
(b)
I l.:r
Q) 0.81><I)
=< 0.6
E 0.4
0.2
0
IlrlI
( c)10
,)c:: 60
. 4"a)
2
0
BtxI
Yeast 15,111-121(1999)
(a)
DepleUon RepleUonAD-COR1
6hr
Figure 4. Percentage labelling of PtdEtn with ftuorescamine (15.6 mM), (a) in the presence of inhibitorsand (b) after depletion and repletion of cellular ATP in AD-CDRI. All the labelling conditions andpreincubation with inhibitors, etc., were similar to those described in the caption for Figure 2.
14hr 4hr
25 r IIIAD-CDR1
20
IS....0
:.= 10CI)'i-?ft. 5
0
Control Azide Vanadate NEM
(W/o) (IOmM) (20 mM) (10 mM)
(b)
I:s 20!gc::
IS-'a
:.= 10
-
?ft.5
0
Ohr
Yeast 15,111-121(1999)

ACKNOWLEDGEMENTS
We are grateful to A. Decottignies, E. Balzi andA. Goffeau for kindly providing the strainAD1234568. We are also grateful to PranabMukherjee for his help in finalizing the manuscriptand to Mr R. N. Saini for photographic work. Thework presented in this paper has been supported inpart by grants to one of us (R.P.) from theDepartment of Biotechnology (BT/R&D/15/02/94), and the Council of Scientific Research(60(0028)98/EMR-II), India. S.D., S.K.M. andV.G. acknowledge the fellowships awarded fromthe University Grants Commission.
REFERENCES
Alarco, A. M., Balan, I., Talibi, D., Mainville, N. andRaymond, M. (1997). API-mediated multidrugresistance in Saccharomyces cerevisiae requires FLRIencoding a transporter of the major facilitator super-family. J. Biol. Chem. 272, 19,304-19,313.
Balan, I., Alarco, A. M. and Raymond, M. (1997). TheCandida albicans CDR3 gene codes for an opaque-phase ABC transporter. J. Bacteriol. 179,7210-7218.
Balasubramanian, K. and Gupta, C. M. (1996). Trans-bilayer phosphatidylethanolamine movements in theyeast plasma membrane. Evidence for a protein-mediated, energy-dependent mechanism. Eur. J.Biochem. 240, 798-806.
Balzi, E. and Goffeau, A. (1994). Genetics and biochem-istry of yeast multi drug resistance. Biochim. Biophys.Acta 1187, 152-162.
Balzi, E. and Goffeau, A. (1995). Yeast multidrugresistance: the PDR network. 1. Bioenerget. Biomemb.27, 71-76.
Bosch, I., Joannopoulos, K. D., Wu, R.-L., Furlong,S. T. and Croop, J. (1997). Phosphatidylcholine andphosphatidylethanolamine behave as substrates of thehuman MDRI P-glycoprotein. Biochemistry 36,5685-5694.
Carlson, M. and Botstein, D. (1992). Two differentiallyregulated mRNAs with different 5' ends encodesecreted and intracellular forms of yeast invertase.Cell 28, 145-154.
Decottignies, A. and Goffeau, A. (1997). Completeinventory of the yeast ABC proteins. Nature Genet.15, 137-145.
Decottignies, A., Grants, A. M., Nicholas, J. W., Wet,H. D., McIntosh, D. B. and Goffeau, A. (1998).ATPase and multidrug transport activities of theoverexpressed yeast ABC protein Yor Ip. J. BioI.Chem. 273, 12,612-12,622.
Diaz, C. and Schroit, A. J. (1996). Role of translocasesin the generation of phosphatidylserine asymmetry.1. Memb. BioI. 151, 1-9.
Goffeau, A., Park, J., Paulsen, I. T., Jonniaux, J.-L.,Dinh, T., Mordant, P. and Saier, M. H., Jr (1997).Multidrug-resistant transport proteins in yeast: com-plete inventory and phylogenetic characterization ofthe yeast open reading frames within the MajorFacilitator Superfamily. Yeast 13, 43-54.
Gow, N. A. R., Hube, B., Bailey, D. A., Schofield,D. A., Munro, c., Swoboda, R. K., Bertram, G.,Westwater, c., Broadbent, I., Smith, R. J., Gooday,G. W. and Brown, A. J. P. (1994). Genes associatedwith dimorphism and virulence of Candida albicans.Can. J. Bot. 73 (Supp!.), s335-s342.
Herrmann, A. and Devaux, P. F. (1990). Alteration ofthe aminophospholipid translocase activity duringin vivo and artificial aging of human erythrocytes.Biochim. Biophys. Acta 1027,41-46.
Higgins, C. F. (1992). ABC transporters: from micro-organisms to man. Ann. Rev. Cell Biol. 8, 67-113.
Hyde, S. c., Emsley, P., Hartshorn, M. J., Mimmack,M. M., Gileadi, D., Pearce, S. R., Gallagher, M. P.,Gill, D. R., Hubbard, R. E. and Higgins, C. F. (1990).Structural model of ATP binding proteins associatedwith cystic fibrosis, multidrug resistance and bacterialtransport. Nature 346, 362-365.
Ibrahim, A. S. and Ghannoum, M. A. (1996). Chroma-tographic analysis of lipids. In Prasad R. (Ed.),Manual on Membrane Lipids. Springer-Verlag, Berlin/Heidelberg, pp. 52-79.
Jethwaney, D., Hofer, M., Khaware, R. K. and Prasad,R. (1997). Functional reconstitution of a purifiedproline permease from Candida albicans: interactionswith the antifungal cispentacin. Microbiology 143,397-404.
Kaur, S., Mishra, P. and Prasad, R. (1988).Dimorphism-associated changes in intracellular pHof Candida albicans. Biochim. Biophys. Acta 972,277-282.
Kean, L. S., Grant, A. M., Angeletti, c., Mahe, Y.,Kuchler, K., Fuller, R. S. and Nichols, J. W. (1997).Plasma membrane translocation of fluorescent-labeled phosphatidyl ethanolamine is controlled bytranscription regulators, PDRI and PDRJ. J. Cell.BioI. 138, 255-270.
Koul, A. and Prasad, R. (1996). Extraction ofmembrane lipids. In Prasad, R. (Ed.), Manual onMembrane Lipids. Springer-Verlag, Berlin, pp. 37-52.
Lewis, K. (1994). Multidrug resistance pumps inbacteria:variation on theme. TrendsBiochem.Sci. 19,119-123.
Prasad, R. (1991). Candida albicans: Cellular andMolecular Biology. Springer-Verlag, Berlin.
Prasad, R., Krishnamurthy, S., Prasad, R. and Gupta,V. (1995a). Multiple drug resistance in Candida albi-cans. Acta Biochem. Polonica 42, 497-504.
Prasad, R., Worgifosse, P. D., Goffeau, A. and Balzi, E.(1995b). Molecular cloning and characterisation of anovel gene of C. albicans, CDRI, conferring multiple
-.
Yeast 15,111-121(1999)

resistance to drugs and antifungals. Curro Genet. 27,320-329.
Prasad, R., Krishnamurthy, S., Prasad, R., Gupta, V.and Lata, S. (1996). Muitidrug resistance: anemerging threat. CurroSci. 71, 205-213.
Rawyler, A., Roelofsen, B. and Op den Kamp, J. A. F.(1984). The use of fluorescamine as a permeant probeto localize phosphatidylethanolamine in intact Frienderythroleukaemic cells. Biochim. Biophys. Acta 769,330-336.
Ruetz, S. and Gros, P. (1994). Phosphatidylcholinetranslocase: a physiological role for the mdr2 gene.Cel!, 1071-1081.
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989).Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press, New York.
Sanglard, D., Ischer, F., Monod, M. and Bille, J. (1997).Cloning of Candida albicans genes conferring resist-ance to azole antifungal agents: characterisation ofCDR2, a new multidrug ABC transporter gene.Microbiology 143,405-416.
Schroit, A. J. and Zwaal, R. F. A. (1991). Transbilayermovement of phospholipids in red cell and plateletmembranes. Biochim. Biophys. Acta 1071, 313-329.
Tang, X., Halleck, M. S., Schlegel, R. A. andWilliamson, P. (1996). A subfamily of P-type ATPaseswith aminophospholipid transporting activity. Science272, 1495:-1497.
.
Utsugi, T., Schroit, A. J., Connor, J., Bucana, C. D. andFidler, I. J. (1991). Elevated expression of phospha-tidylserine in outer membrane leaflet of human tumorcells and recognition by activated human bloodmonocytes. Cancer Res. 51, 3062-3066.
Van Helvoort, A., Smith, A. J., Sprong, H., Fritzche, I.,Schinkel, A. H., Borst, P. and Van Meer, G. (1996).MDR I P-glycoprotein is a lipid translocase of broadspecificity, while MDR3 P-glycoprotein specificallytranslocates phosphatidylcholine. Cel!, 87, 507-517.
Van den Bossche, H. (1995). Chemotherapy of humanfungal infections. In Lyr, H. (Ed.), Modern SelectiveFungicides. Properties, Applications. Mechanisms ofAction. Gustav Fisher Verlag, Jena, pp. 431-484.
Walsh, T. J., Kasai, M., Francesconi, A., Landsman, D.and Chanock, S. J. (1997). New evidence that Candidaalbicans possesses additional ATP-binding cassetteMDR-like genes: implications for antifungal azoleresistance. 1. Med. Vet. Mycol. 35, 133-137.
White, T. C. (1997). Increased mRNA levels of ERGJ6,CDR, and MDRJ correlate with increased azoleresistance in Candida albicans isolates from apatient infected with human immunodeficiency virus.Antimicrob. Agents Chemother. 41, 1482-1487.
Williamson, P. and Schlegel, R. A. (1994). Back andforth: the regulation and function of transbilayerphospholipid movement in eukaryotic cells. Mol.Memb. Bioi. 11, 199-216.
Yeast 15,111-121(1999)