alutazione immunoistochimica della risposta …dspace-unipr.cineca.it/bitstream/1889/3388/1/tesi n...
TRANSCRIPT

1
UNIVERSITÀDEGLISTUDIDIPARMADOTTORATODIRICERCAINSCIENZEMEDICO-VETERINARIE
CICLOXXIX
VALUTAZIONE IMMUNOISTOCHIMICA DELLA RISPOSTAIMMUNITARIAINNATAECELLULOMEDIATANELSUINOIN
CORSO D’INFEZIONE NATURALE DA MYCOPLASMAHYOPNEUMONIAECoordinatore:Chiar.moProf.AttilioCorradiTutor:Chiar.maProf.ssaBenedettaPasseri
Dottorando:Dott.NicolaMartinelli

2
RiassuntoIl Mycoplasma hyopneumoniae (MH) è il principale agente causale dellapolmoniteenzooticadelsuino.49suiniconlesionipolmonari,provenientida5diversi allevamenti sono stati campionati almacello. Per ogni animale è stataeffettuataunavalutazionedellelesionipolmonarisecondoilmetodo.Inoltre,daalcunecarcasse, sonostatiprelevati campionidi tessutoadiposoper rilevare iToll like recptor 4.MH è stato ricercato dai campioni di polmone tramiteRT-PCR. Le sezioni di polmone incluso in paraffina sono state valutateistologicamente, classificandole da 0 a 4, per reattività dei BALT, gravità delladisepitelizzazione bronchiale e bronchiolare, e infiammazione subacuta.Tecnichediimmunistochimicasonostateeseguiteusandoanticorpicommercialianti-MAC387perrivelareimacrofagi,antiCD3perilinfocitiT,anti-CD79αperilinfociti B, Foxp3 per rilevare i Treg. La presenza di cellule immunopositive èstata classificata da 0 a 4 a seconda della quantità di cellule colorate per ilinfociti T e B e per imacrofagi. I Treg sono stati contati individualmente neiBALT. I Toll like receptor sono stati evidenziati nel tessuto adiposo congelatoutilizzando un anticorpo commerciale anti-TLR4 su sezioni di 5µm. La mediadelloscoreistologicoèvariatada0,14a1,98.AllaRT-PCRsonorisultatipositivi32campionimentre17sonorisultatinegativi.Neipolmoni sievidenziavaunapolmonite catarrale bronco-interstiziale con aumento di volume del tessutolinfoide peribronchiale, peribronchiolare e perivascolare. 10 campionimostravano un’infiammazione subacuta e, 22, lesioni indotte da MH senzaapparenti infezioni secondarie. Le cellule MAC387 positive erano numerosenelle lesioni polmonari (classificate come 4) con una grave infiammazionesubacuta,mentrelecelluleB,TeTregeranopocorappresentate.Alcontrario,ilinfocitiTeranomoltopresentineiBALTiperplastici(classificati3e4)daMH.Ilinfociti B erano poco rappresentati nelle infiammazioni subacute,mentre neiBALT con aumento di volume indotto da MH sono stati classificati da 2 a 3.L'espressione dei TLR4 è stata valutata su 20 campioni, utilizzando unaclassificazione semi-quantitativa basata su diffusione e intensità dellacalorazioneinquattrolivelli:assente,moderata,media,intensasu4campivisivi.In questo studio, si rielva che i Treg sono molto rari nei BALT iperplastici,suggerendoneunaminorattivitànel ruolodimantenereunabassaattivazionedel sistema immunitario. I macrofagi sono numerosi nell'infiammazionesubacuta,mentesonomenoneiBALTiperplasticidaMH,suggerendounaminorapoptosicellulareneicentrideifollicoli.L'analisi semi-quantitativa dell'espressione dei TLR4 nel tessuto adiposo delmediastinoeffettuata inquesta ricerca, tendea confermarequanto riscontratoin medicina umana: i soggetti che presentano una maggior infiammazionepresentano anche una più alta intensità di colorazione rispetto a quelli conminorinfiammazione;ulterioristudisonocomunquenecessariperconfermarequest'ipotesi.

3
ImmunohistochemicalevaluationofcellmediatedandinnateimmuneresponseinpigsnaturallyinfectedwithMycoplasmahyopneumoniaeAbstractMycoplasma hyopneumoniae (MH) is the main causative agent of enzooticpneumoniaofpigs.49pigs,presentinglunglesions,from5differentherdsweresampledatslaughterhouses.Lunglesionswereindividually.MediastinaladiposetissuewassampledfromsomecarcassestorevealTolllikereceptors4.MHwaslocallyRT-PCRdetected fromeach lung lesion.Paraffin sectionsof lungswerehistologicallyevaluatedwithascore,rangingfrom0to4, forBALT,severityofbronchi, bronchiolar disepithelization and subacute inflammation.Immunohistochemistry was carried out using commercial antibodies anti-MAC387 to detect macrophages, anti-CD3 for T-cells, anti-CD79α for B-cells,Foxp3forregulatoryT-cells.Theamountofantigendetectedwasscoredfrom0to 4 based on the number of positive T, B andmacrophages. T-reg cellswereindividually counted in BALT. To detect Toll-like receptors in mediastinaladipose tissue immunohistochemistry was carried out using a commercialantibodyanti-TLR4onfrozen5µmsection.Theaveragehistologicallungscoreranged from0.14 to1.98.ResultsofRT-PCRwere32positiveand17negativesamples.Lungsshowedcatarrhalbronco-interstitialpneumoniawithincreasedvolumeof peribronchial, peribronchiolar andperivascular lymphoid tissue. 10samples showed subacute inflammation and 22 showed lesions MH inducedwithout secondary apparent bacteria irruption. MAC387 positive cells werenumerous in lung lesions (scored4)affectedbyseveresubacute inflammationwhileB,TandTregcellswerepoorlyrepresented.Onthecontrary,TcellswerenumerousinhyperplaticBALTMHinduced(scored3to4).Bcellswerescarcelyrepresented in subacute inflammatio,while inMH inducedBALT lesionswerescoredfrom2to3.TheexpressionofTLR4wasevaluatedon20samples,usingasemi-quantitativeclassification according to the spread and intensity of staining on four levels:absent,mild,medium,intensiveonfour,4magnificationsfields.ThisstudyshowsthatTregcellsareveryrareinhyperplasticBALT,suggestingalessactiveroleofTregcellsinmaintainingalowactivationofimmunesystem.Macrophages arenumerous in subacute inflammation,while less inMHBALT,suggestingalowgradeofapoptosisofcellsofthefolliclecenter.Semi-quantitative analysis of the presence of TLR4 in mediastinal fat tissue,carried out in this research, tends to confirmdata shown in humanmedicine:those subjectswhoshowedanmore severe inflammation showedahigh stainintensity than those with mild inflammation; further studies are needed toconfirmthishypothesis.

4
Sommario
INTRODUZIONE...............................................................................................5
TOLLLIKERECEPTORS.................................................................................6GENERALITÀ....................................................................................................................6PRIMAIDENTIFICAZIONE..............................................................................................7STRUTTURAELOCALIZZAZIONEDEITLR..................................................................8ELEMENTIRICONOSCIUTIDAITLR.............................................................................9TLR4.............................................................................................................................12TLRNELSUINO............................................................................................................14
RISPOSTAIMMUNITARIAALL'INFEZIONEDAMYCOPLASMAHYOPNEUMONIAE........................................................................................16PARTESPERIMENTALE.............................................................................20INTRODUZIONE............................................................................................................20MATERIALIEMETODI................................................................................................26Prelievodeicampioni............................................................................................26Istologia.......................................................................................................................27Immunoistochimica...............................................................................................27
RISULTATI.................................................................................................................28CONCLUSIONI..........................................................................................................41
ABBREVIAZIONI...........................................................................................44
BIBLIOGRAFIA..............................................................................................45

5
INTRODUZIONEIlsistemaimmunitariopuòesseredefinitocomel'organodeputatoadifenderel'integrità di un organismo dagli attacchi di altri microrganismi; dal suofunzionamentoedallasuaintegritàdipendelasalutediognimammifero.Lostudiodelsistema immunitarioèandatodiparipassocon leconoscenze incampomicrobiologico.Leprimesignificativescopertesonostate fatteverso lafinedell'800conl'individuazionedeimicrorganismicomecausadimalattia.Neitempimoderni lescoperteincampodell'immunitàhannopermessodirendereinoffensivediversemalattieedieliminarnealtre.L'allevamento del suino in Italia ricopre una notevole importanza socio-economica.Attualmentesonoallevatipiùdi8milionidisuini incirca120milaaziendegarantendocosìlamateriaprimaperlaproduzionedisalumieinsaccatidinotevolepregio.Ilsuinoèinoltreritenutol'animaledestinatoadiventarelafabbricadiorganidiricambioperl'uomo.Lesperimentazionieffettuatefinadorahannopermessodiimpiantarecuore,polmonierenisuprimatinonumani.SebbeneinItaliaquestaricercasiastata fermatadaldecreto legislativo4marzo2014n.26,siprevedecheentroil2020loxenotrapiantodipolmonisuinisuuomosarànellafasedeiclinical trials grazie agli enormi sforzi economici per lo sviluppo di questatecnologiaeffettuatisoprattuttonegliUSA(ReardonS.,2015).Laconoscenzadeimeccanismicon iquali il sistema immunitariosimuovepereliminaregliagentiinfettivi,oltreallaconoscenzadeimicrorganismistessi,èdifondamentale importanza per lo sviluppo di nuovi metodi per combattere losviluppo e la diffusione di nuove e vecchie malattie. In particolare, alleconoscenzeincampoimmunologicoèstrettamentelegatolosviluppodivaccinisemprepiùefficaciesicuri(MurtaughM.P.,2014).In questo lavoro si è voluto descrivere le più recenti conoscenze sul sistemaimmunitariodelsuinoconparticolareriferimentoaitoolllikereceptor(TLR)ealla risposta all'infezione da Mycoplasma hyopneumoniae. Nella partesperimentaleèstatainvestigatalarispostaimmunitariaaseguitod'infezionedaMycoplasmahyopneumoniaecontecnicheimmunoistologichenelsuino.

6
TOLLLIKERECEPTORS
GeneralitàLa difesa dell'ospite dall'invasione di agenti microbici è affidata al sistemaimmunitario che consiste di due componenti: l'immunità innata e l'immunitàacquisita.Entrambe le componenti riconoscono i microrganismi invasori come non-selfinnescando le risposta immunitaria al fine di eliminarli. Ad oggi, la maggiorpartedeglistudihaesaminato inmodo indipendenteentrambe lecomponenti,ma l'interesse principale di ricerca è stato puntato sull'immunità acquisita.Nell'immunità acquisita, i linfocitiB eTutilizzano recettori antigenici come leimmunoglobulineoirecettoridellecelluleTperidentificareelementinon-self.Imeccanismiconiqualiquestirecettoriindividuanoantigeniestraneisonostatiintensamenteanalizzatieimaggiorimeccanismicomelaclonalità,ladiversitàela memoria sono stati caratterizzati a fondo. Tuttavia, questi recettori sonopresenti solo nei vertebrati, di conseguenza non si comprende pienamente ilmeccanismo di riconoscimento dei non-self negli organismi meno evoluti(Takeda e Akira, 2005). Una parziale risposta a questo quesito è stata dataquando,alla finedeglianni '90, fu individuato ilrecettoreTollcomeessenzialeper la difesa contro i funghi nel moscerino Drosophila. Un anno dopo, neimammiferi un omologo del recettore Toll (denominato poi TLR4) è statoindividuato come induttore dell'espressione di geni coinvolti nella rispostainfiammatoria(Medzhitov,1997).I sistemi difensivi innati sono diversi. La prima barriera contro l'invasione diorganismi esterni è rappresentata dalla cute e dalle mucose. Un'importanteazionedi difesa èpoi svoltadalle cellule aventi la funzione fagocitaria, come igranulocitineutrofili, imonociticircolantie lecellulemacrofagichedelsistemareticolo-endoteliale. I neutrofili hanno un’azione rapida ed efficiente che siesaurisce con ladistruzionedelmateriale estraneo, senza chequest'ultimosiacoinvoltonella risposta immunitaria; al contrario imacrofagi simobilizzanoe,nonsolodistruggonoimicrorganismifagocitati,maliprocessanoeripresentanoi loroantigeniallealtrecelluledelsistemaimmunitario, innescandocosìtutti imeccanismidelledifesespecifiche.

7
PrimaidentificazioneNel 1985, Christiane Nüsslein-Volhard del Max Planck Institute a Tübingen(Germania)analizzavalemutazionideimoscerinidellafruttaenotòunastranalarva nella quale la porzione ventrale del corpo era sottosviluppata. Il suocommento spontaneo fu "Das war ja toll!" che significa "com'è notevole!"; ilnomeTollfucosìattribuitoalgenemutato.AllaproteinaprodottadalgeneTollfu attribuita la proprietà di causare ventralizzazione e la normale funzionalitàdelTollènecessariaperlapolaritàdorsoventralenellamosca.DieciannidopolascopertadelToll,LemaitreB.ecoll.scoprironocheilTollnonsolocontrolla lapolaritàdorsoventralemahaancheunruolonelladifesadell'organismocontrole infezioni fungine in Drosophila: senza il recettore Toll le mosche nonsopravvivonoalle infezioni fungine (LemaitreB.etal., 1996). L'attivazionedelTollinnescalacascataNF-κBcheattivaladifesacontroimiceti.Medzhitov R. e Janeway C. all'Università di Yale nel 1997 annunciarono laclonazionediunomologoneimammiferialrecettoreTolldenominatoToll-likereceptor (TLR) (Medzhitov R. et al., 1997). Dall'identificazione del primorecettoreTLR,ilTLR4,diverseproteinesonostatestrutturalmentecorrelatealTLR4equindidenominateTLR.Attualmente,irecettoriToll-likecomprendonouna vasta famiglia di proteine che conta 12membri (KumarH. etal., 2009). ITLR riconoscono diverse molecole associate a patogeni (PAMP) derivate davirus,batteri,funghieprotozoi.

8
StrutturaelocalizzazionedeiTLRITLRsonoglicoproteinetransmembranaditipo1(Fig.1).IldominioextracellulareN-terminaleècostituito da circa 16-28 ripetizioni ricchi dileucina (LRR),eogniLRRsi componedi20-30aminoacidiconilmotivoconservato"LxxLxxL".Il dominio intracellulare C-terminale èconosciuto come Toll/IL-1 (TIR), che mostraomologia con quello del recettore IL-1. Questodominio è necessario per l'interazione e ilreclutamento di varie molecole per attivare ilpercorsodisegnalazione.RecentementeèstatoindividuatalastrutturacristallinadidiversiTLRcon il ligando. è stato dimostrato che questicomplessi formano eterodimeri quali TLR1-TLR2,TLR4-MD2ounomodimerocomeTLR3-TLR3 dopo associazione con i loro rispettivi
ligandieformanounastrutturaaferrodicavallo.Èstatodimostratochequestastrutturaèessenzialeperillegameconilligandoeperilpassaggiodelsegnaleavalle.I TLR sono espressi in comparti cellulari diversi. Il TLR1, TLR2, TLR4, TLR5,TLR6eTLR11 (espresso solonei topi) sonoespressi sulla superficie cellulare,mentreTLR3,TLR7,TLR8eTLR9sonoespressiinvescicoleintracellularicomeendosomi e reticolo endoplasmatico. I TLR intracellulari sono trasportati allevescicoleintracellularimedianteUNC93B1,unaproteinatransmembrana,chesilocalizzanelreticoloendoplasmaticodellacellula(KimY.M.etal.,2008).Parecchi studi hanno rilevato la presenza dei diversi TLR inmolti tessuti. Nelcane, i TLR2 sono stati individuati inmonociti del sangue, linfonodi, polmoni,fegato, milza, pancreas, piccolo intestino, intestino crasso e pelle tramite realtime-PCR (RT-PCR) (Ishiietal., 2006). Le differenzedi localizzazione tissutaledeiTLR4trasuino,bovinoecaneriscontratedaWassefecoll.(2004)neidiversiorganisuggerisconounadistribuzionedifferentenellediversespecie;differenzedidistribuzionedeiTLR3sonostateevidenziateanchetramacrofagialveolarididifferenti specie (Sacco et al., 2011). La distribuzione tissutale dei TLR,soprattuttodiquellicoinvoltinelriconoscimentodeibatteri,ècoerenteconunafunzione di vigilanza dei siti d'ingresso nell'organismo, permettendo ilriconoscimentoprecoced'invasionemicrobica(Alvarezetal.,2008).
Figura1.Schemaproteinetransmembranaditipo1

9
ElementiriconosciutidaiTLRI TLR possono riconoscere diversi componenti della parete della cellulabattericacomeilipolisaccaridideibatterigramnegativi,riconosciutidaiTLR4,ipeptidoglicani da batteri gram positivi, riconosciuti da TLR2, illipoarabinomannan (LAM) dei micobatteri, riconosciuto dai TLR2, i diacil etriacillipopeptidideibatteri,micobatteriemicoplasmi,riconosciutidaiTLR2/1eTLR2/6, laflagellinadibatterifalgellati,riconosciutadaTLR5(Hayashietal.,2001;Uematsuetal.,2006)eilDNAgenomicobattericoriccodiDNACpGnonmetilato,riconosciutodaiTLR9.Diversamente dagli altri TLR, nel topo, i TLR5 non sono espressi nelle celluledendriticheconvenzionalieneimacrofagi(Uematsuetal.,2006).Gewirtzecoll.riportanoche iTLR5sonoespressinella facciabasolaterale,manonnell'apicedelle cellule epiteliali intestinali, suggerendo che la flagellina è riconosciutaquando i batteri passano attraverso l'epitelio (Gewirtz et al., 2001).L'espressione di TLR5 nelle cellule epiteliali intestinali del topo non è alto(Uematsu et al., 2006). D'altra parte, il TLR5 è espresso principalmente sullecelluledellalaminapropriaintestinaleCD11c+(Uematsuetal.,2006).SebbeneilTLR5 induce la risposta immunitaria contro i batteri flagellati, i topi TLR5-deficenti sono resistenti all'infezione orale da Salmonella typhimurium. Iltrasporto di S. typhimurium dal tratto intestinale ai linfonodi mesenterici èdebole in topiTLR5-deficenti.Questi risultati suggerisconocheS.typhimuriumutilizziiTLR5deiCD11c+dellalaminapropriaperattuarel'infezionesistemica(Uematsuetal.,2006).αandεProteobacteria,includingHelicobacterpyloriandCampylobacter jejuni, variano il sito di riconoscimento della flagellina senzaperdere la motilità flagellare. Questa modificazione può essere responsabiledellapersistenzadell'infezione sulla superficiedellamucosa (Andersen-Nissenetal.,2005).Con l'interazione travaricomponentibatteriche iTLR inducono laproduzionedicitochineinfiammatoriee,inspecifichecondizioni,diinterferoniditipoI.Glistreptococchi del gruppo B, che residuano nel fagosoma, inducono uninterferoneTLR7-dipendenteditipoI,suggerendochel'RNAbattericoprodottonelcompartimentolisosomialepossaesserericonosciutodaiTLR7(MancusoG.etal.,2009).AnchespirochetediBorreliaburgdorferi,checausanolamalattiadiLyme nell'uomo, sono riconosciute dai TLR1 e dai TLR2 (Dennis V.A. et al.,2009).Il DNA batterico è un potente stimolatore dell'immunità. Questa risposta èmediatadaisitiCpGnonmetilati.Neivertebrati,lafrequenzamoltoridottadei

10
sitiCpGelametilazionedeiresiduidicitosinadeisitiportanoadunainibizionedell'attività immunostimolatoria. Studi su topi TLR9-deficenti hannomostratocheilriconoscimentodeisitiCpGnonmetilatièmediatodaiTLR9(Hemmietal.,2000).Leproteinedell'envelopediviruscomeilvirusrespiratoriosinciziale(RSV)eilvirusdeltumoremammariomurino(MMTV)sonoriconosciutidaiTLR4.L'RSVinduce la produzione di citochine infiammatorie e chemiochine attraverso iTLR2eiTLR9(MurawskiM.R.etal.,2009).ITLR9sonoattivatidiversivirusaDNAdelle famiglie herpesvirus (Lund J. etal., 2003), adenovirus (Zhu J.etal.,2007),poxvirus(SamuelssonC.etal.,2008)e torquetenovirus(Rocchi J.etal.,2009).ITLR2sisonorilevaticoinvoltinelriconoscimentodelvirusdelmorbillo,delcitomegalovirusumanoedell'Herpessimplexvirusdi tipo1(BiebackK.etal., 2002; Compton T. et al., 2003; Kurt-Jones E.A. et al., 2004). Un analogosinteticodell'RNAadoppiacatena,l'acidopolinosina-deossicitidilico,unpotenteinduttore dell'interferoni di tipo I (Alexopoulou L. et al., 2001). I topi TLR3-deficentisonosuscettibiliall'infezionedacitomegalovirusdeltopo(TabetaK.etal., 2004). D'altra parte, topi TLR3-deficenti mostrano maggior resistenzaall'infezionedavirusdellaWestNileDisease.QuestoflavirusasingolacatenadiRNA innesca la risposta infiammatoria viaTLR3, cheporta adunadistruzionedellabarrieraemato-encefalica,seguitadaunintensa infezione(WangT.etal.,2004). Ciò suggerisce che il virus West Nile utilizzi i TLR3 per penetrareefficientemente nell'encefalo. TLR7 e il TLR8 umano sono in grado diriconoscere ssRNA ricco di guanosina o uridina proveniente da virus come ilvirusHIV,ilvirusdellastomatitevescicolareeilvirusdell'influenza(DieboldS.S.etal.,2004;HeilF.etal.,2004).Attualmente,iTLR10nonsonostaticollegatiadunospecificolegando,inoltrelaloro attività sembra essere inibitoria in particolare sulla produzione di IL-1β,TNF-α,eIL-6(OostingM.etal.,2014).

11
TLR Localizzazione Molecoleassociateapatogeni Citochinaindotta Localizzazionetissutale*TLR1 Membrana
cellulare(superficiecellulare)
Triacil lipopeptidi (batteri e micobatteri)
Citochineinfiammatorie(TNF-α,IL6,etc.)
UbiquitariMonociti/macrofagi,Celluledendritichemieloidi,Mastociti,LinfocitiB
TLR2 Membranacellulare(superficiecellulare)
Peptidoglicani(batteri Gram-positivi) Lipoarabinomannano (micobatteri) Emoagglutinina (virus del morbillo) Fosfolipomannano(Candidaalbicans)Glicosil-fosfofatil-inositolo(Trypanosomacruzi,Toxoplasmagondii)
Citochineinfiammatorie(TNF-α,IL6,etc.)
Monociti/macrofagi,Celluledendritichemieloidi,Granulociti,Mastociti,Celluleepitelialiepatiche,bronchiali,tubulari,Cervello,Cuore,Polmone,Milza
TLR3 Endosoma ssRNAvirus(WNV),dsRNSvirus(Reovirus),virusrespiratoriosinciziale,citomegalovirusmurino
Citochineinfiammatorie(TNF-α,IL6,etc.),IFNtipoI
Celluledendritiche,LinfocitiB,Placenta,Pancreas
TLR4 Membranacellulare(superficiecellulare)
LPS(batteri Gram-negativi), Mannano(SaccharomycescerevisiaeCandidaalbicans),Glicoinositolofosfolipidi(Trypanosomacruzi),proteinedell'envelope(RSVeMMTV)
Citochineinfiammatorie(TNF-α,IL6,etc.),IFNtipoI
Monociti/macrofagi,Celluledendritichemieloidi,Mastociti,Epiteliointestinale,Milza,LinfocitiT,Granulociti
TLR5 Membranacellulare(superficiecellulare)
Flagellina(batteriflagellati) Citochineinfiammatorie(TNF-α,IL6,etc.)
Monociti/Macrofagi,Celluledendritiche,Epiteliointestinale,Ovaia,Prostata,Leucocitiperiferici
TLR6 Membranacellulare(superficiecellulare)
Diacil lipopeptidi (micoplasmi), Zimosano (Saccharomycescerevisiae), Acido lipoteicoico (batteri Gram-positivi)
Citochineinfiammatorie(TNF-α,IL6,etc.)
Monociti/macrofagi,Mastociti,LinfocitiB,Timo,Milza,Polmoni
TRL7 Endosoma ssRNAvirus(VSV,influenzavirus)
Citochineinfiammatorie(TNF-α,IL6,etc.),IFNtipoI
Monociti/Macrofagi,Celluledendritiche,LinfocitiB,Polmoni,Placenta,Tonsille,Linfonodi,Milza
TRL8 Endosoma ssRNAdaRNAvirus Citochineinfiammatorie(TNF-α,IL6,etc.),IFNtipoI
Monociti/Macrofagi,Celluledendritiche,Mastociti,Polmoni,Placenta,Milza,Linfonodi,Midolloosseo
TRL9 Endosoma dsDNAvirus(HSV,MCMV),sitiCpGdibatterievirus,Emozoina(Plasmodiummalariae)
Citochineinfiammatorie(TNF-α,IL6,etc.),IFNtipoI
Monociti/Macrofagi,Celluledendritiche,LinfocitiB,Milza,Linfonodi,Midolloosseo
TRL10 Membranacellulare(superficiecellulare)
nonidentificati inibizionediIL-1β,TNF-α,eIL-6
Monociti/Macrofagi,LinfocitiB,Milza,Linfonodi,Tonsille,Timo
TLR11 Membranacellulare(superficiecellulare)
Molecolesimilprofillina(Toxoplasmagondii),
Citochineinfiammatorie(TNF-α,IL6,etc.)
Monociti/MacrofagiEpatociti,Rene,Epiteliovescicale,Milza,Piccolointestino
Tabella1:ModificatodaKumarecoll.2009edaBrozeMonack2013;*tessutiincuièstatorilevatonellediversespecieanimali,inclusol'uomo.

12
TLR4I TLR4 riconoscono specificatamente i lipopolisaccaridi batterici e la loroattivazioneportaprincipalmenteadunasintesidicitochinepro-infiammatorieechemiochine(Janssensetal.,2003;Kanzleretal.,2007).Recentemente,l'usodiagonistideiTLRperstimolarepreferenzialmentelarispostaimmunitariadeiThelper 1 o dei T helper 2 si è rivelato realisticamente fattibile (Alving et al.,2012). Per esempio ilmonofosforilA, un derivato dei LPS, agonista dei TL4, èstato approvato come adiuvante per i vaccini umani contro l'epatite B e ilpapilloma virus (Ireton et al., 2013; Johnson D.A., 2013). Altri TLR4-agonistisono attualmente coinvolti in studi clinici. Per esempio l'adiuvante lipidicoglucopiranosil, è attualmente in prova come adiuvante in un possibile vaccinocontrolatubercolosi(MeyeretMcShane,2013).Questinuoviesicuriadiuvantipossono facilitare lo sviluppo di vaccini contro bersagli scarsamenteimmunogeni. Gli studi preclinici sono solitamente effettuati inmodelli animalimurini assunti come rappresentativi del sistema immunitario dei mammiferi.Sebbenemolti aspetti del sistema immunitario sia innato che adattativo sianocomuni tra uomo e topo, sono state rilevate numerose differenze tra le duespecienellarisposteall'attivazionedeiTLR(Schroderetal.,2012).Nonostantele differenze delle sequenze che codificano per i TLR tra le diverse specie, lefunzioni biologiche di base e le vie di trasmissione del segnale dei TLR sonosovrapponibilitradiversespecie(Ketloyetal.,2008;RehliM.,2002).Nel 1999, Qureshi e coll. identificarono il gene del TLR4 nella regionecromosomiale LPS come responsabile della mancata risposta ai LPS in alcuniceppidi topi. Furono identificate anche alcunemutazioninei genidelTLR4diceppi di topi iporeattivi ai LPS, dimostrando che il TLR4 è essenziale permediare la risposta ai LPS in vivo (Poltorak etal., 1998;Qureshietal., 1999).OltrealLPS,uncomponentedellamembranaesternadibatteriGramnegativi,iTLR4didiversespeciericonosconomoltialtricomponentideipatogenicomeipolimeri di acido mannuronico di batteri Gram negativi, l'acido teicoico deibatteri Gram positivi e componenti virali come la proteina F del virusrespiratoriosinciziale(Yangaetal.,2001;Haynesetal.,2001;Kurt-Jonesetal.,2000). Oltre a componenti esogene, i TLR4 si legano con molecole endogenecomeproteineliberatedaitessutiincorsodishockdacaloreneltopo,nelrattoenell'uomo(Ohashietal.,2000;Vabulasetal.,2001;Kimetal.,2009;Roelofsetal., 2006). L'extra dominio A della fibronectina e gli acidi grassi saturi sonopotenzialmente riconosciuti daTLR4umani emurini, il gruppo emedai TLR4murini (Gondokaryono et al., 2007; Okamura et al., 2001; Kim et al., 2007;

13
Tsukumoetal.,2007;Figueiredoetal.,2007).Iperossidid'idrogenodell'esteredel colesterile, componente attiva delle lipoproteine a bassa densitàminimamente modificate, sono state recentemente individuati come TLR4agonisti (Choi et al., 2009). Questi studi dimostrano che molecole prodotte ocircolantidurantesituazionianormali, comeundannotessutale,sono ingradodiinnescarel'attivazionedeiTLR4(MedzhitovetJaneway,1997).Strutturalmente, i TLR4 formano un complesso sulla superficie cellulare condiverseproteinenecessariaper il riconoscimentodel ligando(Fitzgeraldetal.,2004;Miller et al., 2005). Nel siero, inizialmente l'LPS è legato dalla proteinaLBPchetrasferiscel'LPSalCD14.IlCD14èunaproteinadimembranalegataalglicosilfosfatidilinositolo che circola anche in forma solubile e che lega icomplessi LPS-LPB con alta affinità. Mentre il CD14, da solo, manca di undominio intracellulare per innescare il segnale, associato al TLR4 forma uncomplessofunzionaleLPS-recettore.IllegamediLPSrichiedeanchelaproteinaMD-2 che è associata al dominio extracellulare del TLR4 (Park et al., 2009).Quindi il complesso attivo recettore-LPS richiede il CD14, il TLR4 e l'MD-2,anche se uno studio suggerisce che il CD14 e la LBP migliorino soltanto larisposta al LPS dipendente dal TLR4 e non siano necessari per il legame e larispostaalLPS(Lizundiaetal.,2008).

14
TLRnelsuinoI TLR nel suino sono stati studiati per la loro importanza nella difesa dalleinfezioni eper verificarne il ruolo. LadistribuzionedeiTLR9neipolmoni e indiversitessutilinfoidièstataindagataindiversistudi(Kuzemtsevaetal.,2015;Shimosato et al., 2005) che ne hanno riscontrato, con tecnicheimmunoistochimicheeconPCR,lapresenzanellecelluleepiteliali,nell'endoteliovascolare,nellecelluledeisettialveolari,indiversecellulepresentantil'antigenedellezoneperifollicolaridellaplacchedelPeyer,neilinfonodi,nellamilzaeneltimo.Nel suino, a differenza del bovino (Werling etal., 2006), imacrofagi alveolariesprimono iTLR3 (Chaungetal., 2010; Sangetal., 2008a; Liuetal., 2009). Insuini infettati con PRRSV e PCV2 sono stati evidenziati due distinticomportamenti nell'espressione dei TLR: un aumento dei TLR2, 4 e 8 e unadiminuzionedeiTLR3,7e9(Tuetal.,2015).ITLR2e iTLR4sonotraipiùstudiatiper il lororuolonelladifesada infezionibatteriche.Indiversicasi,nelsuino,èstatorilevatounaumentodell'espressionediquestiduerecettori:dientrambi,incasodistressdacalore(Juetal.,2014)e,dei TLR2, in caso d’infezionemammaria da E. coli (Zhu et al., 2008). In suiniselezionatiperlaresistenzaaMycoplasmapneumonial'espressionedeiTLR2e4nei tessuti linfoidi si è rilevata più alta rispetto al gruppo di controllo,suggerendo una particolare importanza di questi recettori associata ad altriparametri immunitari nella difesa da infezioni polmonari da micoplasmi(Borjiginetal.,2016).L'utilizzo di anticorpi monoclonali anti-TLR2 ha permesso di evidenziare lapresenza di questi recettori in monociti, macrofagi, granulociti, ma non inlinfociti del sangue periferico. La colorazione di sezioni tissutali ha rilevatol'espressione del TLR2 in cellule epiteliali, dei tratti tracheobronchiale eintestinale,dottibiliari,tubulirenalielaminabasaledell'epidermide(Alvarezetal.,2008).Nelsuino,imonocitielecelluledendritichediderivazionemonocitaria(mo-DC)invitromatureed immatureesprimonotrascrittideiTLR4,mentrequestinonsonostatiriscontratiinmo-DCmatureumane(Alvarezetal.,2006).Nellecellulemononucleari suini, i LPS inducono la trascrizione di TLR4, CD14 e RNAmessaggerodeiMD-2edidiversecitochinecomeIl-1,Il-10,IL12,TNF-αeTNF-γ(Raymondetal., 2005; Sorensenetal., 2011). Sebbene l'espressione dei TLR4

15
aumenti neimonociti umani in seguito all'esposizione al LPS, lo stesso effettononsihaneimonocitisuini(Raymondetal.,2005).IlivellipiùaltidimRNAdiTLR4suinosonostatiriscontraticonl'usodellaPCRnel colon e nella milza. L'mRNA dei TLR4 è stato trovato anche nei polmoni,piccolo intestino, fegato, reni, timo, linfonodi, cervello (ipotalamo, ippocampo,cortecciaecervelletto),tonsille,ovaieecornea,manonnellapelleenelmuscolo(Thomas et al., 2006; Alvarez et al., 2006; Burkey et al., 2007; Wassef et al.,2004). Studi immunoistochimici hanno rivelato che, nei polmoni, il TLR4 èespresso dai macrofagi e dall'epitelio, nel fegato dalle cellule del Kupffer edall'epitelio dei dotto biliari. Comunque né gli epatociti né l'epitelio vascolareepatico lo esprimono. Il TLR4 è espresso anche nelle cellule M e nel tessutolinfoide associato al sistema gastrointestinale. In suini infettati con PRRSV,l'espressionedelTLR4èaumentataneilinfonoditracheobronchiali,nelcervello,nell'ipotalamo, nell'ippocampo, nella corteccia e nel cervelletto, ma non neipolmoni (Migueletal., 2010). Ceppimolto virulenti del virus della PRRS sonoforti induttorideiTLR3,7e8 inmacrofagialveolarienellazonamidollaredelcervello rispetto a ceppi a bassa patogenicità dello stesso virus (Zhang et al.,2013).

16
RISPOSTAIMMUNITARIAALL'INFEZIONEDAMYCOPLASMAHYOPNEUMONIAELa polmonite enzootica suina (PEN), causata principalmente da Mycoplasmahyopneumoniae,èunamalattiacontagiosariscontratainmoltiallevamentisuini(Marèetal.,1966).Lamalattiaècaratterizzatadaaltamorbilitàe,sebbenesiaassociataconunabassamortalità,determinaunadiminuzionedell'incrementomedioponderaledeisuini iningrassoedell'indicediconversione(Morrisonetal.,2000),causandograviperditeeconomiche(Sarradelletal.,2003;Choietal.,2006;Lorenzoetal., 2006;Thomsenetal., 1992; Straw,1989;Noerregaardetal., 2016). Il danno causato da M. hyopneumoniae all'epitelio respiratoriopredispone gli animali ad infezioni secondarie di organismi come Pasteurellamultocida (Amass et al., 1994), Haemophilus parasuis (Williams et al., 2000),virusinfluenzali(Thackeretal.,2001),Actinobacilluspleuropneumoniae(Vigreetal.,2004),virusdellasindromeriproduttivaerespiratoriadelsuino(PRRSV)(Thackeretal.,2000;Thanawongnuwechetal.,2001,2004;ThanawongnuwechandThacker,2003),ecircovirussuinotipoII(PCV2)(Opriessnigetal.,2004).Leinfezioni secondarie sono la principale causa dei danni economiciall'allevamento,poichél'infezionesostenutadaM.hyopneumoniaenoninfluiscesulleperformancedicrescitadelsuino(Escobaretal.,2002).La trasmissione dell'infezione avviene per via inalatoria e successivalocalizzazione nelle prime vie respiratorie. La fase primaria dell’infezione ècaratterizzata da arresto dell’attività ciliare nelle aree colonizzate, adesioneall’apice delle cellule ciliate e loro progressiva distruzione. La colonizzazionedellamucosarespiratoriaavvienegraziealleadesinebatteriche.M.hyopneumoniaehadimensionicellularidi400-120nmconformapleomorficadovuta alla mancanza di parete cellulare. In laboratorio è difficilmentecoltivabileel'isolamentospessorichiede4-8settimane.M. hyopneumoniae induce una risposta immunitaria lenta e inefficaceevidenziatadallalentasieroconversioneedaibassilivellidianticorpineltrattorespiratorio(DjordjevicS.P.etal.,1997;ThackerE.L.etal.,2000;ThackerE.L.etal.,1998).UnacomponentefondamentaledellapatogenesidiM.hyopneumoniaeriguarda i cambiamenti immunopatologici che comprendono una diminuzionedell'attivitàfagocitariadeimacrofagialveolari(CarusoJ.P.eRossR.F.,1990),ladiminuzione della risposta dei linfociti amitogeni non specifici (KishimaM. eRossR.F., 1985) e la produzionedi citochine infiammatorie come IL-1, IL-6 eTNF-α(ThackerE.L.etal.,2000;AsaiT.M.etal.,1993;AsaiT.M.etal.,1994).

17
Inoltre,nelliquidodilavaggiobronchialesirilevanolivelliaumentatidiIL-8,IL-10 e IL-12 (Thacker E.L. et al., 1999). La produzione di TNF-α da parte deimacrofagialveolarièdipendentedallastimolazionedeiTLR2e6(Munetaetal.,2003).Inunostudiodel2011èstatavalutatal'espressionedeiTRL6duediversigruppigeneticidisuiniinseguitoallavaccinazionecontroM.hyopneumoniae.Irisultatihanno mostrato una diminuzione dell'espressione dei TLR6 in seguito allavaccinazione e una significativa differenza di espressione tra i due gruppigenetici (Régia Silva Sousa K. et al., 2011). Inoltre, in un'altra ricerca, è stataevidenziataunadiminuzionedell'infiltrazionedeimacrofagineltessutolinfoideassociatoaibronchiinseguitoavaccinazione(Vranckxetal.,2012).La diagnosi d'infezione può esser fatta conmetodi diretti o indiretti. Imetodidiretti comprendono l'isolamento del battere, l'evidenziazione dell'agente supreparati istologici o freschi mediante tecniche d'immunofluorescenza,l'esecuzionediunaPCRorealtime-PCR(RT-PCR)sutessutiocampionibiologiciquali liquido da lavaggio bronchio-alveolare. Tra i metodi indiretti il piùutilizzatoèiltestELISA.Lelesionirilevabilimacroscopicamentesonocaratterizzatedaepatizzazionedelparenchima polmonare più frequentemente nelle aree cranioventrali delpolmoni (figura 2). Istologicamente è rilevabile un'iperplasia dei complessiBALT (Bronchial associated lynphoid tissue) e notevole essudatointrabronchiolare(vedifigura3).
Figura2:polmoneconlesionimacroscopichedeilobiapicalicaratterizzatedaareedi
consolidamentodelparenchimariconducibiliaM.hyopneumoniae.

18
Figura3:Iperplasialinfocitariaperibronchiolareeperivascolare,macrofagie
neutrofilinellumedellevieaeree(freccia).Ematossilina-eosina,barra:95µm.DaJ.Sarradelletal.,2003.
Diversi studi hanno valutato la risposta immunitaria in caso d'infezione daM.hyopneumoniaenelsuinodelineandoivariaspettichalacompongono.Sarradelle coll. nel 2003 hanno descritto chiaramente le lesioni ascrivibili a M.hyopneumoniae riscontrabili istologicamente. I polmoni infetti da M.hyopneumoniaemostravano iperplasia linfoidedel tessuto linfoideassociatoalbronco (BALT) che provoca obliterazione del lume bronchiale ed atelettasiadeglialcolicircostanti(figura3).MHcolonizzalasuperficieluminaledeibronchiebronchiolisenzapenetrarenelparenchimapolmonare,causandolariduzionedell’attività ciliare, la perdita delle cilia, la formazione di microcolonie el’accumulo di batteri, portando anche all’esfoliazione dell’epitelio. L'epiteliosopra i noduli prominenti si presenta degenerato ed esfoliato. Nellacomparazioneconanimalicontrollo,sievidenziavaladiffusaperditadicigliaindifferenti superfici bronchiali. Nei bronchi e nei bronchioli si evidenziavaun'iperplasia delle cellule caliciformi. I setti alveolari sono ispessitidall'accumulodipiccoliegrandilinfociti,pasmacelluleeneutrofili.NeglialveolisievidenziavaunamoderataiperplasiadeipneumocitiditipoII(Sarradelletal.,2003). Nello stesso studio è stata rilevata, mediante l'utilizzo di anticorpimonoclonali per CD3, un'intensa immunoreattività sulla superficie e nelcitoplasmadi cellulemononucleari. I linfociti CD3+ sono stati riscontrati nelleareeperifollicolarieinterfollicolaricosìcomeall'internod'infiltrazionidicellulemieloidi nelle pareti alveolari (vedi figura 4). Numerosi cellule positive siosservavano nella lamina propria dei bronchi e dei bronchiolo e tra l'epitelio

19
delleviearee(Sarradelletal.,2003).Utilizzandoanticorpianti-SLA-IIeS-100sirilevavano, nei centri germinali dei BALT, cellule stellate con lunghi processicitoplasmatici. Alla periferia dei BALT, nella lamina propria dei bronchi e deibronchioli, nei setti alveolari e attorno alle ghiandole bronchiali submucosalierano presenti plasmacellule contenenti IgG o IgA. Le cellule CD4+predominavanosulleCD8+.Lo stesso quadro istologico è stato riscontrato da Redondo e coll. (2009) chehannoseguitol'andamentodiuninfezionesperimentaleconM.hyopneumoniae.Lelesioniistopatologichesonostaterilevantitrai7ei28giornipostinfezionecoincidendo con un aumento dell'espressione, rilevata con tecnicheimmunoistochimiche,dellecitochine(IL-1a,IL-1b,IL-8,eINF-g)edeimarcatorilinfaticiqualiCD4+,CD8+,muramidasi,IgG+eIgA+(Redondoetal.,2009).Contecniche di RT-PCR è stato rilevato un aumento dell'espressione di IL-1, TNF-alpha e IL-6 in suini infettati sperimentalmente conM. hyopneumoniae doposette giorni dall'infezione (Choi et al., 2006). Lo stato infiammatorio nei sitid’infezione è dimostrato anche dall'aumento di espressione di ciclossigenasi-2nelle cellule epiteliali bronchiali e bronchiolari, nelle ghiandole dellasottomucosaeneimacrofagipresentinell'essudatobroncoalveolare(Andradaetal.,2014).
Figura4:linfocitiCD3+immunocoloratilocalizzatiprincipalmentenelleareeperifollicolarideiBALTenellalaminapropriadeibronchioli(freccia).MetodoABC,ematossilinadiHarris,barra=65µm.DaJ.Sarradelletal.,2003.

20
PARTESPERIMENTALE
IntroduzioneCOMPONENTIIMMUNITÀCELLULO-MEDIATAL'immunità cellulo-mediata è gestita dai linfociti T. Queste cellule nonproducono anticorpi, ma agiscono distruggendo direttamente il "bersaglio" ofacendolo distruggere da cellule accessorie che vengono stimolate dallelinfochine,sostanzesolubilidaesseprodotte.IlinfocitiT,quandoriconosconoesi legano aun antigenemediante il loro recettore specifico, vengonoattivati eproliferano formando un clone di cellule immunocompetenti, nella regionedell'organismo dove è concentrato l'antigene, oltre che nei linfonodi dovel'antigene è trasportato dai macrofagi che lo hanno fagocitato, processato epresentato in superficie. Tra queste cellule che proliferano rapidamente sidifferenziano, su base funzionale, tre distinte sottopopolazioni: a) celluleregolatrici,b)celluledellamemoria,c)celluleeffettrici.Le cellule regolatricimodulano l'intensità della risposta immunitaria e inoltreintervengono nella cooperazione tra immunità cellulo mediata e immunitàumorale.IlinfocitiThelpersvolgonoilruolocentraleinquestaregolazione.Piùprecisamente, la sottoclasse di linfociti T helper, denominata Th1, controllal'attivazionedeimacrofagi,aumentandonel'efficienzanellaneutralizzazionedeimicrorganismi fagocitati; la sottoclasse helper, denominata Th2, interagisceinvece con i linfociti B, al fine di avviare o di aumentare la produzioneanticorpale.Questotipodiinterazionerappresentaunsistemadisicurezza,perevitare che accidentalmente un linfocitaB inizi a produrre anticorpi inmodoincontrollato.UnasecondapopolazionedicelluleregolatricièquelladeilinfocitiTsuppressor,cheagisconolimitandolarispostaanticorpaleequestodaunlatoconsentediimpedirecheunarispostaimmunitariaeccessivapossadanneggiarel'ospitestessoe,dall'altro,evitaun’inutileproduzionedianticorpiquandononsonopiùnecessari.LecelluledellamemoriarappresentanounulteriorepooldilinfocitiTche,dopostimolazionedapartedell'antigene,permangonoperlunghiperiodi,anchedopolascomparsadiquest'ultimo.Questecellule"sensibilizzate"rimangonoquiescentifinchénonincontranosuccessivamentelostessoantigene:inquestocasosi riattivanomoltorapidamentedando luogoallaproliferazionedi un pool di linfociti effettori e regolatori. Questo tipo d’immunità cellulomediata "della memoria" è simile alla risposta umorale anamnestica e ha lafunzionediridurreilperiododilatenzadacirca20giorniasolo48-72ore.Le cellule effettrici o linfociti T citotossici attaccano fisicamente le cellule

21
bersaglio e ledistruggono inducendoalterazionidimembrana cui consegue lalisi. L'immunità cellulo mediata è d’importanza fondamentale nelle difeseimmunitariecontromicrorganismiaparassitismo intracellularequaliprotozoi,funghi, virus e batteri soprattutto dei generi Listeria, Brucella e micobatteritubercolari.Ineffetti,percombattereimicrorganismichesireplicanoall'internodelle cellule dell'ospite, gli anticorpi che non sono in grado di penetrare nellecellulesonoinefficienti.IlinfocitiTcitotossicisonoparticolarmenteefficientinelriconoscere e distruggere le cellule infette o comunque alterate. L'immunitàcellulomediataèinoltred’importanzafondamentaleneifenomenidirigettodeitrapiantienelsistemacheindividuaedistruggelecelluleneoplastiche,tramitelecelluledenominatenaturalkiller.Perrealizzarequestimeccanismieffettori,ilinfociti T, a differenza dei linfociti B, sono capaci di riconoscere e reagire,attraversoillororecettorespecificodimembrana(Tcr)soloconbrevisequenzedi peptidiche, appartenenti al microrganismo invasore, presenti o sullasuperficie di cellule specializzate (ad es. i macrofagi), che fungono da antigenpresentingcells, oppure presenti sulla superficie di cellule infette o comunquealterate che devono quindi venire distrutte. Più precisamente gli antigeniriconosciuti dai linfociti T sono parti di un agente patogeno che vengonopresentatesullasuperficiedellecelluledopoesserestateelaborateemontateinuncomplessoconproteinecellularidellacellulastessa,chesonocodificatedaigenidelcomplessomaggiorediistocompatibilità(MHC).LINFOCITITI linfociti T sono coinvolti nel processo di coordinazione della rispostaimmunitaria cellulo-mediata. I linfociti T sono distinti in due sottopopolazioniprincipali,caratterizzatedaldiversomeccanismod'azioneedalmeccanismodifunzionamento: linfocitiThelper e linfocitiTcitotossici.AltresottopopolazionidiminorimportanzasonoilinfocitiTmemoria,linfocitiTregolatori, linfocitiTnatural killer e linfociti T γ/δ. I linfociti T originano nel midollo osseo ematurano nel timo. Durante la maturazione attraversano vari stadi che sipossonodistinguere inbaseai recettori espressi in superficie. I linfocitiTnonpossiedononéimmunoglobulinedimembrana,nérecettoriperlafrazioneC3bdelcomplemento.Possonoperòessereidentificatiperlapresenzadiunagrandevarietà di antigeni di superficie. Il recettore più importante delle celluleT è ilrecettoreperl'antigene:ilcosiddettoTcellreceptoroTcr.Talerecettoreèmoltocomplesso in quanto è strettamente aggregato ad altre proteine di superficie,quali la CD3, presente su tutti i linfociti T e ,inoltre, o alla CD4 o alla CD8, a

22
seconda che il linfocita T abbia, rispettivamente, funzioni di helper o dicitotossico/suppressor. Il linfociti T helper sono cellule ad attività regolatricedellarispostaimmunitariacheesprimonoilfenotipoCD4+eCD8-.Costituisconocirca un terzo della popolazione delle cellule T mature e hanno il ruolo diamplificare la funzione di altre cellule: aiutano i linfociti B a produrre glianticorpieasecernerli; induconoi linfociticitotossiciadifferenziarsiincelluleeffettrici e stimolano i macrofagi e altre cellule non specifiche ad intervenirenellereazioniidipersensibilitàritardata.Questediversefunzionisonosvoltedadiverse sottopopolazioni di linfociti T helper: i linfociti T helper 1 (Th1)stimolano le cellule citotossiche e attivano i macrofagi, promuovendo quindisoprattutto la risposta immunitariacellulo-mediata; i linfocitiThelper2 (Th2)stimolanoinvecelaproliferazionedeilinfocitiBequindilasintesidellediverseclassianticorpali,promuovendospecificatamentelarispostaimmunitariaditipoumorale.LecelluleTh1eTh2produconodiversecitochine:IL-2,IFN-γ,TNF-αeLT-αlecelluleTh1;IL-4,IL-5,IL-6,IL-10eIL-13lecelluleTh2.I linfocitiT citotossici (CTL) sono indotti dall'IL-2 epossonoespandere il loronumeromigliaiadivoltealpiccodellarispostaimmunitaria.IlsistemaCTLlisacelluleallogeniche,cheveicolanoantigenidiistocompatibilitàdiclasseIdiversidai propri e cellule singeniche, cioè con gli stessi antigeni di istocompatibilità,chesianoperòstatimodificatinellalorostruttura.I linfociti T suppressor, o Treg, sono deputati a sopprimere direttamente oattraverso fattori solubili (TGF-β, IL-10 e IL-35), l'attività di altre celluleimmunitarieepiùprecisamentedei linfocitiB,dialcuni tipidi linfocitiTedeimacrofagi.Il recettore con cui i linfociti T riconoscono l'antigene è il recettore, uneterodimero, denominato Tcr che viene espresso sulla membrana inassociazione con un altro complesso molecolare denominato CD3 che ècoinvolto nella trasmissione del segnale all'interno della cellula. Pertanto lastruttura responsabile del riconoscimento dell'antigene estraneo associatoall'antigened’istocompatibilità è rappresentatodall'eterodimeroα/β associatoadunaseriedipeptididenominatinel loro insiemeCD3.Oltreall'eterodimeroα/βèstatoindividuatounaltrotipodieterodimerotransmembranaconcateneγ/δ. Questi due tipi di eterodimeri sono presenti in diverse percentuali nellediversespecie.Nell'uomoil95%-99%deilinfocitiTveicolaTcrα/β,mentreneiruminatiil30-60%veicolaTcrγ/δ.I linfociti T regolatori, Treg, sono una sottopopolazione dei linfociti T chemodula la risposta immunitaria, mantiene la tolleranza agli auto-antigeni e

23
prevengono le malattie autoimmuni. I Treg sono immunosoppressivi egeneralmenterallentanol'induzioneelaproliferazionedeilinfocitiTeffettori.ITregesprimonoibiomarcatoriCD4,FOXP3,eCD25ederivanodallastessalineadeilinfocitiTCD4+(Broereetal.,2011;PolieRocchi,2005).MACROFAGIImacrofagisonocellulemononucleatetissutalicheappartengonoalsistemadeifagociti. Di quest'ultimo sistema fanno parte anche i granulociti neutrofili e imonociti. Essi svolgono un ruolomolto importante nelle risposte immunitarienaturaliespecifiche.Lalorofunzioneprincipaleèlafagocitosicioèlacapacitàdiinglobarenellorocitoplasmaparticelleestranee,compresiimicrorganismi,edidistruggerle. Secernono inoltre citochine ad attività proinfiammatoria epresentanol'antigeneai linfocitiT-CD4.Imacrofagihannounadimensionechevariada12a25μmdidiametroehannounavitamediadicirca75giornieoltre.Originano nelmidollo osseo da cellule precursori e passano nel sangue comemonociti;questi,dopoessererimastiincircolopercirca10-30ore,migranoneitessuti dove si trasformano in macrofagi. I monociti del sangue, cherappresentano circa il 5% dei leucociti circolanti, quando passano nei tessutidivenendo macrofagi, acquisiscono una maggiore attività fagocitaria, che siaccompagnaaunaumentodimensionaledelcitoplasmaeanchenumericodegliorganuli quali lisosomi, microtuboli, microfilamenti e delle membranedell'apparatodiGolgi;inoltreilnucleodivienepiùirregolareeacquisisceunoopiùnucleoli.Monociti e macrofagi, che sono caratterizzati dalla capacità di aderiresaldamente alle superfici solide, di muoversi attivamente, di rispondereintensamente agli stimoli chemiotattici e di essere capaci di fagocitare lesostanzepiùdiverse,organichecomevirus,batteriprotozoi,complessiantigene-anticorpo, sia inorganiche come carbone, silice asbesto, ecc., realizzano unsistemadifensivomoltoefficiente:ilsistemafagocitariomononucleare.L'intensacapacitàfagocitariadiquestecelluleèdovutaallapresenzasullaloromembranadi recettori specifici per le sottoclassi IgG1 e IgG3: tale caratteristica consentelorodi"catturare"specificatamenteedifagocitaregliantigenirinvenutineivaritessuti linfoidie, inarticolaricircostanzediattaccarecelluledimammiferichepresentinosullalorosuperficiel'antigenecorrispondente.Comeglialtrifagociti,presentanoancheirecettoriperlaporzioneFcdeglianticorpi,dopochequestisisonolegaticonl'antigene,erecettoriper la frazioneC3bdelcomplemento. Imacrofagipresentinelsitod'infezionepresentanounaumentodelnumerodei

24
lisosomicontenentienzimiidroliticieinoltreproduconoaltrienzimitraiqualiattivatoridelplasminogeno, collagenasi,elastasi,proteasie lisozima. Inquestomodoacquisisconoun'eccezionalecapacitàdidigerirematerialidiversi, tracuiproteine, polisaccaridi e acidi nucleici. I macrofagi attivati producono anchelinfochinecheintervengononell'attivazionedeilinfociti.Altrifattoriprodottidaimacrofagi includono le frazioni C2, C3, C4, e C5 del complemento, leprostaglandine, l'IFN-αeunfattorechestimola lareplicazionedellecelluledelmidolloosseo.Questecitochinesonoessenzialiperlarisoluzionedelleinfezionimasonoancheresponsabilididannitissutali.Il richiamo e l'attivazione dei fagociti (macrofagi e neutrofili) sono favoriti dafenomeni di chemiotassi, che gli stessi microrganismi invasori inducono,soprattutto attraverso la reazione infiammatoria. Il contatto tra fagocita eparticella estranea viene facilitato dalla presenza su quest'ultima di sostanzeplasmatiche, quali fibronectina o frazioni del complemento, che tendono alegarsi ai batterie alle sostanze particolate in genere. Il massimo di efficienzadellafagocitosisirealizzaquandoimicrorganismisonoricopertidaanticorpi:ilcomplessoantigene-anticorpo tende infattiadaderiremoltoefficacementeallacellula fagocitaria, per la presenza su quest'ultima di recettori specifici permolecoleanticorpalimodificatedal lorolegameconl'antigene.Inalternativa, ilcomplesso antigene anticorpo può fissarsi al fagocita per l'intervento dicomponenti del complemento che, da un lato, si legano alla porzione Fcdell'anticorpo,dall'altroaspecificirecettori(CD16)cheimacrofagipresentanosulla membrana. I macrofagi possono essere identificati tramite citometria oimmmunoistochimica attraverso la messa in evidenza di specifiche proteinecome il CD14, CD40, CD11b, CD64, F4/80 (topo)/EMR1 (uomo), lisozima M,MAC-1/MAC-3eCD68(Khazenetal.,2005).IlrecettoreCD14riconosceLPSdeibatteriGramnegativi.ImacrofagialveolaridelsuinoesprimonoiTLR3,iTLR2eiTLR6(Zhangetal.,2013;Munetaetal.,2003).LINFOCITIBI linfociti B, responsabili della risposta immunitaria di tipo umorale, sono ingrado, differentemente dai linfociti T, di riconoscere e reagire, tramite ilspecificorecettore(BCR),direttamenteconepitopipresentisullasuperficiedeimicrorganismi invasori o su qualunque altra sostanza estranea solubile,rilasciata dal patogeno. I linfociti B si sviluppano dalle cellule staminaliematopoietichedelmidolloosseoequimaturano. Il linfocitaBmaturoveicolasulla sua superficie immunoglobuline di una determinata classe anticorpale

25
specifiche per un determinato antigene, per cui solo quest'ultimo vienericonosciutodatalelinfocita,mentretuttiglialtriantigenivengonoignorati.Lamaturazione di queste cellule attraversa diversi stadi caratterizzati dallapresenza di diversi isotipi: durante l'ontogenesi, la prima cellula identificabilecomeappartenenteallalineadeilinfocitiB,èlacellulapre-B,caratterizzatadallapresenza di IgM citoplasmatiche, ma non di membrana; lo stadio successivodella cellula B immatura è caratterizzato dalla presenza di IgM anche sullamembrana.Inquestostadioilinfocitisonoestremamentesensibiliall'induzionedellatolleranzaneiconfrontidiantigenichedovesseroincontrare.SipassapoiallecelluleBmaturecheacquisiscono lacapacitàdiprodurreunadeterminataclassa anticorpale (IgG, IgA, IgE, o IgM); questi linfociti presentano sullamembrana,oltrealleIgM,anchel'isotipochesarannoingradodiprodurredopol'incontro con l'antigene. Durante le varie fasi di maturazione sono espressesulla superficie diverse proteine, nelle cellule mature sono espresse la CD4,CD19,CD20,CD21,CD22,CD23,CD32,CD35,CD40,CD72,CD79αelemolecoleMHCdiclasseIediclasseII.L'attivazionedeilinfocitiBpuòessereindottadalsemplice legame con l'antigene col BCR, con l'aiuto di frammenti delcomplementooppuredall'attivazionedeiTLRcheattivanoalorovoltailinfocitistessi. In particolare, per quest'ultima via bisogna ricordare che i linfociti Besprimono i TLR5, TLR7 e TLR9 che riconoscono rispettivamente flagellinabatterica, RNA a singola elica e sequenze non metilate in DNA presenti negliendosomi. Per lamaggior parte degli antigeni proteici l'attivazione non causaunarispostadirettadiproduzionedianticorpi,bensìl'internalizzazionedelBCRlegantel'antigeneperprocessarloedesporlosullemolecoleMHCdiclasseIIinattesacheilinfocitiThelperloriconoscano.Lostimoloaun'intensareplicazioneedifferenziazionedei linofcitiB, giàattivati, avvieneadoperadei linfocitiTH2siapercontattodiretto,siaattraversolaproduzionediIL-4(PolieRocchi,2005;RossereMauri,2015;LeBieneTedder,2008).

26
MaterialieMetodi
PrelievodeicampioniDuranteil2016sonostativalutatialmacellogliscorepolmonaridi5partitedisuinigrassiprovenientida5diversiallevamenti,chepraticano lavaccinazioneperM.hyopneumoniaecondueinoculi,unoa15giornidivitaeunoacirca180giorni.Perognipartita,compostada130-140animali,sonostativalutatialmeno100polmonisecondoilmetodopropostodaMadeceKobish(1982)perdefinireunoscorepolmonare individuale.Daognipartitasonostatiprelevaticampionidi polmoni con lesioni riconducibili a M. hyopneumoniae per la ricerca delmicoplasma tramite real time PCR come descritto da Marois e coll. (2010),eseguitapresso laSezionediDiagnosticadiBresciadell'IstitutoZooprofilatticoSperimentale.Per le ricerche istologiche ed immunoistochimiche, eseguite presso Sezione diAnatomia Patologica e Patologia Generale del Dipartimento di ScienzeMedicoVeterinarie dell’Università degli Studi di Parma, sono stati prelevati campionirappresentativi dei polmoni con lesioni e campioni di tessuto adiposomediastinicodellostessoanimale,veditabella2.I campioni di tessuto adiposo sono stati divisi in due aliquote: una è statacongelata in azoto liquidoeunaè stata inclusa inparaffina. I tessuti congelatisonostatisezionatia5µmalcriostatoepostisuvetrinipolilisinati.
Allevamento NumeropolmonicampionatiperricercaM.hyopneumoniae
Numerocampioniper
immunoistochimicaA 46 13B 11 6C 14 10D 7 7E 16 13
Tabella 2: campioni utilizzati per la ricerca diM. hyopneumoniae e per le indaginiistologicheedimmunoistochimiche.

27
IstologiaIcampionidipolmoni,49traquellicampionati,32RT-PCRpositivie17negativi,sonostatidisidratatieinclusiinparaffina.Successivamentesonostateottenutesezioni di 5µm su vetrini polisinati (Buchwalow e Böcker, 2014). Per ognicampione, dopo colorazione con ematossilina-eosina, sono state valutatel'attivazionedeiBALT,ladisepitelizzazionedell'epiteliobronchialeelapresenzad’infiammazione bronco-polmonare acuta attraverso la valutazione degliaggregati neutrofilici-macrofagici nei lumi bronchiali ed alveolari. Per laclassificazione delle lesioni sono stati stabiliti 4 livelli per ciascun parametrosopraelencato,da0a3.
ImmunoistochimicaLe sezioni di tessuto polmonare incluso in paraffina sono state colorate contecnica immunoistochimica avidina-biotina-perossidasi per fenotipizzare itessuticonglianticorpiriportatiintabella3.Le sezioni ottenute da tessuti fissati in formalina sono state sottoposte asmascheramento antigenico, in quanto la fissazione maschera i determinantiantigenici per la formazione di legami crociati. Le perossidasi endogene sonostatebloccateconidrogenoperossido(H2O230%w/w)al3%inPBSperitessutiinclusiinparaffinaeallo0.3%perlesezioni.Tuttelesezionisonostatetrattatecon siero normale di capra diluito al 10% in PBS per eliminare i legami conepitopi aspecifici. In seguito, tutte le sezioni sono state incubate con unanticorpo secondario biotinilato anti anti-mouse IgG-B (sc-2039 Santa CruzBiotechnology, inc.)1:200 inPBSper30minuti.L’amplificazionedelsegnaleèstataottenutaconVectastainEliteABCkitdiVectorLaboratoriesper30minutie la colorazione della reazione è stata effettuata con diaminobenzidina DAB(DAKOCytomationLiquidDAB+SubstrateChromogenSystem)applicataper5-10minuti.Comecontrocolorazionenucleareèstatautilizzata l’ematossillinadiMAYER per 5 minuti, quindi le sezioni sono state disidratate e coperte convetrinicoprioggetto(BuchwaloweBöcker,2014).La valutazione della presenza dei linfociti Treg è stata effettuata contando lecellulecoloratepositivamenteinalmenoquattroBALTpercampione.Lapresenzadeimacrofagi, dei linfociti T e dei linfociti B è stata valutata su4campivisivipercampionia200 ingrandimenti, conunascalada0a4:0per i

28
campioni in cui non si rilevavano cellule colorate, 1 dove si rilevavanomediamentefinoa20cellule,2da20a60cellule,3da60a100cellulee4pericampioniconpiùdi100cellule.Per la fenotipizzazione dei TLR-4 nei tessuti adiposi mediastinici, è statonecessario utilizzare ilmateriale congelato in azoto liquido perché l’anticorpomonoclonaleanti-TLR-4sièrivelatoinefficaceneicampionifissatiinparaffina.Icampioni congelati in azoto liquido, non subendo la fissazione, presentano ideterminanti antigenici non mascherati sulla loro superficie. La valutazionedell'espressione dei TLR4 è stata effettuata su 20 campioni, utilizzando unaclassificazione semi quantitativa in base alla diffusione ed intensità dellacolorazionesuquattrolivelli:assente,lieve,media,intensasuquattrocampia4ingrandimenti.
Tabella3:anticorpiutilizzatiperl'immunofenotipizzazionecellulare.
RISULTATIIn tutte le partite di animali sono state riscontrate lesioni riconducibilimacroscopicamenteaM.hyopneumoniae;gliscorepolmonarimedidellepartitevisionate hanno variato da 0,14 a 1,98 (vedi tabella 4). La percentuale dicampionidipolmoneperciascunapartitarilevatipositiviallaRT-PCRèvariatadallo0%al100%;sonorisultatenegative2partite(veditabella4).
Anticorpo(Clone)
TipoDiAb Produttore Diluizione SmascheramentoAntigenicoinMicroonde(4CicliA800Watt)
TempiDiIncubazione
Numerodicampioniprocessati
CD3(CD3-12)
Monoclonale AbDSerotec
1:100 TamponepH9 1Ora 16pos7neg23tot
CD79α(HM57)
Monoclonale AbDSerotec
1:30 TamponepH6 1Ora 16pos7neg23tot
MAC387(MAC387)
Monoclonale AbDSerotec
1:100 TamponepH6 1Ora 16pos7neg23tot
Foxp3(FJK-16s)
Monoclonale eBioscience 1:200 TamponepH9 Overnight4°
29pos10neg39tot
TLR-4(76B357.1)
Monoclonale Abcam 1:100 NonNecessario(TessutoCongelato)
1Ora 14pos6neg20tot

29
Partita NumeropolmoniRT-PCRpositivi
Mediascorepolmonare
A 46/46(100%) 1,98B 0/11(0%) 0,16C 0/14(0%) 0,18D 6/7(85,7%) 0,14E 16/16(100%) 0,41Tot 68/94(69,4%) /
Tabella4:RisultatidellaricercadiM.hyopneumoniaetramiteRT-PCRemediadelloscorepolmonarerilevatinellediversepartitecampionate.Le lesioni più significative rilevate istologicamente erano caratterizzate dapolmonitecatarralebroncointerstizialeconaumentodivolumeperibronchiale,peribronchiolareeperivascolaredeltessutolinfoideassociatoaibronchi(BALT)da lieve a notevole. In questo ultimo caso vi era anche lo sviluppo di follicolilinfatici. Nei casi più gravi si osservava la riduzione del lume bronchiale,bronchiolareealveolare indottadall’aumentodivolumedeiBALT.Gliepiteliaridossodeinoduliabnormementesviluppati,sipresentavanospessoesfoliatiinmodopiùomenomarcato.Lecellulecaliciformibronchialiapparivanotalvoltaiperplastiche.Vedifigure5,6e7.In 10 campioni si osservavano aspetti di flogosimista, sia interstizialemapiùspessoessudativa.Lapolmoniteinterstiziale,chemostravaestensionevariabile,da focale a multifocale confluente a diffusa, era caratterizzata dall’accumuloprevalentementealveolo-settale,maancheperibroncovascolare,dimacrofagielinfociti. Non erano infrequenti casi di alveolite macrofagica con numerosimacrofagiessudatinellumedeglialveoliadistribuzionelobularefocale.La componente essudativa della flogosi polmonare era riconducibile ad unabroncopolmoniteo,piùraramenteadunacomponentefibrinosaalveolaremistaadessudatodicellulerappresentatodamacrofagiegranulocitineutrofili (vedifigura7).Nelle partite con campioni risultati positivi alla RT-PCR, sono state rilevatelesioni caratteristiche dell'infezione daM.hyopneumoniae, spesso connotevoliinfiltrati di neutrofili nel lume alveolare. Le alterazioni più intense sono staterilevateneicampioniRT-PCRpositivi.I Treg più numerosi (vedi figura 15) sono stati riscontrati nei campioni conminorinfiammazionealivellopolmonare,mentreladdovequestaeraalta,iTregsonostatimenofrequenti.LemediedellecontedeiTregsonovariateda0a13,5cellule.L'utilizzodell'antiTLR4sutessutoadiposohapermessodidistinguere4livellidiintensitàdellacolorazione(vedifigura16).Icampioniconlacolorazionepiù

30
intensa per i TLR4, campione A13 e E6, entrambi positivi alla RT-PCR, hannomostrato anche un intenso stato infiammatorio a livello polmonare convalutazionidiinfiammazionebronco-alveolarepiùelevataeunaminorpresenzadiTregalivellodeiBALT.Viceversa,10campioniconvalutazionedeiTLR4paria0e1,comprendenti2RT-PCRpositivi,mostravanoungradod'infiammazionealveolo-bronchialeconnessunoopochimacrofagieneutrofilieunamediadellapresenzadiTregneiBALTdi3,9cellule.Nei campioni con più cellule CD3 positive sono state riscontrate anchemoltecelluleCD79αpositive.
Figura5:polmonedisuino.Siosserva iperplasiadelBALTconparzialeriduzionedellumebronchiolare.Colorazioneematossilina-eosina,ingrandimento4X.CampioneA1.

31
Figura6:polmonedisuino.SiosservanotevoleiperplasiadeiBALTconinvasionedelparenchimapolmonare.Colorazioneematossilina-eosina,ingrandimento4X.CampioneD1.

32
Figura7:polmonedi suino. Si osserva infiltrazionemassivadineutrofili emacrofagidei setti alveolari con invasione del lume alveolare. Depositi di fibrina nel lumealveolare.Colorazioneematossilina-eosina4X.CampioneA12.
Figura 8: polmoni di suino. Colorazioni con anticorpi antiCD3 a), antiCD79α b) eantiMAC387c).Morfologiacellulare.a)linfocitiT;b)linfocitiB;c)macrofagi.
a) b) c)

33
Figura9:polmonedisuino.ImmunofenotipizzazioneconanticorpoantiCD3.Iperplasiadei BALT. Si osservano numerosi linfociti T nella zona mantellare dei centrigerminativi. Rari linfociti T si trovano nella zona centrale dei centri germinativi.Ingrandimento40X.CampioneD6.

34
Figura 10: polmone di suino. Immunofenotipizzazione con anticorpo anti CD79α . SiosservanonumerosilinfocitiBalivellodimantelloe,talvolta,nelcentrogerminativo.Ingrandimento4X.CampioneD6.

35
Figura 11: polmone di suino. Immunofenotipizzazione con anticorpo anti CD3. NellasottomucosabronchialesiosservanonumerosilinfocitiTinfiltranti.AnchealivellodiparetialveolarisinotainfiltrazionedilinfocitiT.Ingrandimento4X.CampioneD1
36
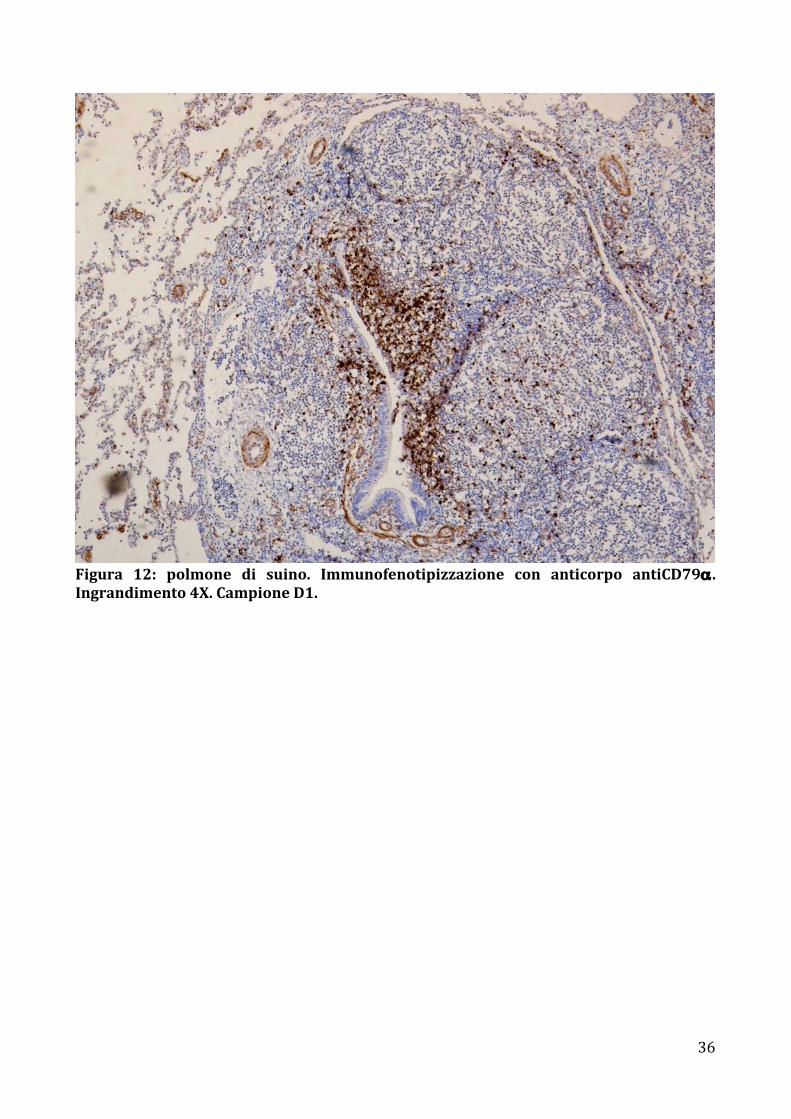
Figura 12: polmone di suino. Immunofenotipizzazione con anticorpo antiCD79α .Ingrandimento4X.CampioneD1.

37
Figura13:polmonesuino.ImmunofenotipizzazioneconanticorpoMAC387.Bronchiolicon infiltrazione di macrofagi nel lume ed intensa infiltrazione di macrofagi neglialveoli.Ingrandimento4X.CampioneA5.

38
Figura 14: polmone di suino. Si osserva iperplasia del BALT con scarsa presenza dicellulemacrofagiche immunocolorate conantiMAC387.Le cellulemacrofagichesonolocalizzateaimarginideltessutolinfoide.CampioneD5.Ingrandimento4X.

39
Figura15:polmonedisuino.ImmunofenotipizzazioneconanticorpoFoxp3.BALTconnumerosiTregcoloratipositivamente.CampioneB3.Ingrandimento10X.
Figura 16: tessuto adiposo suino. Immunofenotipizzazione con anticorpi anti TLR4.L'antigene rilevato dagli anticorpi definisce i margini delle cellule adipose.Ingrandimento4X.a)campioneD1;b)campioneE6.
b)a)

40
Campione RT-PCR TLR4 BALT Epit. Neutr. Treg CD3 MAC CD79α ScoreA1 + n.d. 1 1 0 1,25 1 1 2 1A2 + n.d. 2 0 0 0,5 n.d. n.d. n.d. 1A3 + 1 1 0 2 4 3 3 1 1A4 + n.d. 2 2 3 2,5 n.d. n.d. n.d. 1A5 + 1 3 2 3 2 2 2 3 2A6 + n.d. 2 2 2 5,5 3 3 2 3A7 + n.d. 0 0 0 5,75 n.d. n.d. n.d. 4A8 + n.d. 2 2 3 0 2 3 3 7A9 + n.d. 1 2 3 0 n.d. n.d. n.d. 5A10 + n.d. 3 3 1 5,7 3 2 3 8A11 + n.d. 2 3 3 n.d. 1 4 1 2A12 + 2 1 3 3 0 1 4 1 2A13 + 3 2 3 3 0 n.d. n.d. n.d. 8B1 - 2 0 0 0 3 2 1 1 1B2 - n.d. 0 0 0 3,75 n.d. n.d. n.d. 1B3 - n.d. 0 0 0 13,5 n.d. n.d. n.d. 1B4 - n.d. 0 0 0 6 n.d. n.d. n.d. 1B5 - 3. 0 0 0 n.d. n.d. n.d. n.d. 2B6 - n.d. 0 0 0 n.d. n.d. n.d. n.d. 1C1 - n.d. 0 0 0 2,3 1 1 1 2C2 - n.d. 0 0 0 6,7 n.d. n.d. n.d. 1C3 - 1 0 0 0 7,3 n.d. n.d. n.d. 1C4 - 1 1 0 0 0,7 1 1 1 1C5 - n.d. 0 0 0 4 1 1 1 1C6 - n.d. 1 0 0 4 n.d. n.d. n.d. 1C7 - n.d. 2 1 0 5 n.d. n.d. n.d. 1C8 - 2 0 0 0 n.d. 1 1 1 1C9 - n.d. 0 0 0 n.d. n.d. n.d. n.d. 1C10 - 2 0 0 0 n.d. 1 1 1 1D1 + 0 3 2 0 n.d. 4 1 4 3D2 + n.d. 2 1 0 n.d. n.d. n.d. n.d. 1D3 + 0 0 0 0 n.d. n.d. n.d. n.d. 1D4 + 2 0 0 1 4 n.d. n.d. n.d. 2D5 + 1 3 2 1 0,2 4 1 4 2D6 + n.d. 3 2 2 0,3 4 1 2 3D7 - n.d. 2 0 0 1,3 3 1 1 4E1 + 1 0 0 0 7,3 n.d. n.d. n.d. 3E2 + n.d. 1 2 2 1,7 1 3 1 4E3 + 1 2 1 0 4,7 n.d. n.d. n.d. 1E4 + n.d. 2 0 0 11,5 2 2 1 2E5 + 2 1 1 2 3 n.d. n.d. n.d. 1E6 + 3 1 3 3 0 n.d. n.d. n.d. 3E7 + n.d. 2 2 0 n.d. n.d. n.d. n.d. 3E8 + n.d. 0 0 0 2 3 3 2 7E9 + n.d. 3 3 3 5,5 n.d. n.d. n.d. 2E10 + n.d. 2 3 0 1 n.d. n.d. n.d. 5E11 + 1 1 2 2 4,5 1 3 1 3E12 + n.d. 1 1 0 n.d. n.d. n.d. n.d. 3E13 + n.d. 2 2 2 0,7 3 1 3 3
Tabella5:risultatidellavalutazioneistologicaeimmunoistochimica.PCR:"+"positivoallaRT-PCR, "-" negativo; TLR4: risultato della valutazione del TLR4nei campioni ditessuto adiposo; BALT: valutazione dell'attivazione del BALT; Epit.: valutazione delladisepitelizzazione dell'epitelio bronchiale; Neutr.: valutazione della presenzad’infiammazione bronco-polmonare acuta attraverso la valutazione degli aggregatineutrofilici-macrofagici nei lumi bronchiali ed alveolari; Treg: media delle 4 contedelle cellule colorate dall'anticorpo in 4 differenti BALT; CD3: valutazione dellapresenza di cellule colorate dall'anticorpo (linfociti T); MAC: valutazione dellapresenza di cellule colorate dall'anticorpo (macrofagi); CD79α: valutazione dellapresenza di cellule colorate dall'anticorpo (linfociti B). Score: score polmonare.

41
CONCLUSIONII rilievi al macello hanno confermato che l'infezione daM. hyopneumoniae èdiffusanelsistemadall'allevamentosuinicoloitalianoechelavaccinazionenonpreviene l'infezione,maèutile adiminuirne lapatogenicità, come rilevatodaibassipunteggialloscorepolmonaredellepartiteesaminate.Icampioninegativi,benché tutti presentassero lesioni macroscopiche, compatibili coninfiammazionebroncopolmonaredeilobiapicali,possonoessererisultatitaliinquantolelesioninonsonopatognomonichedell'infezionedaM.hyopneumoniaeesoprattuttoperchéquestopatogeno,abbassando ledifese immunitarie locali,favorisce l'insorgenza di infezioni secondarie, prima di essere eliminato dallarispostadell'ospite.Il quadro istologico rilevato è sovrapponibile a quello precedentementedescrittodaaltriautorineiquadrid'infezionedaM.hyopneumoniae(Kwonetal.,2002;Sarradelletal.,2003;Opriessnigetal.,2004;Choietal.,2006;Lorenzoetal., 2006) caratterizzato da iperplasia linfocitaria peribronchiolare eperivascolare, macrofagi e neutrofili nel lume delle vie aeree. L'infiltrazionemacrofagicaeneutrofilicadel lumeedel settoalveolareevidenziata indiversicampioniè rappresentativadiunostato infiammatorioavanzato, riconducibilesia all'infezione pura da M. hyopneumoniae (Sarradell et al., 2003) sia allapresenza di germi d'irruzione secondaria. Queste lesioni sono riconducibiliall'accumulodimuco e di essudato infiammatorionel lumebronchiale dovutoalladiminuzionedell'attivitàciliare,allabroncocostrizionedamediatorichimicirilasciatidaimacrofagialveolarieallapresenzadiaggregatilinfatici.Lapresenzadimacrofaginellumedibronchiebronchioli,eneisettialveolarida14 a 28 giorni dopo l’infezione daM. hyopneumoniae è stata documentata daRedondo e coll. (2009) confermando che queste cellule hanno un ruoloimportante nello sviluppo della risposta infiammatoria. Oltre che nel lumealveolare in corso di un'intensa risposta immunitaria, nel nostro studio,numerosimacrofagisonostadievidenziatinellezonemarginalidiBALTreattivied iperplasici. Quest’attivazione dei BALT può essere dovuta agli stessimacrofagiche,stimolatidallapresenzadiantigenibatterici,produconocitochinepro-infiammatorie(Thanawongnuwechetal.,2003).Neiquadriistologiciincuivi era una forte componente infiammatoria subacuta caratterizzata dallamassiva presenza di neutrofili e macrofagi, le altre componenti del sistemaimmunitario,linfocitiB,linfocitiTeTreg,sisonomostratepocorappresentate.NelcontempoanchelareazionedeiBALTsimostravablandacontessutilinfoidi

42
pocoopernullareattivi.Assiemeallacomponentemacrofagicasiosservavaunacomponenteneutrofilicachel'accompagnavanellarispostaimmunitaria.Questoè determinato dal fatto che a livello polmonare, oltre alla popolazionemacrofagica stabile, è presente, a ridosso dell’endotelio, una popolazione dimonociti con funzioni di controllo (Auffray et al., 2007), essenziale per uninterventorapidoneltessutoinfiammatoelaformazionedimacrofagitissutali.Nelpolmoneinparticolare,èstatoriportatouninteressantelegametramonocitie neutrofili: utilizzando tecniche d'imaging in vivo, è stato osservato che ladeplezione di monociti del sangue danneggia il reclutamento di neutrofilitramite migrazione transendoteliale nel polmone, dimostrando così che ilreclutamentodineutrofilidurantel'infiammazionepolmonareèdipendentedaimonociti(Kreiseletal.,2010).Analogamenteaquanto rilevatonel tessutoadiposoumano (Bès-Houtmannetal., 2007), è stata confermata la presenza di TLR4 nel tessuto adiposomediastinico del suino. L'aumento dell'espressione dei TLR4 nei tessuti coninfiammazioneinattoèstatorilevatoneltessutomammarioincorsodimastitibatteriche nel bovino (Goldammer et al., 2004). Nell'uomo, diverse citochineinduconol'aumentodell'espressionedeiTLR4ancheintessutinondirettamentecoinvolti da processi infiammatori. Ricerche hanno confermato l'aumentodell'espressionedeiTLR indiverse celluledel sistema immunitario causatodadiverse citochine (Yang et al., 2009; Derkow et al., 2013). L'analisisemiquantitativa della presenza dei TLR4 nel tessuto adiposo mediastinico,svoltainquestaricerca,tendeaconfermarequestidatirilevandovaloripiùaltiin quei soggetti chemostravano un'infiammazione più intensa o più diffusa alivello polmonare; ulteriori studi sono comunque necessari per confermarequestaipotesi.IlinfocitiTsonostatirilevatisoprattutoinquadriistologiciconmoderataoforteiperplasiadeiBALT,conscarsainfiltrazionemacrofagicaoneutrofilicaalivellodegli alveoli. Sianei quadri conmodico interessamentodeiBALT, sia inquellicon forte interessamento, i linfociti T si mostravano diffusi nelle diversestrutturepolmonari,sottomucosabronchiolarecompresa.IlinfocitiBpresentisoprattuttoneicentrigerminativiealivellodelmantellodeiBALTe in vicinanzadei bronchi e dei bronchioli, si sono rivelate scarsamentepresentinelleformeconcomplicazionibattericheemaggiormenteevidenziabilinelleformepiùpure.I linfociti Treg, Foxp3+, hanno, nel suino come nell'uomo, proprietàimmunosoppressive (Kaser etal., 2008) e quest’attività può essere indotta da

43
patogeni,comeilPRRSV(Silva-Campaetal.,2009). IlriscontrodiunamaggiorpresenzadiTregneiBALTdeicampioniistologicichemostravanounquadrodiscarsaonullaattivazionedeifenomeniimmunitari,suggerisceunruoloattivodiqueste cellule per mantenere un basso livello di attivazione del sistemaimmunitario a livello locale. Viceversa nei quadri istologici con maggiorattivazionedelsistemaimmunitario,sisonorilevatipochiorariTregneiBALT.Le tecniche istologiche ed immunoistologiche utilizzate, ci hanno permesso diinquadrare la risposta immunitaria del suino alle infezioni polmonari conpartecipazione di MH, rilevando la complessità dell’intervento delle diversecelluledelsistemaimmunitario.

44
AbbreviazioniBALTtessutolinfoideassociatoaibronchiLPSlipopolisaccarideCTLlinfocitiTcitotossiciMHMycoplasmahyopneumoniaeMMTVvirusmurinodeltumoremammarioRSVvirusrespiratoriosincizialeTRLtoll-likereceptorRT-PCRrealtimepolymerasechainreaction

45
BIBLIOGRAFIA1) Alexopoulou L, Holt AC, Medzhitov R, Flavell RA (2001) Recognition of double-
strandedRNAand activationofNF-kappaBbyToll-like receptor3.Nature413:732–738.
2) Auffray C, Fogg D, GarfaM, Elain G, Join-Lambert O, Kayal S, Sarnacki S, Cumano A,Lauvau G, Geissmann F. Monitoring of blood vessels and tissues by a population ofmonocyteswithpatrollingbehavior.Science.2007Aug3;317(5838):666-70.
3) Alvarez B, de la Riva PM, Handler A, Revilla C, Alonso F, Ezquerra A, Domínguez J.ExpressionofTLR4inswineasassessedbyanewlydevelopedmonoclonalantibody.VetImmunolImmunopathol.2013May15;153(1-2):134-9.
4) AlvarezB,RevillaC,DoménechN,PérezC,MartínezP,AlonsoF,EzquerraA,DomíguezJ. Expressionof toll-like receptor 2 (TLR2) in porcine leukocyte subsets and tissues.VetRes.2008Mar-Apr;39(2):13.
5) Alvarez B, Revilla C, Chamorro S, Lopez-Fraga M, Alonso F, Dominguez J, et al.Molecularcloning,characterizationandtissueexpressionofporcinetoll-likereceptor4.DevCompImmunol(2006)30:345–55.
6) Alving CR, PeachmanKK, RaoM, Reed SG. Adjuvants for human vaccines. CurrOpinImmunol(2012)24:310–5.
7) Amass,S.F.,Clark,L.K.,VanAlstine,W.G.,Bowersock,T.L.,Murphy,D.A.,Knox,K.E.andAlbregts,S.R.(1994).InteractionofMycoplasmahyopneumoniaeandPasteurellamultocidainfectionsinswine.AmericanJournalofVeterinaryResearch,204,102-107
8) Andersen-NissenE,SmithKD,StrobeKL,BarrettSL,CooksonBT,LoganSM,AderemA(2005)EvasionofToll-likereceptor5by flagellatedbacteria.ProcNatlAcadSciUSA102:9247–9252.
9) Andrada M, Quesada-Canales O, Suárez-Bonnet A, Paz-Sánchez Y, Espinosa de LosMonterosA,RodríguezF.Cyclooxygenase-2expressioninpigsinfectedexperimentallywithMycoplasmahyopneumoniae.JCompPathol.2014Aug-Oct;151(2-3):271-6.
10) Asai,T.,M.Okada,M.Ono,T. Irisawa,Y.Mori,Y.Yokomizo,S.Sato. (1993) Increasedlevelsoftumornecrosisfactorandinterleukin1inbronchoalveolarlavagefluidsfrompigsinfectedwithMycoplasmahyopneumoniae.VetImmunolImmunopathol38,253-260.
11) Asai,T.,M.Okada,M.Ono,Y.Mori,Y.Yokomizo&S. Sato:Detectionof interleukin-6and prostaglandin E2 in bronchalveolar lavage fluids of pig experimentally infectedwithMycoplasmahyopneumoniae.VetImmunolImmunopath44,97-102(1994).
12) Bès-Houtmann S, Roche R, Hoareau L, Gonthier MP, Festy F, Caillens H, Gasque P,Lefebvred'HellencourtC,CesariM.Presenceof functionalTLR2andTLR4onhumanadipocytes.HistochemCellBiol.2007Feb;127(2):131-7.
13) BiebackK, LienE,Klagge IM,AvotaE, Schneider-Schaulies J,DuprexWP,WagnerH,KirschningCJ, TerMeulenV, Schneider-Schaulies S (2002)Hemagglutinin protein ofwild-typemeaslesvirusactivatestoll-likereceptor2signaling.JVirol76:8729–8736.
14) BroereF,S.G.Apasov,M.V.Sitkovsky,W.vanEden.TcellsubsetsandTcell-mediatedimmunityinF.P.NijkampandM.J.Parnham,2011PrinciplesofImmunopharmacology:3rdrevisedandextendededition.
15) BrozP,MonackDM.Newlydescribedpattern recognition receptors teamup againstintracellularpathogens.NatRevImmunol.2013Aug;13(8):551-65.
16) Buchwalow IB, BöckerW. Immunohistochemistry: Basics andMethods 31 ott 2014.SpringerEditore.
17) Burkey TE, Skjolaas KA, Dritz SS, Minton JE. Expression of toll-like receptors,

46
interleukin8,macrophagemigrationinhibitoryfactor,andosteopontinintissuesfrompigs challenged with Salmonella enterica serovar typhimurium or serovarcholeraesuis.VetImmunolImmunopathol(2007)115:309–19.
18) Caruso, J.P. & R.F. Ross: Effects of Mycoplasma hyopneumoniae and Actinobacillus(Haemophilus)pleuropneumoniaeinfectiononalveolarmacrophagefunctioninswine.AmJVetRes51,227-231(1990).
19) Chaung,H.C.,Chen,C.W.,Hsieh,B.L.,Chung,W.B.,2010.Toll-likereceptorexpressionsin porcine alveolar macrophages and dendritic cells in responding to poly ICstimulation and porcine reproductive and respiratory syndrome virus (PRRSV)infection.Comp.Immunol.Microbiol.Infect.Dis.33,197–203.
20) Choi SH, Harkewicz R, Lee JH, Boullier A, Almazan F, Li AC, et al. Lipoproteinaccumulation inmacrophages via toll-like receptor-4-dependent fluid phase uptake.CircRes(2009)104:1355–63.
21) Choi, C., Kwon, D., Jung, K., Ha, Y., Lee, Y. H., Kim, O. et al. (2006). Expression ofinflammatory cytokines in pigs experimentally infected with Mycoplasmahyopneumoniae.JournalofComparativePathology,134,40-46.
22) Compton T, Kurt-Jones EA, BoehmeKW, Belko J, Latz E, GolenbockDT, Finberg RW(2003)Humancytomegalovirusactivates inflammatory cytokine responsesviaCD14andToll-likereceptor2.JVirol77:4588–4596.
23) BorjiginL,ShimazuT,KatayamaY,LiM,SatohT,WatanabeK,KitazawaH,RohSG,AsoH,KatohK,UchidaT,SudaY,SakumaA,NakajoM,SuzukiK.ImmunogenicpropertiesofLandracepigsselectedforresistancetomycoplasmapneumoniaofswine.AnimSciJ.2016Mar;87(3):321-9.
24) Dennis V.A., S. Dixit, S.M. O’Brien, X. Alvarez, B. Pahar, M.T. Philipp, Live BorreliaburgdorferispirocheteselicitinflammatorymediatorsfromhumanmonocytesviatheToll-likereceptorsignalingpathway,Infect.Immun.77(2009)1238–1245.
25) DerkowK,BauerJM,HeckerM,PaapBK,ThamilarasanM,KoczanD,SchottE,DeuschleK, Bellmann-Strobl J, Paul F, Zettl UK, Ruprecht K, Lehnardt S. Multiple sclerosis:modulation of toll-like receptor (TLR) expression by interferon-β includesupregulation of TLR7 in plasmacytoid dendritic cells. PLoS One. 2013 Aug12;8(8):e70626.
26) Diebold SS, Kaisho T, Hemmi H, Akira S, Reis E, Sousa C (2004) Innate antiviralresponses by means of TLR7-mediated recognition of single-stranded RNA. Science303:1529–1531.
27) Djordjevic,S.P.,G.J.Eamens,L.F.Romalis,P.J.Nicholls,V.Taylor&J.Chin:Serumandmucosal antibody responses and protection in pigs vaccinated against Mycoplasmahyopneumoniae with vaccines containing denatured membrane antigen pool andadjuvant.AustVetJ75,504-511(1997).
28) Escobar J, VanAlstineWG,BakerDH, JohnsonRW.Growthperformance andwhole-bodycompositionofpigsexperimentallyinfectedwithMycoplasmahyopneumoniae.JAnimSci.2002Feb;80(2):384-91.
29) Figueiredo RT, Fernandez PL, Mourao-Sa DS, Porto BN, Dutra FF, Alves LS, et al.Characterization of heme as activator of toll-like receptor 4. J Biol Chem (2007)282:20221–9.
30) FitzgeraldKA,RoweDC,GolenbockDT.EndotoxinrecognitionandsignaltransductionbytheTLR4/MD2-complex.MicrobesInfect(2004)6:1361–7.
31) GewirtzAT,NavasTA,LyonsS,GodowskiPJ,MadaraJL(2001)Cuttingedge:bacterialflagellinactivatesbasolaterallyexpressedTLR5 to induceepithelialproinflammatorygeneexpression.JImmunol167:1882–1885.
32) Goldammer,T.,Zerbe,H.,Molenaar,A.,Schuberth,H.J.,Brunner,R.M.,Kata,S.R.,Seyfert,

47
H.M., 2004. Mastitis increasesmammarymRNA abundance of beta-defensin 5, Toll-like-receptor2(TLR2),andTLR4butnotTLR9incattle.Clin.Diagn.Lab.Immunol.11,174–185.
33) GondokaryonoSP,UshioH,NiyonsabaF,HaraM,TakenakaH, JayawardanaST,etal.TheextradomainAoffibronectinstimulatesmurinemastcellsviatoll-likereceptor4.JLeukocBiol(2007)82:657–65.
34) HayashiF,SmithKD,OzinskyA,HawnTR,YiEC,GoodlettDR,EngJK,AkiraS,UnderhillDM,AderemA(2001)The innate immuneresponse tobacterial flagellin ismediatedbyToll-likereceptor5.Nature410:1099–1103.
35) Haynes LM, Moore DD, Kurt-Jones EA, Finberg RW, Anderson LJ, Tripp RA.Involvementoftoll-likereceptor4ininnateimmunitytorespiratorysyncytialvirus.JVirol(2001)75:10730–7.
36) HeilF,HemmiH,HochreinH,AmpenbergerF,KirschningC,AkiraS,LipfordG,WagnerH, Bauer S (2004) Species-specific recognition of single-stranded RNA via toll-likereceptor7and8.Science303:1526–1529.
37) HemmiH,TakeuchiO,KawaiT,KaishoT, SatoS, SanjoH,MatsumotoM,HoshinoK,Wagner H, Takeda K, Akira S (2000) A Toll-like receptor recognizes bacterial DNANature408:740–745.
38) Ireton GC, Reed SG. Adjuvants containing natural and synthetic toll-like receptor 4ligands.ExpertRevVaccines(2013)12:793–807.
39) IshiiM,HashimotoM,OgumaK,KanoR,MoritomoT,HasegawaA.Molecularcloningand tissue expression of canine Toll-like receptor 2 (TLR2). Vet ImmunolImmunopathol.2006Mar15;110(1-2):87-95.
40) LeBienTW,TedderTF. B lymphocytes: how they develop and function. Blood. 2008Sep1;112(5):1570-80.
41) LizundiaR,SauterKS,TaylorG,WerlingD.Hostspecies-specificusageoftheTLR4-LPSreceptorcomplex.InnateImmun(2008)14:223–31.
42) Lorenzo H, Quesada O, Assunçao P, Castro A, Rodríguez F. Cytokine expression inporcinelungsexperimentallyinfectedwithMycoplasmahyopneumoniae.VetImmunolImmunopathol.2006Feb15;109(3-4):199-207.
43) JanssensS,BeyaertR.Roleoftoll-likereceptorsinpathogenrecognition.ClinMicrobiolRev(2003)16:637–46.
44) JohnsonDA.TLR4agonistsasvaccineadjuvants:achemist’sperspective.ExpertRevVaccines(2013)12:711–3.
45) Ju XH, Xu HJ, Yong YH, An LL, Jiao PR, LiaoM. Heat stress upregulation of Toll-likereceptors2/4andacuteinflammatorycytokinesinperipheralbloodmononuclearcell(PBMC) of Bama miniature pigs: an in vivo and in vitro study. Animal. 2014Sep;8(9):1462-8.
46) KanzlerH,BarratFJ,HesselEM,CoffmanRL.Therapeutictargetingofinnateimmunitywithtoll-likereceptoragonistsandantagonists.NatMed(2007)13:552–9.
47) Kaser T, Gerner W, Hammer SE, Patzl M, Saalmuller A. Phenotypic and functionalcharacterisation of porcine CD4(+)CD25(high) regulatory T cells. Vet ImmunolImmunopathol.2008;122:153–158.
48) Ketloy C, Engering A, Srichairatanakul U, Limsalakpetch A, Yongvanitchit K,Pichyangkul S, et al. Expression and function of toll-like receptors on dendritic cellsand other antigen presenting cells from non-human primates. Vet ImmunolImmunopathol(2008)125:18–30.
49) Kim Y.M., M.M. Brinkmann, M.E. Paquet, H.L. Ploegh, UNC93B1 delivers nucleotide-sensingToll-likereceptorstoendolysosomes,Nature452(2008):234–238.
50) KimSC,SticeJP,ChenL,JungJS,GuptaS,WangY,etal.Extracellularheatshockprotein

48
60,cardiacmyocytes,andapoptosis.CircRes(2009)105:1186–95.51) KimF,PhamM,LuttrellI,BannermanDD,TupperJ,ThalerJ,etal.Toll-likereceptor-4
mediatesvascularinflammationandinsulinresistanceindiet-inducedobesity.CircRes(2007)100:1589–96.
52) Kishima,M.&R.F.Ross:SuppressiveeffectofnonviableMycoplasmahyopneumoniaeon phytohemagglutinin-induced transformation of swine lymphocytes. Am J Vet Res46,2366-2368(1985).
53) KreiselD,NavaRG,LiW,ZinselmeyerBH,WangB,LaiJ,PlessR,GelmanAE,KrupnickAS, Miller MJ. In vivo two-photon imaging reveals monocyte-dependent neutrophilextravasation during pulmonary inflammation. Proc Natl Acad Sci U S A. 2010 Oct19;107(42):18073-8.
54) KumarH,KawaiT,AkiraS.Toll-likereceptorsandinnateimmunity.BiochemBiophysResCommun.2009Oct30;388(4):621-5.
55) Kurt-Jones EA, ChanM, Zhou S,Wang J, Reed G, Bronson R, ArnoldMM, Knipe DM,Finberg RW (2004) Herpes simplex virus 1 interaction with Toll-like receptor 2contributestolethalencephalitis.ProcNatlAcadSciUSA101:1315–1320.
56) Kurt-Jones EA, Popova L, Kwinn L, Haynes LM, Jones LP, Tripp RA, et al. PatternrecognitionreceptorsTLR4andCD14mediateresponsetorespiratorysyncytialvirus.NatImmunol(2000)1:398–401.
57) KuzemtsevaL,PérezM,MateuE,SegalésJ,DarwichL.ExpressionofToll-likereceptor9(TLR9)inthelungsandlymphoidtissueofpigs.VetJ.2015Feb;203(2):259-61.
58) Kwon D, Chae C. Detection and localization of Mycoplasma hyopneumoniae DNA inlungsfromnaturallyinfectedpigsbyinsituhybridizationusingadigoxigenin-labeledprobe.VetPathol.1999Jul;36(4):308-13.
59) Lemaitre, B., Nicolas, E., Michaut, L., Reichhart, J.M., Hoffmann, J. A. 1996. Thedorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potentantifungalresponseinDrosophilaadults.Cell86:973.
60) Liu,C.H.,Chaung,H.C.,Chang,H.L.,Peng,Y.T.,Chung,W.B.,2009.ExpressionofToll-likereceptor mRNA and cytokines in pigs infected with porcine reproductive andrespiratorysyndromevirus.Vet.Microbiol.136,266–275.
61) Lorenzo,H.,Quesada,O., Assuncao, P., Castro,A. andRodrıguez, F. (2006).Cytokineexpression in porcine lungs experimentally infected with Mycoplasmahyopneumoniae.VeterinaryImmunologyandImmunopathology,109,199-207.
62) Lund J, Sato A, Akira S, Medzhitov R, Iwasaki A. Toll-like receptor 9-mediatedrecognitionofherpessimplexvirus-2byplasmacytoiddendriticcells.JExpMed.2003;198(3):513–520.
63) Madec F, Kobish M. Bilan lesionnel des poumons de porcs charcutiers à l'abattoir.JournéesdelaRecherchePorcine1982,14:405-412.
64) MancusoG.,M.Gambuzza,A.Midiri,C.Biondo,S.Papasergi,S.Akira,G.Teti,C.Beninati,Bacterial recognition by TLR7 in the lysosomes of conventional dendritic cells, Nat.Immunol.10(2009)587–594.
65) MarèCJ,SwitzerWP.Viruspneumoniaofpigs:propagationandcharacterizationofacausativeagent.AmJVetRes1966;27:1687-1693.
66) MaroisC,DoryD,FabletC,MadecF,KobischM.DevelopmentofaquantitativeReal-Time TaqMan PCR assay for determination of the minimal dose of Mycoplasmahyopneumoniaestrain116requiredtoinducepneumoniainSPFpigs.JApplMicrobiol.2010May;108(5):1523-33.
67) Medzhitov R, Preston-Hurlburt P, Janeway CA Jr. A human homologue of theDrosophila Toll protein signals activation of adaptive immunity. Nature. 1997; 388:394–397.

49
68) Medzhitov R, Janeway CA Jr. Innate immunity: the virtues of a nonclonal system ofrecognition.Cell(1997)91:295–8.
69) MeyerJ,McShaneH.Thenext10yearsfortuberculosisvaccines:dowehavetherightplansinplace?ExpertRevVaccines(2013)12:443–51.
70) Miller SI, Ernst RK, Bader MW. LPS, TLR4 and infectious disease diversity. Nat RevMicrobiol(2005)3:36–46.
71) MiguelJC,ChenJ,VanAlstineWG,JohnsonRW.Expressionofinflammatorycytokinesandtoll-likereceptorsinthebrainandrespiratorytractofpigsinfectedwithporcinereproductive and respiratory syndrome virus. Vet Immunol Immunopathol (2010)135:314–9.
72) Morrison,D. F., Foss,D. L. andMurtaugh,M.P. (2000). Interleukin-10gene therapy-mediated amelioration of bacterial pneumonia. Infection and Immunity, 68, 4752e4758.
73) MunetaY,UenishiH,KikumaR,YoshiharaK,ShimojiY,YamamotoR,HamashimaN,YokomizoY,MoriY.PorcineTLR2andTLR6: identificationand their involvement inMycoplasma hyopneumoniae infection. J Interferon Cytokine Res. 2003Oct;23(10):583-90.
74) Murawski M.R., G.N. Bowen, A.M. Cerny, L.J. Anderson, L.M. Haynes, R.A. Tripp, E.A.Kurt-Jones, R.W. Finberg, Respiratory syncytial virus activates innate immunitythroughToll-likereceptor2,J.Virol.83(2009)1492–1500.
75) MurtaughM.P.Advancesinswineimmunologyhelpmovevaccinetecnologyforward.Veterinaryimmunologyandimmunopatology2014;159:202-207.
76) Noerregaard E., C.-J. Ehlorsson, A.-K. Lieber. The economic impact of an outbreak ofMycoplasmahyopneumoniae. Book ofAbstracts of 24th International PigVeterinarySocietyCongressJune7th-10th2016Dublin,Ireland.
77) OhashiK,BurkartV,FloheS,KolbH.Cuttingedge:heatshockprotein60isaputativeendogenousligandofthetoll-likereceptor-4complex.JImmunol(2000)164:558–61.
78) OkamuraY,WatariM,JerudES,YoungDW,IshizakaST,RoseJ,etal.TheextradomainAoffibronectinactivatestoll-likereceptor4.JBiolChem(2001)276:10229–33.
79) OostingM,ChengSC,BolscherJM,Vestering-StengerR,PlantingaTS,VerschuerenIC,ArtsP,GarritsenA,vanEenennaamH,SturmP,KullbergBJ,HoischenA,AdemaGJ,vanderMeer JW,NeteaMG, Joosten LA.HumanTLR10 is an anti-inflammatory pattern-recognitionreceptor.ProcNatlAcadSciUSA.2014Oct21;111(42):E4478-84.
80) Opriessnig, T., Thacker, E. L., Yu, S., Fenaux,M.,Meng, X. J. andHalbur, P. G. (2004).Experimentalreproductionofpostweaningmultisystemicwastingsyndromeinpigsbydual infection with Mycoplasma hyopneumoniae and porcine circovirus type 2.VeterinaryPathology,41,624-640.
81) Park BS, Song DH, Kim HM, Choi BS, Lee H, Lee JO. The structural basis oflipopolysacchariderecognitionbytheTLR4-MD-2complex.Nature(2009)458:1191–5.
82) Poli G. e Rocchi M., Sistemi difensivi acquisiti e specifici struttura e funzione delsistemaimmunitarioinPoliG.etal.MicrobiologiaeImmunologiaVeterinaria,secondaed.2005,UTET.
83) PoltorakA,HeX,SmirnovaI,LiuMY,VanHuffelC,DuX,etal.DefectiveLPSsignalinginC3H/HeJandC57BL/10ScCrmice:mutationsinTlr4gene.Science(1998)282:2085–8.
84) Qureshi ST, Lariviere L, Leveque G, Clermont S, Moore KJ, Gros P, et al. Endotoxin-tolerantmicehavemutationsintoll-likereceptor4(Tlr4).JExpMed(1999)189:615–25.
85) Raymond CR, Wilkie BN. Toll-like receptor, MHC II, B7 and cytokine expression by

50
porcine monocytes and monocyte-derived dendritic cells in response to microbialpathogen-associated molecular patterns. Vet Immunol Immunopathol (2005) 107:235–47.
86) ReardonS.Newlifeforpigorgan.Nature2015;257:152-154.87) Redondo E, Masot AJ, Fernández A, Gázquez A. Histopathological and
immunohistochemical findings in the lungs of pigs infected experimentally withMycoplasmahyopneumoniae.JCompPathol.2009May;140(4):260-70.
88) RehliM.Ofmiceandmen: speciesvariationsof toll-like receptorexpression.TrendsImmunol(2002)23:375–8.
89) RégiaSilvaSousaK,MauricFrossardRibeiroA,RobertoNunesGoesP,ElizaFacioniGuimarães S, Lopes PS, Veroneze R, Gasparino E. Toll-Like Receptor 6 differentialexpressionintwopiggeneticgroupsvaccinatedagainstMycoplasmahyopneumoniae.BMCProc.2011Jun3;5Suppl4:S9.
90) RocchiJ,RicciV,AlbaniM,LaniniL,AndreoliE,MaceraL,PistelloM,Ceccherini91) NelliL,BendinelliM,MaggiF.TorquetenovirusDNAdrivesproinflammatorycytokines
production and secretion by immune cells via toll-like receptor 9. Virology. 2009;394(2):235–242.
92) RoelofsMF,BoelensWC,JoostenLA,Abdollahi-RoodsazS,GeurtsJ,WunderinkLU,etal. Identificationof smallheat shockproteinB8 (HSP22)asanovelTLR4 ligandandpotential involvement in thepathogenesisof rheumatoidarthritis. J Immunol (2006)176:7021–7.
93) Rosser EC, Mauri C. Regulatory B cells: origin, phenotype, and function. Immunity.2015Apr21;42(4):607-12.
94) SaccoRE,NicholsonTL,WatersTE,BrockmeierSL.PorcineTLR3characterizationandexpressioninresponsetoinfluenzavirusandBordetellabronchiseptica.VetImmunolImmunopathol.2011Jul15;142(1-2):57-63.
95) SamuelssonC,Hausmann J, LauterbachH, SchmidtM,Akira S,WagnerH, ChaplinP,SuterM,O’KeeffeM,HochreinH.Survivaloflethalpoxvirusinfectioninmicedependson tlr9, and therapeutic vaccination provides protection. J Clin Invest. 2008;118(5):1776–1784.
96) Sang, Y., Ross, C.R., Rowland, R.R., Blecha, F., 2008a. Toll-like receptor 3 activationdecreasesporcinearterivirusinfection.ViralImmunol.21,303–313.
97) Sarradell J, Andrada M, Ramírez AS, Fernández A, Gómez-Villamandos JC, Jover A,LorenzoH,HerráezP,RodríguezF.Amorphologicandimmunohistochemicalstudyofthebronchus-associated lymphoid tissueofpigsnaturally infectedwithMycoplasmahyopneumoniae.VetPathol.2003Jul;40(4):395-404.
98) Schroder K, Irvine KM, Taylor MS, Bokil NJ, Le Cao KA, Masterman KA, et al.Conservation and divergence in toll-like receptor 4-regulated gene expression inprimary human versus mouse macrophages. Proc Natl Acad Sci U S A (2012)109:E944–53.
99) ShimosatoT,TohnoM,KitazawaH,KatohS,WatanabeK,KawaiY,AsoH,YamaguchiT,SaitoT.Toll-likereceptor9 isexpressedon follicle-associatedepitheliacontainingMcellsinswinePeyer'spatches.ImmunolLett.2005Apr15;98(1):83-9.
100) Silva-CampaE,Flores-MendozaL,ResendizM,Pinelli-SaavedraA,Mata-HaroV,MwangiW, et al. Induction of T helper 3 regulatory cells by dendritic cells infectedwithporcinereproductiveandrespiratorysyndromevirus.Virology(2009)387:373–9.
101) Sorensen NS, Skovgaard K, Heegaard PM. Porcine blood mononuclear cellcytokineresponsestoPAMPmolecules:comparisonofmRNAandproteinproduction.VetImmunolImmunopathol(2011)139:296–302.

51
102) Tabeta K, Georgel P, Janssen E, Du X, Hoebe K, Crozat K, Mudd S, Shamel L,SovathS,GoodeJ,AlexopoulouL,FlavellRA,BeutlerB(2004)Toll-likereceptors9and3asessentialcomponentsof innate immunedefenseagainstmousecytomegalovirusinfection.ProcNatlAcadSciUSA101:3516–3521.
103) Thacker,E.L.,B.J.Thacker,T.B.Boettcher&H.Jayappa:Comparisonofantibodyproduction, lymphocyte stimulation and protection induced by four commercialMycoplasmahyopneumoniaebacterins.SwineHealthProd6,107-112(1998).
104) Thacker, E.L., P.G. Halbur, R.F. Ross, R. Thanawongnuwech & B.J. Thacker:Mycoplasma hyopneumoniae potentiation of porcine reproductive and respiratorysyndromevirus-inducedpneumonia.JClinMicrobiol37,620-627(1999).
105) Thacker, E. L., Thacker, B. J. and Janke, B. H. (2001). Interaction betweenMycoplasma hyopneumoniae and swine influenza virus. Journal of ClinicalMicrobiology,39,2525-2530.
106) Thacker,E.L.,B.J.Thacker,M.Kuhn,P.A.Hawkins&W.R.Waters:Mucosalandsystemic characteristics of protective activity of a Mycoplasma hyopneumoniaebacterin.AmJVetRes61,1384-1389(2000).
107) Thacker, E. L., Thacker, B. J., Young, T. F. and Halbur, P. G. (2000). Effect ofvaccination on the potentiation of porcine reproductive and respiratory syndromevirus(PRRSV)-inducedpneumoniabyMycoplasmahyopneumoniae.Vaccine,18,1244-1252.
108) Thanawongnuwech, R., Thacker, B., Halbur, P. and Thacker, E. L. (2004).Increased production of proinflammatory cytokines following infectionwith porcinereproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae.ClinicalDiagnosticandLaboratoryImmunology,11,901-908.
109) Thanawongnuwech,R.andThacker,E.L.(2003).Interleukin-10,interleukin-12and interferon-gamma levels in the respiratory tract following MycoplasmahyopneumoniaeandPRRSVinfectioninpigs.ViralImmunology,16,357-367.
110) Thomas AV, Broers AD, Vandegaart HF, Desmecht DJ. Genomic structure,promoteranalysisandexpressionoftheporcine(Susscrofa)TLR4gene.MolImmunol(2006)43:653–9.
111) Thomsen BL, Jorsal SE, Andersen S, Willeberg P. The Cox regression modelappliedtoriskfactoranalysisofinfectionsinthebreedingandmultiplyingherdsintheDanishSPFsystem.PrevVetMed199212:287-297.
112) Tsukumo DM, Carvalho-Filho MA, Carvalheira JB, Prada PO, Hirabara SM,Schenka AA, et al. Loss-of-function mutation in toll-like receptor 4 prevents diet-inducedobesityandinsulinresistance.Diabetes(2007)56:1986–98.
113) TuPY,TsaiPC,LinYH,LiuPC,ChangHL,KuoTY,ChungWB.Expressionprofileof Toll-like receptor mRNA in pigs co-infected with porcine reproductive andrespiratory syndrome virus and porcine circovirus type 2. Res Vet Sci. 2015Feb;98:134-41.
114) Uematsu S, Jang MH, Chevrier N, Guo Z, Kumagai Y, Yamamoto M, Kato H,Sougawa N, Matsui H, Kuwata H, et al. (2006) Detection of pathogenic intestinalbacteria by Toll-like receptor 5 on intestinal CD11c(+) lamina propria cells. NatImmunol7:868–874.
115) VabulasRM,Ahmad-NejadP,daCostaC,MiethkeT,KirschningCJ,HackerH,etal. Endocytosed HSP60s use toll-like receptor 2 (TLR2) and TLR4 to activate thetoll/interleukin-1 receptor signaling pathway in innate immune cells. J Biol Chem(2001)276:31332–9.
116) Vigre,H.,Dohoo,I.R.,Stryhn,H.andBusch,M.E.(2004).Intra-unitcorrelationsin seroconversion to Actinobacillus pleuropneumoniae and Mycoplasma

52
hyopneumoniae at different levels in Danish multi-site pig production facilities.PreventiveVeterinaryMedicine,63,9-28.
117) VranckxK,MaesD,MarchioroSB,VillarrealI,ChiersK,PasmansF,HaesebrouckF.Vaccinationreducesmacrophageinfiltrationinbronchus-associatedlymphoidtissuein pigs infectedwith a highly virulentMycoplasma hyopneumoniae strain. BMC VetRes.2012Mar12;8:24.
118) WangT,TownT,AlexopoulouL,AndersonJF,FikrigE,FlavellRA(2004)Toll-like receptor 3 mediates West Nile virus entry into the brain causing lethalencephalitis.NatMed10:1366–1373.
119) WassefA, JanardhanK,Pearce JW,SinghB.Toll-likereceptor4 innormalandinflamed lungs and other organs of pig, dog and cattle. Histol Histopathol. 2004Oct;19(4):1201-8.
120) Werling,D.,Piercy,J.,Coffey,T.J.,2006.ExpressionofTOLL-likereceptors(TLR)by bovine antigen-presenting cells—Potential role in pathogen discrimination? Vet.Immunol.Immunopathol.112,2–11.
121) Williams, J., Torres-Leon, M. A. and Sansor-Nah, R. (2000). Prevalencia,extensionycaracterizacionde las lesionesenelpulmondecerdossacrificadosenelrastromunicipaldeMerida,Yucatan,Mexico.RevistaBiomedica,11,25-32.
122) YangH,WeiJ,ZhangH,LinL,ZhangW,HeS.UpregulationofToll-likereceptor(TLR)expressionandreleaseofcytokinesfromP815mastcellsbyGM-CSF.BMCCellBiol.2009May9;10:37.
123) Yang S, Sugawara S, Monodane T, Nishijima M, Adachi Y, Akashi S, et al.Micrococcusluteusteichuronicacidsactivatehumanandmurinemonocyticcells inaCD14-andtoll-likereceptor4-dependentmanner.InfectImmun(2001)69:2025–30.
124) ZhangL,Liu J,Bai J,WangX,LiY, JiangP.ComparativeexpressionofToll-likereceptorsandinflammatorycytokines inpigs infectedwithdifferentvirulentporcinereproductiveandrespiratorysyndromevirusisolates.VirolJ.2013Apr30;10:135.
125) Zhu J, Huang X, Yang Y. Innate immune response to adenoviral vectors ismediated by both toll- like receptor-dependent and -independent pathways. J Virol.2007;81(7):3170–3180.
126) ZhuY,MagnussonU,FossumC,BergM.Escherichiacoliinoculationofporcinemammaryglandsaffects localmRNAexpressionofToll-likereceptorsandregulatorycytokines.VetImmunolImmunopathol.2008Sep15;125(1-2):182-9.