aem accepts, published online ahead of print on 28...
TRANSCRIPT

REVISED VERSION
1
Mycobacterium gilvum illustrates size-correlated mycobacteria Acanthamoeba polyphaga 1
relationships 2
3
Running title: Environmental mycobacteria-amoeba relationships 4
5
Otmane Lamrabet1, Michel Drancourt1# 6
7
1 Aix Marseille Université, URMITE, UM63, CNRS 7278, IRD 198, Inserm 1095, 13005 8
Marseille, France 9
# Corresponding author: Michel Drancourt: URMITE, UMR CNRS 7278, IRD 198, INSERM 10
U1095. Faculté de Médecine, 27 Bd Jean Moulin, 13385 Marseille cedex 5 France. Tel: 33 (0)4 11
91 38 55 17 Fax: 33 (0)4 91 38 77 72 12
13
Email addresses: 14
OL: [email protected] 15
MD: [email protected] 16
17
18
19
20
21
22
23
24
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.03765-12 AEM Accepts, published online ahead of print on 28 December 2012
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
2
ABSTRACT 25
Mycobacteria are isolated from soil and water environments where free-living amoeba are living. 26
Free-living amoeba are bactericidal, yet some rapidly-growing mycobacteria are amoeba-resistant 27
organisms surviving in the amoebal trophozoites and cysts. Such capacity has not been studied 28
for the environmental rapidly-growing Mycobacterium gilvum. We investigated the ability of M. 29
gilvum to survive in the trophozoites of Acanthamoeba polyphaga strain Linc-AP1 by using optic 30
and electron microscopy and culture-based microbial enumerations in the presence of negative 31
controls. We observed that 29% of A. polyphaga cells were infected by M. gilvum mycobacteria 32
6-hour post-infection. Survived M. gilvum mycobacteria did not multiply and did not kill the 33
amoebal trophozoites during five-day co-culture. Extensive electron microscopy observation 34
indicated that M. gilvum measured 1.4 ± 0.5 µm, and failed to find M. gilvum organisms into the 35
amoebal cysts. Further experimental study of two other rapidly-growing mycobacteria 36
Mycobacterium rhodesiae and Mycobacterium thermoresistible indicated that both measured < 2 37
µm and exhibited same amoeba-mycobacteria relationships as M. gilvum. In general, we 38
observed that mycobacteria measuring < 2 µm significantly do not grow within and do not kill 39
amoebal trophozoites comparing to mycobacteria measuring > 2 µm (p < 0.05). The mechanisms 40
underlying such observation remain to be determined. 41
42
43
44
45
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
3
BACKGROUND 46
Non-tuberculous mycobacteria are environmental organisms (25, 7) found in soil (23), 47
marine environment (21) and fresh water (10, 11). They are recovered from water samples also 48
colonized by free-living amoeba (FLA) (12, 4, 1). Despite the fact that FLA are bactericidal, 49
several non-tuberculous mycobacteria were found to be amoeba-resistant, surviving within FLA 50
trophozoites and cysts (13, 18). Latter act as “Trojan horses” protecting environmental 51
mycobacteria from unfavorable conditions (22, 1, 2). 52
Amoeba-resistant mycobacteria include slowly-growing mycobacteria (SGM) such as 53
Mycobacterium avium (3) and Mycobacterium tuberculosis complex mycobacteria (27, 22); and 54
more than 25 different species of rapidly-growing mycobacteria (RGM) (1, 18). The outcome of 55
such rapidly-growing, amoeba-resistant mycobacteria depends on the mycobacterial species: 56
some Mycobacterium species such as Mycobacterium septicum survive without multiplication 57
into the trophozoites (1), while other species such as Mycobacterium smegmatis and 58
Mycobacterium chelonae multiply within the trophozoite (18, 24). Also, some mycobacteria such 59
as Mycobacterium canettii escape the FLA before encystment (22) whereas the majority of 60
Mycobacterium species survive within the amoeba cysts (22, 3). 61
Mycobacterium gilvum (formerly Mycobacterium flavescens) is an environmental 62
mycobacteria, isolated from river sediments based on its ability to degrade polycyclic aromatic 63
hydrocarbons such as pyrene, as a sole source of carbon and energy (6, 8). It is able to form 64
biofilm and its resists to ampicillin, being susceptible to other antibiotics including isoniazid (26). 65
M. gilvum has rarely been isolated as an opportunistic pathogen (26) and no study regarding M. 66
gilvum-amoeba relationships was developed yet. 67
We therefore studied the relationships between M. gilvum with the FLA Acanthamoeba 68
polyphaga trophozoites and cysts and derived features characterizing amoeba-mycobacteria 69
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
4
relationships. To validate our observations with M. gilvum we further studied two other rapidly-70
growing mycobacteria Mycobacterium rhodesiae and Mycobacterium thermoresistible. 71
72
MATERIALS AND METHODS 73
Mycobacterium and A. polyphaga strains. 74
The type strains of Mycobacterium senegalense DSM-43656T, Mycobacterium 75
conceptionense DSM-45102T, Mycobacterium rhodesiae DSM-44223T, Mycobacterium 76
thermoresistibile DSM-44167T, Mycobacterium chelonae DSM-43804T, Mycobacterium 77
smegmatis DSM-43756T, Mycobacterium abscessus DSM-44196T, Mycobacterium fortuitum 78
subsp. fortuitum DSM-46621T and M. gilvum DSM-45363T were purchased from the German 79
collection of microorganisms and cell cultures (DSMZ, Braunschweig, Germany). Mycobacteria 80
were cultured in Middlebrook 7H9 liquid medium (Sigma-Aldrich, Lyon, France) and 81
subcultured at 37°C on Middlebrook and Cohn 7H10 agar (Becton Dickinson, Le Pont de Claix, 82
France) for three days. A. polyphaga Link-AP1 trophozoites strain (19) was cultured in peptone-83
yeast extract-glucose (PYG) medium at 32°C for 3 day as described previously (22, 18). In brief, 84
A. polyphaga amoebae were suspended twice in Page’s modified Neff’s Amoeba Saline (PAS) to 85
obtain 5.105 cells/mL and 10 mL of such suspension was placed into tube Falcon® 50 mL 86
(Becton Dickinson, Le Pont de Claix, France). 87
88
Mycobacteria-amoeba coculture. 89
The liquid culture of RGM M. gilvum, M. rhodesiae and M. thermoresistible was washed 90
two times with sterile phosphate-buffer saline (PBS) and the pellet was suspended in PAS. Each 91
10 mL of the amoebal culture was inoculated with 1 mL of a 5. 107 RGM/mL suspension 92
(multiplicity of infection =1:10). As a control, A. polyphaga, M. gilvum, M. rhodesiae and M. 93
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
5
thermoresistible were cultured separately in PAS medium. After 6-h incubation at 32°C the 94
coculture was washed two times with PAS to remove any remaining extracellular or adherent 95
mycobacteria (18). After washing, the coculture was incubated in 10 mL of PAS for 5 days at 96
32°C. 97
The presence of intra-amoebal mycobacteria was determined by shaking the coculture, 10-min 98
centrifugation at 100 g and observation using a light microscope after Ziehl-Neelsen staining. In 99
addition, the presence of viable mycobacteria inside amoebal trophozoites was assessed as 100
previously described (18). In brief at 0, 24, 48, 72, 96 and 120 h post-inoculation time points, the 101
A. polyphaga monolayer was lysed with 0.1% sodium dodecyl sulfate (Sigma-Aldrich) for 30 102
min and passed through a 26-gauge needle to ensure complete lysis of the amoebae. A 100-µL 103
volume of lysate was plated onto 7H10 agar and incubated for four days at 37°C to determine the 104
number of colonies (colony-forming units, CFU) of intracellular mycobacteria. The viability of 105
amoeba, with and without bacteria, was done using Trypan Bleu coloration 0.4% (Sigma-Aldrich, 106
Taufkirchen, Germany) and counting in the Glasstic slide chamber (HycoR, Garden Grove, 107
California USA). Experiments were done in triplicate. Negative controls remained negative in 108
each experimental step. 109
110
Encystment of M. gilvum-infected amoeba. 111
Amoebae were cultured with encystement buffer as described previously (18, 22). In brief, 10 112
mL of amoebal coculture (5.105 cells/mL of PAS) were infected with 1 mL (5.107 mycobacteria/mL of 113
PAS) of Mycobacterium suspension in PAS for 6 hours. The supernatant was discarded and the 114
amoebal monolayer was rinsed twice with encystment buffer before being incubated at 32°C for three 115
days in fresh encystment buffer (18). Moreover, cysts corresponding to the time point 0 were 116
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
6
centrifuged at 1,000 g for 10 min and washed three times with PAS before electron microscopy 117
observation. Experiments were done in triplicate. 118
119
Ultrastructural studies. 120
Ultrastructural observations were done as previously described (18). In brief, amoeba 121
monolayer previously infected by M. gilvum and amoebal cysts were washed three times with 122
sterile PAS to eliminate noningested mycobacteria and fixed (18). Then, the samples were 123
successively incubated for 45 min in a 3:1, 2:2, 1:3 (vol/vol) ethanol-Epon suspension, then in 124
100% Epon overnight with continuous shaking before being embedded in an Epon 812 resin 125
(Fluka, St Quentin Fallavier, France) incubated for three days at 60°C. Ultrathin sections (70 nm) 126
were cut from the blocks using an ultracut microtome (Reichert-Leica, Marseille, France) before 127
being deposited on Formvar-coated copper grids (Sigma-Aldrich). Ultrathin sections were stained 128
for 10 min with 5% uranyl acetate and lead citrate before being examined using a transmission 129
electron microscope (Morgani 268D; Philips, Eindhoven, the Netherlands). 130
Mycobacterial size was determined: after 2-day culture in Middlebrook 7H9 medium at 131
37°C and after 2-day preculture in Middlebrook 7H9 medium followed by 2-day culture in PAS 132
medium at 37°C. The size of mycobacteria was measured by electron microscopic observation of 133
50 single mycobacteria to determine the median and standard deviation of cell length. 134
135
Statistical analyses. 136
All statistical analyses mentioned in this study were performed using the chi2-square test with a 137
significance level of p = 0.05. 138
139
RESULTS 140
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
7
Survival of mycobacteria into A. polyphaga trophozoites and cysts. 141
The number of non-infected (negative control) and infected A. polyphaga trophozoites 142
with RGM incubated into PAS at 32°C did not change significantly over the time of the 143
experiment. After six hours co-culture, 29% of A. polyphaga cells were found to be infected by 144
M. gilvum mycobacteria as confirmed by Zielh-Neelsen staining. The number of M. gilvum per 145
trophozoite varied from 1 to 46 (a mean of 17 ± 14 mycobacteria/trophozoite). Electron 146
microscopy revealed mycobacteria into vacuoles surrounded by several mitochondria (Fig. 1A). 147
We observed that the three tested RGM species survived but did not multiply over five-day co-148
cultured with amoeba at 32°C (Fig. 2). No significant difference was observed in the number of 149
mycobacteria with time (p = 0.1). Mycobacteria survived in PAS yet M. gilvum, M. rhodesiae 150
and M. thermoresistible CFUs did not increase from day 0 to day 5 (negative control) (Fig. 2). 151
Electron microscopy revealed pre-cysts and mature-cyst after 3-day co-culture. Careful electron 152
microscopy observation of 300 cysts formed at that time failed to reveal any M. gilvum organism 153
into A. polyphaga cysts (Fig. S1). 154
155
RGM size measurement 156
When it was not available in literature, we measured the length of RGM cells in two 157
different culture conditions (see materials and methods) using electron microscopy observation. 158
In this study, after culture in 7H9 medium the length measured was 2.1 ± 0.7 µm for M. 159
abscessus, 2.3 ± 0.3 for M. chelonae, 2.6 ± 0.1 for M. smegmatis, 2.5 ± 0.5 µm for M. 160
conceptionense, 1.7 ± 0.1 µm for M. fortuitum subsp. fortuitum, 1.9 ± 0.2 µm for M. senegalense, 161
1.3 ± 0.4 µm for M. rhodesiae, 1.1 ± 0.3 µm for M. thermoresistibile and 1.4 ± 0.5 µm for M. 162
gilvum (Fig. 3). After culture in 7H9 medium the length measured was 2.2 ± 0.5 µm for M. 163
abscessus, 2.1 ± 0.5 µm for M. chelonae, 2.8 ± 0.9 µm for M. smegmatis, 2.3 ± 0.9 µm for M. 164
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
8
conceptionense, 1.8 ± 0.4 µm for M. fortuitum subsp. fortuitum, 1.8 ± 0.5 µm for M. senegalense, 165
1.6 ± 0.3 µm for M. rhodesiae, 1.4 ± 0.3 µm for M. thermoresistibile and 1.3 ± 0.2 µm for M. 166
gilvum (Fig. S2). We observed that whatever the medium (7H9 or PAS) there is no significant 167
difference between mycobacteria size for all studied strains (p > 0.05). The size of mycobacteria 168
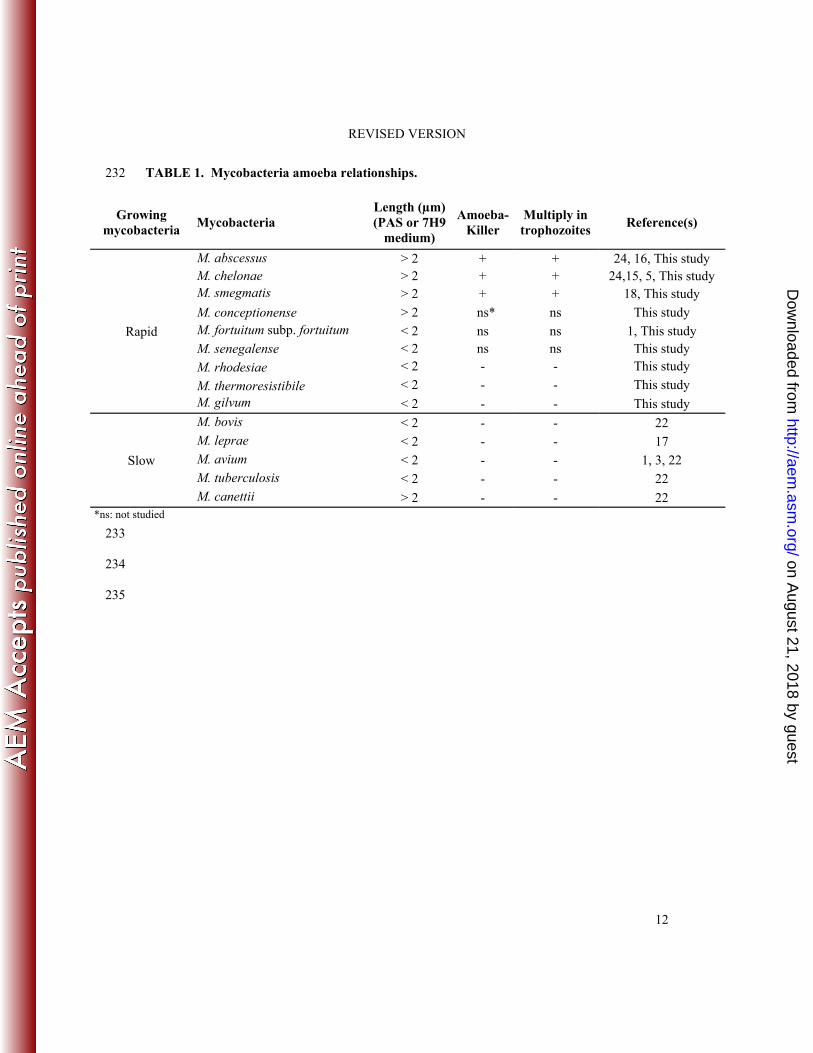
significantly correlated with intraamoebal growth and amoeba killing with mycobacteria 169
measuring < 2 µm are not growing within and are not killing amoebal trophozoites and 170
mycobacteria measuring > 2 µm being growing within and killing amoebal trophozoites (p < 171
0.05) (Table 1). 172
173
DISCUSSION 174
A. polyphaga and Acanthamoeba castellanii are two FLA routinely used to probe 175
mycobacteria-FLA interactions and A. polyphaga was used in this study (9, 20, 29). We observed 176
that rapidly-growing mycobacteria M. gilvum, M. rhodesiae and M. thermoresistibile penetrated 177
into A. polyphaga trophozoites, a reproducible result obtained by using a low (1:10) multiplicity 178
of infection. Previous studies have showed that the majority of RGM penetrated into amoebal 179
trophozoites (1, 18) but our observation that M. gilvum, M. rhodesiae and M. thermoresistibile 180
could also be ingested by amoebal trophozoites has not been previously reported. We further 181
observed that such intra-amoebal mycobacteria survived into the A. polyphaga trophozoites, a 182
fact documented by both microscopic observations and microbial enumerations. This observation 183
agrees with previous demonstrations of intra-amoebal surviving of Mycobacterium septicum, 184
Mycobacterium abscessus (1) and M. smegmatis in A. castellanii (14, 28) and A. polyphaga (18). 185
Furthermore, M. gilvum mycobacteria were observed into vacuoles, as previously observed for 186
other RGM as M. septicum, Mycobacterium mucogenicum, Mycobacterium massiliense and M. 187
smegmatis in A. polyphaga (1, 18). 188
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
9
M. gilvum, M. rhodesiae and M. thermoresistibile mycobacteria did not multiply within amoeba 189
and did not kill the amoeba during the time of the experiment (five-day coculture). This is 190
contrary to other RGM such as M. abscessus, M. chelonae, M. smegmatis, Mycobacterium 191
monacens and Mycobacterium neoaurum which all multiply within trophozoites and kill the 192
amoeba after five-day co-culture (24, 18). These data indicate that not all the RGM are amoebal 193
killers suggesting that factors other than the rapid growth may be involved in the mycobacteria-194
amoeba interactions. Accordingly, we here observed that intraamoebal multiplication and amoeba 195
killing significantly correlated with the size of mycobacteria. Indeed, among mycobacteria 196
belonging to the same species, organisms exhibiting a > 2 µm size do behave differently that the 197
ones that exhibit a < 2 µm size; larger RGM species measuring more than 2 µm penetrate, 198
multiply and kill the amoeba, contrary to smaller RGM such as M. gilvum, M. rhodesiae and M. 199
thermoresistibile measuring less than 2 µm which do not kill amoeba (this study). This puzzling 200
observation could be extended to SGM. We observed that the majority of studied SGM 201
measuring less than 2 µm and do not kill amoeba; the notable exception being M. canettii, a 202
species measuring more than 2 µm (22) which does not kill, but instead escapes out of amoeba. 203
Overall, our data indicate significant correlation between the median size of mycobacteria 204
and outcome of mycobacteria into amoeba. This observation warrants further investigations to 205
understand whether the size of mycobacteria triggers intra-amoebal outcome by itself, or whether 206
size is just a proxy for biological propriety of mycobacteria, which is not studied already. Data 207
herein presented suggest that replication rate is not the biological factor. Also, we observed no 208
correlation between genome size of studied mycobacteria and intra-amoebal survival (data not 209
shown), pending to additional genomic studies. 210
Extensive electron microscope observation failed to reveal any M. gilvum organisms in 211
cysts. This observation extends previous observations made for other RGM such M. smegmatis 212
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
10
(18). Combining morphological and cultural data indicates that the majority of RGM bypass the 213
amoebal cyst after they are phagocyted into the amoebal trophozoites. These data agree with the 214
previous observations that all M. canettii organisms and the majority of M. tuberculosis 215
organisms and non-tuberculous organisms such as M. smegmatis (18) escape from the A. 216
polyphaga pre-cyst before its maturation contrary to M. avium organisms (3). 217
In conclusion, the substitute characteristics of RGM-amoeba interactions may be wider 218
than previously reported. It may partly relied on the size of the RGM species and is comprising of 219
(i) RGM species smaller than 2 µm, surviving in amoebal trophozoites but not in the cysts such 220
as M. septicum (1) and M. gilvum (present work); (ii) RGM longer than 2 µm, surviving into 221
trophozoites and cysts such as M. fortuitum and M. abscessus (1, 24; 16); and (iii) RGM longer 222
than 2 µm which penetrate, multiply and kill the amoeba such as M. chelonae (24) and M. 223
smegmatis (18) (Fig. 4). 224
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
11
ACKNOWLEDGMENTS 225
The authors acknowledge Audrey Borg and Audrey Averna for their technical help with the 226
electron microscopy observations. 227
228
COMPETING INTERESTS 229
The authors declare that they have no competing interests. 230
231
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from
REVISED VERSION
12
TABLE 1. Mycobacteria amoeba relationships. 232
Growing mycobacteria
Mycobacteria Length (µm) (PAS or 7H9
medium)
Amoeba-Killer
Multiply in trophozoites
Reference(s)
Rapid
M. abscessus > 2 + + 24, 16, This study M. chelonae > 2 + + 24,15, 5, This study M. smegmatis > 2 + + 18, This study
M. conceptionense > 2 ns* ns This study M. fortuitum subp. fortuitum < 2 ns ns 1, This study M. senegalense < 2 ns ns This study
M. rhodesiae < 2 - - This study
M. thermoresistibile < 2 - - This study M. gilvum < 2 - - This study
Slow
M. bovis < 2 - - 22 M. leprae < 2 - - 17 M. avium < 2 - - 1, 3, 22 M. tuberculosis < 2 - - 22 M. canettii > 2 - - 22
*ns: not studied 233
234
235
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
13
Figure legends: 236
Fig. 1. M. gilvum mycobacteria are internalized into amoeba. Transmission electron-237
microscopy observation of M. gilvum (►) co-cultivated with A. polyphaga trophozoites at (A) 0 238
hour (B) 72 hours. m: mitochondria. Scale bar: 2 µm (A, B). 239
Fig. 2. Growth of RGM within A. polyphaga trophozoites. M. gilvum (A), M. rhodesiae (B) 240
and M. thermoresistibile (C) co-cultured with free-living amoeba A. polyphaga (black bar), 241
cultivated in PAS medium (grey bar) and cultivated in 7H9 complete medium (white bar). Each 242
bar represents the mean of triplicate experiments. Standard errors are represented by error bars. 243
Fig. 3. Mycobacterial size in 7H9 medium. Size of M. gilvum (A), M. senegalense (B), M. 244
conceptionense (C), M. rhodesiae (D), M. thermoresistibile (E), M. chelonae (F), M. smegmatis 245
(G), M. abscessus (H) and M. fortuitum subsp. fortuitum (I) by electron microscopy. The size was 246
measured in the same cultures conditions for all mycobacteria (see materials and methods). Scale 247
bar: 500 nm (A, B, C, E, F, G, H), 300 nm (D) and 1µm (I). 248
Fig. 4. The spectrum of RGM-amoeba interactions. 249
250
251
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
14
Supplemental material legends: 252
Fig. S1. Transmission electron-microscopy observation of A. polyphaga cysts. M. gilvum 253
present in the outside of a mature form of cyst. Scale bar: 5 µm. Bacteria are represented black 254
triangle. 255
Fig. S2. Mycobacterial size in PAS medium. Size of M. gilvum (A), M. senegalense (B), M. 256
conceptionense (C), M. rhodesiae (D), M. thermoresistibile (E), M. chelonae (F), M. smegmatis 257
(G), M. abscessus (H) and M. fortuitum subsp. fortuitum (I) by electron microscopy. The size was 258
measured in the same cultures conditions for all mycobacteria (see materials and methods). Scale 259
bar: 500 nm (A, C) and 1 µm (B, D, E, F, G, H, I). 260
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
15
REFERENCES 261
1. Adekambi T, Ben Salah S, Khlif M, Raoult D, Drancourt M. 2006. Survival of 262
environmental mycobacteria in Acanthamoeba polyphaga. Appl. Environ. Microbiol. 263
72:5974–5981. 264
2. Barker J, Brown MR. 1994. Trojan horses of the microbial world: protozoa and the 265
survival of bacterial pathogens in the environment. Microbiology 140:1253-1259. 266
3. Ben Salah I, Drancourt M. 2010. Surviving within the amoebal exocyst: the 267
Mycobacterium avium complex paradigm. BMC Microbiol. 10:99. 268
4. Ben Salah I, Ghigo E, Drancourt M. 2009. Free-living amoeba, a training field for 269
macrophage resistance of mycobacteria. Clin. Microbiol. Infect. 15:894–905. 270
5. Bergey DH, Harrison FC, Breed RS, Hammer BW and Huntoon FM. 1923. pp. 1–271
442. In Bergey's Manual of Determinative Bacteriology, 1st ed. The Williams & Wilkins 272
Company, Baltimore. 273
6. Brezna B, Khan AA, Cerniglia CE. 2003. Molecular characterization of dioxygenases 274
from polycyclic aromatic hydrocarbon-degrading Mycobacterium spp. FEMS Microbiol. 275
Lett. 223:177-183. 276
7. Chilima BZ, Clark IM, Floyd S, Fine PE, Hirsch PR. 2006. Distribution of 277
environmental mycobacteria in Karonga District, northern Malawi. Appl. Environ. 278
Microbiol. 72:2343-2350. 279
8. Dean-Ross D, Cerniglia CE. 1996. Degradation of pyrene by Mycobacterium flavescens. 280
Appl. Microbiol. Biotechnol. 46:307-312. 281
9. Douesnard-Malo F, Daigle F. 2011. Increased persistence of Salmonella enterica 282
serovar typhi in the presence of Acanthamoeba castellanii. Appl. Environ. Microbiol. 283
77:7640–7646. 284
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
16
10. Ettinger MR, Webb SR, Harris SA, McIninch SP, Garman CG and Brown BL. 285
2003. Distribution of free-living amoebae in James River, Virginia, USA. Parasitol. Res. 286
89:6–15. 287
11. Falkinham JO, Norton CD, LeChevallier MW. 2006. Factors influencing numbers of 288
Mycobacterium avium, Mycobacterium intracellulare, and other mycobacteria in drinking 289
water distribution systems. Appl. Environ. Microbiol. 67:1225-1231. 290
12. Greub G, La Scola B, Raoult D. 2004. Amoebae-resisting bacteria isolated from human 291
nasal swabs by amoebal coculture. Emerg. Infect. Dis. 10:470–477. 292
13. Greub G, Raoult D. 2004. Microorganisms resistant to free-living amoebae. Clin. 293
Microbiol. Rev. 17:413-433. 294
14. Krishna Prasad BN, Gupta SK. 1978. Preliminary report on the engulfment and 295
retention of mycobacteria by trophozoites of axenically grown Acanthamoeba castellanii 296
Douglas, 1930. Current Science. 47:245–247. 297
15. Kubica GP, Baess I, Gordon GE, Jenkins A, Kwapinski JBG, McDurmont C et al. 298
1972. A cooperative analysis of rapidly growing mycobacteria. J. Gen. Microbiol. 73:55-299
70. 300
16. Kusunoki S, Ezaki T. 1992. Proposal of Mycobacterium peregrinum sp. nov., nom. rev., 301
and elevation of Mycobacterium chelonae subsp. abscessus (Kubica et al.) to species 302
status: Mycobacterium abscessus comb. nov. Int. J. Syst. Bacteriol. 42:240-245. 303
17. Lahiri R, Krahenbuhl JL. 2008. The role of free-living pathogenic amoeba in the 304
transmission of leprosy: a proof of principle. Lepr. Rev. 79:401–409. 305
18. Lamrabet O, Mba Medie F, Drancourt M. 2012. Acanthamoeba polyphaga-enhanced 306
growth of Mycobacterium smegmatis. PLoS One. 7:e29833. 307
19. La Scola B, Mezi L, Weiller PJ, Raoult D. 2001. Isolation of Legionella anisa using an 308
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
17
amoebic coculture procedure. J. Clin. Microbiol. 39:365-366. 309
20. Laskowski-Arce MA & Orth K. 2008. Acanthamoeba castellanii promotes the survival 310
of Vibrio parahaemolyticus. Appl. Environ. Microbiol. 74:7283-7288. 311
21. Liu H, Ha YR, Lee ST, Hong YC, Kong HH and Chung DI. 2006. Genetic diversity of 312
Acanthamoeba isolated from ocean sediments. Korean J. Parasitol. 44:117–125. 313
22. Mba Medie F, Ben Salah I, Henrissat B, Raoult R, Drancourt M. 2011. 314
Mycobacterium tuberculosis complex mycobacteria as amoeba-resistant organisms. PLoS 315
One. 6:e20499. 316
23. Narang R, Narang P, Mendiratta DK. 2009. Isolation and identification of 317
nontuberculous mycobacteria from water and soil in central India. Indian J. Med. 318
Microbiol. 27:247–250. 319
24. Pagnier I, Raoult D, La Scola B. 2008. Isolation and identification of amoeba-resisting 320
bacteria from water in human environment by using an Acanthamoeba polyphaga co-321
culture procedure. Environ. Microbiol. 10:1135–1144. 322
25. Primm TP, Lucero CA, Falkinham JO. 2004. Health impacts of environmental 323
mycobacteria. Clin. Microbiol. Rev. 17:98-106. 324
26. Stanford JL, Gunthorpe WJ. 1971. A study of some fast-growing scotochromogenic 325
mycobacteria including species descriptions of Mycobacterium gilvum (new species) and 326
Mycobacterium duvalii (new species). Br. J. Exp. Pathol. 52:627-637. 327
27. Taylor SJ, Ahonen LJ, de Leij FA, Dale JW. 2003. Infection of Acanthamoeba 328
castellanii with Mycobacterium bovis and M. bovis BCG and survival of M. bovis within 329
the amoebae. Appl. Environ. Microbiol. 69:4316–4319. 330
28. Tenant R, Bermudez LE. 2006. Mycobacterium avium genes upregulated upon infection 331
of Acanthamoeba castellanii demonstrate a common response to the intracellular 332
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from

REVISED VERSION
18
environment. Curr. Microbiol. 52:128–133. 333
29. Thomas V, McDonnell G, Denyer SP, Maillard JY. 2009. Free-living amoebae and 334
their intracellular pathogenic microorganisms: risks for water quality. FEMS Microbiol. 335
Rev. 34:231–259. 336
337
on August 21, 2018 by guest
http://aem.asm
.org/D
ownloaded from



