antimicrobial and cell lytic peptidesshodhganga.inflibnet.ac.in/bitstream/10603/16634/8/08_chapter...
TRANSCRIPT

ANTIMICROBIAL AND CELL LYTIC PEPTIDES

2
1 . 1 INTRODUCTION
During the past fifteen years, proteins and peptides that have potent antimicrobial
and/or cytolytic activity have been isolated in pure form from organisms all along the
evolutionary scale. The ubiquitous presence of these molecules.implies a multitude of defensive
and offensive functions for them. Whereas the antimicrobial proteins and peptides kill the
invading microorganisms and provide protection to the host, peptide toxins perform offensive
functions. However, the size, structure and effects of many proteins and peptides are so
variable, - some of them act by interfering ~ith the metabolic activities of the cells while .
others by altering the membrane permeability - that common molecular determinants of
cytotoxic action and their target specificities have not become apparent, except perhaps
implicating the amphiphilic structural features in membrane-perturbing activity of many
cytolytic peptides. Some of the agents show very narrow target cell specificity and many are
indiscriminate in their cytotoxicity.
This chapter reviews the work on some of the well studied antibacterial and hemolytic
peptides that act by membrane permeabilization as well as the earli.er work on seminalplasmin
(SPLN), an antimicrobial peptide present in bovine seminal plasma. The structure-function
correlations of SPLN forms the subject of the thesis.
1 . 2 ANTIBACTERIAL PROTEINS AND PEPTIDES
Granular proteins and peptides from phagocytes:
Phagocytosis is one of the important mechanisms of the host defence network in animals.
The polymorphonuclear neutrophils (PMNs) are the most abundant phagocytic cells circulating
in the human body and these cells kill the engulfed bacteria by both oxidative and non-oxidative
mechanisms. The oxygen independent killing depends on several large proteins such as
bactericidal-permeability increasing protein (BPI) and cathepsin, as well as a group of small
peptides called defensins.

3
Bactericidal/permeability increasing protein
Bactericidal/permeability increasing protein (BPI) is a non-catalytic, lysine rich
protein with a mol. wt. of 50-60 kDa and has been purified from human and rabbit PMNs
(Weiss et al., 1978; Gray et al., 1989). The proteins from both the sources share similar
structure and function. On a molecular basis, BPI is the most potent of the mammalian
cytotoxins. BPI acts on numerous gram-negative bacterial species but does not seem to affect
gram-positive and eukaryotic cells. This remarkable specificity of BPI to gram-negative
bacteria arises due to its high binding affinity to the outer envelope lipopolysaccharide (LPS).
This property has also been exploited for its purification.
BPI seems to exert its activity in two stages. The first stage is characterized by a
reversible bacteriostatic activity involving discrete outer membrane alterations leading to
permeability changes. The second stage involves the irreversible growth arrest as a result of
impairment of biochemical functions linked to the intact cytoplasmic membrane (Mannion et al.,
1989).
The functional domains of human BPI have been dissected by limited proteolysis. All the
biological effects of BPI are reproduced by a 25 kDa N-terminal fragment. The C-terminal
portion of BPI is devoid of antibacterial activity but may serve as an anchor into the granule
membrane (Weiss and Olsson, 1987).
The action of BPI on its target cells is strikingly similar to that of intact PMN. This
suggests that BPI has a prominent role in the antimicrobial functions of PMN against BPI
sensitive gram-negative bacteria.
Defensins
Defensins constitute the most abundant (- 30%) of antibacterial peplides of the
cytoplasmic granules of the phagocytes (Greenwald and Ganz, 1987). The defensins are highly
conserved, disulfide linked, cationic peptides with a molecular weight < 4000. They exhibit

4
antimicrobial activity against gram-negative and gram-positive bacteria, fungi, mammalian
cells and enveloped viruses. They have been isolated from PMNs of humans, rabbits, guinea
pigs and rats and the lung macrophages of rabbits (Lehrer et al., 1991 ).
The amino acid sequences of fifteen of these, basic, arginine - and cystine -rich
peptides have been determined (Table 1.1 ). Defensin (HNP-3) has a dim eric ~-structure and
an architecture different from other lytic peptides. The structural aspects of defensins from
the point of view of their activities have been discussed in greater detail in Chapter 3.
Defensin-like peptides have recently been isolated from insects (Hoffmann and Hetru,
1992). Todate, seven defensins have been fully sequenced in species belonging to three insect
orders : (A) Diptera, (b) Calcoptera and (c) Odonata.
Bactenecins
Two antibacterial peptides named Bac-5 and Bac-7 (from the general term
bactenecins) have been purified from extracts of granules of bovine neutrophils. They have
molecular weights of 5000 and 7000 respectively (Gennaro et al., 1989). They are rich in
proline and arginine. Proline constitutes about 45% and arginine 20% of the amino acid
residues and the remaining are mainly hydrophobic amino acids like isoleucine, leucine and
phenylalanine. They exhibit a potent bactericidal activity towards several gram-negative
bacteria in vitro. They have been shown to exert their antibacterial activity by increasing the
permeability of both the outer and inner membrane of gram-negative bacteria (Skerlavaj et
al., 1990; Gennaro et al., 1989).
A smaller bactenecin of molecular weight ·(2000 exhibiting bacteriolytic activity
against both E. coli and S. au reus at 1 o-7 - 1 o-5 M concentration has been purified and its
sequence (shown below) has also been determined (Romeo et al., 1988).
RLCRIVVIRVCR l I

Source
Human
Guinea
pig
Rabbit
Table 1.1
Comparison of defensin sequences
HNP-1
HNP-2
HNP-3
HNP-4
GPNP
NP-1
NP-2
NP-3a
NP-3b
NP-4
NP-5
Sequence
ACYCRIPACIAGERRYGTCIYQGRLWAFCC
CYCRIPACIAGERRYGYCIYQGRLWAFCC
DCYCRIPACIAGERRYGTCIYQGRLWAFCC
VCSCRLVFCRRTELRVGNCLIGGVSFTYCCTRV
RRCICTTRTCRFPYRRLGTCIFQNRVYTFCC
VVCACRRALCLPRERRAGFCRIRGRIHPLCCRR
VVCACRRALCLPLERRAGFCRIRGRIHPLCCRR
GICACRRFCPNSERFSGYCRVNGARYVRCCSRR
GRCVCRKQLLCSYRERRIGDCKIRGVRFPFCCPR
VSCTCRRFSCGFGERASGSCTVNGVRNTLCCRR
VFCTCRGFLCGSGERASGSCTINGVRHTLCCRR
Rat RatNP-1 VTCYCRRTRCGFRERLSGACGYRGRIYRLCCR
RatNP-2 VTCYCRSTRCGFRERLSGACGYRGRIYRLCCR
RatNP-3 CSCRTSSCRFGERLSGACRLNGRIYRLCC
RatNP-4 ACYCRIGACVSGERLTGACGLNGRIYRLCCR
5

6
lndolicidin
A potent and structurally novel antimicrobial peptide has been isolated from cytoplasmic
granules of bovine neutrophils (Selsted et al., 1992). 10 Jlg/ml of this peptide, named
indolicidin, has been found to virtuatty sterilize suspensions of gram-positive S. aureus and
gram-negative E. coli. The peptide was found to be very rich in tryptophan and amidated at the
C-terminal. Tryptophan residues have been thought to play an important role in the function of
this unique peptide. Th~ sequence of this peptide is shown below.
ILPWKWPWWPWRR-NH2
Antimicrobial peptides from amphibians
Magainins:
African clawed frog Xenopus laevis secretes a broad spectrum of oligopeptides, two of
which have been identified as strong antimicrobial agents and termed as magainins (Bevins and
Zasloff, 1990). The absence of infection during the healing of surgical wounds of these animals
prompted the search for these antimicrobial agents. The magainin peptides are each 23 amino
acids long and lack sequence identity with any other reported peptide (Table 1.2). They exert
antimicrobial activity against bacteria, fungi and protozoa at molar concentrations comparable to
other known antimicrobial agents.
Several other peptides having antimicrobial activity including Xenopsin precursor
fragment (XPF), caerulein precursor fragment (CPF) have also been identified (Bevins and
Zasloff, 1990) (Table 1 .2). All these peptides show antibiotic activity similar to magainins,
although they differ in their spectrum of activity. They are all 25 or 26 amino acid residues in
length and are lysine-rich basic polypeptides lacking tyrosine and lysine. Even though the
sequences display limited sequence identity except for the similarity between PGLa, XPF and
levitude precursor fragment (LPF), all of them can be configured as amphiphilic alpha helices

Source
Xenopus
laevis
Bombina
variegata
Bombina
orienta/is
Phylomedusa
sauvagii
Table 1.2
Antimicrobial peptides from amphibians
Name of
peptide
Magainin 1
Magainin 2
PGLA
XPF
LPF
CPF
Bombinin
Bombinin
related pep
tide ·(B RP)
Bombinin like-
Sequence
GIGKFLHSAGKFGKAFVGEIMKS
GIGKFLHSAKKFGKAFVGEIMNS
GMASKAGAIAGKIAKVALKAL-NH2
GWASKIGQTLGKIAKVGLKQLIQPK
GWASKIGQTLGKIAKVGLQGLMQPK
GFGSFLGKALKAALKIGANALGGSPQQ
GIGALSAKGALKGLAKGLAEHFAN-NH2
GIFFALLSAAKVGLKGLAKGLAEHFAN
peptide 1(BLP 1)GIGASILSAGKSALKGLAKGLAGHFAN-NH2
B L P- 2 GIGSAILSAGKSALKGLAKGLAGHFAN-NH2
B L P-3
8 L P-4
Dermaseptin
GIGAAILSAGKSALKGLAKGLAEHF-NH2
GIGAAILSAGKSIIKGLANGLAEHF-NH2
ALWKTMLKKLGTMALHAGLAALGAAADTISQ
GTQ-OH
7

8
which direct their surface activity and membrane affinity. Magainins form random coils in
aqueous solution but adopt a-helical structure in aqueous trifluoroethanol and upon interaction
with phosphatidyl serine vesicles and other micelles (Matsuzaki et al., 1989). Magainin 2 has
been shown to organise into macroscopic filamentous structures, presumably reflecting the
ability of the monomeric peptide to self aggregate (Urrutia et al., 1989). The antibacterial
activity of magainins is presumed to arise from their interaction with biomembranes.
Magainins have also been shown to rapidly depolarize the bacterial cell membrane and uncouple
oxidative phosphorylation. Magainin 2 has been shown to interact with synthetic lipid bilayers
to form anion selective channels (Duclohier et al., 1989).
Bombinin, bombinin-related peptides and bombinin-like peptides:
An antimicrobial peptide called 'bombinin' was characterzed by Csordas and Michl
(1969) from the European toad, Bombina variegata. This is a 24 amino acid residue peptide
(Table 1.2) with hemolytic and antibacterial properties and shows many of the general
structural feature of magainins.
A family of bombinin-related peptides with 27 amino acids has also been described by
Simmaco et al., (1991 ). One of them has been synthetically made and shown to possess
antibacterial activity.
Bombinin-like peptides having antimicrobial and hemolytic activity have been reported
from skin secretions of the Asian toad Bombina orienta/is. Three of the peptides BLP-1 to 3 have
been sequenced. All the three peptides share considerable but not complete homology with
bombinin (Gibson et al., 1991 ).
Dermaseptin
A highly potent antimicrobial peptide from skin extracts of the South American arboreal
frog, Phylomedusa sauvagii, which acts against pathogenic fungi has been reported. This peptide
called dermaseptin is 37 amino acid residue long. The peptide is non-hemolytic. The

9
sequence(Table 1.2)of this peptide has the potential to form amphiphilic a-helical structure
and probably acts by membrane permeabilization (Mor et al., 1991 ).
Antimicrobial peptides from insects
Cecropins:
Insects possess efficient immune systems, both cellular and humoral. The immune
systems of Hyalophora cecropia and other silkworms respond very effectively to bacterial
infections by induced synthesis of proteins and peptides consisting of a mixture of lysozyme and
two antibacterial protein families, named cecropins and attacins (Boman and Huttmark, 1987).
Cecropins are small, basic proteins with a broad spectrum of antibacterial activity against both
gram-positive and gram-negative bacteria. The primary structure of the three major cecropins,
cecropin A, cecropin B and cecropin D have been determined. Cecropin sequences from flesh fly
Sarcophaga, the silkworms Bombyx and the tobacco horn worm Manduca have also been reported
(Boman et al., 1991 ). During a search for mammalian antibacterial peptides, a cecropin
homolog was isolated from pig intestine and this shows 64-75% homology with insect cecropin
(Lee et al., 1989). The sequences of 11 known cecropins are given in the Table 1.3. The
sequences are characterized by a strongly basic N-terminal region and a long hydrophobic
stretch in the C-terminal half. Some other typical conserved features include, tryptophan at
position 2, single and double lysine at positions 5, 8 and 9 arginine at position 12. In the N
terminal region, polar and hydrophobic amino acids are interspaced in a regular pattern. NMR
studies have demonstrated the presence of two amphipathic a-helices interrupted by a hinge
region composed of Ala-Giy-Pro-sequence (Holak et al., 1988).
The antibacterial activity of the three main forms of cecropin has been compared
(Hultmark et al., 1983). Cecropin A and B are highly active against several gram-positive and
gram-negative bacteria while the D form shows high activity against E. coli and Acinetobacter
calcoacitans and several other insect pathogens Cecropins have been shown to lyse bacteria but
not eukaryotic cells (Steiner et al., 1981 ). Cecropins also lyse vesicles composed of

10
Table 1.3
Comparison of 11 known cecropins
------------------------------------------------------------------Manduca B-2 W--NPFKEL ERAGQRVRDA VTSAAPAVAT VGQAAAIAR*
Hyalophora D W--NPFKEL EKVGQRVRDA VISAGPAVAT VAQATALAK*
Antheraea D W--NPFKEL ERAGQRVRDA IISAGPAVAT VAQATALAK*
Hyalophora A KW--KLFKKI EKVGQNIRDG IIKAGPAVAN VGQATQIAK*
Bombyx CMIV RW--KIFKKI EKVGQNIRDG IVKAGPAVAV VGQAATI*
Bombyx A RW--KIFKKI EKMGRNIRDG IVKAGPAIEV LGSAKAI*
Antheraea B KW--KIFKKI EKVGRNIRNG IIKAGPAVAV LGEAKAL*
Hyalophora B KW--KVFKKI EKMGRNIRNG IVKAGPAIAV LGEAKAL*
Drosophila A GWLAKKIGKKI ERVGQHTRDA TI-QGLGIAQ QAANVAATAR*
Sarcophaga lA GWLKKIGKKI ERVGQHTRDA TI-QGLGIAQ QAANVAATAR*
Porcine P1 SWLSKTAKKL ENSAK-KR---ISEGIAIAI QGGPR
* Indicates amidated C terminus

11
phospholipids which are negatively charged .or zwitterionic in a stoichiometric way (Steiner et
al., 1988).
Antibacterial peptides from horseshoe crabs
Tachyplesins I and II and polyphemusin I and II, cationic peptides isolated from the
hemocytes of horseshoe crabs show antibacterial activity with similar efficiencies for both
gram-negative and gram-positive bacteria (Miyata et al., 1989). Tachyplesin I, a 17 residue
peptide was found in the hemocytes of Japanese horseshoe crab Tachypleus tridentatus as an
inhibitor of the bacterial lipopolysaccharide (LPS)-mediated activation of factor C, the
initiation factor in the limulus clotting cascade. Tachyplesin Ill was isolated from the south east
Asian horseshoe crab (Muta et al., 1990), Carcinoscorpins rotundacauda and Tachyp/eus gigas.
Polyphemusins I and II were isolated from the American horseshoe crab Limulus polyphemus.
The sequences of these related peptides are shown in Table 1.4.
These peptides are characterised by two intramolecular disulfide bridges and three
tandem repeats of a tetrapeptide, hydrophobic amino acid-Cys-hydrophobic amino acid-basic
amino acid, suggesting an amphiphilic nature of the peptide. A two dimensional NMR study of
tachyplesin I has shown it to form a rigid antiparallel ~- sheet structure.
Tachyplesins exhibit broad spectrum antimicrobial activity (Nakamura et al., 1988;
Miyata et al., 1989). The strong binding of these peptides to LPS cannot explain activities for
gram-positive bacteria and fungi. The mechanism of action thus appears to be different from
that of polymyxin B. Tachyplesin I has been shown to interact with lipid bilayer membranes
(Matsuzaki et al., 1991 ). The interaction induces permeability changes and results in the
leakage of entrapped calcein suggesting that the peptide~induced membrane permeability changes
may bring about its antibacterial activity. Tachyplesin indeed causes a rapid efflux of K+ ions
from E. coli cells. Subinhibitory concentration of the peptides was shown to sensitize gram
negative bacteria to the bactericidal action of novobiocin and nalidixic acid showing that
tachyplesins permeabilise the outer membrane of bacteria. As polymyxin B resistant bacteria

Tachyplesin
Tachyplesin II
Tachyplesin Ill
Polyphemusin
Polyphemusin II
Table 1.4
Antimicrobial peptides from horseshoe crabs
KWCFRV<(YRGI<(YRKtR - NH2
RWCFRVCYRGICYRRtR - NH2 l l
KW&;RV<(YRG1fYRR6R - NH2
RRW6FRVCYRGFCYRKCR - NH2 ' l
RRWtFRVCYKGFCYRK~R - NH2 h ,
l2

13
are also susceptible to the action of tachyplesins, the target of its action seems to be
constituents in the outer membrane other than LPS (Ohta et al., 1992).
Plant antimicrobial peptides
Two antimicrobial peptides designated as Mj-AMP1 and Mj-AMP2 have been isolated
from seeds of Mirabilis jalapa (CamrrJUne et al_11992). These peptides are highly basic and 37
and 36 residues in length. Both peptides contain three disulfide bridges and differ from one
another only by 4 amino acids.
The peptides appear to associate into dimers in the native form. Mj-AMPs exhibit broad
spectrum antifungal activity and also act on gram-positive bacteria. They are non-toxic to gram
-negative bacteria and cultured human cells. Although Mj-AMPs show sequence homology to J.L
agatoxins a class of insecticidal neurotoxic peptides isolated from venom of spiders, they do not
possess any neurotoxic effect.
Mj-AMPs, have structural similarities to defensins and sequences are shown below.
Mj-AMP1
Mj-AMP2
OCIGNGGRCNENVGPPYCCSGFCLRQPGGYGYCKNR
CIGNGGRCNENVGPPYCCSGFCLROPN.QGYGVCRNA
1.3 HEMOLYTIC PEPTIDES
Mel itt in
Structural aspects
Melittin is the main toxic component of the venom of the honey bee, Apis me/litera
(Haberman, 1972). It is a short, 26 residue peptide and has multiple effects on membrane
structure and functions. It is perhaps the best studied membrane perturbatory peptide. Even
then the mechanism of membrane lysis still remains to be proven and no consensus exists on the
nature of its interaction with membrane lipids. The sequence (Table 1.5) of melittin appears to
have two structural regions (De Grado et al., 1982). The No-terminal 20 amino acids have the

Peptide
Melittin
Mastoparan
Bombolitin
Bombolitin II
Bombolitin Ill
Bombolitin IV
Bombolitin v
o-toxin
(human ·strain)
Table1.5
Sequences of cytolytic peptides
Sequence
GIGAVLKVLTTGLPALISWIKRKRQQ-NH2
INLKALAALAKKIL-NH2
I KITTM LA K LG KV LAH V-Nti2
SKITDILAKLGKVLAHV-NH2
rKIMDILAKLGKVLAHV-NH2
INIKDILAKLVKVLGHV-NH2
INVLGILGLLGKALSHL-NH2
formyl MAQDilSTIGDLVKWIIDTVNKFTKK
o-t ox in formyl MAADIISTIVEFVKLIAETVEKFIKK
(canine strain)
14

15
potential to form an amphiphilic a-helix with a proline link at position 14 and the C-terminal
hexapeptide is highly cationic with 2 lysines1 2 arginines and amidated o- terminus. Neither the
peptide fragment 1-20 nor the C-terminal!hexapeptide shows hemolytic activity (Dawson et
al., 1978; Schroder et al., 1971 ).
Melittin exists as a monomer both at low concentration and low ionic strength but
aggregates at high concentration and high ionic strength, forming tetramers as the only
detectable form (Brown et al., 1980). The structure of melittin has been investigated by X-ray
diffraction (Terwilliger and Eisenberg, 1982), NMR (Pastore et al., 1989), CD and many
other techniques. The monomer has been found to be in a-helical conformation with a bend at
proline 14. The crystal structure of the tetramer contains pairs of dimers facing each other
along the hydrophobic faces thus shielding them from the aqueous environment. Within each
dimer the monomers are arranged antiparallel to each other , minimising the electrostatic
repulsion between the cationic C-termini.
Hemolytic activity of melittin
Melittin is known for its marked cytolytic activity. Its hemolytic activity has been
extensively studied. Although considerable controversy exists regarding the aggregation state of
melittin bringing about hemolysis, it has been well established that binding of the peptide to the
membrane is essential for bringing about lysis. The apparent dissociation constant has been
found to be 3 x 10-8M and a maximum binding capacity of 1.8 x 107 molecules/cell (Tosteson
et al., 1985). Melittin induced hemolysis follows reproducible biphasic kinetics - a fast phase
of hemoglobin release triggered by binding of melittin to the membrane followed by a slow steady
state release.
Studies done by Tosteson et al.1suggest that the hemolysis by melittin occurs by a colloid
osmotic mechanism. The evidence for such a conclusion include (i) hemoglobin release from the
melittin treated cells is preceded by 200-2000 fold increase in permeability to Na+ and K+
ions (ii) erythrocytes show swelling before the release of hemoglobin and (iii) osmotic
protectants of higher molecular weights are capable of providing protection to lysis by melittin.

16
In order to understand the structure-function correlations of melittin, various synthetic
analogs have been used. Studies done with chemically modified or synthetic analogs have shown
that hemolytic activity and surface activity are not directly correlated. Studies done by De Grado
et al., (1982) using a model peptide which is homologous to melittin only at five positions (6,
9, 13, 16 & 19) in the N-terminal hydrophobic stretch and the C·terminal hexapeptide showed
that neither the sequence nor the presence of proline in the centre of the helix are required for
the hemolytic activity. The model peptide had been designed to have a higher potential to form an
amphipathic a-helix than melittin and composed solely of leucine on the hydrophobic side of the
helix and of serine and glutamine as hydrophilic residues. These results showed that the N
terminal 20 residues have only a structural role which can be duplicated by non homologous
sequence having a potential to form a amphiphilic a-helix. A synthetic analog of melittin
(Ala 14 melittin) containing a Pro to Ala substitution at residue 14 also showed 2.5 times
higher hemolytic activity than melittin confirming the lack of importance of the helix bend for
the hemolytic activity of melittin (Dempsey, 1990). But this analog had poorer voltage
dependent ion channeJ activity than native melittin suggesting the importance of the central
flexible region for the stability of voltage-gated channels. This also supports the idea that
hemolytic activity of melittin is not directly related to its channel forming property.
Formylation of N-terminal methionine did not result in the loss of hemolytic activity of
melitrin (Lubke et al., 1971 )indicating that free N-terminal a-amino group is not essential
for activity. Photooxidative destruction of tryptophan or replacement of tryptophan with
leucine resulted in reduced hemolytic activity suggesting an important role for tryptophan in its
hemolytic activity (Haberman and Kowallek, 1970; Blonde lie and Houghton, 1991 a).
Two models have emerged from the structural and functional studies for the molecular
mechanism of action. In one type of model, melittin forms aqueous channels by spanning the
membrane forming channel oligomers exposing its hydrophobic side to the lipid and its
hydrophilic face to the aqueous pore (Tosteson and Tosteson, 1981 ). The pore size of these
channels will not be big enough to allow the hemoglobin to pass through. The model predicts that

17
these channels produce ionic imbalance which leads to colloid-osmotic lysis. Some of the
experimental data which support such a mechanism have been mentioned earlier. An analog of .
melittin lacking Lys7 (Biondelle and Houghton, 1991b) lacksits activity. The omission of
lysine 7 in the analog is likely to cause formation of smaller pore channels and make it
selecNve for anions over cations. Replacement of lysine 7 with leucine also results in loss of
activity. These two observations also are in agreement with the melittin pore theory.
The second set of models proposes that melittin produces lysis by disrupting the
phospholipid structure of the membrane by forming a wedge like conformation (Dawson et al.,
1978). The resulting weakening of the overall membrane structure allows the leakage of
hemoglobin.
Several specific models of this type have been proposed (Dawson et al., 1978; DeGrade et
al., 1982; Terwilliger et al., 1982). The observations suggesting that melittin lies parallel to
the membrane plane or that melittin penetrates only a few angstroms into the bilayer tend to
support this type of model (De Grado et al.11982; Terwilliger et al, 1982).
Mastoparan
Mastoparan is a toxic tetradecapeptide (Table 1.5) isolated from wasp venom (Hirai et
al., 1979). It stimulates exocytosis from several mammalian cells, of histamine from mast
ceils, serotinin from platelets and insulin from pancreatic islet via a mechanism that involves
activation of GTP-binding regulatory proteins (G-proteins) (Higashijima et al., 1988).
Mastoparan was first isolated from Varpula lewisii and chemically synthesised by Hirai and co-
workers in 1979 (Hirai et al., 1979).
Mastoparan has also been found to bind to calmodulin with high affinity. Like melittin,
mastoparan is also potent stimulator of phospolipase activity (Male nick and Anderson, 1983).
However melittin is a more potent hemolytic agent than mastoparan. Mastoparan exists in a
random coil conformation in aqueous solution but takes up a a-helical conformation in
hydrophobic environment (Higashijima et al., 1983). Mastoparan was shown to increase the

18
cation permeability of planar lipid bilayers, indicating its possible ionophoric properties
(Okumura et al, 1981 ). Mellor and Sansom (1990} have demonstrated the existence of two
types of voltage dependent ion channel property of mastoparan in planar lipid bilayers. In the
presence of 0.3 - 3.0 J..LM mastoparan, type I activity was characterized by discrete channel
openings exhibiting multiple conductance levels in the range of 15-700 ps. Type II activity was
characterized by transient increases in the bil~yer conductance upto a maximum of about 650
ps. Mastoparan channels are weakly cationic in character. A twelve residue analog of
·mastoparan lacking the N-terminal dipeptide 1-N also showed Type I form of ion channel
activity.
Bombolitins
Five structurally related heptadecapeptides which are hemolytic have been discovered in
the venom of bumblebee, Megabombus pennslyvanicus . The sequences of these peptides, named
bombolitins, have also been determined (Argiolas and Pisano, 1985) (Table 1.5).
In addition to their structural similarity they have functional similarities also. They
lyse erythrocytes and liposomes, release histamine from rat peritoneal mast cells and stimulate
phospholipase A2 from different sources. Bombolitin V has been found to be the most potent of
hemolytic agents and its activity compares very well with that of melittin. Its mast cells
degranulating activity is also five times higher than mastoparans.
The conformationsof bombolitin I & Ill have been determined by using circular dichroism
(CD) nuclear magnetic resonance (NMR) and computer simulations (Bairaktani et al., 1990).
Bombolitin was found to lack an observable secondary structure in aqueous solution independent
of temperature, pH and concentration. In striking contrast, bombolitin Ill adopts a well defined
a-helix at concentrations greater than 3 mM. The a-helix of bombolitin was found to be pH
dependent, with a great decrease in the observed secondary structure at pH values below 3.5.
This was probably due to protonation of the Asp residue which could prevent the formation of
salt bridges and consequent molecular aggregation and secondary structure. In the presen-ce of

19
SDS micelles, the characteristic bands in CD spectra indicated an a-helix content of 60% for
bombolitin Ill and 70% for bombolitin I. When the sequences of bombolitin are depicted in the
a-helical conformation they appear amphiphilic. The amphiphilic nature of bombolitin
probably determines their biological activities.
Delta toxin [a-toxin, a-hemolysin]
Staphylococcus aureus secretes several peptide toxins that are membrane active. Delta
lysin is the smallest of them with 26 residues and has been extensively studied. Delta lysins
from two different strains of S. aureus have been sequenced (Table 1.5).
The structural requirements of a-toxin to show its activity have been investigated by
Alouf et al. (1989} by synthesizing the sequence identical to the natural toxin from the human
strain along with five 26 residue analogs and two shorter peptides corresponding to residues 1-
11 and 11-26. The 26 residue analogs were designed in such a way that they maintain (a) the
hydrophobic/hydrophilic balance, (b) the a-helical conformation and, (c) common epitopic
structure. These analogs were, (i) deformylated toxin, (ii) an idealised amphipathic sequence
for residues 1-20, (iii) an analog which closely resembles (ii) but with the tryptophan moved
from position 16 to position 5, (iv) unformylated peptides of the above analog, (v) F-moc
analog.
All 26-residue analogs were hemolytically active. The peptide 11-26 bound weakly to
lipids and was weakly hemolytic. The peptide 1-11 did not bind to lipids and was not hemolytic.
a-hemolysin shares many of its membrane-modifying properties with melittin. Both the
peptides are 26 residues in length with a single tryptophan residue and lacking cystine. Both
peptides ·have potential to form amphipathic helices with a cationic tail. They lyse erythrocytes
and many other mammalian cells including fibr:oplasts, intracellular organelles and bacterial
protoplasts. They bind to phospholipids and penetrate phospholipid monolayers and induce
permeability changes in phospholipid vesicles. Both of them activate phospholipase A2 of 3T3

mouse fibroblasts. Both of them have similar solubilities in organic solvents, aggregate to
multimeric forms and are heat stable.
However, there are many differences between these and toxins. Their amino acid
sequences do not show any homology and differ in the distribution of charges. a-toxin does not
have a long hydrophobic sequence. The functional differences include, (i) melittin is more
hemolytic than a-toxin, (ii) melittin shows a preference for negative lipids whereas o-toxin
has no such preference, (iii) melittin causes aggregation of gel phase liposomes unlike o-toxin.
Specificity of antibacterial and hemolytic peptides
Although amphiphilic a-helix or amphiphilic p-structure motif has been identified as
an essential feature of antibacterial and cell lytic peptides which act by membrane
permeabilization, nothing is known as to what determines their target specificity. For example,
cecropins and magainins are relatively less active against mammalian cells but more specific on
bacterial cells. Defensin on the other hand has broad cytotoxicity which include bacteria, fungi
and tumour cells. No clear cut rules exist to explain the broad cytotoxicity of melittin. o-lysin
for example is highly cytotoxic but relatively less antimicrobial (Dhople and Nagaraj, 1993).
A large number of functional properties have been attributed for amphiphilic helices.
1 . 4 SEMINALPLASMH'J
Seminalplasmin (SPLN) is a protein present in bovine seminal plasma that is known to
exert antimicrobial effect (Reddy and Bhargava, 1979). The presence of an inhibitory factor of
spermatozoal RNA synthesis in bovine seminal plasma had been postulated by Bhargava and
colleagues from the observation that bovine seminal plasma inhibited the synthesis of RNA by
spermatozoa. Preliminary studies had also indicated this factor to be macromolecular and
susceptible to proteolytic activity. As the RNA synthesis in spermatozoa was found to be
exclusively mitochondrial (Premkumar and Bhargava, 1972& 1973) and susceptible to the
same inhibitors as the corresponding synthesis in prokaryotes, it was reasoned that the
inhibitory factor in seminal plasma would act as an antibacterial agent. Bovine seminal plasma

21
indeed inhibited the growth of E. coli. Using this as the assay system, a protein named
seminalplasmin (SPLN) was purified from dialysed bovine seminal plasma (Reddy and
Bhargava, 1979).
Extensive work has been done by Bhargava and co-workers on SPLN Following are some
of the properties of SPLN.
( i ) It has been shown to be a potent antimicrobial agent acting on a variety of gram-positive
and gram-negative bacteria as well as yeast (Bhargava, 1981, Reddy and Bhargava,
1979; Scheit et al., 1985; Scheit and Bhargava, 1985). It is bacteriostatic at low
concentration and bactericidal at high concentrations. It appears to activate autolysins in
E. coli at higher concentrations (Chitnis et al., 1987). The effect of SPLN on the growth
of various microorganisms is tabulated in Table 1.6.
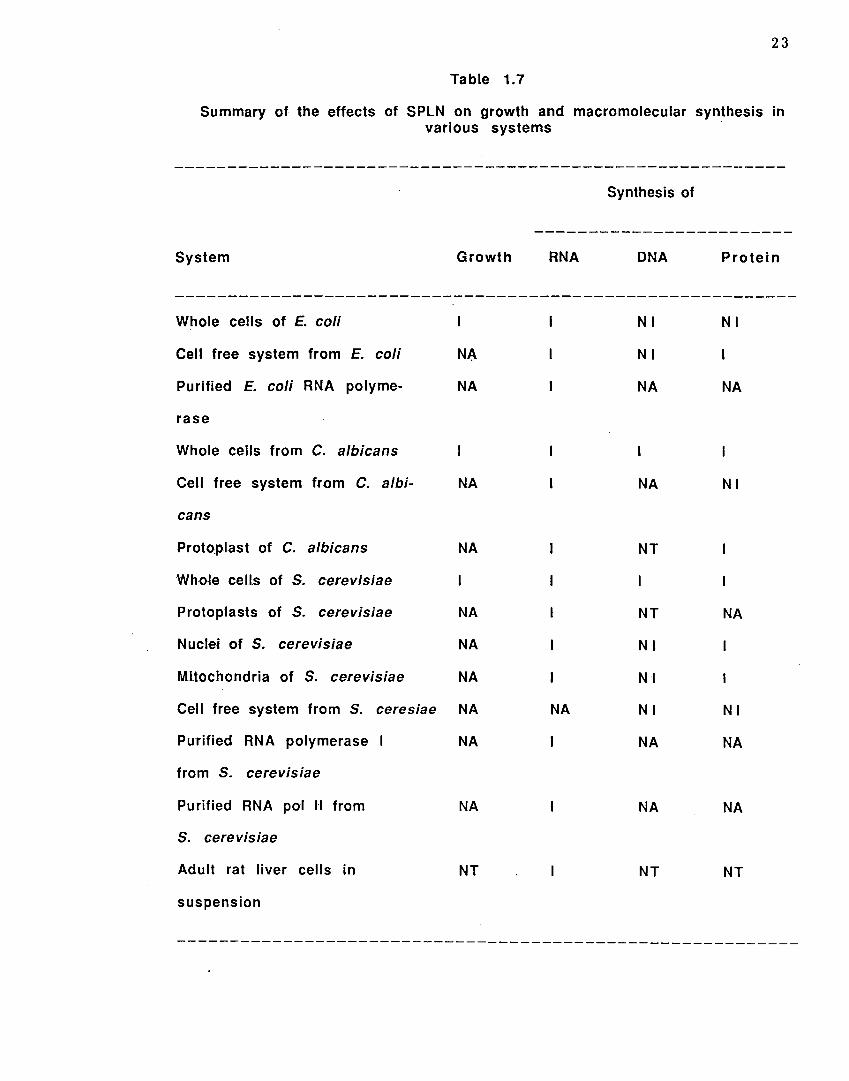
( i i ) The effects of SPLN on the synthesis of macromolecules have been studied in whole cells,
protoplasts, mitochondria and cell free systems (Table 1.7). The authors have concluded
that SPLN is likely in to penetrate into susceptible cells and inhibit RNA synthesis
(Reddy and Bhargava, 1979; Scheit and Bhargava, 1985; Scheit et al., 1979 & 1985).
(iii) Studies on the interaction of SPLN with bovine spermatozoal plasma and acrosomal
membranes have shown that SPLN binds to both of these membranes .increasing their
fluidity (Shivaji, 1986).
( i v) SPLN incorporates into lipid bilayers of phosphatidylcholine and phosphatidic acid
vesicles and as a consequence of such an interaction the a-helical content of SPLN increaseS
(Galla et al., 1985).
( v ) SPLN binds to calmodulin and acts as a powerful calmodulin antagonist (Gietzen and Galla,
1985). SPLN forms a ca2+ dependent 1 :1 complex with calmodulin which is resistant to
urea (Comte et al., 1986).

Table 1.6
Effect of SPLN on the growth of microorganisms
Microorganism
E. coli B
E. coli K -1 2
E. coli 1 6 0 . 3 7
Euterobacter aerogenous
Streptococcus faccalis
Cryptococcus neoformans
Salmonella typhemurium
Candida albicans 1
Candida a1bicans (din)
Bacillus subtilis
Saccharomyces cerevisiae SM202
S. cerevisiae Y V 11 0 6
Proteus vulgaris
Pseudomonas aeruginosa
Minimum inhibitory concentration of
SPLN J.Lg/ml
25-40
2 5-4 0
40
100
100
25
400
4 0
1 0 0
20
240
14
5
20
22
23 .
Table 1.7
Summary of the effects of SPLN on growth and macromolecular synthesis in various systems
System
Whole cells of E. coli
Cell free system from E. coli
Purified E. coli RNA polyme-
rase
Whole cells from C. albicans
Cell free system from C. albi-
cans
Proto:p.last of C. albicans
Wh-o~e ceiJs o-f S. cerevisiae
Protoplasts of S. cerevisiae
Nuclei of S. cerevisiae
Mitochondria of S. cerevisiae
Growth
NA
NA
NA
NA
NA
NA
NA
Cell free system from S. ceresiae NA
Purified RNA polymerase NA
from S. cerevisiae
Purified RNA pol 1-1 from NA
S. cerevisiae
Adult rat liver cells in NT
suspension
Synthesis of
RNA DNA Protein
Nl Nl
Nl
NA NA
NA Nl
NT
NT NA
Nl
Nl
NA Nl Nl
NA NA
NA NA
NT NT
----------------------------------------------------------

(vi) SPLN binds strongly to DNA and unwinds it (Reddy and Bhargava, 1979; Gopal et al.,
1988). DNA carrying as much as 1 mol. of SPLN per 8 base pairs can however be
transcribed normally by E. coli RNA polymerase (Scheit et al., 1979).
Many of the properties of SPLN appear to be membrane related. Most of the above
studies have been done when the primary structure of SPLN was unknown and hence no
attempts could be made to correlate its activities with its structure.
Objectives of the work discussed in the thesis
The presence of antimicrobial proteins/peptides in organisms all along the
evolutionary scale emphasizes their importance in defence. The primary target of many of
these peptides like defensins, magainins and cecropins is the bacterial membrane. They act by
permeabilizing the membrane. In spite of the broad antimicrobial spectrum of the above
peptides, their use as therapeutic agents has not been thoroughly investigated due to their
relatively long length or complex structure and consequent difficulties in large scale
synthesis. Hence it would be desirable to get short-antimicrobial peptides without cytolytic
activity. One of the pre-requisites for synthesizing short and simple antimicrobial peptdies
without cytotoxic properties is to understand their structure-activity relationship. ln the
case of long peptides, it would be desirable to define the minimal requirements for biological
activity.
SPLN is one of the first antimicrobial peptides to be isolated and characterized from
mammalian sources (Reddy and Bhargava, 1979). SPLN has been postulated by Reddy and
Bhargava to possibly protect the male and/or female reproductive tract against
microorganisms and retroviruses. Recently, a male specific antimicrobial peptide 'andropin'
has been detected in the extracts of male reproductive tract of Drosophila (Samakovlis et al.,
1991 ). Thus, antibacterial peptides may play an important role in protecting the male
reproductive tract. In fact, caltrin (which was later shown to be identical to SPLN) like
24

peptides have been shown to be widely distributed in seminal plasma of various animals and
have been purified and characterized from guinea pig, rat and mouse.
Since SPLN is a noncytotoxic peptide with potent, broad spectrum antimicrobial
activity, determining its structure will conceivably help in studying its structure-activity
relationship. Once the primary structure is known, it can possibly be analysed to understand
the structural basis for its antibacterial activity and delineate regions responsible for it In
this study, SPLN has been used as a model peptide to study structure-activity correlations of
cytolytic and antibacterial peptides. SPLN has been sequenced and its antibacterial activity
further characterized. Two putative segments responsible for its antib~cterial activity have
been identified by analysing its sequence. These segments have been chemically synthesized
and their antibacterial and cell lytic activities characterized. In order to further understand
the structure and charge requirements for their biological activity, several active and
inactive analogs have been synthesised. Interaction o.f SPLN and several of these peptides with
model membranes has also been studied in an effort to rationalize the basis of their biological
activity.
25