advances in marine biology, volume 1
TRANSCRIPT


Advances in
MARINE BIOLOGY VOLUME 1

This Page Intentionally Left Blank

Edited by
F. S. RUSSELL Plymouth, England
Advances in
MARINE BIOLOGY
VOLUME 1
ACADEMIC PRESS, INC. (Harcourt Brace Jovanovich. Publishers)
London Orlando San Diego New York Toronto Montreal Sydney Tokyo

ACADEMIC PRESS INC. (LONDON) LTD.
BERKELEY SQUARE HOUSE
LONDON, W.1
US. E d i t h , published by
ACADEMIC PRESS, INC. Orlando, Florida 32887
Copyright 0 1963 hy Academic Press Inc. (London) Ltd.
All rights reserved
NO PART OF THIS BOOK MAY BE REPRODUCED IN ANY FORM, BY PHOTOSTAT,
MICROFILM OR ANY OTHER MEANS, WITHOUT WRITTEN PERMISSION FROM THE
PUBLISHER
Library oj' Chyri?ss Cutu1o.q Cwd Nwmbw: 63-14040
SBN: 12 026101 4
PRINTED IN THE UNITED STATES OF AMERICA
8 4 8 5 8 6 8 7 9 8 7 6 5 4

CONTRIBUTORS TO VOLUME I
J. H . 8. BLAXTER, Marine Lizborw, Aber&een, Scotland
ANTON F. BRTJUN, formerly of Copenhagen, Denmark
b y C. DAVIS, U.S. Bureau of C m w c i a l Fisheries Biological Laboratmy, Iliilfot.d, Connecticut, U.S.A.
F. G. T. HOLLIDAY, Department of Natural History, Aberdeen Univereity, 8cfAHul
VIUTOR L. LOOSANOFF, U.S. Bureau of Commercial Fi8heries, She& $aheries Laboratmy, Tiburon, California, U.S.A.
J. A. C . NICOL, Marine Biological A88ociation, The Laboralory, Citadel Hill, Plymouth, Demn, England
C. M. YONOE, Department of Zoology, The University, M a g o w , Scotland

This Page Intentionally Left Blank

PREFACE
The very great oxpansion of marine rcsearch in recont years haa resulted in a mass of published results scattered through a very wide range of periodicals. In consequence i t is becoming increasingly diffi- cult to obtain a general picture of the overall advance that is being made in our knowledge of the many aspects of life in the sea.
It is hoped that the production of this new serial publication will help biologists to keep abreast of knowledge in the different lines of research on the biology of marine organisms. It is intended that each annual volume shall contain comprehensive review articles summarizing the general position of our knowledge in individual fields. Attention will be given to recent advances in fisheries biology, the results of research in which are often published in periodicals that may not normally be available in the librarics of univorsity biology depart- ments. These investigations are, however, of vcry goneral interest since they usually concentrate on the biology and ecology of a few individual specics in greater detail than for other marino organisms.
When possible shorter roview articlcs may also be publishoti drawing attention to new dovelopmcnts and growing points in marino biology.
General articlcs on the biology of marine organisms will include information on the environment only in so far as it is nccessary for an understanding of their habits. Articles will not be published which relate only to the physical and chemical conditions in tho flea in relation to water movcmcntn and doop-sea occanoqraphy.
Any suggestions from readers on fields of resoarch that nccd reviewing and might form subject matter for future volumes will be welcomod. IMitorial corrcspondeiice should be addressed to me at Wardour, Derriford, Crownhill, Plymouth, Devon.
April, 1963 F. S . R.

This Page Intentionally Left Blank

CONTENTS
CONTRIBUTORS TO VOLUME 1 . . .. .. .. .. PREFACE . . .. .. .. .. .. .. ..
Rearing of Bivalve Mollusks
A. B. C. D. E. F. G. H. I. J. K. 1;.
M.
VICTOR L. LOOSANOFF AND HARRY C. DAVIS I. Introduction .. .. .. * . .. ..
11. Equipment . . .. .. . f .. .. .. 111. Conditioning Mollusks for Out-of-Seaon Spawning . . IV. Cultivation of Eggs and Larvae of Bivalve8 . . ..
Abnormal Eggs and Larvao . . .. . . General Description of the Development
Methods of Cultivation of Eggs and Larvao
. .
. . Larval Period . . .. 1 . .. .. Hardiness of Eggs and Larvae . . .. .. Effects of Temperature on Eggs and Larvae . . Effects of Salinity on Eggs and Larvae.. Effects of Turbidity on Eggs and Larvae . . Effects of Foods on Growth of Larvae . . Effeects of Crowding . . .. .. ..
..
.. -
Metamorphosis . . .. .. .. .. Diseases of Larval and Juvenile Mollusks and their
..
..
..
..
.. I .
..
..
. .
..
..
..
..
..
..
..
.. Treatment . . .. .. .. ..
Selective Breeding and Hybridization . . .. V. Rearing of Different Species . . .. .. ..
A. Crassoetrea virginicu (Gmelin) . . .. .. C. Arm tralzgveraa Say . . .. .. . . D. Modiolzcp demis8us (Uillwyn) . . .. . . E. Mylilwedulie Linn6 . . . I .. .. F. Anomia eimplex 1YOrt)igny . . . . * .
G . Pecten irrudium Larnarck . . .. . . H. O&eu vdulia Idinn/, . . .. .. .. I . Odrm lurida Carperrbr . . .. .. .. , I . f !rtr,rwdrctc vigm (Thunberg) . . .. .. K. Laevicurdium mortoni (Conrad) . . .. .. L. Mercenaria (= Venw) campechiensia (Gmelin) M. Tapm mnddecusmiu Reeve . . .. ..
B. Mercenaria (= Venua) mermnrlrio (Linnb) . .
11
. .
..
..
. .
..
. .
. .
. .
. .
. .
..
..
..
..
..
..
PAOL
V
vii
2 4
14
26 26 -30 35 38 41 47 52 53 55 68 71
76 80 81 82 84 HB H7 1)o ! j f i
101 104 106 107 109 110
m

X CONTENTS
N. Pitar (= Callocardia) morrhmna Gould .. 0. Petricola pholadiformis hmarck .. .. P. Emis directus (Conrad) . . .. .. . . Q. Mactra (= Spisula) solidissima Dillwyn . . R. N y a arenaria Linn6 . . .. .. .. S . Teredo navalis Linnd . . .. .. ..
VI. Acknowledgments .. . . .. .. .. VII. References . . . . .. . . 1 . .. ..
The Breeding of the North Atlantic Freshwater- Eels
The late ANTON F. BRUTJN I. Introduction . . . . . . .. .. ..
PAGE
.. 112
.. 115
.. 117
.. 120
.. 123
.. 127
. . 129
.. 130
.. 137 11. Anguillu anguilla L.-A. rostrata Le Sueur, the Taxonomic
Situation . . .. .. . . .. . . . . . . 139 I11 The Distance to Cover during the Migration . . .. .. 141 IV. How Does A . anguilla Reach the Breeding Place? . . .. 142
Why Have No Migrating Eels Been Caught in the Strait of Gibraltar?. . .. .. . . .. . . . . . . 145
VI. The Return of the European Eel to the Sea . . .. . . 147 VII. Schmidt’s CoIlections of Anguillu Leptocephali. . . . . . 164
Possible Temperature Effects on the Number of Vertebrae in
V.
VIII. Anguilla . . .. .. .. .. .. . I
IX. Parallel Cases among North AtlaRtic Apodea . . . . X. Other Specie8 of Apodert BrwJiny in the H~ganno Heti
XI. Reference8 . . . . . . . . . . . . . I
Some Aspects of Photoreception and Vision in Fishes
q J . A. (!. NICO~, . . . . . . .. . . I. Introduction . .
11. Extra-ocular Reception . . . . . . . . . . 111. Kegalntion of Light Itenching SScnnory Surfaces ..
.. . . A. I’ineul . . .. . . ..
C. The Tapetum Lucidum of Chondrichthyes . . ,. * . B. Pupillary Movement . . ..
I). Retinomotor Changea in Teleosts . . . .
.. 156
.. 162
. I la4 . I ICH)
. . 171
. . 172
I . 174 . . 174 .. 176 .. 176 .. 178

CONTENTS
Refraction, Accommodation and the Receptor Layer . . Visuul Pigments and Spectral Sensitivity . . . . . . Transmiesion.of the Lens . . .. .. . . .. .. Photosensitivity and Visual Thrcxholde . . .. I . . . Thc Chorioitlnl Gland . . .. .. . . . . . . Nnologicnl cind Bchavinnral StudieH . . .. .. . . Synopsix . . .. .. .. .. .. .. . .
..
References . . . . .. .. .. . * .. ..
IV. V.
VI . VII.
VITT. IX . X.
XI.
I. 11.
111. IV. V.
VI. VII.
VIII. IX .
X. X I .
XII. XIII.
I.
11.
The Biology of Coral Reefs C. M. YONGE
Introduction .. .. .. Reviews . . .. .. .. Systematics and Distribution . . Settlement of Planulae . . .. Ecology of Atolls . . .. .. Atlantic Reefs . . .. .. Eroeion . . .. .. .. Physiology . . ,. .. .. Zooxunthellae . . .. ..
A. Nature .. .. .. R. Significance of the hociut ion
Growth . . I . . . . . lm(!t I d I, iKilt . . . . . . Productivity .. . . . . Itcferciiccs . . 9 . . . ..
.. .. .. ..
.. .. .. ..
.. .. .. ..
.. .. .. ..
.. .. .. ..
.. .. . . . .
.. .. .. . .
.. .. .. . .
.. .. .. . .
. . . . . . . .
.. .. .. . .
. . . . . . . .
. . . . . . . . * . .. . . . . . * .. ..
The Behaviour and Physiology of Herring and Other Clupeids J. H. S. BLAXTER AND F. G . T. HOLLIDAY
.. .. .. .. * . Introduction .. .. A. General .. .. .. .. .. .. . . B. Characteristics of Clupeids . . a . . . . .
A. Baxic Structure and Componition . . .. 13. To1er:mcc to External Coriditioiili . .
. . . . . . The Gametes .. .. . . ..
xi p.im
182
186 101
191 104
196
200 20 I
209
213 214 217
219 224
229 232 23 2 232 236 246 21H
260
266
262 262 264 2fM 264 2t;f;

xii CONTENTS
C. Viability of the Gametes and Artificial Storage . . D. Fertilization .. E. Parthenogenesis . . .. .. ..
111. The Developing Egg . . .. .. .. A. Embryology . . . . .. ..
ing Egg . . . . .. . .
.. .. .. .. * . .. .. .. .. .. ..
P. Effect of Temperature on Rate of Development . . C. Salinity Tolerance and Osmo-regulation of the Develop
D. Effect of Temperature, Pressure and CO, on Egg8 . . . . .. ..
. . . . .. .. E. Egg Mortality .. . . .. .. .. . . IV. TheLarva .. .. ..
A. Development of Organ Systems.. .. .. B. Feeding of Larvae.. .. . . . . .. C. Growth of Larvae.. .. .. . . . . D. Rearing of Larvae. . .. .. . . . . E. Farming . . .. .. F. Mortality of Larvae . . .. .. .. G. Predation on Larvae . . .. .. .. H. Salinity Tolerance and Osmo-regulation of Larvae I. Oxygen Uptake . . .. J. Dermal Receptors. . .. .. .. ..
Limiting Factors * .
. . .. . .
.. .. ..
K. Temperature, pH, Oxygen, Pressure and Light
L. Loeomotory Behaviour and Rheotropic Response .. .. ..
M. Vertical Migration of Larvae . . .. .. N. Response of Larvae to Light . . * . ..
.. . . .. .. V. Metamorphosis . . . I
VI. Post-metamorphic Stages . . . . . . . . . . A. 13. (;. I ) . 12. It'.
G . H. I. J. K. L. M.
N. 0. P.
PAQE
267 267 268 268 268 269
27 1 272 273
Siilinity Tolermce and OHmcJ-regulution Temperature, Oxygen, C02 atid H,Y n~ f,imitirig IhtorH The B r h . . . . . . .. . . . . . . Vision . . . . . . . . . . . . . . Olfaction . . . . . . .. .. .. .. The Labyrinth, Hearing, the Effect of Sound, and
Sound Production . . . . .. .. .. Buoyancy and Equilibrium . . .. .. .. Swimming . . .. . . .. I . . . . . Activity . . .. .. . . . . . . ..
. . 312 313 314 316
316 320 322 325
. . 274
. . 274 ,. 276 .. 277 .. 278 . . 280 . . 280 . . 283 .. 283 .. 286 .. 287 as .. 287 . . 289 . . 289 . . 291 . . 292 . . 294
294 . , Wfi . . 297 . . 2 w . . :so2 . . 3OG . . 307 . . 3(H)

CONTENTS
.. .. .. .. Q. Shoaling .. .. .. R. Migrations .. .. S. Vertical Migration. . .. T. Effect of the Moon .. .. .. .. .. U. Attraction to Artificial Lights . . .. .. .. V. Reaction to Nets and Other Obstacles . . .. .. W. Learning .. .. .. .. .. .. .. X. Maturation of the Gonads .. .. .. .. Y. Spawning .. .. .. .. .. .. .. Z. Racial Characters, tho Genotype and tho Environment
VII. Conclusions . . .. .. .. .. .. .. ..
.. .. .. .. .. .. .. .. ..
.. .. .. .. .. VIII. References . . .. ..
xiii
326 332 342 347 348 361 366 366 364 367 370 372
P A W
AUTHORINDEX .. .. .. .. .. .. .. . . 396
SUBJECT INDEX . . .. .. .. .. .. .. .. 406

This Page Intentionally Left Blank

REARING OF BIVALVE MOLLUSKS
VICTOR L. LOOSANOFF* AND HARRY C. DAVIS U .S. Bureau of Commercial Fisheries Biological Laboratory,
Milford, Connecticut
I. Introduction . . . . * . .. .. . . .. .. .. 11. Equipment .. .. .. . . .. . . .. .. ..
1x1. Conditioning Mollusks for Out-of-Season Spawning . . .. .. .. IV. Cultivation of Eggs and Larvae of Bivalves +. A . . . I .. ..
-4. General Description of the Development . . .. .. .. R. Abnormal Eggs and 1,arvae . . . . .. .. .. .. C. Methods of Cultivation of Eggs and Larvae . . * . . . D. 1,arval Period . . . . . . . . . . . . . . .. E. Hardinees of Eggs and Larvae . . . . . . . . . . F. NHocts of Temperatiire on Eggs and Larvae . . . . . . G. E:ffects of Salinity 0 1 1 Eggs and Larveo . . . . . . .. H. Effects of Turbidity on Eggs and Larvae . . . . . . .. I. Effects of Foods on Growth of Larvae . . .. .. .. J. Effects of Crowding . . . . . . .. .. .. .. K. Metamorphosis .. . . . . .. . . .. . . L. Dimasos of Larval arid Juvenile Mollusks and their Treatment . . M. Sdectivo Brooding arid Hyhridizatioii . .
V. ltmririg of J)ifferent Spociew . . .. . . . . A. B. C. D. E. F. G. H. I. J. K. L. M. N. 0. P. Q. R. S.
Gruuaoatrea virginim (Grnelin) . . . .
Arm tmnaveraa Say . . .. .. . . ModWlu.4 demiasuo (Dillwyn) . . .. .. Mytilus edulia Linn6 . . .. . . .. A m i a mmpkx DOrbigny . . .. .. Pcctcn irradiana Lamarck . . .. . . Oatreu edulis Linn6 . . .. .. . . Oatmu lurkah Carpenter . . .. .. Cmasoatrea gigaa (Thunberg) . . . . .. Luevieardium mortoni (Conrad) *. -. . ..
T a p aemideeueaata Reeve . . . . . .
Petriwlu p h a l d i f m b Lamarck . . . Emia directw ( ( h r e c l ) . . . . Mrrclrrr ( . = Npinula) aoluhnnarn4i I)illwyrt M?/cr rcrenurirr l.irin6 . . . . . . . . lsrrtlo i u i w l i n Liiinh . .
Merwnanb (= Venw) mcmenuriu (LinnB) . .
Memenuria (= Venus) campechiensie (Grnelin)
Pitar (= CaUoeardia) mmrhwnu Gould . .
. .
. . . . . . I ,
..
. .
..
..
..
..
. .
. .
. .
..
..
. .
..
..
..
. .
. .
. .
. .
. .
..
..
..
..
. .
..
..
..
..
..
..
..
..
..
..
..
..
..
. .
. .
. .
. .
. .
..
.. * .
..
. .
..
..
..
..
..
..
..
..
..
..
..
..
.,
. .
. .
. .
. ,
..
. .
..
2 4
14 26 26 30 36 38 41 47 52 53 66 68 71 76 80 81 82 84 86 87 90 96 98
101 104 106 107 109 110 112 116 I I7 120
127 129 130
I . -....... I , ....... ... .*.... I.. . . . . . . . . .. .. . . 1*I
V I. A ~ ~ l l ~ i ~ ~ ~ l i ~ i ~ ~ i i ~ ~ ~ l ~ i ~ .. .. . . . - .. .. .. .. 129 Vl1. Iiofmwircw. . . . . . .. . . .. . . .. * . .. 130
Y.11. I 11

2 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
I. INTRODUCTION Until recently rearing of larvae and juveniles of marine bivalves,
on a basis where repeatable results could be expected, was virtually impossible because of the lack of satisfactory, reliable methods. Thus, although culturing of larvae of bivalves was first attempted in the last century, few workers succeeded in rearing them to metamorphosis and, as a rule, they were rarely grown beyond early straight-hinge stage. Even though, in the twenties, Wells (1927) was able to rear the American oyster, Cramoetrea virginica, from artificially-fertilized eggs to epat, and Prytherch (1924) raised larvae of the same species in large numbers, their results could not be consistently repeated by other investigators. The failures were usually due to poor culture methods and want of good food for the larvae, especially when they were grown in heavy concentrations. It is also possible that diseases, including those caused by fungi, were responsible for the persistent failures.
Attempts to rear larvae of bivalves were not confined, of course, to C. virginica. Cultivation of larvae of several other species was a h tried by early workers. For example, Belding (1912) attempted to raise larvae of clams, Hercenuria nzercenaria (formerly Venm mrcen- aria), but without success. He concluded that there was no practical method for raising clam larvae to straight-hinge stage because of the small size and delicate nature of the egg. Wells (1927), however, was more successful and carried the clam larvae in his cultures until they metamorphosed.
Even in more recent years the situation remained practically the same. This is well demonstrated by the work of Yoshida (1963) who, in his attempts to identify larvae of Japanese bivalves, had to depend upon obtaining the larvae from plankton, instead of trying to grow them from fertilized eggs under controlled laboratory conditions where their identity would be assured. The difficulties experienced as recently as 1953 by Nikitin and Turpaeva (1959), in their attempts to raise l&rves of some bivalves of the Black Sea by using old methods, vouch for the inefficiency of these now obsolete approaches.
Obviously, as the general studies of marine organisms progress, the necessity for methods by means of which bivalve larvae can be reared successfully becomes more and more urgent. The availability of such methods would immediately offer the opportunity to study the effects of numerous environmental factors, singly and in combination, upon the growth of larvae, thus helping to determine the physiological requirements of these organisms. It would also offer the means for studying the genetics of bivalves and initiating properly controlled experiments on selective breeding of these mollusks. Moreover, by

REARINQ OF BIVALVE MOLLUSKS 3
growing larvae under different conditions their diseases and parasites oould be studied and methods for their control developed. Finally, because the larvae of many species of bivalves are much alike in size and appearance, it was virtually impossible to identify them, with any degree of accuracy, in plankton collections. With the recent dovelopmelit of mtrthods of rearing larvan in the laboratory, howaver, this difficulty Rhoiiltl Noon disappear bccautw larvae found in plankton can now be easily and accurately compared with preserved samples and photomicrographs of larvae grown from known parents under controlled conditions.
By using successfully conditioning and rearing methods, many aepects of which were developed at Milford Laboratory (Loosanoff and Davis, 1950; Loosrtnoff, 1964) and are described in this article, larvae of approximately twenty species of bivalves have been cultured at Milford. Not all of these species are indigenous to New England waters or even to our Atlantic coast. Several are native to the Pacific and one species came from Europe. The non-indigenous forms were representa- tives of commercially important species in which we were interested. The bivalves, the lmae of which have been reared from fertilization to metamorphosis, included the transverse arc clam, Arca tranevert~a ; the ribbed mussel, Modiolue demhsua; the common mussel, Mytilue eduli8; the bay scallop, Pecten iwadiam; the jingle shell, AnomM; eimpkx; tho European oyster, Ostrmz edulie; the native Pacific coaat oyster, Ostrea lurida; the American oystir, Cras8oedreu Virginia; the Japanese oyster, Crmsmtrea glgm ; Morton's cockle, 7 Laehrdium morloni; the hard shell clam, Mercenaria (-Venue) mereenaria, and its relative, Mercenuria (- Venue) ca9wpechiensi.9 ; hybrids of thew two species; the Japanem obm, Tupu ~emi&maah; t h wmcll clam, Pitar ( - - -Callocardia) mMThwLnu ; tho rock borer, Petrieokcplroladi- f m b ; the razor clam, lim.8 directw ; tho aurf clam, Nactra (-8pi8u&) 801idissi?na ; the soft shell clam, Mya arenuria ; and the common ship- worm, Teredo navalia.
Of the above species the larvae of Crassostrea Virginim and Mercen- aria rnercenaria have been studied most intensively and, as a result, we h v e accumulated an extensive knowledge of their physiological and ecologioal requirements (Loosanoff and Davis, 1960; Looscmoff et al., 1961 ; Loosanoff and Dmis, 19528; Loosanoff and Davis, 1962b; Davis, 1963; Loosanoff, 1964; Loosanoff et al., 1955; Davis and Chanley, 1956a; Davis, 1968; Davis and Guillard, 1958; Loosanoff, 1958a ; Loosanoff, 1958b ; Loosanoff, 1969 ; and Davis, 1960). Fhveral other species, such &R the European oy~ter, O&ea r iht in, and tho Olympia oyster, O&m lurida, have alno roceivcd mudl nttrwtion.
II a

4 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
Most of the other species, however, were studied less intensively, work on them being confined to culturing their larvae and observing the appearance and general behavior of the latter. Naturally, our knowledge of the requirements of larvae of such species is still fragmentary but, nevertheless, we shall present the information already available even though it is admittedly incomplete.
11. EQUIPMENT The rearing of larval and juvenile bivalves requires an adequate
supply of sea water of proper salinity and free of substances that may interfere with their normal development. The water used at Milford Laboratory is pumped from the Wepawaug River a t a point about 100 yd from its entrance into Long Island Sound. Because the tidal rise and fall in this area is from B to 10 ft, the flushing rate of thiR comparativcly narrow and shdlow inlet i N relatively high.
The sca water is pumped into a 8000-gal wooden storage tank located in tho laboratory attic. Because pumping normally takes place 16 hr before and after the high tide stage, the salinity of the water is usually near 27 parts per thousand, which is virtually the same as in Long Island Sound, where the majority of the forms, the larvae of which are described in this article, exists. To assure a supply of water of high salinity the intake of the salt water system is located approximately 4 ft below the mean low water mark; therefore, it is at a safe distance from the surface layers which, after periods of heavy rains, may be greatly diluted.
The main pump providing the laboratory with salt water is rubber- lined. The intake and distribution lines, as well as the check and cutoff valves, are made of lead. The faucetn, however, are of hard rubber. The storage tank is of cyj)r(:sH wootl'and irs peiritd innid,? with asphalt paint.
We prefer lead pipe8 because, although p i p mcufc! of Hnveral new plastics are nontoxic, light and inaxpennive, they ponrrtnn mvctrel important disadvantages. One of them ia that n i n w it in oftcrtr rremwnrrry to reduce fouling inside of tho pipw by trmtjng t h m with hot we& or steam, t,hiH trontment, Commoniy uned with 1cta.d pipcH, cannot be employed in HywtarnR contairiing plaHtic parts ae i t may cau~e damage, especially a t the joints of the pipeline.
Another serious disadvantage in wing plastics is that they adsorb and absorb many chemicals, including insecticides, and once con- taminated can themselves become a murce of later contamination of the sea water. Moreover, since somc pla~tics are permeable to inseoti- cides and other compounds, these materials might enter from the sur-

REARING OF BIVALVE MOLLUSKS 6
rounding soil into pipes oarrying sea water. Finally, some laboratories that have plastio sea water systems have complained that since these pipes are not electrically self-grounded, they present a serious element of danger in laboratories with wet floors.
We are finding an increasing usage for plastic pumps and pipes, especially in our temporary installations. We have also found that tmks made of Fiberglas, instead of wood, can be advantageously used, especially in areas where wood-boring organisms, such aa Teredo, am common.
Fro. 1. Diegrain of wattw filter designed to remove dl perticu~ete matter larger than 15 p in diameter. Description in text.
Outlet t u b
Normally, in addition to small algae on which larvttl arid juvenile mollusks feed, sea water oontains many large diatom, frw-nwimming crustaccoans, gaatropods, wormn, otc., arid t h c k uggn md lervao. Many of 1,tieHn I'orrnn (:olii[~4m with bivalve lsrvw for f o d , pruy on thorn or may w o n hsrbor diwxww or pamites ttirit oould bo tranumittd to larvae. We prevent undesirable organismH of larger sizm from entering our larval cultures by filtering the water and later killing the smaller forme with ultmviolati light.
'I'ho tiltcrr s l m o i i t coneists of a polyvinylohloride (PVC) core wound with Orlon. 'l'he complete unit (Fig. 1) is manufactured by Commer- cial Filters Corporation, Melrose, Massachusetts (filter no. CFX1-10-5

6 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
with an 015-RlOX filter element). These filters, designed to remove all particulate matter larger than 15 p in diameter, are made with a variety of core and winding materials. We chose the PVC core because it is nontoxic, and the Orlon winding because it is inexpensive, nontoxic and does not support bacterial growth.
To prevent fungus diseases in clam larvae and juveniles we began treating sea water with ultraviolet light in 1954 and, within a short time, had some evidence that such treatment, even of running, unfiltered sea
Rubber squeeze gasket
'- 1.25 in. 1.0. PVC pipe
FICI. 2. I ' h o t ~ o K r t q t t ~ ( C l h n t ! ) 1 i t ~ 1 ~ l r t ~ w l l i ~ (/dew) l,f iili~rfiviolvt, w1rt.w i~ rwtml t~ t l i~ I l t l i i ,
i t w i t rrl. Mill iwtl !$ictlrtyic.rtl t , t d r t r r / ~ l . f ) q ~ . !)f*~crl~lt,tftrl ill t . 0 ~ 1..
w c ~ k ~ , WILH t w l p f i i l in provmting mortdity of juvenile clams. I n the sumnicr of 1!)59 it was definitely demonstrated that larval cultures, receiving treated water and untreated phytoplankton from the outdoor mass culture, developed fungus, whereas larval cultures in which phytoplankton and sea water were both treated did not. Since that time, i t has become a routine practice to treat with ultraviolet light all sea water used for our larval culture8 and for keeping recently-eot clams and oysters. Moreover, we are attempting to supply ultraviolet- treated running sea water to all containers ip which later stage8 of juvenile clams are grown.

REARING OF BIVALVE MOLLUSKS 7
Ultraviolet treatment of sea water for purification of shellhh has been described by several workers in Japan (Sato, 1964; Satoh, 1960) and Wood (1961) in England. As is the practice in our laboratory, Waugh (1958) also used ultraviolet-treated sea water for rearing larvae of the European oyster, 0. edulig. Several of these authors have described the equipment used but, because of certain considerations, we constructed our own units, a description of whioh is offered here.
The ultraviolet water treatment unit consisfs of a 1)-in inside diameter PVC pipe, 30 in long, threaded at each end for caps (Fig. 2). A small ring of PVC is cut to fit inside of each end of this pipe and reamed to act as a spacer for a 26-mm Vycor tube. A squeeze gasket is wed to make a water-tight seal between the Vycor tube and the end of the PVC pipe. An inlet tube is located on the side at one end of the PVC pipe and an outlet tube is located on the opposite side at the other end. The 33-in-long, slimline ultraviolet tube lays free in the 324411- long Vycor tube and extends slightly beyond at each end.
In practice we use two such units connected in a series so that the water passes the length of both tubes. Since there is only about a 4-in layer of water surrounding the Vycor tube, this apparatus, when used with filtered sea water, should give practically sterile water at the rate of flow of about 10 gal per min. With unfiltered sea water the efficiency is not expecid to be as great, but our experience has shown that even then the treatment is of considerable help in reducing mortality of juvenile clams and in preventing fouling by tunicates, worms and bryozoa.
To condition mollusks for out-of-season spawning it is necessary to keep them in running sea water at temperatures of 18" to 20°C or sometimes higher. Warm sea water is also needed for rearing larvae and juveniles during the cold season. Since the water must not contact toxic metals, conventional water heaters cannot be used. Therefore, to heat the water we use a type of heat exchanger (haanoff , 1949). The sea water is heated as i t passes through a coil of lead pipe immersed in hot fresh water, which fills the tank of a conventional gas water heater that has had the top removed to permit insertion of the lead coil (Fig. 3). However, because the thermostatic controls of a conven- tional water heater are not sufficiently accurate, the gas flame is con- trolled through a solenoid gas valve by a Minneapolis-Honeywell thermostat (T415A323XA3). The thermostat-sensing bulb is e n c d in a lead well in the warm sea water line and maintains the temperature at 37°C f 0-5"C.
By mixing varied amounts of cold and heated sea water any tem- perature between that of the unheated water and 37°C can be main-

8 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
tained. In our winter work, when the temperature of the water in Milford Harbor is near freezing, we often simultanaously employ streams of water a t 5", lo", 15", 20", 25", 30", and even 35°C. Of course, any other temperature within this range can also be maintained by using constant level jars of cold and warm water and regulating the
FIG. 3. Hoat exchanger to provi~le laboratory with warm Ma wabr. A, thormortai. xunniriK bnib ; 13, thermostat; C, air j)ump Lo provent alratificatiorr of frcmh water in tank ; 1), tcrrik ; E, gan watnr hest.orn ; F, nolorioicl KM vdvctn.
flow from thcxe jam into a mixing ahsmtwr from which watcr c ~ f d ~ i i r t ~ l ttmpc?ratim flow8 in to trayH or aqimria whom expc?rirnotttal anirnalH IAN! k q ) t (Ng. 4).
'1'0 k w p larvd cultures at tlrsired temperatures vltrious constant temperature devices are used. Since a temperature of about 24°C is

FIQ. 4. Racks of trays for conditioning bivalves for spawning. Racks are provided with running ma water of differoiit tomperat,ures. Constant bvol jars for warm urid cold wutor are H W I ~ i i i upyw left corncr. Watw from these two jarn ie rnixod in required proportionH in the smalle~r glum jure located on lower sticdf. Ultra- violet uiiit for tmatrnerrt, of water is located at right.

10 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
near the optimum for growth of algae, such as Isochrysis galbanu (Ukeles, 1961), which are the best larval foods, this temperature level ia often maintained. However, because 24°C is somewhat above normal room temperature, our simplest and most commonly used temperature control devices are lead-lined water tables, 3 f t x 12 ft, that serve &B
constant temperature water baths (Fig. 6). These tables are filled to a depth of 3 to 4 in with tap water, which is kept in constant circulation by a pump that takes water at one end of the table and discharges it
Expansion tonk
Thermostat
Clrculotion pump
FIG. 6. Two methods of heating water for constant tempereture baths. Diagram of closed system above, and open system below.
at the other end. A Minneapolii+Honeywell thermofitat (T41 SA323XA3) with the sensing bulb enclo~ed in a lead pipe, immered in the water on the table, controls a 1000- or 1500-W immerfiion heater. Tho heater may be either inserted in the pipeline, through which the circu- lating pump transfers-water from one end of the table to the other (open system) or it may be inserted in a small hot wabr tank anti tho heat transferred to the water on the table by passage of the watcr from the tank through a lead coil or loop immersed in the water on the table (clot~ed or hcat exchangc: sy,rtcm) (Fig. 6).

REARING OF BIVALVE MOLLUSKS 11
Whenever the larval cultures are to be kept at about 24OC any type of container, the lower part of which is immersed in the bath, will maintain this temperature because convection currents within veaaels prevent temperature stratification. Even in oontainera of
ho. 7. Constant temperature apparatue coneistirig of 6 unita. Ternperaturn of each unit can be adjusted independently and mairitaiiid at any desired level within the range from 5* to 37°C. If necosnary. all iiriita may be maintained at the oame temperature.
Merent sizes and shapes the temperature will vary only dightly, while in a series of individual contaiiier~ of the same type the water will be maintained at alrnoRt procirrely tho same tomperaturoe.
A similar rtrrungcrrnont can bo I I H ~ ~ to maintain temporatwee below that of tho &om by omploying tb liquid cooler, instead of a

12 VICTOR L. LOOSANOFF A N D IIARRY C. D A V I S
heater. Units that combine a heater and cooler and will maintain temperatures above or below room temperature are also available. How- ever, when cooling devices are used it is necessary to keep the water in cultures continuously agitated to prevent temperature stratification.
To study the effects of different temperatures e n dcvelopmeitt of eggs and larvac of bivalves anothcr upparatus wa8 devisctl (lpig. 7). This apparatus, which can also be used for studies of many other forms, consists of a series of six lead-lined tanks, each 15 in wide by
FIQ. 8. Constant ternpersture sir chsrnbrn in which tray8 or othur I?oIltAbifJlm with larval or juvede mollusks are held. p%?otric? heater and fan for CirClJhhJl( Lhn air can be seen in right hang corner of unit.
Constant temperature air chambrn in which tray8 or othur I?oIltAbifJlrn with larval or juvenile mollusks m e held. Electric? heater and fan for ~irciihitiiig Lhn air can be seen in right hang corner of unit.
Constant temperature air chambrn in which tray8 or othur I?oIltAbifJlrn with larval or juvenile mollusks m e held. Electric? heater and fan for ~irciihitiiig Lhn air can be seen in right hang corner of unit.
26 in long and 13 in deep and f i l l r d with frwh wntm to IL ( I ~ t i w I level. Vessels containing experirneritul animal8 &re imrnamud i r i therw, tanks.
To mriiritain wat,er in thc tmiks at desired temperature8 each tank is equipped, idoiig i t H walls, with l o o p of tubing to circulatc cold and hot water. The amount of watcr paHsing through each of them tube8 is corit,rollcvi I)y dout)Ip-actiort tlic*rrrtontsts which activate Holenoid vi t lvw n o thtt if' i,1w tc~~npc~iit,rir(t i t 1 iti1.V tatik falls i)elow the thermo- n t ~ ~ t , ~ ( $ 1 ~itig. t l i t . ~ I b I y t ? i 1 1 thc Iiiw, tlwollgfll which hot water circulates, opsus. allowing hot, water to flow through the loop. If , o n the other

REARING OF BIVALVE MOLLUSKS 13
hand, the temperature exceeds that indicated by the thermostat, the solenoid valve in the cold water line opens, allowing circulating cold water to reduce the tank temperature. To have the entire mass of water at uniform temperature a circulator pump is employed. The temperature controls of the entire unit are so arranged that all tanks may be maintained a t the same temperature, within the range from 5" to 37°C or, if necessary, at different temperatures.
Fro. 0, Tompertrturo rtppnratus for rirnahtnour r t u d h of c:ortairi wrpmb of bsturvlor of juvonihi rnoll~inkn in rtinnii~~ wutor of clifTorenb but oonstnnt tomperaturn. A, wwnp t i a h for tLir trrppod in nea wator linen; B, oold and warm water constant Ifivcil jaw tiwin wlrinti wutor in diffnrtlnt proportions enters mixing jam (C). Ewh jrrr i m y Iw nisic~lainod nt any teinportrtwt ranging from 6" (in winter) to 35OC; D, ultfrHvirdtrtl mi& thi-ougti whirli all norr wetor peeeee to corinhnt level jam; E, conntant love1 jars from whirli phytoplankton ia added at a definite rate to running sea water ; F, floats controlling levels in constant level jam.
To control the temperature of the air chambers in which culture vessels are kept in some experiments thermostatically-controlled eleo- tric heaters are used. For example, in experiments, where a sories of four banks of five trayu each of standing water are umd to hold juvenilo clams, a uniform constant temperature ie maintaincd by encloNing ail twenty trays in a chamber where thcrmo~tatically-controlled c!lectrio heaters are installed (Fig. 8). When heated air is uwd, however,

14 VICTOR L. LOOSANOFF A N D HARRY C. DAVIS
special Imcautions art' noct?smry to prevent its Ht,rntilict~tiol\. 'l'o achieve this in the enclosure where our racks of trays aro kcpt a large fan forcing air through has been found sufficient.
Still another temperature apparatus is used at our laboratory for simultaneous studies of growth of juvenile mollusks in running water of different but constant temperatures (Fig. 9). The entire apparatus consists of seven independent units, each insulated so as not to be affected by outside temperatures. As many as five trays may be placed in each chamber. By mixing, in winter, different proportions of warm and cold sea water, temperatures ranging from about 5" to 35°C can be maintained quite accurately.
The amount of water entering each tray can be adjusted to a desired rate and, when necessary, the trays in all seven chambers may receive the same quantity of water and plankton food per hour. Ae a rule, sea water and food, before entering trays containing juvenile mollusks, are passed through the special unit where they are sterilized by ultraviolet rays.
In addition to various apparatus and devices discussed in thia section there are several others that have been used in special studies. A description of these will bc given elsewhere.
111. CONDITIONING MOLLUSKS FOR OUT-OF-SEASON SPAWNING
Before the present method of providing laboratories with warm water in winter was developed, experiments on most of the bivalves and, especially, their larvae were confined, in New England waters and similar areas, almost exclusively to the short periods of natural propagation, usually lanting for'only the '24 or 3 Hummer month However, since i t was found that in many hivalvc:n, by uning p r q w conditioning methods, norms1 de-elopmen t of gonuln can b: n t h u b t w l and spawning induced during late fall, winter and Rpring, the e x p r i - mental period has been greatly expanded (Looanoff, 1948).
Conditioning of bivalves to develop mature gonads during the cold part of the year is relatively simple. It consists of placing mollusks, brought from their natural environment where water temperature may be near freezing, into somewhat warmer water and then gradually increasing the temperature several degrees each day until tho desired level is reached (Loosanoff and DaviR, 1060). Sometimc~, especially towards tho spring, instoad of a gradual conditioning the mollusks can tw p l ~ t d dircctly in wrktcr of about 20°C. As a rule, the gametes obtainotl from theee mollusks were no less viable than from those conditioned gradually. We have often employed this more rapid

REARING OF BIVALVE MOLLUSKS 15
approach, thus shortening by several days the length of the con- ditioning period which, for oysters kept a t 2OoC, is approximately 3 to 4 weeks.
The conditioning period can also be shortened by keeping mollusks at temperatures higher than 20°C (Loosanoff and Davis, 1962b). For example, Crtlssostrca virginica kept a t 26°C developed ripe spermatozoa and fertilizable eggs by the 6th day, and light spawning could be induced on the 7th day. When kept a t 30°C ripe spermatozoa and fertilizable eggs were found in oysters which, only 3 days before, were brought from the ice-covered harbor where they were hibernating. Some oysters of this group were induced to spawn on the 6th day.
Obtaining spawn from another common bivalve, the hard shell clam, Mercenuria mercenaria, is also relatively simple in summer. It is often accomplished merely by raising the water temperature a few degrees and by adding a sperm suspension (Loosanoff, 1937a). Pre- viously, as already mentioned, this could be accomplished only during a short period, whereas, using our recently-developed methods, it ie now possible to obtain ripe gametes and raise lmae of this speoiee on a year-round basis (Loosanoff and Davis, 1960, 1961).
The method for conditioning clams for spawning in winter is the same as that described for oysters. The entire conditioning period takes approximately 2 to 3 weeks, but can be made even shorter towards or during spring. On several ocoasions clams brought directly from natural beds during early spring could be induced to spawn without any preliminary conditioning. However, this method often failed and cannot be considered reliable. Usually, only males responded on suoh occasions. As a rule, some conditioning of clams is necewry, even towards spriiig, to have a reliable nourm of "RWH arld nptrm.
Our Htiitl i(w hrtvo (ktmonstrrttod that bivalve8 can be conditioned for late fall and early winter spawning only after they reoover from the natural spawning activities of the preceding summer. This recovery oonsiets of I K I R I ~ Y aomplox physiological proattsses leading, in general, to t i i ~ ~ ~ i i i ~ i ~ i l ~ ~ , i ~ ) i ~ of roxorvtk mnt,ericilN, of wliioli glyoogen is probably h ino~t , iinpor(mrt, (t,ooetm~ff, 1937t1,, 1f~4p). Since many speoies of bivalvox of Long Island Hound, iiicluding oysters and clams, some- times continue to spawn until lato August or even the middle of Sep- tember and are not completely recovered from these activities until the end of November, they cannot be conditioned for spawning during these months.
We solved the problem of supplying ripe mollusks during the period from late August to late November by delaying their gonad develop- ment and spawning until late fall (Loosanoff and DaviR, IA961). Clams,

16 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
M . mercemria, and oysters, C . virginica, are taken from Long Island Sound early in the season, usually late in May, long before the beginning of their natural spawning, and transplanted to the waters of Maino, where the summer temperature averages about 7" lower than in our waters. This temperature, while permitting slow development of gonads, is, nevertheless, low enough to prevent spawning. Thus, when oysters and clams in Long Island Sound are already spent, those transplanted to Maine still retain their spawn. In the fall, small groups of these mollusks are routinely shipped back to our laboratory, where they are easily induced to spawn, providing normal gametes which are unobtainable locally during that time of the year.
By using the above method spawning of C. virgiiticu can be post- poned only for 6 or 8 weeks after oysters of Long Island Sound are completely spent. After that period the oysters, even if they are still kept in the waters of Maine, begin to resorb undischarged gonad material and, thereafter, become useless as spawners. We overcame this difficulty by developing another useful method, which postpones early gonad resorption. It consists of conditioning oysters early in the spring and spawning them at Milford by early June. After that they are transferred to the colder waters of Maine. Oysters treated in this manner must resorb old gonads and build up glycogen before developing new gonads. Because they are compelled to go through these processes, these oysters reach ripeness much later in the season than those that are planted in Maine without spring conditioning and spawning and, aa a result, they do not begin to resorb gonad material aa early as do unspawned oysters transferred to Maine at the same time. Taking advantage of this Rituation we have: h n obtaining normal larvaa from
I3conu.rc~ M. mrrcenaria d o o H not, rtrnorh untlinchargod gonad material in tho fall, an oynterH do, tranuforritig them to the coldor wtltern of Maino in tlw xpritlg p v o d to he n highly HatiAfactory method of delaying spawning. Under thea: conditions the clams retain sperm or eggs throughout the summer imd, a8 a reuult, can be induced to spawn throughout the next fall, winter and even during the following spring, always producing gametes which develop into normal larvae.
We have a160 delayed spawning of clams and oysters by taking these mollusks early in the summer from their natural habitat and keeping them in insulated boxeH through which mochttriic~ll,y-oooled sea water flowed. U~ually, only a uomparativcly Hrrinll numhor of adult mollusks could be convenicn tly kept urrt1r:r ttieru: cc~rirlitior~ and, as a rule, bivalves so treated wcre in much jworcr conditiorr thn those kept under natural surroundings in the waters of Maine. More-
0Ck)tMtr .IfLflll&r,J' fr0m HpfbWrl Of OyHtA'rH HO !Jcl&h%l.

REARING OF BIVALVE MOLLUSKS 17
over, a failure of the artificial refrigeration system may cause the entire stock to spawn prematurely.
By combining our two methods, one consisting of conditioning mollusks for spawning during the cold periods and the other of delaying gonad development and preventing spawning during their normal reproducti.re seamn, ripe bivalves may now be available throughout the entire year.
We have also found (Loosanoff and Davis, 195%) that C. Virginia end Af. mercenaria are able to reproduce several times a year, provided that changes in ecological conditions, especially temperature, are 80
controlled that these mollusks can rapidly recover from spawning, accumulate in their bodies material needed for gonad development, and begin the cycle again. As a result of the discovery of these new approaches and methods, as much can now be accomplished in one yeax in certain fields of the biology of bivalves as could formerly be done in three or four.
It should be emphasized that our conditioning methods are not equally successful or applicable to all groups of oysters, C. virgi?cica, and, perhaps, certain other species of bivalves of our Atlantic and Gulf coasts. This is probably because populations of these species are not genetically homogeneous, but consist of different physiological races. We began to suspect the existence of such races in C. virgin& as early as 1937 (Loosanoff and Engle, 1942). Stauber (1960), in reviewing the literature on spawning of the American oyster, also cqme to the oonolusion that oysters from different areas along our Atlantic coaat may belong to different geographical races. Our experiments in this field strongly supported this assumption by demonstrating that, even though all these oysters belong to the same species, the temperature requirements for gonad development and spawning of the northern populations are definitely lower than those of the southern group (Loosanoff and Nomejko, 1951).
The results of our later, more extensive studies, in whioh several thousand specimens representing populations of different areaa of the oyster-producing belt extending from the Gulf of Mexico to Cape Cod were used, fully supported our original conclusions (Loosanoff, 1968e). The oysters used in these experiments and observations were from Florida (Gulf of Mexico), South Carolina, Virginia, New J e m y and New England. They were received in the fall, after they had conlpletely spawned in their native environment, and were kept in Milford Harbor throughout early winter. Some time in January the first groups of these oysters were transferred to the laboratory to be conditioned for spawning.
A.X.B. C

18 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
We employed two criteria to evaluate ripeness of the oysters. The first was to ascertain the number of days needed for 50% of the oysters constituting a sample to develop active spermatozoa or fertilizable eggs. Secondly, we had to determine the length of the conditioning period before spawning in 50% of the oysters could be induced by our usual method. Each sample contained fifty adult individuals.
The experiments showed conclusively that Long Island Sound oysters develop gonads and can be induced to spawn after considerably shorter conditioning periods than those required by southern oysters. When kept at the same temperatures oysters from New Jersey, although slower than those of Long Island Sound, showed, nevertheless, much faster gonad development than oysters of Virginia, South Carolina and Florida. In averaging the results of the experiments it was found that 50% of Long Island Sound oysters, conditioned at 21", 24" and 27"C, contained mature gametes after only 15,s and 5 days, respectively. The corresponding groups of New Jersey oysters reached this stage only after 65, 32 and 22 days, thus requiring three or four times as long at the three above-mentioned temperatures as did the northern race.
In certain experiments we were able to induce spawning in 50% of Long Island Sound oysters after only 18 days of conditioning at 21°C. To achieve the same results with New Jersey oysters 78 days were needed. The more southern groups kept under the same conditions failed, as a rule, to produce 50% spawners.
The most striking differences were noticed when oysters of different geographical regions were kept a t relatively low temperatures. For example, after 68 days of conditioning at 12"C, 67% of Long Island Sound oysters contained mature eggs or spermatozoa. In this group we were able to induce spawning in one male and, 10 days later, in one female. Oysters of the other groups kept at the same temperature contained not a single individual with mature gonads, even after 78 days. Moreover, in the majority of New Jersey and Virginia oysters and in all of those from South Caroliria and Florida the gonads were so poorly developed that the sexes could not be distinguished, even by microscopic examination of the raw gonad material.
The method of inducing spawning of oysters and clams in summer has already been described in detail (Galtsoff, 1930, 1932; Loosanoff, 1937a, 1954). The same method, as a rule, haa also been used to induce spawning in other bivalves. Tn general, our present method can be described as follows : After the proper conditioning period ripe bivalves are placed in glass spawning dishes, each containing approximately 1 liter of sea water of the same temperature as that a t which mollusks

FIG. 10. Ripe oysters in spawning dishes. Male oyster is shown spawning in center dish.
11 . Inducing spawning of clams, oytern and other bivalve8 by irnrnerninK dishes of sea water containing anirnt~lx iri warm water on qmwriitrg table.
3

20 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
were conditioned (Fig. 10). These dishes are partly immersed in a large tray or sink, which is filled with hot water, thus quickly raising the temperature in the dishes to the desired level (Fig. 11).
I n some instances thermostimulation alone is sufficient to induce spawning. In other cases, however, mollusks need additional stimula- tion, which consists of adding to the water small quantities of sperm or egg suspension made from gonadal material of ripe individuals of the same species. Many forms quickly respond to combined thermal and chemical stimuli ; others, such as the common mussel, Mytilm edulis, do not usually respond to this method but can, nevertheless, be induced to sFawn by other means, which will be discussed later.
In a special series of experiments we tried to cause artificial discharge of reproductive elements by injecting weak solutions of Mn,OH and other chemicals into the bodies of bivalves that could not be spawned by other means. The results were usually not gratifying, except in the case of M. edulis, when injection was made in its adductor muscle.
At the time of our experiments to induce spawning in ripe bivalves we were already aware of the success of Japanese workers in inducing spawning in mussels by giving them a mild electric shock. We repeated these experiments but, unfortunately, with indifferent results.
In still other species, for example, Modiolus demissus, all our methods, including those that were successful in the case of Mytilwr edulis, proved to be ineffective in inducing spawning. Therefore, unlese ribbed mussels spawn naturally, thus providing normally fertilized eggs, no other means, except perhaps stripping, are left for obtaining their spawn.
Fertilizable eggs of many q)ecics, including thom of C. uirginica (BrookB, 1880), can be ohtainvd Ijy fitripping matiirr: fr:mitbn hiit, since many of these forms spawn HO readily in rcqJonwc: to chemical and thermal stimulations, it is seldom necessary to resort to thie mectne. However, when working with other species, especially those that cannot be spawned by conventional methods, stripping may be the only way to obtain ripe eggs. It i H a Himplc procem arid i~ carried an as follows: After removing the outer memhranc? that cr)vorH thH gonda, the mollusk is gently r i n d in nea water. Thiw ar:t,ion Hc!lJwat,.en front the gonad large number8 of egg8 without rw:rioon irrjiiry b them. Using a series of sieves of proper size meRh tho ~ g g ~ arc later frmd of bIood cells, pieces of tissue, etc., and then placed in 8ea wator to which sperm is added. The fertilized eggs can then be placed in culture vessels.
This approach is possible only for eggs of those forms in which the germinal vesicle dissolves after stripping. In many species, however,

REARING OF BIVALVE MOLLUSKS 21
including Mercenuria mercenaria and Pitar morrhuana, attempts to fertilize stripped eggs usually fail because in these eggs the germinal vesicles remain intact and, as a result, fertilization does not occur. Under normal conditions the germinal vesicles in eggs of such species dissolve while they are still in the ovaries of the female, just before they are discharged in the process of spawning. Upon dissolution of the germinal vesicle the germinal spindle is formed and the discharged egg is ready for fertilization (Loosanoff, 1953).
Recently, following the suggestion of Mr. David Tranter of Australia, we uHed a weak solution of ammonium hydroxide to break the germinal vesicle of eggs of certain bivalves. By employing this method we succeeded in raising normal larvae from eggs stripped from Mereenaria mercenaria, Tapes sernidecussata and several other species.
After the eggs were washed from a gonad they were passed through a cotme, SO-mesh screen to remove debris, large pieces of tissue, etc. T,;tter, thcy were washed OIL a 3%-mesh screen which retained the eggs but let pass the body fluids that might pollute the water in culture vessels. After that, 3 ml of 0.1 normal solution of ammonium hydrox- ide were added to every 100 ml of the prepared suspension of eggs in sea water. After the eggs were in this solution for some time they were wmhed again on a 325-mesh screen, being finally ready for fertilization. A more detailed description of handling fertilized eggs will be given in tlm section on methods of cultivation of eggs and larvae.
'I'hc lwgth of caxposure to the solution of ammonium hydroxide may vary soniewhat from species to species. The following table shows the ratio between length of exposure and percentage of normally developing eggs of M. rnercenuria :
15 minutas---32'%) 30 minutee-] 6%) 45 minutes- !Yg, 60 minutes- 3% 75minutes- 2% 90 minulm- 07)
Even after 90 minutun of cxpoHiirr: to l h nolution of' tbrrifnorriutn
hydroxidc! somc eggs became ji:rtiliml, h u t their dcvolopmer~t ww not normd.
The percen tage of normal larvae obtained from chemically-treated eggs was low compared to that of naturally spawned eggs but, never- theless, it was high enough to permit successful culturing of larvae of those species in which we were not able to induce spawning. Perhaps by changing the concentration of ammonium hydroxide, using other

-- a j - 7 VICTOIC L. LOOSANOFI.’ A N D IIAHRY C . DAVIS
chemical agents, or by improving the methods of stripping wen better results may 1~ obtained.
Finitlly. t l ic ixl ucre scvcrnl qwcit~s of’ I)ivalvcs wliic.11 W P ooulti ncitlirr spawn artificidly nor c w l l w t t I i c h i r normally tlisc~lmrgcd eggs. Moreowr, eggs strippcd from somc. of thew: forms coultl not b c . ftirtiliacd regardless of various prtxparatory measures, which includcd the chemical treatment described above.
Fecundity of many lamellibranchs, especially those of commercial importance, has been speculated upon for a long time. Brooks (1880) estimated that C. virginica could produce between 18 750 000 and 125 000 000 eggs. He based his estimate upon volume of material removed from the ripe female, but stated that this figure should be reduced by approximately 50% because of other matter that was measured together with eggs. Churchill (1920) stated that a large oyster may discharge 60 million eggs, while Galtsoff (1930) estimated that the number of eggs released in a single spawning may range between 15 and 115 million. He concluded that the maximum number of eggs that can be released by a single female during the entire spawning season is approximately half a billion. Burkenroad (1947), without offering any experimental observations of his own, suggested that Galtsoff’s estimate was approximately ten times too high. Belding (1912) estimated that M . mercenaria, 2 $ in long, produces an average of 2 million eggs, a figure not substantiated by experimental studies.
Since reliable informat,ion on the fecundity of even the most com- monly studied pelecypods was unavailable, experiments were under- taken by Davis and Chanley (1956h) to determine total numbers of eggs actually produced by individual oysters, C. virginica, and clams, M . mercenaria, under natural and artificial conditions.
The first series of observations was made on seventy-five oysters, measuring from 39 to 4q in long, and on the same number of clams approximately 3 to 4 in long.
The experiments were conducted in the laboratory during the win- ter, a most convenient period for proper conditioning of both clams and oysters. Each bivalve was individually numbered and a complete record was kept of its behavior during the entire experiment. The first group of oysters composed of twenty-five individuals was spawned at 3-day intervals, the second group at 5-day intervals and the third at 7-day intervals. In clams, which were also divided into three groups of twenty-five individuals each, spawning was induced a t 3-, 7- and 14-day intervals. Spawning of these groups of clams and oysters was continued a t the specified intervals for more than 2 months.

REARING OF BIVALVE MOLLUSKS 23
Experiments have shown that, as a rule, an individual oyster or clam does not discharge all its eggs or sperm in a single spawning, but will continue to spawn at intervals over extended periods. One female oyster spawned on sixteen occasions and a clam, eleven times. The number of spawnings per female oyster ranged from two to sixteen. The highest number of eggs was produced by an oyster that spawned nine times, while a female that spawned sixteen times ranked second. The lowest total number of eggs relemed by an oyster was by an individual that was induced to spawn seven times.
No significant difference was observed in the average number of eggs released during the entire experimental period, whether the oysters were induced to spawn at 3-, 5- or 7-day intervals, although the average number of spawnings per female oyster decreased progres- sively aa the intervals between spawnings were incremed. It waa also determined that female oysters having larger numbers of eggs tended to spawn more frequently than did females with smaller numbers.
The highest number of eggs released by any female clam in a single spawning waa 24.3 million and the total number released by individual clams during the entire experimental period of about 2 months ranged from 8 million to 39.5 million, with an average of about 24.6 million.
There waa no significant difference in the average number of eggs r e l d in a mason, whether the clams were spawned at 3-, 7- or 14-day intervals. It waa also found that correlation bebween the number of times a female clam spawned and the number of eggs pro- duced was not significantly different from zero.
An auxiliary ,experiment consisted of observations on spawning of fifty oysters taken from Milford Harbor early in April, brought into the laboratory and. placed in conditioning trays at temperatures of about 2OOC. Three weeks later these oysters were induced to spawn daily for 5 consecutive days, and seventeen females and twenty-four mdea responded during the first day. Altogether, this group contained twenty-four females and twenty-six males. Of the twenty-four females, fourteen spawned on 2 or more consmutive days, eight spawned on 3 or more consecutive days, five spawned on 4 or more conwcutive days, and three females spawned on each of the 5 days of the experiment. Eight males spawned each day.
The important contribution of this experiment was the clear-cut demonstration that there is no 2- to 5-day refractory period during which female oysters cannot be induced to spawn, aa maintained by Galtsoff (1930). On the contrary, the results suggest that upon proper
24 vIcTon L. LOOSANOFF A N D HARRY c . DAVIS
stimulation both male a d femsic oystcrs can spawn ;my time they have physiologically -ripe sex cells to discharge.
The final experiment consisted of observations o n nine female oysters developing gonads under normal conditions in Long Island Sound and ijlduced to spawn at t lw c ~ n c l of June. The total number of eggs dischargd by thc3sc. oyst(>i*s rangcd from 23.2 million to 85-8 million and averaged 84.1 millioir ~ g g s per female. Thus, both average number of eggs and maximam number per female of the summer
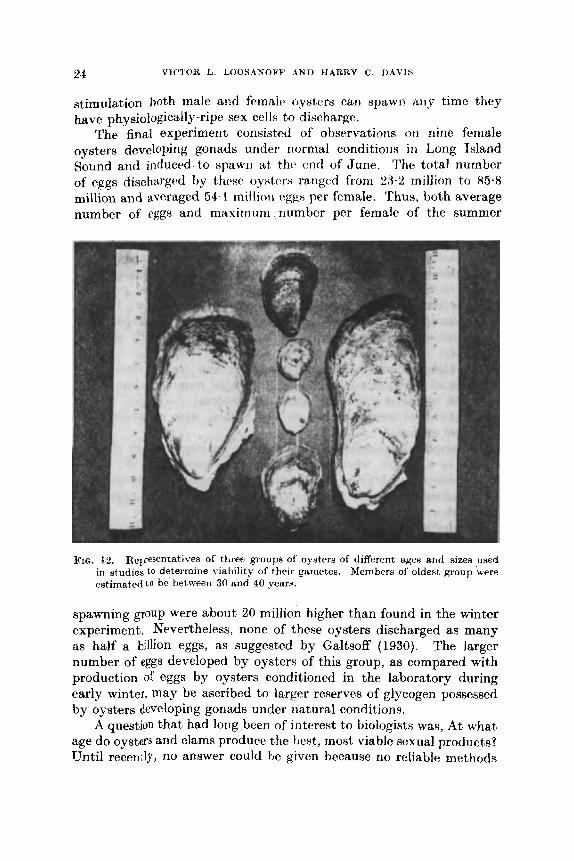
FIG. 1s. Representatives of three groups of oyiters of different uges arid A I Z ~ R uwd Members of oldeat, group 'were in studies to determine viability of their gametes.
estimated t o be between 30 arid 40 years.
spawning group were about 20 million higher than found in the winter experiment. Nevertheless, none of these oysters discharged as many as half a billion eggs, as suggested by Galtsoff (1930). The larger number of eggs developed by oysters of this group, as compared with production of eggs by oysters conditioned in the laboratory during early winter, may be ascribed to larger reserves of glycogen possessed by oysters developing gonads under natural conditions.
A question that had long been of interest to biologists waR, At what age do ovsters and clams produce the hest, most viable sexual products? until recently, no answer could be given because no reliable met})&

REAXINQ OF BIVALVE MOLLUSKS 26
were available to conduct critical experiments on development of eggs and growth of larvae to setting or post-setting stage. Since develop- ment of these methods, such studies havo become poaeible and recently were undertaken at our laboratory.
Three groups of oysters of different ages and sizes were conditioned for spawning (Fig. 12). The average age of individuals of the oldest group was estimated to be between 30 and 40 years, some of them being over 9 in long and over 4 in wide. The intermediate group of oysters of marketable size was from 5 to 7 years old, while the youngest p u p was composed of small oysters approximately 2 years old. These groups were conditioned and induced to spawn under controlled conditions, their larvae grown to setting stage, and rates of survival and growth of larvae from the three size groups compared.
The results showed no significant difference between oysters of the different age groups in the time needed to develop ripe gonads. We were somewhat surprised, however, to find that oysters of the oldest group responded to spawning stimuli more rapidly than individuals of the two younger groups.
There wks also no significant difference in percentage of fertilizable eggs because almost 100% of the eggs of all three groups became fertilized. Furthermore, the percentage of fertilized eggs developing to straight-hinge larval stage showed no consistent variation that could be ascribed to size or age of parent oysters. Finally, no consistent difference was found either in the sizes of the early straight-hinge larvae originating from eggs of different age-group oysters or in survival and rate of growth of their larvae.
Similar studies on hard shell clams, N. mercenaria, measuring from 37 to 110 mm in length, also showed that there was no significant difference in viability of spawn produced by clams of Werent sizes and ages. Often the differences between the progeny of individuals of the same size groups were as great m the differences between those of Werent ages and sizes. Larvae grown from eggs of clams of all three sizes were successfully carried to setting stage.
On the basis of the above-described experiments we came to the conclusion that since there was no Significant difference in the quality of spawn developed by individuals of different ages or sizes, mature oysters and clams of all age groups may be safely used as spawners.
Of special biological interest was the observation that the sexes among the oldest oysters were about evenly divided. This discovery was contrary to the old conception that in the oldest groups females should decidedly predominate in numbers. We also noticed that many of the largest and oldest oysters, while kcpt in the laboratory to be
,

26 VICTOR L. LOOSANOFF AND HARRY C. DAVIS '
conditioned for spawning, formed normal, new shell growth, thus indicating that even at that age and size the oysters did not lose their ability to grow (Fig. 12).
IV. CULTIVATION o w Earn A N D IARVAE OY IJIVAI~VES A . (Imertd descript.ioii~ (4 flu: dwdopmerd
Kggs of' l , i vdv( !x clitTcr i r i tiitLily w - q j o c t H , i~tclu~litig thcir H i m , color and spcxific gravity. They also tlifier in thickness of the membrane surrounding them (Costello et nl., 1957). In oysters and certain other forms this membrane is only a few microns thick. In others, however, such as M. mercenaria, the egg proper measures only 70 to 73 p, while the total diameter of the egg and surrounding gelatinous membrane is about 170 p. This membrane, in many instances, continues to sur- round the embryo past blastula stage and, on some occasions, until late trochophore stage is reached (Loosanoff and Davis, 1950).
We shall describe specific characteristics of the eggs later on, when discussing cach of the specien Atdied. Here, because the descrip- tion of a typical hivalve egg and its development to straight-hinge stage or, as i t is often called, early veliger has been given on many occasions, including Brooks (1880), MacBride (1914) and others, we shall present only a general picture of changes occurring from the moment the egg is discharged, or stripped, until it becomes a straight-hinge larva. This description is based upon observations made on eggs and early embryos of Mactra (= Spisula) solidissima, the surf clam, which is the largest bivalve of our Atlantic coast. It measures up to 79 in long and can be found in considerable numbers from Labrador to Cape H atteras. Additional informat,ioti on spawning of these clams and rcaririg of' their I i i rv i ic in i t i i : l i i i I w l i r r f.tw wi:t,icur f l 4 i t i K wi1,li rwwjtig
I tit. diiitii(+v of' IL i t i t ~ i ~ i r c (:gg of M. .wlidiwirn.ii, avcragw Mi.5 p (Fig. 13a). Costello et al. (1!)87) give the diameter of the unfertilized ovum of the same spocies as ranging hetwwn 53 and 56 p, thuti agreeing with oiir mrnsurements. According to Cahn (1851), who bases his conclusions o t i thc work of thc Japanese investigators, Kinoshita and Hinulo ( 1!)34), wtiosc ptq)cr WILS not nviiilable to us for consultation, the diatrictcr of ttic ~ g g of a cloxoly rc:lated form, Spisula sachalinensis, is only 50 p. Another group of ,Japanese workers (Imai et al., 1953), studying the same species reports that the diameter of the mature egg of this clam varies from 70 to 75 p, thus being considerably larger than the size given by Cahn. Jmgensen (1946) states that eggs of Spisula subtruncata of European waters vary in diameter from 50
O f IHI'VILI! f J 1 ' ~ ~ l f ~ f ! f Y ! l l ~ ~ HlN!(!i(S4.' I ,
to 55 p.

REARING OF BIVALVE MOLLUSKS 27
Early development of the egg of M . 80&ii88inza is basically the =me as that of many other bivalves. After dissolution of the germinal vesicle (Fig. 13b) the size and shape of the egg remain the same. If
Mactra (Spisula) solidissima Fro. 13. Development of Mactra (= Spisula) S o ~ i d i . ~ m m a from unfertilized egg (A) to
Diameter of egg is about 58 p, while length of early straight-hinge larva (H). streight-hinge larva is about 79 p. Dettrilrct dewription in text.
the fertilized egg is kept in water of about 20°C, the polar body is formed in about 45 min (Pig. 13c) end the two-cell stage, meaeuring about 65 p along the longest axis, is reached in 90 min (Fig. 13d).

28 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
Development of the egg of a bivalve, as described above, is typical only of a group in which the germinal vesicle breaks upon discharge
F f 53 x 137
I 206x 184
J 219 x I93
K 233 x 207
L 257x 231
Mactra (Spjsula) solidissima FIG. 14. Development of Mactra (= S p i ~ u h ) . Y O ~ i r l i 8 m W A from atraight-hinge Htage (A)
Meesurernentq of length arid width of larvae of different to metwnorphosis (L). stages are given in microns.
or upon stripping, thus rendering the egg ready for fertilization. Eggs of Crassostreu virginica and many other species belong to this category. In the other group stripped eggs continue to retain their

REARING OF BIVALVE MOLLUSKS 29
germinal vesicles intact and fertilization does not occur. However, as elready mentioned, eggs of such species can be chemically treated and thereby become ready for fertilizaticp.
As cleavage progresses and more micromeres are formed (Fig. 13e), the embryo gradually develops into a swimming, ciliated larva which eventually reaches trochophore stage (Fig. 13f ). Under favorable conditions this stage may be reached in 12 to 16 hr, depending upon the original condition of the eggs, culturing methods and, of course, water temperature.
During late trochophore stage (Fig. 139) the cell gland begins to secrete the shell. When the shell completely encloses the soft parts the larva has reached early straight-hinge stage (Fig. 13h).
Development of larvae of Mactra 8olidissima, from early straight- hinge stage until metamorphosis, and their length-width measure- ments during this entire period are shown in Fig. 14. Very early, normal straight-hinge larvae measure only about 79 p in length and usually between 63 and 65 p in width. In some cultures, composed mostly of abnormal individuals, somewhat smaller, slightly deformed straight-hinge larvae can be seen occasionally, but it is doubtful that they survive to metamorphpsis.
Individual larvae of M . solidissima display considerable variations aa to the size at which certain organs of their bodies begin to develop and at which metamorphosis occurs. For example, in some individuals the foot can be seen when they are only about 160 p long. Approxi- mately 80% of the larvae show a well-developed foot by the time they are 215 p long, and at a length of 240 p practically all possess this organ.
Disappearance of the velum is another step in larval $evelopment that is not strictly correlated with a definite size. In some larvae measuring only 219 p in length the velum wm already completely resorbed, while in extreme cases a diminishing, but still functional velum was seen in larvae about 257 p long.
A few larvae begin to metamorphose when they are about 220 p long, but the majority are between 230 and 250 p before metamor- phosis occurs. At this time the velum is resorbed, rudimentary gills develop, and a powerful ciliated foot, which when expanded is as long as the young clam itself, serves as the only means of locomotion. Individuals memuring 262 p in length were the largest true larvae recorded. In this respect our observations are in agreement with those of Imai et al. (1953), who found that in Mactra sachalinen& the foot begins to develop at a length of about 200 p and that larvae set at about 270 p.

30 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
B. Abnormal eggs and larvae Abnormal development of eggs and larvae of bivalve mollusks may
be due to any one of a variety of factors or to a combination of such factors. It is our practice, however, to discard cultures in which less than 50% of the eggs develop into normal straight-hinge larvae. This is done because batches of eggs giving a low percentage of larvae may be abnormal in some respects, and these abnormalities may lead to aberrant experimental results and wrong conclusions. We do not know, as yet, what factors are responsible for poor eggs and feeble embryos. In some instances abnormal larvae, or failure of eggs to develop to straight-hinge larval stage, may be the result of incom- patible genetic combinations. Our experience indicates, however, that such combinations are comparatively rare.
Some abnormalities of larvae may be ascribed, no doubt, to the poor physical condition of spawners. Several investigators have believed that eggs released late in the season were less viable and pro- duced less vigorous larvae than those from earlier spawnings. Loosanoff and Davis (1950) were under the impression that the last batches of eggs discharged by virtually spent females gave feeble larvae that grew slowly and showed high mortality. Cole (1941) offered evidence that the brood strength of Ostrea edulis may decline during the course of t i
breeding season and he believed that this was due to a depletion of food reserves in the bodies of parent mollusks. Walne (1956) thinks that the lack of " vigour " in larvae may result from poor condition of the parent oyster and believes i t possible that the vigour of larvae may be affected by the quantity of food reserves laid down in the eggs.
More recently, Davis and Chanley (1956b) have shown conclusively that the last batches of eggs of both clams, M . mereenaria, and oysters, C. virginica, discharged by virtually spent females were cultured with no apparent diminution of either percentage of eggs developing into straight-hinge larvae or rate of growth of these larvae. Accordingly, we now believe that abnormal or feeble larvae do not occur more frequently in later spawnings than in other spawnings throughout the season. As has already been mentioned, our experiments have shown that there is no correlation between viability of spawn and age of parents.
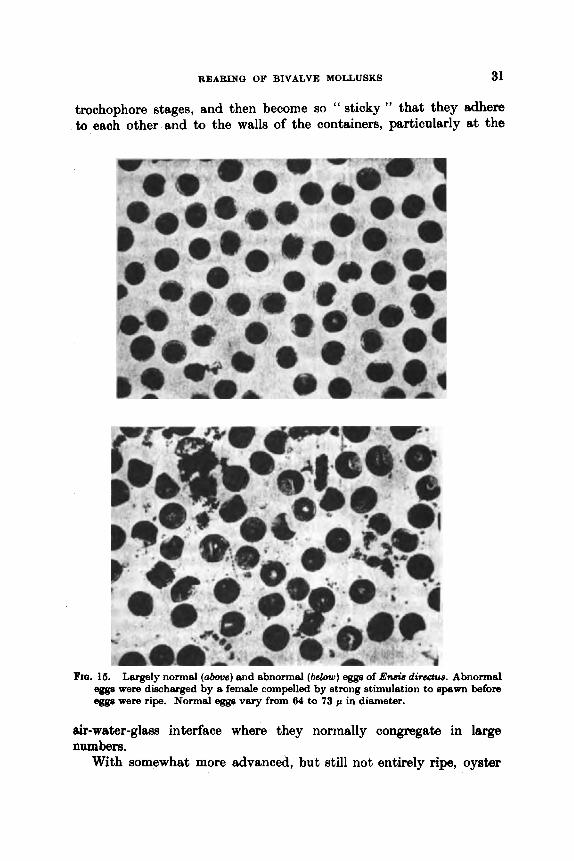
There is evidence that bivalves can be induced, by strong chemical and thermal stimulations, to abort eggs even though they are not fully ripe (Fig. 15). In some cases such spawnings appear to be quite normal and a large number of eggs may be released. More often, however, comparatively few eggs are shed. In the case of C. virginica immature oyster eggs usually develop only to late gastrula or early
REARING OF BIVALVE MOLLUSKS 31
trochophore stages, and then become so " sticky " that they adhere to eaoh other and to the walls of the containers, particularly at the
FIQ, . 16. Largely norm81 (above) and abnormal (below) egga of En& dir&w. Abnormal eegs were discharged by a female compelled by strong stimulation to 8p8w1-1 before eggs were ripe. Normal egga vary from 64 to 73 p in diameter.
air-water-glass interface where they norrnally congregate in large numbers.
With somewhat more advanced, but still not entirely ripe, oyster

32 VICTOR L. LOOSANOFE’ AND HARRY C. DAVIS
eggs the larvae develop more normally but are quite small, measuring only 60 to 70 p at the 48-hr stage (Oavis, 1949). Finally, in induced spawning of oysters late in the season after resorption of their gonads has begun, embryonic development of eggs is frequently abnormal and only a low percentage of them develop into healthy straight-hinge larvae.
Subjecting eggs and spermatozoa to temperatures higher than 30°C may injure or even kill them. Maintaining rcrcently discharged eggs in heavy concentrations, a condition that leads to formation of a thick layer of them on the bottom of rearing vessels, may result in a sufficient
Q
deplet’ion of oxygen and accumulation of catabolic products that will affect the eggs and their further clevalopment. l f zygote8 and early embryos are badly overcrowded, thoir shell develctpmcn t u,uually proceeds only as far as the shell gland Htage. ‘I’hun, inntr:ad of nhelln being fully formed 48 hr after fertilization, as owurn i n normal 1arvti.e which, at that time, can retract their soft tiotlien complc:tc:ly wi th in t h new shell, overcrowded larvae have a small, dark oval arc!& clsrroting the position of the #hell gland o r i i snirtll tirtntl of’ Hhdl mcttc:rittl r i o t more than t w i w ttic clianrc.tc!r of t h : ? i h c * l l ~I i~i l (1 .
Iii Ii~ss-cri iw(h~(1 c i i I t , i i r ( b H IILrviu. iimti ( ~ t i i i i i g I i H t i v I I ti) i w ( i t r i i ~ I c from t.lic*ir Iio(1ic-s give- t , l i c * ii.f~iii*ii,rii,iic’i. (11‘ ~ ~ i i r ~ l l wiriKH ( ” wiriK:c-cl Irrrvao ”). Uiidvr s o i i 1 i w h i L t t)c.i.tcr (:ot)cliLionx $1. I a r p r , h u t till iricornlileh xhcdl

REARING OF BIVALVE MOLLUSKS 33
is formed, but the hinge line, instead of being straight, is concave, characterizing “ saddleback larvae ”, or convex, typical of “ hump- back larvae ” (Fig. 16). In both of these abnormalities much of the ventral pcrtion of a larva’s body will extend beyond the shell. The results of overcrowding on development and growth of larvae will be more fully described in a later section of this article.
Occasionally, in some cultures many larvae have abnormally small vela. This abnormality may, sometimes, be due to mechanical injuries to the velum when larvae are screened before their shells are fully developed to protect the soft parts. In other cultures it has been aasociated with tho presence of numerous ciliates. It is possible that velar deformities in these c a m were the results of injuries by ciliates, but it is more probable that the ciliates were feeding on particles of vela cast off by larvae in response to adverse conditions, such aa wtificially-created concentrations of certain chemicals.
The same type of abnormalities, as observed in overcrowded cultures, occurs when eggs are cultured in sea water in which adult oysters have previously been kept. Probably because of the same reason, eggs carried along with water from tanks or trays in which a mass spawning has occurred seldom develop into normal larvae unless the original water is greatly diluted with fresh sea water.
Failure of larvae to develop normal shells when overcrowded, or when grown in water in which adult oysters had previously been kept, may indicate a depletion of certain substances, normally present in sea water, that are needed for shell formation. In experiments devised to verify this possibility some of the eggs discharged by a single female were placed in fresh sea water, while others, fertilized with sperm from the same male, were placed in set3 water taken from an aquarium in which adult oysters had been kept. The latter group of eggs gave a much lower percentage of normal larvae.
One interesting clms of abnormal larvae consists of those that do not feed, even though they do not show clear-cut anatomical malforma- tions. These apparently normal larvae, which developed to straight- hinge stage under our standard conditions and are kept in the same cultures with other larvae which are feeding and growing normally, seem unable to feed, do not grow, and eventually die., Before death, the larvae become emaciated so that most of the space inside their shells is empty with only the retractor muscles, a small velum and a shrunken visceral maas remaining. In some cultures this abnormality is found in more than 25% of the larvae.
In several experiments this type of abnormality has been associated with the kind of food given. In these instances approximately 50%

34 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
of larvae of M . mercenaria receiving Chlamydomonm sp. displayed this abnormality, while the remaining 50% fed and grew at a normal rate, as did all of the sibling larvae in other cultures receiving other foods. A similar phenomenon was observed in American oyster larvae fed Phaeodactylum tricornutum (Davis and Guillard, 1958).
While some anatomical abnormalities may interfere with the ability of larvae to gather or ingest food, resulting in poor growth and eventually death, other abnormalities, such as badly deformed shells, may still permit larvae to feed and grow. Sometimes, as they grow, such larvae gradually become more normal in appearance, but usually remain distinguishable even when they are nearing metamor- phosis.
Experimeiits on tolerance of eggs and larvae of bivalves to such factors as turbidity and salinity, and to chemicals, such as pesticides, antibiotics, and bacteriostatic compounds, have also shown that if any one of these falls outside of the tolerated limits, embryonic develop- ment becomes affected, resulting either in death of the zygotes or in abnormal larvae. These matters will be discussed in more detail later in the article.
Some of the larvae, particularly those of the clam, M . mercenaria, that are abnormal because of overcrowding, exposure to low tempera- tures or high turbidity during early stages of development often grow to metamorphosis if returned to favorable conditions and given good food.
We have frequently observed, during blooms of dinoflagellates in Milford Harbor, that in our laboratory cultrire~ only a Rmall prrc:rlntag:e of clam or oyster eggh d ~ ~ e 1 0 j ~ t : d inho rrormal ntmigtit tiirigc! Isrvrrc~. Eggs placed in, water from which dgae w m : rt$movcvl hy Millipor(! filters showed only a slightly higher rate of normal d~:vc:loj)rnr:nt ttiari eggs grown in unfiltered water containing dinoflagc:Il~~t~:~. Egg8 from the same spawning8 but cultured in sea water co1lectr:d prior h the bloom gave considerably higher percentages of normal IltrvrLc: ( IJavin and Chanley, 1956b).
It may IJC addod that plrtnkt,ori sarriplm co1l~ctc:tl i r i I ~ ) t i l r ; l h n d Sound t l u r i r i K o r iniawtliata~ly followitlg Iicvtvy dgnl t ) I o o r n H aro usiitLlly rliuructcrixcd I b y tho nciwiby or wcm complete abwnce of early straight- hinge stagex of bivalve larvae (hosanoff, 1958a). We believe that reduction in numbers of normally developing bivalve eggs and larvae ill t h a h v v iiistaiiccs is primarily caused by highly toxic metabolites of algno thst may pcrsist for several days after the blooms have ended. It is possible, however, that this phenomenon is due, at least in part,
Dense algal blooms may also cause abnormal development.

REARING OF BIVALVE MOLLUSKS 36
to removal by algae of certain chemicals from sea water that am essential @ k a l development.
C. Me&& of cultivdion of eggs and h m Methods of culturing eggs and larvae of bivalves under laboratory
and small-sde hatchery conditions have been tested by many workem for over 100 years. Costki, a Frenchman, was probably the first to attempt this mound 1868. In the United States a number of extremely capable men, including Brooks (1880), Ryder (1883) and Winslow (1884), continued these efforts on C. virginica, but were unsuccessful. Perhap the beef summary of theae efforts ie given by Winslow, who states, “ But after my experience of the peet spring and summer I am oonvinced that it will require a series of painstaking experimenfs, extending over oonaiderable time and conducted under many diesimilar conditioqs, before the artificiel production and culture of the oy&x is made a matter of prrtctitx.1 importance.”
Inter& in artificial propagation of bivalves was revived when Prytherch (1924) and Wells (1920, 1927) succeeded in carryhg oyster h a e to metamorphosis. This success wm probably due to the practice of renewing the water in which oyster larvae were kept. Wells used 8
milk separator for this purpose, while Prytheroh ueed filtros plates. Other suocessful workers in this field included Hori and Kusakabe (1926), Cole (1936), Bruce et d. (1940), Lindsay and Woelke (1960), Woelke (1960) and, especially, Imai et d. (1950b). In our,- some oysters were carried to metamorphosis as early as 1932, but efforts to repeat this succ8g~ usually failed until about 1946-1947 when we began to develop and improve the methods used at our laboratory (Loosanoff and Davis, 1960 ; Davis, 1963 ; Loosanoff, 1964).
It is our practice to fertilize eggs m soon a8 they am dhhrged. Usually it happens automatidy because since we use a sperm sue- pension to stimulate spawning, spermatozoa are already present in the water when females begin to discharge eggs. b h , actively-moving sperm are used to assure normal fertilization of eggs and development of zygotes. Sufficient quantities of sperm are always added, but when working with small eggs, such as those of C. virgin*, which cannot be retained even by fine screens, we limit the quantity of suspension. In this way excessive quantities of sperm are not carried into our culture vessels, and the undesirable effects of decomposing sperm on developing eggs are avoided.
The bivalves are usually spawned in Pyrex glass dishes containing about 1.6 litemof water(Figs. loand 11). As haealrdybeenmentioned, to separate the eggs from the debris and, later, from the excess sperm, 4

36 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
blood cells and body fluids accompanying the spawning, we use a series of stainless steel sieves with screens containing different numbers of meshes per linear inch (Fig. 17) . The finest screen that was found practical in our operation has mesh openings averaging 44 p, but since many mcahes arc actually larger and exceed the diameter of an oyster cgg, which is i ~ l i o i i t !iO p, 1riiiii.y qpi pn~n throuKh tho Hievc!. I t i H
true t h a t I)y using No. 26 bolting silk n smaller Hize meHh is available but in that case the openings are so small that they eaxily get clogged,
rt~ridi:riiig tl iv a i ( ~ w s u ~ i : l w n . As th rwi~lt , in our practice we UHC a Hcrics of N i w i L s , tjlw f i w n t of which h w H iiominal opening of about 44 p, followed, WJNW ~ic~cc~ssnry, t)y an!, of thv coarser screens with openings 0 f 5 3 , ti2, 74, t(H, 120, 126, 14!) nlid 177 p.
In spccies having eggs too small to be retained even by our finest screen, the eggs can be partially freed of body fluids, sperm, etc., by letting the eggs settle on the bottom of a dish and then syphoning or decanting most of the fluid. By repeating this procedure several times most of the undesirable substances t,hat are dissolved or suspended in the water will he discarded.

REARINU O F BIVALVE MOLLUSKS 37
We have used a variety of containers to culture larvae. Some of them were large glass vessels, including Downing and McDonald jars used in fish hatcheries for incubation of semi-buoyant eggs, lobster jars, 5-gal earthenware jars, and 76-gal polyethylene and Fiberglas containers. We have also grown larvae in large, outdoor, concrete tanks containing several thousand liters of sea water (Loosanoff, 1954). In all these instances the larvae were grown successfully.
For precise experimental work Pyrex glass beakers of 1000- to 1500-ml capacity are perhaps the most satisfactory because they are not toxic and are readily cleaned and sterilized (Fig. 5) . Polyethylene and other plastic containers are also convenient and non-breakable and some can be sterilized. However, some of them are permeable to certain insecticides and, perhaps, to other substances and are known to adsorb a variety of toxins. Because of these considerations poly- ethylene and some other plastic containers, while convenient as culture vessels, cannot be used in experiments involving certain toxic sub- stances, such as insecticides.
New, soft glass vessels may contain substances which are toxic to eggs or larvae of oysters and clams. Even though these vessels are conditioned in sea water for Reveral days, culturing larvae in them is always haphazard because, although larvae in different vessels are presumably grown under identical conditions, their rates of growth are often distinctly different and they show mortalities unrelated to the treatment. A t the same time sibling larvae grown in earthenware jarsor Pyrex glass containers suffer no mortality and different cultures receiv- ing the same treatment show good duplication in their rate of growth.
Although we could not identify the substances responsible for poor growth of larvae gro;wn in soft glass jars, we found, nevertheless, that young bivalve larva, especially those of C. virginica, are sensitive to presence in the water of e&n minute quantities of certain chemicals. For example, we noticed that washing of glassware and other imple- ments with tap wa'ter that passed through a pipeline containing copper unfavorably affected larval development. Apparently, even minute quantities of these metals are sufficient to interfere with normal development of eggs and larvae.
Our experience in growing bivalve larvae has ehown that they cannot be kept in recently-built concrete tanks and that, usually, i t is necessary to age the tanks with sea water for a long time before this can be done.
Since most of our studies are quantitative, as well as qualitative, definite numbers of eggs or larvae per ml of culture are needed from the start. This is achieved in the following manner: eggs are placed

38 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
in a tall, narrow glass jar and the water in it is thoroughly agitated with a perforated plastic plunger to distribute the eggs uniformly. A sample is then taken with a volumetric pipette, and the number of eggs or larvae per ml is determined by counting them on a Sedgwick- Rafter cell. After that the eggs or larvae are again agitated, to ensure their even distribution in the vessel, and the necessary volumes of water carrykg larvae are transferred to culture vessels.
We usually begin our experiments with 10 000 to 15 000 straight- hinge larvae per liter of sea water. However, because not all fertilized eggs develop into normal larvae, i t is our practice to piace approxi- mately twice this number of eggs into each culture vessel to produce a sufficient number of larvae. Accordingly, about 30000 eggs per liter are used in starting cultures. These eggs are placed in containers filled with 8ea water that is first filtered through an Orlon filter and then subjected to the sterilizing action of ultraviolet rays. The eggs are then left to develop undisturbed for 48 hr and no food is added during this period.
All culture vessels are immersed in a common water bath table, the temperature of which is controlled within f I.O"C (Figs. 5 and 6). Usually, neither aeration nor mechanical agitation is employed because we have established that clean, well-attended cultures do not require aeration if the water is changed every second day. In special experi- ments requiring mechanical agitation we use a number of devices, including a rotating wheel, paddle-agitator (Fig. 51, or regular shaking machine.
At the end of 48 hr young larvae, now protected by fully formed shells, are collected by screening the cultures through sieves having 325 meshes per lineal inch (44 p opening). Larvae retained by the screen are gently washed and placed in a tall jar. Using the same method as that employed with eggs, the number of larvae per ml is determined, and the appropriate volumes of water containing larvae are placed in each culture jar to create desired concentrations.
Samples of larvae are taken whenever needed, usually a t 2-day intervals. This is again accomplished by collecting all larvae from a container on a 325-mesh screen and then transferring them to a graduated cylinder of I-liter capacity from which, after proper agitation, required samples are taken, while the remaining larvae are returned to the culture vessel.
D. Larval period
morphosis is affected by many conditions. Rate of growth of veligers from straight-hinge stage to meta-
In our laboratory work

REARING OF BIVALVE MOLLUSKS 39
the chief controlling factors have been food and temperature. The role of these conditions will be more fully discussed in special sections later on, and also in the sections dealing with development and growth of larvae of different species ; therefore, here it will be sufficient to mention only general observations.
Our experiment8 have shown that larvae of different bivalves display different food requirements (Loosanoff and Davis, 1951). Until they reach a length of about 125 p larvae of C. virginim, for example, are quite restricted in types of food they can utilize (Davis, 1953). Certain naked flagellates are the only organisms, thus far tested, that may be included in this category. Chlorella is one of the many genera of algae having thick cell walls that oyster larvae either cannot utilize or utilize only to a very limited extent during early stages, although i t seems to be quite a satisfactory food for older larvae (Davis and Guillard, 1958). Thus, if during the early stage of development of oyster larvae specific food organisms are either entirely absent or are uncommon, the larval free-swimming period may be greatly prolonged or the larvae may never reach metamorphosis. For example, during our earlier efforts of raising larvae of C. virginica, when little was known about their food requirements, approximately 50 days were required before the most advanced individuals began to metamorphose in some cultures. In similar cases, with larvae of C. gigas, the cultures were discarded after 53 days because the largest larvae at the time measured only about 100 p. Now, using good food organisms, such as Ieochryeis galbana and other naked flagellates, and mairitaining the temperature at about 23"C, larvae of C. iiirginica have been reared to metamorphosis in our laboratory in 18 days. This is, probably, the approximate time required by larvae to grow to setting size under natural conditions in Long Island Sound (Loosanoff and Engle, 1947). At 30°C well-fed oyster larvae, grown under laboratory conditiong, began to metamorphose 10 days after fertilization.
The importance of the second factor, water temperature, on length of larval period of bivalves has also been well demonstrated in our etudies of larvae of M . mercenaria (Loosanoff et al., 1951). These studies showed that, under identical conditions, larvae kept at a tem- perature near 30°C began to set as early as the 7th day afkr fertilization, while cultures maintained at 18°C contained the first metamorphosing individuals only after 16 days.
In the caae of larvae of all the species we reared it has been clearly demonstrated that even though larvae originate from the same spawning and, sometimes, from the same parents, and are kept in the same vessel under identical conditions, individuals grow at widely

40 VICTOR L. LOOSANOFF A N D HARRY C. DAVIS
different rates and, therefore, metamorphose at different times (Fig. 18). For example, in a recent experiment a healthy culture of larvae of c. virginicn fed a mixture of I . p l b a z n and Monochrpis lutheri and kept at, about ?:3.R"C hcgnn t,o set 1 H (jays af%cr f+rtilizntion. Setting gradually increased in intcnsity :tnd remained heavy for the first 17 days, but some larvae continued to swim, before metamorphosing, for another 10 days. Thus, setting of this, presumably, homogeneous culture continued uninterrupted for it period of 27 days.
A number of similar observations on larvae of M . mereemria obtained from the same parents and grown under identical conditions, but showing considerable individual variations in rate of growth and in
tinit? 11c~ctlt!d to rcwh meta~norphoui~. can be given. Perhaps the most detailed description of this phenomenon appears in the paper of Loosanoff et al. (1951) on growing clam larvae at five constant but different temperatures. These authors gave the minimum and maxi- mum sizes of larvae recorded in each culture every 2nd day from time of fertilization until the majority of the larvae metamorphosed. During the early life of a culture, on the 2nd or 3rd day, the larvae differed in size by only a few microns, but swcral day8 1atf:r. ckpf:ridirig i i p n the temperature, the size ranged from mall Ntraight-hirip Iltrvw of approximat.eIy 100 p long to f ' i ~ l l grown, rcrufy to r r i ( ~ t , ; ~ r r i ~ ~ r i l t i ~ ) ~ ~
individuals. For examplc, ~ j z r : ~ r ~ f ' ja,rv;w grown at. 30''f; rmigwl o r 1 thrt 8th day from 107 to 2'Lf) 11. I J J IL ClJkltJ'r! k i q h tit, 21''(; t h * rnitlirnurn ;$rid

REARING OF BIVALVE MOLLUSKS 41
maximum sizes recorded on the 2lst day were 107 and 221 p. Our colleagues working in the same field, especially Imai et al. (1954), fully share our experiences.
At present, no well-based explanation can be advanced for these differences. Perhaps, as has been suggested in connection with survival and growth of certain fish, vitality of the individual eggs and larvae that emerge from them depends to some extent upon the position of the eggs in the ovaries and the amounts of nutritive materials that have been stored in the individual eggs before they are discharged.
Chanley (1955) assumed that differences in sizes of larvae in the same cultures must be due, a t least in part, to inherited characteristics. He also reported evidence of significantly different rates of growth of larvae originating from eggs of the samt' female crossed with different males and larvae grown from eggs of two females individually crossed with the same male. He tentatively concluded that inherited differences from either parent may he responsible for differences in rate of growth of different larvae.
In some cultures, especially those kept at comparatively high temperatures, the range of larval sizes usually diminishes several days after beginning of metamorphosis. This is due to the disappearance of larger individuals because of setting and, partly, because abnormal, undeveloped, slow-growing larvae are rapidly dying.
There were periods in our practice, for example, in growing larvae of Ostrea edulis, when regardless of all efforts they would not grow at all or ceased growing soon after reaching a size of about 220 p. The reasons for cessation of growth still remain an enigma because, at times, these larvae refused to grow wen when given foods on which, in previous experiments, they grew well.
E. Hardiness of eggs icnd larvae According to Nelson (1921) larvae of C. virginicu are extremely
sensitive to a sudden change in water temperature. A drop of only 3" to 5°C within 24 hr may be followed by the disappearance of a majority of the larvae. According to the same author rain storms, as well as strong winds, cause death of large numberu of bivalve larvae. Nelson, however, failed to offer experimental evidence to support his contention of the unusual sensitivity of bivalve larvae to relatively minor changes in their environment. Our observations, reported partly in this section and partly in the sections to follow, lead us to disagree with Nelson's point of view because they have clearly demonstrated. that bivalve eggs and larvae, if protected against disease-causing organisms and toxic substances, are rather hardy.

42 VlCTOlt L. LOOSANOFF A N D HARRY C . DAVIS
Laboratory and field observations lead us to believe that oyster eggs that are still in the ovaries are hardy and capable of withstanding sharp physical changes in their environment. For example, on several occasions oysters with mature gonads have been kept for various periods of time in the refrigerator at about 2°C to delay their spawning. Some of them, kept at this low temperaturq for 7 days, have spawned copiously later on, when subjected to proper stimulation, and larvae from these spawnings have been reared to metamorphosis. Other groups kept in the refrigerator for 15 days also spawned normally and pro- duced healthy larvae. However, oysters that were kept in the refrig- erator for 30 days spawned feebly, and only a portion of the eggs developed into normal larvae. This semifailure was probably due to severe dessication of the oysters and their gonadal tissue. This conclu- sion is supported by the observation that the best spawnings occurred when refrigerated oysters, prior to attempts to spawn them, were kept in running sea water at room temperature for at least 6 hr. During this recuperation period they probably restored their water loss.
Another experiment on effects of low temperature upon ovarian eggs of C. virginica of Long Island Sound was performed only last winter (1961-62). Oysters measuring from 3 to 6 in long were brought into the laboratory early in January from their natural beds and placed in conditioning trays to be ripened for spawning. After conditioning at about 20°C for about 1 month, these oysters, now ripe, were trans- ferred to outdoor tanks where the water temperature was near 0°C and where, at times, a layer of ice \$as formed.
Twenty days later, on 26 February, the first group of oysters was returned to the laboratory and placed i11 water of the same temperature as that outdoors. Then the temperature was lowly raised to ahout 17OC for 2 days. Following this recovery period fiftcen oy.rtr:rx were placed in spawning dishes and our usual method of iiidueing spawning was applied. Eight of fifteen oysters responded, of which five were females and three were males. Bpawnings were light to medium with a total of 27 million eggs released. These eggs were cultured by our usual method, but only a comparatively small number of larvae developed to straight-hinge stage.
On 6 March another group of the oysters was brought into the laboratory from the outdoor tanks and later induced to spawn. Four out of eight oysters spawned, three of which were females. The total number of eggs discharged by these females was 57 800 000. The majority of the eggs were normal in appearance, although a few were deformed and some were small.
I n one of the containers, in which 375000 eggs from this spawning

REARING OF BIVALVE MOLLUSKS 43
were placed in 5 gal of water, a count of straight-hinge larvae was made 48 hr later and showed that the culture contained 165 000 normal, straight-hinge larvae, 11 000 abnormal ones and 2 000 dead individuals. Therefore, approximately 168000, or about 4,50/, of the eggs placed in the culture developed into larvae. 111 another culture, whero approxi- mately 7,50000 of those eggs worc plucod in 10 gal of water, t h o count made 48 hr later gave 424000 normal, 46 000 abnormal and 32 000 dead larvae; therefore, approximately 502000, or 61% of all the eggs that were originally placed in the culture developed to straight-hinge stage. From then on, however, larval development was poor, showing high mortality.
On 19 March, 43 days after the ripe oysters were placed in the icy water, another group was brought in and, after being kept for 4 days in running sea water at a temperature ranging between 14" and 17'C, was induced to spawn. Both males and females spawned. One of the females released 18 million and another, 27 million eggs. Larvae obtained from eggs of one of the females grew well, increasing approxi- mately 10 p in length per day.
This experiment demonstrated the remarkable fact that oysters artificially ripened in the middle of winter can be transferred abruptly from warm to freezing water, retained there for over 40 days, and then returned to warm water and induced to spawn, producing viable eggs and sperm that eventually develop into normal straight-hinge larvae. Even though mortality among larvae obtained in this unusual manner was relatively high and many larvae were abnormal, the, experiment demonstrated, nevertheless, the remarkable power of oysters to retain their ripe sex cells under extremely adverse conditions. Results of histological studies of gonads of oysters involved in these experiments will be described later in a special publication.
Regardless of the ability of ovarian eggs of C. virginicu to withstand exposure to low temperatures for long periods, recently fertilized eggs, in the polar-body stage of development, do not display the same tolerance. This was demonstrated by an experiment in which eggs, within 1 or 2 hr after fertilization, were placed in a refrigerator main- tained at a temperature of about 2'C, and kept there for 6,24 and 48 hr. Samples were then returned to room temperature and further develop- ment of the eggs subsequently observed. In all samples a few eggs developed into abnormal ciliated blastulas, but practically all of them failed to develop further and soon disintegrated.
Healthy shelled larvae of oysters, C. virginim, are, nevertheless, capable of withstanding sharp changes in temperature of the surround- ing water. In a special series of experiments, designed to verify Nelson's

44 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
conclusion of extreme sensitivity of these larvae to temperature changes, beakers of 1-liter capacity containing larvae grown at a tem- perature of about 22°C and measuring about 200 p in length were placed in a refrigerator at 2°C and returned to room temperature following 6, 12 and 24 hr of refrigeration. Within a few hours the predominating majority of larvae exposed to the low temperature for 6 and 12 hr were swimming and feeding normally. However, many of the larvae which were refrigerated for 24 hr lost a portion of the velum and eventually died. The mortality in each of the above groups at the end of one week after return to room temperature was: control, 4.2%; 6-hr chilling, 6.5%; 12-hr chilling, 4.9%; and 24-hr chilling, 44.9%. Thus, even though exposed to a near freezing temperature for a 24-hr period, more than half of the larvae survived and continued to develop.
In still another experiment oyster larvae lived and grew when subjected every 48 hr to a sharp drop in temperature, from 20" to 10°C, for periods of 15 to 30 min, followed in a few minutes by an equally abrupt return to 20°C. A majority of these larvae subsequently reached metamorphosis. It would seem unlikely, therefore, as claimed by Nelson (1921), that ordinary short-term temperature fluctuations of only a few degrees, occurring in natural waters, could be responsible for an appreciable, sometimes total mortality of larvae.
It is also certain a t this time that bivalve larvae may survive long periods with Iittle or no food. In many of our experiments several control cultures survived from 2 to 3 weeks with little or no mortality, even though they did not receive any food except that which was present in the filtered sea water where they were kept. Moreover, in our earlier experiments, before such good food forms as naked flagel- lates became available, many oyster larvae cultures were kept for more than 40 days, although they did not show any growth. As already mentioned, in some of these cultures setting began only after 50 days. These observations demonstrate that bivalve larvae may tolerate comparatively long periods of semi-starvation and some may even reach setting size and metamorphose regardless of poor feeding condi- tions. It is improbable, therefore, that under natural conditions larval populations of such mollusks as oystms will die within 2 or 3 days because of a lack of sufficient quantities of Sood. It is clear, however, that lack of food will prolong the larval period, thus increaaing the loss of larvae because of predation and dispersal.
Larvae are also able to tolerate very low oxygen concentrations, at least for short periods. For example, on several occasions a number of

REARING O F BIVALVE MOLLUSKS 45
larvae were accidentally left overnight in a small pipette of sea water, yet they were found alive and healthy the following day.
Recent studies of Davis (1958) have clearly demonstrated that eggs and larvae of at least some estuarine species, such aa C . virginicu, can endure sharp changes in salinity. This matter will be discussed more extensively in the section dealing with the general aspects of changes in salinity upon development of eggs and larvae.
Studies of effects of turbidity upon eggs and larvae of C. virginicu (Davis, unpublished) and those of M . mercenaria (Davis, 1960), which will be discussed later in greater detail, have demonstrated that larvae of these two species can endure and even continue to grow in water that is quite turbid. For example, Davis has shown that larvae of C. virginica may survive for a t least 14 days in a concentration of 2 g of silt per liter of sea water. Such a heavy concentration seldom occurs in nature.
Fertilized eggs and larvae of many bivalves can also withstand vigorous mechanical disturbances without ill effects. For example, to obtain a representative sample of the population from our culture vessels, the water is strongly agitated by means of a plunger to assure a homogeneous distribution of larvae. Such relatively rough treatment, usually performed every day or every second day, does not cause an increase in mortality or decrease the rate of growth of larvae. Observa- tions on the behavior of larvae in nature also support this conclusion because, as shown by our studies of plankton samples and by observa- tions on intensity of setting of oysters on natural beds, i t has been definitely established that strong winds accompanying New England hurricanes and churning the water of Long Island Sound steadily for several days do not noticeably diminish larval populations. This was especially well demonstrated in August 1955, when a marked increase in intensity of setting of oysters occurred immediately after hurricane (( Connie ”. This increase continued for 2 weeks, thus showing that larvae of all ages survived the hurricane. It is also of interest that setting of oyster larvae occurred during the hurricane, thus indicating that strong water turbulence does not easily destroy larvae or seriously interfere with their metamorphosis.
Recent studies have repeatedly demonstrated the sensitivity of bivalve larvae to traces of certain substances in the water. These observations showed that sea water, in which our larval cultures are grown, sometimes contains substances, so far unidentified, which determine whether larvae will grow normally (Loosanoff et al., 1951 :
1953). Wilson (1951) found similar differences between natural sea water collected from widely separated areas of the ocean. We are

46 VICTOR, L. LOOSANOB’E’ A N D HARRY C . DAVIS
still not certain whethcr it is the presence of deleterious materials or absence of growth-promoting siibstances in sea water that, slows growth or prevents normal development of larvae.
W’e have observed that somc substances which interfere with normal development of larvae may originate from sources to which we have previously paid little attention. Under certain conditions these substances may be released mechanically from bottom soil. This was noticed during a winter when a deep channel was dredged in Milford Harbor, from which our laboratory obtains its water. During that period the water acquired certain properties whch strongly interfered with normal development of eggs and larvae. These sub- stances were apparently in solution or in fine colloidal suspension because they were still present in the water after i t was filtered. Neither aeration nor ageing appreciably improved the quality of the water.
Sensitivity of eggs and larvae to different substances dissolved in sea water was further demonstrated by Davis and Chanley (1956b) in a series of experiments which showed that, while low concentrations of antibiotics may increase rate of’ growth of larvae, even a slight excess of them reduces rate of growth. Progressively increasing con- centrations of these substances corrrspondingly decrease rate of growth of larvae and eventually cause their mortality. This mattcr will be discussed in greater detail in the section devoted to larval diseases and their treatment.
Recently. extensive studies on effects of r~umerous insecticides, weedicides, oils, organic solvents and detergents on mollusks have been undertaken at Milford Laboratory. While these studies are still in progress, it has already been found (Davis, 1961) that within each group of these compounds there are great differences in toxicity of individual chemicals to eggs and larvae of bivalves. For example, DDT was found to be one of the most toxic of the commonly used insecticides because even at a concentration of 0.05 parts per million i t caused almost total mortality of oyster larvae. On the other hand, another common insecticide, Lindane ( 1 , 2 , 3, 4, 5. 6 hexachlorocyclo- hexane), even at a concentration of 10 ppm, which is essentially a saturated solution in sea water, caused no appreciable mortality of larvae. On the contrary, growth of clam larvae in 5 ppm of Lindane was somewhat faster than that of larvae in control cultures.
Certain concentrations of phenol, chloramphcnicol and Dowicide “A”, among the antibiotic, bactericide and disinf‘cctant compounds a180 appreciably improved rate of growth of bivalvc: larvae. Thiu iu attributed to the action of these compounds which inhibits growth of

REARING OF BIVALVE MOLLUSKS 47
bacteria toxic to larvae. Other compounds which, in certain concen- trations, probably improve rate of growth of larvae by partially inhibiting growth of toxic bacteria are acetone and trichlorobenzene among the organic solvents, Monuron and Fenuron among the weedi- cides, and Guthion among the insecticides. Davis (1961) appropriately suggested that, in some phases of shellfish culture, a sufficient concen- tration of such insecticides as Lindane may be maintained to destroy all undesirable crustaceans, while not affecting growth of bivalve larvae or their food organisms.
Our laboratory and field observations have shown that metabolites released by some microorganisms, especially dinoflagellates, seriously affect not only adult bivalves (Loosanoff and Engle, 1947), but also development of their eggs and larvae (Loosanoff et al., 1953). Such toxicity of external metabolites and their physiological effects on aquatic organisms have been recognized by many biologists, some of these studies having been summarized by Lucas (1947, 1961) and Korringa (1952). More recently, Loosanoff (1955) reported that a heavy bloom of dinoflagellates in Milford Harbor caused abortion of embryos and immature larvae of gravid European oysters, 0. edulis. Davis and Chanley (1956b) found that a dense bloom of dinoflagellates caused abnormal development of eggs and larvae of the clam, M . mer- cenaria, and oyster, C . virginica. Under these conditions only a few developed into shelled veligers. During that summer concentrations of dinoflagellates in some areas of Milford Harbor were as high aa 300000 cells per ml. Placing eggs of clams or oysters in this water, even after it was passed through a Millipore filter to remove dino- flagellates, resulted in only a slight increase in the percentage of clam or oyster eggs that developed normally.
Although we assume that the effectH noted above were due to external metabolites emitted by dinoflagellates, it is possible that they were caused by removal from sea water, by these cells, of certain substances necessary for normal development of clam or oyster eggs and larvae. Another possibility is that the presence of a certain sub- stance, favoring rapid growth of dinoflagellates and preventing normal development of larvae, was simultaneously responsible for both phenomena.
F. Effects of temperuture on cggs and larvae
Certain observations and experiments devoted to studies of effects of sudden and extensive changes in temperature on eggs and larvae of several bivalves have already been described in the preceding section.

Here, we shall briefly discuw the results of observations on effwts of temperature within a much more limited range.
Larvae of most of the species cultured at our laboratory were grown under routine conditions, i.e. at room temperature, which was normally near 20°C. Because of this no extensive information is available as to temperature ranges within which larvae of different species may survive or their optimal growing temperatures. I n a few species, nevertheless, rather extensive observations on effects of temperature on development of their eggs and on growth of the larvae were under- taken. Studies of this nature on M . mercenaria and C. virginica have been the most complete.
M . mercenaria has been grown from egg to metamorphosis at con- stant temperatures ranging from 18" to 30°C (Loosanoff et al., 1951). If, within 3 hr after fertilization, eggs of these clams were placed in water of 15"C, virtually none of them developed normally to straight- hinge stage. If eggs were kept a t room temperature from 6 to 9 hr after fertilization and then subjectod to a temperature of about 15"C, some developed into straight-hinge larvae. The majority of these larvae, however, were abnormal and many of them Boon died, although some continued to grow at a very slow rate.
If eggs and, later, larvae developing from them were kept at room temperature for the first 2 days after fertilization, until straight- hinge stage was well formed, and then placed in water of 15"C, some of the larvae survived for 12 days or even longer. It is possible that if given good food, some of the individuals might eventually reach metamorphosis. However, if larvae grown at room temperature for the first 2 days of their existence were placed in water of 10°C (Fig. 19), they would not grow.
Bt the other end of the temperature range, at about 33"C, abnormal development and heavy mortality usually occurred if recently fertilized eggs were transferred to water of this temperature. However, if eggs and, later, larvae developing from them were kept at room temperature for the first 48 hr after fertilization and then transferred to water of 33"C, rapid normal development, similar to that observed in cultures kept at 30°C followed. Thus, our observations on development of eggs and growth of larvae of M . mercenaria a t temperatures from 15" t o 33°C support the view expressed by Pelseneer (1901) that normal early cleavage stages of molluscan eggs are limited to a narrower temperature range than can be tolerated by more advanced stages of the eggs or larvae.
Larvae of M . mercenaria, developing from eggs within the tem- perature range of 18" to 30"C, grew to metamorphosis, growth being
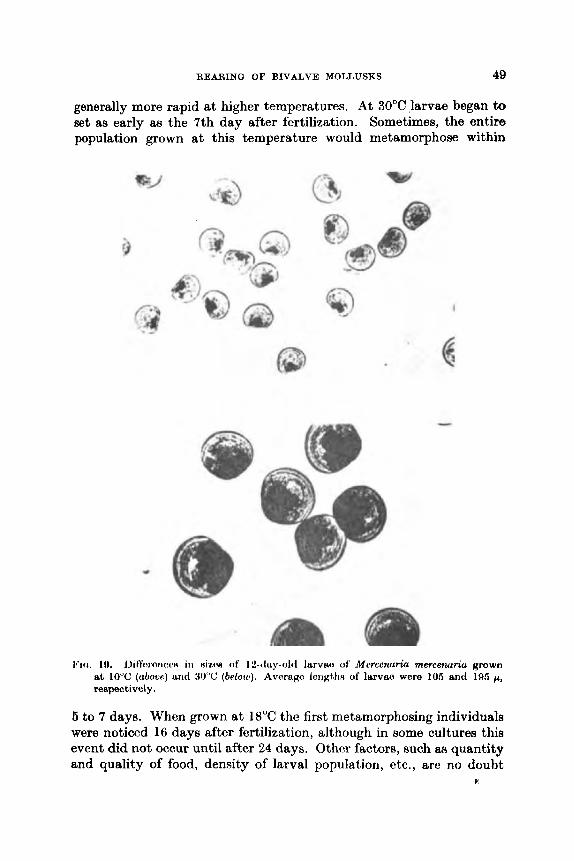
REARING OF BIVALVE MOLLUSKS 49
generally more rapid at higher temperatures. A t 30°C larvae began to set as early as the 7th day after fertilization. Sometimes, the entire population grown a t this temperature would metamorphose within
la'i(&. 1% Ihfforoii(-tw iu H i w R of 18-tiay-old Iarvtw of Mrreencirda merceruiriu grown at 10°C (ubovr) wid 30°C (below). Avorago lorlgth of lervao were 105 and 195 f i , respectively.
5 to 7 days. When grown at 18°C the first metamorphosing individuals were noticed 16 days after fertilization, although in some cultures this event did not occur until after 24 days. Other factors, such as quantity and quality of food, density of larval population, etc., are no doubt
Y
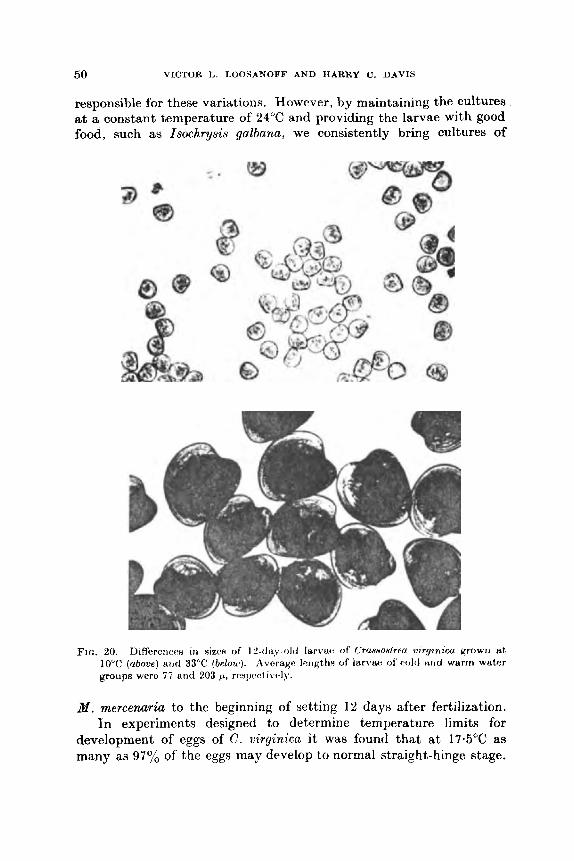
50 VICTOR L. LOOSANOFF A N D HARRY C. DAVIS
responsible for these variations. However, by maintaining the cultures at a constant temperature of 24°C and providing the larvae with good food, such as Isochrysis galbana, we consistently bring cultures of
M . mercenaria t o the beginning of setting 12 days after fertilization. In experiments designed to determine temperature limits for
development of eggs of C. virginicu i t was found that at 17.5"C as many as 97% of the eggs may develop to normal straight-hinge stage.

REARING OF BIVALVE NIOLLIrSKS 51
At 15"C, however, none of the eggs roached this stage although a few developed as far as early shelled larvae.
In some experiments 100% of recently fertilized oyster eggs trans- ferred directly to 30°C developed into normal straight-hinge larvae, but, a t 33"C, only 48% or less reached th is stage. The abnormal larvae of this group were unable to feed or grow even when returned to tx
temperature of 24°C. Although 2-day-old larvae placed in water a t constant tempera-
tures of 10" and 15°C for 12 days did not grow (Fig. 20), their rate of mortality during this period was comparatively low. The larvae kept at 10°C for 12 days could not feed even after being returned to a temperature of 24°C. However, some of the larvae kept a t 15°C for the same length of time and then returiied to 24°C fed, but their growth was negligible. Larvae kept a t 17-5"C took some food, but also showed little growth. The majority of these larvae, however, began to grow rapidly when returned to water of 24°C.
At temperatures of 20°C and higher growth of oyster larvae was, to a large extent, dependent upon the food given. When fed Chlorellu sp. (580), which is a relatively poor food, the larvae grew less rapidly than they did at the same temperatures when given better foods. Nevertheless, even when fed Chlorellu sp. growth of larvae within the range from 20" to 33°C increased progressively with each increase in temperature.
Recent experiments suggest that one of the ways in which low temperature may affect growth of bivalve hrvae is through inactivation of certain enzymes. For example, clam larvae kept a t '10°C can ingest food organisms but are apparently unable to digest them. This is well shown in the upper photograph of Fig. 19. Larvae kept at 15°C can digest and assimilate naked flagellates and grow slowly, but are unable to utilize Chlorellu sp. Those kept at 20°C were able to utilize both the naked flagellates and Chlorellu sp. Similarly, larvae of C. virginica kept a t a temperature of 20°C or lower cannot utilize Chhel la sp. How- ever, at 25°C these larvae receiving Chlorelh sp. showed some growth and at 30°C they were apparently able to utilize ChloreUa sp. much more efficiently and, as a result, grew quite rapidly.
Larvae receiving Dunaliella euchlora, a moderately good food organism, showed a sharp increase in growth bvtween 20" and 25°C. However, within the temperature rangt: from 25" to 33°C the rate of growth remained virtually the same.
Larvae given a mixture of our best food organisms, M . lutheri and I . galbanu, together with Dicrateria sp. (BII) and Chlorellu sp. (580), grew better at the same temperatures than when fed only ChloreEh
S

52 VICTOR I,. LOORANOFF A N D HARRY C. D A V I S
sp. (680) or D?LnaZidln euchloru. I n gcwrritl, 1,1~~,1lg]1 t h rangt from 20" to 3 0 T growth incrcastcl paraIIc~1 with tho iticrcvisc* iri tcmpcrature. At 30" and 33"C, however, tho Iarvirc. grc'w virtually th(. samt: and meta- morphosis at both temperatures bcgari consistently between the 10th and 12th days.
G. Effects of salinity on eggs and larvae
Bivalves, even though they belong to the same class of mollusks, display extremely wide differences in their salinity requirements and in ability to withstand sharp or gradual changes in salt content of sea water. Therefore, in determining minimum, maximum and optimal salinities for their existence each species, especially those populating estuarine regions, must be studied individually. For example, deltas of rivers, where salinities are relatively high most of the time, may be populated by both C. virginica and M . mercenaria, while a short distance above this line, where the salinity of the water is consider- ably lower than 20 parts per thousand, only oyster beds can be found because clams are unable to survive under such brackish conditions. To demonstrate the differences that, may exist betwecn two upecies that often may be found in the samc cwvironment, we HhalI hiefly tliscum the diffcrencos in salinity rcquircmrntR of larvae of (,'. virginicn and M . mercenaria.
Loosanoff (1952) found that the lowe& salinity at which normal development of gonads of C. virginica of Long Island Sound max proceed is near 7.5 ppt. Continuing the study of various aspects of variations in salinity on propagation of American oysters, Davis (1958) demonstrated that 22.5 ppt was the optimum salinity for development of eggs of oysters that had grown in Long Island Sound and had developed gonads at a salinity of about 27 ppt. Some normal larvae developed, nevertheless, in salinities as low as 15 ppt and as high as 85 ppt. At salinities below 22.5 ppt the percentage of eggs that deve- loped to straight-hinge larval stage steadily decreased until, at 15 ppt, only 50 to 60% of the eggs developed normally. At 12.5 ppt practically none of the eggs developed into normal shelled larvae.
In another experiment Davis used Maryland oysters that had grown and developed gonads in the upper parts of Chesapeake Bay where the salinity, at the time the oysters were collected, was only 8.7 ppt. These oysters were spawned at Milford Laboratory in Salinities of 7.5, 10 and 15 ppt. Under these conditions some eggn developed into normal larvae even at 10 ppt and 7.5 ppt although, i n the latter, slightly smaller than normal larvaca were common. In ganeral, the optimal salinity for normal devetopment of eggs of these oysters from

very brackish water was bctwccn 1% aiid 15 ppt, while a Ralinity of 22.5 ppt was the upper limit.
When oysters that had developed gonads a t a salinity of 27 ppt were used as parents, the optimal salinity for growth of their larvae, after they had reached straight-hinge stage, was 17.5 ppt. Good growth was also recorded a t a salinity of 15 ppt, but at 12.5 ppt growth was appreciably slower, although some larvae grew to metamorphosis. A t 10 ppt growth was practically at a standstill and i t is doubtful that any larvae could reach setting stago a t this salinity. The older the larvae, however, the better they withstood the salinity of 10 ppt. Larvae that were reared almost to setting stagc: a t our normal salinity of about 27 ppt continued to grow and even metamorphosed when transferred to a salinity of only 10 ppt.
Davis (1958) also showed that the optimal salinity for development of eggs of &I. mercenaria of Long Island Sound was about 27.5 ppt. No normal larvae developed at salinities of 17.5 ppt or lower. The upper salinity limit for development of clam eggs was 35 ppt, but only an occasional normal larva developed a t {,hat concentration of ualt.
Straight-hinge clam larvae grew reasonahly well a t 17.5 ppt and many reached metamorphosis, but at 15 p1)t none of them reached that stage, although some lived for 10 or more days and showed a slight increase in size. At 12.5 ppt straight-hinge clam larvae showed no growth and all were dead by the loth day.
As can be seen from this brief comparison, eggs and larvae of C. vir- ginica can normally develop and grow to metamorphosis in a much lower salinity than those of M . mercenaria. Undoubtedly, using present methods of cultivation of larvae, similar studies will soon be performed on other species of bivalves and prove to be as informative and useful as those reported in the recent article by Davis and Ansell (1962) on development of eggs and growt$ of larvae of 0. edulia in water of different salinities.
H. Effects of turbidity o n eg:p and larvae One of the least studied factors of molluscan environments is that of
turbidity (Loosanoff and Tornmers, 1948 ; ,Imgensen, 1949 ; Loosanoff, 1962a). A review of the literature in this field (Jarrgensen, 1960) shows that even though some work has been performed on adult mollusks, until the recent coritributionv of Ilavis (1‘360, and unpub- lished), virtually nothing was known of thr: ability of bivalve eggs to develop or larvae to survive in turbid waters. Daviu employed a rotating wheel, to which culture vessels were attached, to maintain turbidity a t definite constant levels. The turbidity-creating subdances

54
used in his experiments were the Same as those enxplogfh(l i)y horanoff and Tominers (1948) in their st1ltljc.s 0 1 1 behavior of atfillt O.yst(’rs. Thry wwc fine silt collected frorii t j(jjtl f{ats, kml i i i (alr~mitriurn silicatcb), powdered chalk and Fuller’s cart h .
Davis showed that silt was considerably more harmful to eggs of oysters, C. cirginica, than to those of clams, M , mmenaria. For example, in concentrations of 0-25 g/l of silt only 737& of oyster eggs survived, while more than 95% of clam eggs developed to straight- hinge stage. Practically all clam eggs developed to straight-hinge stage in concentrations of 0.5 g/l of silt, while only 31% of oyster eggs survived.
In a suspension of kaolin and Fuller’s earth, on the other hand, clam eggs showed much higher mortality than eggs of oysters. Thus, in concentrations of 1 g of these substances per liter of sea water, practically all oyster eggs developed to straight-hinge stage, while only 31% and 57% of clam eggs Survived.
Strangely enough, of the materials which were tested in these experiments, silt, a natural substance, was more harmful to oyster eggs than either kaolin or Fuller’s wrth. While practically none of the eggs exposed to 1 g/l of silt rcachtd straight-hinge stage, some eggu developed normally even iri conccntrations as heavy as 4 g/l of kaolin or Fuller’s earth.
As in the case of eggs of thcsc: t w o specieB, silt was more harmful to oyster larvae than to clam larvae. At a concentration of 0.75 g/1 of silt growth of oyster larvae was markedly decreased while, a4 a striking contrast, clam larvae grew normally even in 1 g/l of silt. Moreover, the majority of the clam larvae survived for 12 days and even showed lome growth in 3 and 4 g/l although it is doubtful that they could reach metamorphosis under these conditions.
Kaolin and Fuller’s earth were considerably more harmful to clam larvae than to oyster larvae. Concentrations of 0.5 g/l of kaolin caused about 5076 mortality of clam larvae in 12 days and, while Practically no clam larvae survived in concentrations of 1 g/1 of either kaolin 01‘ Fuller’s earth, growth of oyster larvae was not appreciably affected by 1 g/l of kaolin. Davis also found that some oyster larvaR may live as long as 14 days in concentrations of 2 g/l of silt and up to ‘ d1 Of either kaolin or Fuller’s earth. These observations demon- strated an unusual ability of larvae of C. virginica to withstand highly
Of rivers where natural oyster be& arc found.
Creating IUdXrials were added to Water containing straj&-hjnge larvae
VICTOR L. LOOSANOFF ANI) HARRY c . D A V I S
turbid water, a situation often exifiting summer at o r near mouths
In several exPeriments, where small quantities of turbidity-

REARING O F BIVALVE MOLLTJSKS 56
of oysters or clams, growth of these organisms was stimulated, often becoming considerably more rapid than in control cultures. Possibly, this was a result of adsorption, by particles of such materials, of toxic substances formed in larval cultures. It is also possible that some of these materials that were added to the water contained a positive growth factcr, as do certain soil solutions.
In summarizing the observations on the effect of turbidity on oyster and clam larvae, i t may be concluded that; larvae, as well as dults , are affected by turbidity-creating substances, although larvae seem to display a considerable tolerance towards some of these materials. Moreover, larvae of different species react differently, one species may be more tolerant or less resistant to the same material under the same conditions. It is significant, nevertheless, that in some instances comparatively light concentrations of common silt may strongly interfere with normal development of eggs of some bivalves.
I. Effects of foods on growth of larvae
In any laboratory, where large numbers of adult and juvenile mollusks are kept, these animals, becaust. of limited space, are often crowded, receiving insufficient quantities of water and, therefore, food. To improve these conditions we add large quantities of artificially- grown plankton to the water flowing through troughs and trays.
Because of the scope of our work several hundred gallons. per day of relatively rich phytoplankton are often needed. Obviously, it is impractical to grow such large quantities of plankton in glass flasks using common laboratory techniques. Fortunately, by merely adding complete commercial fertilizers to sea waher, rapid growth of phyto- plankton can be initiated and later maintaiiied in heavy concentrations. Our experiments have shown that fertilizers designated by formulas 5-3-b and 6-3-6, both used by Connecticut tobacco growers, gave the best results, although lawn fertilizer, 10-6-4, was also good (Loosanoff and Engle, 1942). Using these fertilizers, mass cultures of rich, mixed plankton have been continuously grown at our laboratory since 1938. A wooden, 2000-gal oval tank is used for this purpose, although on several occasions the cultures have been grown in outdoor, concrete, 10000-gal tanks. The sea water used in these tanks was passed through a sand filter.
To supply the laboratory in winter. with a sufficient quantity of plankton we designed a special enclosure, resembling a greenhouw, in which a plankton-containing tank is instalhd (Fig. 21). By providing artificial light, when needed, and maintaining the temperature of the

56 bI( ’TOl1 I , t . O O ? A N O V b ’ A h l ) I IAl l l lY ( $ . l > A b l \
cnclosrirc~ at t l w (li-sitv(I h.vc.1, 1 hi, 1rrt)or;rtor.y ix rlow sirpplicd with rich, mixcd I)~‘~tnpliLtlkt,ot~ ~ I I i L ycw-routi(i basis.
A common difficulty expt~icwwcl in growing phytoplartkton i t1
open tanks of sevcral-thousarttl-1itc.i. ciipscity, as is donc st Milford, is invasion of these cultures by zooplankton organisms. In our mass cultures the most common offenders are crustaceans, especially cope- pods. These forms multiply so rapidly in rich phytoplankton that they soon consume most of the plant cells, rendering the culture useless.
FIG. 21. Mass culture of mixed phytoplankton grown in large nooden tank of about 2 000-gal capacity under s~rni-outdoor conditions. Desrription in text.
In the past, several methods were tried to prevent such contamina- tion of open-air algal cultures, but they usually were unsuccessful because some crustacean eggs were always left behind and eventually hatched, reinfesting the cultures. Iiecently, we devclol)cd an extremely simple and safe method to control t h s v infestations tJy mc.rc:ly adding to our culturc~s, whtw ~lccc~ssa~ty, slrliill cjuantiti~s of’ insecticides (Loosanoff et al., 1957). Several of thc:sc. substarlocs l~svc t)eeri tried and found successful in conccntratiolls as low as 0.1 ppm. A t present, we use a commcrcial preparation known as ‘l’li!Pl’, which coritairiH 40% of tetraethyl pyrophosphatc. ‘I’hc. advantages of uxirig 7’EI’f’ are

REARING OF BIVALVE MOLLUSKS 57
that it hydrolyzes within 24 to 48 hr and it has no permanent effect on algae ; therefore, it does not impair the usefulness of the culture as a food for mollusks.
The mass culture grown in our tank is not a single species but a mixture, the composition of which vnrics from day to day or cvm hour to hour. l'his ciiltirrc irsrinlly cont i b i i i s viirioiis s p c i w of (Ihlorella-like organisms, hiit hcwtiist. (:hbrrlla is not o i i v ot' the hest foods for larval
FIG. 22. Battery of spcially-fitted. 5-gal P y r e x carbuys ~ e r v i n g HR Krowth chnmhntn for m w culture of photorynthatie rnrwoorKnnmme. Dwript ion in text.

r~~)~)r.oxirn;rt,c. ly :I y ' w s ago irt i (1 h s siiicc. c*otrxist,c.iitIy c ( iv (S i i siii isfiid,ory rexults ( D a v i s and IJkc.lrx, 1! )61) .
The culture apparatus consists of sixteen 5-gal Pyrex carboys as growth chambers (Fig. 2 2 ) . Vigorous agitation, by bubbling a mixture of air and CO, through the cultures, keeps the contents of the chambers thoroughly mixed. This prevents stratification and helps to expose all cells to cqual periods of strong illumination. The carboys are immersed to a depth of 3 or 4 in in a water bath kept at a desired temperature, usually 19" f 1°C. About 3 liter of the algal culture from each growth chamber are harvested each day, yielding about 1.5 ml of packed wet cells. The present system, thus, produces daily approxi- mately 50 liter of algal suspension, averaging 0.5 ml of packed cells per liter.
A volume of sea water, nutrient salts and antibiotics, equal to the volume of culture drawn off, is added daily to each growth chamber. The sea water used is first passed through Orlon filters, previously described, to remove larger particles. Nutrient salts are then added and this solution is forced through a ceramic bacteriological filter (Selas No. FP-128-03, maximum pore size 0.6 p) into the growth chambers. Recently, wc have been adding to the sea water and nutrient salts, prior to final filtration, O-002yo Acronize (approximately loyo chlorotetracycline) to reduce bacterial growth on the ceramic filters. This concentration of Acronize does not interfere with algal growth and helps prolong the life of the cultures.
Although Chlorella sp. and a number of other algae will grow on a media made from 5-3-5 or 6-3-6 fertilizers, a more elaborate media is needed for our bacteria-free cultures. Since the requirements of each of the more than eighty species of marine algae maintained at our laboratory have not been determined, we use the following as '' univer- sal " media, although it is recognized that the necessity for the various ingredients has not been ascertained. Two stock solutions of nutrient salts are prepared and 1 ml of each is used per liter of sea water.
Solution A
Dissolve in 1 liter of distilled water NaH,PO, 20.0 g Thiamine H C1 0.2 g Biotin 0401 g BIZ 0401 g Pyradoxine HCl 0.1 g Calcium pantothenate 0.2 g

Solution B Dissolve in 1 liter of distilled water
NaN03 150.0 g *NH,Cl 50.0 g Ferric sequestrine 10.0 g * Mociia for Isockyxiu gulbunu shoci l t l omit the NH,CI.
Experiments conducted at our laboratory have shown that some species of bacteria are harmless, others are strongly pathogenic, and still others produce toxic metabolites. It is quite possible, therefore, for some bacterized cultures of algae to be good larval foods, while the same algae, with a different bacterial population, may be acutely toxic. Consequently, for critical evaluation of any phytoplankton organism as a food for larvae it is necessary to use a bacteria-free culture of it. Furthermore, in using mass cultures of algae, where there is always danger of bacterial contamination, i t should always be ascer- tained that the cultures have not become contaminated with toxic o r p t hopwic: 1)nctmia.
As already mentioned, certain species of algae also produce meta- bolites that are toxic to bivalve larvae, while other species produce little or none (Fig. 23). Some of the algae produce so much toxic material that they are useless as foods because their toxins kill larvae even when concentration of algal cells is too light to satisfy larval demand for food. Others, such as Chlorella (580), produce some toxic products but are still usable foods, provided that their concentrations are not too high.
Recently, Davis (1953) and Davis and Cjuillard (1958) concluded that presence and thickness of cc:ll walla t i r i d dngrtu: fd frtxic:it,,y of metabolites are important factors in determining uarthility of jdioto- synthetic microorganisms as larval food. 7'hcy Rhowed that tht: m k ~ d flagellates, I . gulbana and M . Zutheri, were of apjmxirnetcly qud value as food for larvae of the American oyster and inducc:d marc rapid growth than any of the other species tested. Davis also believw that I . gulbuna and M . Zutheri produce little, if any, toxic external metabolites which unfavorably affect Iarvac:. This is supported by his ot)sc:rvntions that thcb optimal conccntrationr;l of the two formS for either clam or oyster larvae wore a t lcast tloublt: the optirnul coricen- tratiom of Chlorelkz RP. (Lewin's isolate).
The food value of microorganifima also tlopend~, in part, upon how
for i > x i m p l v , t I I I L ~ , IL misf i i r ~ ot' I . p r / / ) ( i / / u , M . h t h ~ r i , f'l~ikrl~/mo~~as sp. aiitl 1)uncrliolLr vuchloru iiitluccd morc rapid growth of both clam
(.ornplc.ltdy tht1.V I I I W ~ , t h foot1 ~ . c ~ ( l ~ i i ~ . c . r n c . ~ ~ t , q of I IL~VIU! . It, w w ~ O I I I I ~ ,

F I ~ , 23. Twelve-day-old larvae of (I'rfAwo&eu oi7yir~icn givwl diffcr~!nt algw us) food. Group fed M . lutheti ( u p p e ~ 7 h > t o g ~ ~ i p h ) uveragctl 189 p in Iwigtti, while the mitldlo group, serving aa control and receiving I I O supplemerltary f i d , averaged 96 p. The lower group, containing Only dead larvae, was givrrl Stichococcm Hp. iwleted from Great South Bay, Long Island, S e w York, which produces toxic metabolites.

REARMQ OF BIVALVE MOLLUSKS 61
and oyster larvae than did equal quantities of any of these foods separately.
D . euchlora and Dunalidla sp., both naked flagellates, as are 1. galbarn and N. lutheri, also induced better growth of oyster larvae during the first 6 days of their development than did any of the forms having cell walls. Thus, Davis concliidetl that, with the exception of Prymnesium parvum, which is toxic, evm the poorest of the naked flagellates is a better food for young Iirrvrbe of G . virginica than any of the organisms with cell walls. This coriclusion supports the earlier one of Cole (1939) and Bruce et al. (1940), that nanoplankton may differ in their value as food for larvae of 0. edulis. Davis found, never- theless, that Chlorella (580), Platymows sp., Chlorococcum sp., and Phaeodactylum tricornutum, all forms having cell walls, were utilized by larvae of C . virginica, but growth, particularly of younger individuals, was slow and might have occurred because of presence in the water of other food materials.
Observations on behavior of larvae of approximately twenty different species of bivalves grown at our laboratory have shown that, in general, as far as their qualitative food requirements are concerned, they can be roughly divided into two or, perhaps, three groups. The first group, well-represented by larvae of oysters of the genus Crass- ostrea, is able to utilize, during early straight-hinge stages, only a few of the many food forms (Davis, 1953; Davis and Guillard, 1958). The second group includes larvae of such species aa Mereenaria mercenaria and Mytilus edulis, which seem to be able to utilize most of the micro- organisms, provided that they are small (.nough to be ingested. The third, an intermediate group, also can be tentatively recognized. This group includes such larvae as those of larviparous oysters of the genuu Ostrea, which are much less restricted in their qualitative food rcquire- ments than larvae of the genus Cruseostron, yet they are unable to grow quite as well on some of the foods as larvae of M . mercenaria.
The food requirements of the two marginal groups were clearly shown in our experiments where larvae of the oyster, C. virginica, and clam, M . mercenaria, were placed simultaneously and kept together in the same laboratory culture vessels or in large outdoor tanks. All cultures were given the same food, which consisted of mixed phpto- plankton in which small, green algae, such as Chlorella, normally pre- dominated. Under these conditions larvae of 21.1. nlercenaria grew rapidly and metamorphosed approximately in 12 days, while larvae of C. vir- ginica, after attaining straight-hinge stage, showed virtually no growth and eventually died. The results of theje early experiments were confirmed by Davis (1953), who showed that while young larvae of

62 VICTOR L. L O O S A N O W A N D HARRY C. D A V I S
C . virginica are unable to utilize forms having cell walls, such as Chlorella, older larvae of the same species become able to do so after they reach a larger size of approximately 110 p.
I n general, these studies have demonstrated several important points. One of them is that larvae of M. mercenaria can live and grow to metamorphosis on a very restricted diet, consisting of a single species of algae, such as Chlorclla, and that unlike larvac of C. virginica, they can utilize these algae during all stages of clevalopment. Our conclasions, therefore, disagree with those of Cole ( 1938) who maintained that bivalve larvae, in general, (lo riot possess the enzymes needed for digestion of cellulose, of which the cell walls of algae, such as Cklorella, are made.
As our techniques improved and we were able to evaluate the food value of different forms of phytoplankton, we found that larvae of M. mercsnaria can be grown not only on a pure culture of Chlorella, reaching metamorphosis in some cases in 12 days, but that they can also be grown to metamorphosis on pure cultures of any one of the following three flagellates : Chlumydomoitas sp., Chromulina pleiades or I . galbana. (Davis and Loosanoff, 1953).
Our studies have also shown that organic detritus, at least of the types tested, cannot be utilized by larvae of clams, M. mercenaria (Loosanoff et al., 1951), or oysters (Davis, 1953).
Larvae of M. mercenaria seem to be capable of both mechanical, or quantitative, and chemical, or qualitative, selectivity in feeding. They arc athlo to regulate tho amount, of food ingested : ~ n d t h u s survivc: in heavy conccntrations of food cc:IIs, often containing IWH food in their stomachs than larvtu: kcJ)t in lighter food conct:ritrationn. AJ)pari:ntly, c l a r r i larvrrc: w i : tiot rric:rc:ly i i w ( h ~ t t i c d f i ~ t ( I i : r ~ , twt ~ O H H C H H
a mccIiruiisrri tiy I Y I C ~ ~ I ~ H of which th:y can coiitrol the fiJOd intake by rejecting algal cells, when necessary. However, if kept in heavy con- centrations of algae for a long time, the larvae loso this regulating ability, become choked with food cells and, eventually, die.
We also observed that larvae can select certain food organisms from a mixture of several forms of phytoplankt,ori. ]+'or c:xample, whcm given a mixture of Porphyridiurrb SJI. and Chlrlm~~ilortrori,r,.nH H I ) , Inrvtw of M. mercenuria ingested the much 1argt:r cells of' Chllnr.?ldi~u)ri,ax, while rejecting the cells of the smaller Porphyridium (1,oonarioff at d , 1953).
An important problem faced in connection wi th cultivatiort of bivalves was to ascertain the cffects of different, ooiic:c:rit,rRt,iotIH of' food organisms upon larval survival and rate of growth. 'I'hc first series of experiments was conducted with larvae of M. mercsn,nria in

ItEARINQ OF BIVALVE NOLLUSKS ti3
concentrations of approximately seven larvae per ml, but fed different quantities of food, consisting principally of small Chlorella measuring only about 3 p in diameter. These cells were fed to larvae in concen- trations ranging from 6 500 to 1 million cells per ml of water in culture vessels. Simultaneously, another series of larval cultures was fed a unialgal strain of another Chlorella, the cells of which were about 8 p in diameter (Loosanoff et aE., 1953).
Results showed that optimal concentrations of food cells clearly depended upon their size. When large Chlorella was given, the optimal concentration of this form for best survival and growth of larvae of M. mercenuria was approximately 50000 cells per ml, while approxi- mately 400000 cells per ml of the smaller Chkwella were needed to achieve the same results. This suggests that the food value of 400 000 small Chlorella cells closely approached that of 50000 cells of the larger form, both concentrations being near optimum. If the cells are considered as perfect spheres, the volume of 400000 cells, 3 p in diameter, is approximately equal to the volume of 50000 cells, 8 p in diameter.
A concentration of approximately 750000 cells per ml of small Chlorella was already above the optimum because, when given so much food, the larvae grew more slowly than when given only 400 000 cells per ml of small Chlorella.
Larvae of M . mercenaria and many other bivalves can be killed if concentrations of food cells, such as Chlorellu, become too heavy. Again, these concentrations depend upon the size and kind of cells. For example, approximately 90% of clam larvae were killed within a few days and those that survived grew very slowly or not a t all when given approximately 300000 cells of large Chlorella per ml of water. When the concentration was increased to 500 000 or more cells per ml, all clam larvae were kilbd within 24 hr. However, when given the much smaller form of Chlorella, which measuretl only about 3 p, the larvae grew comparatively well in concentrations as high as 750000 cells per ml.
It is of interest that larvae that rnan;Lged to survive in heavily overfed cultures usually displayed certain anatomical ahnorrnalitie~~ which, probably, made the larvae unable to ingest food. Perhaps the% abnormalities were responsible €or survival of these larvae under the conditions where normal individuals were killed.
We realize that, in studying the food requirements of bivalve larvae, the quantity of algal cells in the surrounding water constitutes only one factor needed to determine the adequacy of a food because this value may be subject to considerable ~ariations according to the

age of the algal cultures, their density, chemical coml)osition, hactrrial flora and, of course, the media in which they arc grown. These difi- culties have been eliminated, to some extent, in our more recent experi- ments where production of food cells has been standardized (Davis and Ukeles, 1961).
As has previously been showii for adult bivdves (Loosanoff and Engle, 1347), their larvae can be killed either by a heavy concentration of algal cells alone, by the filtrate of algal cultures or by a combination of the two (Loosanoff et al., 1953). In other words, dense concentrations of certain food organisms, such as Chlorella, affect larvae of M . mer- cenaria, as well as those of several other species, both mechanically, by interference of food cells with larval swimming and feeding mecha- nisms, and chemically, by producing external metabolites which are toxic to larvae. As an illustration, the larvae grew comparatively well when control cultures received approximately 100000 cells per ml of large Chlorella, even though this concentration was somewhat above the optimum for this strain of algae. However, cultures of larvae receiving cells of Chbrella, t h a t had been removed from the culture medium by Millipore filters and later resuspended in sea water at the. rate of 1 million cells per ml, were rapidly killed. Similarly, larvae receiving the filtrate only from a certain volume of algal culture originally containing one million cclls of Chlorella per ml also quickly died. These studies further showed that a filtrate containing heavy concentrations of metabolites of Chlorella cells may be even more detrimental to larvae than heavy concentrations of the resuspended cells themselves. As has already been mentioned, the ecological effects of external metabolites have long been recognized by aquatic biologists (Lucas, 1947).
Recently, Davis and Guillard ( 1958) conducted extensive experi- ments to determine the relative value, as larval food, of representatives of ten different genera of microorganisms. I . galbana and M . Eutheri were the best foods and were approximately of equal value. I n some experiments, nevertheless, Chlorococcum sp. was the best food for larvae of M . mercenaria. Clam larvae were also able to utilize several species of Chlorellu, Dunaliellu euchlora, Dunaliella sp., P l a t y m o w sp., Chlamydomonas sp. and Phaeodactylum tricornutum. However, they could not utilize one species of Stichococcw or Prymnesiuni parcum.
Experiments also demonstrated that, as in the case of larvae of C. virginica, a mixture of I . yalhana, M . lutheri, Platymclnas. sp. and D. euchlora promoted somewhat more rapid growth of clam larvae than did equal quantities of any of these food8 separately.
Some of the algae tested at Milford Laboratory are given below with

REARING OF BIVALVE MOLLUSKS 65
the species listed in their approximately descending order of value as foods for larvae of M. mercennrin and G . virginica. Several other specks tcstctl arc’ riot listcd hcm lwcau~(+ t h y were' tither poor foode or wwc toxic.
M . m wcenaria
Nonochrysis lutheri Isochrysis galbana Dicrateria sp. (B 11) Chlorococcum sp. Platymoms sp. (1)
Carteriu sp. Chlamydomonas sp. Cyclotella sp. (0-3A) Chlorclla sp. (580)
Ph~rotlac t y lum tricornutum Skalctonrma co,ytnlum Chlamydomonas sp. (D) Rhodomonas sp. Dunaliella sp. Olisthodiscus sp. Dumliella euchlora
A. Good Foods:
B. Medium Foods:
Stichococcus 8P. (0- 18)
C . virginica
A. Good Foods: Monochrysis lutheri Isochrysis galbana Chromulinu pleiades* Dicrateria inornata* Pyramirnonas grossi* Hemiselmis refesced
Dunaliella euchlora I’latymonas sp. ( 1) C‘yclotella sp. (0-3A) 1)unaliella sp.
Chlorella sp. (UHMC) Phueodactylum tricornutum Cryptomonas sp.
B. Medium Foods:
‘lilorocomu?n up.
* Not tested on clam larvae,
The most recent contribution to oiir kriclwlcclgc. of fowl rc*cjtiirv
ments of the European flat oystm, 0. P ~ u Z ~ H , WRH rnadf: hy Walne ( 1!)56) who also rcviewcvi cfforts in this fivld by earlier investigators. Walnr contliirtc~ti o w r ‘LOO twtg wing many species of algae. Although he statcs that rnr~riy of his experiments wero failures, often for no appnrrnt, r(vwoii, his rr8iilts 8liowod that t h Chrysophyceac, eHpccially lwchr,t/.si<v !l(i l lmt(i tL i i ( I ( ‘hromulinn pleirirleqs, gavo good results, not- ~ i t ~ l i ~ t i ~ i i d i i i ~ 1 l i d , /’. p i n l t i i r i W I ~ H toxic to larvae. Member8 of the ~‘Iiloroc~cic~c~ril~~s. wliidi Iiavc- 41 t trick ( ~ 4 1 wnll, wcrc found to be poor f0Od for t h I W V I L O of‘ 0. edulis. Of tho (%lorophyccm, only I’yrnn& nionns p , w i W ~ L H good food.
In our experiments we found that larvae of 0. edulis can be grown on a species of Chlorella, even though i t has a thick cell wall. In this respect our results disagree with those of Walne and Home of the earlier investigators. We agree, howevrr, that naked flagcllrttn~,

66 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
especially I. gaBana and M. Eutheri, are the best foods for larvae of this oyster.
Imai et al. (1953) mentioned in several articles the successful use of the colorless naked flagellate, Monas sp., as a food for oyster larvae. Through the kindness of Dr. Imai we received a sample of a culture of this organism and grew i t on a large scale. However, we have consis- tently been unable to raise larvae on a diet of this flagellate alone. In our controlled experiments larvae of M. mercenaria and C. virginica grew and survived better in unfed controls than when given 4 000 cells of M o w sp. per ml of water in culture vessels. Use of the same flagellate by oyster biologists of the State of Washington (Lindsay, personal communication), who also obtained these flagellates from Dr. Imai, always resulted in failure. Our conclusion that colorless flagellates are not a good food for bivalve larvae is also supported by recent observations of Walne (1956), who could not raise larvae of 0. edulis by using another colorless flagellate, B& sp.
Growing of algal food for larval or juvenile mollusks, even if these cultures do not need to be kept unialgal and bacteria-free, presents a number of difficulties and, naturally, an easier method of producing food for bivalves, including adults, is desirable. Our experience, in the late 'thirties, in attempting to increase the glycogen content of adult oysters by feeding them pulverized dried algae, such as different species of Ulva and Laminaria, led us to try the same approach in feeding larvae with dried algae. In the initial experiments dried and pulverized Ulva, which we preserved and kept in a glass jar for over 20 years, was used. Surprisingly enough, many cultures of larvae of M. mercenaria, given this food, grew to metamorphosis. However, several problems were encountered in connection with this method. First, i t was found difficult to grind these algae into particles small enough to be ingested by larvae. Secondly, the material quickly settled on the bottom, thus becoming unavailable to swimming larvae. Finally, because of rapid decomposition of unconsumed algae, the larval cultures became fouled and the bacterial flora in them quickly increased.
Fortunately, we have been able to overcome these difficultiea. Through the courtesy of Dr. Hiroshi Tamiya of The Tokugawa Institute for Biological Research in Tokyo, Japan, and Dr. Hiroshi Nakamura of the Japanese Nutrition Association, we received samples of dried Scenedamus sp. and ChloreUa sp. Because the average size of the individual cells of both of these algae is small, they can be easily ingested by larvae. By feeding proper quantities of dried, ground Scenedesmus in larval cultures, which are continuouely agitated by

REARING OF BIVALVE MOLLUSKS 67
plungers or horizontal-action paddles (Fig, 5), and by controlling tho bacterial flora in the cult,iirt~s by ih addition of Siilmct, our associntc, t l . 11 idu, to whom t h s o ~ t u t l i c w w t w ~uggested, liati I)cc.n
YJrr. 24. Twelve-day-old larvae of Mercenaria marconaria. Larvas shown in upper photomicrograph were fed freezo-dried celh of' Imr.hryais galbarn, while thoae in the lowor picture wore given the Rame quantitiocc Of l ivo n t h of tho RBIf10 d g m . 1,crrpst larviu, two t h ) i i t , 215 p loiig.
i ~ l ~ l o to grow IILI-VIL(~ 01' A t . ?nwrrnririii : L I I I \ O H ~ ax c?ffectivcly I ~ H when Llwy arc focl o u r Iwnt, l ivo l o o t l orgtmixms, Hueh JJH naked flagellates. In some experimcnts olarn Iarvse fed freeze-dried 1. gnlbn?zu grcw aN rapidly as larvae in the control cultures given the same quantities of live cells of the same form (Fig. 24).
6

68 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
When fed Seenedesmus clam larvae grew best in concentrations of 0.02 ml packed food per liter of water per day. However, dosages as low as 0.005 ml packed food per liter gave just as good results in feeding larvae during their first 4 days of existence (Hidu, personal communi- cation). Agitated cultures treated with Sulmet gave 50 to 100% better growth of larvae than identical unagitated cultures. The type of motion and size of agitating devices appeared to bc of little significance t o larval growth and survival. Mortality in all agitated cultures was negligible. I n some situations, nevertheless, agitation may not be desired.
Experiments on growing larvsr of American oysters on dried Scenedesmus have been unsuccessful thus far. We are, however, continuing experiments in this field using, at present in a dried state, such organisms as I . gulbuna and M . lutheri which, when alive, are readily utilized by larvae of C. virginica. Results of these and related studies will be reported in the near future by Mr. Hidu. It should be realized, nevertheless, that this sphere of research is now only in its initial phase and, therefore, i t is too early to form general conclusions. We hope, however, that i t will be extremely productive.
In discussing larval foods we would like to emphasize that color of the larvae themselves depends, to a large extent, upon color of food consumed. By feeding larvae differently colored food organisms we were able, especially during early straight-hinge stages, to change their color within a few hours.
Although the color of the shell proper was not always affected, the soft parts of the body quickly changed their coloration which, of course, reflected on the color of the larvae as a whole. It may also be added that according to our observations even the color of shells of recently set oysters growing in running water is darker t h a n that of oysters of the same age that are kept in filtered sea water and fed naked flagellates. However, if these pale young oysters are transferred to natural water containing color-carrying phytoplankton, they quickly develop a much darker color. It seems logical to expect that in nature, where combinations of forms constituting phytoplankton change frequently, correspondingly frequent and rapid changes in the general color of larvae may also occur. We disagree, therefore, with workers who maintain that color is a dependable criterion for identification of bivalve larvae.
J. Effects of crowding An important problem that faced us in early attempts to rear
molluscan larvae was to determine maximum concentrations at which

REARINQ OF BIVALVE MOLLUSKS 69
they could survive and optimal concentratioiis for their growth. We expected these concentrations to depend upon species and quantity and quality of food, but soon found that such hardy varieties as M . mercenaria could be grown in concentrations of 50 to 100 larvae per ml of water. If these cultures were properly attended and fed, the majority of the larvae survived and, despite their slow rate of growth, they could, nevertheless, be reared to metamorphosis.
Although these were much heavier concentrations than those advocated by other investigators, who usually emphasize the danger of overcrowding, we sometimes reared larvae to metamorphosis even under more crowded conditions. However, since these observations were of a general nature, special quantitative experiments were undertaken (Loosanoff et al., 1955). In these experiments larvae of M. mercenaria were grown in concentrat ions of about six, thirteen, twenty-Rix and fifty-two individuals pc’r ml. Each culture received the same quantity of food, namely, 100000 cells of a small strain of Chlorellu per ml of water in the rearing vessels.
Larvae in all cultures showed good survival and grew to meta- morphosis, although the mean rate of growth in the different cultures had an inverse relationship to population density. For example, on the loth day the mean length of larvae in the series of cultures containing six, thirteen, twenty-six and fifty-two individuals per ml was 162, 156, 151 and 144 p , respectively. Experiments showed that, as reported earlier (Loosanoff et aZ., 1951), overcrowding clam larvae may not be as dangerous as formerly believed but, nevertheless, rate of growth in overcrowded cultures is slower and time of metamorphosis is corres- pondingly delayed.
Slower growth in the moro crowrh~tl cattiires may hc attributed, i i i part, to mow frc.qiiciit (:ollisionH Iwtwcwi tho larvae and, in part, to t h t l c l ~ ~ t c w i o i w c 4 h t of groatw ~ o J i ( ~ v i t r~itions of cbxcrcttory producte o f Iiwviw. l~iiiiill.v, L h v f w t tlirLt t l i t w wns ICRR food per larva in the IWrv t l n s v ity ~~op t i t~b t~d cwlturtw wits suNIwctCd &R the most, probable (*WINO of slow(^ growl It. If this w ( w trricb, rato of growth of larvae in denecly I)o~)uliit ,t~d cultures slioirld be Iirought up to normal by a proportional increase in amount of food supplied. Accordingly, a new series of experiments was conducted in which number of ccll~ of Chlorellu was increased in proportion to increase in larval population. This was achieved by adding daily to the culture vessels containing six, thirteen, twenty-six and fifty-two larvae per ml of water, approxi- mately 100 000, 200 000, 400 000 and 800 000 cell8 of ChZoreZla per mi.
After 10 days larvae in the leafit crowdrd culture averaged 1 NZ / L in length, while larvae in the culture contairrirrg thirteen l w v w pw rrrl

70 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
averaged only 148 p. The two other cultures, containing twenty-six and fifty-two larvae and given 400000 and 800000 Chlordh cells per ml, died within a few days. This and similar experiments showed, therefore, that crowding of larvae beyond certain limits cannot always
D FIG. 25. Forty-elght-hour larvae of Mereenaria mercenaria developing from egg8
cultured at different denaities. A, 250 eggs per milliliter of water ; R, 500 egg6 per ml ; C, 1000 eggs per ml ; and D, 3000 eggs per ml.
be remedied by a proportional increase in quantity of food. It w88 also demonstrated that lethal concentrations of food organisms, 88
was the case when 400 000 and 800 000 cells of ChkweUu per ml were given, cannot always be reduced to a safe level by the large number of larvae that presumably would rapidly consume these cells.

REARING O F BIVALVE MOLLUSKS 71
Still another experiment showed that eggs and larvae of M . m,ercen- aria can develop under even much more crowded conditions than we have indicated thus far. In thiR expcrimcnt cultiirerr werc begun by placing in rearing v e s d s 2b0, fj00, 7b0, 1000 rind 3000 ~ g g s pvr ml of water. The cultures wcre incu t)ntctl r h t R tcrnpcmtirre of ahout 24'C and examined st the end of 48 hr. 'I'hc majority of eggs rcarccl a t a density of 'LBO per ml developed into normal straight-hinge larvae (Fig. 25), while in cultures that were begun with 3000 eggs per ml, most of the eggs failed to develop past morula stage and those few that managed to develop to shelled stage were abnormal. In inter- mediate concentrations of BOO, 750 and 1000 larvae per ml some eggs managed to dcvolop into normal straight-hitigo larvae Init thc' w num- hcrs clecreasctl proportionally with each iricroase in the initial concen- tration of eggs.
Experiments on effects of crowding on growth of larvae of the American oyster, C. virginica, were described by Davis (1953). He used 640, 2 785, 18 518 and 32 918 48-hr-old larvae per liter and gave all of these cultures the same quantity of food, namely, 50000 cells of Chlorella per ml of water. A t the end of 14 days larvae from all cultures werc! examined and measured. Rceults showed that there was an invc:rst. relationship bctwcen conccntrations and rates of growth of larvae. Thus, observations on Inrvac! o f (1. viryinica were i n agrcc- ment with similar observations on clam Iarvac.
On the basis of our results we can conclude that the danger from overcrowding larvae in well-maintained cultures may not be as acute as believed. Nevertheless, crowding is undesirable because it decreases rate of growth of larvae and, ot)viously, overcrowded larva(. are more susceptible to diseases than are larvao i n IPHS ~ P I I H P I Y ~ ~ ) p i i I ~ t C ! d cultures.

72 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
or, perhaps, even days. This opinion is indirectly supported by the suggestion of Thorson (1946), Wilson (1958) and other investigators that pediveligers of many bivalves are able to postpone metamorphosis if conditions of environment are unfavorable for completion of this important stage.
Literature contains few references to anatomical changes accom- panying metamorphosis of bivalves. This is understandable because, until recently, few forms were successfully and repeatedly grown to this stage. Prytherch's (1934) description of metamorphosis in C. virginiea, Cole's (1938) in 0. e d u b and, especially, Quayle's (1962) in Venerupie pullustra should be mentioned as contributing to our knowledge in this field. However, since a description of the major anatomical changes occurring during metamorphosis does not con- stitute a part of this paper, no references will be made to the details of this phenomenon in larvae of the mollusks studied by us.
As already mentioned, the number of days required for larvae of the same species to reach metamorphosis depends upon a complex combination of ecological factors, of which temperature and quantity and quality of food are the most important. Each species, of course, has its optimal temperature range for larval development, as well as minimum and maximum levels within which development is possible. Since all species that we cultured were grown to metamorphosis at room temperatures ranging from 18" to 24OC, it is quite certain that this part of the temperature range is comparatively favorable for development of most bivalves of our waters, as well as some of the exotic species with which we worked. However, information aa to complete temperature range within which development of each of these species is possible is still lacking in most instances.
For example, aa already mentioned, in the case of M . mercenaria i t has been ascertained that larvae grown at 18°C began to metamorphose 16 days after fertilization and continued to set as late as the 30th day. Thus; at thh temperature, the period between beginning and completion of meta- morphosis of larvae of the same cultures extended for 14 days. At a temperature of 24°C the earliest metamorphosing larvae were noticed on the 11th day, and the latest, 22 days after fertilization. At the considerably higher temperature of 30°C setting of clam larvae began as early as the 7th day after fertilization and was normally completed within 5 to 7 days, or by the 14th day, although in some of the more erratic cultures the last setting larvae were observed on the 16th day after fertilization (Loosanoff et al., 1951).
Similar observations on larvae of C. virginicu showed that, at a
We have obtained such data for some species.

HEARING OF BIVALVE MOLLUSKS 73
temperature of 30°C setting usually began on the 10th or 12th day but, again, i t extended for several days, as was the case with clam larvae. At lower temperatures the beginning of setting was delayed and its duration prolonged. For example, in one culture kept a t a temperature of 24°C the larvae metamorphosed continuously for 27 days.
The number of days needed for larvae of the same cultures to reach setting stage is, therefore, not sharply dejined even for a culture reared under standard, well-controlled conditions. In poorly fed, crowded or generally neglected cultures the beginning of setting is usually delayed and the time difference between beginning and end of setting is considerably extended. Sometimes, in cultures kept under poor conditions, towards the end of the experiment large numbers of larvae were found which were apparently unable to grow beyond a certain stage and eventually died without metamorphosing.
In earlier studies (Loosanoff and Davis, 1950) we were already aware that a t the time of metamorphosis the size of larvae varied, even among those of the same species. At first, the smallest meta- morphosing larvae of M . mercenaria were thought to be 210 p long but, later, larvae measuring only about 175 p but already going through this process were seen in the cultures. However, setting occurs most commonly when larvae measure between 200 and 210 p in length.
Often, in the same cultures comparatively small individuals only about 180 p in length will complete metamorphosis, while many larger larvae, exceeding 220 p in length, still continue to swim.
Since there are several suggestions in the literature that invertebrate larvae reach a larger size before metamorphosis if they develop at low temperatures, we conducted a number of experiments to verify these suggestions (Loosanoff, 1959). Thjs information was desired because if i t were true that there was a definite relationship between setting size and temperature a t which the larvae were grown, then i t fihould be possible to devise a formula to predict setting size for each tem- perature.
The studies were conducted with larvae of M . mercenaria a t tem- Peratures ranging from 18" to 30°C. Within this temperature range there were no significant"differences with respect to mean length of larvae at time of setting. Thus, although larvae grown at lower tem- Peratures required more time before reaching metamorphosis, in all Cases the larvae attained approximately the same mean lengths before setting. Furthermore, there was no apparent relationship between
The largest larvae do not always metamorphose first.

74 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
maximum lengths of larvae at time of setting and water temperature. Finally, no relationship was found between shape of larvae (ratio of mean lengths to mean widths) at the time of setting and water tem- perature.
Incidentally, these studies gave us an opportunity to verify the opinion expressed by some authors that forms Ijving in warmer waters have just as long a pelagic life as related species living in northern waters, and that eggs and larvae of southern species develop more slowly than those of northern species of the same genus if both species are kept under identical temperature conditions. We conducted these studies with M. mercenaria of Long Island Sound and M. eampecfiienais imported from the Gulf of Mexico. Both groups were ripened in our laboratory under identical conditions and induced to spawn by the usual methods. Eggs and larvae of both species were gro& at a temperature of 21°C in triplicate cultures, and random samples of 100 larvae from each culture were taken for length and mortality measurements every two days from fertilization to metamorphosis.
Results clearly indicated that rates of growth and mortality of larvae of the two species were practically identical and that setting of larvae of both groups began simultaneously. On the basis of these observations we may conclude that the belief that eggs and larvae of southern species, when grown at the same temperatures, develop more slowly than those of the northern species of the same genus is not true in all cases.
As has already been mentioned, in some bivalves the beginning of metamorphosis may be delayed because of unfavorable conditions. In addition, in a t least one case, namely, that of A m i a simplez, we observed what appeared to be partial or interrupted metamorphosis (Loosanoff, 1961). While in larvae of this species metamorphosis commonly occurs at a length of approximately 200 p, hundreds of individuals considerably larger than 215 p were seen in some cultures crawling on the bottom by means of a powerful ciliated foot. One of these individuals megsured 577 x 514 p.
Partial metamorphosis was characterized by disappearance of the velum but retention of a functional foot. Moreover, examination of these crawlers showed another common characteristic, namely, a definite narrow band on their shells, which probably indicated the edge of prodissoconch, or larval shell, and the beginning of dissoconch, or post-larval shell. We assume that the band dividing the two portions of the shell was formed during an important event in the life of the young mollusk, possibly when the velum had entirely disappeared or some other equally important anatomical or physiological change

REARING OF BIVALVE MOLLUSKS 75
had taken place. This conclusion is supported by measurements of the inner prodissoconch shells surrounded by the band, whiah were usually approximately 200 p in length. This is the size a t which setting of tho majority of normal larvae of A. simplex occurred.
L. Diseases of larval and juvenile mollusks and their treatment Even under the best conditions that can now be maintained in the
laboratory there have been occasional heavy mortalities of larval and juvenile mollusks that cannot be accounted for by experimental treat- ment or errors in technique. Although in most instances this mortality was light, weak or dying larvae could usually be seen harboring heavy populations of bacteria or protozoans: in some cams the mortality was epizootic, killing nearly all young bivalves within a few days.
The field of diseases and parasites of larval and juvenile mollusks is entirely new because studies of this nature were impossible until successful methods for rearing larvae were developed. No information was available, therefore, as to whether the bacteria, protozoans, etc., observed in weak and dying larvae, were the primary cause of their death or merely secondary invaders and, perhaps, only scavengers. Although Davis (1953), in attempts to feed oyster larvae on bacteria, had shown that bacteria retarded growth of larvae and eventually killed them, i t was not until 1954 that a fungus, Sirolpidium zooph- thorum (Fig. 26), was found responsible for some of the epizootic mortalities in our cultures of clam larvae (Davis et al., 1954; Vishniac, 1955). So far, fungus has been found to affect larvae of M. mercenuria, M . campechiensis, the hybrids of these two species, Teredo nuvalis, Pecten irradians, Tapes semidecussata and G. virginica. However, since it was observed and identified only recently, wc have not established whether i t can infect larvae of those bivalves which were reared at our laboratory prior to its discovery.
Not all of the forms mentioned above are equally susceptible to fungus. For example, while larvae of M. mcrcenaria and Teredo nuvalis are easily infected, the infection often assuming epizootic proportions, infection of larvae of C. virginica is extremely rare. The only case of this type occurred when a large number of fungus-infected clam larvae was introduced into a vessel containing young oyster larvae. Larvae of other forms, such as the bay scallop, Perten irradians, and the Japanese clam, Tapes semidecussata, frequently became infected, but the disease seldom reached epidemic proportions.
In M. mercemria and several other species, larvae of all ages, from very early free-swimming stage to thofie ready to undergo metamor- phosis, can be parasitized by fungus. .Juvenile clamu up to 400 p in
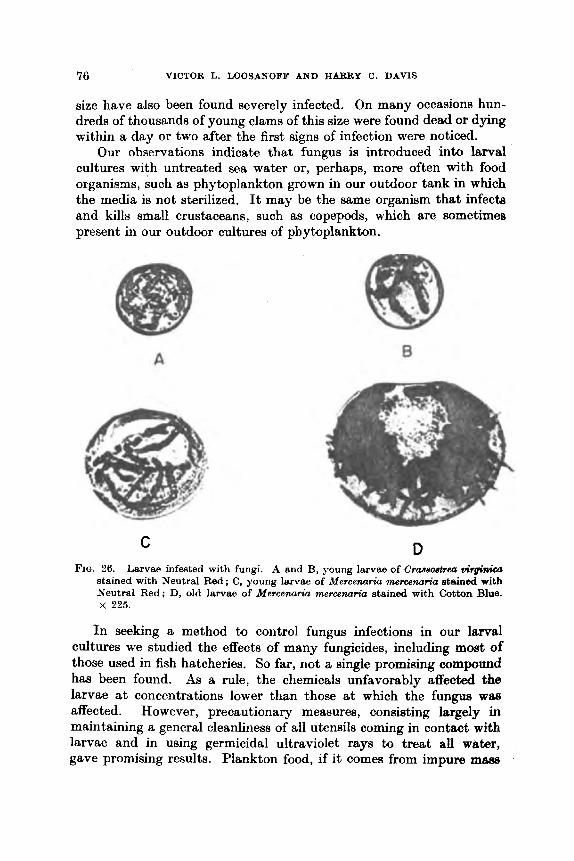
76 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
size have also been found severely infected. On many occasions hun- dreds of thousands of young clams of this size were found dead or dying within a day or two after the first signs of infection were noticed.
Our observations indicate that fungus is introduced into larval cultures with untreated sea water or, perhaps, more often with food organisms, such as phytoplankton grown in our outdoor tank in which the media is not sterilized. It may be the same organism that infects and kills small crustaceans, such &s copepods, which are sometimes present in our outdoor cultures of phytoplankton.
8 A
D C FIG. 26. Larvae infested with fungi. A and B, young larvae of Crassostrea VitginiCa
stained with Neutral Red ; C, young larvae of Mercenuria mercenaria stained with Neutral Red; D, old larvae of Mercenarka memenaria stained with Cotton Blue. x 225.
In seeking a method to control fungus infections in our larva1 cultures we studied the effects Gf many fungicides, including most of those used in fish hatcheries. So far, not a single promising compound has been found. As a rule, the chemicals unfavorably affected the larvae at coneentrations lower than those at which the fungus wae affected. However, precautionary measures, consisting largely in maintaining a general cleanliness of all utensils coming in contact with larvae and in using germicidal ultraviolet rays to treat all water, gave promising results. Plankton food, if i t comes from impure rn-

REARING OF BIVALVE MOLLUSKS 77
cultures, should also pass over the ultraviolet tubes. This measure, in addition to controlling the fungus, also potects larval cultures against invasion by undesirable forms, such as small crustaceans, larvae of different worms, rotifers and protozoa, which compete with bivalve larvae for space and food. I n some instances raising the water tem- perature to approximately 32.5"C for several hours may kill fungus without causing serious injury to young mollusks.
Ultraviolet treatment of sea water for purification of shellfish has been described by several workers in Japan, and by Wood (1961) in England. Waugh (1958) and Walne (1958) also used ultraviolet-treated sea water for rearing larvae of 0. edulis. To prevent fungus diseases of young clams we began using ultraviolet light in 1954 to treat sea water and soon found that this treatment was helphl in preventing mortality of juvenile clams kept in trays of running water. In the summer of 1955 it was found that larval cultures receiving ultraviolet-treated sea water, but untreated phytoplankton from the outdoor tank, developed fungus, while cultures receiving the same phytoplankton and sea water, both of which were treated, did not become infected.
Because of these observations and because i t had long been recog- nized at our laboratory that many mortalities in larval cultures were caused by bacteria (Loosanoff, 1954; Davis and Chanley, 1956b), i t became our standard practice, since the summer of 1955, to use filtered and ultraviolet-treated sea water in all larval cultures and in standing water trays containing recently set clams.
Walne (1958) also reported that bacteria may seriously affect larvae and Guillard (1959) demonstrated, while working at our labora- tory, that certain bacteria produce toxins that can retard growth of larvae or kill them. Guillard isolated two virulent clones, one of which appeared to be a species of Vibrio, and the other, of P s e u d m m s . His experiments showed that larvae of M . memenaria, when exposed to virulent bacteria and treated simultaneouslg with antibiotics, such as penicillin and streptomycin sulfate, remained as healthy as the control cultures. High temperature, near 30"C, favors growth of bacteria, thus emphasizing some of the disadvantages of growing larvae at com- paratively high temperatures. Bacterial mfttabolites, when present in high concentrations, may stop larval growth entirely. Cuillard also noticed that bacterial contamination of algal food cultures sometimes caused a sharp decrease in growth of larvae without, however, causing immediate extensive mortality.
More recently our associates, Chanley and Tubiash, again isolated bacteria from moribund and dead clam larvae. When these pathogens were seeded into cultures of clam or oyster larvae they caused almost

78 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
complete mortality of larvae within 12 hr (Fig. 27). Later, these strains were reisolated from moribund larvae to satisfy Koch’s postulates. These studies are still in progress and it appears that the recent iso- lation deals with different clones from those observed by Guillard.
FIQ. 27. Effect of pathogenic bacteria upon larvae of Mercenaria mercenuriu. Healthy larvae (above) ; larvae from the same culture 12 hr after inoculation (below). Largmt larvae in this photomicrograph are approximateIy 160 p long.
Following recognition of the pathogenicity of fungi and bacteria we began testing a number of antibiotics and fungicides, in addition to using physical measures for prevention of diseases, determine their effects on suryival and growth of larvae (Davis and Chanley,

REARING OF BIVALVE MOLLUSKS 79
1956b). It was found that at low concentrations streptomycin, aureo- mycin, Combistrep (a mixture of dihydrostreptomycin and strepto- mycin sulfate), and Sulmet (sulfamerazine) increased the rate of growth of clam larvae but that, as concentrations were increased beyond certain limits, the rate of growth was retarded and, a t still higher concentrations, the larvae were killed.
The most rapid growth of clam larvae occurred in cultures contain- ing about 100 pprn of streptomycin or Combistrep, in about 33 ppm of Sulmet and in only 3 ppm of aureomycin. The increase in rate of growth of clam larvae in cultures receiving 3 pprn of aureomycin was not as great as in cultures receiving optimum concentrations of other antibiotics, such as streptomycin, Combistrep or Sulmet. The com- mercial preparations of terramycin, sulfathiazole and sulfanilamide were somewhat toxic in minimum concentrations tested and retarded growth of larvae. Combistrep, even at 100 ppm, which is the optimal concentration for clam larvae, retarded the rate of growth of oyster larvae.
In some tests streptomycin did not reduce larval mortalities but, for certain pathogens a closely related compound, Combistrep, in con- centrations ranging from 125 to 200 ppm has proved to be quite effective. Acronize (chlorotetracycline HCl) at 10 pprn and chloro- mycetin a t 20 ppm have also been found effective for certain pathogens, Furthermore, cultures of clam or oyster larvae, routinely receiving 33 ppm of Sulmet with each change of water. consistently showed either better survival or more rapid growth, or sometimes both, than larvae in cultures receiving no treatment. Although at this concen- tration Sulmet did not appreciably decrease the number of bacteria, in some instances, where larvae in untreated cultures grew poorly, treatment with Sulmet improved their growth remarkably. I n these cases, we believe, poor growth of larvae in untreated cultures was caused by toxins produced by bacteria introduced with food cultures. The much better growth of Sulmet-treated larvae, therefore, must have been due either to a reduction in number of the bacteria that produced the toxins or to a removal of the toxin from solution by forming a nontoxic complex with the Sulmet.
We find that, in addition to antibiotics, several substances, including such unrelated compounds as phenol, the insecticides Lindane and Guthion, the weedicide Endothal and such presumably inert substances as silt and kaolin, when present in proper concentrations cause a similar improvement in rate of growth of larvae. We suspect that it is the result of removal of toxins from the water in larval cultures. This is, probably, accomplished either by adsorption on particles, such as

80 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
silt and kaolin, or by formation of nontoxic complexes with large organic molecules of antibiotics and pesticides.
We have also found that routine use of certain chemicals can improve survival of larvae and aid in control of many fouling and competing organisms. One of these extremely useful compounds is polyvinjdpyrrolidone-iodine, known commercially as PW-Iodine. Recently set clams will remain open and feed for at least 24 hr in concentrations of 100 ppm of this compound, and they can tolerate 250 pprn of it for 24 hr, although in this concentration they will remain closed most of the time. We have also found that as little aa 50 ppm of this compound can protect juvenile clams from certain pathogenic bacteria and that, in cultures free from organic debris, 250 ppm of PVP-Iodine will kill bacteria, free-swimming and attached ciliates, some annelid worms and even crustaceans.
Good survival and growth of juvenile clams can also be helped by the following treatment: The cultures are treated overnight, for approximately 18 hr, with 100 ppm of PVP-Iodine. During this time the flow of water is stopped ; the young bivalves, therefore, remain in standing water. Upon completion of this part of the treatment the water is turned on for a period of 48 hr, and then the second 18 hr of treatment with 250 ppm of PVP-Iodine in standing water is applied.
Pentachlorophenol at I ppm, formalin at 40 ppm, and dichlorophene at 10 ppm are other compounds that have been found useful in con- trolling certain bryozoans, ciliates and ascidians that sometimes infest trays or other containers of hatchery-reared juvenile mollusks.
M. Selective breeding and hybridization Development of reliable standard methods for cultivation of larvae
opened fields of studies of heredity of bivalves and their selective breeding. Using these methods i t is now possible to cram individuals showing specific characteristics and attempt to develop s t r a b , or races, of oysters, clams, mussels, scallops, eta, with such desirable characteristics as rapid growth, high glycogen content in their bodies, resistance t o certain diseases and, finally, ability to propagate and exist under less favorable conditions. For example, the ability to reproduce at lower temperatures, say, within the range of 13"-16OC, may extend the oyster beds of such species as C. virginica several hundred miles northward of their present geographical limits.
Davis (1950) and Imai et a2. (1950b) were among the first to start cross-breeding experiments of commercial bivalves. Davis found that crosses of C . virginicuQ x C. gigmd and C . gigas? x C. virginimd showed a lower proportion of eggs developing to veliger stage than did

REARING OF BXVALVE MOLLUSKS 81
non-hybrids. Moreover, neither group of hybrid larvae could be reared to metamorphosis, while under the same conditions non-hybrid larvae of C. virginica and C . gigas grew to setting stage. I n both hybrid crosses fertilization and early larval development apparently progressed normally but about 5 days later, after reaching straight-hinge stage, the larvae died.
More recently, Imai and Sakai (1961) presented results of their extensive studies of crossing different strains of Japanese oysters, C. gigas. According to these authors the hybrids displayed a greater adaptability to environmental conditions than the inbred strains. Imai and Sakai also reported that hybrid oysters can be grown from crosses of C. gigas and C . angulata, while in the case of crossing C . gigas with C . virginica, C. echinata or C . rivularis, cross-fertilization might take place but hybrid larvae will not mrvive.
It has also been demonstrated that two closely related species of the hard shell clam, M. mercenaria, of Long Island Sound and M. campechiewis from the Gulf of Mexico can be successfully crossed (Imsanoff, 1964). Larvae resulting from thew crosses wore often grown to mettimorphoBix, porhaps au a icnult of hybrid vigor, a t rt
faster- t han -normal ra tc. Many thousands of juvenile clams from these crosses, after they
reached sizes ranging from &- to &in, were distributed to a number of marine laboratories along our Atlantic and Gulf coasts for observrt- tions on their survival, behavior and growth under different conditions. Usually, together with the two groups of hybrids, non-hybrid juveniles of M. mercenaria and M . campexhiensis of the uame age aR the hyhridrr were also inclndcd. A number of j)nj)cru, iricliiding thono of Havc:n t t t id
Ari(lr(!WH ( I f)b7), a r i t l (%crnt,niJt, el d , ( I fhj7), cl(:H(:rilwd t h ! otJHcrviit,ionn 011 survivsl a i i c l growth of' these four group of young clams.
Chanley (1961) using the techniques for rearing larvae from crosses of selected clams showing certain peculiarities demonstrated that shell markings of M . mercenaria notata were inherited as a sirnplt: Mc?ndelian character with incomplete dominance. T n cLriot,hr:r mriw of c:xp(:rirnc:iit~ he crossed two unselected clam8 and 11~4et l thc: fantcr growirtg offtqring obtained from that cross as parentH for the next genc?ration. Hr: com- pared growth of progeny of these clam8 with growth of progwiy of t w o randomly selected individuals. When grown urider t h c uaml: corditiorrw for 15 month8 progeny of the fast, growing iriclivitlurtln ww: W x , Irtrpr than progeng of tho rtmclomly Hclnc!tc!cl jlldiVifhlL1H.
V . I~ICAILIN(I ov I ~ ~ W E I C E N T SIWCIEY Since much of the material diacusued earlier in this article was

82 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
based on our observations and experiments on larvae of C. virginica and M . mercenaria, these species will be considered first. However, to avoid repetition of the material already discussed, information on rearing of larvae of t,hese two bivalves will be presented in the form of a comparatively brief review.
A. Crassostrea virginica (Gmelin) A discussion of the literature on various aspects of propagation of
this species has already been given in the earlier part of this article. By using different methods these oysters can be conditioned for
spawning throughout the year and, if necessary, several times annually (Loosanoff and Davis, 1952a). Spawning is induced by raising the temperature within the range from 20" to 32"C, depending upon the previous temperature r6gime. Addition of suspensions of sex products helps to stimulate the spawning. Eggs can be stripped and fertilized.
Eggs of C . virginica vary in size from 45 to 62 p, but the majority measure between 50 and 55 p. Costello et al. (1957) stated that the size of the oyster egg varied between 45 and 54 p. The size of the egg is not influenced by the size of the mother oyster. For example, eggs dis- charged by females 30 to 40 years old and over 9 in long averaged 50.4 p and ranged between 48 and 54 p in diameter, while the eggs from much younger and smaller females, approximately 6 years old and 4 in long, a#veraged about 51 p, the difference being insignificant.
Under favorable conditions 90 to 957" of ripe, properly handled eggs can develop within 24 hr to shelled veliger stage. The smallest normal straight-hinge larva,e measure approximately 68 x 55 p. In 48 hr, at a temperature of about, 22"C, the larvae measure about 75 x 67 p. At this stage they already begin to take in particulate food.
The length of the larvae, measured parallel with the hinge line, continues to be approximately 5 to 10 p greater than the width, measured from the hinge to the ventral edge of the shell, until the larvae reach the size of 85 x 80 p. A t 95 to 100 p the length and width are approximately equal, but after that stage the increase in width is more rapid. When the length reaches 125 to 130 p the width already e x d it by about 10 p. This 10 p disparity between the two measurements remains virtually constant until the larvae reach metamorphosis.
The length-width ratio of larvae of C. virginica is almost linear, except for the section roughly confined to the length between 105 and 130 p where it displays a tendency to form a flat curve. It is of interest that the length-width curve of the closely related species, C. qigae, the Japanese oyster, clearly shows the same tendency.

REARING OF BNALVE MOLLUSKS 83
The majority of larvae of C. virginica metamorphoses between 275 and 315 p in length, but occasionally free-swimming larvae may be 365 p long. Larvae were grown to metamorphosis at temperatures ranging from about 17" to 33°C ; at the latter temperature some larvae metamorphosed in about 12 days. Naked flagellates, such as I. galbana and M . luthpri, are the best foods, whereas some of the green algae with thick cell walls, such as certain Chlorelh species, are not readily utilized by early veligers, especially at comparatively low temperaturea.
Stafford (1912), in his paper on recognition of bivalve larvae in plankton collections, offered a description and drawings of larvae of C. virginica of different stages. His description and, especially, the length-width relationships of the larvae are widely different from ours. For example, Stafford gives the following length-width (depth) measurements, in micron8, as typical for oyster larvae: 82-8 x 69.0;
193.2 ; 310.5 x 276.0 ; and 345.0 x 303.6. A comparison with our data shows that the majority of the above length-width ratios cannot apply to larvae of C. virginica. For example, we found that among the 130 of our larvae measuring 110 p in length the width varied only from 105 to 125 p. I n other words, none of these larvae were only 96-6 p wide, as indicated by Stafford. His discrepancies in length-width ratio are even greater when he gives measurements of larger larvae. Finally, his larvae always measure more in length than in width (depth) although, as has already been mentioned, when a length of about 125 p is reached, the width of the larvae begins to exceed the length by about 5 to 1 0 p arid contiIiuwi t20 do 80 until metamorphorri8. Siillivari'fi ( I ! b t H ) ~ n ( ~ ~ ~ ~ i i l ' ( ~ i i i t r i i t ~ t ~ l r i c t I ~ t i o ( . ~ ) t i ) i r r o ~ ~ ~ ~ ~ t ~ R (if' Iiirvrw of ( '. viq/i i / iui JWI- I ~ I I I ~ t * ~L(~II. ILII*.
'l'hie color o f larvae varies somewhat with the food, but healthy larvae are usually a golden brown, particularly in the area of the digestive diverticulum. Young larvae are usually pinkish-orange en masse, but this color changes to brown and, later, to almost black as the larvae grow larger..
Larvae are highly active and remain in suspension throughout the free-swimming period. Large larvae, 200 p or more in length, have a. tendency to gather on the surface of the water and may appear as small " rafts '' floating just beneath the surface. A pronounced " eye " spot develops, when larvae are about 270 p in length, and is present throughout the remainder of the free-swimming period.
Larvae of C'. virginica may be affected by fungus, rYirdpidium zoophthorum, and die, but they are much less RUHceptible to this disease than larvae of M. mercenaria and some other formH.
110.4 x 96.6; 124.2 x 110.4; 151.8 >( 131.1; 165.6 X 144.9; 207.0 X
7
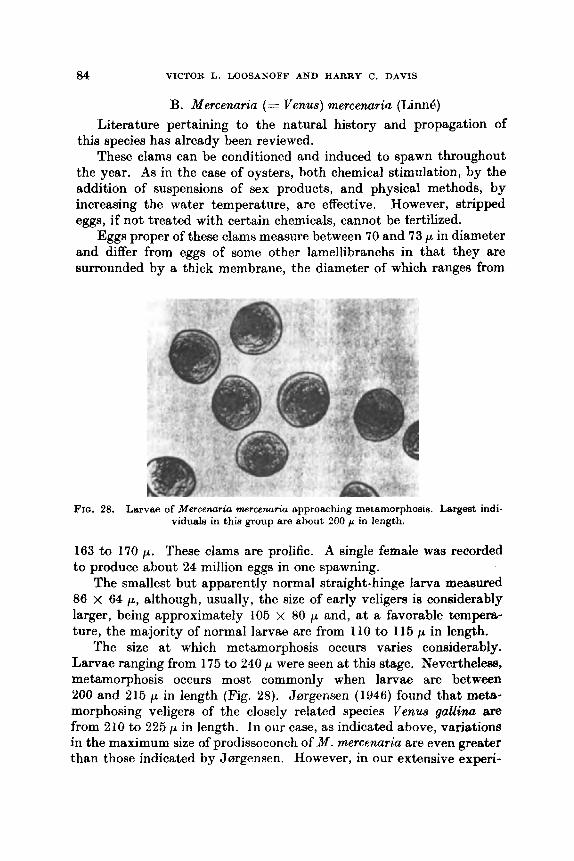
84 VTCTOR L. LOOSANOFF AND HARRY C. DAVIS
B. Mercenaria (= Venus) mercenaria (Linnt5) Literature pertaining to the natural history and propagation of
this species has already been reviewed. These clams can be conditioned and induced to spawn throughout
the year. As in the case of oysters, hoth chemical stimulation, by the addition of suspensions of sex products, and physical methods, by increasing the water temperature, are effective. However, stripped eggs, if not treated with certain chemicals, cannot be fertilized.
Eggs proper of these clams measure between 70 and 73 p in diameter and differ from eggs of some other lamellibranchs in that they are surrounded by a thick membrane, the diameter of which ranges from
FIG. 28. Larvae of Mercemfiu rnercewria approaching metarnorphosie. Legeat indi- viduale in thie group are about 200 p in length.
163 to 170 p. These clams are prolific. A single female was recorded to produce about 24 million eggs in one spawning.
The smallest but apparently normal straight-hinge larva measured 86 x 64 p, although, usually, the size of early veligers is considerably larger, being approximately 105 x 80 p and, a t a favorable tempera- ture, the majority of normal larvae are from 110 to 115 p in length.
The size a t which metamorphosis occurs varies considerably. Larvae ranging from 175 to 240 p were seen at this stage. Nevertheless, metamorphosis occurs most commonly when larvae are between 200 and 215 p in length (Fig. 28). Jsrgensen (1946) found that mets- morphosing veligers of the closely related species Venus gallina are from 210 to 225 p in length. In our case, as indicated above, variations in the maximum size of prodissoconch of M . mercenaria are even greater than those indicated by Jmgensen. However, in our extensive experi-

REARING OF BIVALVE MOLLUSKS 85
ence larvae of these clams have never approached the size of 320 p, as reported by Sullivan (1948). In a letter to us Miss Sullivan explained that she had confused larvae of Pitar morrhuana with those of M . mer- cenaria and ascribes the error to the circumstances that newly set spat of both species were present in her collection (Loosanoff et al., 1951).
Our observations on maximum size of larvae reached before meta- morphosis also show that i t is highly improbable that Stafford (1912) was actually describing larvae of M . mmenaria because he stated that veligers of this clam grew to a size of 448.6 p before setting. Further- more, when the shapes of the larvae shown in Stafford's figures are compared with those of larvae grown from eggs at our laboratory, it becomes clear that Stafford mistook larvae of some other bivalve for those of M . mercenaria.
Food requirements of larvae of M . mercenaria are less restricted than those of some other species, such as C. virginica. Clam larvae can be grown to metamorphosis on a diet consisting almost exclusively of a unialgal culture of Chlorella sp. However, such foods as I . galbana and M . lutheri promote better growth.
Larvae have been grown at temperatures ranging from 15" to 33OC. At the latter temperature setting occurred 6 to 8 days after fertilization.
During early stages the color of the larvae can be significantly changed by differently colored foods. However, as the larvae grow and their shells become darker, this effect becomes less noticeable. Larvae of the same cultures greatly differ in size, especially if they are not properly attended or are too crowded.
Larvae of M . mercenaria are not too selective with regard to the substratum on which their metamorphosis is completed. They meta- morphose in containers made of a variety of substances, including glass, plastics, earthenware, Plexiglas, concrete and several other materials. This is contrary to observations on other forms, such aa echinoderms (Mortensen, 1938) and polychaetes (Wilson, 1948), where i t was found that mature larvae postponed metamorphosis until they located a substratum possessing the necessary physical and chemical characteristics. In our cultures setting usually began when some larvae were approximately 180 p in length, regardless of the size of containers or material of which they were made.
Setting of clam larvae occurred at approximately the same time, whether they were grown in containers from which all light wa excluded or in transparent glass vessels. Turner and George (1955) found that clam larvae were rapid swimmers capable of moving upward at the rate of 8 cm/min. However, they showed no detectable reaction to light.
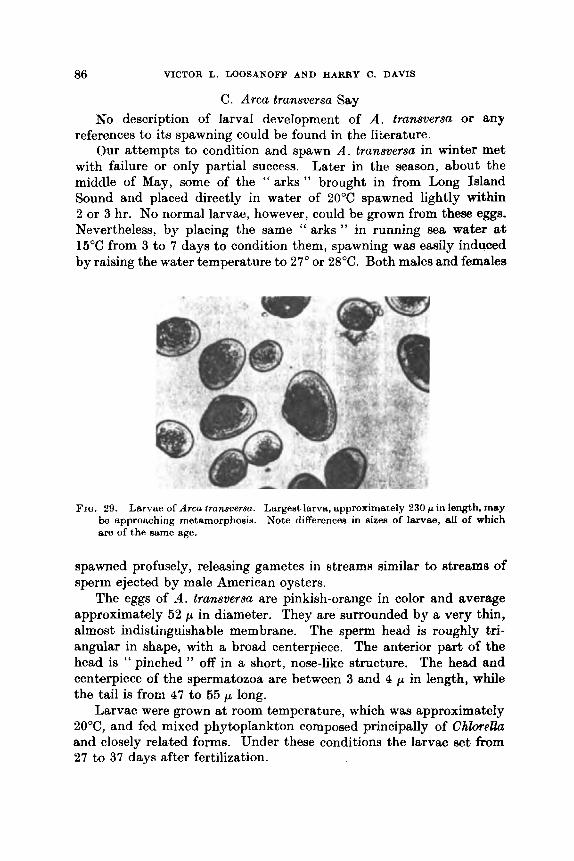
86 VICTOR L. LOOSANOFF A N D HARRY C. DAWS
C . Arca transversa Say No description of larval development of A . tramversa or any
references to its spawning could be found in the 1iLerature. Our attempts to condition and spawn A . transversa in winter met
with failure or only partial success. Later in the season, about the middle of May, some of the “a rks” brought in from Long Island Sound and placed directly in water of 20°C spawned lightly within 2 or 3 hr. No normal larvae, however, could be grown from these eggs. Nevertheless, by placing the same ‘‘ arks ” in running sea water a% 15°C from 3 to 7 days to condition them, spawning wag easily induced by raising the water temperature to 27” or 28°C. Both males and females
FIQ. 29. Larvae of Arca tramversa. Largest larva, approximately 230 /.I in length, m y be approaching metamorphosis. Note differences in sizea of larvae, etl of which are of the same age.
spawned profusely, releasing gametes in streams similar to streams of sperm ejected by male American oysters.
The eggs of ,4. tramversa are pinkish-orange in color and average approximately 52 p in diameter. They are surrounded by a very thin, almost indistinguishable membrane. The sperm head is roughly tri- angular in shape, with a broad centerpiece. The anterior part of the head is “ pinched ’’ off in a short, nose-like structure. The head and centerpiece of the spermatozoa are between 3 and 4 p in length, while the tail is from 47 to 55 p long.
Larvae were grown at room temperature, which was approximately 20°C, and fed mixed phytoplankton composed principally of Chlmellu and closely related forms. Under these conditions the larvae set from 27 to 37 days after fertilization.

ILEAICING OF I % I V A I , V E MOI,I,lJSKY 87
Larvae of A . transversa are quite distinctive in appearance resem- bling, in general, their adults, particularly as they grow larger. The smallest normal straight-hinge larva measured 70 p long and 55 p wide. Thus, even in the youngest larvae, the difference between length and width is quite considerable. However, when larvae are near setting, the length may exceed the width by about 70 p (Fig. 29).
As they grow larger, their color becomes darker and one end of their shells becomes more pointed than the other. Under the microscope larvae appear quite variable in outline because their odd shapes cause them to rest in a variety of positions.
The first indication of an umbo begins to appear soon after the larvae pass 105 p in length, and at 130 p i t usually is quite conspicuous. When the larvae reach approximately 140 p in length the pointed, or less rounded end of the shell becomes distinctly reddish-brown, while the opposite, more rounded end remains almost brown. This color difference is quite pronounced in many individuals.
An " eye " spot appears when the larvae are approximately 225 p long. In some individuals this spot is not too conspicuous at first, hut as the larvae approach metamorphosis i t becomes quite prominent. Slow-growing larvae may develop the " eye " spot at a much smaller size. For example, in at least one case it was observed in larvae that were only 205 p in length.
As is usual in rearing larvae of bivalves, considerable variation was noticed in setting sizes. The smallest, virtually metamorphosed individuals measured only 215 p in length, while one larva in the same culture measured about 310 p and, although it had a large foot, was observed actively swimming. This may have been a somewhat abnormal individual because a majority of the normal larvae in our cultures set when only 240 to 260 p long.
Upon reaching metamorphosis recently set clams develop a strong byssus by which they attach to particles of sand, shells or other material.
Unlike larvae of M . mercenariu and several other species, which are easily affected by fungi, no epizootics that could be ascribed to these organisms were observed in our cultures of A . transversa.
The larvae are dark brown in color.
D. ,Vodiolus demissus (Dillwyn) M . demissus, the common ribbed mussel of our waters, is a180
referred to by some authors as Volsellu demissa, Volsella ylicalula and Modiolus plicatulus. A review of the literature shows that, except

88 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
for Sullivan’s (1948) article, there are no other references to larval development of this species. Considerable literature, nevertheless, exists on larvae of Modiolus modiolus and closely-related forms. These references can be found in the papers of Jmgensen (1946) and Rees (1950).
Various methods used to stimulate spawning of other bivalves proved unsuccessful for ribbed mussels. On two occasions, however, mussels kept in aquaria spawned without artificial stimulation. In both cases spawning occurred at night md, when the eggs were found in the morning they were already undergoing segmentation. Thus, no measurements of recently discharged eggs were made. Jsrgensen
FIG. 30. Larvae of Modwlus derninsue. Note differencee in sizos of larvae that originated from the same group of eggs and were reared in the earno veaeel. Lsrgeet larva in center is approximately 300 p long.
(1946) states that the diameter of an egg of a closely related species, Modiolus modiolus, is about 100 p.
Larvae developing from eggs released during two normal spawnings were reared to metamorphosis. At room temperature straight-hinge stage was reached in about 24 hr. The majority of the l a ~ a e at that time measured from 110 to 115 p in length and 85 to 90 p in width. The smallest normal straight-hinge larva was only 105 p long.
If kept a t a temperature of about 22°C and given good food, such as I . galbana, many larvae reached a length of 200 p by the 10th day. A few, upon reaching this size, developed a prominent “ eye ” spot and functional foot. Nevertheless, although completely metamorphosed juveniles of a size slightly above 220 p in length were observed in some cultures, the majority of the larvae did not lose their vela until they

HEARING O F BIVALVE MOLLUSKS 89
reached 275 p, and a few individuals measuring between 295 arid 305 p in length still retained a functional velum.
While some larvae metamorphosed in about 12 to 14 days after fertilization, many others in the same cultures were still free-swimming when the experiment was discontinued after 43 days (Fig. 30). I n one of these cultures, while there were still free-swimming larvae from 250 to 29.5 p long, there were also several thousand completely meta- morphosed juvenile mussels that ranged up to l mm in length. Why so many of the larvae were still in the free-swimming stage, while others in thc same cultures had mctamorphoscd and showed considerable post-larval growth, remaincd uncxplaitied.
According to Sullivan (1948) the smallest straight-hinge larvae of M . demissus measure approximately 112 x 90 p. This size quite closely agrees with our measurements. However, the maximum size given by Sullivan, 205 x 180 p, is considerably smaller than that observed in our cultures, which was about 303 x 260 p. We also differ with Sullivan on the color of ribbed mussel larvae because she states that these larvae are a deep yellow in the early stages, whereas we found that throughout their development they are brown.
Jarrgensen ( 1946) discussed larvae of closely-related species. In his references to Modiolus modiolus Jsrgensen stated that larvae of this species have not been identified with certainty in plankton samples taken in Danish waters. He thinks that this is probably due to the very close resemblance of this species to veligers of Mytilus edulis. We disagree with Jarrgensen since, in our studies of larvae of these two species, we found them to be distinctly different in their length- width measurements. Furthermore, the difference in their general appearance, especially during later stages of development, is too great to confuse these two groups of veligers.
Rees (1950) stated that in the smallest veligers of Modiolus and Mytilus the shape is similar, although in the former the narrow end is less pointed. Nevertheless, in later stages the shapes of the two genera become more differentiated. Rees also thought that the small specimens of the two mussels are distinguished more easily by their texture than by their shape. He concluded that Jsrgensen’s drawing (Fig. 167d) is probably of Modiolus modiolus, instead of Mytilus edulis, as claimed by Jsrgensen. However, examination of the photomicro- graphs of, presumably, lclodiolus modiolus offered by Rees clearly shows that their shape significantly differs from that of Modiolus demissus grown in our cultures from known parents. This is especially true of his larvae, approximately 250 p in length, which have very little resemblance to our larvae of the same length.

90 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
E. Mytilw edulis Linn6 A study of the literature on lamellibranch larvae shows that
veligers of M . edulis have been described more frequently than those of other bivalves. Borisjak (1909) was probably the first to describe these larvae while, more recently, descriptions of presumably the =me veligers were given by Jmgensen (1946), Sullivan (1948), Rees (1960) and others.
Mussels with apparently morphologically-ripe gonads are found in our waters in early spring, but under laboratory conditions, using our method of maturing animals in warm water, ripe mussels can be made available in January.
Field (1922) reported that rough handling of mussels, such as shaking them in a dish of sea water, will induce spawning within an hour, but Costello et al. (1957) found this method unsuccessful. In our case, during studies conducted for 3 yr, except for brief periods in April and early May, all efforts usually employed to stimulate bivalves to spawn failed to work with M . edulis. These included sharp incremes or decreases in temperature, addition of sex products to the water, keeping mussels in air overnight, placing them in refrigerators for various periods, increasing or decreasing salinity of the water, and even applying light electrical shocks. This was true also of mussels that were conditioned in the laboratory for a considerable time and whose gonads appeared to be morphologically ripe.
Fortunately, i t w4s noticed by two of our associates, first, W. Arcisz, and several years later, J. Uzmann, that some ripe female mussels released large numbers of eggs after their adductor muscles were cut.
These observations suggested a number of experiments on inducing spawning in mussels. The first attempt consisted of opening a large number of these mollusks and placing each individual in a separate vessel. The results were rather gratifying because, in the majority of cases, the mussels discharged large quantities of normal sex products. The method worked even on mussels that were not laboratory con- ditioned. For example, on 26 April 1951, mussels were brought in from Milford Harbor, where the water temperature waa 8°C. The adductor muscles of twelve females were severed and the mussels were placed in water a t room temperature. The first two females spawned within 12 hr; four others deposited masses of eggs within 24 hr, and six remaining females released eggs toward the end of the 48-hr period. This experiment was repeated on several occasions giving virtually the same results.
Eggs released by females with severed muscles were usually dis- charged in " strings ", as occurs in normal spawning. The majority

REAILING O F BIVALVE MOLLUSKS 91
of the eggs released appeared normal, but many abnormal larvae were found later (Fig. 31). However, such larvae were also common in cultures grown from eggs discharged in normal spawnings. Mussel eggs are orange-yellow in color. ‘l’hsy are quite heavy and usually settle quickly on the bottom.
We also noticed that mussels which were dying from some undeter- mined cause often spawned soon after they relaxed their shells, even when their muscles were not cut. This observation suggested that the stimulating center, which induces discharge of eggs or sperm, is prob- ably located in the adductor muscles. If these muscles are abnormally relaxed or stimulated, spawning follows. This reasoning led to another series of experiments designed to obtain eggs and sperm from ripe
FIG. 31. Larvae of Myti lus edulis. Largest larvae are approximately 170 p long. Several small, abnormal individuals can be seen in lower right comer.
mussels without killing them by cutting their muscles. This modifica- tion of the method would prevent fouling of eggs with blood cells, decomposition products, large numbers of bacteria, etc., that usually accompanies collection of eggs from females whose muscles are cut.
The new method consisted of inserting a small wooden wedge between the mussels’ shells, thereby stretching the adductor muscle so that the two shells would be forced open about Q- t o t-in. In experi- ments where this principle was used the majority of pegged mussels spawned, while control mussels kept under identical conditions did not. Some began spawning within 2 hr after pegs were inserted and several spawned three times during a single week.
No mortality was noted among pegged mollusks, even when the pegs remained between the shells for 10 days. Mussels fed in an

92 VIFTOR L. LOOSANOFF AND HARRY c. DAVIS
apparently normal way, forming true feces and pseudo-feces. Mussels that were placed together in large trays with running sea Water even- tually attached to each other by their byssus.
Since the second series of experiments has also showed that dis- charge of spawn in mussels of both sexes can be induced by stretching the posterior adductor muscle, i t became more certain that the nerve locus causing the discharge is located there. Accordingly, i t w&8 decided to stimulate this region, by touching or pricking it with a sharp needle, and note whether this type of stimulation would a h induce spawning reaction.
In the middle of March two groups of mussels, each composed of ten individuals, were placed in running water at a temperature of 19°C. The first group, the control, contained individuals that were not stimu- lated. In the second group each individual waa gently stimulated by pricking the muscle. The first male in the stimulated group spawned in about 2 hr. An hour later the first female spawned and continued to discharge eggs for almost 75 min. The eggs were discharged in orange-colored rods, 3 to 4 mm long. These rods were often expelled with such force that they travelled 4 to 5 in away from the spawning female.
Seven hours after the stimulation another male spawned, and after 9 hr two more females began to discharge spawn. Within 12 hr every individual in the stimulated group had spawned, while none in the control group had done so. Thus, it was demonstrated that needling the adductor muscle is an effective method of inducing spawning in M. edulis.
All mussels that had spawned were later placed in running @a water to see whether they would survive the stimulation treatment. All of them lived for over a month and were then discarded.
Although the previously-described experiment had demonstrated that pricking of the adductor muscle stimulated spawning of mussels, the fact that all ten animals were kept in the same dish left some doubt aa to the exact conditions that caused spawning during the latter Part of the experiment. This doubt existed because toward the end both sperm and eggs were present in the water and, therefore,
have helped to stimulate the mussels that had not yet spawned. matter another experiment was designed in which we
kept each mussel in a separate dish. Again, ten mussels were used in the g o u p and ten in the group that was stimulated. The
'pawned 30 min after stimulation and the first female began to discharge eggs hr after its muscle had been gently touched several t h e s with the point Of a needle. The second female spawned only

REARING O F BIVALVE MOLLUSKS 93
2 or 3 min after the first one. Both individuals discharged eggs in large numbers for a considerable period. Within 12 hr eight of ten stimulated mussels spawned, while none of the control group discharged any sex products.
The experiment was repeated four times involving, altogether, 120 mussels. Of the sixty stimulated mussels, fifty-four responded, while none of the sixty control animals spawned. All mussels of the stimulated group remhined alive for at least 2 weeks, after which they were discarded. Thus, a very simple method for stimulating ripe mussels to spawn has been developed. To make the method even easier we filed a small notch at the edge of the shcll through which the needle was inserted.
According to Jargensen (1946) one of the most remarkable pecu- liarities of larvae of M . edulis is “ i t s excessive variability both as regards the color of the shell, its shape, and stage of development as compared with larval size, in which respects it is not surpassed by any of our other common Lamellibranch veligers ” (p. 287).
Our cultures of M . edulis were grown during the early period of our studies of molluscan larvae ; in other words, before the good food organisms, I . galhana and M . lutheri, became available to us. The larvae were fed an algal mixture, consisting chiefly of Chbrella and other green forms, and grew remarkably well. Strangely enough, i t was virtually impossible to injure them by adding to the water large quantities of food microorganisms, a sitiiation that often arises in culturing larvae of other species, such as clams Snd oysters.
The smallest normal straight-hinge larvae mcasured approximately 93 x 64 p, and the largest, about 300 x 286 p. Thus, our measure- ments are very different from those given by Sullivan (1948), who stated that larvae of M . edulis range in size from 155 x 120 p to approximately 355 x 320 p. The smallest size given by Sullivan is, therefore, almost 60 p greater than that of‘ the smallest larvae in our cultures, and the maximum size is considerably larger than that which we ever recorded among swimming larvae.
Jargensen also apparently mistook other larvae for advanced stages of M . edulis because he speaks of individuals which are 400 p long. Werner (1939), whose length and width measurements of larvae of M . edulis somewhat resemble ours, still fails to give correct measure- ments for the smallest straight-hinge stage larvae. In his growth Curve for these organisms he gives the size of the smallest larvae as approximately 112 p long and 84 p wide, which is considerably larger than the size of the smallest larvae ordinarily found in our cultures.
The so-called “ eye ” spot usually begins to appear in larvae when

94 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
they reach 215 p in length. This spot is usually located approximably in the center of the larvae and measures from 6 to 7 p in diameter. In one individual measuring only 205 p in length a well-defined ‘‘ eye ” spot was found, but this was the only case of an “ eye ” in such a small larva recorded after examining thousands of individuals. When a length of 230 p was reached the “ eye ” spot was present in all larvtle.
The size at which mussel larvae metamorphosed varied by almost 90 p. The smallest metamorphosed individual, attached by a b y m s to an oyster shell laid in one of our culture jars, was only 215 x 201 p. Individuals measuring 225 p in length, but fully metamorphosed and attached by a byssus, were not uncommon. On the other hand, some larvae continued to swim until they were almost 300 p in length. Large variations in the size of M. edulis larvae at the time of meta- morphosis were also recorded by Jmgensen (1946). He thinks that variable size at metamorphosis is not entirely due to the fact that veligers may pass into the bottom stage after attaining a different stage of organization, but also because of the varying ratio between the rate of differentiation of the tissues and rate of growth in different individuals .
The factors which govern appearance of the foot and disappearance of the velum in larvae of M. edulis are rather baffling. In some instances a well-developed foot was observed in animals as small aa 185 p in length. Such cases, however, were not too common and, usually, the appearance of a developing foot occurred as the larvae approached 210 p in length. Most larvae possessed a well-developed foot by the time they reached 230 p in length, but a functional velum has been observed in larvae as large as 288 p.
The great differences in the state of organization attained at a certain size by larvae of M . edulis are also emphasized by Jmgensen. He gives examples of larvae measuring 250 p long and nearing meta- morphosis with well-developed gills and foot and with a velum in 8
reduced condition, while in other cases larvae of the same size will have only a vestige of a foot but a large velum, thus indicating that they are still far from metamorphosing.
The variation in size of larvae at the time of metamorphosis sug- gested a more critical analysis of the data on growth of larvae in the same cultures. Fortunately, among our cultures there was one in which all larvae originated from eggs of the same female and were fertilized by sperm of the same male. Regardless of this, 2 days after fertiliza- tion the larvae varied in length from 93 to 120 p, with an arithmetic mean of 106 p. When the culture was 8 days old the length varied from 107 to 162 p, the average being 130 p. At 14 days some of the

REARING O F BIVALVE MOLLUSKS 95
larvae were already metamorphosing and the mean length was 181 p, but some larvae only 128 p long were still present.
This experiment demonstrated, as we have pointed out on many occasions, that it is obviously incorrect to relate the size of larvae to their age because variations in size, even among larvae that originate from the same parents, are of considerable magnitude. In cultures that are grown at different temperatures and fed different foods these differences are usually even more striking.
Some workers explain variations in size of larval fish originating from eggs produced by the same female as dependent upon develop- ment of these eggs while still in the ovaries, including proximity of an egg to large blood vessels which provide the best feeding conditions for a single egg. Others explain the different rates of survival and growth of fin fish larvae as dependent upon quantities of yolk in different eggs. Possibly, the same considerations hold true and, in part, explain variations in rate of growth of bivalve larvae that originate from eggs discharged by the same female in the same spawning.
As is usually the case with larvae of most bivalves, their color varies greatly in accordance with the color of the food microorganisms that they are able to obtain from water. Larvae of M . edulis given heavy doses of purple bacteria rapidly acquired a pinkish color, which was most pronounced in the area of their digestive organs. If these larvae were later fed green Chlorellu, their coloration changed to a greenish tint and the livers became dark green. Contrary to Sullivan’s (1948) contentions, no purple tint was observed around the shoulders of the shell, even in advanced larval stages.
F. Anomia simplex D’Orbigny A . simplex is a common bivalve of Long Island Sound, where it
propagates a t approximately the same time as C . virginica. Because of its more rapid growth immediately after setting, however, this mollusk is a dangerous competitor, since it overgrows and completely covers large numbers of recently set oysters (Loosanoff and Engle, 1941).
Descriptions of larvae of several species of Anomia are given by students working in widely separated parts of the world. Stafford (1912) gave a description of A . acubata; Miyazaki (1935) described the larvae of the Japanese species, A . Zischkei, and Lebour (1938), as well as Jmgensen (1946), described those of A . sqwlmula. More recently, Sullivan (1948) also described larvae of A . aculeata, while Rees (1950) made a brief reference to three species of Anomia. One

96 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
of these, A . ephippium, is a synonym for A . simplex. Other refer- ences can be found in the above-mentioned articles of Jrargensen and Sullivan.
Large A . simplex can be conditioned in Deoember and induced to spawn during the first part of January. Some conditioned groups were induced to spawn several times between the middle of January and the beginning of August. To induce spawning presented no problem because, usually, ripe animals began to spawn as soon aa the tem- perature of the water was raised only a few degrees above 20°C. Even in summer i t was not necessary to raise the temperature much above 25°C. In many instances spawning began without addition of any sex products, a circumstance which is unusual in our experiences with spawning of other bivalves.
Artificial fertilization of A . simplex eggs is possible. Stripping can be easily accomplished by gently washing the light yellow gonad tissue of a female in sea water. The eggs separate easily and many remain uninjured. Therefore, a high percentage of them ordinarily develop into normal larvae.
Compared with eggs of other bivalves, eggs of A . simplex are com- paratively small, ranging only from 42 to 45 p in diameter. Because of their small size the eggs are difficult to handle, as they easily pase through the mesh of the smallest screen used for egg retention. Becauee eggs and early larvae cannot be strained and, thus, separated from the surrounding water, as is done with larger eggs, in fertilizing eggs of A . simplex care should be taken not to use too much sperm so as not to pollute later cultures of developing eggs and early larvae.
In our cultures of A . simplex the smallest larva with completely formed shells measured approximately 58 x 47 p. Young larvae are more fragile and transparent than those of C. virginica of corresponding stages. The " straight-hinge " is actually sloped about 5" so that i t is somewhat convex. This is difficult to observe under a microscope unless larvae are seen precisely in profile.
The most common size at which larvae of A . simplex metamor- phosed was at a length of 195 to 210 p. The largest swimming larva8 measured approximately 215 x 220 p. We cannot compare the dimen- sions and shapes of our larvae with those given by Stafford (1912) because his drawings and measurements bear little resemblance to the larvae of A . simplex in our cultures.
Sullivan ( 1948) gave the smallest size of A . acuEeata aa 1 10 x 100 p, and the largest as 285 x 280 p. Jrargensen (1946), in referring to his own observations on A . sqwzmuh, reported that the smallest larvae measured 65 to 75 p in length, and the largest attained a length of

REARING OF BIVALVE MOLLUSKS 97
260 to 270 p. Lebour (1938) stated that A. squumula may metamor- phose at a length of only 180 p, which coincides with our observations on A. simplex, while Stafford maintained that the veliger of A. aculeata is still pelagic a t about 350 p.
The size of larvae of A. simplex, when the “ eye ” spot appears, is difficult to indicate. In a few cases the ‘‘ eye ” was observed in indivi- duals measuring only 136 p in length although, as a rule, it appeared when the larvae were near 160 p. The “ eye ” seems to be composed of from four to six dark bodies grouped together. In many individuals, including some that were near Betting, no “eye” spot could be distinguished.
Even in early straight-hinge stages there is considerable difference in the shape of the two shells. The right shell, or the one that will
FIQ. 32. Larvae of Anomia simplex. .Largest larvae in photomicrograph are about 160 p long. Note transparency of larval shells.
eventually be the upper, is conspicuously rounded, while the left, or the one that will become the lower shell is virtually flat. This distinc- tion remains throughout larval developmen t. J s r g e n ~ n ( 1946) reported that in A. squamula the umbo of the right shell is not developed. We made the same observation on the shell of A. simplex.
An observation made during the rearing of larvae of A. simplex concerned the inconsistency of the appearance on its shell of the inden- tation known as the byssus notch which, presumably, in diagnostic of larger larvae of this species. Since this was one of the features which we expected to be rather prominent and to appear in all older larvae, we were somewhat perplexed not to find this notch in many individuals. It is true that in some individuals this notch was present

98 VICTOR L. LOOSANOW AND W Y C. DAVIS
in larvae only about 180 p long, but often it was only a thickening of the shell edge on the side opposite the foot. In our extensive studies of living material and numerous preparations of larvae imbedded in balsam, in permanent slides, we were unable to find such " deformed " larvae as those shown by Sullivan (1948).
Early larval stages of A. simplex are very light, almost trans- parent in color (Fig. 32). Even larvae nearing metamorphosis remain silvery transparent except, of course, their digestive organs which, aa a rule, take on the color of the food that they contain. In this respect our observations are rather different from those of Sullivan (1948), who stated that, at first, Anomia larvae are pale yellow but, later, become deep yellow, except for the viscera, which are dark gray.
Since larvae of A. simplex may be mistaken for those of oysters, especially in advanced stages when umbones in both species are quite prominent, one way to distinguish the two species is to remember that in A. simplex the digestive organs are situated much higher, lying almost under the umbones.
I n the early stages of our work on cultivation of A. simplex larvae, which was conducted in 1950 and 1951, the only food that was regu- larly available waa a mixture of green algae composed largely of Chlorella. Although the larvae reached metamorphosis when fed this food, their growth, as a rule, was slow and mortality heavy. Later, when cultures of I. galbana and M. lutheri became available, the larvae responded well to the new food, growing much faater and showing lower mortality. When fed these flagellates at room temperatures, many larvae of A. simplex began to metamorphose after the 12th day although, in the same cultures, some larvae were still swimming 33 days after fertilization.
As already mentioned, one of the most interesting observations made on cultures of A. simplex was what appeared to be partial meta- morphosis, or metamorphosis without attachment to substratum (Loosanoff, 1961). Partial metamorphosis was characterized by disappearance of the velum, but retention of a functional foot. More- over, these partially metamorphosed individuals were unable to attach to the substratum, and their shells showed a distinct demttrca- tion line between larval and post-larval portions.
G. Pecten irradians Lamarck The early embryonic development of the bay scallop, P. iwadians,
also called Aequipecten irradians, has been described by a number of authors, including Fullarton (1890), Drew (1906), Belding (1910),

ItEAltINU OF BIVALVE MOLLUSKS 99
Gutsell (1930) and, more recently, by Costello et al. (1957). The later development of the larvae, beginning with the formation of the straight- hinge stage, however, has never been adequately described because virtually no one succeeded in growing these larvae to metamorphosis. The only exception was Wells (1927)) who grew scallop larvae past setting stage, but gave only a superficial description of them, although he aupplkd good photographs showing the different stages of development.
Descriptions of eggs and early stages of several European Pectinidae were offered by Jmgensen (1946). Efowever, as in the case of the American bay scallop, virtually nono of the European workers, except Odhner (1914), gavc photomicrographs of older veliger stages that may be helpful in identifying the larvae.
The scallops used in our studies were brought, in February, from Long Island, where they are found in a number of bays, harbors and inlets. They were placed in running sea water, the temperature of which was increased within a few days to 20°C. The first spawning was attempted after 23 days of conditioning. The scallops responded readily to a temperature stimulus of about 30°C and spawned profusely.
Scallops usually spawn first as males and, later, as females. During one spawning, fourteen scallops started spawning as males. However, 10 or 15 min later seven of these fourteen individuals began to discharge eggs, but within a few minutes two of these seven again reverted to spawning as males, releasing large numbers of spermatozoa. On one occasion the same scallop was observed releasing sperm and eggs simultaneously.
Scallop eggs obtained in our experiments measured from 5 5 to 65 p in diameter, the average size being near 60 p. Belding 11910) and Gutsell (1930) gave the size of the scallop egg as 63 p. Fullarton (1890) reported that the egg diameter of a rolntetl European species, Paclen opercularis, is 68 p. The scallop egg is surrounded by a thin membrane and is usually of a pale orange color. Brief remarks on fertilization, cleavage and rate of development of the fertilized ovum were given by Costello et al. (1957), who based his descriptions on the works of Belding ( 19 10) and Gutsell ( 1930).
The smallest straight-hinge scallop larvae observed in our cultures were 80 p long and 65 p wide, and the largest swimming individuals, approximately 200 x 195 p. Some larvae metamorphosed upon reaching 175 p in length. Jmgensen (1946) stated that the largest planktonic stage of the European bcallop, P. opercularis, is approxi- mately 250 to 260 p long, while the larvae of another European scallop, P ~ c t ~ n striatus, approaching metamorphosis may be only 200 to 2 10 p
8
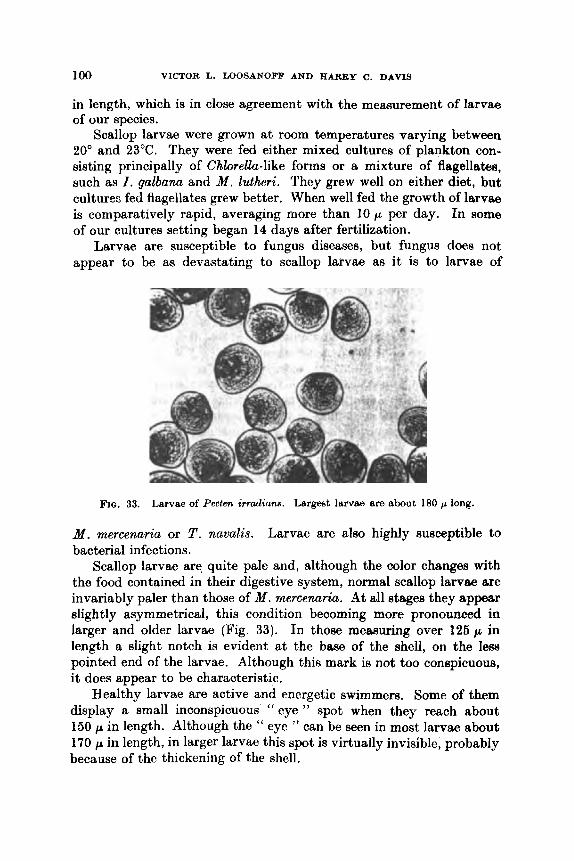
100 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
in length, which is in close agreement with the measurement of larvae of our species.
Scallop larvae were grown at room temperatures varying between 20” and 23°C. They were fed either mixed cultures of plankton con- sisting principally of Chlorella-like forms or a mixture of flagellates, such as I . galbarn and M . luthari. They grew well on either diet, but cuiturss fed flagellates grew better. When well fed the growth of larvae is comparatively rapid, averaging more than 10 p per day. In some of our cultures setting began 14 days after fertilization.
Larvae are susceptible to fungus diseases, but fungus does not appear to be as devastating to scallop larvae as i t is to larvae of
FIQ. 33. Larvae of Peclen irrdionn. Lergeht larvee are about 180 p long.
H. mercenaria or T . navalis. Larvae are also highly susceptible to bacterial infections.
Scallop larvae are. quite pale and, although the color changes with the food contained in their digestive system, normal scallop larvae are invariably paler than those of M . mercenaria. At all stages they appear slightly asymmetrical, this condition becoming more pronounced in larger and older larvae (Fig. 33). I n those measuring over 126 p in length a slight notch is evident a t the base of the shell, on the lem pointed end of the larvae. Although this mark is not too conspicuous, i t does appear to be characteristic.
Healthy larvae are active and energetic swimmers. Some of them display a small inconspicuous “eye” spot when they reach about 150 p in length. Although the ‘‘ eye ” can be seen in most larvae about 170 p in length, in larger larvae this spot is virtually invisible, probably because of the thickening of the shell.

REARING OF BIVAI,VE MOLLUSKS 101
H. Ostreo edulis Linni5 We are interested in this species because its northern races prop-
agate at somewhat lower temperatures than the American oyster, c. virginica. Because of this we imported 0. edulis into the United States in 1949 in the belief that certain areas along the shoreline of Maine and our Pacific coast states were promising for natural propaga- tion of these oysters (Loosanoff, 1951, 1955, 1962b). Since then, 0. edulis have grown and propagated naturally in New England waters and have become established in Maine. Young oysters reared at Milford, Connecticut and sent to the State of Washington have also grown well there.
Literature on the sexuality and propagation of the European flat oyster, 0. edulis, has been well reviewed by Orton (1937), Korringa (1941), Walne (1956) and Yonge (1960) and, therefore, need not be discussed here in detail. Using our methods for ripening mollusks out of season we have obtained larvae of 0. edulis from the end of January to the end of the normal spawning period which, in Milford Harbor, extends into September. Our attempts to induce spawning of ripe oysters were not always successful. On several occasions, nevertheless, by raising the temperature of the water containing conditioned oysters and simultaneously adding a suspension of gonad material, spawning has been initiated in both sexes. This was ascertained by finding sperm balls or recently discharged eggs on the bottom of the aquaria. Well-developed eggs ranging in size from 114 to 126 p were often lost when, during the process of spawning, they were released from the gonad to enter the gill chamber where, normally, they are incubated until larvae are released a t swarming.
The manner in which eggs are lost was observed on several occasions during normal spawnings. These eggs were released from the exhalant chamber in a thin stream, almost in ‘‘ single file ”. Because only a weak current was carrying them, the eggs settled next to the shells of the mother oysters, eventually forming a small mound. By a gentle moving of the spawning oysters from one area of the aquarium to another and observing the formation of new piles of eggs, i t waa established that a spawning female may continue a steady discharge of eggs for as long as 4 hr.
Since we could not always induce spawning in conditioned oyntere, we depended quite often on unprovoked normal spawning8 and subsequent release, or swarming of the larvae. Therefore, when a gravid female was detected, by the presence of a small pile of eggo near its excurrent side, i t was placed in a separate aquarium with aerated standing water and kept there until larvae were released. The

102 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
water was changed daily and the oysters were given sufficient quantities of food to keep them and the larvae alive.
At room temperatures swarmings took place from 6& to 10 days after spawning. The size of larvae released by a single female in the same swarming varied considerably. On one occasion samples of 100 larvae were collected from swarms released by each of six females which had been kept in separate aquaria. Larvae released by female (A) ranged in length from 142 to 199 p, with a modal size of 186 p ; by female (B), from 164 to 192 p, with a modal size of 184 p ; by female (C), from 164 to 203 p, with a modal size of 188 p ; by female (D), from 149 to 203 p, with a modal size of 187 p ; by female (E), from 164 to 192 p, with a modal size of 180 p ; and female (F), from 164 to 195 p, with a modal size of 183 p. Occasionally, broods of smaller larvae measuring 175 x 160p were released. The average size of such broods closely resembled those reported by Boury (19281, Voisin (1931) and Korringa (1941).
In rare cases the modal sizes of larval broods were unusually large, over 200 p in length. Because all female oysters incubating larvae were kept a t the same temperature, it is improbable that the difference in size of the liberated larvae of the different broods was governed by the temperature a t which incubation occurred. It seems more reason- able that size of larvae at liberation depends upon how long larvae are retained in the mantle cavity. This may be governed by the physio- logical condition of the parent oysters or the conditions in the water in which oysters are kept, the larvae being retained longer and, hence, are larger at time of release when these conditions are not at their optimum. This is supported by observations on liberation of larvae by two females that appeared sick. The first was used in an experiment where induced spawning was attempted by quickly raising the temperature to 37°C. The oyster did not spawn but was apparently injured by this treatment because it gaped for several hours after its return to sea water a t room temperature. It did spawn the next day, however, but the spawning was not normal because too many eggs were found on the bottom of the aquarium near the oyster. The eggs, nevertheless, were fertilized.
Ten days later this oyster discharged, simultaneously, a brood of larvae and many empty larval shells, indicating that many larvae hacl died within the mantle cavity during incubation. There was also a considerable number of abnormal larvae with defective vela. Normal larvae, which still were in the majority, measured from 199 to 212 p in length, with a modal size of about 2 0 8 p . Regardless of some abnormality observed in the development of this brood, the larvae were reared to metamorphosis.

ItEARINO OF BIVALVE MOLLUSKS 103
The second release of liirger-thari-;ivc.raRc larvae, measuring between 200 and 210 p i n length, was also observed in an oyster that was kept at high temperature for several hours and was probably injured by this treatment.
Another possibility is that the size upon release may be governed by the numb of incubating larvae in the mantle cavity, with the larvae growing faster if the riumber is small.
Swarming or, more correctly, releasf) of larvae by parent oysters was observed on numerous occasions. 1 t i general, i t resembled dis- charge of eggs by a spawning female of C virginica. The larvae were expelled in cloud-like masses by a forccful closing motion of the shells. Discharge of larvae by a single female niay continue for 3 or even 4 days. Usually, during the 1st day only a few larvae are released. The chief masses of larvae are expelled during the 2nd day and then the number of newly released larvae sharply decreases. Larvae that had just been released usually contained in their stomach large quan- tities of algal cells which had been added to the water during their incubation.
In many of our experiments, including the early one when larvae were fed phytoplankton composed principidly of Chlorella, the veligers grew and survived well. At temperatures ranging between 18" and 20°C " eyed " larvae measuring approximatrely 270 p, with a prominent pigmented spot, began to appear about the 9th or 10th day. Setting usually began on the 15th or 16th day, altltough in several cases. when fed I . galhana and M . lutheri, the larvae began to set between the 7th and 8th days.
Setting size of the larvae in our cultures was most commonly between 280 and 300 p. In this respect, (aur ohrva t ions agree with those of Cole (1!+3$)), who reported t,hat metamorphosing larvae in his experiments measured from 290 to :rIO p. However, we have never observed larvar as large ns 350 p which Cole found in some of his tanks. Imai et 01. (1953) reported that none of thc larvae of 0. edulis in their cultures reached 300 p in width, and most of the individuals in Walne's ( 1 956) broods also set before reaching 300 p.
Throughout the experiments the rate of growth of individual larvae of the same troods showed considerahle v<triations. For example, at the time of srttitig, ~ h c n some larvae i i i ;t culture already measured 300 p in length and were undergoing mrtamorphosis, other larvae still measurcd only about 175 p, thus bciiig of approximately the same size as many larvae at the time of swarming.
When recently released larvae were plac*ed in culture jars many of them ascended to the surfact. and formed la~ge, brown, floating masses,

104 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
often measuring several square millimeters in area. This congregating tendency apparently did not unfavorably affect larvae. If these m a w s of larvae were disturbed, the larvae would separate and continue to swim for some time but, later, would again form floating groups. Similar floating masses of larvae are frequently observed among older larvae of other species, including C. virginicu.
I. Ostrea luridu Carpenter The native oyster of our Pacific coast is the Olympia oysfer,
0. lurida. It is considerably smaller than its close relative, 0. duli8, or the Atlantic coast oyster, C. virginicu, seldom exceeding a length of 23 in. Unlike C. wirginica which, when adult, are of separate sexes, the Olympia oyster is hermaphroditic. Its reproduction is also different from C. virginicu in that the eggs are not discharged directly in the water but, as in 0. edulis, they remain within a special brood ohamber in the mantle cavity of the mother oyster.
Gonad development and spawning of 0. lurida can be induced out of season by keeping them in water at about room temperature for several weeks. Spawning can be detected, as in 0. edulis, either by clouding of the water with discharged sperm or by presence on the bottom of a few eggs that were lost during spawning. The eggs measure from 100 to 110 p in diameter.
According to Coe (1931) Olympia oysters begin to spawn when the temperature reaches 16°C. Hori (1933) indicated the temperature as 14OC and Hopkins (1937) thought that spawning occurred soon after the minimum daily water temperature reached 13°C.
In our experiments the incubation period lasted from 7 to 9 days after which swarms of larvae were discharged in the water. Stafford (1914) estimated that in 0. lurida the period between spawning and swarming was 164 days. Coe (1931) reported this period aa 10 to 12 days, and Hopkins (1937), as about 10 days. The difference in time is probably due to environmental conditions, especially temperature, under which the different observers conducted their studies.
We were unable to rear successfully recently fertilized cggs that were occasionally lost by females. However, embryos that were lost after they had reached the ciliated blastula stage were cultured to straight- hinge stage,
In one of our experiments larvae as small as 160 x 149p were relertsed at swarming, but these larvae appeared immature as they were too transparent and unable to withdraw the velum completely. It was possible, nevertheless, to rear even these small larvae to meta- morphosis.

NEARING OF BIVALVR MOLLUSKS 106
The size of normal larvae, at the time oS swarming, waa about 185 p, but on several occasions groups of smaller larvae, measuring from 165 to 170 p were released. Hori (1993) reported that larvae at the time of release are between 175 and 185 p. Hopkins (1937) gave 180 p as the average. Imai et al. (1954), in culturing 0. Zurida in Japanese waters, fonnd that the length of larvae immediately after swarming ranged from 174.6 to 189 p. In general, all these figures closely agree with ours.
The larvae are quite dark, even when first releaaed, and appear black when seen en m s e (Fig. 34). When given good food and kept at a temperature of about 24°C growth of these larvae is quite rapid, some of them beginning to set only 7 days after swarming. At 18'C,
FJQ. 34. Larvw of Omea lur ih . 1,argest h v a . rmar cerlter, is 204 x 185 p.
however, setting does not begin until thfn 16th day (Davis, 1949). Imai et al. (1954) reported that in their best cultures setting began on the 10th or 11th day at temperatures averaging 21.8OC.
Unlike larvae of the genus Crassostrea, larvae of 0. lurida can utilize Chlorelln from early straight-hinge stage and, in general, appear to be less restricted in types of phytoplankton organisms that they can utilize as food.
Larvae of 0. luridtr., particularly the 1argc:r ones, when seen under a microscope appear as thick wedges. Individiials 250 to 300 p in length may measure as much as 'LOO p in thickness Lf measurements are taken near the hinge, tapering rapidly to the sharp ventral or " bill " side. This makes i t difficult to obtain accurate wiclth measurements.
The setting size of larvae grown in our laboratory cultures was approximately 300 p, which is not significai ttly different from setting

106 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
sizes of larvae of C. virginica and C. gigas. No free-swimming larvae measuring more than 312p in length were observed. Growth after metamorphosis was, for a short period, somewhat faster in 0. lutida than in G. virginica.
J. Crassostrea gigas (Thunberg) C. gigas, the common commercial species of Japan, was introduced
to the waters of the State of Washington in 1902. During the last few decades it has become one of the most important mollusks of our Pacific northwest. Although the industry still depends largely upon seed oysters imported from Japan, a natural set of C . gigae occurs occasionally in such areas as Quilcene, Willapa and Dabob Bays and Hood Canal.
FIG. 35. Larvae of Crassoslreu q i p a . Largest larva, near center, is approximately 175 /L long.
Extensive studies conducted by Japanese workers on early develop- ment and propagation of C . gigas are reviewed at length by Cahn (1950). We may add to this review the articles of Hori and Kumkabe (1926) describing their successful efforts in growing larvae of C . gisQs. More recently, Imai and Hatanaka (1949) and Imai et al. ( 1 950b) grew larvae of these oysters on a much larger scale.
Our experience with C. gigas received from the State of Washington showed that their spawning behavior, egg development and the s h a p of their larvae seem to be identical to those of C . wirginica (Fig. 35). The similarity in shapes and sizes of the larvae of these two species is so great that we doubt if even an experienced investigator, familiax with both species, would be able to differentiate them in a mixed sample,

REARING O F BIVALVE MOLLUSKS 107
The general behavior, food requirements and growth rate of larvae of C. gigas also appear to be identical to those of larvae of the American oyster. Even the size of the eggs arid the size at which larvae of the tmo species begin to metamorphose are alike. A comparison of the length-width relationship of larvae of the two groups will still further indicate their similarity. Yet, regardless of this similarity, the attempt8 of Davis (1950) and Imai et al. (1950b) to hybridize these two species were unsuccessful. Although fertilization occurred and the larvae developed into veligers, they all died before reaching metamorphosis.
Gonad development and spawning of Japanese oysters kept at Milford have not been as thoroughly investigated as those of the American oyster. Nevertheless, there appear to be several important differences. First, Japanese oysters require higher temperatures to begin spawning. Groups of these oysters kept in the laboratory all summer did not spawn, although the temperature at times was over 26°C. Furthermore, C. gigas either does not resorb the gonad material that remains undischarged at the end of the spawning season or does so at a much later date and at lower temperatures than C. virginim. Japanese oysters, brought to Milford from Piiget Sound in October with undischarged gonads, held this material far into the winter and active sperm was found in some oysters late in December.
In our waters growth of post-setting stages of Japanese oysters is much more rapid than that of the local species. C. gigas that set in the laboratory in the spring and then were planted in Milford Harbor were 25 to 3 in long by the end of the growing season, while native oysters, which had set, at the same time and were grown under identical conditions, were only 1 to 1 4 in long.
K. Lnevicardium wwrtoni (Conrad) This bivalve, which is called Cardium nmrtoni by some authors and
is also known as Morton's cockle, is quite common in the compara- tively shallow waters of Long Island Sound. If brought into the laboratory in late January and conditioned at temperatures between 15" arid 20°C for several weeks, these cockles are easily induced to spawn by temperature stimulation alone. On several occasions spawning of ripe cockles began in less than 5 min after the water temperature was raised only 1" or 2" above 20°C. The majority spawned within fr hr and, &s a rule, all began spawning before the water reached 30°C. Some cockles spawned at least three times during a single week.
The hermaphroditic cockles often acted first as males and then as females in the same spawning. Sometimes, after first discharging sperm, they released large quantities of eggs and finally ejected sperm

108 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
again. Probably because of this condition, which resulted in lmge quantities of sperm in the spawning dishes, many cases of polyspermy were seen.
The eggs of L. mrtoni measure from 60 to 65 p in diameter. Lov6n (1848) reported that eggs of the related species, Cardium exiguum, have a diameter of 64 p and are deposited in a thick gelatinous mantle within which development goes on until the shell-bearing veliger hss attained a length of 90 p. Orton (1924), who performed succeesful artificial fertilization of a closely related species, Cardium fawidurn, and Jmgensen (1946) reported that eggs of that bivalve messure about 80 p in diameter, thus being considerably larger than those of our species. In still another closely related form, Cardium ed&, according to Lebour (1938), the egg diameter is only 50 p.
The smallest normal straight-hinge larvae in our cultures measured about 85 x 70 p. Sullivan (1948) stated that very small larvae of Cardium pinnulaturn are 90 x 80 p, a size closely corresponding to our smallest normal larvae. In early stages the margin of the larval shell is outlined by a dark band, similar to that seen in Teredo larvae, but not quite as prominent. Usually, one end of the shell is slightly more pointed than the other.
The umbo begins to appear when larvae are between 135 and 150 p in length and is usually quite prominent when a length of about 160 p is reached.
The larvae were grown at room temperature of about 20°C and fed a mixture of I . galbana and M . lutheri. The first metamorphosing individuals were observed 8 to 10 days after fertilization. The smallest individual, having a definite foot but no velum, measured only 205 p, while the largest swimming larvae with a still well-developed velum measured 245 p. Some metamorphosed cockles with well-developed gills and a siphon were only 220p long. No “eye ” wafi observed in any of the larvae.
The size of the largest swimming larva ever recorded in our cultures was 250 x 220 p, but only one such individual was seen. The maximum size of our larvae, therefore, closely agrees with that described by Sullivan (1948) for C. pinnulaturn, which measured 250 x 230 p. Jarrgensen (1946) mentioned that Lebour grew larvae of Cardium echinatum which, at metamorphosis, measured about 480 p in length. We accept this figure with considerable reservation. Jmgensen also assumed that the maximum length of larvae of C. exiguum is about 250p: although he never observed this veliger with certainty in the Sound, where he carried on his experimenta. His observation on C. fasciatum indicated that the largest larval stages

REARING OF BIVALVE MOLLUSKS 109
of this species found in plankton were about 300 p long. However, he d d e d that the veliger may leave the plankton at a length of about 256 p, which is not too different from our maximal size.
Jergensen (1946) also offered extensive observations on the length of larvae of C. edulp. He stated that in rearing experiments at Kristine- berg the length at metamorphosis varied from about 275 to 345 p. Length measurements of prodissoconchs in oyster ponds near Limfjord varied from 200 to 275 p, averaging 240 p. Jergensen was under the impression that larvae of this species are distinguished by an excessive variation in size a t the time of metamorphosis. This condition coincides with our observation that usually in cultures of larvae approaching metamorphosis there is a great variation in size. For example, in our
cultures of L. mortoni, while some of th(: larvac: had a1rc:mly wt arid others approaching motarnorphosin mcaxurc4 245 / 805 p, thtr nrnrrlle~t individualR in thc samc! oulture, growit titidt:r itl(:tit,ical coii(Jit,ions, were otily 140 x 120 p.
J,. Mercenariu (== V e n w ) canipechienoiu (Gmelin)
1 heso ohins, cornrnoiily knowti ILH noullic:rri ( l l I t thO#R, ww(! Im)tight to Miffortl f k o m t h ( iulf of M(:xic:o, iiwr itliriluahic:olrL, I4orida. Although thcv q p : a r to differ wrnc!wtirht from northcwr quahogu, Mercenaria (=-: Yenw) mercanarin, thcir spawriing behavior anti appear- ance of their eggs and larvae are identical and, therefore, they will not be described (Fig. 36).
On several occasions groups of M. m,ercrnariu and M . cumpechiensis
t l

110 VICTOR L. LOOSANOFF AND HABEY C. DAVIS
were conditioned for spawning and later induced to spawn simul- taneously but in separate dishes. The eggs obtained from theae spawnings were cultured under identical conditions. Observations showed that egg development and rate of growth of larvae of thee two species were virtually identical throughout the experiment and that metamorphosis in both groups began at the same time.
Reciprocal crosses of the two species produced viable larvae, which grew to metamorphosis; a t times these hybrids showed somewhat more rapid growth, possibly as a result of hybrid vigor. bhny of these hybrids have been grown to maturity in several locations along our South Atlantic coast and also in the Gulf of Mexico, but not in cold axeas, such as Milford Harbor, where the low water temperature in winter usually caused their death.
M. Tapes semidecussata Reeve T . semidecussata is one of the most important commercial species
of clams in Japan, where it is known under the common name of Assri. It was accidentally introduced to our Pacific coast when Japanese oysters, C. gigas, were brought there early in the century. It spread rapidly and is now one of the most important speciea of clams in the State of Waahington, where i t is called the Japanese little neck, or Manila clam. In the literature T. semidecusslata appears under a variety of names, including Venerupis semidecussata, Venerupia philippimmm, Paphia p h i ~ i p p ~ ~ ~ m , Tapes japonica, and others.
The fist group of T. sernidecussata was received in Milford from Puget Sound in December 1955. Upon arrival the clams were con- ditioned at about 20°C for 2 to 3 weeks and then induced to spawn. Cultures of larvae were successfully grown from January until the end of the summer. According to Cahn (1951) the Spawning period of T. semidecussata of the Hokkaido region of Japan extends from June to late August, when the water temperature is betiveen 20" and 23°C.
In our spawning experiments ripe clams quickly opened and pumped vigorously when the temperature of the water was increased to about 25°C. However, raising of the temperature even to 30°C waa often insdiicient to induce spawning, and addition of sperm or egg suspension waa usually necessary. Even then, on some occasions, although their gonads were filled with mature gametes, the clams did not respond to the combination of stimuli.
Spawning usually occurred at temperatures between 20" and 27.5"C. The act of spawning closely resembled that of M. mercenwia.

REARING OF BIVALVE MOLLUSKS 111
Sperm or eggs were released in a constant stream from the excurrent siphon, sometimes continuously for 30 min or longer.
Eggs of T . semidecussata range from GO to 7 5 p in diameter, in- cluding the transparent membrane around the eggs that measures from 5 to 10 p in thickness. The average diameter of the egg, including the membrane, is approximately 6 9 . 6 ~ . According to Cahn (1951) .Japanese workers found that eggs of this clam memured between 63 and 6 6 p in diameter with a perivitelline space measuring from 3-3 to 4-6 p in width and surrounded by a gelatinous coating approxi- mately 23 p in thickness.
Most of the cultures of T . semidecussata were grown at room tem- perature. They were fed a variety of algal cultures, but no systematic
PIC:. 37. Larvae of Tape8 sam~dem.88ala. Largest larvae in photomicrograph are approximately 17.5 p long.
studies of their food preference were made. Nevertheless, it wm noticed that, while sometimes they grew well on mixed green algae consisting mostly of Chlorella, at other times, when given practically the same food, growth was poor. When git en a mixture of Chlorella sp. and P . tricornutum, their growth also varied on different occasiona. When fed Chhococcum sp. the larvae usu:dly grew well. Best growth waa achieved when they were given 1. galbana or N. Zutheri.
Very early stages of straight-hinge larvae of T . semidecussata are remarkably similar to those of M. mercenaria. Some of the smallest straight-hinge larvae in our cultures were only about 9Op long, but such individuals were uncommon ; the majority measured approxi- mately 95 x 70p . The umbo begins to develop when the larvae are approximately 120p long and becomes fairly prominent by the time

112 VICTOR L. LOOYANOFF AND HARRY C. DAVIS
the larvae are about 140 p in length although, even then, the hinge line may appear straight when seen from certain angles. As the larvw grow, the umbo becomes more prominent and in older larvae it is somewhat more pronounced than in M. mercenaria (Fig. 37).
When fed Chlorella sp. larvae reached metamorphosis in about 2 weeks. However, by giving them better food, such as naked flagel- lates, and by increasing the temperature of the water, setting size could be reached several days earlier.
The smallest metamorphosed individuals with a large, functional foot and fully resorbed velum measured about 175 p in length, or about the same as that reported for M. mercenaria. The majority of the larvae metamorphosed when they were between 200 and 220 p in length, but some much larger larvae measuring about 235 p in length still possessed a functional velum and were able to swim. Yoshida (1960) gave the size of fully grown veligers of T . variegata as 215 x 195 p.
As in most of our larval cultures, the color of the larvae depended, to a large extent, upon the food given. Furthermore, differences in size among larvae of the same cultures were of about the same magni- tude as in cultures of other species. It has also been definitely established that larvae of T. semidecussata are susceptible to fungus diseases.
Because of our prolonged studies of T . semidecus~ata we have had an opportunity to observe their adaptability to changes in their surroundings. These clams were able to survive considerable variations in temperature and salinity, which occur over the tidal flats of Milford Harbor in front of our laboratory. During one winter, when the tem- perature of the water remained below zero for several weeks and the shores were covered with a thick layer of ice, only about 3% of the clams planted on the flat, near the mean low water level, died. The clams also survived periods of freshets, such as were caused by hurricane " Donna '' in 1960, when salinity of the water in Milford Harbor was reduced almost to zero and remained low for several days.
The clams spawned and their larvae set normally in our experi- mental ponds. The small clams grew to a size of about 1 cm by the middle of September.
N. Pitar (= Callocardia) MT~UUTUZ Godd This small mollusk, which somewhat resembles H. mercenaria, is
found along the northern section of our Atlantic shore, roughly from Prince Edward Island to Cape Hatteras. In some sections of this range,
in Narragansett Bay, it is quite numerous. Recent literature contains few references to propagation or larval

REAXNU OP BIVALVY MOLLUSKS 113
stages of P . morrhuana. Sullivan (1948) is virtually the only author to devote some attention to veligers of this bivalve. Costello et al. (1967), calling the same form Callocardia convexa, gave a good descrip- tion of procuring gametes of these mollusks.
Our own experience with P. morrhuanc, has been extensive but not always satisfactory because this clam is one of the few bivalves that we could not induct? to spawn. All the methods known to us, including increasing of the water temperature, adtlition of ripe sex products, changes in p H or salinity, and use of mechanicrtl and electrical stimu- lation failed to induce spawning. Moreover, wc have never observed natural spawning among the hundreds of these clams that have been kept in our laboratory under different conditions, sometimes for periods of several months.
Animals brought into the laboratory from their natural beds in January already contained what appeared to be morphologically-ripe gametes. The color of the female gonad was usually creamy white and that of the male a light yellow. Even this early in the season active sperm could be taken from male gonads. The spermatozoa resembled those of M. mercenaria in shape. Eggs could be easily obtained by stripping, but they could not be fertilized so early in the season. This was true even when P . morrhuana was slowly conditioned in the laboratory by day-by-day increases in temperature. However, as the season progressed, the eggs became more responsive and the germinal vesicle in some of them dissolved soon after introduction of spermatozoa.
Our larval cultures were grown from eggs obtained by stripping. The size of the eggs taken from females with well-developed gonads ranged in diameter from 49 to 00 p, with t h c modal size-group betwoen 50 and 55p. These measurements, however, refer only to the egg proper; if the entire diameter of the ovum, including its thick, gela- tinous membrane were measured, it would vary between 92 and 128 p, with the modal class near 117 p.
On several occasions we tried to improve the condition of stripped eggs by adding 3 ml of 0.1 N ammonium hydroxide to every 100 ml of egg suspension. After 25 min of this treatment the eggs were screened to discard the fluid. The eggs were then rinsed with sea water, resus- pended, and sperniatozoa added in proper quantities. In some instances, after this treatmciit about 40% of all eggs underwent cell division, but llsually not more than 5% developed to normal straight-hinge larvae.
Early development of eggs and larvtw of P . mrhuana is virtually identical to that of other mollusks of the same group. Progress of development, as usual, depends upon the temperature a t which the

114 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
organisms are grown. I n some cases straight-hinge larvae were formed after 24 hr at a temperature of about 21°C.
The size of the smallest normal straight-hinge larvae waa approxi- mately 78 x 64 p. Several smaller larvae meafluring only 75 p b length were also observed, but appeared to be abnormal. Some larvae began developing an umbo when 95 p long, and the majority possessed a well-developed umbo by the time they reached 8 length of 125 p. No " eye " spots were observed in either larvae or recently set juveniles. Larvae of P. morrhwznu closely resembled those of M. mercenaria, but were considerably less active (Fig. 38).
Metamorphosis occurred when the larvae were still comparatively small. The smallest young clam possessing a functional foot and
pc
Fio. 38. Larvae of Pitar morrhuunn. Largest larvae shown are approximately 150 p long. Small, abnormal larvae can be seen in the group.
attached by a byssus thread measured only 160 p. However, a majority of the larvae did not begin to metamorphofle until reaching a length of approximately 180 p. The largest free-swimming larvae still pos- sessing a functional velum measured 192 x 179 p.
According to Sullivan (1948) the smallest larvae of P, morrhuunu were 120 x 95p. This size is considerably larger than that of the smallest larvae in our cultures. The maximum size offered by Sullivan, 220 x 210pL, was much larger than ever recorded in our cultures. It seems improbable, therefore, that the larvae considered by Sullivan to be those of P. morrhuanu were actually of this species.
Growth of larvae in different cultures varied considerably, depend- ing upon temperature and quality of food given. The majority of the cultures fed a mixture of plankton, composed mostly of several species

of ChloreZEn, and kept at room temperature began to set about 14 days after fertilization. Probably if given twttc:r foods, such as I. galbanu and M. lutheri, which at the time of rearing P . morrhuanu were not available at our laboratory, a better rate of survival and growth could have been achieved.
Many cultures suffered mass mortality, which usually occurred during the late stages of larval growth. These mortalities were probably caused by the same fungi that were later found responsible for heavy mortalities of larvae in ciiltures of M. mercenaria and several other species.
0. Petricola pholadiformis Lamarck Even though the range of this clam extends from Prince Edward
Island to the West Indies and Texas and their larvae should be common in plankton samples during the reproductive period, Sullivan (1948) seems to be the only author who has desvribed them. According to this author the first larvae of P. p?wladiformis normally appear in the waters of Malpequo Bay, Prince Edward Island, about the 2nd week of July and, after reaching peak numbers in early August, disappear from plankton in early September.
In our laboratory, groups of P . pholcrdiformia were conditioned for spawning from the middle of December until summer. I n the early winter these efforts were not as successful as later in the season and conditioning required a much longer time. For example, clams placed in conditioning trays a t about 20°C in December required 4 to 6 weeks before the majority responded to stimulation and spawned. In the middle of April, however, only a week of vonditioning a t about 20°C was sufficient, to bring these clams to spawiling condition.
Conditioned clams were induced to spawn by our usual method, consisting of subjecting them to a quick increase in the water tempera- ture from about 20" to 27°C and by the addition of a sperm suspension. Later in the season spawning, especially of males. could be induced on some occasions a t a temperature as low as 15°C. Raising the water temperature to about 32°C always affected dams unfavorably.
Early in the season aevcral hours at the increased temperature were often required before the first individiials, usually males, would resp'md, whereas late in March and in A i d spawning was usually provoked within tho first hour. Even young clams, leas than 1 yr old, and individuals with partially damaged shellr responded to stimulation and discharged large quantities of spawn. Often the same individual would spawn for a while, cease spawning and then resume it. Many individually-marked clams spawned several times during the Reason.
9

110 VICTOR L. L O O S ~ O F F AND H m Y C. DAVIS
Attempts at artificial fertilization of eggs .of P . p h o l d i f m b were partially successful, indicating that this can be done. As usual in the& cases, the majority of stripped eggs were somewhat injured.
Ripe eggs meclsured immediately after discharge varied in diameter from 61 to 58 p, but the modal size was quite clearly defined at 52 p . No speciai sttdies were made as to the number of eggs produced by a single female during the season. However, ip one case a large female discharged approximately 1 125 000 eggs in a; single spawning.
According to Sullivan (1948) straight-hinge larvae of P. p h o b diformis are quite large, measuring 115 x 100 p. We found, however, that young straight-hinge larvae are considerably smaller than reported by Sullivan. With the exception of one, apparently an abnormal lama
FIG. 39. Larvae of Petrieola pholadifomia. Largest larva in this photomicrograph is
which memured approximately 71 x 67 p, the majority of normally formed young larvae were 79 x 65 p in size. The largest free-swimming larva wm about 193 x 174p, although many underwent metamor- phosis before reaching this size, being approximately 186 x 164p (Fig. 39). These dimensions are somewhat larger than indicated bg Sullivan (1948).
Early straight-hinge larvae are very light in color and even their digestive organs me comparatively pale. In general, they are more transparent and thinner than larvae of most of the species described in this paper. Some of them remain transparent up to the time of metamorphosis. However, when given purple sulphur bacteria belong- ing to the genus Chromatium, larvae quickly acquire a pinkish color. If they are given such foods as Chlore.eUa, their digestive glands rapidly become a greenish color.
about 164 X 143p.

REARING OF BIVALVE MOLLUSKS 117
Since studies on larvae of this species were carried on when good food organisms, such as the naked flagellates, were not available, the larvae were fed mixed cultures of microorganisms in which ChloreUa was usually the predominating form. Larvae grew well on this alga, although it is quite certain that they would have grown faster if given better foods.
Larvae of P. pholadifomnis were grown at temperatures ranging from 18" to 30°C. The best cultures were observed at room temperatures ranging from 20" to 25°C. A t somewhat lower temperatures, between 17' and 18"C, eggs developed and larvae grew, but growth was slow. At 30°C egg development was rapid, but larvae were feeble and the majority died within 2 to 4 days. Perhaps mortality was not due exclusively to temperature alone, but to the heavy bacterial flora that quickly established itself under these conditions.
The shortest period from fertilization of the egg to metamorphosis at room temperature was 13 days. In several instances, however, the larvae of these cultures did not begin to set until the 20th or 21st day. The differences were probably due to variations in quality and quantity of food.
Two characteristics were noticeable in young clams soon after metamorphosis. First, the color of their shells becomes muoh denser and, secondly, the edges of the shells opposite the umbo acquire a serrated, roughened appearance.
P. Ensis directus (Conrad) The general characteristics of larvae of E. directus were given
recently by Sullivan (1948). Costello et al. ( I 957) gave a brief descrip- tion of early HtageH of (:gg dsvnlopmctnt, whilc l,ohoiir ( 1 IW) tirul nLiidiod larvae of fiaxin diquu. I n gt:nc:ral, howt:vc:r, lrirvao of 1C. djrectw have bcon l c s ~ iritcnsively studied than thoso of other common bivalves of our coast.
In our laboratory, observations on E'. directua and its larvae were conducted for three winters. Each year these clams were brought from the field and placed in conditioning trays in late December. It was found, however, that when these clams arc removed from the oil they soon begin to die because the powerful hinge pulls the clam shells apart. This can be prevented by placing tight rubber bandu mound each clam to counteract the action of the hinge and kccp itH shells closed. Using this simple method thew clam8 havo hcen kept alive without any soil for periods aH long
Attempts to induce spawning of E. directm were madc during tho middle of January after 2 or 3 wecks of conditioning. Although at
A month.
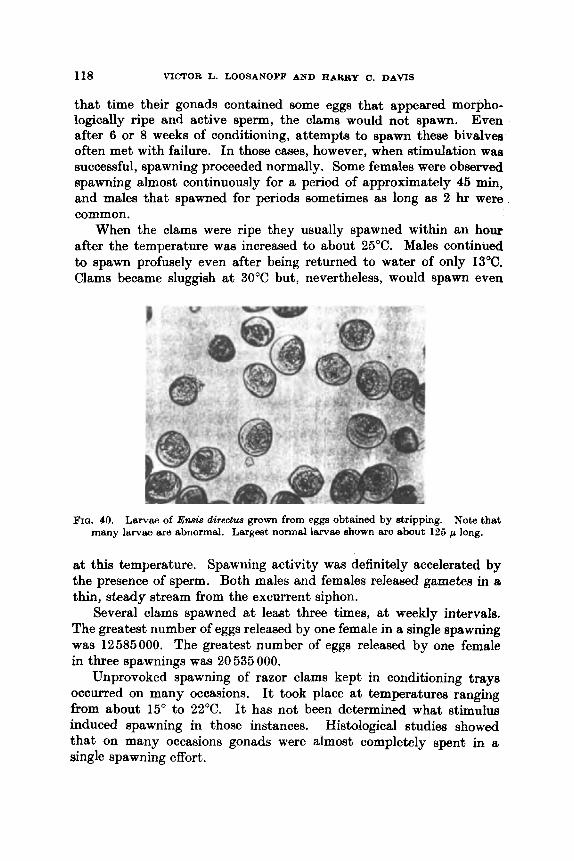
118 VICTOR L. LOOSAXOFF AND H A R R Y C. DAYIS
that time their gonads contained some eggs that appeared morpho- logically ripe and active sperm, the clams would not spawn. Even after 6 or 8 weeks of conditioning, attempts to spawn these bivalves often met with failure. In those cases, however, when stimulation was successful, spawning proceeded normally. Some females were observed spawning almost continuously for a period of approximately 45 min, and males that spawned for periods sometimes as long a8 2 hr were common.
When the clams were ripe they usually spawned within an hour after the temperature was increased to about 25°C. Males continued to spawn profusely even after being returned to water of only 13°C. Clams became sluggish a t 30°C but, nevertheless, would spawn even
FIG 40. Larvae of E d directw, grown from eggs obtained by stripping. Note that many larvae are abnormal. Largest normal larvae shown are about 125 p long.
at this temperature. Spawning activity waa definitely accelerated by the presence of sperm. Both males and females released gametes in thin, steady stream from the excurrent siphon.
Several clams spawned at leaat three times, a t weekly intervals, The greatest number of eggs released by one female in a single spawning was 12585000. The greatest number of eggs released by one female in three spawnings was 20 535 000.
Unprovoked spawning of razor clams kept in conditioning trays occurred on many occasions. It took place at temperatures ranging from about 15" to 22°C. It has not been determined what stimulus induced spawning in those instances. Histological studies showed that on many occasions gonads were almost completely spent in a single spawning effort.

ZLEARINU OF BIVALVE MOLLU8KY 119
Success of fertilizing eggu stripped from conditioned females de- pended upon the degree of their ripeness. Early in the season, when the largest of the eggs were only 6611. nolie were fertilizable. Later, when the clams were riper, a large perccntage of stripped eggs under- went cleavage but, as is often the cast in using eggs obtained in this manner, most of the larvae were abnormal (Fig. 40). Costello et al. (1957) also reported that eggs of E . directus can be fertilized artificially and they remain fertilizable for about 5 hr.
The egg of a razor clam is surrounded by a narrow perivitelline space enclosed in n thin vitelline menibrane. Measurements of eggs discharged in a normal spawning showod that they varied in diameter from about 64 to 73 p, the majority being 71 p, thus indicating that this is the modal size of iiormal eggs. Lebour (1!)38) gave the diameter of eggs of Ensis Riliqua as about 7 0 p, which closdy resembles measure- ments of eggs of our species. Jcirgensen (1946) gnve references to other workers who studied eggs and larvae of En.& and related forms.
Eggs and larvae were usually cultured at about 22°C. The smallest straight-hinge larvae were 80 p in length, but only two individuds this small were observed. Even larvae as small as 85 p in length were scarce, while those measuring approximately 92 x 78 p were common. This size is considerably smaller than the 140 x 120 p indicated by Sullivan (1948) as the smallest larvae of this species.
The larvae are light in color and more transparent than those of M . mercenaria. The umbo appears in litrvne only 116 p in length and becomes well developed in larvae 135 p long. Larvae of this size have definitely passed the straight-hinge stage. The umbo is so pronounced in larger larvae that it is impossible for them to lie flat. As a result, considerable variation in the length-width ratio is obtained when large larvae are measured under a microscope.
Setting of E . directus began soon after some larvae reached a length of about 210 p. A t 24°C this often occiirred only about 10 days after fertilization. However, some larvae rtbmained swimming until they reached a much larger size before undergoing metamorphosis. For example, in one culture fed Chlorella sp. larvae measuring 250 to 260 p in length were still swimming 27 days after fertilization and possessed both a powerful velum and a well-developed Soot. The largest swimming larva measured 270p, while in the same culture many fully meta- morphosing individuals showing a well-developed foot, gills and siphons were only 220 p long.
The appearance of the " eye " was irregular and i t was not located where this structure is usually found in most bivalve larvae. The smallest individual possessing thig struct ure was only 126 p long, but

120 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
many other larvae, even those nearing metamorphosis, did not display it. Sullivan (1948) did not mention the presence of an " eye '' in E. directus larvae and Werner (1939), working with a species that he called Cultellus pellucidus, a form closely related to En&, stated that its larvae do not possess " eyes ". This observation is quite important because, according to Rees (1960), the description by Werner of C. pellucidas was really that of Ensis. Our observation that the "eye "-like structure is sometimes present in larvae of E. directw and Werner's, that it is always absent in the species that he studied, may indicate that Reed assumption is not correct and that the C. pellucidus of Werner was not an Ensis.
One of the characteristics of larvae of E. directus which may be of diagnostic value is the presence of a clear area around the edge of the shell. This area, which does not occur in larvae of other speciea tihat we have studied, is well defined and clearly seen. It appeared in larvae of about 120 p long and persisted until they reached a length of 180 to 190 p . Later this area became less definite and more or less merged with other parts of the shell. Near setting, however, a purple wst became apparent along the edge of the larval shell.
In rearing of E. directus the presence of many abnormal larvae was quite common, not only in cultures that were started from stripped eggs, but also in those that were grown from normally discharged o m .
In several cultures, usually those where larvae were approwhing setting stage, mortalities on an epizootic scale were recorded. In several instances practically all of the larvae died within a short period, usually less than one day. Since this happened before we learned the devastating effect of larval diseases caused by fungi or bacteria, we did not establish the cause of death of these larvae but strongly suspect that these mortalities were principally the result of fungus infestations.
Q. dlactra (= Spisula) 80&h3ima Dillwgn The clams used in these studies were taken from the sandy beclchee
of the south shore of Long Island, New York. They were shipped to Milford and kept for many months, either in the laboratory or in the harbor, without any appreciable mortality, although the salinity at Milford is about 27 ppt, or approximately 8 ppt lower than in the areas where the clams were dredged. The clams survived equally well whether they were allowed to dig into sand or were kept in wire baskets containing no soil. To prevent mortality, however, it was necessary to expel bubbles of air which were trapped inside the clams while they were out of water. Clams placed on their sides cannot expel these bubbles and, eventually, die.

REARING OF BIVALVE MOLLUSKS 121
Our experience in growing larvae of &I. solidissirnu was confined to the period from the middle of January to the end of June. During that time morphologically-ripe eggs and spermatozoa were always present in the gonadal folficles. Possibly, equally well-developed sex cells could have been found in the gonads several weeks earlier than January because, apparently, the most important period of gametogenesis in M . solidissima begins immediately after the end of spawning, as in M . mercenuria, in which gonads of both sexes present a ripe appearance in December (Loosanoff, 1937b). Allen (1953) obtained
Spermatozoa, taken from clams brought from their natural beds early in January, when placed in a drop of water at room temperature quickly began to move. However, eggs taken from clams of the same groups usually could not be fertilized, probably because they were not physiologically ripe. Nevertheless, placing female clams in water of about 15°C for only a few days was sufficient to ripen them. Later in the season, when the temperature of the water in Milford Harbor increased to 12"C, no conditioning was needed.
Fertilizable eggs could be obtained either by stripping or by inducing spawning. Sometimes, stripped eggs obtained from a ripe female were as viable as those discharged during a normal spawning. More often, however, a culture started with stripped eggs contained many abnormal individuals. Costello et al. (1957) suggested a different method of obtaining gametes, which consisted of straining the eggs through a cheesecloth.
Spawning was induced by the usual method of thermal and chemical stimulation. On several occasionH t h s r m d stimulation alone w w sufficient. Normally, addition of a wspwwiorr of H(:X C ~ I H ~ i i ~ rtquin:d and, even then, many conditioned clams did not Hpawn.
Spawning was induced at temperatures ranging from 18" to 25°C. Temperatures of 30°C and over unfavorably affected the clams. They gaped and abnormally extended their feet which, like the rest of their bodies, became limp and soft. If, after exposure to mch high temperatures the clams were quickly returned to cooler watm of about 25"C, they often began to spawn, hut the egg8 ohtaincA in uuch a manner did not develop paRt the formatior) of polfir b d i w Appar- ently, exposure of clam^ to temperaturr!x ovcr W C caunc:rl Hc,r' ' I O U R
injuries to their eggs. Oar ohflorvatiorci in t h i H rc!~pcct agrco with those of Imai ot nl. ( I ! K 5 ) who, while working on II closely-relafcd species, Muclru Yachulinensis, found that at a temperature of about 32°C none of the eggs underwent normal cleavage.
Observations were made on the growth of larvae of M . solidissima
eggs of M. solidissima from early spring until late autumn.

122 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
at two different temperatures. Eggs obtained from several females and fertilized with spermatozoa of three ripe males were mixed together and then divided into two groups, one to be grown at a temperature of about 22°C and the other at 14OC. The concentration of eggs per rnl of water was the same in both cultures. During the experiment the larvae were fed equal quantities of phytoplankton composed largely of ChEorella sp.
Eggs placed in watkr at 14°C were in the trochophore stage at the end of 40 hr. After 72 hr the culture contained normal and active straight-hinge larvae averaging 85 p in length, which were quite uniform in size and appearance. At 22°C this stage was reached after only 28 hr. Young straight-hinge larvae of M . solidissirnu were usually
FIQ. 41. Larvae of Mactra 80l idb8hZ. Largest larva, 219 p long and 193 p wide, is Note differences in sizee of larvae that Originetea approaching metamorphosis.
from the same group of eggs and were reared under identical conditions.
almost transparent >and had a granular appearance with the internal organs at this time not too well defined. Later, as in the caae of most larvae, the color began to deepen.
On the 19th day, when larvae in the warmer culture began to metamorphose, the length of the modal class of larvae grown at 14OC waa only 117 p, and the larger individuals in the cultures were ody 153p long. The first metamorphosing clams were observed in thie culture after 35 days. When the experiment waa discontinued on the 60th day the remaining larvae showed a wide range in size, the small& being only 109 p long (Fig. 41).
A partial description of development and dimensions of larvae of M. solidissirnu was given earlier in this article. Here we may add

REARING O F BIVALVE MOLLUSKS 123
that the most recent description of what were assumed to be larvae of M d r a (= Spisulu) solidissinla was reported by Sullivan (1948). She gave the minimum size of straight-hinge larvae as about 95 x 80 p, which is approximately 15 p longer than usually found in our cultures, while the maximum size of 270 x 2 4 5 p closely agreed with our measurements. In both instances the length-width relationship given by Sullivan falls near the median line of length-width relationships which we found for larvae of M. solidissima.
In describing larvae of a related form, Spisula subtruncata, Jlargen- sen (1946) reported that veligers are about 400 p at the time of metamorphosis. Kandler (1926) stated, however, that the length of this species a t metamorphosis is only 3lOp. Considering that lmai p t d. (1953) found the maximum size of' larvae of M a c h ( Spisuln) sachalinerisis to be about 270 p, virtually the same as we found for M . solidissima, and because Imai's conclusions and ours are based on measurements of larvae of a known origin, we think that Kandler's measurements are more realistic than Jsrgensen's and that the latter was probably describing larvae other than those belonging to the genus Hactra (= Spisula). The same consideration leads us to believe that the descriptions and measurements offered by Rees ( 1950) of advanced stages of larvae presumably of the superfamily Mactracea, including Spisula solida (360 p) and Spisula plliptica (355 p), are really those of some other species.
R. Mya arenaria Linn6 Our efforts to induce spawning of M. arenaria were confined largely
to the period extending from March until the middle of July. Several groups of these clams were also conditioned and Hpawnwl i n winter. Moreover, to induce spawning of' evcn well-coriditionr:~l arid rtpjiitrwi t l y ripe M. arenaria is difficult, nevr:rthc,l A f!:HH.
In developing a method to indur:c: fipawrlirig of t l i c : ~ : cli irrifi wv f.rtc:tj
many approachw, iiicliitlirig niichlwr i w l grii~liid d i r m p f i 111 wrr1,t.r tc*m p r R 1 , i i rv , (4 i i ~ r I ~ P H i r i Ii f I , ~ i i l I i I i 1,y , tiy(lroHtiLtic prt:wu rf., light inkiwities and the addition of sex protlucis. Usually, none of theRe worked. The only method that proved to he successful with some regularity consisted of subjecting ripe clams to water of relatively high temperature, of about 26" to 28"C, for long period8 often extending from 6 to 8 hr and adding, during this time, a suspension of sex prod- ucts. Many clams spawned profusely when this method was employed and discharged a large number of cgp, I)ut a high perccmtage ( J f these eggs usually dt~velopeti into abnormal larva(..
Belding (1931) reported the diametcr of the average egg of the

124 VICTOR L. LOOSANOFF AND HARRY 0. DAVIS
soft shell clam as 62.5 p, while Battle (1932) gave the egg size as varying from 70 to 8 0 p . Our measurements of hundreds of eggs discharged by different females and on different occasions showed that the majority were between 68 and 73 p in diameter, with a modal size of 70-5 p.
Beldirig (193 1) expressed the opinion that artificial cultivation of M . arenuria is virtually impossible because the eggs either fail to develop normally or else never pass the young veliger stage. Never- theless, Belding was able to show that, unlike other pelecypods, eggs stripped from M . arenaria can be artificially fertilized.
The smallest normal straight-hinge larvae recorded in our cultures measured only about 86 x 71 p. These were, however, extremely uncommon and normal, fully-formed straight-hinge larvae were usually about 93 x 77 p. As in most pelecypod larvae, they were light in color at this stagc and their internal organs were not well defined. They remained quite light, almost transparent, until a length of about 110 p was reached. As the larvae grew, they became darker. Never- theless, as mentioned on several occasions, these larvae do not possess characteristic colors that would help to distinguish them from members of other genera or species of bivalves. In our experiments, where larvae of this species were fed different foods, their color ranged from a dark reddish-brown to dark green. We cannot, therefore, agree with Sullivan (1948) that brown pigmentation in large larvae of M. arenaria is diagnoRtic of that species.
In older larvae measuring about 175p and longer we noticed the presence, in the margins of the mantle, of irregular opaque spots varying in size from 5 to 15p. These granules occurred with such regularity that we are inclined to consider them a8 aharackristic of thc speciali, a t kaHt during Irttf. I&rvti t Ht&gc:H. .jwrgf.rinf:n ( I fr/tfj) flot,ic4i a somewhat different pigmcntatiori of the soft parts of larvae of M . nrertarici mtmuring ahout 200 p i d larger. He also suggested that thiR may 1)c a reliablc specific character.
'Thc size of larvae of M . arennria at setting is extremely varied. Metamorphosis may occur a t any lerigth from 170 to 228 p. The latter is the size of the largest free-swimming larva ever recorded in our cultures. The majority metamorphosed at a length between 200 and
The smallest larva in which the foot was present waH about 166 p long but many of the larvae had a well-developed foot by the time they reached 175p in length. The presence of a large foot doeR not neces- sarily indicate that the velum ha8 already hccome non-fiinctional. Larvae as long as 210 p have hcen w:m at timcH nwimmirlg rttmit cuing both the velum, which still nppc:ared to ht: of ncJrrrirrI ~ i m , arid t l h
210 p.

REARING OF BIVALVE MOLLUSKS 126
the large foot. However, the velum normally begins to disappear soon after a length of 172 to 175 p is reached and sometimes even earlier. In most individuals 200p in length the velum is already resorbed. Some larvae measuring only 175 p in length, and having no velum, were seen actually crawling, using their feet.
The balancing organ, the otocyst, can be clearly seen at the base of the foot of larvae measuring about 175p in length. The byssua gland also can be seen in larvae less than 200p in length, and the gills may be clearly discerned in some individuals of about the same size. The byssus thread is strong, and our attempts to break it by directing a strong jet of water from a pipette caused the larvae to sway from side to side, but did not break the thread.
FIQ. 42. Young larvas of Myo arena&. Largest larva shown ia about 140 p long.
No systematic studies on the rate of growth of larvae of M. arenuria at different temperatures, such as those conducted with larvae of Y. mercenaria, were made. Our attempts to grow larvae at low tem- peratures ranging from 12" to 15°C were usually unsuccessful because of slow growth. For example, at a temperature of about 14°C larvae, even after 15 days, were only about 110 to 115 p long. Larvae grown at low temperatures, probably because of the slow rate of growth, were usually of extremely uniform size.
Most of our cultures were grown at room temperatures which ranged from about 19" to 24°C. Under those conditions the rate of growth was quite rapid, although it varied, of course, from culture to culture, depending upon the temperature, concentration of larvae, and quality and quantity of food given. A t about 23°C the average length of larvae 2 days after fertilization was approximately 1 0 9 . 6 ~

126 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
and the maximum 117 p. After 5 days larvae averaged 120 p in length and the largest individuals were 140p long (Fig. 42). After 10 daye some of the largest larvae approached a length of 180p at which setting is possible. By the 15th day many individuals had already set, and the average size of the larvae in the cultures had i n c r e w to about 175 p. A few large larvae, measuring about 225 p in length but still free swimming, were also found in our samples. In some cultures setting began when the larvae were about 10 days old and continued until the end of the 35th day.
As has already been mentioned, the smallest straight-hinge larvae of M . arenaria in our cultures measured about 86 x 71 p. The largest free-swimming individuals were 228 x 207 p, although most of them metamorphosed before that size was reached. Our measurements, therefore, differ from those given by Stafford (1912), who stated that the smallest straight-hinge larva of M . arenaria that he found wzu only 75.9 x 62p, while the largest measured 414 x 3 4 5 ~ . Of the latter he said, “The largest measurement I have is 64 x 53, and I have seen them attached by a byssus-thread, their siphons protruded, and the big hinge-tooth on the left valve.” Since, according to Stafford, each unit of his measurements was equal to 6*9p, the dimensions of his larvae were as given above. It is quite possible, if his measurements were correct, that Stafford was working with larvae of a species other than M . arenaria and, possibly, his large individuals were already juvenile mollusks and not free-swimming larvae. Another possibility is that Stafford’s microscope was not correctly calibrated.
The dimensions of larvae in our cultures were not too different from Sullivan’s (1948). Nevertheless, her smallest size of 105 x 90 p is considerably larger than we found and her largest, 250 x 230p, also somewhat exceeds ours. Yoshida’s (1938) observation that larvae of M . arenaria in Japanese waters metamorphose upon reaching a size ranging from 240 to 300 p also disagrees with ours because, while the length of 240 p does not differ radically from the measurements of ow largest larvae, the maximum size of 3 0 0 p given by Yoshida exceeds ours by about 70 p.
The length of early straight-hinge larvae of M . arenaria given by Jsrgensen (1 946) is similar to ours. This is to be expected because his early larvae were laboratory-reared, as were ours, and, therefore, there is no doubt that we and Jsrgensen were working with the same species. However, we disagree with Jarrgensen that larvae of M . arenaria may reach 300p in length before metamorphosis. His conclusion regarding the setting size is based not on laboratory-reared M . arenaria, but on specimens collected in the field and only assumed to be this

REARING OF BIVALVE MOLLUSKS 127
species. The fact that Jsrgensen stated on several occasions that metamorphosis of M. arenuria may occur upon attainment of a much smaller size than 300p strongly suppo~%s our opinion. For example, Jsrgensen mentioned that in Ringkbing Fjord the size of larvae at metamorphosis in shallow water varies between 200 and 225p, thus being within the size range observed in oiir cultures.
S. Teredo nuvalis Linn6 Larvae of T . navalis have been described by many authors, including
Jsrgensen (1946), Sullivan (1948) and, ako, Imai, Hatanaka and Sato (1950), who gave a good description of the method of rearing them.
In our laboratory adult T . navali9 wcre conditioned to spawn as early as the fist part of December. This was done by placing pieces of wood, containing wood-borers, in sea water maintained at a tempera- ture between 16" and 20°C. Spawning occurred at temperatures of 14°C and higher, and larvae were released at temperatures ranging from about 16" to 20°C. Grave (1928) reported that spawning of T. nuvalis began when the water temperature reached 11" to 12°C. Imai et al. (1950b), however, found that spawning begins when the water temperature reaches 18°C. Sullivan's (1948) data closely agree with ours, that spawning and swarming niay occur a t approximately 15°C.
Although T . navalis is naturally larviparous, both recently- fertilized eggs and immature larvae removed from the gill chambers of the parents developed normally past metamorphosis. The diameter of unfertilized eggs varied between 50 and 60 p, agreeing with measure- ments given by Jsrgensen (1946) and Gofitello et al. (1967). In the diasected adults, however, most of the egg(i found were either already fertilized or immature, thus presenting difiiculty in obtaining reliable egg measurements.
The smallest larvae released in natur a1 H warm ings at our laboratory measured only 80 x 70p, whilo the largejt larvae found in the gill chamber of the mother.were 9Op long, or approximately lop longer than reported by Jrargensen (1946). Evidently, the length of larvae at the time of release may vary by at least 20 p. Imai et al. (1950b) indioated that the mean size at the time of release is 85 x 72p. Our observations that the average size of just-released Teredo laxvae is between 86 and 9 6 p &re in agreement with those of Sullivan (1948), Jmgensen (1946) and Imai et al. (195Oa). We cannot, however, accept the conclusion of Lane et al. (1954) who maintained that larvae are about 250 p in size when released from the gill chamber.
Eggs, in early stages of development, taken from the gill chamber

128 VICTOR L. LOOSANOFF A N D HARRY C. DAVIS
of a female Teredo were cultured to metamorphosis in 28 days at a temperature of about 20°C. Grave (1928) thought that the entire period of development of Teredo from the moment of fertilization to mela- morphosis takes about 5 weeks. Judging by data offered by Imai et al. (1950a), setting in their cultures occurred between the 24th and 34th days.
A brief description of early stages of development of eggs and larvae of T. navalis are given by Costello et ul. (1957) and of later stages, by Sigerfoos (1908). Imai et aE. (1950b) gave a good account for all stages. In our cultures the shells of straight-hinge larvae appeared heavy and thick. The larvae were also characterized by a dark band
FIG. 43. Late larval stages of Teredo nuvuli.9. Large&. larva in the group h about 185 p N o h dark hand tworind edge of nhtll charachristic of hrvw long and 300 p wide.
of this species.
around the edge of the shell from one end of the hinge to the other (Fig. 43). A light band was quite conspicuous inside of this dark band. Although these bands are probably optical illusions red t ing from curvature of the shell seen under the microscope, they are well pro- nounced. The bands are quite narrow and lees conspicuous in l&rvaa smaller than 9Op in length but, nevertheless, they are preAent even in these small individuals. As larvae approach Betting H i m , the ban& become less sharply delineated, although they remain quite, prominont.
The color of larvae hogiriH to dnrkan Hoon aftm ttwy rcriwh 100 p in length. Tmai at d. ( I MOu) t t l ~ ~ aamn to thtt namo aonalirniort. flowcwrr, whilc I rnai r.c.l,ortsc*tl thnt. t i c : i t h c v tho foot,, oto(:.yHt or gill filamerlt eppnw bc*f’orct larvno rcmh thv H i m of‘ ‘LOO x 216 p, wo observed their appearance in larvac at least 16 p smaller.

REARING OF BIVALVE MOLLUSKS 129
Imai et al. (1950b) gave a table showing growth of T. navcdis Iarvcle from day to day, indicating length-width relationships during different stages of growth. These data closely resemble ours. Other observations of these authors on appearance and behavior of larvaa are also in close tlgreement with ours.
Larvae of T . navalis are extremely active and usually swim vigor- ously and virtually continuously. This is particularly true of younger stages. We noticed that the larvae have some substance on the outside of their shells by means of which they adhere readily to glassware and, as a result, i t is extremely difficult to rinse them from beakers, pipettes, slides, etc.
Larvae began to metamorphose soon after a length of 200p was reaohed. However, several fully-metamorphosed individuals measuring only 190 p in length and 206 p in width were seen. The largest swim- ming larvae were approximately 200 x 231 p. Our maximum size of free-swimming larvae of the wood-borer is, therefore, somewhat smaller than the 220 x 250 p reported by Sullivan (1948), but closely approaches that given by Imai et al. (1950).
Larvae of advanced stages do not develop an ‘* eye ” that is characteristic of larvae of corresponding stages of other species, such as C. virginica. The foot of recently set borers is extrcmely slender and worm-like. The set attach themselves to the substratum by means of a byssus.
The time required for larvae in our cultures to reach metamorphosis varied. Early in our work, before good food organisms became avail- able, the first metamorphosing larvae were observed 20 days after swarming, when grown at room temperature. If better growing con- ditions were provided, the free-swimming pcriod could undouhtodly be shortened. Nevertheless, we strongly disagree with the conclusions of Lane et al. (1954) that the normal free-awimming period of Teredo larvae does not exceed 4 days.
Such infections were observed on numerous occasions and were probably responsible for the complete mortalities of Teredo larvae in some of our cultures.
Teredo larvae are quite susceptible to fungus diseases.
VI. ACKNOWLE DQME NTS
Our studies, which provided most of the material for thig articln, have been continuous since 1944 and, nrrturally, during t h i H timct many people have participatod eithcr directly or indircctly. Wc want to express our appreciation to all biologists of our laboratory, especially to Mr. Paul E. Chanley, who grew larvae of the several species of

130 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
bivalves discussed in this article ; to Mr. Herbert Hidu, who contributed a pea t deal of information on utilization of dried algae a larval food ; to Miss Phyllis B. Smith, who assisted us in a number of the earlier experiments ; and to our microbiologists, Dr. Ravenna Ukeles and Dr. Robert R. Guillard, who provided the phytoplankton for our feeding exporirnents. Wo aru grctteful to Dr. F. 8. Rusmll and Dr. Mary Parke of Plymouth Laboratory, England, for sending us inocula of algae, several of which were extensively used in our studies.
We also want to acknowledge the technical help of our Laboratory Mechanic, Mr. Joseph F. Lucash, whose ingenuity in designing and building various apparatus for our experiments made possible the successful completion of many of our physiological studies. Miss Rita S. Riccio and Mrs. Florence S. Munz were extremely helpful in preparing and editing this manuscript, while to Messrs. Manton Botsford and Charles Nomejko we are indebted for preparation of the photo- graphs and other illustrations.
VII. REFERENCES Allen, R. D. (1953). Fertilization and artificial activation in the egg of the
surf-clam, Spisukz solidhsima. Biol. Bull., Woo& Hole, 105, 213-239. Battle, H. (1932). Rhythmic sexual maturity and spawning of certain bivalve
molluscs. Contr. C a d . Biol. 7, 256-276. Belding, D. L. (1910). A report upon the scaIlop fishery of Masaachusotte,
inc:liitling t h o habits, life history of Pecten i radiam, its rate of growth and othor factors of oconomic value. Kpec. Rep., Comm. E'idh. unrl Qame, Mwn. 1-160.
Belding, I). I,. (1912). A report upon the quahog and oyster %herim of Maaaa- clirisui ts, pp. 1-1 34, Boston.
[ h l d i n ~ , I ) . 1,. ( l0Sl f. &p. hlil.88. ( h n m . ( l h p . ) Conarrv. firrbr. Piah. 8or. No. 12, pp. 1-66.
Ihrisjnk, A. (1909). l'elecypodu tlu planlcton do la Mer Noire. BuU. eci. Fr. Be&.
Boury, M. (1928). lb ude sur la reproduction des huitres. Rev. Trav. OJ. P6che8
Brooks, W. K. (1880). The developmtmt of the American oyster. Stud. Biol.
Bruce, J. R., Knight, M., and Parkc, M. W. (1940). The rearing of oyster larvrte
Burkenroad, M. D. (1947). Egg number is a matter of interest in fiahel'y biology.
Cahn, A. R. (1950). Oyster culture in Japan. Fish. Leafl.. Waeh. 383. 1-80. Cahn, A. R. (1951). Clam culture in Jclpan. Fish. Leap., Waah. 399, 1-103. Cerriker, M. R. (1966). Biology and propagation of young hard clam, Mercerulria:
mrcenaria. Chenley, P. E. (1955). Possible c a u f l ~ of growth variritionn in clam hrvm,
Proc. nut. SheUfih Ann. (1964), 45, 84-94.
'J'hrt 1v~t'f~.fii114J~~~l c.l&rn flMfJttfy of Mnwrnr!hiiauth~.
42, 149-181.
Marit. 1, 87-99.
Lab. Johns Hopkim Univ. IV, 1-104.
on an algal diet. J. Mar. biol. As&. U.K. 24, 337-374.
Science, 106, 290.
J . Eliaha Mitchell nci. Soc. 72, 1, 67- 00.

REAEINGI OF BIVALVE MOLLUSKS 131
Chanley, P. E. (1961). Inheritance of shell markings and growth in the hard clam, Venus mercenaria.
Cheatnut, A. F., Fahy, W. E., and Porter, H. J. (1967). Growth of Venus mer- -ria, Venus campechiensia and their hybrids. Proc. &. SheUjeSh Ass. 47, 50-56.
Churchill, E. P., Jr. (1920). The oyster and the oyster industry of the Atlantic and Gulf coasts. Docum. U.S. Bur. Fish. no. 890; app. 8 to Rep. U.S. Comm. Fkh for 1919, pp. 1-61.
Coe, W. R. (1931). Sexual rhythm in the California oyster (Oatreu lurida). Science, 74, 247-249.
Cole, H. A. (1936). Experimenh in the breeding of oyatere (Ostreo eddb) in tanks, with special reference to the food of tho larva and spat. Fkh. Invest., Lond. Ser. 11. 15, 1-28.
Colo, H. A. (1938). The fate of the larval orgun8 irr t,ho rnotamorphosis of Ostreu edulia. J . Mar. bid. Ass. U.K. 22, 409-4HH.
Colo, kE. A. (1939). Further experimentR in broetling of oysters (Oslrea eddiS) in tanks.
Cole, K. A. (1941). The fecundity of Ostrea e d d k J. Mar. biol. Ass. U.K. 25, 243-260.
Coatello, D. P., Davidson, M. E., Eggers, A., Fox, M. H., and Henley, C. (1957). Methods for obtaining and handling marine eggs and embryos, pp. 1-247. Mar. Bwl. Lab., Woo& Hole, Mass.
Davis, H. C. (1949). On cultivation of larvae of Ostreu lurida. A&. Reo. 105, 111.
Davis, H. C. (1950). On interspecific hybridization in Ostreu. SciencC, 111, 622. Davis, H. C. (1953). On food and feeding of larvae of the American oyster,
C. virgkicu. Davis, H. C. (1958). Survival and growth of clam and oyster larvae at different
salinities. BWE. Bull., Wood8 Hole, 114, 296-307. Davis, H. C. (1960). Effects of turbidity-producing materkle in 888 water on
eggs and larvaa of the clam (Venus (Mercenurk) W Y ~ ? Z W & Z ) . BWl. Bd.,
Effect8 (If w m e pcHticich on c?ggn and Iurvw ( i f oyrrhrr thmn. IGh. I ~ I .
Ihvis, H. C., and AnRell, A. D. (1962). 8urvivriI and growth of Lrvru, of tha Europoan oyHLor, 0. edulir. at Inwored HalinllioH. fiiol. fiull., Woods Hole,
I)lbviH. I I . t!., t r r i t l (!triubloy, 1’. JZ. (196th). Spttwiiing and egg production of
DaviH, 11. C., ant1 Chanley, 1’. 15. (1956b). Effects of some dissolved substances 011 bivalvo larvao. Proc. nat. ShellfMh Ass. 46, 59-74.
Davis, H. C., and Uuillard, R. R. (1958). Relative value of ten genera of micro- organisms a3 foods for oyster and clam larvae. Fieh. B d . , U.S. no. 136, 58, 293-304.
Davis, H. C., and Loosanoff, V. L. (1953). Utilization of different food organisms by clam larvae. Anat. Rec. 117, 646.
H. C., and Ukeles, R. (1961). Mass culture of phytoplankton as f& h r metazoans. Science, 134, 662-564.
Proc. &. SheUjeSh Ass. 50, 163-168.
Fbh. Invest., Lond. Ser. 11, 16, 64.
Bwl. BuU., Woods Hole, 104, 334-350.
WOO& Hole, 118, 48-54. Davis, H. C . (1961).
(Craasmtrea virgin&) and clam ( Yenu8 m.?rmru~ff~). 23, 8-23.
122, 33 :ilL
OYHhPH Ibll(1 ClfWlH. I#Wl. hdc!., WOO& Hole, 110, 117-128.
10

132 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
Davis, H. C., Loosanoff, V. L., Weston, W. H., and Martin, C. (1954). A fungus disease in clam and oyster larvae. Science, 120, 36-38.
Drew, G. A. (1906). The habits, anatomy, and embryology of the giant scallop (Pecten tenuicostutua Mighels). Stud. Unav. Maine, no. 6, 3-71.
Field, G . A. (1922). Biology and economic value of the sea mussel Mytilue edulis. Bull. U.S. Bur. Fish. 38, 127-259.
Fullarton, J. H. (1890). On the development of the common scallop (Pecten opercularis). Rep. Fish. Bd. Scot., 1889 (1890). Part 111. Scient. Invest., 290-298.
Galtsoff, P. S. (1930). The fecundity of the oyster. Science, 72, 97-98. Galtsoff, P. S. (1932). Spawning reactions of three species of oysters. J . Wmh.
A d . IS&. 22, 65-69. Grave, B. H. (1928). Natural history of shipworm, Teredo Wwtli8, at Woods
Hole, Massachusetts. Biol. Bull., Woods H o b , 55, 260-282. Guillard, R. R. (1959). Further evidence of the destruction of bivalve larvae
by bacteria. Bwl. Bull., Woo& H o b , 117, 258-266. Gutsell, J. S. (1930). Natural history of the bay scallop. B d . U.S. Bur. Fish.
Haven, D., and Andrews, J. D. (1957). Survival and growth of Venua mercenaria, Venus campechiemis, and their hybrids in suspended trays and on natural bottoms. Proc. nut. Shellfish Ass. 47, 43-48.
Hopkins, A. E. (1937). Experimental observations on spawning, larval develop- ment and setting in the Olympia oyster, Ostrea lurida. Bull. U.S. Bur. Fish.
Hori, J. (1933). On the development of the Olympia oyster, Ostrea Zuridcl Car- Bull. Jap. SOC. aci.
Hori, J., and Kusakabe, D. (1926). Preliminary experiments on the artificial culture of oyster larvae. J . Imp. Fish. Imt . 22, 47-52.
Imai, T., and Hatanaka, M. (1949). On the artificial propagation of Japaneao oyster, Ostrea g i g m Thun., by non-colored naked ffagellatea. BuU. Imt. agrk. Res. Tohoku Univ. 1, 33-46.
Imai, T., and Sakai, S. (1961). Study of breeding of Japanese oyster, Craas- oatrea g i g m . Tohoku J . agric. Rea. 12, 125-163.
Imai, T., Hatanaka, M., and Sato, R. (1950a). Breeding of marine timber-borer. Teredo nuvalis L., in tanka and its use for anti-boring test. Tohoku J. agric. Res. 1, 199-208.
Imai, T., Hatanaka, M., Sato, R., Sakai, S., and Yuki, R. (1950b). Artificial breeding of oysters in tanks. Tohoku J . agrk. RM. 1, 69-86.
Imai, T., Hatanaka, M., Sato, R., and Sakai, S. (1953). Tank breeding of the Japanese surf clam, Mactra sachalinemis Schrenk. Sci. Rep. Ree. Inat. Tohoku Univ. 4, 121-131.
Imai, T., Sakai, S., Okada, H., and Yoshida, T. (1954). Breeding of the Olympia oyster in tanks and culture experiments in Japanese waters. Tohoku J . agric. Res. 5, 13-25.
Reproduction and larval development of Danish marine bottom invertebrates. 9. Lamellibranchia. Me&. Komm. Havun- dersog., Kbh Ser.(tl): Plankton, 4, 277-311.
Jsrgensen, C. B. (1949). The rate of feeding by M y t i h in different kinds of suspension. J . Mar, biol, Ass. U . K . 28, 333-344.
46, 569-632.
48, 439-503.
penter, transplanted from United States to Japan. Fish. 1, 269-276.
Jsrgensen, C. B. (1946).

REARING OF BIVALVE MOLLUSKS 133
J~rgensen. C. B. (1960). Efficiency of particle retention and rate of water tram- port in undisturbed lamellibranchs. Extr. du J. C m . int. Explor. Mer. 26, 94-116.
Kiindler, R. (1926). Muschellarven aus dem Helgolander Plankton. Bestim- mung ihrer Artzugehorigkeit durch Aufzucht. Wies. Meeresuntermch., N. F., Abt. Helgoliind, 16, 1-8, Kiel & Leipzig.
Kinoshita, T., and Hirano, Y. (1934). A study on tho optimum tcmpcrature for the devolopment of Mactra eachulinene~.
Korringa, P. (1941). Experiments and observations on swarming, pelagic life and setting in the European flat oyster, Ostreu edulie L. Arch. need 2002.
Quart. Rev. Biol.
Tho respiration of normal larvae of Teredo bartechi Clapp. Biol. BuU., Woo& Hole, 106, 323-327.
Lebour, M. V. (1938). Notes on the breeding of somc lamellibranchs from Ply- mouth and their larvae. J. Mar, biol. As8. U.K. 23, 119-146.
Lindsay, C. E., and Woelko, C. E. (1960). Production of clam and oyster seed. Fish., Fish Farm., Fish. Mgmt. Wwh. State Dept-. of Fish., Seattle, Waah..
Loosanoff, V. L. (1937a). Seasonal gonadal changes of adult clams, Venzra mewenaria (L.). Biol. Bull., Woods Hob, 72, 406-416.
Loosanoff, V. L. (1937b). Spawning of Venus mercenuria (L.). Ecology, 18, 606-616.
Loosanoff, V. L. (1942). Seasonal gonadal changes in the adult oysters, Oslreu v i rg inh, of Long Island Sound.
Loosenoff, V. L. (1945). Precocious gonad development in oystcrs induced in midwinter by high temperature. Science, 102, 124-125.
Loosanoff, V. L. (1949). Method for supplying a laboratory with warm sea water in winter. Science, 110, 192-193.
Venue. 4, 368-372.
5, 1-249. Korringa, P. (1952).
Lane, C. E., Tieniey, J. Q., and Honnacy, R. E. (1954).
Recent advances in oyster biology. 27, 266-308, 339-365.
3, Ch. 8, pp. 81-85.
Biol. Bull., Woods Hob, 82, 195-206.
LOORanOff, v. L. (1951). fhlturing phytoplariktiin 0 1 1 8 hrKo H C f d I * . /h‘dllIJ@/, 32, 748-750.
Loosannff, V. L. (19.52). / V W ,
TAoosannff, V. I,. (lean) / h l . /Mi . ,
~ ~ ~ ~ ~ I N I w ~ ~ , V. I,. ( l l v i 4 ) . Now I L ~ V I L ~ ~ C O H in t l i n HI irily ( I F I)ivalvo Iirrvtuj. Amcr. i % h t . 42, (107 1\24.
IlQoHILnoff, v. I,. (IfG5). Tho bhiropan oyHlor iri Ainorican watom. b’cience,
I*owimoff, V . L. (lD68e). Challepging problems in uhellfish biology. I’erspeo. in Mar. Biol. Part IV, (1966), 483-495.
Loosanoff, V. L. (1968b). Environment and larval behavior of several pelecy- pods. XVth Internatl. Congress of Zool., London, England, Sect. 111, Paper 10, 1-3.
bosanoff, V. L. (1959). The size and shape of motamorphosing larvae of Venw (Mercenasia) mereemria grown a t different temperatureu. Biol. Bull.. Woods Hole, 117. 308-318.
I<ihavior of oywtr:rM iri watr.i ( i f J O W NII~I~IIIII*N,
nal. Shelljiah Am. 43, 136- 151. Itq)rrduct ivo c*yr:lo in {‘yprituh itilfandica.
C~’fJ0d.q !lOb. 104, 148 166.
121, 119-121.

134 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
Loosanoff, V. L. (1961). Partial metamorphosis in Anomia eimp2ex. Science,
Loosanoff, V. L. (1962a). Effects of turbidity on some larval and adult bi- valves. Proc. Gulf & Carib. Fish. Inst., 14th Annual Session, pp. 80-96.
Loosanoff, V. L. (1962b). Gamet,ogenesis nnd spawning of the European oyster, 0. edulis, in waters of Maine. Uiol. Bull., Woods Hole, 122, 86-94.
Loosanoff, V. L., and Davis, H. C. (1950). Conditioning V . mercennria for spawn- ing in winter and breeding it,s lnrvne in the laboratory. Biol. Bull., Woods Hole, 98, 60-66.
Loosanoff, V. L., and Davis, H. C. (1951). Delaying spawning of lamellibranchs by low tempcrature. J . Mar. h a . 10, 197-202.
Loosanoff, V. L., and Davis, IT. C. (1952a). Rcpoated semiannual spawning of northern oysters. Science, 115, 675-676.
Loosanoff, V. L., and Davis, H. C. (1952b). Temperature requirements for maturation of gonads of northern oysters. Biot. Butt., Woods Hofe, 103, 80-96.
Loosanoff, V. L., and Engle, J. B. (1941). Little known enemies of young oysters. Science, 93, 328.
Loosanoff, V. L., and Engle, J. B. (1942). Use of complete fertilizers in culti- vation of microorganisms. Science, 95, 487-488.
Loosanoff, V. L., and Engle, J. B. (1947). Effect of different concentrations of microorganisms on the feeding of oysters (0. virgin&). Fish. BuU., U.S.. no. 42, 51, 31-67.
Existence of physiologically- different races of oysters, Craasoatrea virginica. Biol. Bull., Wood8 Hole,
Loosanoff, V. L., and Tommors, F. 1). (1948). Effect of suspended silt and other mbstunces on rate of feeding of oysters. Science, 107, 69-70.
Loosanoff, V. L., Miller, W. S., and Smith, 1’. B. (1951). Urowt.h and setting of Inrviio of V ~ U N mrrcetuwin i r i idst,iori to t , 6 I l i ~ i ~ ! ~ U t , l l ~ i 3 . ./. mur. Ilea. 10, r,l) HI.
IhtInvior of‘ c h r i
ItLrvtw i t 1 tliVforont coirc:oiiLriit.iotiH of’ looi l orgunisrnH. Anat. Ztec. 117, GHU --fiH7.
Loosclnoff, V. L., IlnviR, 11. C., und ( h n l e y , 1’. 12. (1955). Food requirements of some bivulvo larvae. Proc. Nut. ShellJsh As8. 45, (1954), 66-83.
Loosanoff, V. L., Hanks, J. E., and Ganaros, A. E. (1957). Control of certain forms of zooplankton in mass algal cultures. Science, 125, 1092-1093.
LovBn, S. (1848). Bidrag till Kannedomen om Utvecklingen af Mollusca Aceph& Lamellibranchiatu. 1-39, Stockholm.
Lucas, C. E. (1947). The ecological effects of external metabolites. Bio2. Rev.
Lucas, C. E. (1961). Interrelationships Iietwecn aquatic organisms rnodiated by external metabolites. Publ. Amer. Ass. Adwtnc. Sci. 67, 499-4517.
MacBride, E. W. (1914). Text-book of embryology. Invertelrufu, 1 , 1-692, Macmillan t CO., New York.
Miyazaki, I. (1935). On the dovelopmnnt of ~onie marine bivaIvt:R, with rcyu:c:ial reference to the shefled larvae. J . Imp. Fi8h. Znxl. 31, I - i 0 .
133, 2070-2071.
Loosanoff, V. L., and Nomejko, C. A. (1961).
101, 161-156.
. ,
I4ooHniioff, V. I,.. IhviH, 11. ( J . , ILII~I (!hiilltry, 1’. 15. (l!j!i:i).
22, 270-295.

REARING OF BIVALVE XOLLUSKS 135
Mortensen, T. (1938). Contributions to the study of the development snd l ~ s l fonna of Echinoderms IV. K . dumke vid.emk. Selsk., Naturh. & Math. Afd., Raekke 9, 7 , 1-59, Kebenhavn.
Nelson, T. C. (1921). Aids to successful oyster culture. I. Procuring the seed. Bull. N . J . apic. Exp. Sta. no. 351, pp. 1-69.
Nikitin, V. N., and Turpaeva, E. P. (1959). The euryhalinity of some species Of
the Black Sea Benthos and the possibility of their settlement in the Sea of Azov. Trans. of Imt . of Oceanology, 20. 4b-66. Mar. Bio1.-USSR Acad. of Sci. Press, Moscow, 1957. Published by: Amer. In&. Biol. Sci., 1959.
Odhner, N. H. (1914). Notizen uber die Fauna der Adria bei Rovigno. Beitriige zur Kenntnis der marinen Molluskenfauna von Rovigno in Istrien. ZooZ. Anz. 44, 156-170.
Orton, J. H. (1924). On early sexual maturity in the molluscs. Syndomya a h and Cardium jmchtum. Nature, Lond. 114, 244.
Orton, J. H. (1937). Oyster biology snd oyster culture. The Buckland Lectures for 1935, pp. 1-211. Edward Arnold & Co., London.
Pelseneer, P. (1901). Sur le degrt5 d'eurythermie de certainea larves marinea. Bull. Acad. Belg. C1. Sci. 279-292.
Prytherch, H. F. (1924). Exporiments in the artificial propsgation of oysters. Docum. U.S. Bur. Fish, no. 961 ; app. 11 to Rep. U.S. Comm. Fiah, pp.
I'rytherch, H. P. (1934). The role of copper in the setting, metamorphosin, and Ecot. Monopr. 4,
Quaylo, I). B. (1952). Structriro and biology of the larva snd spat of " Venerupiu
Rees, C . B. (1950). The identification and classification of lamellibranch larvae.
Ryder, J. A. (1883). Rearing oysters from artificially fertilized eggs, together
Sato, T. (1954). Artificial purification of oysters. Shokuhin-Eisei-Kenkyu, 6 ,
Satoh, T. (1960). Purification of edible oyster. Rep. Fur. Finh, Pre/eclural tlniv. of &f i e , 3, 627-032.
sigerfoos, C. P. (1908). Natural history, organization. and late tlavclo~moat of the Teredinidae, or ship-worms. Bull. U.S. Bur. Fieh. 27, 191-231.
stafford, J. (1912). On the recognition of bivalve larvae in plankton collcctionx. Contr. Canad. Biol. (1906-1910), 221-242.
Stafford, J. (1914). The native oyHter of Ilriti~h Columbia (Ohlrto Zurwh
1-14.
distribution of tho American oyster, 08 tTeCl virginicu. 47-107.
pullastra " (Montagu). Trans. roy. SOC. Edinb. 62, 256-297.
Hull Bull. Mar. Ecol. 3, 73-104.
with notes on pond culture. Bull. U.S. Ficlh. Comm. 3, 281-294.
51-56.
Carpenter). Rep. Comm. Ficlh. -B.C'. for the ycm ending 31 JJcc., 1013, 79-102.
Staubcr, L. A. (1950). Tho problom of phyHiologicat qwcios with apocial rofomnce to oyHtm mi11 oyt.itor clrilh. Ecology. 31, 109 118.
slillivtiii, (I. M. (IM!+). Ilivalvo ltrrvutr of Malimlrio lhy . P.E.I. Fbh. Rw. Bd (?ten. 77, I - 30.
'hornon. C;. (1 948). Itoproiluction and larval dovolopment of Danish marine bottom iiivnrtotinbtoH, with qmciel roforonco to tho planktonic larvae in the Sound (Orusrind). Medtl. Komm. Havundermg.. Kbh Ser.(d): Plankton. 4, 1-523.

136 VICTOR L. LOOSANOFF AND HARRY C. DAVIS
Turner, H. J., and George, C. J. (1966). Some agpects of the behavior of the guahaug. Venua wrcenaria, during the early stages. 8th Rep. Invest. Shellfsh. Mass.. Dept. Nut. Ree., Div. Mar. Fa'ah., Commonwealth Maw., 6-14.
Ukeles, R. (1961). The effect of temperature on the growth and survival of several marine algal species. Biol. Bull., Woo& Hole, 120, 255-264.
Vishniac, H. S. (1965). The morphology and nutrition of a new species of Sirol. pidium. M ycologia, 47, 5, 633-645.
Voisin, P. (1931). Recherche8 sur la reproduction des huitres indigknes dane le Morbihan. Rev. Trav. 08. Pbhes Marit. 4, 369-378.
Walne, P. R. (1956). Experimental rearing of larvae of Ostrea edulk L. in the laboratory. Fish. Invest., Lond. Ser. 11, 20, 1-23.
Walne, P. R. (1958). The importance of bacteria in laboratory experiments on rearing the larvae of Ostrea edulis (L.). J . Mar. bwl. Ass. U . K . 37, 416-425.
Waugh, G. D. (1958). Ultra-violet sterilization of water for rearing oyster larvae. Nature, Lond. 181, 1747.
Wells, W. F. (1920). Growing oysters artificially. Conservationist, 3, 151. Wells, W. F. (1927). Report of the experimental shellfish station. Rep. N.Y.
St. Conserv. Dep. no. 16, 1-22. Werner, B. (1939). ffber die Entwicklung und Artunterscheidung von Muschel-
larven des Nordaeeplanktons unter besonderen Beriicksichtigung der Schalen- entwicklung. Zool. Jb. Abteilung Anatomie und Ontogenie, 66, 1-54.
Wilson, D. P. (1948). The relation of the substratum to the metamorphosis of Ophelia larvae. J . Mar. biol. Ass. U.K. 27, 723-760.
Wilson, D. P. (1951). A biological difference between natural sea waters. J . Mar. b id . As8. U.K. 30, 1-26.
Wilson, D. P. (1958). Some problems in larval ecology related to the localized distribution of bottom animals. Perspec. in Mar. Biol., Pert I, pp. 87-103.
Winslow, F. (1884). Report of experiments in the artificial propagation of oysters, conducted a t Beaufort, N.C., and Fair Haven, Conn.. in 1882. Rep. U.S. Fish. Comm. 1882 (1884), 741-761.
Woelke, C. E. (1960). Preliminary report of laboratory studies on the relation- ship between fresh sulfite waste liquor and the reproductive cycle of the Olympia oyster, Oetrea lurida. Res. Bull. St. Wmh. Dep. Fiah. no. 6,
Wood, P. C. (1961). The principles of water sterilisation by ultra-violet light, Fkh. Invest. Lond.,
107-148.
and their application in the purification of oysters. Ser. 11, 23, 1-48.
Yonge, C. M. (1960). Oysters. pp. 1-209, Collins, London. Yoshida, H. (1938). Notes on the veligers and the young shells of Mya arenaria
Yoshida, H. (1953). Studies on larvae and young shells of industrial bivalves
Yoshida, H. (1960). On the early life-history of Tape8 variegata Sowerby.
japonicu. Venue, Kyoto, 8, 13-21.
in Japan. J. Shimonoaeki Coll. Fish. 3. 1-106.
J . Shinwnoseki Coll. Fish. 10, 115-118.

THE BREEDING OF THE NORTH ATLANTIC FRESH WAT E R- E E LS
The late ANTON F. BRUUN . Copenhagen, Denmark
Editor'a Note In August 1961 I wroto to Anton Bruun asking whether he would prepare an article
for the first volume of ' I Advances in Marine Biology". I saw him in Copenhagen the following Octobor and ho told me that he had nearly fininlied a manuscript in support of S<*hinidt,'H Ilitwry (111 tho iiiigriitioiin of tho id. A fow wookn lator ho diod. 11, March I !NV~ tho ~ I I I L I I I I H I ~ ~ I ~ P ~ . WIIH H o i i L to rho froin Copiihiigwi by 'I'orhnn Wolff.
It wa.~ originally iritcndotl that t l i t : I q m r ~11oulil bo undor the joint authorship of Bruun and Bertelsen, but subsequently it was decided that there should be one author. The manuscript waa in the form of draft sections which Bruun had intended to knit together and round off with a concluding section.
It was obviously unfinished, but nevertheless since Anton Bruun was one of the foremost authorities on the subject and had worked side by side with Johs. Schmidt I felt that i t should be published if possible. The manuscript waa then seen by N. B. Marshall, who agreed.
KO alterations have been made to the original toxt other than minor grammatical amcndments to the English. Bruun was a maater of languages and the paper haa been left much aa his friends would have liked to hear him say it.
It is a pleasure to be able to publish this posthumous work aa a tribute to a great zoologist and oceanographer who had the esteem and affection of so many in all parta of the world.
I wish to thank Dr. E. Bertelsen, Dr. Torben Wolff and Mr. N. B. Marshall for their assistance. Mr. J. Boiitius kindly helped in the preparation of the lkt of references.
1 um indoliid t,o 1.;. IhrtolHoIi for tho followiriy information.
F.S.R.
I. 11.
111. IV. V. VI.
VII . V I I I .
I S . s .
xr.
I 137 . . I ari . . 141 I . 142
. . 147 & . I54 . . 156 .. 162 .. 164 .. 166
. . 1411
I. INTRODUCTION
Sincc the publication of " The Urcoding Places of the Eel " by Johs. Schmidt (1925), and even before that, no scrious objections have been raised against Schmidt's well-founded conclusions from 20 years of extensive and intensive observations, until D. W. Tucker (19594 put
137
I. 11. 111. IV. V. VI.
VII . VIII. IS. s .
xr.
I 137 . . I ari . . 141 I . 142
. . 147 & . I54 . . 166 .. 162 .. 164 .. 166
. . 14n

138 ANTON F. BRUUN
forward “A New Solution to the Atlantic Eel Problem”. In this article Tucker (p. 496) rejects Schmidt’s assumption that the European fresh- water eel (Anguilla anguilla L.) returns to the Sargasso Sea and ‘( pro- poses a new and drastically different interpretation of the observed phenomena, namely: (1) That the European eels need not and do not succeed in returning to the ancestral spawning-area, but perish in their own continental waters. (2) That the American and European eels are not distinct species, but merely eco-phenotypes of Anguillu anguilla, their apparent distinguishing characters being environmentally determined (in the manner of numerous precedents) by demonstrable differences in temperature conditions encountered during the ascent from different parts of the American eels’ spawning area to the surface, and their distribution by demonstrable coincident differences in the subsequent transport of the surface water-masses. (3) That the populations of the so-called ‘ European ’ eels, A . unguilla, are therefore entirely main- tained by reinforcements of larvae of American ‘A. rostratu ’ parentage.’’
Tucker’s hypothesis thus leaves us with the implication ‘( that Europe and North Africa are regularly colonized by eels of American ancestry doomed to perish in a fruitless suicide-migration.”
No wonder this statement has had a great appeal to the general public and has been very widely spread by magazines and newspapers. With Dr. D. W. Tucker’s background, the British Museum (Natural History), it is quite natural if teachers of biology have come to doubt whether Schmidt was right in his solution of the problem about the breeding places of A . anguilla. They may think this is still a problem in which two hypotheses are possible; for Tucker claims at least parity with Schmidt. But I propose to provide sufficient evidence to prove that this is far from being the case.
Three scientists have already put forward @XJd argurricritH iqpinwt Tucker‘s ideas (D‘Ancona, 1959; Jones, 195!); l ~ c c l ~ l ~ ~ r , I O A O ) . I n r:ac:ti case Tucker (195Yb, lSWc, 1960) has fully mttintairicd n r i r l c:xtc:iltIt4
his own ideas. 1 shall restrict mync?lf to dincuHnirig what ‘l‘iwkw hirnnc*ll‘ has put forward. I h v c fourid it impowitilc to i11(:11itIc rwicwe in g(wml (:VCII i f t h y havc h r i i i i favour of ‘I‘uckor’n i(I(:a~ (Jde ‘J’ucker, I!G!k, 1’. 1283).
Tucker has founded his hypothcsis on part of thc cxisting literaturo without any new observations of any kind; so although I have hecn working with Schmidt and Schmidt’s collections since 1924 I shall not use anything unpublished from this material; but will only mention that any new observations which may have been made or old observa- tions studied in more detail than already published by Schmidt, have only given further support to Schmidt’s conclusions.

THE BREEDING OF THE NORTH ATLANTIC FRESHWATER-EELS 139
11. AnguiUa anghilla L.-A. rostrata LE SUEUR THE TAXONOMIC SITUATION
Ege (1939) has shown that the two species are more closely allied in every respect than either is even to A . japonica Temminck and Schlegcl, which is quite closely related to both. The two Atlantic species are most easily distinguished by thc number oi vertebrae. Schmidt (1913) examined sixteen samples between Iceland and Cyprus, altogether 2775 specimens of A . anguilla, and found from 110 to 119 vertebrae, average 114-7, only one having 110 and seventeen 111 vertebrae. Furthermore in four samples of A . rostrata between St. Laurence and St. Croix, J7irgin Is., 361 specimens had vertebral complements varying between 103 and 110, average 107.1 vertebrae; twelve had 110 verte- brae. Later on Schmidt (1916, p. 6) and Ege (1939) added further material of A . rostrata, so that 962 specimens have been examined, showing a variation of from 103 to 111 vertebrae, average 107.2: thirty- three had 110 and three had 11 1 vertebrae.
The differences in the average numbers of fin rays and branchio- stegals are too small to be useful in identification. While Schmidt was fully aware of the fact that the numbers of vertebrae and fin rays may vary to a certain extent under the influence of the environment, this has not been shown €or one character, the dentition, which Schmidt also used. Ege (1939, p. 122) has studied the proportion between the length of the intermaxillary-vomerine band from the foremost point to the hindmost tip, and the length from the same foremost point to the posterior end of the right maxillary band. This character varies to a certain extent according to sex and the total length of the fish. Out of Ege’s detailed analysis we may select a5 an example the females of a length between 30 and 40 cm. The percentage values are, for A . japonicu 77-7, A . rostrata 69.0 and A . anguilla 76.1.
When Schmidt (1927, 1928) divided A . australis Richardson into two subspecies he had a similar case, later studied in detail by Ege (1939). Here the total number of vertebrae is 112.6 in A . australis australis and 111.7 in A . austraZis schmidti. Using the same character of the dentition (Ege, 1939, p. 184) as above for A . anguilla and A . rostrata, corresponding values of specimens measuring 50-60 cm are 69.3 and 71.2, thus indicating a much smaller difference, if any, than that between A . rostrata and A . anguilla.
Tucker treats A . anguilla and A . rostrata as though they are dis- tinguished only by the number of vertebrae. It ahould, however, not; be forgotten that Johs. Schmidt made a closc study of the taxonomy of the genus Anguilln Shaw. This was brought together in a monograph

140 ANTON F. R R U U N
by Ege (1939) after Schmidt’s untimely death in 1933, and it may very well be maintained that no genus of fish has been treated taxonomically in so thorough a way. I may briefly summarize some observations from Ego and at the same time mention A . japonica, which is so closely related to the two North Atlantic eels.
TABLE I*
Anguilla unyuilln A . roatrah A . japonica
_ _ - _ _ _ ~ _._______ ____.__. ___ Pronnnl lcmgth as yo of totul
lo11gth . . 41-56 43-83 42’73 43.90 38.85-40.03 _ _ _ _ _ _ - - - __------- --I_
Prortrictl length - h o d lcngth total longth
as % . . 28.33-30.40 29’70-30.10 26.5626‘94 -~
Tlistanco from origin of dorsal fin to anus us yo of hotal longt,h . . 10.31-1 1.62 8.80-9.63 9*01-9*35
~
Prodorsal length without head as yo of total length . . 17.93-19‘32 20.23-21.01 17.52-17‘72
Head length as % of total length . . 11.20-13.39 12.90-13.78 12.24-13.16
Length of gape as % of head length . . 22.73 -26.51 25.88-27.23 23‘95-27.53
Nurnrr id characters
I’rnliuciiinl vort. . . 44-45.2-47 41-42.8-45 42-43-6-45 Total vortobruo . . 110-114.7-119 103-107.2-111 112-115.8-119 Branchiostegal rays . 10.8 11.0 11.4 Pectoral rays . 17.4 16.7 17.3
data havo been given by N. B. Marshall instead. For further details me Ege (1939). In his manuscript Bruun had ueed the expressions “high” m d “low”. Numerical

THE BREEDIPTC O F THE NORTII ATLANTIC FRESHWATER-EELS 141
Altogether I find that Schmidt had very good reasons for assuming two breeding populations of the North Atlantic eels, whether they be called two distinct species ur geographical races. He made these in- vestigations before he knew the location of the breeding places, but it explained to him why he had failed in finding any eaxly larval stages of A . anguilla in the Mediterranean on his two expeditions 1908-1910.
111. THE DISTANCE TO COVER DURING THE MIGRATIQN The ceiitral arc:& for the distribution of’ A . anguilla is the British
Jsles and tlic coasts of tho Buy of I<iscny, judging from the mam occurrence (JS c4vcrs nmcnding thc rivers (Schmidt, 1000, p. 196-213). $0 Tuckcr (1!15!h, p. 4!)(i) is corrcct in giviiig the distance Sor this area from the breeding places as at lcast 3500 miles on a great circle course. But in his argument against Jones (1939, p. 1281), who thinks that A . anguilla is able to cover this distance, he ovcrlooks the fact that the Azores have a population, about which Schmidt (1909, p. 139) had the information that A . anguilla is common in all the islands. This reduces the distance to cover for the eel of the Azores to about 2000 miles, or no more than A . rostrata from the New England states and Canada have to cover.
As is known (Bruun, 1937, p. 26), a single A . rostrata has been found in northern Spain; it was found among several hundred A . anguilla of which the number of vertebrae were counted. Rather a few specimens only of eel from the Azores have been examined, altogether 131 (Schmidt, 1913, p. 11) and they were all A . angzcilla. It would, in fact, be very interesting to have several hunclreds examined to we whether A . m.stmta might rr:ach thc: AzrmH morc: oflw t h i h i Ih !r to wijqjowc~. Tucker (1‘359a, p. 495) in rcferrirrg to Vaillatrt, I &!)4) rwrirr tni tui h h f , “Anguilla adults have yet to he takc:ri in t h : o p r r f w w i ( ( t i i t : firm Lhc stomach of a cadidot off thc Awrw i H O/JVkJll!ilJ’ wJfiltccl , o f JocnJ ~ ) r o v ~ ! i i ~ ~ i i ~ ( ! ) . ” 11, rnny w r y w d I 1 ) ~ of Iocd provonamw, hut it in r ivvwt I i ( * I ( w [)roof f A i d 8 t~ r.iI)oriiiig 001 IIILH 1 ~ w ol”-mcd in tho opon occaii, over vvry t1cc-j) wnlvr, whore tho qwnnwhalo is hunting other fuwt-swimming crcatirrw like tho sqiritl~ (Clarke, 1058).
I f wc sul)1)osc that tlirrc i N ono Hiiiglo : m m t r d species of Anguilla in the North Atlantic and that this has a distribution more or less like that which A . rostruta now has, then the North Atlantic islands, first the Azores, and later on Madeira and the Canaries, may very well have served as stepping stones towards the development of A . anguilkz; Jones (1959, p. 1281) is therefore right in assuming “ intense selection in favour of eels which found the ‘ right ’ spawning ground.” Such

142 ANTON F. BRUUN
selection must have been a t work, and probably still is, especially in A . rostrutu. From the known distribution of the A . rostruta lepto- cephali it may be concluded that a considerable number fail to reach the coasts, especially those found north of 40" N. lat. and east of 55" W. long. But even some of these do reach Europe as mentioned above.
IV. How DOES A . unguilla REACH THE BREEDINU PUCE? As already mentioned Tucker is of the opinion that A. anguillu
cannot reach the breeding places. He then asserts (Tucker, 1969a, p. 497) : " The east-coast American migration, by contrast, presents little difficulty. Along that coast the coastal water and immediately adjacent slope water are moving in a south-westerly direction (Fig. 3). Recent Discovery I1 work (Swallow, 1957) has confirmed the hypo- thesis of Stommel and proved the existence, 150 miles off South Carolina, of a strong south-going current (up to 84 miles a day) deep under the Gulf Stream. This system provides a simple, rapid and direct path, and without demanding any differences of behaviour among the different eastern groups of American eels. Temperature and salinity gradients encountered on this route (Sverdrup et ul., 1942) are such that active migration would reinforce passive transport. The fate of the Mississippi eels need not concern us.
" We may here meet Bertin's demand (1956) for a migration theory containing all the observed phenomena of physiology and behaviour."
We do know that the European eel shows positive reactions to higher temperature, as Tucker (1959a, p. 497) states on the authority of Bertin (1956). This is well known to all fishermen and biologists who have studied the eel. The last silver eels are caught in Denmark when the season with frost starts, but the great majority#are caught during the dark nights of the interlunar periods of August, September and October, and male silver eels may start migration as early a8 in the interlunar periods from June onwards (Otterstr~m, 1948, p. 296). In the spring the fishermen cannot use baited hooks for catching yellow eels until the water temperature reaches about 10" C, simply because the eel does not eat a t lower temperatures (Otterstrsm, bc. cit .) .
According to Matsui (1952, p. 242) the Japanese eel becomes inactive and loses its appetite when the water temperature falle below 15O C, and he advises that the feeding of stocks in eel-ponds should be stopped at temperatures below 10" C. This is in good agreement with the fact that twelve of the seventeen species of Anguilh are tropical and subtropical in distribution, and all of them breed in tropical or sub- tropical seas,

THE BREEDINQ O F THE NORTH ATLANTIC FRESIIWATER-EF1.S 143
We do not know exact temperatures in relation to the behaviour of A . rostrutu, but Bigelow and Schroeder say (1953, p. 152): " Eels tolerate a wide range of temperature. But i t is common knowledge that those inhabiting the salt marshes and estuaries of our Gulf, and its tributary streams, mostly lie inactive in the mud during the winter.)' On the basis of this statement I feel myself justified in concluding that the reaction of A . rostratu towards temperature is much the same as that of the two other northern temperate eels, the Japanese and the European.
First I may mention then that temperature conditions in the Katte- (Danish waters) as shown by Tucker ( 1959aJ p. 496, Fig. 2) have no
relation whatsoever with the migration of the European silver eels, because they do not migrate at all a t the temperatures shown, 0" to 5" c.
But, most important, Tucker, maintaining that there can only be one inherited pnttcrn of behuviour, uscs this postulih in his arguments &ut tho 1Ciiropwii id, lmt for@ nll nlmi t it ~ I i m hc ApenkR ahout t l i v Aniericrin wi.
His scntencc. thn t tlic: fiitc of the hiissiwi1pi wls riced not concern us rnakes onc ask why 1'11cIwr uses so m w h space in arguing about the Mcditerranean stork wlicn considcring t he 12uropcan eel; even if the observations ON p. 4!)7 slioiild be of minor jml)ortance in the light of new and well-established facts in favour of Ekman's (1932) and Tucker's hypothesis, i t might just as well be stated that the fate of the Mediter- ranean eels need not concern us.
But as regards the simple, rapid and direct path of the East American eel, leading to the breeding places, I fail to wc its simplicity, keeping in mind that the breeding must take placc ahol I: the occanic tlicrmoclinc, probably not deeper than 500 m, a t tempcr"ttures of about 15°C. The deep countercurrent of the Gulf Stream off South Carolina is a true abyssal current. Instead of referring to Swallow ( 1 957), as Tucker does, I may be allowed to cite Swallow and M'orthingtc~n ( 1 959), which waR published well before Tucker" last article ( 1 !,tIO). SwJlow rmrl Worthington write (1956, p. 444): " Southwly volocition of tJ€3tW(!f:tl
2 and 9 cm/sec were mcasiired at i t dc:pth of :!58O 111 arid twtwcmn I) r ~ n ( 1 18 cm/Hec at 2840 m.
" The Icvc!l of no motion ww dctc*rrriiiicd from tho floatn o i i fivo occasions. It fcll between 1649 arid 21MJ m mtl txtwocn tho pohntial temperature surfaces 3.43" and 3-68'. "
So Tucker, assuming that the American eel, like the European, shows positive reactions to high tempenLture and salinity, disregards the fact that the American eel, in order to tuke advantage of the deep

144 ANTON F. BRUUN
countercurrent of the Gulf Stream, will have first to head for higher temperature and salinity, then to dive to abyssal depths with a very low temperature and salinity, and finally to rise again to the level about 500 m below the surface with water of a higher temperature and .salinity.
I do not intend to discuss fully how the American and European eels may find their breeding places, but will only refer to Hasler’s excellent review (1956). After mentioning his experiments with the minnow which can smell the difference between water from Georges Bank and samples from the Sargasso Sea, he writes (p. 461): “ Salinity, gases, or odors a t any one placc at sea xeeni to me to be signals tha t would be more meaningful as appropriate (‘UCR for informing the fish it is, for exaniplc, in a place to spawn, and not for directional orientation. To help in reaching a distant goal, the cdge (sheer) effcct presumcs that the fish might follow the border of two water masses, but it leaves unex- plainrd what might direct i t straigltt ahead or the reverse.” In the case of the eel he says (p. 462): “ I t niight be conceivably stimulated by diffwcnccs in sheer between the Gulf Stream and the water masses on each side. But more likely a chemical difference might also serve as signals, in which case by swimming straight ahead, with the odor of the Gulf Stream on the left, it could arrive in the vicinity of the spawning grounds of the Sargasso Sea.”
I would also like to call attention to Waterman’s (1959) paper on animal navigation in the sea. He reminds us (p. 253) of experiments showing that aquatic animals can use the Sun and perhaps the Moon for determining compass directions, as do many terrestrial animals, and he also mentions the possibility of an addition to the direct effect of the Sun’s image from the polarization of natural unr1c:rwrrtor illumitta- tion; furthermore that laboratory studies yicldorl g r d cvid~rico tfrnt the fish’s directional sense is regulated by an intcmial clock mcdrunintn which compensates for the Fjun’H movcmcnt through thc: Hky an i i i many terrestrial forms.
Mtiiir (1947) has given a detailed analysk of marking experiments on silver ecls, or as ho very propcrly calls thcm, Wandoraale, migrating ecls. A total number of 882 were marked nnd no fewrr than 167 or 17.7%, were rrcaptitrcd. Miiiir docs not study the migration speed, but n fow cctlculations of migrations longer than some 500 km support the views of Jones (1959) and Deeldcr (1960) of a considerable speed, much higher than thoso accepted by Tucker. Tucker’s low estimates may have several reasons; only one may be mentioned here, that the silver eel does not migrate between sunrise and sunset in shallow water. In the open Baltic or in the Atlantic Ocean, howcver, the eel

THE BREEDING OF THE NORTH ATLANTIC FRESHWATER-EELS 145
may find its preferred layer of illumination and migrate continuously day and night.
Besides discussing his own results in comparison with all earlier experiments, Maar pays special attention to the question whether the silver eel is guided by some sense of direction, and his answer is con- vincingly in the positive ; the migration is essentially directed towards the west as seen in the open Baltic, “ in der Eigentlichen Ostsee hat sie im freien Wasser immer eine westliche Richtung gehabt, mit einer gewissen Tendenz nach dem Suden ” (Maar, 1947, p. 33). Naturally the migrating eel from the Baltic must at some point turn to the north for some time to get out of Danish waters, but here salinity, tempera- ture, currents and, perhaps, the smell of oceanic water may interfere with the directional sense.
We would like to see many more experiments carried out on the silver eel, either in aquaria or by marking experiments and other observations in free waters. But with the evidence a t hand it is not too difficult to see a pattern of the migration leading to the Sargasso Sea. A directional sense towards the west with a deviation towards south ; a preference for a small amount of dim light, which is indicated by the beheviour of the migrating fish and by the change of visual pigment in the eye from a mixture of rhodopsin and porphyropsin in the yellow eel to deep-sea rhodopsinin the silver eels (Wald, 1958, p. 1487, with further references) ; a preference for a temperature of about 15°C and the hydrostatic pressure at 500 m ; these factors combined can be found only in the Sargasso Sea, which in addition may attract the eel by its smell long before it reaches there.
The olfactory sense of the apodal fishes is well known to be highly developed, even to the extent that Gosline (1959, p. 161) asserts : “ As to sensory systems, the eels have undoubtedly the largest olfactory surface relative to body size of any fishes, and their sense of smell is notorious.” By “ eels ” Gosline in this connection means apodal fishes in general.
V. WHY HAVE No MIGRATING EELS BEEN CAVQIIT IN THE
STRAIT OF GIBRALTAR? Tucker (1959a, p. 496) writes: “The expected passage of the vast
Rlediterranean populations through the sharp focus provided by the Strait of Gibraltar has never been witnessed, despite much study ”; Jones (1959, p. 1281) replies to this: “ It is no cause for astonishment that eels are not caught in the Strait of Gibraltar, for no commercial fishing gear in use there can be expected to catch eels.” Then Tucker

146 ANTON F. BRUlJN
(1959~) p. 1282) continues : ‘( A surprising variety of fishes, including other Apodes, have been taken in deep-sea trawls by scientific expedi- tions ; I have before me 29 Synaphobranchus from a single haul of an Agassiz trawl at 1300 metres.” And further: (( I did not write of ‘ commercial fishing gear ’ in the Strait of Gibraltar but of an ( intensive study ’ ; still more explicitly, scientific investigations by Danish, French, and Monagasque expeditions.”
With an intimate experience of the gear used by the Danish expedi- tions and having been on board when they were used in the Strait of Gibraltar on four occasions I can state that no one on board ever dreamt about catching migrating eels in the utramin-nets intended for catching eel larvae, and that no other gear was used. Nor does any French or Monrtgasquc publicatioii indicate that suitable gear was ever used.
111 fact a trawl which could catch the migrating eel in the Danish waters has still to be invented. The fishing of the bottom-dwelling yellow eel is a characteristic feature of the Danish eel fishery, but all experiments aimed at catching the pelagic migrating eel with a kind of trawl have been failures ; such eels are taken in all sorts of traps.
Ehrenbaum (1930, p. 171) has commented on this problem (trans- lated from German) : ‘( Outside the Skaw no marked eels have been recaptured, as in general the capture of eels in the open North Sea, not to speak of the ocean, is extremely rare.
( ( Probably the migrating eel swims at moderate or changing levels above the bottom. In recent years, when German trawlers have used a trawl, which only lightly touches the bottom, for catching the herring, eels are occasiorially caught; t h u e.g. on 18 Novcmbcr 1!)24, N. of the Skaw i f ] 90 arid 180 m of dcptti, arid o r 1 ‘LO I M w r r r tm I W 4 nrh far from thc: Swcdish cowit off Viiigu.”
lJRir ig the 8amC kind of geru as u ~ e d for catchinK t h c ycllow celn in Danish watws, but in hthyrtl and abyueal dopthH it i H caHy to catch the i)ottom-dwc.lling Syiiuz)fiobrtrnchuR, r t ~ 1 havo cxperiencd R ~ R O in the Indian aiitl Pacific Occak. So ‘J’ucker’s reference to the capture of twenty-nine Synaphobranchus is no surprise, but not to the point, because so far thcrc is no evidence that a trawl, which was not towed along the bottom, ever brought up a single Symphobranchus.
If the silver eel migrates a t depths of 180 m, as the German observa- tion may indicate, it may even be carried out of the Mediterranean by the out-flowing current in the Strait of Gibraltar (Schott, 1928, p. 158, Fig. 10).

THE BREEDINQ OF THE NORTH ATLANTIC FRESHWATER-EELS 147
VI. THE RETURN OF THE EUROPEAN EEL TO THE SEA
Tucker’s statement (1959a, p. 496) : “ Differences between mor- phological and physiological states of the European and American eels on their descent to the sea are remarkable in a context quite apart from taxonomy, The European eel is already well advanced towards being a reproductive oceanic fish, and this despite the long journey which sup- posedly lies before it. The American eel, with a much smaller distance/ time to go, is yet relatively retarded. The two conditions are the reverse of what any reasonable consideration of adaptive characters and natural selection would predict.”
In support of this, Tucker firstly claims that the American eel is on average a larger and four-times heavier fish.
Tucker cites only Vladykov (1955), who has studied the female eels from the province of Quebec, Canada, one of the northernmost ranges, where the American eel is fairly frequent (Schmidt, 1909, p. 125). Vladykov has seen a female of 131b 5 0 2 , but also gives the average weight of 1500 adult females as close to 3$ lh (1585 g), with an average length of 36 in (91 cm). These are certainly large eels, but they do not represent any typical population ; it is much more likely that this is the case with specimens from the Gulf of Maine or Chesapeake Bay. Hildebrand and Schroeder (1928, p. 115) have given the length and weight in a series of which the two largest eels are 30 in (76 cm), 44.5 oz (1276 g) and 30.1 in (76 cm), 33.9 oz (960 g).
But without entering into too many details I would just like to cite Bigelow and Schroeder (1953, p. 151): “ Eels are said to grow to 4 f t (122 cm) in length and to 164 lb (7460 g) in weight. Full-grown females average only 2 to 34 f t (61 to 107 cm), however, and males are ~mallc;r. Any eel more than 18 in (46 cm) long would probably be a female, and one more than 24 in (61 cm) in length would certainly be one. The smallest males are about 11 to 12 in (28 to 31 cm) long, females about 18 in (46 cm).” (The metric values added by me.)
This may be compared with the conditions of A . anguilh in Denmark ; Otterstram (1914, p. 308) says (translated from Danish) : ‘‘ The eel is said to reach a length of 150 cm and perhaps even a little more and a weight of about 6 kg. Eels larger than 120 cm, 3 kg, are rare, a t least in Denmark (Saltbaekvig, 1909, 123 cm, about 4 kg). The full-grown females are a t least 42 cm ; usually they are 40 to 90 cm and weigh 1 to 14 kg; males are very rarely longer than 50 cm (the largest known was 51 cm) and weigh a t most 250 g ; the smallest full-grown males observed were 29 cm.”
$ome figures from the Mediterranean may be mentioned. Heldt I I

148 ANTON F. BRUUN
(1931), speaking about the silver eel, writes (p. 21) : “ Pour le lac de Tunis, par esemple, on est siir d’avoir, en debut de saison, une grando majorit6 de petits exemplaires: anguilles de 30 ZL 40 cm, d’un poids de 50 B. 100 g, et qui constitueront aussi la presque totalit6 des prises. Puis des individus plus grands sont cspturh: 40 ZL 60 cm, 100 A 400 g, et enfin ds tr&s gros sujets, long d’un mktre et plus et pesant 1 ou 2 kg. En Algkrie, par contre, le lac Fczzara, en communication avec la mer par la Seybouze, fournit presquc exclusivement de grosses anguilles d’un kg et plus; le lac Melah donne regulikrement 50% d’anguilles moyc.niic.s d’cnviron 300 g ct SO‘%) do grosses anguillcs clc 80 -90 cm, tl’criviroii 1 kg:.”
1 1 1 lakv ‘l’iiiiis t 1 1 ~ snidlwt group comprises mostly I I~ILICR, arid it is intcrcstiiig to riotc that the small nialcs depart before the higger females, and that the biggest females are the last to leave for the ocean ; this is in very good agreement with Tucker’s argument (1959c, p. 1282) that larger individuals are faster swimmers, except that he uses it about A . anguillu versus A . rostrata, under the wrong supposition that A . rostrata is essentially larger than A . anguilla. For comparison of length- weight I cite some figures from Gandolfi-Hornyold (1930, p. 34): 96 cm-1675 g ; 99 cm-1620 g ; 99 cm-2210 g : 100 cm-2190 g ; 101 cm-1780 g ; 101 cm-1890 g. These figures also show a very great variation of weight of fish of the same length.
Much more material has been published, but I have only wanted to show that there is no very great size difference between the two species, and that Tucker has also neglected to discuss the fact that A . rostrata males are just as small ns the Euiopem.
Nc~xl, ‘I’iictkcAr i H t,oLally wrong t~hoi i t , what, t i in t,r,rmN Hilver eel and 1)rt~t~zt~ (vQJ i r t w r i w t i r l i i I t ( * t J i i i i l t r r t z l i d , in A . u q w ’ / / u 1 J i r t H i l v w twl in morc. ~ L ~ I v ~ L I I C ( ~ ( I t h i ~ i i t h Lronw ( v b l , M y own r x p : r i t w x * orily conlirtns Sviirtlson’s ( 194‘3) slid I t u s m u ~ s ~ r i ’ ~ ( 1‘352) obwrvations, which clcarly state that i t is just the opposite; or as Sviirdmn sums it up, we should distinguish between four different stages of the eel :
1. Larval stage (Leptocephalus stage) 2. Yellow stage 3. Silvery stage 4. Nuptial stage
Svardson has observed two females and two ma1c.q in what he calls partial nuptial dress, and Rasmussen reports on two fcmaien. Natur- ally, nobody knows so far what is really the nuptial drew, the morc RO
because in none of them were thc gonads essentially more advanced than in the normal silver eel. In any case these largo-cycrl bronzu W I H

TIIE BREEDING OF THE NORTH ATLANTIC FRESHWATKR-EELS 149
are very striking in appearance and thereforc reported by the fishermen, so we can safely conclude that they are cstremely rare, considering the thousands of tons of silver eel caught in the Baltic area every year.
Vladykov (1958) assumes that the bronze eel stage in North America corresponds to that of the silver eel for the European species. I would suggest rather that it is more advanced, but no decision can be made without (t study of thc cgg-diameter in relation to the change of colour before and during the migration. I linve seen ten live bronze eels, received through the kind assistance of Dr. Vladykov, and they cer- tainly were more advanced than the European silver eel. The egg- diameters were from 0.20 to 0.35 mm in four specimens, which died within 27 days after their arrival from Canada, while those of the European silver eel very rarely exceed 0.20 mm.
Tucker (1959s, p. 496) mentions that the eel shows positive reactions to higher temperature and salinity and continues : “ there can be only one common inherited pattern of behaviour, yet this has hitherto been assumed adequate to direct the return journeys of eels leaving points all over Europe and North Africa and succcssfully traversing a great variety of conditions to a common oceanic torminal.”
We may agree with Tucker that there can only be one inherited pattern of behaviour ; but it may very well be much more complicated than we think.
As has been long known, the elvers (glass eels) work their way towards brackish or fresh water and against the currents ; this may be observed anywhere in the world where Anguilla occurs. Later on, when the yellow eel has turned into tho silver eel, they leavct the brackish water or frmh water and go for ocr~atiic salinitinfi. 1:ut not rtlwrryrr rro : Natiirn has mado tho grc:rtt e x pcrirri(mt of t(:d,iriK thc: ocl towtlrdw highcr salinities than those of tho ocean. We may cite Heldt and Heldt (1928, p. 7 ) : “ En ce qui concerne le lac de Tunis, les conditions hydro- logiques sont tout autres. Aucun apport d’cau douce no se fait dans ce lac oh, par contre, l’kvaporation concentre cn sels une masee d’eau peu profonde etalbe sur une grande euperficie.” But lake Tunis produces every year up to 110 tons of dve r eel (Gandolfi-Hornyold, 1930, p. 6), and Heldt and Heldt (1930) have shown how the elver8 enter (p. 7): ‘ I La plus frappante est que, normalcment, lcs civelh marchent dans le sen9 du courant, soit qu’il cntre, soit qu’il R O ~ .
Elles Be laissent porter par h i . 11 eUt rare qu’ellcs no prhentertt d f : b O { I C
au courant que leur vigueur pourrait aidment remontcr. NOUN n’avon~ noti5 le fait qu’8 de rares occasions et particulihrement lors do no8 premibres observations, ce qui nous fit prendre d’abord I’exception pour la rhgle gdnkrale.

150 ANTON F. BRUUN
‘‘ I1 est en outre trhs net que, si les courant qui entrent portent aouvent des civelles en grand nombre, il est exceptionnel d’en trouper en quantitb notable, aux abords du lac, par courant sortant.” On several occasions Heldt and Heldt caught from 2000 to 4000 elvers in 2 to 3 hr, and once 10000 in 14 hr, but with an outgoing current they most often caught none at all, arid only on two occasions more than 60, namely 300 and 400.
I have rcpcntcdly visited lake ‘Cunis and been shown the elvers by Or. Hcldt and Madame Dr. Heldt, who also demonstrated how literally every single silver eel was caught when the elvers had grown up. Heldt (1931, p. 23) describes the departure: “ Dans les lacs cbtiers, commes aux esbuaires, l”appe1 du large’ est nhcessaire pour decider les reproducteurs ii prendre la mer. I1 faut que l’eau de l’O&an les vienne baigner, les sollicite, les dirige. I1 faut du gros temps, une nuit obscure et un courant de flot. Si ces conditions se produisent au moment favorable, la passe sera bonne. Des lacs se vident ainsi en quelques nuits de toutes leurs anguilles matures. C’est par centaines de tonnes qu’on lea capture alors dans nos lacs de 1’Afrique du Nord.”
If for nothing else these observations, together with the well-known fact that the majority of males grow up only in waters of a salinity higher than about 10 parts per thousand, should have made Tucker hesitate writing (1959a, p. 498) : “ The evidence shows that the debility of the (European) eel which is obliged to drift passively downstream in the current is due to demineralization of the starving body a t the inception of gonadial and other ho~montt activity, and that ro-mineral- izatiori arid rcsumed livalincn~ occiir whon tho eel reaches tho ma.”
I havo Reen thousandn arid hnntllcd hundrcdn of nilver ueln, from fresh water, brackish water tlnd sea water aa well, and can fully confirm the statement of D’Ancona (1959, p. 1405) and J. W. Jones (1959, p. 1281) that the European silver eel is “ vigorous, extremely energetic and in good condition.” But Tucker (1959, p. 1282) is not “ subdued by Dr. Jones’s expericnco with thousands of eels. The groat curse of the voluminous work upon eels, upon tho Salmonidno and upon sundry other animals, has been that too much of it ha^ been mochanical and repetitive, replete with experiments unintelligcntly planned and mountains of data inadequately pondered, parochial alike in the range of its geographical experience and in it8 isolation from relevant literature from alien countries and related disciplines.”
This is just after citing Keys (1 933) whose experimental results are used as a support for Tucker’s ideas, but Krogh (1939, p. 160) has called attention to the fact that Keys unfortunately did not pay any

THE BREEDING OF THE NORTH ATLANTIC FRESHWATER-EELS 151
attention to the urine production, when discussing the change in weight taking place in eels transferred from fresh to salt water.
Apart from the facts from lake Tunis and the male silver eels in brackish wattbr, cxperimonts-whioh I l i i d i i r t dligontly pli~nii~~d- havc definitely shown that neither a dchytlration iior a deniineralization of the female silver eel in fresh water is a fact in the normal play of Nature (Koch, 1949; B’ontainc and Koch, 1950). In passing I have been permitted to cite Prof. Koch’s reccnt letter to me : “ Not a single one of the silver eels that I manipulated in Denmark did show any sign of weakness. Some of the specimens came from the fresh-water of the lake Furesi), other ones were caught in thc Baltic. Afterwards I had many specimens caught in the Meuse and Scheldt in Belgium and they too were very vigorous. The same remark was made to me for fresh- water silver eels by Miss W. Frost (Windermere) and Miss R. M. Lowe who at Windermere (in the English Lake District) manipulated a very large number of specimens caught in the Lake District on their down- stream migration. Even after having been kept without food for months in fresh water, silver eels are still very strong, &B I saw myself in Belgium.”
It is also important to note that the European eel sometimes starts its migration towards the sea before i t is in the typical silver phase (Frost, 1946, j p 47-48). Frost has mado observations on all eels caught in a trap fishing the whole width of a strcam which is the outflow of a lake: “ Some of the eels caught, although clearly seaward migrante, had not developed fully these characteristics but were turning from the yellow to the silver phase. When the trap was first used no distinction between the fully silver and the transitiond form waa made, although later the two forms were dintinguinhccl, with thc reAult thd, of thc? no migrants containing food 21 (IS? and 3dl may be takcn a8 silver eel#, the remainder being in the transitional forms, although probably including some silver specimens.”
This does provide some evidence that the silvor eel takcs food on its migration towards the sea, and, thereforc, this is anotttcr proof that it is not in a state of debility as Tucker (I95Oe, p. 498) anxcrtn.
Tucker (1959a, p. 500) cites Callamand and Fontaine ( l ! ~ ) in nupport of his view that the debility of the female d v e r eel in due to dnmirlcrsl- ization. This is only possible if Tucker can prove that Koch (194O) is wrong when he shows that the apparent dehydration is only due to the deposition in the muscles of an increasing amount, of fat which is a non- solvent phase for water. If the degree of hydration is calculated relative to tho amount of protein prestmt in tho musoIes, there is no differonco bctweeii a yollow and a silver crel,.whoth from freuh water or

182 ANTON F. BRUUN
sea water. Nor did Koch find any sign of demineralization in the female silver eel, when leaving fresh water and freshly caught.
Fontaine and Callamand, in fact, reviewed the problem about &mineralization on the female silver eel in a paper (1948, p. 318) of which it is sufficient to cite their comparison of new experiments with earlier oms : “ Toutefois l’intensitk de la d6minhralisation du milieu inthrieur nous Btait apparue, sup ces shies, en liaison directe avec dhveloppement ponddral des ovaires. Ce n’est cependant pas l& une rhgle gdn8raIe. En effet, au cours de I’hiver 1947-48, nous avons r e p du lac de Grand Lieu des Anguilles jaunes et des Anguilles argent6es Bans qu’apparaissent des diffhences significatives entre les unes et les autres. Les chlorBmies Btaien; assez variables, comprises entre 4 et 7 grammes x0 (Toutes les valeurs sont exprimees en NaCl %,), mais les valeurs obtenues sur leu Anguilles jaunes et les Anguilles argentdes 6taient Btroitement m&16es. Ainsi, l’hypochlorbmie n’est pas une caracthistique biochimique obligatoirement like Q. l’argenture de 1’Anguille et A sa transformation en Anguille argentke.”
Again, if Tucker had cited only just the last sentence, all his argu- ments about this probIem would have given his readers a more correct impression about the facts.
Fontaine and Koch (1950, p. 302) also explain the findings of Callamand and Fontaine (1940) and Callamand (1943). Fontaine and Koch also found a demineralization, when female silver eels are kept in fresh water without food for 4 to 5 months. But, as they point out, this is the situation first observed by Fontaine and Callamand, a consequence of two factors working towards the same result :
1. The absence of alimentary contribution of mineral matter
2. The non-existence in the eel of an extra-enteral mechanism for because of the starvation.
the absorption of mineral ions (Krogh, 1937).
Tucker (1959b, p. 1406) findsimportant support from Schnakenbeck’s (1934) and Berndt’s (1938) descriptions of autolysis and phagocytosis of the gut. The fact is, however, that nobody has ever found a high mortality of silver eels which could be ascribed to malfunction of the gut as an osmo-regulatory mechanism in salt water.
As Tucker (1959a, p. 497) points out, mass-mortalities of eels have been observed; but instead of mentioning a case from 1655 (Nicoll, 1836, 2, p. 234,JLide Tucker) and 1914 (Mazzarelli, 1914) it seems to me that some of the many later cases where the cause was examined should have been included. ’ In 1655 very little was known about the biology of the eel, but we now know that cold winters may kill immense num-

THE BREEDMU OF THE NORTH ATLANTIC FRESHWATER-EELS 163
bers of eels (Johansen, 1929). The eel is simply the species of the Danish salt-water fishes which suffers most from severe winters ; several other species, like Qobius niger L., Coryphopterus jIaveacena (Fabricius), Solea soka L. and others, may also suffer, but the im- portant point is that among the eels both silver eels and yellow eels are kilted.
Furthermore, bacterial diseaaes quite often cause mass kills, and great losses to the fishermen (Bergmann, 1909, 1912 ; Schiiperclaus, 1927, 1934 ; Bruun and Heiberg, 1932, 1936 ; Nybelin, 1936).
Tucker (1969a, p. 497; 1959b, p. 1408) seems to assume that the European eel dies off mass-wise not very far from the mmts ; Schnaken- beck‘s (1934) specimen, with complete occlusion of the vent, from the North Sea “ is to be taken as representing the state attained in Euro- pean continental waters by eels which survive thus.” In view of the many marking experiments, showing recaptures of eels which have migrated 600-600 nautical miles, this seems & rather sweeping state- ment, also because Schnakenbeck’s specimen was only one out of four, and that nobody else ever reported on a similar specimen.
If there really were a mass-mortality in European continental waters, not least in the North Sea and the English Channel, such a yearly occurring phenomenon would certainly have been observed. In nature weaklings and dead animals are rarely left to rot on the sea- floor ; they are eaten. This has, in fact, also been reported after cold winters in Denmark. Johansen (1929, p. 91) reports a remarkable case of a cod (Gadus mrrhucc L.), 89 cm long, containing eight large eels; this is remarkable because the cod gencrally cannot catch large eels, so they must have been weakened by the cold. Johansen also cites a letter from a fisherman, who states: “ The eel is the fish which suffers most here during ice-periods. When fishing operations are carried on with the Danish seine or otter seine on the last few days before the water is covered with ice, the cod which am caught are full of eels, and feeble eels are now and then taken up with the seines. The same OCCWH just after the ice has gone.”
The sperm-whale is no scavenger so even if Vaillant’s observation (1898) is unique so far, it is not contrary to the idea that healthy silver eels do occur in the open Atlantic, in this case in the vicinity of the Azores.
Another observation pointing to a pelagic life of the silver eel is found in the classical paper of Grassi and Calanhccio (1897, p, 427), here translated from the German : “ Eels . . . are also found in the stomachs of sword-fishes, caught in the Straits of Messina. They even seem to be their favourite food.”

154 ANTON F. BRUUN
VII. SCHMIDT’S COLLECTIONS OF Anguillu LEPTOCEPHALI Tucker (1959a, p. 499) gives the following statement : “ In fairness
to Schmidt, it must be said that he lacked much of the information now available and that his terms of reference from his Government were to locate the birthplace of the European eel. The ‘A. rostrata ’ larvae were, to him: an unexpected nuisance. In the area now suggested as the source of the ‘A. rostrata’ phenotypes the Danish Expeditions took only a score of stations (Schmidt, 1919) ; those of the R. V. Margrethe were worked in during November-December 1913, too late in the year, whiIe the remainder, a t miscellaneous dates and shallow depths, were worked by amateurs from commercial vessels, in a region of rapid currents and, on my hypothesis, minute deep-lying prelarvae. Closing nets were not used.
“ It may be objected that ‘A. rostrata’is said to breed during January- February, A . anguilla during February-March (Schmidt, 1922 ; Thing, 1938). Against this simple statement must be balanced the fortunes of collecting, the lengths of overall journeys from various starting-points, disparities in sexual maturation, velocities of currents a t various possible migration-depths, duration of spawning periods and annual variations in all these ; effects of temperature on rates of hatching and early development and rates ofascent of larvae over different parts of the spawning area. The criticism is more easily answered than made.”
But Schmidt (1925, p. 307) has really given some of the answers. He was very much aware of the lack of material from the early months of the year; therefore the Dana expedition 1920-21 and the Dana I1 expedition 1921-22 were sent out with the following result (Schmidt, 1925, p. 307):
TABLE I1
Leptocephali of A . rocltrata A . anguillu
Captain Andrea (about 1865) 1 2 Trading vessels (191 1-1916) 3 120 Margrethe 1913 24 714 U.S.S. Bache 1914 6 31 Dana 1920-21 1868 6446 Dana 1921-22 > 400 >4500
Thus Tucker’s statement gives the impression that Schmidt had only thirty-four A . rostrata leptocephali and 867 A . anguilla lepto- cephali from the Atlantic whereas he really had more than 2200 A . rostrata and more than 11 800 A . anguilla. In these figures are not

THE BREEDINQ OF THE NORTH ATLANTIC FRESHWATER-EELS 155
included the numerous A . anguillu leptocephali which Schmidt collected on his Thor expeditions in the Mediterranean and the ad- jacent seas in 1908-10 and earlier. Details of the stations of the two Dana expeditions were given by Schmidt (1929), a paper which is included in Tucker's references (1959a, p. 501).
In any case the background for Schmidt's considerations about the breeding of A . rostrata is that he had seen tiny larvae among the more than 2200 leptocephali examined. So he could write (Schmidt, 1925. p. 300) : " The early tiny larvae, 7-8 mm long, of Anguilla rostrata were taken in February. In April the average length was aSout 20-25 mm, in June about 30-35 mm, in July about 40 mm, and in September 50-55 mm (cf. Fig. 14). Toward the end of the year the larvae have attained their full length, about 60-66 mm ; metamorphosis hkee place during the winter months, and in the spring the hosts of elvers move up into fresh water." And further: " From the data to hand, then, we may conclude that the American eel spawns earlier, that its larvae grow more rapidly (cf. Fig. la), and that the full-grown larva is smaller than is the oase with the European species."
Schmidt's Fig. 14 furthermore shows that a few tiny larvae of 7 mm may still be found in April, while in July the smallest is 25 mm. The position of Dana station 948, 22"14'N., 67"22'W., is also in good agreement with Schmidt's conclusion in reference to his Fig. 5 : " From the position of the curves we can conclude that the breeding area of the American eel lies along the entire range north of the West Indian Islands. Its central portion lies west and south of the central breeding grounds of the European eel; the areas embraced by the two species, however, are apparently not separated, but seem to overlap." I would, however, emphasize that Dana station 948 lies well outside, towards the S.W., of the area where A . anguilla of any size was ever caught.
Tucker repeatedly (1959a, p. 495, p. 500) mentions the alleged finding of the eggs 'of the American eel by Fish (1927). These were taken near Bermuda Is. (32*02'N., 65"OO'W.) by the " Arcturus " on 16 July 1925. Fish, herself, was very well aware of the possibility that she might be wrong about the identification. Happily her description is very careful, and it gives good reasons for Schmidt (1929, p. 16) and especially Thing (1938, p. 315) to discard her identification. Bigelow and Schroeder (1953, p. 153) also doubt it. I would like to add that the '* Arcturus " also caught Anguilla leptocephali from May to September at the same place as Fish's eggs. The sizes of these leptocephali fit very well with the picture given by Schmidt; using the figures given by Thing (1938) it should be kept in mind that the " Arcturus " meterid W&S preserved in alcohol and therefore was more shrunken than

166 ANTON F. BRIiUN
Schmidt’s larvae, which were preserved in formalin. The ‘‘ Arcturus ” did not catch A. rostrata leptocephali until July, and ther only two specimens, 39-43 mm, which is exactly what would be expected from a glance at the catch of Dana station 891 fairly near t o Bermuda IS. (Schmidt, 1925, Fig. 14). That Tucker (1959a, p. 500) should consider a find of eggs so far north and lacking a concentrated oil-globule as explained by the assumption that they merely represented one of the many mistimed and misplaced spawnings, seems utterly improbable to me. The more so because the ‘‘ Arcturus ” also caught seven A . anguilh leptocephali in July, which also fitted into Schmidt’s pic- ture, but none was smaller t,han 18 mm, and this was a single specimen, while the rest were larger than 28 mm.
Incidentally I (Bruun, 1937, p. 9) have pointed out that Uchida (1 935) also misidentified a leptocephalus close to Leptocephalm similis as the larva of A . japonica. Uchida overlooked the possibility that another genus of apodal fish might have the same number of myomeres as the Anguilla, as is the case with Lept. similis and L. euryurus in the North Atlantic; Uchida’s larva may thus be a Pacific parallel to A. japonica, as are L. similis to A . rostrata and L. euryurus to A . anguilla.
VIII. POSSIBLE TEMPERATURE EFFECTS ON THE NUMBER OF
VERTEBRAE IN Anguilla
Out of the sixteen species of the genus Anguilla examined by Ege (1939) A. mossambica has the lowest number of vertebrae, namely an average of 102.9, varying between 100 and 106. The highest number is found in A . japonica, with an average of 115.8 and a variation between 112 and 119. This makes a total variation of 100 to 119 within the genus.
From Table 111 i t may be seen that A. japonica and A . anguilla represent the extreme high numbers, while A . rostrata in some way may be said to show a number rather close to the average number of the rest of the species.
Furthermore it may be noted that only in A. bicolor the subspecific difference may be about 2.
I do not want to discuss in detail whether one may be justified in expecting that the variation of the number of vertebrae should be directly proportional to the total number of vertebrae, when com- paring species of higher taxons than a genus. Tucker (1959a, p. 498) supposes this when comparing Salmo trutta (order Isospondyli) with Anguilla anguilla and A. rostrata (order Apodes). I would just mention

THE BREEDING O F TIIF, NORTH ATLANrIC FRESHWATER-EELS 157
TABLE 111. Average numbers of vertebrae in the species of Anguilla. Fractions disregarded. (After Ege, 1939.)
Average no. of vertebrae
1 If\ 115 I14 113 112 111 110 109 108 107 106 105 104 103
Species or aubspeciee
j(cponicn angwilln
australis australis, dieffeubachi australis schmidti, megastoma nebulosa labiata bicolor bicolor nebulosa nebulosa reinhardti rostrata, bicolor paeijca bornemis, marmorata interioris obscura mossambica, ancestralis, celebesensis
that Zoarces viviparus (order Percomorphi) is an example showing that this is not the case. I n Danish waters i t has been shown (Schmidt, 1917 ; Ege, 1942) that Zoarces viviparus cannot be divided into well-defined geographical subspecies ; there is a great variation even within the Danish waters. even so great that they represent tho highest and lowest values found within the wholc gc!ographical range of this spccies, West a n d North 1Siiropxtti consts. 'Tho lowa~t numlwr foiml i H 100 and the highcst 126 (ISgc:, 1!)42, pp. 281- 282). ThiH meann t l ~ t the species 2oarce.q vivipur"s hag R jyc:atr:r vnriittiort t h t I L ~ J npr:i:ic?w t o p t h r (Jf thr: g:c:n I I H An,!yuillri.
If any conclusion mtty bc drawn from this it would be that number of vertebrae in tho spccics of the gcnus A?ipi l la are less affected by vnristions of tlit: (vivironiiiciit than in thc: single sl)ccics of Zonrccs.
A fcw oxrtmplos of t,hc vdriation in thc numl)cr of vertebrae may show this quitc: clearly. A . marmoruto (Quoy :tnd Gnimard) has a goographicnl tlistributioti right from S. 15. Africa and Madagascar across the Indian Ocean to Indonesia arid 'J'niwan, arid farther in the Pacific to the Marquesas Is. fiot thc total variation in the numbcr of vertebrae in this huge area is only between 100 and 110, the average extremes being 104.6 (136 specimens, Celelm) and 106.4 (162 speci- mens, Tahiti). These figures are given by Ege (1939, p. 73). Other large samples from RBunion and New Guinoa are in between, a8 alRo smaller samples from many places, except cight specimeits from t h

158 ANTON F. BRUUN
Marquesas having 106.9, but this is naturally not statistically sig- nificant.
These general observations may serve as a background for the following.
With Tucker (1959a, p. 498) we may assumc that Anguilla shares the common susceptibility to factors, such as temperature, as has been shown to inf-Iuvnce the numher of vertebrae in other fishes, like Zoarca and Salmo (Schmidt, 1920, 1921 ; Tbning, 1952).
Schmidt (1!)27, 1928) was aware of this when he divided Anguilh australis Richardson into two geographical races, an eastern one from Ncw Zcaland, and a western from East Australia. After Schmidt’s death, Ege (1939) worked this up in greater detail, showing that the eastern race, 9nguilla australis schmidti Philipps, has on an average 111.7 vertebrae (1666 specimens) and the western race, A . australi,q Richardson has 112.6 (559 specimens). Similarly the Indian Ocean population of A . bicolor bicolor McClelland, distributed from East Africa, India, and Java to N.W. Australia, has 109.5 vertebrae (1251 specimens from nine localities in the whole range of distribution) and the Pacific population, A . bicolor paci$ca Schmidt, distributed from Celebes along the north coast of New Guinea, has 107.1 vertebrae (324 specimens from four localities in the whole range of distribution). This is what would be expected in a species of fish with several breeding places, but i t is certainly in sharp contrast to the conditions in the two North Atlantic species, A . anguilla having 114-7 vertebrae and A . rostrata 107.2 vertebrae. Even different year classes of A . anguilla show no variation as seen in Table IV.
TABLE IV. S o . of vertehrr: in clvcrs, caught at Epncy, ltiver Severri, England. (After Schmidt, 1913, p. 9.)
1908 1909 1911
Averageno. ofvertebrae . 114.7 114.7 114.7 Totalno. ofspecimens . 138 184 163
Now to assume that the differences between the American and the European eels are merely eco-phenotypical seems to me to be utterly improbable, even if only with the information above as a general background.
But even more so when we look into the details of how Tucker has combined results from experiments with Salmo with the observations above and the hydrography of the breeding places of the eel. When

THE BREEDIN(: O F THE WORTH ATLANTIC FltESHW'ATEIE-EELS 161)
Tucker (1959a, p. 498) writes : " Means of 56-86 and 60.06 vertebrae were obtained in batches of young from the same parents, a difference proportionally similar to that pertaining between whole populations of Atlantic Anguilla," he correctly refers to T~~ning's (1962, p. 15) observa- tion on temperature shock-effect, but omits to give tho character of the shock and the temperatures before and after the shock a t which the eggs were reared.
T h i n g (1952, p. 15) gives the results of two experiments, one in which the extreme variation of average values is 68.69 and 60.12 vertebrae or a difference of 1.43, and another in which i t is between 56.86 and 60.06 vertebrae ; this latter is the one in the quotation from Tucker above and is the very extreme example of ail Thing's experiments.
T h i n g had two control samples, one reared a t 2.7"C averaging 58.38 vertebrae and one at 12°C averaging 59.36 vertebrae.
Now the great difference, cited by Tucker, of 3.20 vertebrae waa only obtained in a very special way. The batch with 66.86 vertebrae was first reared at 2.4"C until the sensitive period, then exposed to a shock treatment of 20 day-degrees, or about 30 hr, a t 16OC, and then again reared at 2-4°C. The other batch averaging 60.06 vertebrae was first reared a t 12"C, then in the sensitive period exposed to 20 day- degrees, or about 177 hr, at, 2.7'6. In short, the extreme result was only obtained if the eggs were reared a t a certain low temperature, then in the sensitive period suddenly exposed to a quite different, very high temperature in a rather short time, after which the eggs were moved back to the original low temperature. These are conditions which cannot possibly be similar in any way to temperatures influencing the eel eggs during their development.
Here I would like to cite Schmidt (1929, p. 16) : " The larvae recently emerged from the ova (praelarvae and quite young larvae) are extremely fragile, so much so that the majority undoubtedly perish in the nets even under the most favourable weather conditions, and our hauls therefore are not likely to present a true picture of the quantitative proportions between praelarvae and the somewhat larger larvae. Of the European eel, we have comparatively many praelarvae and quite young larvae, in which the oil globule of the yolk sac has not yet been re- sorbed. These are found somewhat deeper down than rather older larvae, but still in what may relatively be called surface strata, abt. 200-500 metres down."
This means that the number of myomeres in the larvae are so well developed that they can be counted and thus a distinction be made between A . anguilla and A . rostrata larvae ; and furthermore that we

160 ANTON F. BRUUN
are a t most allowed to consider temperature occurring between the oceanic thermocline, at about 10°C and 200-500 m below the surface; the hundreds of thousands of leptocephali of species of the order of eels, Apodes, have always been caught in the oceanic thermosphere, the upper layers of the ocean warmor than about 10°C (Bruun, 1947, pp. 164-165) ; even leptocephali with tubular eyes, like those belonging to the deep-sea eel Synaphbranchus live in the thermosphere (Bruun, 1937). But it is likely that the larvae of the genus Anguilla are hatched well above the oceanic thermocline, firstly because, as mentioned above, the newly hatched larvae were actually found between 200-500 m, and secondly because the temperature preference of Anguilla is probably at least 15°C. As may be seen from Schmidt (1929) and Jacobsen (1929) 15°C was found about 500 m below the surface all over the area where the Dana found the tiny larvae of both species of Anguilla. If we take the actual observations made in the area on the stations in the area from Schmidt (1929), we get Table V.
TABLE V. Temperature observatioils at the surface, 700 m, 500 m and 200 nl ("C).
Station No.
~~
Date 700 m 500 m 200 m
1239 1241 1261 1319 1321 1322 1 :w 1335
11. ii.1922 13. ii. 22 9.iii. 22
26.iv. 22 28.iv. 32 30.iv. 22 0. v. 22 8 . v. 22
10.9 12-5 9.5
10.3 10.8 1 I . h 1 1,(1 i 2.fi
14.70 16.35 13.32 14.36 13.88 1 k46 I f i ~ l I 5 I G 4 G
20.76 21.43 19-83 19.93
I11.M 18.1 1
18.67
i n m
Surface
23.42 23-92 24.83 24.51 23.37 22.56 21 frl Z2,fiO
This agrees fairly well with thv picture given by Tucker (l959a, Fig. 4), exccpt that no newly hatchtd larvae have becii taken north of 28"N. ; liis suggestion of spawning taking place right down to 700 m
I can only be founded on Fish's wrong identification of four eggs from an open net which had been down at to0 fathoms. But furthermore the temperatures below 500 m are probably Loo low, only about 9 to 12OC or well below the preference of Anpi l la . But apart from this Tucker's conclusions arc against the observed facts. He says (p. 498) : " Assume a linear spawning area extending from 20-30"N. and at a depth (400- 700 m) below the influence of surface currents. Eel eggs, rising through the lower layers, will encounter a substantially identical temperature

THE BREEDING OF THE NORTH ATLANTIC FRESHWATER-EELS 161
gradient until they strike the 18°C isotherm a t about 300 m. From then on several conditions become differentiated simultaneously, roughly in relation to latitude. Eggs and larvae a t the southern limit pass through the gradient 18-25°C during the last 250 m ascent; in particular, they encounter an abrupt 4" rise between 200 and 100 m which may well have the effect of ending segmentation and so fixing the number of somites/vertebrac in thc range 103-1 1 1 rccognizcd as typifying 'Anguilla
" At the northern limit, however, the temperature gradient remains gradual right up to the surface, where a maximum temperature of barely 20°C is encountered. No temperature effect, certainly no shock effect, is likely, wherefore the addition of somites presumably continues to the genetically determined limit and results in the population with 110-119 somites/vertebrae which we recognize as 'A. anquilla '."
As pointed out above all newly hatched larvae had the full number of myomeres, and they were caught between 200-500 m, never in nets fishing closer to the surface ; I may add that even older larvae are never caught close to the surface, but always a t about 25 m or deeper. Tucker's remark that closing nets were not used does not disqualify these statements because the newly hatchcd larvae were only caught when the nets had been down between 200 and 500 m, and older larvae were only caught quite rarely if the nets had not been down at 25 m or deeper. Finally I may add that there is no indication of any sort of a secondary thermocline anywhere which could possibly cause some shock effect even of the type mentioned by T&ning (1962, p. 15); here TBning had two control samples, one kept a t 4°C averaging 88-94 verte- tmc, arid one at 10°C nvcrfigi~ig 59.60 vc!rt.ol.)rlu~
Ily 1mLtidiwiiig IL H I L ~ U I ~ , firHt r w r w l i d , do(!, i t 1 tho wln i t ivc t IwriCrcl to 10" aid kocying it Lhcre hc obtairwl i ~ i i avcrago of litC.liD, or only 0.26 vcrtebrao lowcr than thc sample kept f iw the whole time at 4'C.
Apart from this slight lowering of thc average number of vertebrae, fair too small to satifify Tucker's idea, it i8 impossiblc to imagine that thorc would be II sharp limitation betwccii tho conditions in tho sea crwtitig 'I'ucIwr'H iniaginary shock arid where tliey do not exist. The distance betwcw 'LOON. and 25"N. is 300 m:a-miJes, so the slope of the isotherms is in fact very gradual, and if 'I'ucker were right we would have very many intermediate figures, mabing i t impossible to distin- guish between the two species of leptocephali.
Therefore I fail to see any support for Tucker's main assumption, that the difference between the American and the European eel is only eco-phenotypical. Field studies and experiments are both against it.
r08h'dU *.

162 ANTON F. BRUUN
Ix. PARALLEL CASES AMONG NORTH ATLANTIC APODES While studying the leptocephali of the two North Atlantic species of
Anguilla, Schmidt at the same time collected probably at least 100 000 specimens of leptocephali of other species of Apodes. He published many studies on these ; but here we may only discuss his findings regards the two North Atlantic species, Conger conger and C. oceanicw Mitchell (Schmidt, 1931).
Schmidt had about 2500 larval Conger and gave the number of myomeres in 201 C. conger as varying between 154 and 163, average 158.2, while 288 larvae of C. oceanicus showed 140 to 149, average 144-6 myomeres. The distribution of the leptocephali of the two species is a striking parallel with that of the two species of Anguilla, except that Conger conger, as Schmidt points out, breeds along the whole extent of the area from the Sargasso Sea to the eastern part of the Mediterranean ; throughout this extent he found tiny larvae, only 10 mm in length, always over great oceanic depths. As to C. oceanicw, Schmidt found quite tiny larvae in the West Indies, indicating a breed- ing place there.
From this i t is obvious that Conger requires a much less special environment for its spawning than Anguilla. This is also borne out by the fact that Schmidt found leptocephali of a third species of Conger in the South Atlantic, off Argentine, where no Anguilla species occurs.
Tucker (1959a, p. 497) states: “ In the Mediterranean, again, are several deeps which offer conditions apparently identical to those of the Sargasso, yet eels do not spawn in them nor anywhere in that Sea. In the open Atlantic the Cape Verde Basin, Monaco Deep and West Indian Basin, all on the obvious migration route, are likewise devoid of young larvae.”
It is very easy to show that all these areas each have quite charac- teristic features of their own ; it is in fact more easy to point a t differ- ences than a t similarities in their hydrographic structures. For the time being we can only consider Conger and Anguilla as one example out of very many which discriminate in different ways in the choice of breeding places.
In 1937 I added two more cases similar to Anguilla and Conger, namely that of Synaphobranchus kaupi Johnson and S . infernalis Gill and of the two very closely related Leptocephalus species L. euryurw Lea and L. similis Lea.
The parent species of the two last mentioned species are not known with certainty ; but as regards the adult Synaphobraltchus I definitely know that they are of bathyal and abyssal distribution; so i t is no wonder that the bottom-dwelling adolescent and adult stages have

THE BREEDING OF THE NORTH ATLANTIC FJEESHWATER-EEI,S 163
partly overlapping distributions. Both species breed in the Sargasso Sea close to the West Indies; the parallelism to the two AnguiUa species is found in the maximum size of the unmetamorphosed lepta- cephali, being about 90 mm in the western species, S. infernalis, and about 120 mm in the eastern species, 8. kaupi. Furthermore the duration of the leptocephalus stage is about 8 months in 8. infernalis and about 20 months in S. kaupi. The number of myomeres counted in 40 S. infernalis was 131-140 and in 262 S. kaupi 143-154.
Tucker (1959c, p. 1283) has commented on this as follows : “Possibk parallel cases. Bruun (1937) has commented on the hitherto unex- plained coincidence that, of four pair8 of Atlantic apodal ‘ species ’, having distributions roughly similar to those of the two types of Anguilla larvae and, moreover, larvae which can be taken ‘ at the same place and same depth within a certain area of the Sargasso Sea ’, the American ‘ species ’ of each pair has the lower number of vertebrae (Table 1). This situation may well be due to a common eco-phenotypy rather than to coincident genetic effects.
“ TABLE 1. Numbers of Vertebrae or Myomeree in some North Atlantic Eels, mainly after Bruun (1937).
AnpiUaanguillu . AnguiUa rostrata . Conger conger . Congeroceanicue . Synaphobranchue kaupi . Synaphobranchw infernalis Leptocepholue euryunu, . Leptocephalua similie .
Ewt Attantic
110-119
164-163
143-164
111-1 19
- - -
-
West Atlantic
- ’ 103-111
140-149 - -
131-140 - 1CM-113 ”
This table is identical with Table 10 of Bruun (1937), except that I had used the Figures given by Schmidt (1 916, Table 11) for two random samples of 266 specimens of each of the two Anguilla species ; Tucker, more correctly, has given the full variation, probably from Schmidt (1913), thus adding the two very rare variants of 110 in A. anguiUa and 103 in A. roslrata. But this does not alter the picture in any way. In Synuphobranchw and Conger there is no overlapping whatsoever in the numbers of myomeres or vertebrae, while thwe is a very slight one in the two other genera; in Anguilla and Syri.phol~ranchw there is a certain horizontal overlapping of the Imeding places, as also for
12

164 ANTON F. BRUUN
Conger. To what extent there is a vertical overlapping is not known, but Tucker's hypothesis, that each of the genera is represented by one species only, excludes the possibility that the eastern and western species might have different preferences aa to temperature when spawning, hence spawning at different levels. It seems impossible to understand the case of the two Conger species without assuming the existence of two well established species. About C. conger we know that we have at least two well-defined breeding populations, one Meditm- ranean and one Atlantic (Schmidt, 1911). But nothing indicates tha,t the Sargasso Sea spawners of C. oceanicus should be the parents of the leptocephali of C. conger occurring in this area. So here i t seems quite impossible to agree with Tucker that this situation may well be due to an eco-phenotypy rather than to genetic effects. But if this is 80,
Syqhobranchus can be no different; never have any experiments shown shock effects of a magnitude such aa the differences in the num- bers of vertebrae in the two species of Sywpiaobranclius, not to spertlr of the Conger species.
So when A. anguilla-A. rostra& and Leptocephalus euryurw-L. sirnilis are considered, it seems most logical to consider them as paralIe1 cases in the sense that they also represent pairs of genetically different populations, and that they may not well be due to a common em- phenotypy as Tucker suggests.
x. OTHER SPECIES OF APODES BKEEDINU IN THE SARQASSO SEA
Out of the many thousands of leptocephali collected in the North Atlantic by Schmidt's efforts, only a few species more than hitherto mentioned have been examined.
One of them was treated by Schmidt himself in four papers (1912, 1930, 1931 ; Schmidt et al., 1932),; this was the leptocephalus of Nessoritamphus ingoljianus. '' Nessorhumphus ingolfinus must be one of the commonest species of the North Atlantic. We have not yet counted up the total number of larvae of this species from all om cruises in the Northern Atlantic, but I may mention, that on the summer cruise of 1920 with M/S Dana I, no fewer than 4103 specimens were taken (against 5959 of Anguilla mlga9&).
" Nessorhumphus ingoljianus spawns in the Sargasso Sea at the same period as the Eel, i.e. in spring and early summer, somewhat later h the east than in the west, and the metamorphosis of the larva tak- place especiaIly in the months of August, September, and October." (Schmidt, 1930, p. 374). For this species Schmidt also identified the egg, and hatched the larva, and was thus able to make firm connection

THE BREEDING OF THE NORTH ATLANTIC FRESHWATER-EELS 165
with the larger leptocephalus, which he could follow through meta- morphosis stages to a t least adolescent specimens ; the type specimen, caught mesopelagically, is 249 mm in length.
For our discussion the important point about this species is that, although breeding in the same area in the North Atlantic as the geminate species of Anguilla, Conger, Synaphobranchus, and Lepto- cephclus similis-euryurus, there is one and only one species in the North Atlantic; its number of myomeres being 150-160.
In fact, one could also speak about geminate species in Nesaor- hamphus, but here the breeding places arc totally separated, the other species in the Atlantic Nessorhamphus danae Johs. Schmidt breeding around the Equator and having 135-145 myomeres. And then again a single leptocephalus found in the South Atlantic indicates a subtropical population of N . ingoljianus corresponding to the North Atlantic (Schmidt et at., 1932, p. 252). Finally we may refer to the world- wide, but sporadic occurrence in all oceans; it is equally important to note the slight variation in number of myomeres from one population to another within the same species, and that the occurrences, except for the Equatorial and South Atlantic, coincide with areas where leptocephali of Anguilla species also occur.
The other apodal fish to be mentioned here is Serrivomer beani Gill and Ryder 1883, also of a meso- or bathypelagic occurrence, while its leptocephalus is epipelagic.
Schmidt (1930, p. 374) mentions its leptocephalus, L. lanceolatus Strbmman, as one of several other Muraenoids spawning in the Sargasso Sea, but in this case spawning takes place in autumn and winter. Bauchot (1959) has given a very detailed account of the whole material, 5624 specimens, from which it is quite clear that there iR only one breeding population; the number of myomeres is 156 to 163.
It is also possible, however, here to speak about geminate species, the other species being Serrivomer brevidentatus Roule and Bertin 1929, and its leptocephalus L . lanceolatoides Johs. Schmidt 1916 with the number of myomeres of 155 to 165. Thus the number of myomeres are essentially the same in L. lanceolatus and L. lanceolatoides, but several other characters make it easy to distinguish between the two (Schmidt, 1916 ; Bauchot, l959), and the smallest leptocephali were caught between April and August. Bauchot studied a total of 1653 specimens of this species. Figures 42 and 43, in Bauchot (1959), show clearly that L. lanceolatoides spawns farther north than L. lanceolatwr. But none of them show the slightest sign of splitting up into eco- phenotypes.
This seems in good agreement with the fact that the adults live meso-

1 GG ANTON F. BRUtTN
or bathypelagically in the huge North Atlantic eddy system centering in the Sargasso Sea.
As in the case of Nessorhamphus, similarly distributed, the biology of the adults does not induce a segregation into a western and an eastern population.
In contrast all the species of Anguilla, Conger, Synaphobranchus, and Leptocephalus sirnilis-euryurus lead an adolescent life as bottom- dwellers, more or less close to the coasts or the continental slopes. This opens the possibility of having two populations and in time--as it is- pairs of closely related species.
XI. REFERENCES
Bauchot, M.-L. (1959). Etude des larves leptocephales du groupe Leplocepha2wr lanceolatus Stromman et identification 21 la famille des Serrivomeridae. Dana Rep. no. 48, 1-148.
Bergmann, A. M. (1909). Die rote Benlenkrankheit des Aals. Ber. buyer. bid. VeraStu. 2, 10-54.
Bergmann, A. M. (1912). Kine ansteckende Augenkrankheit, Keratomalacie, bei Dorschen an der Sudkiiste Schwetfens. 261. Bukt. I, (Abt., Origin.) 62, 200- 212.
Morphologie und Histologie des Rumpfdarmes von Anguilla Juviatilia und die Veranderungen desselben im Individualzyklus. 2001. Jb. ( h a t . Ont.) 64, 4, 347-510.
Berndt, Oswald (1938).
Berth, L. (1956). ‘‘ Eels. A Biological Study,” London. Bigelow, H. B., and Schroeder, W. C. (1953). Fishes of the Gulf of Maine. Fiuh.
Bruun, Anton Fr. (1937). Contributions to the life histories of the Deep Sea
Bruun, Anton Fr. (1947). In “ Naturforskeren Johannes Schmidt,” Copenhagen. Bruun, Anton Fr., and Heiberg, Barge (1932). The “ Red DiaeMe ” of the
eel in Danish waters. Medd. Komm. Havunderaog. Kbh. (Scrie : Fiskeri) 9, 1-19.
Bruun, Anton Fr., and Heiberg, Barge (1935). Weitere Untersuchungen iiber die Rotseuche des Aals in den danischen Gewksern. 2. Fiech. 33, 374-382.
Callamand, Odette (1943). L’hguille europ6enne (Anguilla anguilla L.) Les b m physiologiques de sa migration. Ann. Inst. ochn. 21, 361-440.
Callamand, O., and Fontaine, M. (1940). Sur le determinisme biochimique du retour $ la mer de 1’Anguille femelle d’avalaison. C.R. Acad. Sci., Park
L’activit6 thyroidienne de l’angdle
Bull., U.S. 53, 1-577.
Eels : Synaphobranchidae. Dana Rep. no. 9.1-39.
211, 357-359. Callamand, O., and Fontaine, M . (1942).
Clarke, R. (1956). Sperm Whales of the Azores. ‘Dbcovery ’ Rep. 28, 237-298. D’Ancona, U. (1959). Old and new solutions to the eel problem. Nature, L o d e
183,1405. Deelder, C. L. (1960). The Atlantic eel problem. Nature, L o d . 185, 589-691. Ege, V. (1939). A revision of the genus Anguilkz Shaw. A systematic ph@-
EU coups de son d6veloppement. Arch. 2002. exp. gdn. 82, 129-136.
genetic and geographical study. Dana Rep. no. 16.

THE BREEDING OF THE NORTH ATLANTIC FRESHWATER-EELS 167
Ege. V. (1942). A transplantatim experiment with Zoarcee m t . + i p ~ L. C.R. Lab. Carlsberg (S6r. Physiologique). 23, no. 17, 271-385.
Ehrenbaum, E. (1930). Der Flusseal. Handbuch Binmnjbh. Mittehrop. 3, 169-217.
Ekman, S. (1932). Prinzipielle uber die Wanderungen und die tiergeographische Stellung des Europaischen Aales. Zoogeographicia, 1, 86-106.
Fish, M. P. (1927). Contributions to the embryology of the American Eel.
Fontaine, M., and Callamand, 0. (1948). Nouvelles recherchea sur le deter- minisme physiologique de l’avalaison des poissona migkateurs amphi- biotiquea. Bull. Mua. Hist. nat. Paris 2 ser. 20, 317-320.
Fontaine, M., and Koch, H. (1950). Les variations d’euryhalinit6 et d’osmor6gu- lation chez les poiseons. J. Physiol. Puth. gdn. 42, 287-318.
Frost. W. (1946). Observations on the food of Eels (Anguilla anguilla) from the Windermere catchment area. J. Anim. Ecol. 15, 43-53.
Oandolfi-Hornyold, A. (1930). Recherches su I’age, la croisaance ct le sexe de la petite anguille argent& du Lac de Tunis. Bull. Sta. ocbnogr. Salammnb8, no. 17, 1-50.
Gosline, W. A. (1959). Mode of life, functional morphology, and the classification of modern teleostean fishes. Syatemutic Zool. 8, 160-164.
Grassi, B., and Calandruccio, S. (1897). Fortpflanzung und Metamorphose dea Aales. Allg. Fimh. Ztg. 22, 402-428.
Haaler. A. D. (1956). Perception of pathways by fishes in migration. Quart. Rev. Biol. 31, 200-209.
Heldt, H. (1931). Le fumage de l’anguille, industrie possible dans les pays MBditerranBens. Notes Sta. ocbnogr. Salammb6, no. 21, 1-27.
Heldt, H.. and Heldt, H. (Mme.) (1928). Premieres captures de civelles dam le Lac de Tunis. Notes Sta. ocbnogr. Salammbb, no. 9, 1-8.
Heldt, H., and Heldt, H. (Mme.) (1930). Sur les modalit6s de l’empoissonnement en anguilles du lac de Tunis. Notes Sta. ochnogr. Salammb6, no. 13, 1-12.
Hildebrand, S. F., and Schroeder, W. C. (1928). I’ishes of Chesapcake Bay. Bull. U.S. Bur. Fish. 43 part 1.
Jacobsen, J. P. (1929). Contribution to the hydrography of the North Atlantic. Oceanogr. Rep. ‘ Dana ’ Exped. 1920-22, no. 3, 1-87.
Johansen, A. C. (1929). Mortality among porpoises, fish and the larger CRIII- taceans in the waters around Denmark in severe winters. Rep. Danish blol. sta. 35, 59-91.
Jones. J. W. (1959). Eel migration. Nature. Lond. 184, 1281. Keys, A. (1933).
ZoologicA, 8, 289-324.
The mechanism of adaptation to varying salinity in the Common eel and the general problem of osmotic regulation in W e e . Proc. roy. See. 112, 184-199.
Koch, H. J. (1949). Quelques caracteristiques osmotiquea de l’mguille femelle bune et argent&. Arch. int. Physiol. 57, 125-132.
b g h A. (1937). Osmotic regulation in fresh water fishes by active absorption of chloride ions. 2. vergl. Physiol. 24, 656.
K%h. A. (1939). “ Osmotic Regulation in Aquatic Animals.” Csmbridge University press.
Mat8Ui~ I. (1952). Studies on the morphology, ecology and pond-culture of the Japanese Eel (Anguilla japoniccr). J. Shimonoeeki CoU. Fish. 2, 1-245.

168 ANTON F. BRUUN
MWr. A. (1947). Uber die Aalwanderung im baltiechen Meer auf Grund der Wanderaalmarkierungsversuche im Finnischen und Livischen Mwrbusen i.d.j. 1937-1939. Medd. Undersokndnat. Sotvattenajiak. Stockh. no. 27.
Mazzarelli, G. (1914). Note critiche siilla biologia dell’ anguiila. Ztiv. Peeca Idrob. Anno 9, 16, no. 2.
Nicoll, J. (1836). ‘‘ Diary of Public Transactions . . . Scotland,’’ Edinburgh, 1836. (Quoled in Chambers, R. “ Domestic Annals of Scotland,” 2,234, Edinburgh and London, n.d.).
Nybelin, 0. (1935). Untersuchungen ubm den bei Fischen krankheitserregenden Spaltpilz Vibrio angudlarum. n4edd. UndereoknAnat. Sotvatteneji8k. Stwkh. no. 8, 5-62.
Otterstr~m, C. V. (1914). “ Fisk 11. Dmmarlts Fauna.” Copenhagen. Otterstrsm, C. V. (1948). I n “ E’iskcrict i Danmark,” Vol. 2. Rasmussen, C. J. (1952). Size and age of the Silver Eel (Anguillo anguiZZu L.)
in Esrum Lake. Rep. Danish Biol. Sta. 54, 3-36. Schaperclaus, W. (1927). Die Rotseuche des Aals im Bezirk von Rugen und
Strabund. 2. Fiach. 25, 99-128. Schiiperclaus, W. (1934). Untersuchungen iiber die Aalseuchen in deutschen
Binnen- und Kiistengewiicxmrn 1930-33. Z. Fiech. 32, 191-217. Schmidt, J. (1909). On the distribution of fresh-water eels (Anguilla) throughout
tho world. I. Atlantic Ocean and adjacent regions. Medd. Komm. Havunder- aopdaer. (Scrio Fiskcri) 3, no. 7.
Schmidt, J . (1911). Biology of the 15cI-l~%h0s, especially of the Conger. Nature, Lond. 86, 61-63.
Schmidt, J. (1912). Contributions to tho biology of somo North Atlantic species of f:els. Vidmwk. hledd. danak. naturh. Foren. Kbh. 64, 39-61.
Schmidt, ,J. (1913). First roport o r 1 I S c ~ l invostigations 1913. ltapp. Cone. Explor. Mer. 18, 1-30.
Schmidt, J. (1915). Rapp. Corz~r. Explor. hfer. 23, 1-26.
Schmidt, J. (1916). On tho early larval 3tages of the Fresh-water Eel (Anguih) and some other North Atlnrit ic Murctonoid~. Medd. Kwnm. Hnuundersegeber. (8w. FiHkori.) 5, I 20.
Schmidt, J . (1917). (Rwial investigationx I). C.Z1. Lub. Carlaberg, 13, 3 : 279-397.
Schmidt, J. (1920). Experimental investigiLtions with Zoarcea Vivipurw, L. (Racial investigstions V). C.R. Lab. Carluberg, 14, 1- 14.
Schmidt, J. (1921). Annual fluctuations of racial charactcm in Zoarcea v i v ipm8 L. (Racial investigations VII). C.R. Lab. Carbberg, 14, 15: 1-24.
Schmidt, J. (1922). The breeding places of the eel. Phil. Trann. 11. 211, 179-2015. Schmidt, J. (1925). The breeding p1acc.s of the eel. S’mithaon. Ilep. for 1924
Schmidt, J. (1927). The fresh-water Ec 1s of Now Zcaiciricl. l’ranw. I ’ ror . N . Z .
Schmidt, J . (1928). ‘I’tro fronh-wd(:r w+ (jf Aiirrlrrtllr~. Wlih wtrrrrcl rwrlwkM 01i I l l s
Ychrniclt, .J. (1020). Iritro(lu~*tiori t o t I io ()(~~.trrlc~~rtLpiiical ICoprts. Ocmnogr. Rep.
Schmidt, J. (1930-31). Nea80rhumphrcn, n. ncw cosmopolitan gerrux of ocoanic eeh
Second report on Eel investigations 1915.
Zoarcex oiriipa9ua I.. arid loral raws of tho xamo.
279-316.
Inat. 58, 379-388.
short-finrind Hpciw of‘ Arcyui lh . It1 r . nuat. M w . 16, 179 210.
‘Ilrcna ’ Ik?.pctl. I920 22, 1 1 0 . I, 1 80.
Videnuk. Medd. Dan8k nalurh. Forell. Khh. 90, 371-37G.

THE BREEDINQ OF THE NORTH ATLANTIC FRESHWATER-EELS 169
Schmidt, J. (1931). Eels and Conger Eels of the North Atlantic. Nature, Lond.
Schmidt, J. et d . (1932). “ Dana’s Togt omkring Jorden 1928-30.” Copenhagen. Schakenbeck, W. (1934). Verlinderungen im Verdauungstraktus bei Blankaale.
Schott, G. (1928). Die Waaserbewegungen im Gebiete der Gibreltarstreese.
Sviirdson, Q. (1949). Sex differentiation in oel (AnguiUa angtiilta) and occurrence
sverdrup. H. V., Johnson, M. W., and Fleming, R. L. (1942). “The Oceans.’’
Swallow, J. C. (1967). The Listener, 57, 997. swallow, J. C., and Worthington, L. V. (1969). “he deep countercumnt of the
Gulf Stream off South Carolina. Intern. Ocean. Cmgreue. 1969. Preprinte ed. by Mary Seers. Washington, D. C. 1959 (Amer. Am. Adv. Science).
Thing, A. V. (1938). Deep-sea fishes of the Bermuda oceanographic expeditions. Family Anguillidae. Zoologica, 23, 313-318.
Thing, A. V. (1952). Experimental study of moriatic charactera in hhw. BWZ. Rev. 27, 169-193.
Tucker, D. W. (19598). A new solution to the Atlantic eel problem. Ndure, Lond. 183,495-601.
Tucker. D. W. (1969b). (Answering D’Ancona, 1969) Nature, Lon&. 183, 1406. Tucker, D. W. (1969~). (Answering Jones, 1969) Nature, Lon&. 184, 1281-1283. Tucker, D. W. (1980). (Answering Deelder, 1960) Nature, Lond. 185, 691-692. Uchida, K. (1936). First record of the Anguilla-larva found in Japaneae watera.
(In Japanese, known from the author’s reprint only.) Ref. Bruun, 1937. Vaillant, L. (1898). Sur la p rknce de 1’Anguille commune en haute mer. C. R.
A d . Sci., Pa&, 126, 1429-1430. Vladykov, V. D. (1965). “ Eel-Fishes of Quebec.” Department of Fieherim,
Province of Quebec. pp. 1-12. Wald, G. (1968). The significance of vertebrate metemorphoeis. Science, 128,
Waterman, T. H. (1969). Animal navigatioir in the MA. (~UMUA ./. t twf. picia Y,
128, 602404.
2001. Am. 108,85-91.
J . CW. 3, 139-176.
of male eels in the Baltic. Rep. Inst. I”reshw. Res. DroUning, no. 29, 123.
New York.
1481-1490.
3 : 243-262.

This Page Intentionally Left Blank

SOME ASPECTS OF PHOTORECEPTION AND
I. 11. 111.
IV. V. VI. VII.
VIII. IX. X. XI.
VISION IN FISHES
J. A. C. NICOL
&urine Biological Assoeia$ion, Plymmth, Zqp!iznd
Introduction . . . . . . . . . . . . Regulation of Light Remhing Sensory Surfecea . .
A. Pineal . . . . . . . . . . . . C. The Tapetum Lucidum of Chondrichthyee . .
Refraction. Accommodation and the Receptor Layer
Extra-Oculer Reception . . . . . . ..
B. PupiUary Movement . . . . . . .. D. Retinomotor Chanp in Teleosts . . ..
Vieud Pigments and Speotral Semitivity . . .. Transmiseion by the Lens . . .. .. .. Photosensitivity and Visual m h o l d e . . .. Eaological and Behavioural Studies . . .. .. 8 ~ 0 ~ .. .. .. .. . . . . .. The Chorioidal Gland . . .. .. .. ..
Refemncea . . . . . . . . . . ..
.. ..
. . . . . . . .
. . . .
.. ..
.. ..
.. ..
.. ..
.. ..
. . . .
.. ..
. . . .
.. ..
. . . .
.. ..
.. 171 .. 172
.. 174
.. 174
.. 176
.. 176
.. 179 .. 182
.. 186
.. 191
.. 191 .. 194 .. 195
.. 200
.. 201
I. INTRODUCTION The most important organ for photoreception in fishes is the eye,
although other photosensitive sites are known. The eye, moreover, haa especial significance because it is a visual organ, capable of image+ formation. Like many other organs of fishes, the eye exhibits much structural and functional variation end specialization, the features of which have been related to environmental factors to which the hhea are normally exposed.
Descriptions and reviews of the pisoine eye are now available in goodly number, the latest of which by Rochon-Duvigneaud (1943, 1968) and by Brett (1957) point the way to the earlier literature. The present review does not aim at being comprehensive. I have restricted myself to several aspects of photoreception in fishes, especially marine fishes ; these aspects are ones in which I am most interested or in which there have been notable new advances. In particular, I have gathered together work dealing with photobehaviour and the functioning of the eye in marine environments. I have not attempted to deal with the gathering momentum of electrical studies on the tetina, which could be reviewed more profitably in conjunction with similar studies on other animals.
171

172 J. A. C. NICOL
II. EXTRA-OCU- RECEPTION Photoreception in many fishes may be mediated by structures other
than the eyes. Skin, spinal cord, brain and pineal complex &re involvd in various degrees. Cyclostomes are light-sensitive but the reoeptor- regions are not identical in different forms. Hagfish nnd lamprey8 respond to light by general or increased locomotory activity after a rather lengthy reaction period (several seconds). The hag is practically eyeless, its photoreceptors are located in the skin and occur mostly at the anterior end of the head and in the cloaca1 region. Afferent impulsee from the posterior photoreceptors travel through spinal nerves to the spinal cord. Thence they proceed anteriorly to the brain, but local reflexes can still be elicited after spinal section (Newth and Ross, 1955). The ammocoetes (larva of the lamprey) is virtually blind whereas adults have eyes. The caudal region is especially sensitive to light; photo- receptors occur in the skin and afferent pathways lie in the lateral line nerve. The middle region of the body shows some photoeeneitivity, due apparently to direct stimulation of the spinal cord by light, and there is a photosensitive region in the head in the region of the pineal complex. When the head of a lamprey is illuminated, its responses are lea8 regular and reaction times are longer than those elicited by stimulation of the tail. It still responds to illumination of the head after removal of the pineal and parapineal bodies and the paired eyes; there are obviously other photoreceptors in the head, either in the skin or in the brain (Young, 1935a).
The experimental approach just described involves detecting sensitivity of photcmcctptors nrd ddmmining operative prtthwayf3. on a different pl t i rw w! to h C O I I H J ~ ~ * ~ f h t kirrrlw of rwpori@~;~ svcikd by different phc,torcceptora. l r i thc. untrnococteR, for oxwn pie, illurninahion of the tail initiate8 Hwimmirig niovcrnentx leading to burrowing ; in the adult, simply swimming (negative photokinefh). When paired eye6 of the adult lamprey (Lampetru) are illuminated, however, the oon- sequences are uneasy movements of the animal and altered breathing rates. Moreover, lampreys undergo colour-changes, controlled by incident light. The colour-response di.sayJpears in the larva and is disturbed in the adult after destruction of the pineal complex, when the melanophores become expanded, a1 td it is completely abolished in the adult by removal of the pineal complex and paired eyes. It seems, therefore, that dermal photoreceptors of the tail are primarily respon- sible for generalized swimming movements (leading to burrowing in larvae) ; the pineal organ controls mlour-responses in the larvae and pineal plus paired eyes act synergetiaally in controlling colour-responses

SOME ASPEOTS OF PHOTORECEPTION AND VISION IN FISHES 173
of adults. No doubt the paired eyes play other roles in phototactic and directed activities (Young, 1935a, b).
Integumentary photoreceptors are presc-nt in blind cave fishes, but none was found in marine fishes examined by Parker (1909). Howover, there is evidence that the pineal body (Fig. 1) or the pineal coniplex (which term refers to the pined body anti adjoining brain tissus) is photosensitive in many teleosts. It has long been known that the melanophores of the minnow Phozinua disperse when its pineal is stimulated by light, and the pineal also influences photokinesis. Since photosensitivity is not completely abolished by destruction of the
FIQ. 1. Vertical section through the middle of tho pined region of the cstfih (A7nleiwurs). Legend : ES, spiphyeeal stalk ; EV. epiphyw a1 vwiclo ; I'C, pontrtrior wir irr i inaurt i ; S, euperior commimre. ~ 2 9 0 . (From Terry, l(r10.)
pineal, it has been postulated that thcrc: arc additional ph"tort:c~ptr,m in the wall of the diencephalon (vnn f'rbch, I!)] I ; Hchrtrror, I!t2H). Breeder and Rasquin ( 1 947) have Hhowii that t h o pirical in conwrrit~d with photokinwis in Mind characim ( A noptichlhp) from O ~ V U N ant1 they (1950) have distinguished three group of i,cleoHtH according to the character of the pineal covering. Thcw iLrc . ( I ) f i H f \ m with tranrtliit:t:rit
tissue over tho pineal (mostly light-ponitivc) ; (2) fisheH that have chromstophorm on the surface of the head, which control light reaching the pineal (these fishes have changeable responses to light) ; (3) fishes with a permanently covered pineal area (light-negative) ; and some exceptions. In some fishes, which were blinded and which had the pineal area covered, light still exerted some effect, albeit to 8 reduced

174 J. A. C. NICOL
degree. The results suggest that light was acting on some other p h o b receptors, presumably in the skin.
The responses noted by Breder and RMquin (1950) to stimulation of the pineal were photokinesis (apparently) and colour change, and responses of a like nature have been observed in soukeye salmon srnolh (Hoar, 1955). The smolts showed avoiding reactions (when sttlrtled by an observer during the day, or by a light at night), negative p h o w (or photokinesis), and colour-responses (pallor on a white baokground), Blinded fish showed no avoiding reaction by day, were startled by a light at night, were negatively phototactic (or photokinetic), and wem darker than normal fish. Fish lacking pineal organs displayed avoiding reactions by day and by night ; chromatophore-responses were variable. Fish blinded and lacking the pineal organ were very dark and were insensitive to light. The results demonstrate that stimulation of the pineal area in the salmon smolt influences the movement of pigment in melanophores and behavioural responses to light. The experiments involved variable degrees of injury to the pineal organ and to oon- tiguous regions of the brain in some fish; a minor degree of photo- sensitivity not connected with the pineal organ and the paired eyes is not excluded by these experiments. Germane to this work are resulte obtained on frogs in which action potentials have been recorded from the pineal nerve after photostimulation (Dodt and Heerd, 1962). The responses show obvious parallels to those detected in the iish retina (p. 190), and an extension of this experimental approach to the fiah pineal would be interesting.
111. REGULATION O F LIGHT ItEACHING SENSORY SUFlFACES
In fishes which inhabit regions where great diurnal changes of light-intensities take place, there are usually devicee for controlling the amount of light reaching the photoreceptors, especially the eyes. We are concerned here with photomeehanical changes ; equally important, of course, are those changes in responsiveness of retinal neural elements and content of visual pigments, which come under the concept of dark- adaptation, in its more restricted meaning (Rushton, 1959).
A. Pineal In many fishes the tissues in the roof of the skull over the pineal
region (Fig. 1 ) are translucent, thus permitting light to reach the pineal organ and, perhaps, contigupus areas of the brain (see p. 173). Often there are chromatophores in the skixi over the parietal foramen and, by their concentration and dispersion, they determine the amount of light reaching the pineal complex. Experiments involving removal of the

SOME ASPECTS OF PHOTORECEPTION AND VISION m FISHES 175
overlying chromatophores, or covering the pineal region, have p r o d u d changes in the responsiveness of Uambwia to light (Breder and Rasquifi, 1960; Raaquin, 1958; Breder, 1959).
B. Pupillmy Movement Closure of the pupil in light occurs in many elasmobranchs and in a
few teleosts (see Walls, 1942), and some degree of pupiIIary movement, albeit small, probably occurs in a great many bony fishes. Gilbert (1961) haa confirmed that pupillary closure is not consensual in sharks ; i.e. illumination of one eye causes closure of the pupil of that eye and not of its fellow. Young (1933a) was able to give a physiological explanation of this effect when he showed that the iris sphincter muscle of dogfish was not innervated, and it contraded in direct response to illumination. In rays (Raia) there is contralaterill lowering of the operoular flap of an eye when the opposite eye is illuminated (Bateson, 1890b), and this presupposes the existence of a different kind of con- trolling system in these animals. Pupillary closure is very slight, variable and negligible in some species. e.g. S q w l w acunthias.
Investigating two teleosts (Uranoscopw and Lophim) having ex- tenaive pupillary excursions, Young (1931a, b, 1933b) found that the responses are under the control of the autonomic nervous system. The sphincter is controlled by the sympathetic and the dilatator muscle by the oculomotor (parasympathetic nervous system). There am some speoies, however, notably the eel, in which there is a direct response to light, snd the pupil of the isolated eye closes when it ia illuminated. In these animals the pigmented smooth muscle fibres of the iris contraot when stimulated by incident light. Studies of the isolated iris of the eel have shown that the action spectrum for contraction has a major peak at 600 mp, and resembles the absorption spectrum for eel rhodopsin (Seliger, 1962).
C . The Tapetunb Lucidurn of Chondrichthyea There is a niirror or tapetum lucidum (L. tapete, carpet) in the eye
of chondriohthian fishes, which is responsible for the remarbble eye- shine that they exhibit. Eyezdhine, of course, is most evident in those species whose pupils remain open when illuminated and in bathypelagic species with large pupils. The tapetum lucidum contains reflecting cells that lie in the inner layer of the chorioid, immediately underneath the ohoriocapillaris (Fig. 2). It is present in most elasmobranch and it also occurs in holocephalans (Franz, 1905 ; Walls, 1942). It appears to be absent in the bathybenthic torpedo Benthobat& with rudimentary eyes ; at least one is not mentioned by Brauer (1908). A tapetum is said to be lacking in Laemargua (=Somniosw, an abyssal shark), aayliobatM
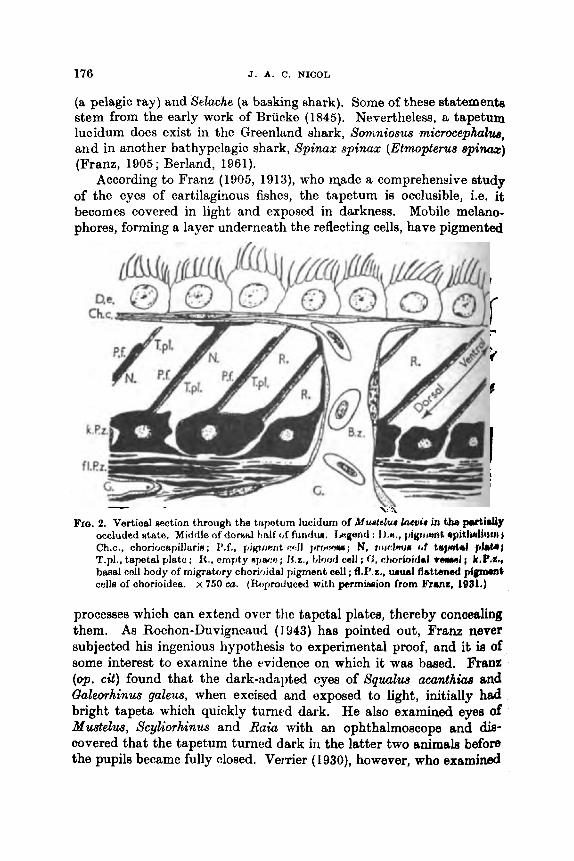
176 J. A. C. NICOL
(a pelagic ray) and Selache (a basking shark). Some of these statemente stem from the early work of Briicke (1845). Nevertheless, a tapetum lucidum does exist in the Greenland shark, Sonmiosus microcephub, and in another bathypelagic shark, Spinax spinax (Etmopterus s p i m ) (Franz, 1905; Berland, 1961).
According to Franz (1905, 1913), who made a comprehensive study of the eyes of cartilaginous fishes, the tapetum is occlusible, i.e. it becomes covered in light and exposed in darkness. Mobile melano- phores, forming a layer underneath the reflecting cells, have pigmented
Fro. 2. Vertical section through the tripetliin lucidum of Muatelur bevie in the putidy occluded atata. Middle <If dornnl tielf c,f fimdrla. Legand : I),%, pigtwmt opithitliwn I Ch.c., choriocapillerin ; I9.f,, pilpnnnt cdl pmm; N, i i i i d m ~ ~ ~ J T tayd*l plat&# T.pl., tapetal plato; It., empty spaat; H z . , blood cell ; (1, atrorbidal veml; L.P.C., baaal cell body of migratory choriaidal pigment cell ; fl.P,z., wusl fisttuned pirfgmrmt cells of chorioidea. x 760 cu. (Roproduced with permhion from Frsnz, l@al.)
processes which can extend over the tapetal plates, thereby concealing them. As Rochon-Duvigneaud (1943) has pointed out, Frau never subjected his ingenious hypothesis to experimental proof, and it ie of some interest to examine the evidence on which it was based. Franz (op. cit) found that the dark-adapted eyes of Squalus acunthiaa and Galeorhinus galeus, when excised and exposed to light, initially had bright tapeta which quickly turned dark. He also examined eyes of Mustelus, Scyliorhinus and Raia with an ophthalmoscope and dis- covered that the tapetum turned dark in the latter two animals before the pupils became fully closed. VerTier (1930), however, who examined

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 177
chiefly Sqwclua and Mustelus, observed no difference in the disposition of chorioidal pigment of dark-adapted and light-adapted dogfish.
Obviously, there is some doubt about the occlusible properties of the elasmobranch tapetum, and its presence in some species still awaits confirmation. After long neglect, interest in this structure recently haa been renewed. It has been found that in some benthic selachians from the neritic zone, the tapetum is fixed, i.e. non-occlusible ; examples are Scyliorhinua and Raia. The eyes of these animals always exhibit a bright fundus, and a black ventral field below the optic stalk. In the latter region reflecting material is absent and there ie s layer of blwk chorioidal pigment outside the retina. Pupillary movement ie rapid and extensive. In some pelagic sharks, on the other hand, the whole interior of the dark-adapted eye shines brightly, but soon beoomes very dark when exposed to light, owing to migration of black pigment which obscures the tapetum lucidum. Examples are Mustelus and Squalus (Nicol, 1961a, c). In these animals pupillary movement is slight or absent. Gilbert (1961) observed that the tapetum of the sharp-toothed shark Negaprion became occluded in less than an hour in bright light, but required 4 hr or more to become uncovered (when the shiny surfam is exposed).
The chorioidal tapetum of selachians contains guanin, to which it owes its reflecting properties. Guanin was discovered in the ohorioid of Sqwzlus, Mugtelua and Scyliorhinua by Kuhne and Sewall (1880), who found that extracts gave a positive reaction to a purine test, involving the successive use of dilute nitric acid followed by sodium hydroxide. Extracts treated with sulphurous acid gave rise to crystals of gypsum, and it has been suggested that the guanin is compounded with calcium in vivo. The presence of guanin in the chorioid of Squalus acunthiae was confirmed by Pirie and Simpson (1946), who movered guanin hydrochloride from extracts. Also, a green fluorescent substance is present, which appeara to be a pterin, xanthopferin.
Many intriguing problems, presented by the tapetum luoidum of elasmobranchs, still await clarification and resolution. Among them are the functional significance of the tapeturn in terms of visual sensi- tivity and acuity ; the mode of regulating the movement of chorioidel pigment to and fro over the reflecting plates ; the significance of the orientation of the plates in diverse regions of the eye ; and the efficienqg of the system.
It is generally believed that elssmobranchs find food ohidy by smell, and it haa been uncertain to what extent they make use of vieion. Benthic species (5cyliorhinus, R a h , Rhinu) hunt by soent, generally without making use of their eyes, but simple observation shows that

178 J. A. 0. NICOL
the common dogfish (Sc$iorhinzcs canicula) aeas an approaching net and is quite adept at avoiding capture (Bateson, 1890~). And recent work hw shown that in clear water vision plays a predominant role in guiding sharks to prey at distances less than 50 f t (Gilbert, 1962).
D. Retinomotor Changes in Tekosts In many teleosts movements take place in the retina which am
collectively termed retinomotor movements. They involve dgration of retinal pigment and displacement of rods and cones under the influence of light and other factors. The retinal pigment is contained in cells of the pigment epithelium, which bear processes that extend inwards between the receptor cells. In darkness, the retinal pigment moves outwards, towards the base of the pigment cell (withdrawd); in light, the pigment moves inwards (expansion). When movement of the rods and cones occur^, the rod myoids contract and the oone myoids lengthen in darkness ; in light, the rod myoids lengthen and the cone myoids contract. Considered in conjunction, these several move- ments cause the outer segments of the rods to be exposed in darknese ; in light, the outer segments of the con- are withdrawn towards the interior of the eye, whereas those of the rods are obscured. These changes, seemingly, are devoid of visual significance in total darknepls ; they are concerned with enhancing vision in bright and dim light.
Retinomotor changes are to be found in fishes inhabiting inahore waters and surface waters of the oceans. They do not occur in bathy- pelagic and bathybenthic species. In eels (Anguilla anguilla) the rode migrate whereaa the cones do not; the eye of the conger (Conger conger) contains few or no cones, and it is uncertain whether the rode move (Garten, 1907). It is believed that only the pigment m o m in flat-fishes (pleuronectids), and that rods and cones are stationary (Bayliss et al., 1936). It has since been discovered that cones of flat-fish extend and retract in darkness and in light, and a report concerning soles ( S o h solea) has been published (Nicol, 1964b).
The movement of pigment, cones and rods takes place rather slowly compared, for example, with the swift course of pupillary movement. Timea given in the literature for maximal contraction or expamion range from 20 to 70 min for various species and conditiona of observa- tion. In Ameiurus and Fundulus the retinal pigment attaine full expansion or retraction in 1 hr ; cones shorten and rods lengthen (in light) in 45 min ; reverse changes in cones and rods occupy 30 min or more. In Abrumis the pigment requires 45 min to expand and 30 min to retract ; cones shorten in 45 min and extend in 30 min (Arey, 1916). In Leuciscus pigment expands and retracts in 20 min to I hr; cones
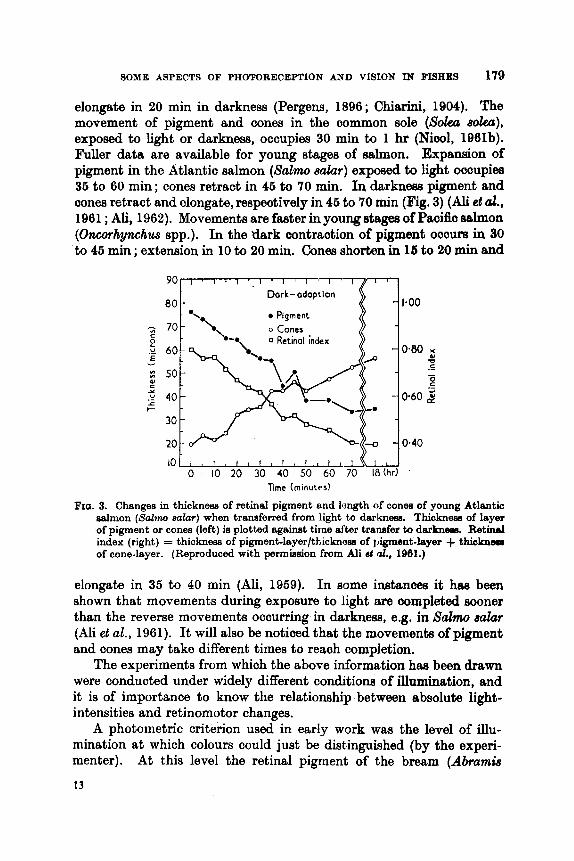
SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 179
elongate in 20 min in darkness (Pergens, 1896; C h i d , 1904). The movement of pigment and cones in the common sole ( S o h e o h ) , exposed to light or darkness, occupies 30 min to 1 hr (Niml, 1961b). Fuller data are available for young stages of salmon. Expansion of pigment in the Atlantic salmon (Salmo salar) exposed to light oocupiea 36 to 60 min ; cones retract in 46 to 70 min. In d a r h w pigment and mnes retract and olongate, respeotively in 45 to 70 rnin (Fig. 3) (Ali et d., 1961 ; Ali, 1962). Movements are faster in young atages of Paoific salmon (Oncorhynchwr spp.). In the dark contraotion of pigment ocoum in 30 to 46 min ; extension in 10 to 20 min. Cones shorten in 16 to 20 min and
Dark- adoprlon
Pigment
o Cones 0 Retinal index
20 t- N to I I 1 I l t l 3 l * i r I t i
0 10 20 30 40 SO 60 70 1 8 h ) l ime (minutes)
FIQ. 3. Changes in thickness of retinal pigment and longth of cones of young Atlantic aalmon (Sabru, a-ala7) when transferred from light to darkness. Thickneee of layer of pigment or cones (left) is plotted against time after tranefer to darknem. Retinal index (right) = thickness of pigment-layer/thicknoss of )igment-layer + thicknws of cone-layer. (Reproduced with permission from Ali st d., 1961.)
elongate in 36 to 40 min (Ali, 1959). In some instanoes it has been shown that movements during exposure to light am completed sooner than the reverse movements occurring in darkness, e.g. in S d m salar (Ali et al., 1961). It will also be noticed that the movements of pigment and cones may take different times to reach completion.
The experiments from which the above information has been drawn were conducted under widely different conditions of illumination, and it is of importance to know the relationship between absolute light- intensities and retinomotor changes.
A photometric criterion used in earIy work was the level of illu- mination at which colours could just be dist.inguished (by the experi- menter). At this level the retinal pigment of the bream (Abrumie
13
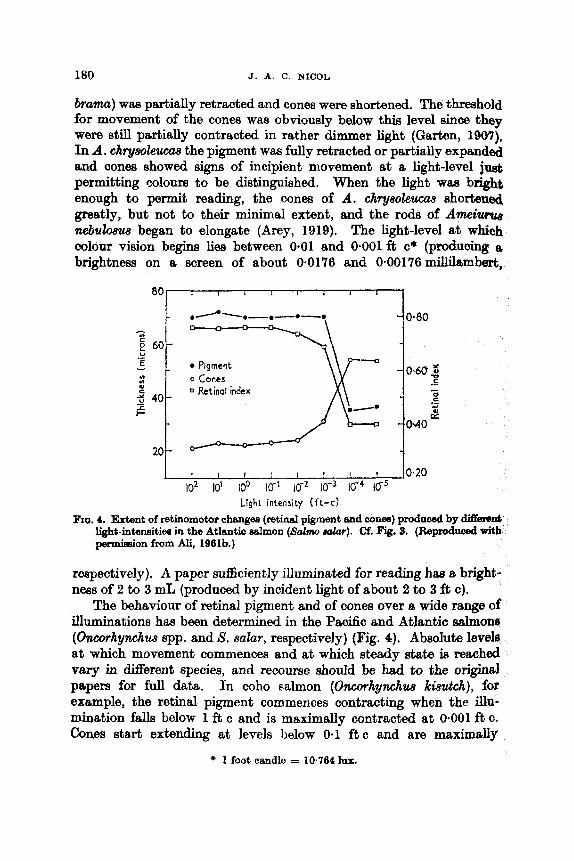
180 J. A. C. NICOL
./*--.-.--.-. -
brama) was partially retracted and cones were shortened. The threshold for movement of the cones was obviously below this level sinoe they were still partially contracted in rather dimmer light (Garten, 1907). In A. chrywleucas the pigment was fully retracted or partially expanded and cones showed signs of incipient movement a t a light-level jnst permitting colom to be distinguished. When the light wm bright enough to permit reading, the oones of A. chryaokum.9 shortened greatly, but not to their minimal extent, and the rods of Ameium nebuhus began to elongate ( k e y , 1919). The light-level at which colour vision begins lies between 0.01 and 0.001 ft c* (produohg a brightness on a screen of about 0.0176 and 0.00176millil8jmbert,
0.80
60- U .- E v -
U Y)
k 40- E t
20
Pigment 0 Cones 0 Retinal index
c d .- 2 - 0.40
- I 1 1 I I I I 1 0.20
i 0-60 2
* -1 .- I s
Light intensity ( f t -c) FIG. 4. Extent of retinomotor ohengee (retinal pigment and con-) produced by diSmr&
light-intensities in the Atlantic &on (Scrlmo &r). Cf. Fig. 3. (Reproduced with permhion from Ali, 1961b.)
reepectively). A paper su%kiently illuminated for reading has a. bright- ness of 2 to 3 mL (produced by incident light of about 2 to 3 f t c).
The behaviour of retinal pigment and of cones over a wide range of iUuminations has been determined in the Pacific and Atlantic aalmonn (Oncot.hynchus spp. and S. salar, respectively) (Fig. 4). Absolute levels at which movement commences and at which steady state is maohed vary in Merent species, end recoume should be had to the ori@ papers for full data. In coho salmon (Oncorhynchus kimtch), for example, the retinal pigment commences contracting when the illu- mination falls below 1 ft c and is maximally contracted a t 0.001 ft c. Cones sta.rt extendmg at levels below 0.1 f t c and are maximally
* 1 foot candle = 10.764 lux.

SOME ASPECTS OF PIIOTOREC~RPTION AND VISION IN FISIIES 181
extended at 0.001 f t c (Mi, 1959, 1961a, b). The threshold for the retinomotor shift to the “da rk” position drops during tho night in Lateolabrax and Cyprinus: from 0.004 to 0.001 ft c before and after midnight in Lateolabrax, and from 0-00005 to 0-000006 f t c in Cyprinw, (Tamura, 1957). There are obviously considerable differences in light- levels controlling the photomechanical changes of different fishes, and Rome degree of diurnal rhythmicity may exist (Welsh, 1938). In general it appears that pigment, cones and rods begin to adopt a position characteristic of darkness when the illumination falls below 1 ft c and positions of maximal retraction (pigment, rods) or extension (cones) are attained at levels below 0.01 f t c. This level of illumination is encountered during twilight, and in a landscape lit by the moon at phases between full and quarter. Full retreat of pigment, elongation of cones and shortening of rods are attained in dim light conditions that correspond to those associated with the switch to scotopic and rod vision in man.
Since these retinomotor changes in duplex retinae are almoet certainly conccrncd wit,h enhancing vision, it would be interesting to know at what light-intensity the change from phototopic to scotopio vision occurs in fish. The data for retinomotor changes oould then be interpreted more adequately in terms of vision. Since retinomotor changes are so extensive in fishes, one wonders if the change from cone to rod vision and vice versa depends primarily on the ability of cones and rods themselves to respond ; or if it depends rather on screening or exposure of the receptors.
Most observers have concerned themselves with only one level of illumination (and darkness), but Ali (1962) has considered the effect of different light-intensities on the extent and temporal cmrw of ratim- motor changes. He exposed yearliiigs of Atlantic salmon to two different intensities, viz. 900 and 0.1 f t c and determined the extent of move- ment of the retinal pigment and cones. In the stronger light the pigment became fully extended in 40 min and the cones fully retracted in 46 min. In the weaker light the pigment expanded, reaching a steady state in 36 min, and the corm shortened to a steady state in 66 min. The pigment was expanded to a greater degree and the cones mora retracted in the stronger light. Placed in darknem, thct pigmcnt and mnen h)ok longer to return to the I ‘ dark ” position u f lm sxpowuro to the, ntrotlg light than after expomrc to the weak light.
Possibly a fish does not often oncounter abrupt ohmgon from light to total darkness, arul it i H worthwhilo dimovering the s p d at whioh movomerit of pigrncnt and recoptom procoob undor oonditionH of changing light-intonaitios cncounturod in nature, Hay at twilight 1)olow

182 J. A. C. NICOL
1 ft c. There is an early experiment concerning Abramis brama, whioh were kept in failing daylight one afternoon until darkness, 8 period of 14 hr ; at the end of this time cones were found to be retracted (Ener and Januschke, 1905). Attempts have been made to relate the courm of retinomotor changes in salmon with changes in environmental light-levels and fish behaviour. Ali (1969) has found that retinomotor changes of juvenile Pacific salmon (various spp.) lag behind the decrease of ambient light-levels occurring in nature (from 1 to 0-002 ft 0
in 20 min): this estimate was based upon the time required for da& adaptation after exposure to light of 400 ft c. Under natural conditions; of decreasing illumination during the evening, cones and pigment of pink salmon began shifting towards the " dark " conditions at levela below 1 ft c and movement was complete between and ft c. The natural illumination fell from 1 to lo4 ft c in 45 min. In the labora- tory retinae were in the " dark " condition after exposure for 90 min to 10-1 (cones) and 10-2 ft c (pigment). It is not clear from the descrip- tions what intensities the fish experienced before the experiment began (Ali and Hoar, 1959). In Atlantic salmon yearlings, exposed to 0.1 ft 0,
the retinal pigment retracted after 45 min and the cones became maximally elongated after 25 min. Retinomotor changes towards the dark condition commence in this species when the light falls to 0.1 ft o and are complete at an illumination of 0.0001 ft c. Environmental illumination falls to this extent in about 45min (varying of course with cloud, etc.) and it might be expected from the data. presented above that retinomotor changes would keep pace with this dec.rrem (Brett and Ali, 1958 ; Ali, 1961a, b, 1962).
Photometric data for levels of illumination in the environment, and in experiments relating to retinomotor changes, have been PI%- sented as foot candles, and both sets of measurements are comparable. But other measurements relating to visual threshold, light-intensities in the sea, animal luminescence, etc., are given in energy units, ergs/ sec, watts, or photons, and some day these various kinds of infor- mation will have to be collated and integrated. In view of the un- specified composition of the lights and illumination employed end measured in the first kirds of investigation mentioned above, thia W k may prove to be very difficult.
IV. REFRACTION, ACCOMMODATION AND TEE RECEPTOB LAYEE It is generally believed that elasmobranchs are hypermetropic, and
that they accommodate for near vision by moving the lens towards the cornea through contraction of a protractor lentis muscle. Teleosta, on the other hand, are believed to be myopic, and to aocommodate for

SOME ASPECTS OF PHOTORECEPTION AND VISION I N FISHES 183
distance. The teleost eye contains a retractor lentis muscle (wmpanula HaUeri), contraction of which displaces the lens backwards along the axis of the fish’s body and more or less inwards, according to the species. Hypermetropia in elasmobranchs, according to Franz, amounts to 10 to 15 dioptres, and they can accommodate from 15 to 20 dioptres. Verrier, moreover, found that hypermetropia was also characteristic of the eyes of teleosts, even when accommodated (references in Baylor and Shaw, 1962). Hypermetropia wm detected in the eyes of a wide variety of fishes by Baylor and Shaw (1962). Some mean values for refractive error were 8 dioptres in alewife (Absa psezldoharengue) and 14.6 to 15.8 in silversides (Menidia menidia). Dogfish and skate had a hypermetropia of 6 and 8 dioptres. Accommodation in some of the teleosts they examined amounted to 6 to 10 dioptres. In A h a the refractive error in a line antero-posterior (main axis) waa lower than in lateral view (3.6 vs 8.4 dioptres). The eye of the alewife is ellipsoidal, and is slightly longer (by 10%) in the antero-posterior axis than in the minor optic axis, from lens to centre of the fundus.
A most useful treatment of vision and lens functioning in the normal fish eye has been offered by Pumphrey (1961). The lens is spherical and its dioptric strength is high, ranging, for example, from several thousand dioptres in a juvenile fish a few inches long to several hundred in an adult 1 ft long. The focal length is short, and measurements of refraction have shown that there is a constant ratio of focal length to radius having limits of 2.5 to 2.6 ; this ratio is named after Matthiessen, its discoverer. It is obvious that a refractive error of, say, 10 dioptrea in 1000 is quite small, and amounts to a displacement of focal length of only loop. Unfortunately, it is far from certain what refleoting surface is being examined in retinoscopy of the fish eye ; Beer thought that retinorrcopio reflexion occurred at the front of the retina ; Baylor and Shaw (op cil.) from behind. The retinal pigment layer itself amounts to almost loop in eyes of salmon fry, the retina inside the external limiting membrane is 200p thick, and rods, cones and pigment make considerable rrtdiaJ excursions. It still has to be demonstrated, convincingly, that the plane of focus in the eye at rest lies elsewhere than in the receptor layer; the so-called error may be one not of refraction, but rather of method.
The fish lens is virtually aplanatic and its focal length is remarkably short (2.5 x radius). It is argued that a lens having these characteristics and made of biological material whose refractive index cannot exceed 1.53, must have an index that decreases from a maximum of 1-53 at its centre to 1.33 at its periphery, where the index approximatea that ofthe surrounding medium. This change of index is difficult to measure, but it is believed to be the only explanation that fits the facts.

184 J. A. C. NIUOL
The typical teleost eye is semi-ellipsoidal, about 3 to 10% longer in its antero-posterior axis than in the vertical or oornea-fundus axis and the lens is a concentric sphere. Let it be assumed that images of far objects are in focus on the anterior and central fundus when the lens ir at rest, i.e. that these regions are emmetropic. Ths fish is then myopio for forward vision and when it accommodates by moving the lens backwards, it brings distant objects ahead into focus. Since the lens is displaced along an antero-posterior axis, emmetropia in lateral view is unaffected, but the range of dear vision behind the fish is simultaneously shortened.' The extenb to which the retractor lentis muscle is developed variea in different speoies; possibly the degree of accommodation is correlated with habits. The situation in elagmobranchs is still obaonre because both the retina and lens are slightly ellipsoidal, and the lens is protracted.
Many nektonic teleost fishes possess adipose eyelids, which lie over part or all of the eye ; when the eye is only partially covered the eyelids leave a vertical aperture over the pupil. Adipose eyelids are regarded as structures which enhance stream-lining (Walls, 1942). They are usually clear fleshy structures consisting of densely arranged parallel fibres which are oriented dorso-ventrally. Because of their structure they are strongly birefringent and act aa an analyser for polarized light. The amount of light they absorb varies with the plane of polarization of the light, and some 40% less light is transmitted when the plane of polarization is parallel to the dorso-ventral axis than when it is parallel to the antero-posterior axis. The adipose eyelids thus provide a meass by which fish can detect polarized light, at least in the peripheral field (Stewart, 1962). The possibility that this faculty may be utilized in the behaviour of fish has not escaped notice ; depending upon refkaotbn indices, the eyelih may also affect the focusing power of the eye.
A fovea is rare in fish, the outer segments of the rods and mnea are fairly long and a considerable depth of focus is consequently to be expected. Cones in the fish eye tend to be large, well spread out tmd arranged in mosaic patterns ; because of these factors the quality of an image, in focus, may be much inferior to that perceived by the human eye in air. In addition, the mattering properties of water reduce the ability to resolve images.
The structure and arrangement of cone8 in gadoid fishes have been described by Engstrt)m (1961), who haa found an " accessory element " connected with the outer segment. Intraspeoific differences have been reported in the retinae of Sebmtw wrinua, a benthic scorpion fish from deep water (Hanyu and Ali, 1962). In a specimen of S.m. menteEIa from 275 m, cones were longer than in a specimen from 549 m; rode w m ~

SOME ASPECTS OF I'IIOTORECEPTION AND VISION IN FISHES 186
larger in the fish from deeper water. Another subspecies, S.m. marinus, also from 275 m, had larger cones and smaller rods than S.m. mentelh from the same depth. These results suggest the existence of intra- specific variation of retinal structure correlated with habitat differences ; the sample as reported, however, is so small that oonfirmation is desirable ; dctails concerning size of the fish and time of catches are not reported.
V. VISUAL PIGMENTS AND SPECTRAL SENSITIVITY Scotopic visual pigments in great variety have been identified in
fishes : in general, they are of three main types. Rose-coloured pigments known as rhodopsins are the principal visual pigments in the rods of
I-
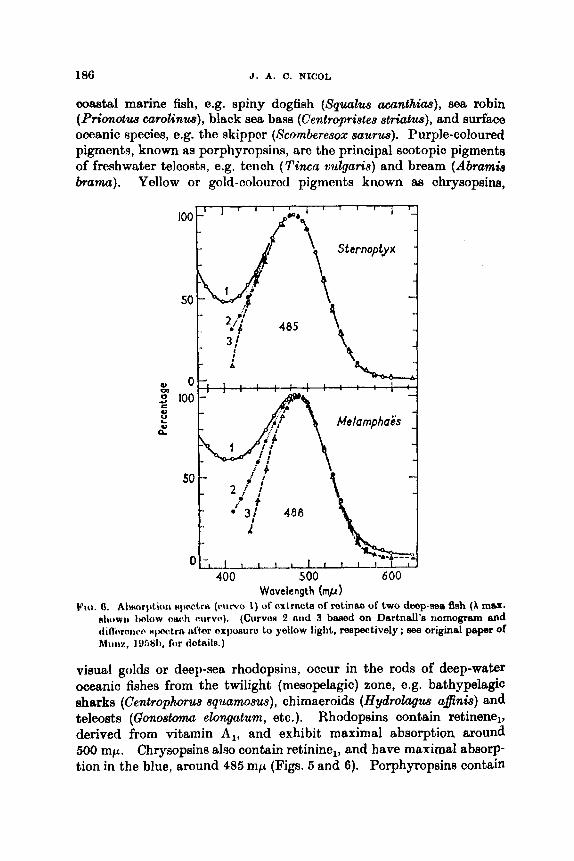
186 J. A. C. NICOL
ooastal marine fish, e.g. spiny dogfish (Sqwlus acanthias), sea robin (Prionotus carolinus), black sea base (Centropristes striatus), and surface ooeanic species, 0.g. the skipper (Scomberesox saurus). Purple-coloured pigments, known as porphyropsins, are the principal scotopic pigments of freshwater teleosts, e.g. tench (Tincu vulguris) and bream (AbramL bramu). Yellow or gold-coloured pigments known aa chrysopsins,
Wavelength (mp) k’ict. 0. A1worptio11 n p w t r e (nrwo I ) ot’nxtriuh of rotinno of two deep-see Ash (A mas.
nirowii I A o w 0w1i (wrvo). (Curves 3 nnd 3 baeed on Dnrtnall’a nomogrnm and clilTnroncir Hpotro irftnr exposuro to yellow light, mspeotively ; me original paper of Hiiilz, 1Y68b, for dOtnilR.)
visual golds or deep-sea rhodopsins, occur in the rods of deep-water oceanio fishes from the twilight (mesopelagic) zone, e.g. bathypelagk sharks (Centrophorus squamom), chimaeroids (Hydrolugus ufinis) and teleosts (Gonostoma elongatum, etc.). Rhodopsins contain retinene,, derived from vitamin A,, and exhibit maximal absorption around 500 mp. Chrysopsins also contain retinine,, and have maximal absorp- tion in the blue, around 485 mp (Figs. 5 and 6). Porphyropsins contain

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 187
mtinene,, derived from vitamin A,, and have absorption curves shifted some 30 mp towards longer wavelengths than do the rhodopsins. These distributions, although generally true, are not without exaep- tions ; the striped mullet (Mugil cephalzut), for example, is provided with a porphyropsin having maximal absorption at 620 mp (Wdd, 1939a, b, 1946; Dartnall, 1957; Denton and Warren, 1967; Munz, 1958a, b ; Denton and Shaw, 1963).
Most workers have concerned themselves with extracting the visual pigments and then measuring their absorption characteristics. Denton and co-workers (op. cit.) have also measured the absorption of the intact retina; their results give relative spectral absorption and a direct estimate of density of visual pigment in the retina.
The terms rhodopsins and porphyropsins are regarded as generic designations for pigments based on retinene, and retinene, respectively, and within each group X max exhibits 8 range of values. Maximal absorption of rhodopsins based on retinene,, including visual golds and typical rose-coloured pigments, ranges all the way from 478 to 525 mp. Dartnall and others have obtained various pigments from the eyes of pike and other freshwater fishes that have absorption maxima in the range 524 to 532 m p (Dartnall, 1957; Rushton, 1959). The change from vitamin A, to vitamin A, brings about a major shift in absorption spectra towards the red ; differences in absorption spotra within each system are determined, presumably, by the character of the protein moiety (i.e. the opsin) of the molecule. Some fishes possess two scotopic visual pigments, i.e. have a dual system. Wald (1946) has emphasized that an admixture of rhodopsin and porphyropsin is characteristic of certain euryhaline fishes, especially anadromous and katadromous fishes moving to and from fresh water. Eels (AnguiUa rostra%) in the freshwater phase and killifish (Fundulus heterocbi2ta) have pre- dominantly rhodopsin ; salmonids, predominantly porphyropsin ; and the anadromous white perch (Norone americana), exclusively por- phyropsin. A marine puffer (Sphoeroides annuhtw) has rhodopsin and porphyropsin in about equal proportions. Other dual systems found in marine teleosts involve admixtures of two rhodopsins or of two por- phyropsins. In the deep-sea teleost Batlhyylagw wesethi there are two retinene, pigments having X max at about 478 and 500 mp and occurring in proportions 4: 1 (Wald, 1939a, 1941, 1958, 1960; CresciteUi, 1968; Munz, 195813).
Changes from one environment to another carried out by oertain fish have prompted enquiries into the problem whether transformations of their visual pigments also occur at the same time. This does not appear to be so in salmon (Sulmo salur) for Denton and Sham (1963)

188 J. A. C. NIOOL
found that smolt (from sea water) and an adult wlmon that ww migrating upstream had purple-coloured retime reeembling those of freshwater fish, i.e. they retained the predominantly porphyropeia system also discovered in freshwater phases of salmonids. The eel (AnguiEla) in its immature freshwater phaae has a purple-coloured retina (dhiefly rhodopsin, vide supra). But when the eel beoornm silvery, preparatory to migrating, its retina, becomes yellow and it acquires a visual pigment closely similar to that of deep-sea f%h, with an absorption maximum shifted some 33 mp towards shorter wave- lengths (Fig. 7). This change involves the combination of mthene, with a new opsin. Conger eels pass part of their life in shallow coastal waters and return to deep water when sexually mature. The immature
I I I
L-L- 400 500 600 700
Wavelength (mp)
FIG, 7. Absorption characteristics of the retinae of immature (yellow) end mature (silver) eels. Spectral curves ehow difference in denaity between unbleached dark- adapted retinee and retinee bleached by strong white light. (Reproduced with permission from CmMe and Denton, 1969.)
eels have a yellow visual pigment and they probably possess this form of pigment throughout their life (Wald, 1958; Carlisle and Denton, 1969). Evidence for lampreys is still incomplete. In the Pacific c& lamprey Entosphnus tridentatus, both up-stream and down-stream migrants contain rhodopsin. In the sea lamprey (Petromyzon wrinua) up-stream migrants of the sea-run form contain porphyropsin, but down-stream migrants of the land-locked form contain rhodopsin. From this somewhat equivocal evidence it has been suggested that a change from rhodopsin to porphyropsin occurs in the ma lamprey aa it approaches sexual maturity (Crescitelli, 1958 ; Wald, 1958, 1960).
It is impossible t o do justice to the complexity of the subject and to the great amount of research which has been devoted to it in thi6 brief summary ; fortunately, detailed information may be obtained in the excellent reviews of Wald (1960), D a r t d (1967) and Creecitelli

SOME ASPECTS OF PHOTORECEPTION AN D VIRION I N FWIIES 189
(1958). One important idea which has come of this work and one whioh has been emphasized by some of the workers concerned, is that the visual pigments, of marine fishes at least, are adapted to the quality of light in their environment. There are considerable differences in the extent to which attenuation of different wavelengths oocurs in natural wutore. 111 const:tI wittttrs whero thero i H much suqmnded matter and dissolved yellow substance, the water is green and maximal tram- parency lies in the region of the spectrum between 500 and 600 mp, as does that of fresh waters. A set of normal transmission curves for
lo-'
I
425 450 475 500 525 Wavelength
Fro. 8. Relative spectral curves of transmitted daylight at doptha shown in the b y of (Reproduced with per. Biscay (46"29'N, 7"59'W, 10.30-11.30 h, 26 September).
mission from Boden ct al., 1960.)
different types of coastal waters, with rather flattened maxima between 500 and 570 mp, is given by Jerlov (1951). To waters with these spectral qualities, the rhodopsins and porphyropsins of inshore, estuarine and anadromous species appear to be well suited, a t least aa regar& their absorption characteristics a t wavelengths between 500 and 636 rnp (Munz, 1958b). Clear owanic water8 arc blue, and hnvo maximal transparency in thc region 400 to 800 mp. With incrsming dopth, there is preferential nhorption of Nhort m d lorig wnvnlongthH tmlow and ~ h o v o tho Iduc~-gr(w rcgioii of tho Hpcctrunl, nrid tho ronidud light which rc!aclion c l o q w wator lioH in tho rogion twoiirid 476 to 480 mp (Ng. H) (Jorlov, I9fi I ; I M o n cl i d . , I WO). 'I'huro is obviolur biological

1!)0 J . A. C. NICOL
value in the shift of absorption maxima of rhodopsins, from values around h max 500 mp in surface species to h max around 485 mp in bathypelagic species (Denton and Warren, 1957; Munz, 1958a, b). Moreover, there is a great deal of bioluminescence in deep waters, and the light emitted is generally blue, with emission peaks around 480 mp. It is nice question whether one is to consider the colour of the luminescence adapted to the spectral sensitivity of the receptor, or versa.
Deep-sea fishes have pure rod retinae, whereas duplex retinae encountered in coastal and freshwater fishes ; colour vision is known to occur among the latter. For scotopic vision, at least, the relative sensitivity to different parts of the spectrum may be reasonably inferred from the absorption curves of the visual pigments, and this may be the only approach possible for bathypelagic species that are dead oa recovery. However; when action spectra have been obtained, either from behavioural studies or electrophysiological recording, they have shown a very good fit with absorption spectra of sootopic pigments of the species in question (freshwater tench and sunfish, having por- phyropsins) (Dartnall, 1957). The hagfish (Myzine gZutinosa), (MI
deduced from behavioural experiments, shows maximal spectral sensitivity between 500 and 520 mp. It contains vitamin A, and ita photosensitive pigment presumably is a rhodopsin containing retinene, (Steven, 1955).
Spectral sensitivity curves based on electrical activity of the retinae of several species of marine and freshwater fishes have been obtained. Potential changes (known as graded photopic responses) have been recorded from some units situated between the ganglion cells and the receptors. One type of response (the luminosity response, a hyper- polarization) is evoked by white light of any wavelength, with a p k response between 500 and 600mp (various shallow water hIm&). Other (chromatic) responses are paired positive and negative polariza- tions recorded from slightly deeper levels (from the receptor layer) than the luminosity responses. These po$entiala are usually evoked by light of long and short wavelengths, respectively, providing oppoaed red- green and yeliow-blue colour mechanisms. On and off discharges corresponding to these potential ehanges have been recorded, and it has been found that the off process (generated by red light) inhibite the on process (generated by blue light). These electrical responeeg reflect indirectly the activity of receptors, presumably cones; photo- stimulation of single cones has yielded luminosity responses ; however, it has not yet been possible to link the sensitivity curves with known visual pigments (MacNichol and Svaetichin, 1968 ; Svaetichin and

SOME ASPECTS O F PHOTORECEPTION AND VISION I N FISIIES 191
MacNichol, 1968 ; Wagner et al., 1960 ; Mitarai, 1960 ; Svaetichin, et al., 1960 ; MacNichol et al., 1961).
VI. TRANSMISSION BY THE LENS The lenses of fishes are singularly freo from chromatic rtberration
(Pumyhrcy, I!)(; I ) . If the IcrisoH woro piginonted atid absorbcd certain components of the incident light, they would alter the composition of light reaching the retina. It has been discovered that those of diurnal and of surface oceanic fishes, whose eyes are exposed to some near ultra-violet light, usually contain pigments absorbing short wave- lengths (far blue and near ultra-violet, below 400 mp). In nocturnal and deep-sea species the lenses are usually transparent to short wave- lengths down to about 310 mp. Among certain benthic species of the neritic zone (Mustelus canis, Tautoga onitis, etc.), the lenses show a gradual rise in density a t wavelengths below 400 mp to 320 mp. Ultra- violet light is absorbed rapidly with increasing depth ; values for absorption of light of 375 mp range from 5%/m in clear oceanic water to 90%/m in turbid coastal water. The difference discovered in the transparencies of lenses are clearly related to the habits of the fishes and, furthermow, :ire nt1qkcd to tho dvptlis R t whioh they live (Ksn- wtly iind M i l k i t i i L i i , I!),Vl; I)rritoii, I05li: Mot,ibiM. 10G7 ; Armstrong and halch, 196 1 ; Clurko ant1 Denton, 1062).
VII. PHOTOSENSITIVITY AND VISUAL THRESHOLDS The minimal light which fishes can perceive, the minimal light in
which fishes can see objects, and the minimal light which regulates their various photoresponuivc activitieu arc frtctoru which have recoivod some attention.
Reliable values for minimal intensities of light which fishes can detect, and which govern their activities, are not easy to obtairi, and recourse has sometimes been made to estimates based on human vision. The fully dark-adapted human eye can just detect a steady light from a large field having an intensity of about 3 X pW/cm2 on the cornea; this is light of h 510 mp, which is the wavelength of maximal efficiency for scotopic vision (Pirenne, 19.56 ; Clarke and Dcnton, 1902). If the eye of a fish is a8 efficient as the human eye in absorbing t h o - green light, it can just detect daylight having a flux u t the pupil of 3 X 10-8 pW/cm2. IJnfortunately, then, &)OH not mom to tm tiny rncnmrod valuo nvailrhlu for f iho l i i t ( t tfrrcnhold in fifih. Hirioo Rome 0thr.r vortotwates niitl at least ono invertd)rato hsvo abtrolutcr throe- h lds as low UH, or lower than, that of mcw, it is highly probablc that

192 J. A. C. NIOOL
many fishes are equally sensitive. Densities of retinal pigment in the human eye are only about 0.15 and some 30% of incident blue-green light is absorbed ; densities in fish retinae are usually much greater than this, 0.6 in the skipper (Scontberesox t?auru.s), and 0.58 in the conger eel (Conger conger). The retinae of these fish absorb about 75% of the light reaching their surfaces, a gain of over twice that of the human. Indeed, in deep-sea fish such as Gonostoma and Xenodermichthys, densities reach 1.0 or more, permitting absorptions of 90% or greater (Denton, 1959). It is known that the owl, nocturnal, has an absolute threshold one tenth lower than that of man, and fishes habitually living in dim light may have thresholds as low or lower (Pirenne, 1956). Denton and Warren (1957), following their investigation of visual pigments of deep-sea fish, have discussed this problem at length. Tho scotopic pigments or visual golds of such fish absorb maximally in the blue around 485 mp, corresponding to the region of maximal transmission of clear oceanic water. After considering a number of advantages which the eyes of bathypelagic fish may possess-high pigment density, relatively large pupil, reduced “ noise ” level-they suggest that the gain in sensitivity of the eye of a deep-sea fish over that of the human may be as much as 100 times. This means that in very clear oceanic water, having an extinction coefficient of, say, 0.04, fish may be able to just detect daylight down to depths of 900 to 1000 m for a short time each day,
Recordings made with bathyphotometers in deep waters have shown that the flashes of luminous animals may be so frequent that they could conceivably provide background illumination having levels of lo-’ pW/cmZ or more (see, for example, Clarke and Huhhard, 1969).
Light is heavily absorbed by water ; trammission vrtluo~ mrrgo from around 65%/m for turbid coaHtnl waters t ( ~ 03%/m for oloar ooeariia waters. The eyo ie so eenHitive, howovcr, that in oomtal wutor8, hrtving abeorption c:ocdTioiuntH of arouiid 0. I :I, a fish at 100 IKI o m Htill porooivs objock froill H I I I ~ ~ ~ H O to H u i w o t ~ I I tho darkofh wintor dtty (region of J’lyi~~o~ith). 1 1 1 111 uoli docyo19 wtdmH, oiilinnood ~,hotoHotiRitivity should iiioroiwo LIIO t i i t l o outrli day wlioii thylight is still purcoptiblo t80 a fish.
In tho Iiumaii baing visual acuity incromes quickly as the light rises above absolute threshold. Similarly we may presume that a fish can begin to distinguish objects at some intensity only a few times brighter than absolute threshold (Clarke and Denton, 1962). In deep waters Of the ocean this means that a fish can begin to perceive objects by fd t ed daylight at depths slightly less than those when it can just detect background light. In some experiments involving rheotactic reapom of the sunfish (Lepomds) it was shown that it would just respond to

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 193
moving bars (subtending an angle of 11' at the eye) in a minimal illumination of 1 x 10-6 mL. It has been estimated that a t this brightness the equivalent of about 1 x pW/cm2 would be falling upon the eye (Clarke, 1936). Lepomis is a shallow-water fish with a duplex retina ; fishes living habitually in dimly-:it waters may be able to distinguish objects at much lower intrnsities. Thus, the mackerel continues to school at very weak light-intensities, which approach visud threshold for humans, and which are well below those at which aggre- gations of sunfish break up (Schlaifer, 1942).
Some observations on young salmon (Oncorhynchw) reveal that they cease feeding at light-intensities below lo-' to ft c (roughly IO-'to 10-6 pW/cm2) (Ali, 1959), and that they rise off the bottom and begin migrating at intensities below 0.01 ft c (approx. pW/cm2) (McDonald, 1960). The down-stream movement of young rainbow trout is influenced by much lower light-intensities (<0.0001 f t c) (Northcote, 1962). Both these responses involve visual discrimination (of food objects or objects on the bottom). Kinetic movements of trout and salmon larvae (Salmo trutta, S. irideus and S. salar) have a light thres- hold of 0.005 m c (=O-00047 f tc) (Woodhead, 1967). The results demonstrate the importance of ambient light-levels on behaviour patterns, but they do not necessarily reveal absolute threshold values. Activities at or near absolute thresholds may be important, however, only in some special environments.
There are enormow populations of luminescent animals in the oceans, hardly any in fresh waters (see, for example, Harvey, 1952.). Since the light-organs of these animals arc often tiny-indeed many of thc light-producirig animal8 themr;lc!lvos arc! quito mall- it i R pri~otical t,o coiiHidor l , l i ( ~ i i i IIH I ) o i i i t , H ~ U ~ W H , i~ii(l ho wfirnrd,o Irow flu thoy onn tw porwivctl. 'I ' l io t i u i t i ~ ~ t i vycr ( m i j u H t p(m:vivo II ~ r n a l l , Htcady ligtit-source hwviiig IL fliiu o f I . ( ! x I 0 l o pW/oniJ at tho surfttco of the cornoa (l'irotitto, I!)%; : Nic:ol, I!)T,H). Oliirko (in Clnrku nnct Donton, 1962) states that I L miall light could just bo detectrtl in clear oceanic water (trans- mission '37%/m) by a human observer at a depth of 66 m, when the intensity was 2.8 x 10-0 pW/cm2 at the eye. Pirenne's value used by Nicol obtained as it was under ideal conditions by a co-operative human observer, is probably too high for natural conditions in the tima. Clarke's figure, on the other hand, is likely to be too low because of the difficult conditions under which it was obtained (foveal vision, partial dark-adaptation, etc.). As previously considered, many fish may have thresholds as low as or lower than that of man. Moreover, in deep oceanic waters, luminescent lights are umally blue and Rhould have maximal eBciency for stimulating scotopic visual pigments of hathy-

194 J. A. C. NICOL
pelagic fishes (visual golds). Measurements made of the lights of lantern fish show that the radiant flux ranges from 0.1 x 10-9 pJ (ca. 0-1 sec) to 52 x pW/cm, at 1 m distance (Nicol, 1960; Clarke et al., 1962). In clear oceanic water i t has been estimated that another fish would see the light of a lantern fish at distances of 10 to 16 m. However, if a fish had a scnsitivity 100 x that of man, it might dctcct luminescence at 60 m or more. 1,uniincscotit flashctl are usually intermittent, and this fciLture msy crihancc thrir ability to be detected (Nieol, 1958).
Absolute thrcsholds for retinomotor changes to tho " dark ' I con- dition in the eyes of various tdeoxts lie in the range 10 to 10 -3 pW/cm, (p. 1x0). Since regulation of retinomotor responses has not been worked out satisfactorily, it is uncertain what receptors arc involved.
Experiments on behaviour of the hagfish show that it responds by general activity to illumination down to 6 m c (perhaps lower) ; this is approximately equivalent to 0.5 pW/cm2. Light levels of this order occur in coastal waters of 30 m or so where Myzine is found (Gustafson, 1935; Newth and Ross, 1955). The light is detected by generalized integumentary photoreceptors and a light level of 0.5 pWlcm2 may be a representative value for absolute thresholds of such unspecialized receptors.
VIII . THE CHORIOIDAL GLAND The chorioidal gland found in the chorioidal layer of the eyes of
most teleosts is a horseshoe-shaped structure containing a rete mirabile. Its blood supply comes from the pseudobranch via the ophthalmic or efferent pseudobranchial artery, The arterial capillaries entering the rete itltcrn:~t,c* rc~guli~rly with thr v c ~ ~ r o t i n wjillariiw 1wLviiig it , , W A I I the vmoun oi iL f l (~w pmi i t l l o t h ~ ~ t i ~ ~ r i ( ~ ~ ~ i ~ , ~ ~ i I I i i r i ~ ! 'I'lioro in I I ~ O 11. I~v~t , tJt~r i t~ ljo(iy 111 tIi(t vIiorioi(1 i i ( w 1h ftiloiforiti iirovtw. It i H Hiipp1it-d by tho retinal I I rl.c~y r i r i t l i1.s v w o i i s oit14ow nlnt) gow to the ohorioixpillnris. A ( ~ o t i i i i ~ ~ t i ftuhtro of both o1iorioidiLl gland and lentiform body is the rcyrular dt~cmuLtk)ir of rtrterinl uiid venous capillaries within them, arangenient that permits maximal exchange of material on the counter- current principle (Barnett, 1951).
Parry and Holliday (1960) found that extirpation of the pseudo- branch produced rapid darkening of the skin and, after some weeks, degeneration of the chorioid gland (in Salmo, Clupea, etc.). They sug- gested that the pseudobranch produced a hormone capable of causing expansion of ehromatophores, and that the chorioidal gland regulated the entry of this substance into the general circulation. Retia arJRociated with the swim-bladder are concerned with secretion of oxygen and it has been found that the chorioidal rote seem8 actively to secrete oxygen

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 195
into the fish eye. Direct measurements have demonstrated the existence of high oxygen tensions in the vitreous humour of many marine species ; tensions ranged from 250 to 800 mm Hg (about 1.5 to 6 times that in the blood) in benthic and pelagic forms, all of which had well-dovoloped chorioidal glands. In two fast-swimtning pclagic fishes, bluofish and remoras, tatisions up to 1320 mm Hg wcre rneasured. Fishes lacking or having but small retia had low tensions -of oxygen in the vitrcous humour. The chorioidal gland is well developed in active predatory teleosts ; possibly its function of supplying oxygen to the retina is to be correlated with visual acuity, density of receptor elements and thickness of the retina (Wittenberg and Wittenberg, 1962). It is possible that the darkening associated with pseudobranch removal is the result of depreRsed retinal activity.
IX. ECOLOGICAL AND BEHAVIOURAL STUDIES Breder (1959) has presented an extensive review, containing much
original information, of the responses of fishes to light. The following discussion is concerned especially with experiments and observations dealing with quantitative factors and the operation of photoreceptors.
Fishes living habitually in lighted environments make colour changes; the receptors for these responses are the pineal organ and paired eyes (see Section 11). Thcse responses can take place very quickly, for example in fishes living among coral reef (Longlcy and Hildebrand, 1941). Waring ( 1942) has presented an evolutionary scheme in which slow chromatic responses, mediated by hormones (in elesmobranchs), have been superseded by fast responses mediated by the nervous Rystem (in tcleosts). I':(~H were thought to occupy a t ransitional fit age, with rcd at i vcl y d o w rc>q I( IIIHCH ; r r i a l r L n c q 1 t 1 oron ww(: innervated, but the nervous Hyetem played no part in the rcspotises of intact animals. This is highly unlikely because Bateson (18904 observed long ago that white spots in the conger eel come and go very suddenly.
A related visual reaction is shown by many fishes which will swim only over a matching (or contrasting) background. Breder (1959) haa described how schools of clupeids, swimming over light sand, refused to paas over dark beds of weeds. Field observations indicate that this kind of reaction is correlated with the degree of pigmentation of the fish. Analogous behaviour is shown by various !%hes that are more repelled by vertical nets reflecting light of long wavelengths than by those reflecting short waves, i.e. they pass in increasing numbers through nets coloured as follows : red, orange rind yellow, green and bluo. The responses are governed by visibility and contraat againnt tmck
14

106 J. A. C. NICOL
ground, and colour is more important than brightness ; net colour becomes less important at twilight (Kanda and Koike, 1958a, b ; Kanda et al., 1958a, b ; Breder, 1959). Fishermen have observed that pilcha& avoid nets in " phosphorescent " waters at night; they call this phenomenon " brining ". The luminescence seemingly reveals the presence of the nets to the fish (Wilcocks, 1883).
Vision is the main factor governing the formation and maintenanm of fish shoals. In general, as shown by numerous laboratory experiments and by observations in nature, shoals break up when the illumination falls below a certain level (Imamura, 1953 ; Breder, 1959). In aquaria, the schooling of mackerel ceases in darkness and after blinding (Schlaifer, 1942). The threshold level for schooling in the Pacific sardine (Sardinops caeruleu) is well below 0.01 ft c (Loukashkin and Grant, 1959) ; in Menidia, Hepsitia, etc. schooling ceases a t 0.05 ft c (Shaw, 1961). A check was kept on cod shoals in deep water (100 m) of the Barents Sea by means of echo sounder and i t was observed that the shoals dispersed at sunset and reformed at sunrise (Ellis, 1956).
Much work has been done on the photo-responses of young salmon, especially in relation to their migratory movements. Juvenile salmon move downstream to salt water either as fry or smolts, and this movement is largely nocturnal. Various species of Pacific salmon exhibit considerable differences in their photo-responses, and Hoar (1958) has traced an evolutionary sequence among them on the bllsis of behaviour to light and other factors. Katadromous progress is associated with nocturnal activity : coho fry are active by day and by night, and their downstream progression at night is probably the r e d t of moro cliHpluactmont with than again& tho current. Pink salmon, cxemplifying tho other oxtrorne, bocomo inteneivsly aotive at night and rise to the surface ; on losing visual contact with the bottom, they swim rapidly with the current, and their exit from the river takm place rapidly. Coho, on transformation to smolt, show increased nocturnal activity and strong concealing behaviour (Hoar, 1953 ; Hoar et al., 1957; McDonald, 1960). Hoar (1958) has pointed out that tho photo-reactions of young salmon depend both upon absolute intensitiea and on rates of intensity-change; the onset of nocturnal migration coincides with rapid decline of light-intensity. The movements of juvenile rainbow trout in and out of a freshwater lake, and of brook trout to and from salt water similarly aro controlled by fall and rim of light-intensity (Smith and Saunders, 1958 ; McDonald, 1900 ; North- cote, 1962). Migration of silver eels downstream is also largely noc- turnal (Deelder, 1954).
Correlating field-observcttion wit h retinomotor changes in juvenile

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 197
pink salmon, Ali and Hoar (1959) have found that crepuscular migration ie initiated when the retina is only partially dark-adapted (by this they mean that elongation of cones and withdrawal of retinal pigment are only partially completed). The light-intensity in the evening falls rapidly (in 46 min) from 1 to 10-4 f t c while retinomotor changes and downstream movement are taking place. They have suggested that the young salmon rise toward the surface at twilight because they are only partially dark-adapted and can no longer see the bottom, i.e. the light-intensity is changing faster than the eye can adapt by retinomotor movements. Once visual contact with the bottom is lost, downstream transport follows. This interesting theory, baaed on a correlation, does not take into account the progress of dark-adaptation 8emu stricto, and there is a notable dearth of information about the movement of the rods. Indeed, the radial movements of photoreceptors and retinal pigment obviously are adaptations to high and low levels of illumination, but their exact significance in terms of visual functioning has still to be established.
Some fish are affected more by an intermittent light than by a oontinuous light at night. Paaific sardine in a dark aquarium avoid a beam of intermittent light (illumination < 1 ft c, flashing rate 60 times/ min), and they display a typical fright reaction when such a light is used (Loukashkin and Grant, 1959). An intermittent light is med in California purse-seine fishing in order to keep the fish within the seine until the net has been closed (California Co-operative Oceanic Fisheries Investigation, 1968 ; von Brandt, 1969). Juvenile silver salmon (Oncorhynchw kisutch) give fright reactions when a light is quickly turned on and off the fish (Dunkan, 1950). And in some experiment8 involving migrating young spring salmon (0. tsuwytscha), a twam of flashing light was found to be more effective in deflecting the course of the fish than a continuous light and a curtain of air bubbles (Brett and MacKinnon, 1953). On the other hand, yellow fish and little tunny, in aquaria, showed no differences in regponaes to continuous and inter- rupted lights (Tester, 1959). Special responses to flashing lights may be significant in connexion with luminous flashes of animals.
Many pelagic fishes diurnally make vertical migrations and these are probably regulated in large part by daily changes in light-intensity, as is known to be the caae in planktonic invortobrates. Vertical movo- ments from deep waters towards the surfnco r b t night, and doscent at dawn, have beon doteoted in pilohards or surdirios (Sardina pilchardw), anchovies (Engrauh8 encrmichlw) and herring (Clupea hurengw) (Dragesund, 1968 ; Cushing, 1959 ; Sara, 1962).
Of quite a different nature is the attractive influence which strong

198 J. A. 0. NICOL
artificial light exerts upon fish. This response, shown by many fishes, haa been exploited by fishermen all over the world in various kinds of " light fishing ", and a large literature has accumulated about the phenomenon (see Kristjonsson, 1959; von Brandt, 1959; Isa, 1961; Imamura, 1961; Sara, 1962). Herring, pilchards, sardine, cod and mackerei are some of the commercial fishes attracted by lamps at night. The behaviour is obviously peculiar and is not to be attributed to normal phototaxis. It is comparable to the attraction of moths to a lamp and involves some sort of visual disorientation connected with the darkness of the surrounding environment in contrast to the bright- ness of the lamp. Verheyen (19694, analysing this behaviour, believes that i t is an abnormal telotaxis in which the fish exchanges normal random movements for directed drift towards the light-source. Normal movements, in so far aa they are affected by photic stimuli, are guidod by differences in the light-intensitius falling upon the two eyes and on different parts of tho same retina. An artificial light is most offective in clear water at night when there iR no reflexion from a background that would reduce contrast. Under these conditions the light is coming mostly from the source and usually affects only one eye at a time, producing grossly abnormal stimulation. In the absence of any other modifying stimuli, that from the lamp seizes control of the lower visual centres and directs muscular activity. When there is much scattering of the light, natural moonlight, or reflexion from the bottom, mm- petitive photic stimulation of a more normal character intervenea and the effectiveness of the artificial light is reduced or nullified (Verheyen, 1956, 1958, 1959a, b).
There is accumulating evidence that behaviour is affected to aomo extent by wavelength as well as intensity. Pacific sardinot3 (8ardinopa mermlea) in an aquarium prefer blue or green light to white, and red light produces confusion and alarm in schools ; in these experiments the fish were responding to the colour of the lights (Loukashkin and Grant, 1959). 'l'eRtR nrudo with colourecl lighta on various RpooicR of marine finli in tnnkH rcrvcvrlod that tho,y woro attracted by gram and blue anti woro idifTorcrnt to rod-flyhwoid~a, Monochantus, Pegu, ctc. A n p i & wtu exccptionnl in that i t was indifferent to bluo and green and w&8 attracted by red light. These experiments took into account both relative radiant energy and radiant visual perceptibility (or spectral sensitivity) (Kawamoto, 1969; Sara, 1902). Mention has been made previously of how colour influences the effectiveness of nets. Most colours, of course, are soon eliminated by differential absorption with increasing depth, leaving monochromatic blue ur green light (@ Section V). The distinct retinal responses to lights of different wave-

SOME ASPECTS OF PHOTORECEPTION AND VISION I" FISHES 199
lengths, mentioned in Section V, may have their counterparts in overt behavioural responses.
The sensory cues and factors which influence migrating fish when moving to and from their breeding and feeding a r e a have fascinated biologists, and evidence is now accumulating that fieh can orientate on the sun and thereby follow a fixed compass direction. In one investiga- tion some white bass (Roccw chysops), a freshwater fish, were captured on their spawning grounds near shore, transported to the middle of the lake (a distance of 2.4 km) and there released. When this was done on clear days they subsequently made their way back to their particular spawning areas, but they failed to orientate and return on days that were overcast (Hasler et al., 1958). The same investigators found that sunfish (Lepomis) could be trained to enter a box in a certain compass directioii at any time of day. Both these fishca can certainly orient by the sun ; in fact, they cannot oriont themselves when it is not visible, and they must be able to compensate for movement of the sun during the course of the day. Moreover, it has been discovered by training experiments with Lepomis that they also adapt their diurnal sun- orientation rhythm to conform to seasonal changes in connexion with alterations in day length. This latter compensation, of course, is necessary to enable a fish to make use of sun-compass reactions through- out the year in the same latitude. Sun-compass reactions have becn demonstrated in young silver salmon and other freshwater fishes (Hasler, 19608 ; Schwassmann, 1960 ; Schwassmann and Braemer, 1961).
The experiments just mentioned deal with homing in local areas. Braemer (1 960) simulated longitudinal diqhcement by conditioning freshwater fish to light-cycles that were delayed or advanced from those of the normal day. When they were tested subsequently out of doors, their sun-compass reaction now deviated by a certain angle from the previous direction, and this doviation was roughly cquivnlcnt to the amount of Hhift in time or longitudo. If II f i ~ h i H dinplaced in latitude, it encounters n chrrngo iii lcrigth of t ho (lay and IL ohango in tlm inulinn- tion of tlio ~ 1 1 1 i ' t i ILW. SiiiilidI traincd to orient, to tho Hun in WiaconHin (43"N) were tlowii to Iirazil ( I "S). A t tho oqiiiitor tho eunfish oriontctl during the fortwoon hut becamo disoriented in the aftornoon, apparently because of the high position of the sun. A sunfish flown to Montevideo (30'5) continued to compensate for the azimuth curve of the sun in the way that would have been correct in the northern hemisphere. Fishes living in the tropics must reverse the direction of the sun compenaating mechanism during the course of the year, owing to the annual reversal of the position of the sun's azimuth. CichlidA (tropical and subtropical

200 J. A. 0. NIOOL
fish) apparently can do this whereas sunfish (from the northern hemis- phere) cannot (Hasler and Schwassmann, 1960).
Salmon, tuna and eels perform long sea migrations which occur at a relatively slow late and during which they may be able to adjust gradually to changing photoperiod and sun-arc. Moreover, salmon in pelagic waters are known to carry out some of their migrations at night,, when sun-compass reactions are not possible ; other guiding factma must then be operative. On approaching parent streams or spawning grounds, other sensory cues must come into play (Hasler, 1960a, b).
X. SYNOPSIS Light-sensitive areas of cyclostomes are the skin, spinal cord, brain,
pineal complex, and paired eyes when present. The hag is practically eyeless and has integumentary photoreceptors at the anterior end of the head and in the cloaca1 region. The caudal region of the ammometea is especially sensitive to light ; photoreceptors additional to the pineal complex and the paired eyes occur in the head of the lamprey.
Integumentary photoreceptors occur in blind cave fishes and may be present in some marine fishes. The pineal region of many teleosta is sensitive to light and mediates some behavioural and colour responses.
Chromatophores over the pineal region control the amount of light reaching the pineal complex. Pupillary movement takes place in aome benthic selachians and teleosts. An occlusible tapetum lucidum liea in the chorioidea of pelagic elasmobranchs and movement of pigment across the tapetum, in light or darkness, takes place slowly. Retino- motor changes-radial excursions of receptors and retinal pigment-in teleosts also occur slowly, in 30 to 60 min, and are related to change8 in light-intensity. Threshold value8 of mxwd epecies have been deter- mined.
A slight degree of hypermetropia is postulated from retinoscopy of selachian and teleost eyes, the significance of which awaits explanation. The aplanatic condition and very short focal length of the teleost lens are thought to be the result of its having a changing refractive index from the centre to the periphery. When accommodating, the lens ia displaced backwards along the antero-posterior axis and brings distent objects into focus on the posterior retina,
Rhodopsins, based on vitamin A, aldehyde, are characteristic of marine fishes. They have a great range of absorption maxima, ranging from maxima around 480 mp (visual golds or chrysopsins of deep-sea species) to purplo rhodopsins with maxima around 610 mp in fishes of turbid coastal waters. The absorption characteristics of theee various scotopic visual pigments seem to be related to the quality of light in the

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 201
environment-residual blue daylight or luminescence in clear Oceania waters, filtered blue-green or green light in inshore watera. The eel, when metamorphosing, changes purple rhodopsin for chrysopsin.
Lenses of diurnal and surface dwelling fishes, that encounter ultra- violet light, absorb short wavelengths, whereas those of nocturnal and of deep-water fishes are usually transparent to ultra-violet.
Despite great interest in photo-responses of fishes, estimates of visual thresholds are derived largely from observations on man and other animals. Fish may be able to just detect daylight having a flux of pW/cma; for a small, steady light-source the threshold may be still lower. Various factors increasing the efficiency of the eye of fishes may improve these estimates by several degrees of magnitude.
Measurements of oxygen tensions in the vitreous humour indicate that the chorioidal gland of the teleost eye is a rete mirabile, concerned with supplying oxygen to the retina.
Recent ethological and ecological studies concerned with photio stimuli have deait with colour changes, schooling, migratory movemente of katadromous salmon and trout, diurnal vertical migrations, responsea to intermittent light, coloured lights and strong steady lights, and the operation of sun-compass reactions in horizontal migrations.
XI. REFERENOES Ali, M. A. (1969). The oculftr structure, retinomotor tind photobehavioral
Ali, M. A. (1961a). Retinal histiophysiology of the yearling Atlantic salmon
Ali, M. A. (1961b). HistQphysiological etudies on the juvenilo Atlantic s a l m 0 1 1
(Sulmo eahr) retina. 11. Responses to light intensitier, wavchgthr, temperatures, and continuous light or dark. C u d . J . Zool. 39, 61 1-628.
Mi, M. A. (1962). Influence of light intensity on retinal h p t e t i o n in Atlaiitin salmon (Su lm uahr) yearlings. C a d . J. Zool. 40, 661-670.
Mi, M. A., and Hoar, W. S. (1959). Retinal responme of pink ealmon wwchtwl with its downstream migration. Nature, L d . 184, 106-107.
Mi, M. A., Stevenson, W. R., and Press, J. S. (1961). Hhtophyeiologicel rrtudiw on the juvenile Atlantic salmon (Sulmo mhr) retina. I. Rates of light- and dark-adaptation. C a d . J. Zool. 39, 123-128.
k e y , L. B. (1916). The movements in the visual cells and retinal pigment of the lower vertebrates. J. m p . Neurol. 26, 121-201.
Awy, L. B. (1919). A retinal mechanism of efficient vision. J. c q . Neurol. 30,
Annetrorlg, F. A. J., and Boalch, 0. T. (1961). The ultra-violet ebaorption of
Barnett, C. H. (1961). The struoture and function of the choroidel gland of the
responses of juvenile Pacific salmon. C a d . J. Zool. 37, 965-996.
(Salmo mlar). Praid. Coll. ZOO^. Mag. 8, 1-11.
'
343-363.
888 wator. J. Mar. biol. Ass. U.K. 41, i'iQI-GQ7.
teleostean fish. J . Anat., Lo&. 85, 113-1 19.

202 J. A. C. NICOL
Bateson, W. (1890a). Sudden colour changes in conger. J.Mar. bwl. A8.9. ux. 1, Bateson, W. (1890b). Contractility of the iris in &hea md cephalopode. J . Mar.
Bateson, W. (1890~) . The sense-organs and perceptions of fishes; with remarks on the supply of bait. J. Mar. biol. Ass. U.K. 1, 225-256.
Bayliss, 1,. E., Lythgm, R. J., and Tansley, K. (1936). Some now forms of visual purplo foiitul iri H O ~ L fishos with a nota on the visual cells of origin. I’roc. roy. SOL, U , 120, 05-113.
Baylor, E. R., arid Sliaw, E. (1962). Itofrirctive error and vkion in hhon. Sc&Mce,
Berland, B. (1961). Coppod Ornmatokoitu elongatu (Grant) in tho eyes of the Greenland shark-a possible case of mutual dependence. Nature. Lond. 191,
Boden, B. P., Kampa, E. M., and Snodgraas, J. M. (1960). Underwater daylight measurements in the Bay of Biscay. J. Mar. biol. Ass. U.K. 39, 227-238.
Braemer, W. (1960). Versuche zu der im Richtungsgehen der Fische enthaltenen Zeitschnt’zung. Verh. dtsch. zool. Ges. Jarg. 1959, 276-288.
von Brandt, A. (1959). Fishing methods in world sardine fisheries. Proc. World Sci. Meet. Biol. Sardines related epeciea (F.A.O.) 2, 663-623.
Brauer, A. (1908). Die Tiefsee-Fische. Wiss. Ergebn. “ lraldivia ”, 15, Lief. 2,II . Anat. Teil, 266 pp.
Breder, C. M., Jr. (1959). Studies on social groupings in fishes. Bull. Amer. M U S .
nut. Hist. 117, 393-482. Breder, C. M., Jr., and Rasquin, P. (1947). Comparative studies in the light
sensitivity of blind characins from a series of Mexioan caves. Bull. Amer. Mus. nut. Hist. 89, 319-352.
Breder, C. M., Jr., and Rasquin, P. (1950). A preliminary report on the role of the pineal organ in the control of pigmcnt cells and light reactions in recent toloost fishos. Science, 111, 10-12.
Brott, J. It. (1957). Tho ~ e r i ~ o orgms: tho oyo. In “ The I’IryHioloRy of PiRheR ”, ml. Jirowii, M. 15. Vitl. 2, Ih:liitvior, 1 1 1 1 , 121 164. Awrtlivtiic: I ’r iw , Ndqw York.
lhul,t, . I . It., i t1111 Ali, M. A . ( I M H ) . Srmr! o I ) H i w i b t . i t v i t . t o n 1.h ui.riivl.rim and ~ ~ ~ ~ ~ ~ l ~ o t ~ ~ ~ ! ~ ~ i ~ t ” i c : n ~ r t l H I ~ O n H l ~ S of f,tio I’iicific Hcilrrion rot ilia. ./. Finh. Re#. Bd Can. 15, 815-829.
Brett, J. Et., and MacKinnon, I). (1953). Yrcliminary expcrimenh using lights and bubbles to deflect migrating young spring salmon. J. Fish. Rer. Bd Can. 10, 548-559.
Brucke, E. (1845). Anatomischc Untermchunyon ukJC!r c1c:r sogcmnclnton‘ louchteir- den Augen beidor Wirbolthierou. Arch. Anat. I’hyeid. winw. Miid. ( J . Muller), Jahrg. 1845, 387-406.
Calif. Coop. Oceanic Fish. Invest. (1958). Effect of an intcrrrrittwit h a m of hJ4)it. Progress Rept., State Calij. Mar. Hea. Committee, 19U, fip. 8 - b .
Carlkle, D. B., and Denton, E. J. (1959). On tho rnelamrJrjhJwiw fJf t h v k u d pigments of Anguilla anguilla ( L . ) . J . Mar. b i d . Am. iJ.K. 30, 07 .102.
Chiarini, P. (1904). Cambiamenti morphologici cha si verificsrlv ricllrr rotine, dci vertebrati perazione della luce o dcll’ oscurita. Yarto 1. ;tietine dei p c i 6
degli a f i b i . Boll. Accad. med., R o w , 30, 75-110. Clarke, G. L. (1936). On the depth a t which fish can see. Ecology, 17,462-466.
214-215.
b i d . Ass. U.K. 1, 215-216.
136, 157-158.
n29-830.

SOME ASPECTS OF PHOTORECEPTION AND VISION IN FISHES 203
Clarke, U. L., Conover, R. J., David, C. N., and Nicol, J. A. C. (1902). Com- parative studies of luminescence in copepods and other pelagic marhe animals. J. Mar. biol. Aee. U.K. 42, 641-664.
Clarke, G. L., and Donton. E. J. (1962). Light and animal life. I n “ The Sea ”, ed. Hill, M. N., Vol. 1 (Physical Oceanography), pp. 456-468. Intolwcienco, London and New York.
Clarko, G. L., and Ilubbard, C. J. (1969). Quantitative rocords of tho luminescent flashing of ocoanic anirnals a t great depths. Limnol. Oceanogr. 4, 103-180.
Crescitelli, F. (1958). The natural history of visual pigments. I n “ Photobiology”. 19th Ann. Biol. Colloquium, Oregon State College, pp. 30-61.
m h i n g , D. G. (1959). Fishing gear and fish behaviour. Proc. World Sci. Meet. on Biol. Sardinee related Speciea, 3, 1307-1326.
Dartnall, H. J. A. (1957). “ The Visual Pigments ”, 216 pp. Methuen, London. Wlder , C. L. (1964). Factors affecting the migration of the silver eel in Dutch
inland waters. J. Cona. int. Explor. Mer, 20, 177-186. Denton, E. J. (1966). Recherche6 sur l’absorption de la lumi6re par le Crkt8lh
des Poissons. Bull. Imt . o c h m g r . , M m c o , 53, no. 1071, 10 pp. Denton, E. J. (1969). The contributions of the orientated photosemitive and
other molecules to the absorption of the whole retina. Proc. roy. SOC. B,
Denton, E. J., and Shew, T. I. (1963). The vkual pigments of Borne deep-sea ehmobranchs. J . Mar. biol. Ae.9. U.K. 43, 66-70.
Denton, E. J., and Warren, F. J. (1967). The photosensitive pigments in the retinae of deep-sea fish. J. Mar. biol. Aea. U.K. 36, 651-062.
Dodt, E., and Heerd, E. (1962). Mode of action of pineal nerve fibera in frogs. J. Neurophyaiol. 25, 406-429.
Dragesund, 0. (1968). Reactions of fish to artificial light with special reference to largo herring and spring herring in Norway. J. Corn. iyt. Expkw. Mer, 23,
Dunkan. R. E. (1966). Use of infra-rod radiation in the study of fiah behavior. Spec. Sci. Rept.-Fish., no. 170, U.S. Depl. Interior, Fiuh wildlife se&e,
Observations on tho shoaling br!hevioiir of cod ( U h mZlarCm) in deep water relative to daylight. 3. Mar. baol. h a . U .K . 31, 4 1 6 4 1 7 .
Engstrom, K. (1961). Cone types and cone arrangement in the retina of wme gadids. Acta rool., Stockh. 42, 227-243.
Exner, S., and Januschke, H. (1905). DM Verhslton doe Ouanintaptume vcm Abramia b r a m gegen Licht und Dunkelheit. S.B. Akad. Wiss. Wien, Math.- Naturm’es. KZ. Abt. 111, 114, 693-714.
Franz, v. (1905). Zur Anatomic, Histologie und funktionellen Gestaltung dee Selachierauges. Jena 2. Naturw. 40. 697-840.
Franz, V. (1913). Sehorgan (Chorioidea, Selachior). In “ Lehrbuch der vergl. mikr. Anat. Wirbeltiere ”, ed. Oppel, A., Teil 7, pp. 166-109. Fischer, Jene.
Franz, V. (1931). Dio Akkommodation des Selachierauges und seine Abblen. dunpapparato, riob~t IkAu-idon an dor Rotina. 2002. Jahrh, Abt. a&. dool. Phyaiol. Tiere, 19, 323-462.
von Frisch, K. (191 1 ) . UeitrBige zur PhyrJiologie dor Pigmentzollen in der Finch- haut. Pjliig. Arch. ge8. Phyeiol. 138. 310-387.
150, 78-94.
213-227.
16 PP. Ellis, G. H. (1956).

204 J. A. C. NICOL
Garten, S. (1907). Die Verhderungen der Netzhaut durch Licht. I n " Graefe- Saamisch Handb. gesam. Augenheilkunde ", 20 Aufl., Teil 1, Bd. 3 (Physiol. Optik. Anhang, Kap. l2), 260 pp.
Uilbert, P. W. (1961). The visual apparatus of sharks and its probable role in predation. Abst. Papers, Pacif. Sci. Congr. (Honolulu), 1961, 176-177.
Gilbert, P. W. (1962). The behavior of sharks. Sci. Amer. 107, 60-68. Gustsfson, C,. (1935). On the biology of Myxine glutinom L. Ark. 2001. 28A,
no. 2, 8 pp. Hanyu, I., and Ali, M. A. (1962). Intra-sub-specific variation in retinal structure
in Sebaatea marinwr mentella. Nature, Lond. 196, 554456 . Harvey, E. N. (1952). " Bioluminescence ", 649 pp. Academic Press, New York. Healer, A. D. (1960a). Homing orientation in migrating fishes. Ergebn. Biol. 23,
Hasler, A. D. (196Ob). Guideposts of migrating fishes. Science 132, 786-792. Hasler, A. D., Horrall, R. M., Wisby, W. J., and Braemer, W. (1958). Sun-
Hasler, A. D., and Schwassmann, H. 0. (1960). Sun orientation of fish at different
Hoar, W. S. (1953). Control and timing of fish migration. Biol. Rev. 28, 437-462. Hoar, W. S. (1955). Phototactic and pigmentary rosponses of sockoye salmon
J . Fish. Rea. Bd Can. 12,
Hoar, W. S. (1968). The evolution of migratory behaviour among juvonile sctlmon of the genus Oncorhynchua. J. Fish. Res. Bd Can. 15, 391-428.
Hoar, W. S., Keenleyside, M. H. A., and Goodall, R. G. (1957). Reactions of juvenile Pacific salmon to light. J. Fiuh. Res. Bd Can. 14, 816-830.
Imamura, G. (1953). Study on the disposition of 6sh towards the light. Rept. no. 1. Studies on the behaviour of Trachuruajaponicus to the light. J . Tokyo Univ. Fish. 39, 224-229.
Imamura, G. (1961). Studies on the 6sheries with lureing lights. 11. Studies on the angling for mackerel, sea bream and grunt. Bull. Jnp. Sue. 8ci. Fhh . 27, 437-439.
ha, G. (1961). A &udy on effect8 of different kindn ctf flrctrtrtg lnrcip. /lull. Jfip, soc. aci. Fish. 27, 493-500.
Jerlov, N. G. (1951). Repf. Sued. Ueep-8eu Exped. 1947-48, 3, Physics. Chem., Fasc. 1, 1-59.
Kanda, K., and Koike, A. (1958a). The study on the colour of fishing net. 1. Observations on the passage of fishes through a coloured net. Bull. Jap. Soc. aci. Fish. 23, 612-616.
Kanda, K., and Koike, A. (1958b). Tho study on the colour of fishing not. 4. A change in illumination affecting the behaviour of a Aeh school near a colourod net. Bull. Jap. SOC. sci. Fish. 23, 680-683.
Kanda, K., Koike, A., arid Ogura, M. ( 1 958a). The study on the colour of fishing net. 2. Behaviours of f%h schools in the neighbourhood of a coloured net. Bull. Jup. SOC. aci. Fish. 23, 617-620.
Kanda, K., Koike, A., and Ogura, M. (1958b). The study on the colour of hhing net. 3. Effect of the depth of colour of a net on the behaviour of a flsh school near the net. Bull. Jap. SOC. aci. Fkh. 23, 621-624.
94-1 15.
orientation and homing in fishes. Limnol. Oceanogr. 3, 353-361.
latitudes. Cold Spr. Harb. Symp. p a n t . Biol. 25, 429-441.
smolts following injury to the pinoal organ. 178-186.
Optical studies of ocoan water.

SOHE ASPEOTS OB PHOTOBEOEPTION AND VISION IN FIBHES 206
Icewarnoto, N. Y. (1969). The eignificenoe of the quality of light for the ettreotion of a. In “ Modem Fishing Oesr of the World ”, ed. Krbtjonnmn, H. (q.o.), pp. 663-566.
Kennedy, D., end Milkmsn. R. D. (1966). Sele&ive light ebeorption by the lmaw of lowem vertebretee, end ita iduence on speotrel d t i v i t y . Bid. BuU., Wood’s Hob, 111, 376.386.
Krietjonsson, H. (1969). “ Modem Fiehing Gear of the World ”, 607 pp. Fbhhg Nem (Books) Ltd., London (F.A.O.).
Kiiluw, W., end Sewall, H. (1880). Zur Phyeiologie de@ Wepithelm w- der Fksche. U n t w d . Phydd. Univ. Heidelhev, S, 221-227.
Longley’, W. H., end Hildebmnd, 8. F. (1941). SystemstiO oet810gue of the 6 h 0 of Tortugee, Florida. With obeervetiom on color, hebite, and looal dL- tribution. Pap. Tortugas Lob. 34, 331 pp.
Loukaehlnn . , A. S., end Grant, N. (1969). Behsviour end reactiom of the P.Cifi0 . eardine, Sar&twp~ cum&,ia (Girerd), under the iduenca of white end oo lod
light8 and derkneee. Pro& Cdu. A d . god. Ser. 4, 29, 609448. MoDoneld, J. (1960). The beheviour of P d 6 c &on fry during their down-
stream migration to freehwater and dtwater nuraery areee. J. Pi&. Rsr. Bd Can. 17, 656.696.
WNiohol, E. F., Jr., and Sveetichin, a. (1968). Elmtrio reepo- from the ieoleted retinae of fishee. Amcr. J . Ophthal. 46, 26-46.
YecNichol. E. F., Jr., Wolbereht, M. L., and Wagner, H. a. (1961). Electro- phyeiologid evidence for a mechenbm of color vision in the goldfbh. In “ A Sympeium on Light end Life ”, ed. McElroy, W. D. end G h , B., pp. 796816. John Hopkina Preee, Beltimore.
Mi-, C3. (1960). Determinetion of ultramicroeleotrode tip podtion in the retina in reletion to S potentid. J. gen Phyuid. 43, Suppl. no. 6, 96-99.
Moteie, R. (1967). Sur l’ebeorption de le lumih par le oristollia de quelqua poieeOne de grsnde profondeur. BuU. IW. oobanogr. Mtmim, 54, no. 1004,
Munz, F. W. (19688). The photoeeneitive retinal pigmenta of flehee from relatively turbid coaetel waters. J. pen. Physiol. 42, 446448.
Mum, F. W. (1968b). Phohmmitive pignerita from the ratiwd of d 8 h thy. ma fkhee. J. Phyeiol. 140, 220-236.
Newth, D. R.. end Rose, D. M. (1966). On the M i o n to light of M p i t w glutinom. J . czp. Biol. 32, 4-21.
Niool, J. A. C. (1968). Observetione on lumineeconce in pelagio eninmle. J. Mar. biol. Ass. U.K. 37,706772.
Nicol, J. A. C. (1960). Spectral compoeition of the light of the lantern-fth (Mgctophum puncfutum). J . Mar. Got. Aas. U.K, 3@, 27-82.
Nicol, J. A. C. (19618). The tapetum in Scyliorhinus wnicub. J. mar. bid. Am.
Nicol, J. A. C. (1961b). Photomeohanicel ohangee in the eyes of Meu. 1. Retino- motor chengee in S o h dea . J . Mar. bid. Ass. U.K. 4I,696698.
Niool, J. A. C. (1961~). Physiology of Wine orgeniems. In Report of the Council. J. Mar. biol. As#. U.K. 41, 861.
Northcote, T. Q. (1962). Migratory behsviour of juvenile rsinbow trout, Sabno @rhd , in outlet end Met atre8me of Loon Luke, Britth Columbh. J . PW. Rw. Bd Can. 19, 201-270.
4 PP.
U.K., 41, 271-277.

206 J. A. 0. NICOL
Parker, G. H. (1909). The integumentary nerves of fishes aa photoreceptors and their significance for the origin of the vertebrate eyes. Amer. J . Phyeiol.
Parry, G., and Holliday, F. G. T. (1960). An experimental analysis of the function of the pseudobranch in teleosts. J . exp. Biol. 37, 344-364.
Pergens, E. (1896). Action de la lumidre sur la r6tine. Ann. SOC. roy. Sci. d. nut. Bruxelles, 5, 33 pp.
Pirenne, M. H. (1956). Physiological mechanisms of vision and the quantum nature of light. Biol. Rev. 31, 194-214.
Pirie, A., and Simpson, D. M. (1946). Preparation of a fluorescent substance from the eye of the dogfish (Spualus acunthias). Biochem. J . 40, 14-20.
Pumphrey, R. J. (1961). Concerning vision. I n “The Cell and the Organism. Essays Presented to Sir James Gray ”, ed. Ramsay, J. A,, and Wiggles- worth, V. B., pp. 193-208. Cambridge University Press.
Rasquin, I?. (1958). Studies in the control of pigment cells and light reactions in recent teleost fishes. Bull. Amer. Mus. nat. Hiat. 115, 1-68.
Rochon-Duvignoaud, A. (1943). Les yeux et la viRion des VertBbrBs. 719 pp. Masson, Paris.
Rochon-Duvigrieaud, A. (1Bb8). L’ooil ot la vision. Zn “ Trait6 do Zoologie ”, ed. Grass6, P.-P. 13, Fasc. 2, 1099-1142. Masson, Paris.
Rushton, W. A. H. (1959). Visual pigments in man and animals and their relation to seeing. Progress in Biophysics and Biophysical Chemistry, 9, 239-283.
Sara, R. (1962). Light fishing. Stud. Rev. Uen. Fish. Camrn. Medit. no. 19, 33 pp. Scharrer, E. (1928). Die Lichtenphdlichkeit bleinder Elritzen. (Untersuchungen
iiber das Zwischenhirn der Fische. I.). 2. vergl. Physial. 7 , 1-38. Schlaifer, A. (1942). The schooling behavior of mackerel: a preliminary experi-
mental analysis. Zoologica, N . Y . 27,.75-80. Schwassmann, H. 0. (1960). Environmental cues in the orientation rhythm of
fish. Cold. Spr. Harb. Symp. quant. Biol. 25, 443-460. Schwassmann, H. O., and Braemer, W. (1961). The effect of experimentally
changed photoperiod on the sun-orientation rhythm of fish. Phyeiol. 2001.
Seliger, €1. H. (1962). Direct action of light in nnturrilly pigrrmtod rnu~clo fihros. J. yen /%g8d. 46,
Shaw, E. (1961). Minimal light intenfiity ant1 the disper8al of HChOolirig fhh. null. Znst. oce‘anogr., Monaco, no. 1213, 8 pp.
Smitli. M. W.. arid Sauders, J. W. (1958). Movements of brook t,rout (Sulvelinw Jmt; tu i lk i Mit.cMl). betwoon and within fresh and salt water. J . Fish. Iles.
Stuvon, U. M. (1056). Experimmts on tho light R o m e of tho ling (Myzine ylulinosa L.) . J . e zp . Ijiol. 32, 22-38.
Stewart, K. W. (1 982). O b ~ e r v a t i o n ~ on t.lio morphology and opticul properties of tho adiposo cyelid of fishcs. J . Fish. lies. Ud Can. 19, 1181--1162.
Svaetichin, G., Kratteninauhur, W., arid Imifor, M. (1960). l’hotostiinulation of single cones. J . gen. Physiol. 43, Suppl. no. (I, 96-99.
Svaetichin, J., and MacNichol, E. F., Jr. (1958). Retinal mechanitlins for chro- matic and achromatic vision. Ann. N . Y . A d . Sci. 74, 386-404.
Tamura, T. (1957). On the rolation txtween tho intensity of illumination and the shifting of conos in the fish rutiria. Bull. Jap. SOC. aci. E’irh. 22, 742-746.
25, 77-80.
34, 273-286.
1. Action spectrum for contraction in col iris sphirictcr. 333-342.
lltl (Jajt. 15, 1403-1449.

SOME ASPECTS OF PROTORECEPTION AND VISION IN FISHES 207
Terry, R. J. (1910). The morphology of the pineal region in teleoats. J . Morph.
Tester, A. L. (1969). Summary of experiments on the response of tuna to stimuli. I n “ Modern Fishing Gear of the World ”, ed. Krietjonason, H. (q.v.). pp.
On a method for collecting and keeping clupeids for experimental purposes, together with some remarks on fishing with light- sources and a short doscription of free cupulae of the lateral line organ on the trunk of the sardine, Clupea pilchardw Walb. Pubbl. Siaz. 2001. Napoli, 28, 226-240.
Verheyen, F. J. (1958). The mechanisms of the trapping effect of artificial light- sources upon animals. Arch. nderl. Zool. 13, 1-107.
Verheyen, F. J. (19594. Attraction of fish by the use of light. I n “ Modem Fishing Gear of the World ”, ed. Kristjonason, H. (4.v.). pp. 648-649.
Verheyen. F. J. (195913). Aquarium studies: new possibilities in sardine research. Proc. World. Sci. Meet. Biol. Sardines related Species (F.A.O.) 3, 1015-1031.
Verrier, M.-L. (1930). Contribution a 1’6tude de la vision chez les SBlacienR. Ann. Sci. nut. Zool. 13, 5-54.
Wagner, H. G., MncNichol, E. F., Jr., and Wolbarnht, M. L. (1960). The response properties of single ganglion cells in the goldfkh retina. J. gen Phyaiol. 43, Suppl. no. 0, 45-02.
Wald, G. (1939a). On the distribution of vitamins A, and A,. J. gen. Phyaiol. 22,
Wald, G . (1939b). The porphyropsin visual system. J. gen. Phyaiol. 22, 776-794. Wald, G. (1941). The visual system of euryhaline fishes. J. gen. Phyaiot. 25,
Wald, G. (1940). The chemical evolution of vision. Harvey Lect., Ser. 41, pp.
Wald, G. (1958). The significance of vertebrate metamorphosis. Science, 128,
Wald, G . (1960). Tho significance of vert~hrato rnohnlorphocrk. C‘wct&don, 21,
W ~ H . G . 1,. (1942). 7’hr C’c,rtcIir& Eye trntl itti .4(&rptiw Itdialion. ?h& No. 19,
W I L ~ I I I ~ , I I. ( I %I%). ‘1’110 c*o-circlitint i o i i of vortobdo nislanophoro responses.
\VolHh, J. H. (193s). I h n i t r l rliyt.hriis. Qtmrt. Rev. Ifid. 13, 123-129. Wilcockn. J. C. (1883).
21, 321-368.
638-642. Verheyon, F. J. (19b6).
391-415.
236-245.
1 17-1 60.
1481-1490.
916-938.
C’mdniok Inat. Sci. 7H6 pp.
I l i o l . / t i?v . 17, 1%) lh0 .
l‘ho history arid statistics of the pilchard fiRhery in England. I n “ Fish nrid Fisheries ”, ed. Herbert, D., pp. 290-303. Wm. Blackwood & Sons, Edinburgh and London.
Wittenberg, J. B., and Wittenberg, B. A. (1962). Active necretion of oxygen into the eye of a fish. Nature. Lo&. 194, 106-107.
Woodhead, P. M. J. (1957). Reactiom of nalmonid larvae to light. J. czp. B i d 34, 402416.
Young, J. Z. (1931a). On the autonomic nervous syatern of the toleostean fish Uranoacopua acaber. Quart. J . micr. Sci. 74, 491-535.
acaber. Proc. m y . SOC. B, 107, 464-485. YOUng, J. z. (1931b). The pupillary rnechariism Of tho tdlx,Rte8rI flnh ~ ~ Y / J W J H C O ~ U H

208 J. A. C. NICOL
Young, J. Z. (1933a). Comparative studies on the physiology of the iris. I.
Young, J. Z. (1933b). Comparative studies on the physiology of the iris. 11.
Young, J. Z. (1935a). The photoreceptors of lampreys. I. Light-sensitive fibres
Young, J. Z. (1935b). The photoreceptors of lampreys. 11. The functions of the
Selachians. Proc. soy. SOC. B, 112, 228-241.
Uramaopua and Lophiua. Proc. roy. Soc. B, 112, 242-249.
in the lateral line nerves. J . exp. Bwl. 12, 229-238.
pineal complex. J . ezp. Biol. 12, 264r270.

THE BIOLOGY OF CORAL REEFS
I. II. 111. Iv. V. VI. VII. VIII. Ix.
X. XI. XII. XIII.
C. M. YONCIE
University of Qlusgow, Swtlund
Introduction . . . . . . .. .. Reviews . . . . . . .. .. .. Systematics and Distribution . . .. .. Settlement of Plandm . . .. .. .. Ecology of Atolls. . . . . . .. .. Atlantic Rsefs . . .. .. .. .. Erosion . . .. . . . . . . . . Physiology . . . . . . .. .. Zooxanthellse . . . . .. .. ..
A. Nature .. .. .. .. .. B. Significance of the Association . .
Growth . . . . . . .. .. .. Effect of Light . . .. .. .. .. Productivity . . . . . . .. .. References. . .. .. .. .. ..
.. .. I .
..
..
..
..
..
.. ..
..
..
..
..
..
..
..
..
.. ..
..
..
. *
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
.. 209
.. 213
.. 214
.. 217
.. 219
.. 224
.. 229
.. 232
.. 232
.. 232
.. 236
.. 246
.. 248
. . 250
. . 265
I. INTRODUCTION Early experience of coral reefs, for over a year on Low Isles and
elsewhere on the Great Barrier Reef of Australia with a later summer at the former Carnegie Laboratory on the Dry Tortugas in the Gulf of Mexico, has left the writer with an enduring intercat in them tropical marine communities. Although rewrit r:oritrrc:hn with oord rwf4 h i v n been largely from the air with some superficial study of the fringing reef at Zanzibar, advances in knowledge have been followed with great interest. This article, therefore, represents the general impressions of one who now views coral reef investigations from some distance but also with somo realization of the gaps in knowledge which impede progress on tho biological side. Many of these demand prolonged observation or cxpcrimentation and cannot be filled until some per- manent centre for biological research, well staffed and well equipped, is established on a suitable coral formation, preferably a small atoll. There, work can be conducted not only throughout the uomons but also over a long period of years so that the broad pattern of reef changes may become known and major oontroversies eettled.
In no branch of biology are we so close to the earth sciencea m we are in the study of coral reefs. From tho standpoint of a geologist, tho
, 209

210 C. M. YONOE
position has recently been admirably expresaed by Ladd (1961) who states that “ The building of reefs is primarily a biological process, but geological processes such as erosion and sedimentation enter as soon as the first reef organism is damaged by wave action. Thereafter, reef building is a combination of organic and inorganic growth. Ultimately the effects spread to many other scientific fields.” It may be added that one effect of erosion and sedimentation is to produce new environments for life in which corals and other reef organisms have become adapted. There is no sphere in which the continual interplay between animal and environment is so well displayed or more worth while studying.
In what follows no attempt has been made to summarize all recent literature which has biological implications if only because the whole of the voluminous geological and geographical work on reefs, especially on the atolls of the Marshall Islands, has such implications. Attention has been restricted to what, in the author’s personal opinion, appear to be the more significant aspects, or implications, of recent biological studies, these ranging from taxonomy to productivity. The occasion has also been taken to point out lines along which research might profitably be directed.
By corals we are here considering those coelenterates which are epifaunistic and form massive calcareous skeletons, namely members of the Hydrozoa (notably the Nilleporina) and of the Anthozoa (the octocorallian Heliopora and Tubipora and the Scleractinia or Madre- poraria). Attention is further restricted to the hermatypic (i.e. reef- building) species which live in shallow tropical waters. As living organ- isms these cannot be considered without relation to the highly complex marine communities or coral reefs of which they are the most chrachr - istic, if not always the most numerous or even the most important, members. These reefs may be divided into fringing reefs, barrier reefs, atolls and also, as suggested by Wells (1957), patch reefs. The latter would apparently cover all but the first three categories, i.e. anything from a coral patch in an atoll lagoon to much more complex coral formations such as the low wooded islands in the lagoon channels within barrier reefs or the ring-shaped faros originally described in association with the atolls of the Maldives. While knowledge of the mode of growth of corals and of the effect on this of the forces of the physical and biological environment is essential to our understanding of coral reefs, the origin of the platforms on which reefs have grown np-and which constitutes the real “coral reef problem”-lies within the province of the geologist and the geographer and is not a matter with which a biologist is directly concerned. It will not receive more than inaidental mention here.

THE EXOLOOY OF CORAL REEFS 21 1
Historically, the scientific study of coral reefs begins with the work of D m . In a centennial survey of his work this author was im- pressed by the paucity of Darwin’s personal contacts with reefs. While on the Beagle he ‘ I had viewed atolls and barrier reefs from afar in the Pacific ; he had examined one atoll with some care in the Indian Ocean where he had also viewed the fringing reef at Mauritius. He had ueen living corals of many types and obtained some impression of their distribution in depth and also in relation to exposure on windward and leeward surfaces. And he had obtained some information, not all of it, as it eventually turned out, correct, about the submarine contours of coral reefs” (Yonge, 1958a). It was on the soundness of Darwin’s observations on the distribution in depth and on the growth in exposure and shelter of living corals (coupled, of course, with his appreciation of the possibility of widespread subsidence) that the fundamental sound- ness of his conclusions rests. Hence a modern definition of a coral reef, such as that given by Wells (1957), represents essentially no more than an amplification of Darwin’s original description.
In later nineteenth century and early twentieth century work on coral reefs, based largely on geographical surveys of reefs and sub- sequent museum description of coral skeletons, the living animal was largely overlooked. The geologist, T. Way land Vaughan, in association with A. G. Mayor, was the first to study corals in relation to their environment and largely in the then recently established Tortugas Laboratory of the Carnegie Institution of Washington. His survey of this work in his I ‘ Corals and the Formation of Coral Reefs ” (1919) may be regarded as the starting point in the modern study of living herma- typic corals. Undoubtedly, work had been hampered by lack of marine laboratories within coral reef areas. Ikttwrtm t h r Firfit, m r j H w : r w l World Wars this WaN partly made good ~ J Y t h : r:stlitili~hmcnt of& h J t c : h Laboratory at Batavia from which invcfitigations were conductcd on reefs throughout the East Indies, by the work of the Great Barrier Reef Expedition which, in 1928 to 1929, established a marine laboratory for 13 months at Low Isles, N. Queensland, and by the foundation by the Japanese in 1935 of the Palao Tropical Biological Station with a reeearch programme based largely on the results of the Great Barrier Reef Expedition and so primarily biological. Meanwhile, the Tortugaa Laboratory was opened for 3 summer months annually although work on corals diminished. Corals were little studied at the Bermuda Laboratory, admittedly at the northern limit of coral growth although, despite similar limitations, at Hawaii much valuable work came from the largely unaided efforts of C. H. Edmondson.
In this author’s account of “ The Biology of Coral Reefs ” which 15

212 C. Y. YONOlD
forms the final paper in Volume I of the Reports of the Great Basrim Reef Expedition (Yonge, 1940), the work of these laboratories waa reviewed. Only the two last, and least important, survived the war. The Tortugaa Laboratory was closed in 1940, the Dutch and Japan- lost not only their laboratories but the territories on which they were situated.
Since the war new activities have appeared. In the Atlantic the Lerner Laboratory at Bimini in the Bahamas and the Marine Labora- tory of the University of Miami have replaced the Tortugas Laboratory, although no great attention has been paid to corals. The major activities in this field have come from the highly important work of Norman D. Newell and co-workers on the form and nature of West Indian and Bahamian reefs and that of T. F. Goreau of the University College of the West Indies, Jamaica, on the physiology and ecology of corals. His researches, with those coming from the Haskins Laboratory, New York, on zooxanthellae, undoubtedly represent the major reoent contributions to our knowledge of living corals. And it should here be noted that a distinction must be drawn between the neede of an individual coral colony and those of the community (even if considered to be no more than the sum of all coral colonies) of which i t forms a Part-
Activities in the Pacific have been almost entirely American. They have been carried out by the United States government and been largely concentrated on the Marshall and Caroline Islands which became U.S. Trust Territory after the last war. There was immediate military need for information following a war fought in coral s e a when too little waa known about reefs and especially about atolls of which these islands largely consist. These investigations were hteaei6ied before and after the nuclear bomb tests at Bikini and adjacent atolls in the Marshall Islands. A massive series of reports, primarily geologial and oceanographical but with many important biological data, have appeared, together with the Atoll Research Bulletins produced by the Pacific Science Board of the National Research Council and National Academy of Sciences. Papers in these Bulletins have covered 0V6V aspect of life on atolls, from anthropology to the ecology and systematics of the terrestrial and marine fauna and flora. Fortunately much of this work has recently been brought together in " Atoll Environment and Ecology " by Harold J. Wiens (1962).
Other major activities in the Pacific have included the important expedition to Raroia (Kon Tiki) Atoll in the Tuamotu Archipelrtgo in the south-west Pacific, led by Norman D. Newell which, although primarily geological, provided most valuable information about reef

THE BIOLOGY OF CORAL REEFS 213
formations and ecology in the south Pacific and enabled first-hand comparisons to be made with Atlantic reefs. It is personally pleasant to note the continued activity of the Great Barrier Reef Committee at Brisbane which has established a small marine station at Heron Island in the Capricorn Group. Further observations have also been made at Low Isles, the most intensely studied coral formation in the world.
In addition, a, variety of papers dealing with some aspect of the biology of corals and coral reefs have come from varied sources such aa the Marine Biological Station at Ghardaqa (Red Sea), from various expeditions and above all from the notable series of systematic reporta on coral collections mentioned later.
It is far from easy to deal adequately with the maas of information represented by these investigations in the major tropical oceans and pursued from such very varying standpoints. The two major themes are undoubtedly the intensive work on atolls in the Pacific, primarily geological and oceanographical but dealing with formations that owe their existence to the activities of living organisms, and the more intimate researches into the physiology of corals and their symbiotic zooxanthellae being carried out by T. F. Goreau at Jamaica. Some link between the two is provided by the attempts at evaluating productivity on Pacific reefs, notably by Odum and Odum (1955).
It would, however, appear best to deal with the general before the particular, i.e. to discuss reviews of coral reef problems and systematic work (including development) and then deal with the post-war work on Pacific atolls (including ecological findings). The comparative work of Newel1 gives a link with Atlantic reefs. On the other hand, the various estimates of productivity carried out on Pacific reefs aro moat suitahly diwussed nftor considcratioti of modern work on zooxanthellae coming from the Haskins Laboratory and of Goreau’s investigations at Jamaica. These represent the present growing point in research on the individual coral on which, in the last resort, all else depends. It is gratifying to know that Dr. Goreau will himself be dealing with this in more authoritative detail in a forthcoming review.
11. REVIEWS The great revival of interest in coral reefs is indicated by the number
of review articles, all by geologists or palaeontologists writing from first-hand knowledge of reefs, which have appeared relatively recently. Ladd and Tracey (1949) give an authoritative sketch of post-Darwinian theories concerning coral reef formation with ~uggestiona for the mont promising lines of future work. Ladd (19.50) givm a aurvey account of

214 C. M. YONOE
recent reefs with a description of borings up to that on Bikini (the deeper one at Eniwetok which struck the basalt foundations was made in 1962 (Ladd et at., 1953)) ending with the generalization that “ the more thet is discovered about the geological history of any reef or reef-enuiruld island, the more complicated its history appears.” Very recently (1961) he has published an admirable general survey of uoral reefs. But his only direct reference to living corals concern growth with a mention of organic productivity. The more popular, but no less authoritafive, paper by Newell (1959) again outlines the history of the cord reef controversy but contains an excellent comparison of Pacifiu and Atlantic reefs (to be discussed later) with a general account of the biology of coral reefs.
Umbgrove (1947), another geologist with exuept iody wide experience of coral reefs, this time in the East Indim, reviewed the results of 15 years of work before 1940 on reefs widely scattered through that region. Although he covered the work of other Dutch investigators, notably Kuenen, Verwey and Boschma (the last two being zoologistS) the main biological implications of his review concern the environmmta,l forces which influence coral growth.
There is finally the invaluable account of coral reefs by W e b (1957) in the first volume of the “ Treatise on Marine Ecology and Paleo- ecology ” and which, in relatively short space, gives the most com- prehensive general account of coral reefs, in all their varied asp&, known to this author. He begins by noting that coral reefs “ are scat- tered over an area of 190,000,000 square kilometres (68,000,000 square miles) wherever a suitable substratum lies within the lighted waters of the tropics beyond the iduence of continental sediments, and away from the cool upwellings of the aea in the eaetern parts of the mane’ basins.” He refers later to “ The coral-reef biotope ” &B ‘‘ a fa&a of the marine tropical biochore ”, with its “ essential fauna and flom ” &B
consisting “ of corals and calcareous algae whioh dominate in numbers and volume and provide the ecological niches essential to the existenCte of all other reef-dwelling animals and plants.” It follows from this that “ The existence and potentialities of reefs are largely uonditioned by the ecological requirements of hermatypic corals and calcareous algae.” Here we have Darwin’s original definition amplified and brought up fo date.
111. SYSTEMATICS AND DISTRJBUTXON Passing to works of systematic importance there come fht the
“ Rsvision of the Suborders, Families, and Genera of the Scleractinia ” by Vaughan and Wells (1943) and the comprehensive account of the

THE BIOLOGY OF CORAL REEFS 215
Sckractinia (Wells, 1956) in the “ Treatise on Invertebrate Paleon- tology”. These deal with structure (notably of the skeleton), physiology, ecology and distribution of corals. Both vertically and horizontally, starting from shallow mid-tropical waters, hermatypic corals diminish in numbers, giving place eventually to ahermatypic species. The probable course of evolution, in terms of skeletal elaboration, is out- lined, but the difference in the sequence of development of organs and skeletal parts noted by Atoda (1953) may also be relevant here. It would also be illuminating if skeletal changes could be related to greater efficiency and wider powers of adaptation. Modern corals are highly adapted for life in the many different environments present on modern reefs and also for capturing zooplankton of all sizes. The form of the skeleton plays its part in these adaptations.
Knowledge of the coral fauna of the Eastern Pacific, from the Gulf of California, the Galapagos and other offshore islands, has been greatly extended by the work of Durham (1947, 1962), Durham and Barnard (1952), Hertlein and Emerson (1957), Squires (1959) and Durham and Allison (1960). Although apparently somewhat modified by later work, Durham and Barnsrd list a coral fauna of ninety-eight species rep- resenting thirty-nine genera. Although twenty-seven of these species are hermatypic, conditions, largely of temperature, prevent the for- mation of reefs. In a very full description of the recent (and also Pleistocene and Pliocene) coral fauna of the Gulf of California, Squires (1959) notes the gradual impoverishment going north. A southern fauna consisting of species of Pavona, Pocillopora and Porites loses first the species of Pavona and then of Pocillopora and finally those of Poritee, which give place to kelp in the northernmost and coldest waters of the Gulf. Squires also notes that tho coral fauna of the ewitern Pacific has little affinity with that of the WeHt Indian region, being essentially Indo-Pacific. The effect of Ekmau’s Eastern Pacific Barrier is negligible, a t any rate for scleractinians, the more hardy of which are distributed over the entire width of the Pacific. The major recent contributions to coral systematics by Bowhma, one of tho most distinguished of modern workers on corals, are a scries of papers (1948a, b, 1949, 1950, 1951) on the Hydrocorallia (not a xystematic unit but a convenient term covering the very different Milleporina and Stylaaterina formerly grouped togethcr in thc Hydrocorallinae). AH discussed below, he has made a major coiltrihution to tho HpecieH problem in Millepora, deciding that infitead of the one fipecieH nuggf!~t,cd by Hickson, there are at least ten.
Major collections of corals from two of the richest area8 in the world, the Great Barrier Reef of Australia and the Marfihall WartdrJ, hnvc! heen

216 C. M . YONOE
reported on by Crossland (1952) and Stephenson and Wells (1966) and by Wells (1954) respectively. These reporta are the more valuable because of the wide field experience of the authors. Crossland spent much of his life studying first Indian Ocean and then Pacific corala and coral reefs. The collections of the Great Barrier Reef Expedition, which could cover only small sections of the 1250 miles of the Barrier aeries, consisted, Crossland found, of 174 species belonging to 54 genera and of these he states “ 30 species are new, 2 of Barnard’s have been given new names, and there are several curious varieties, 3 of which have been given names.” Later, in the course of a survey of the coral fauna of Low Isles still affected by the cyclone of 1960, Stephenson and Wells (1956) estimated a possible 150 species. Wells (1955) haa further shown how the number of coral species diminishes from north to south along the BaTier. No new species appear ; hermatypic corals are always most abundant in the mid-tropics.
Regarding the Marshall Islands, Wells (1954) statea that the coral fauna “ includes 240 speoies and varieties (22 new) representing 62 genera (1 new) of hermatypic scleractinian corals, 15 species and varieties (3 new) representing 10 genera (1 new) of ahermatypio forme, and 11 species representing 6 genera of nonscleractinian corals.” We& had personal knowledge of the Marshall Islands and supplies important information, summarized later, about the distribution of comh on the seaward and lagoon reefs. He also discusses the wider question of the general distribution of Indo-Pacific corals.
Wells (1950) and Searle (1956) have described the coral fauna, of Cocoa-Keeling atoll and of Malaya and gradually the major task of describing the Indo-Pacific fauna is being accomplished. It is queetion- able whether there will again be papers equal in length to those of Crossland (1952) and Wells (1964) although major taxonomic problems remain. Knowledge about Atlantic corals is probably less complete ; iq a personal communication, Dr. T. F. Goreau states that publication of recent work in the West Indies is held up because of taxonomic difliculties.
The major difficulty persists of deciding the limits of variability within sessile animals forming large colonies, the growth form of which is inevitably-but also differentially- influenced by weter movemenfa. The tendency of the nineteenth century museum taxonomist studying coral skeletons was to multiply Bpecies. The lilter reaction of the field worker was to go to the other extreme and convert these museum speoies into growth forms. Thus Milleporcc waa reduced by Hickmn fa the single species, M . alciwrnis Linnams. More recently taxonomy hae been largely in the hands of those who are also field workers and with

THE BIOLOGY OF COB& REEF8 217
interesting results. Thus Wells (1954) reduoed five species of POT&% to one, P. lichen. On the other hand, Boschma (1948b, 1949, 1950) reversed Hickson’s ruling by re-establishing, on the basis of the form of the skeleton and of the ampullae, eight formerly deacribed species of Hillepora and adding a new species, making ten in all. Crossland (1962) added yet another species.
Boschmi~’8 explanation (1948a) of the diversity of forms in Milkpora -it may well serve for many other genera of hermatypic corals- involving both species and growth forms deserves to be quoted. ‘‘ There are a number of species of Hilkpora, each of which is so strongly variable that under the influence of external conditions it may w u m e a form which is more or less typical for another speciee. If a Nillepora larva happens to become fixed on a spot where the conditions of exis- tence are ideal for the species, it grows out to a colony of the typical form. If it happens to become fixed on a spot where the conditions of existence for this particular species are unfavourable it may grow out to a colony with a growth form quite different from that of the typical form. If it happens to become fixed on a spot where the conditions of existence are altogether unsuitable for this particular species (although other species of Millepora may find here ideal conditions for a luxuriant development) the young colony dies. It is not to be denied that certain species which have a pronounced preference for distinct parts of the reefs in these parts only develop into colonies of the typical shape. A good example forms M . platyphyUa that attains ite most vigorous growth (the “ honeycomb facies ”) on the surf-swept edge of the reefs only, and changes into a leafy form when living in the quieter water of the lagoon.”
As formerly stressed (Yonge, 1940), the great mccms of the Scleractinia may well be due to the presence of speciee highly adapted for a particular habitat dong with others capable of modification to produce a variety of growth forms capable of life in a variety of environments. The relation between form and the two factors of light and water movement is shown in the work by Abel(l969) on the aher- matypic Mediterranean coral, Claducera cespito8a. SimiIar observations could very suitably be made on tropical reef-builders.
IV. SETTLEMENT OF F’LANULAE Final distinction between growth forms and species can only be
achieved by observations in the field such as were carried out in the course of the Great Barrier Reef Expedition when marked cords were moved from one environment to another or colonies were split and the

218 C. M. YONOE
two halves then exposed to different environments. A long-term pro- gramme of such experiments would be of great taxonomic value. Further possibilities are presented by the fusion of recently settled planulae which then give rise to colonies having the same form aa those which arise from a single planula. Fusion of planulae has been observed in species of Siderastrea, Pocillopora and Porites and also in Manioina areolata. Identically shaped oval colonies of the last of them may consist of one, two or three ‘‘ valley ” systems indicating formation from one to three original planulae (Yonge, 1935a). There irs here, it seems, a means of determining the effects of genetic constitution and of the environment. If planulae of known origin could be induced to settle together (and although in many planulation does occur at different phases of the moon there appears to be considerable overlap (me Atoda, 1947a, b, 1951 b, c)) successful fusion would indicate origin from different growth forms of the same species, failure to fuse origin from different species. It is, of course, possible that composite colonies might be produced, particularly if allied species with different breeding seasons could be induced to breed at the same time. This would be of no leas interest, especially if the growth of the composite colony could be compared with that of each parental species.
Factors influencing the settlement of planulae have never been adequately analysed. Working on species of Pocillupora, BeriatopOra, Acrhelia and Euphyllia, Kawaguti (1941b) found that within a certain range of light intensity, progressively lower in the order listed, all were positively phototactic, the reaction reversing in higher light intensities. All were negatively geotactic. Distribution of the adult colonies bore some relation to the reactions of their plariulae. These in turn appeared to be correlated with the concentration of zooxanthellm. Edmondson (1946) tested the reaction of plmulae of Pocillupora &micorn&, C y p h t r e a ocellini and the ahermatypic Dendrophyllda man&. Hie results indicate the greater abundance in surface waters of planulae during darkness or subdued light, i.e. a major effect of negative geotaxk. Like Kawaguti, he noted positive thigmotaxis. He observed reactions to temperature and salinity changes but more significantly noted that while the larvae of Cyphastrea and Dendrophyllia would both settle in total darkness, the yoimg polyps of the latter (without zooxanthellae) survived for up to seven months, whereas those of the former soon died. Planulae of these two species, but not of P . damicornis, tended to settle in aggregations.
In a series of papers (1947a, b, 1951a, b, c, 1953), Atoda has described planulation, planulae, settlement and early development in P m i w a damicornis cespitosa, Stylophora pistillata, Acropma brtiggemanni,

THE BIOLOGY OF CORAL REEFS 219
Galaxea aspera and Seriatopora hystrix and also (1951d) asexual reproduction in the Seriatoporidae. Planulation occurs throughout the year and in all except Acropora brwgemanni in relation to some phase of the moon (a condition already known in some other corals). Although usually settling quickly, planulae can remain swimming for a con- siderable period. He confirmed and extended the observations of Edmondson that darkness delays settlement and found that it also retards subsequent development. With suitable facilities such studies could be greatly extended.
Factors governing settlement, including the nature of the sub- stratum and aggregation, are of prime importance for sedentary animals. They have been extensively studied in temperate waters, notably in connexion with acorn barnacles, with Spirorbis and with species of Ostrea and Crassostrea. In corals there is the added problem of the effect of aggregation on the formation of compound colonies. There is also the later loss of attachment in certain genera and species, in the Indo-Pacific fungids and in the Atlantic maeandrine Hanicina areolata, and in the ahermatypic Turbinoliinae which are adapted for life on an unstable substratum which, as noted by Vaughan and Wells (1943), gives little surface for settlement of the planulae.
V. ECOLOQY OF AroLLs
The major contribution to the classic coral reef problem since World War I1 has come from the extensive United States investigations in the Marshall and Caroline Islands. Many atolls, notably Bikini and Eniwetok, have been examined in grc?rA c,c:c?ariop;raphioal, gwhgioal and geographical detail. The major h ~ J l f J ~ i f : d wrm kri6vc l w m dt:dy defined. A comprehensive account of all tqect8 of thiR, extending of course to land flora and fauna and every aqmt of human habitation, has recently been made by Wiens (1962) who has himself made notable contributions in the geographical field. These investigations may be said to have culminated in the final boring on Eniwetok where basalt waa encountered at 4630 ft, the deepest coral being of late Eocene formation (Ladd et al., 1953). The atoll is revealed as resting on a truncated volcano possibly similar in nature to the numerous sea mounts or guyots present in thc north-west Pacific (Hamilton, 1956). Certainly to the east of the andesitc line which represente the wcHtcrn limit of tho Pacific basin, Darwin’s view8 appear triumphtlntly juRtificd. To the west of the line where thcrc i H uxterwive clevation, aH at ‘I’orqp rmd Fiji, the coursc of rccf formution must have hocn different.
The initial rcsult of thcsc survoys has boon to confirm and oxtond

220 C . M. YONGE
previous observations on the influence of trade winds on the con- figuration of reefs (see Yonge, 1951). The often striking differences between windward and leeward reefs were clearly demonstrated by the Dutch surveys in East Indian waters which have been summarized by Umbgrove (1947). In more intimate detail, a t Low Isles and elsewhere, they are described in the reports of the Great Barrier Reef Expedition. The wide diversity of ecological niches on coral reefs has been revealed. This is even greater on atolls where in addition to exposed and sheltered outer shores (NE and SE and SW and NW respectively north and south of the equator) there is the enclosed lagoon with its exposed and shel- 'tered shores (here SW and NW and NE and SE respectively). The consequent further increased environmental complexity influences the distribution of corals and other reef organisms. There can be no region of comparable size in the sea with such a diversity of ecological niches aa an atoll.
The first stage in the analysis of such conditions must be descriptive. The geological background to Bikini and adjacent atolls has been provided by Tracey et at. (1948), Ladd et al. (1953) and Emery et al. (1954) while Munk and Sargent (1954) have described the interactions between the growth of the living reef surface and the force of the waves, The major biological survey is by Wells (1954) and can be only briefly summarized.
Viewed in section, as shown in Fig. 1, these Pacific atolls may be divided into the following regions (Tracey et al., 1955) starting from the windward. (1) The outer seaward slope with a pronounced reduction in corals and coralline algae below about 10 fathoms. (2) A rshallow '' sublittoral " region with more gradual slope, rich coral population and often locally terraced. (3) The seaward reef margin with its dgd ridge of Pwolithun (= Lithothamnion) and characteristic surge channels (in region indicated by x). (4, 5) An outer and an inner reef flat with characteristic zones of coral population described below. (6) The seaward beach of the island, usually buttressed with beach rock. (7) The more sheltered lagoon beach. (8) The shallow lagoon reef flat or lagoon shelf. (9) The lagoon reef margin. (10) The sheltered lagoon slope. (11) The lagoon floor. (12) Coral knolls or pinnacles (patch reefs) that rise in great numbers from the flat lagoon floor which is itself oharac- tenstically covered with the green calcareous weed Halimeda. Intense echo sounding has revealed the great abundance. of these coral r n m , over 2000 in the 24-mile-long lagoon at Eniwetok (Emery, 1948). (13) The more irregular and richer expoaed lagoon slop leading to the lagoon reef margin (14). (16) The reef flat on the leeward side. (16) The seaward reef margin here without an algal ridge and surge

16 4 c l A . . . . . . I .? . . . . . , . . . .
Fro. 1 . Significant features of an atoll shown in section. Arrow indicates direction of prevailing trade winds; x region of algal ridge; dotted lines tidal range. For explanation of numbers, see text. (After Treoey el al., 1955.)
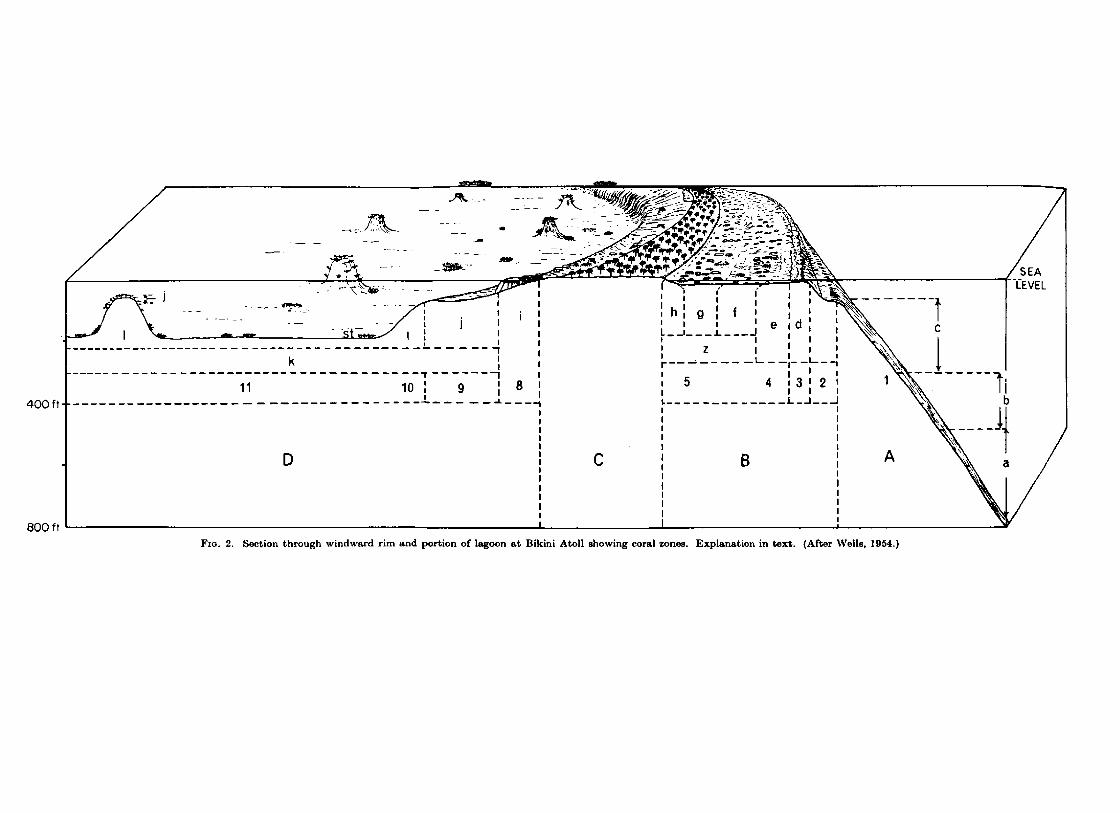
C
FIG. 2. Section through windward rim and portion of lagoon at Bikini Atoll ahowing coral zones. Explanation in text. (After Wells, 1954.)

THE BIOLOOY OB CORAL REEF8 221
channels. (17) The reef slope with coral growth notably decreasing at about 10 fathoms but without the local terracing of the correspond- ing windward depths.
The further analysis of the more windward environments in terms of the coral species most chraoteristic of the different zones on Bikini ie supplied by W e b (1954) and is illustrated in Fig. 2. With numbers indimtkg the same zones aa in the preceding figure, the major areas are : A (=l), the seaward slope ; B, the seaward reef; C, the island ; D, the lagoon. Wells distinguished the following zones (indicated by the corresponding lettering in Fig. 2) in terms of their dominant corals. (a) Sclerhelia-DendrophyUia zone of ahermatypic oorals below 80 fathoms. (b) Leptmeris zone (between 50 and 80 fathoms) " marked by special species or a facies which differs from those of the surface reef forms but is not truly ahermatypic ". (0) Echimphylliu zone from 10 to 50 fathoms and the lowest to which " surface reef species extend and grow with any degree of strength". Then on the upper surface of the reef in regions of decreasing exposure come (d) the Acropora: cuneuta zone, (e) the Acropora digitifera zone, (f) the Acrqma palifera zone, (g) a highly characteristic Heliopora zone with large colonies of this blue octocorallian, and (h) the Porites lutea zone. The three laat occupy the greater part of the just-submerged reef flat (449, a region which Wells describes as that of microatolls (z). Their formation is the result of limited possibilities of upward growth causing the colonies, killed off at their summits, to enlarge peripherally around a dead and often partly excavated centre. The sheltered margins of the lagoon are characterized by (if a Porites udrewsi zone and then (j) an Acropwu reticukda zone found both on the lagoon reef flat (9) and at c o r n - ponding shallow depths near the summits of the coral patches. This overlaps with (k) an Acrqorufomzosa zone which extends generally over the lagoon floor and terr9ce (1) an Acropru ruyneri subzone entirely confined to the floor.
The highly significant environmental differences between the reef margins on the windward and leeward sides of reefs, already noted elsewhere, e.g. in East Indian reefs (Umbgrove, 1947) and at Low Isles and reefs of the Great Barrier series, have been described in great detail for these atolls, notably by Tracey et al. (1948), Mu& and Sargent (1964) and Wells (1954, 1957). The windward margin, fully exposed to the oceanic surf, invariably possesses a mmive algal ridge composed largely of Porolithon (= Lithothumnh) which extends sea- ward to a depth of up to 10 fathoms in the form of a series of buttresses or spurs with deep intervening grooves or surge channels (Fig. 3). Especially in regions of greatest exposure the upper end of these
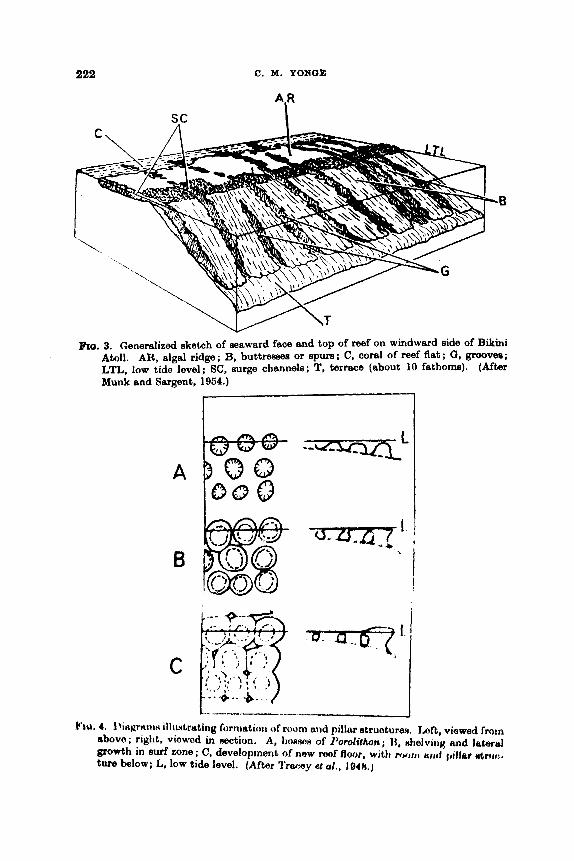
222 C. M. YONQE
sc "P
Fro. 3. Generalized sketch of seaward face and top of reef on windward side of Bikini Atoll. AR, algal ridge; B. buttrewe or spurs; C, coral of reef flat; 0. groovee; LTL. low tide level; SC, surge channele; T, terrace (about 10 fathom). (After Munk and Sargent, 1954.)
A
B
C
ho, 4. Jhgrtt iw illiiN1rutiny fornratiorr of room and pillar structures. raft, viewed froin abovo; right, viowod in section. A, ~JOSSON of I'ootolithon; 1%. shelving and labrul growth in surf zone ; C, development of now reof floor, with r o o r t l rrcd I,illar wfr111:.
ture below; L, low tide level. (After Trsr:ey et ul., 1948.)

TEE BIOLOGY OF CYORAL REEFS 223
channels may become overarohed by the upward growth of mushroom- topped masses of calcareous algae which then fuse with one another leaving channels below. Wahr entering along the surge channels is violently expelled through a series of blow holes on the reef surf-. Details of these “ room-and-pillar ” structures are given in Fig. 4. As described in detail by Munk and Saqent (1964), the effect of the surge channel8 is to counter the force of entering sew. They are, aa shown very clearly at Bikini, confined to the windward surfaoes of the reef and play a clearly essential role in the maintenance of the reef mass against the f d force of the oceanic surf.
-- - -- FIG. 6. Leeward reef edge at Rongelep Atoll showing lerge dd re-tmfrsnta and
almost vertioal descent to depth of more than 30 fathoms. (After ‘Jhxiy eL d., 1948.)
Marginal reefs on the leeward aide, e.g. on the western side of Bikini, show evidence of some mechanical erosion due to summer change in winds. There is here a steeper seaward slope while embayments, often with contained boulders, are formed by collapse of portions of the reef margin (Fig. 6). This is essentially similar (on a far larger scale, admittedly) to conditions in the lee of Low Isles. Coral grows there with great profusion and usually under no constraint from the force of wind and weather. It is therefore in no condition to withatcrad the force of occaaional summer storms of cyclonic force when greet II~&BB~B of coral are dislodged and thrown on to the surface of the reef fiat. There it forms a conspicuous “ boulder zone ” but on the leeward, not

224 0. M. YONQE
the windward, side of the reef flat. While conditions on the lee of an atoll, which cannot provide the same degree of shelter, are much more stable, this evidence of erosion does indicate the greater effect of s t o m on this usually more protected side of the reef.
There is general confirmation of these over-all findings in a variety of other surveys, e.g. in the Caroline Islands, by Tracey et al. (1961) working on Ifaluk Atoll, in the Tuamotu Archipelago of the South Pacific, following theimportant survey by Newell (1956) and co-workers* and, although here modified to a greater extent, in Caribbean reef formations off British Honduras (Stoddart, 1962a).
Detailed studies of particular areas, let alone work on the autecology of important species of corals-such as the study of Uonbtreu myera at Palao carried out by Motoda (1940b)-have hardly yet been attempted. Again, reference should be made to the relevant chapters in Wiem (1962) and in particular to the survey of Raroia (Newell, 1956) whioh involved descriptions of molluscan and other invertebrate popdationa (Morrison, 1954) with an account of the interrelationships of these organisms by Doty and Morrison (1954) and the plotting of the environ- mental niches of coral reef fishes by Harry (1953). However, it is impossible, at any rate on the basis of these Pacific studies which alone he quotes, to agree with Gerlach (1961) that “ only recently has there been concern with detailed studies of the living habits of coral polyps, of their nutrition, and of their dependence on different environmental conditions ”. It is precisely such work, a t present only being conducted by Goreau at Jamaica (see below), which now needs to be re.sud if we are to analyse the factors influencing the distribution of corals in the varied environments presented by a reef, and especially on an atoll. But we may accept the general conclusion of Gerlach that a reef does correspond, aa Stephenson (1958) had already indicated, to the sub- littoral Laminarian forests of colder seas-which does literally occur at the northern end of the Gulf of California (Squires, 1959). Above it, both a, mid-littoral balanoid zone and a higher littorine zone may on occasions be identified.
VI. ATLANTIC REEFS It hcts been generally accepted that Atlantic reefs are very much
poorer than those of the Indo-Pacific with 8 more limited fauns of corals and associated animals and with a slower growth rate. In the light of recent information, these assumptions require wme qualifica- tion. There is first the question of the coral fauna generally described by Smith (1948). According to Wells (1957) this includes “cmly 26
*Who report the presence of surge channels on the leeward side alao at Raroia.

THE BIOMOY OF CORAL REEFS 225
genera and some 35 species, compared with 80 genera and about 700 species in the Indo-Pacific ”, adding that although Acropora and Porites are the most important genera in both regions, each is repre- sented by only three species in the Atlantic compared with 150 and thirty species respectively in the Indo-Pacific. However, in 1961, Goreau extends the number of species, for the West Indies, to 46. He now considers (in correspondence noted earlier) that these numbers should be further increased.
Newell has made important comparisons between coral reefs in these two great faunistic areas, on the basis of personal experiences at Raroia (Newell, 1956) and in the Bahamas (Newell, 1955, 1958, 1959; Newell et al., 1951 ; Newell and Imbrie, 1955). Unlike Pacific reefs, those of the West Indies are seldom on the margin of the submarine platforms from which they rise and which Newell thinks were probably formed by erosion during periods of low sea level in the Pleistocene. The reefs appear immature, seldom reaching the surface and being largely confined to more favourable, i.e. windward, are-. 0- to the long continued coolness of the West Atlantic during the Quaternary when conditions in the Indo-Pacific were tropical, the re-establishment of reefs was longer delayed and Newell does not think that West Indian reefs are more than between 3000 and 6000 years old. But they are situated on extensive platforms bounded by precipitous escarpments which very probably represent coral formations possibly originating as far back as the Jurassic and only finally killed off in the glacial periods of the Pleistocene.
Diagrams illustrating Newell’s comparison between Weat Indian (A) and Pacific (B) reefs are shown in Fig. 6. The seaward slopes in the former are more gradual but in both he identifim a deeper ‘‘ mush- room zone ” (1) and a higher “ elkhorn zone ” (2) whioh reaohes the always submerged reef crest in A. An algal ridge (3) which breaks the surface at low water of spring tides (the level shown in the figure) with its enclosed reef flat (4) is present only in B. What corresponds in A to the latter region but is continually submerged forms an extensive lagoon ( 5 ) much reduced in B. Recent corals form no more than a veneer over the older rocks (shown stippIed) in A ; this rock has been extemively increased by such coral in B (heavier stipple).
Goreau (1959b), in the first of a series of contributions which will greatly widen our knowledge, has given a detailed description of a fringing barrier formation (typical of many such) off the northern coast of Jamaica, i.e. on the weather side facing the NE trades. All is effectively submerged. By surface examination and by diving he divides the reef into the following series of zones (Fig. 7). (1) A s h e
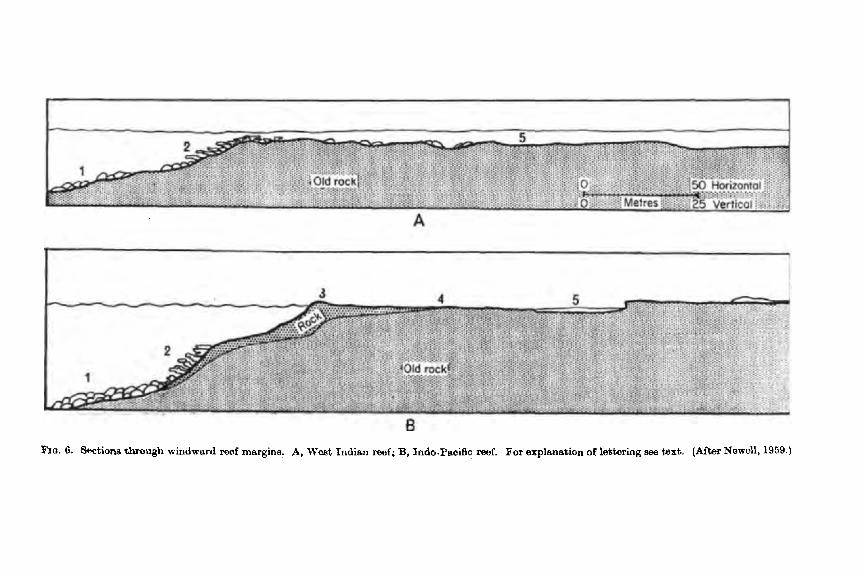
&a. 6. Sections through windward mef margine. A, West fndian reef; B, 1ndo.Pacific reef. For explanation c$ lettering see text. ( A h r Newoll, 1959.)
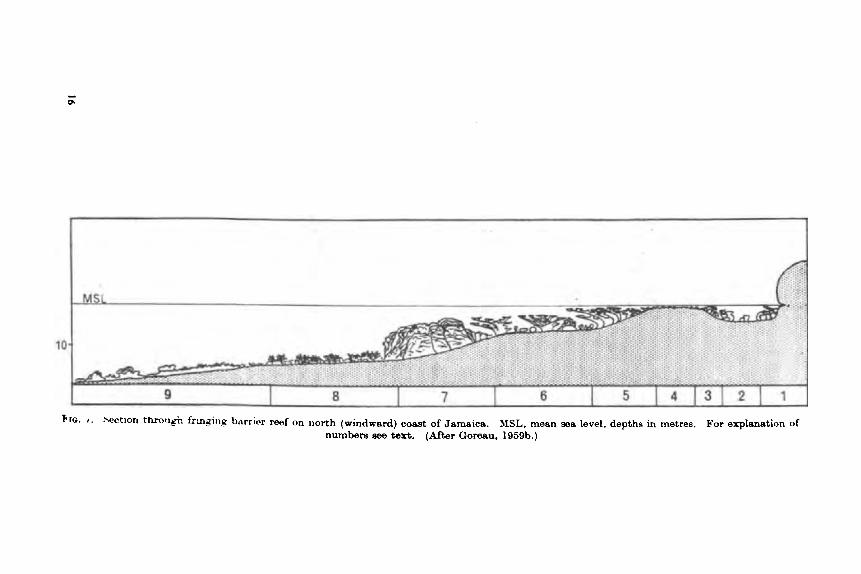
Fro. 7. Section tbl?& frmPiW bmit.r reef on north (windward) coast of Jamaica. MSL. mean ma level, depths in metres. For explanation of numbers see text. (After Gomu. 1959b.)

228 C. M. YONQE
zom down to a depth of 3 m which has a mixed population of corals, including the most hardy species. (2) A channel or lugoon zone which is between 100 and 300 m wide and from 2 to 15 m deep. The bottom is sandy and, apart from the sand-dwelling Hanicinu areolata, commonest amongst eel grass, corals are sparse although gcrgonids, molluscs and echinoderm3 are common. ( 3 ) A rear zone which represents the inner slope of the reef crest and over a depth of 2 to 3 m carries a rich population of corals including both solid and branching colonies, with the former the more numerous. (4) The reef flat or Zoanthus zone, most of which is covered by no more than half a metre of water and is about 40 m wide ; it consists almost entirely of the unconsolidated skeletons of Acropora palmata on which there is a characteristic green growth of the zoanthid Zoanthus sociatzls with a few corals largely confined to the shallow channel and pools. ( 5 ) The palmata zone which is the upper pert
5-
10 20 30 FIQ. 8. Cross-sectional, i.e. east-west, transect through buttress zone (7 in Fig. 7).
MSL, mean ma level. Depths and horizontal distances in metres. (After Goremu. 1959b.l
of the seaward slope descending t o a maximum depth of 6 m and carrying a characteristic and almost exclusive population of “huge tree-like colonies of Acropora palmata which take the full force of the surf ”. The colonies are orientated in the direction of the prevailing seaa forming ‘ I a great jagged comb with irregular teeth ”. Its deeper areas (6) consist of a moat in which A . palntata is still dominant although more spaced out and with much dead coral below. There is also a greater wealth of other corals. (7) The buttress zone, which is described as ‘‘ a region of spectacular underwater scenery l’, consists of huge buttresses of living coral facing seaward and separated by narrow canyons up to 10 m deep, as shown in Fig. 8. More than 90% of the surface is covered with living coral colonies, the most important being the encrusting A arick ugaricites, branching Acropora palmata and massive growths of
with Porites spp. and Milkpora. This is succeeded by (8 ) the ce7vicom& a9 ontastreu annuEaris which form the sides of the buttresses together

THE BIOLOGY OF CORAL REEFS 229
z m which is from 30 to 100 m wide and from 8 to 15 m deep and is largely covered with immense beds of Acropora cervicornis although large colonies of M. annularis and Porites spp. also occur. (9) The annularis zone, which was the deepest surveyed, contains a wealth of massive coral colonies of which M . annularis is the most conspicuous with few Acropora. This reaches a depth of about 30 m.
In his account of reefs off British Honduras, Stoddart (19624 agrees with Goreau in considering M . annularis as the principal reef builder with A. palmta, there attaining heights of 15 ft, a~ specialized for life in the shallower areas protected by these massive corals.
Examination of reefs off the southern coast of Jamaica also revealed zonation with increasing depth but with extensive hurricane damage. I n general, therefore, there is the same pattern in these West Indian reefs aa there is a t Low Isles and other Australian reef formations (see Fairbridge, 1950) or a t Bikini. There are elaborate windward reefs forming, to quote Goreau (196lb), " an organized coherent structure adapted for maximum attenuation of mechanical stresses set up by the constant battering of the seas " and possessing a vigour probably due to a combination of clear water and abundant planktonic food. The former, by its effect (to be described later) on the zooxanthellm, increases the rate of calcification, the latter provides for growth of the tissues and feeding surface. The major difference between fully exposed reefs in the Indo-Pacific (e.g. the Outer Barrier or NE of Bikini) and the Atlantic is the absence in the latter of a massive algal ridge breaking the surface at low water of spring tides. There is almost complete resemb- lance between the leeward reefs. All are less consolidated and grow in more turbid waters containing less zooplankton. They do not develop under such continuous mechanical restraint and for that reason are easily devastated by occasional cyclonic seas.
Unlike the Indo-Pacific, little of the West Indian coral fauna is ever exposed between tide marks. In his account of the ecology of the shores of Florida, Stephenson and Stephenson (1950) notes the presence of only eight, all notoriously hardy, species in this zone.
VII. EROSION The growth processes of reef-building organisms, of which herma-
typic corals are the most important, must be great enough to offset, probably normally more than offset, the effects of erosion. Destruction is the result of biological and non-biological factors, the relative im- portance of which is often diflicult to determine. Newell and Imbrie (1955) note the conflict of views concerning the relative significance of

230 C. M. YONOE
solution and of organic agencies in the formation of the characteristio intertidal “ nick ”.
We can here only concern ourselves with boring organisms, about which there is still very much to be learnt. Effeotively nothing is known about the effect of the filamentous green algae invariably found BB 8
green zone some distance within the surface of living coral oolonies. Ramon (1955) notes the importance of the blue-green algae HyeUa sp. and Ostreobium sp. which live on and just within the surface of coral rock while he also considers that Porolithon does not merely grow over but actually dissolves its way into underlying dead coral rock.
Thanks no doubt to the presence of nematocysts, corals are little affected by predators. In the West Indies, Marsden (1962) describes the activities of a coral-eating amphinomid polychaete, H e m m d h curunculata, which crawls over Porites porites when it is expanded at night. It eats the polyps, fragments of which with nematocysts and zooxanthellae are found in the gut. The skeleton is presumably not affected and the tissues doubtless soon regenerate. A variety of parrot fishes (Scaridae) and file fishes (Monacanthidae) scrape or nibble living corals according to Motoda (1940b), Schultz (1948) and Bardach (1961). The latter estimated the amount, and discussed the probable impor- tance, of the calcareous matter which passes through the gut of these omnivorous browsing fishes.
Although nematocysts protect living coral, the dead skeleton is penetrated by many organisms. Studies on Raroia by Doty and Morrison (1954) and Newell (1950) stress the significance there of blue- green algae which cover the intertidal rock surface and, poseibly, of the deeper penetrating algal filaments. Together they consider these form the food of a series of gastropods which occupy very definite zones in the intertidal and supralittoral, in the order, from above downwards : Melaraphe - Tectarius -iVerita - ThaM/Morula - Turbo. Of these, Thais and Morula are neogastropoda and will be carnivorous, but the remainder, members of the Archaeogastropoda and Neritacsa and so primarily herbivorous, doubtless scrape deeply with the radub into the rock which has previously been softened chemically by the algae. In the Atlantic, Newell and Imbrie (1955) found similar snaitils grazing in the intertidal “ nick ”.
A notable assemblage of animals bore deeply into the rock, including sipunculids (about which very little is known) and boring barnacles of the genus Lithotrya. Between them, Newell and Imbrie considered these worms and barnacles remove ‘‘ aa much as 50% of the rock substance in the intertidal nick ” at Bimini. Umbgrove (1947) noted the im- portance of the urchin Echinometra mathaei in the same region in the

THE BIOLOGY OF CORdL REEFS 231
East Indies. Certainly in Pacific reefs, bivalve borers are no less important. Recent work on probably the most important genus, [email protected], has clearly demonstrated the initial chemical nature of the boring process and the nature of the modifications which have fitted members of the Mytilidae for boring (Yonge, 1955 ; Hodgkin, 1962).
Crossland (1952) associated the more vigorous coral growth charac- teristic of seaward reefs with “ decreased activity of destructive organisms ”. This is probably correct. The greatest surface of living coral occurs there and this we have seen to be largely immune from attack. Moreover, dead patches due to exposure or collections of sediment are not found on these exposed colonies. The results of an experiment involving local destruction of such seaward coral could be very informative.
Although consideration of the mechanical effects of the sea on the form of reefs lies outside the scope of this review, some of the effects of cyclones may be briefly mentioned. After its careful survey in 1928-29, Low Isles has suffered two cyclones, in 1934 and 1950. The effects of the first were almost immediately studied by Moorhouse (1936) who noted great destruction to branching corals such as Acropora and relatively greater survival by colonies of the more rounded Favia, Porites and the meandrines. He also observed peculiar effects in Porites, due apparently to stimulation of skeletal growth after lowering of the water level in the moats where these colonies live, which caused extensive damage. His paper contains a map showing the principal changes caused by the cyclone. A further, more detailed, survey was made in 1945 by Fairbridge and Teichert (1947, 1948) and Fairbridge (1950). They were particularly concerned with the formation of the shingle ramparts which move across the reef flat from the seaward side and of which four, the youngest probably originating after 1928, perhaps even after 1934, were then recognizable. They regarded them as in a state of ‘‘ progressive evolution ”. Damage by the cyclone had been largely repaired. A further map was produced.
The effects of the second and less severe cyclone in 1950 were studied 4 years later by Stephenson and Wells (1956) and Stephenson et al. (1958). Here again branching coral had suffered heavily but maa- sive colonies had largely maintained themselves. The unstable frag- ments from the broken corals were still hampering recolonization on the seaward side. Further changes in the shingle ramparts were noted. These papers represent major additions to knowledge about this now well-known reef, changes in which have been so carefully studied sinoe its original survey in 1928-29.
Turning to Atlantic reefs, Stoddart (1962a, b) has followed up an

232 C. M. YONOE
account of reefs and cays off British Honduras with a preliminary account of re-survey following the major hurricane of 1961. Over a stretch of some 5 miles north of the storm centre he found that 80% of the reef corals had disappeared while the summits of patch reefs were denuded of living coral. Even the groove-buttress system had been completely destroyed for 10 miles. The only surviving corals in the most devastated areas were massive forms, notably Hd&rea annztlaris, already noted as the most important reef-builder amongst Atlantic corals (Goreau, 1969b).*
VIII. PHYsIoLoaY
Recent increase in knowledge about the physiology of corals has been largely confined to the notable work of Goreau on calcification and growth. The influence on this of the zooxanthellae demands prior deseription of the important recent work on these symbionts. Them has been no further study of feeding, digestion, respiration or excretion since that carried out by the Great Barrier Reef Expedition (Yonge, 1930-31) and by the Palao Tropical Biological Station.
From that laboratory Kawaguti (1944b) has given an account of the pigments in various hermatypic corals. While the zooxanthellae (in the endoderm) provide a brown background, there is a wide range of pigments-green fluorescent, pink, purple, vermilion, yellow and white-in the ectoderm. The first probably serves as a protective light filter which could be the function of others. Pigments in the aherma- typic Dendrophyllia and Balanoph yllia occur in both ectoderm and endoderm and are different.
Horridge (1957) studied co-ordination of protective retraction in various Scleractinia and Octocorallia. Responses to electrical sthuh- tion in individual polyps, best seen in Fungia, resemble those in Actini- aria. In colonial genera, repeated stimulation may never, aein ( I r o n i u p ~ ~ and Porites, affect more than a local group of po!yps or may eventually cause retraction of polyps throughout the entire colony.
IX. ZOOXANTHELLAE A. Nature
Unicellular zooxanthellae are invariably present, usually, although not quite exclusively, confined to the endoderm, in all hermatypic corals. They lie within a type of wandering or carrier cell (see Goreau, 1961a, Fig. 8). They are normally present in the extruded p l a n u b although, according to Atoda (1951a) not necessarily so in Acrqpora
*In the course of recent correspondence, the writer hes been informed by Dr. aOreeu that boring qmnges are major agents of erosion on West Indian reefs.

THE BIOLOGY OF CORAL REEFS 233
bvilggemanni. They are present in many other reef coelenterates- hydrozoans and scyphozoans as well as anthozoans-but not in ahermatypic corals. Knowledge about them has greatly increased within very recent years. They are now known to represent the vegetative stage of dinoflagellates. Hovasse (1937) had already, on morphological groundfl, claimed their relationship with this group but direct proof came initially from Kawaguti (1944a) who cultivated zooxanthellae from Acropora coryrnbosa in Miquel-Allen’s solution and found that some changed from spheres to motile organisms 8-12p long and 5-8p in diameter with the characteristic two flagella of the dinoflagellates (Fig. 9). He concluded that they were a species of G y m d i n i u m . It followed from his discovery that zooxanthella are not necessarily confined to the animal tissues and do not need to be conveyed from generation to generation by way of the egg.
FIQ. 9. Qymnodinium sp., dinoflagellate stage of zooxanthellae from Acrqpwa corymboaa. Length of scale 5p. (After Kawaguti, 1944a.)
Following support of this identification by Pringsheim ( 1 955), Zahl and McLaughlin (1957) and McLaughlin and Zahl(1957), from the Haskins Laboratory, reported similar success at the Lerner Laboratory, Bimini, with the cultivation of zooxanthellae from the scyphozoan, Cassiopeia sp., and an anemone, Condylactis sp. Later (McLaughlin and Zahl, 1959) they added zooxanthellae from the Pacific anemone, Anthopleura xanthogrammica. They employed the most modern methods of isolation and cultivation, including the use of antibiotics, full details of which, together with a considerable bibliography, are given in the last paper. They found that, in vitro, (1) vegetative cells (i.e. zooxan- thellae) may give rise to new vegetative cells ; (2) such cells may give rise to motile cells which, in their possession of a transverse girdle, two flagella, one in the girdle and the other extending backwards, together with spiral movement, were undoubtedly dinoflagellate ; (3) motile, i.e. dinoflagellate, cells were, for the first time, observed to change into

234 C. M . YONCE
vegetative cells. They also confirmed previous impressions about the toughness of the outer membrane in the vegetative phase and gave much information about the maintenance and the nutritional re- quirements of these organisms. The most significant of these is the demonstrated ability “ to utilize some normal animal metabolites, particularly waste products ”. This, of course, lies a t the base of their success as symbionts, enabling them to act as automatic agents of excretion for corals (Yonge, 1957).
The symbionts in Cassiopeia and Condylactis were identified as
AP
B A
G r
FIG. 10. Symbiodinium microadriaticum, vegetative cells. A, young vegetative cell con- taining numerous chloroplasts (Ch) and few products of mntabolifim; B, slightly older cell showing the charactoritjtic brownifih-orange “axsirr~ilatir~n product” body (AP) and two vacuoles (1’) containing nipidly mnving griiriulcw ( ( j r ) ; c, older cell showing a larger assimilrrt.iori product rind oil drops (0) ; D, vwy old r :d l fmntuining greatly enlarged amimilntion rJrrJdUCt, oil drops, nunloroils K I I I U ~ ~ gruriulw, and no chloroplasts. CW, cell wail ; N, nucleus. (After Freudenthal, 1962.)
Gynmodinium adriatieurn but this has more recently h e n changed by Freudenthal (1962), working on those from C’assiopeiu, to Sym- biodinium microudriuticum. By observations of his cultures he has provided much the most complete account of the life history. St,ages in the development of ‘‘ the vegetative single thin-walled autotrophic cell, commonly recognized as the zooxanthella ” are shown in Fig. IOA-D with details in the legend. Probably cells with a very large “ assimilation product ’’ (AP) have no further history. Others readily &vide (Fig. 11B) with equal distribution of the cell inclusions apart from the “ assimila-
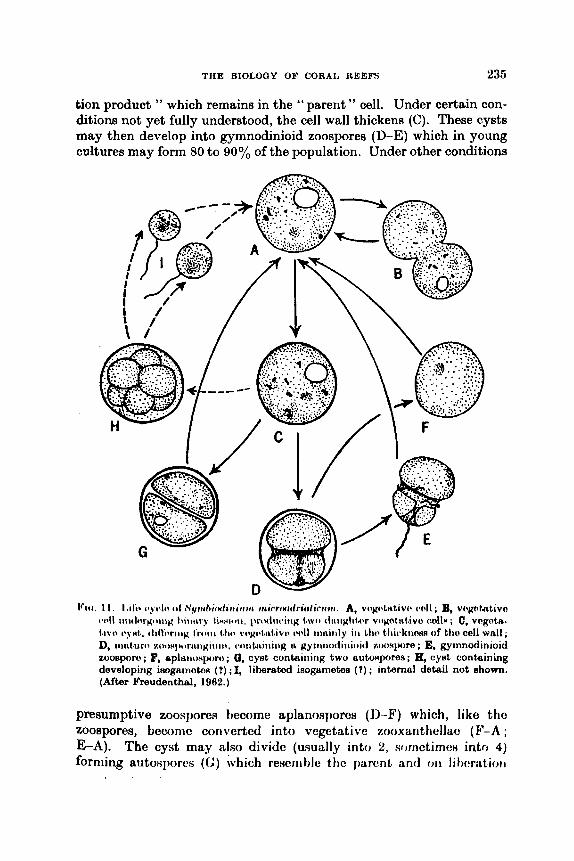
THE BIOLOGY OF CORAL REEFS 235
tion product " which remains in the I' parent " mil. Under certain con- ditions not yet fully understood, the cell wall thickens (C). These cysts may then develop into gymnodinioid zoospores (D-E) which in young cultures may form 80 to 90% of the population. Under other conditions
presumptive zoospores hecome aplanospores (11-19 which, like the zoospores, becomc coriverted into vcgetative zooxunthellac (F-A ; E-A). The cyst may also divide (usually into 2, sornctimt:H into 4) forming autosporcs (C) which rescni M e the purcnt and 011 I i t)cr;Ltiorl

236 C. M. YONOE
form zooxanthellae (G-A). Both in culture and also in the field, spherical bodies, each with a delicate flagellum, may appear and swarm (H, I). These, Freudenthal thinks, may be isogametes whioh on fusion may also give rise to zooxanthellae (this less certain sequence is indicated by the broken arrows C-H-I-A).
While all this work has been done on zooxanthellae from Scyphozos and Actiniaria, the observations of Kawaguti (1944a) leave little doubt that the zooxanthellae in the hermatypic corals are either the same species or one with a similar life history. This assumption is here being made.
B. Signi$cance of the Aumciation
The advantage to the algal symbiont is clear. As “imprisoned phytoplankton ” they gain protection. Immediate access to a source of CO, may be significant where algal concentrations rise to 30000 per mm3; this could otherwise produce severe local deficiency (Droop, 1963). Even more significant are supplies needed for protein synthesis. We now know much more than that nitrates and phosphates may be utilized. McLaughlin and Zahl (1959) have shown that as well as these inorganic sources, the zooxanthellae of Symbiodinium can utilize urea, uric acid, guanine, adenine or any of the twelve amino acids m a source of nitrogen, and also a wide range of phosphoric acids. Much, however, remains to be done on this subject in the experimental field. Zooxanthellae have been shown to intercept ad phosphate that would normally Be excreted by hermatypic corals (and is excreted by the ahermatypic Dendrophyllia). They will aIso remove all phosphate from 2.5 litres of water in which the corals were kept. When the content waa raised from the normal figure of 3.41 mg/ms to the altogether abnormal one of 2036 mg of phosphate almost all of this was removed at the end of 5 days (Yonge and Nicholls, 1931a). The removal of ammonia from the surrounding water has also been demonstrated by Kawaguti (1963). On the other hand, lowered metabolism-resulting from starvation, exposure to sub-lethal temperatures or low oxygen tension-causea immediate ejection, via the “absorptive” zone just within the mesenterial filaments, of great numbers of algae. In other words, a major limiting factor in the concentration of zooxanthellae is the supply, from the animal in which they live, of suitable (and obviously widely ranging) sources of nitrogen and phosphorus. How much the content of zooxanthellae can be increased by external addition of such substances was not, but clearly could be, determined. This has bearings on the problem of calcification ( ~ e e later).

THE BIOLOGY OF CORAL REEFS 237
For the animal the significance of the association has in the past been considered in relation to the possibility that the algae represent (1) a possible source of food, (2) a significant source of oxygen, (3) a significant contribution to excretion, (4) an aid to calcification.
It is no longer disputed that individual hermatypic coral colonies can live in darkness and so effectively without zooxanthellae (Goreau, 1959a). There is no sound evidence that hermatypic corals are not specialized carnivores (like possibly all coelenterates with the exception of the Xeniidae and related Octocorallia mentioned below). While there has been no recent work on the subject, it was shown (Yonge, 1940) that corals collect exclusively animal prey by means of (a) tentacles, (b) tentacles aided by temporary reversal of cilia, (c) orally directed cilia probably aided by extruded mesenterial filaments. There has been no further work on digestive enzymes but all that is known about those of coelenterates generally, notably sea anemones (Nicol, 1959), indicates action exclusively on animal matter.
(1) It is now known that in cultures algae excrete a wide range of organic substances, much of it mucilaginous but including peptides and glycolic acid possibly amounting in all to 50% of the carbon aoquired in photosynthesis (Allen, 1956). The " juvenile " stages in the life history of the zooxanthellae (Fig. 11A) have a thin wall which would facilitate diffusion. They might even, i t is suggested by Zahl and McLaughlin (1959), be actually digested by the animal. This seems improbable, the former the more likely although in this early stage little material may be extruded. It has, however, been demonstrated by Muscatine and Hand (1958), using the anemone Anthopleura elegantis- sima, that material can pass from the zooxanthellae into the animal tissues. They exposed the anemones to sea water labelled with "CO, and by the use of autoradiographs showed the initial fixation of the labelled carbon in the zooxanthellae (confined to the endoderm) and its later presence widely dispersed throughout the body. Passage of material from the zooxanthellae is thus demonstrated but neither its nature nor its significance. Goreau and Goreau (1960a) have taken the matter further using the common Atlantic hermatypic corals Manicinu areolata and Montastrea annuhris. They ran experiments both in light and in darkness with corals containing zooxanthellae and with other corals which had lost their zooxanthellae following 3 months in complete darkness. After exposure for 50 hr to water containing Na,Cl4O0,, they found no significant uptake of radio-carbon in the dark or in the light where zooxanthellae were absent. Where these were present the tissue background activity was about five times higher than in corals kept in the dark. But they considered the level of the transfer

from the zooxanthellae to be “ surprisingly low, especially if the pos- sibility that the zooxanthellae supply the coral host with food materials is considered.” Indeed, the level of radio-activity wits much lower than that found by MusccLtinc ilnd 1 L i d , indicnt~ing t hil t cont1ition.s in Actiniaria iiiity not bc a soiintl critcrion for thosc in thc Sclcmctinia. Goreau and Ooreau conclude that at best thc amount of transferred material could be of little significance as food. However, apart from their effect on calcium metabolism considered below, they reiterate an earlier conclusion (Goreau, 1959a), “ that the zooxanthellae may have a general stimulating action on the coral host’s metabolism, possibly mediated through vitamin or hormone-like trace factors which are secreted in small amounts by the algae but which by themselves do not contribute significantly t o the nutrition of the coral ”. Muscatine (1961) has shown that green Chlorohydra viridissima withstand starvation or reduced feeding better than do albino individuals. He considers that the zoochlorellae may represent a source of coenzymes or coenzyme precursors to the animal and that this may also be true of the zooxan- thellae in hermatypic corals.
The obviously very different conditions existing in many of the tropical Alcyonacea, especially the Xeniidae, demand mention here. Species of the Xeniidae are common on Indo-Pacific reefs, being easily distinguishable owing to the soft velvety appearance of the large polyps. These show characteristic rhythmical pulsationR recently studied by Horridge (1‘3.56). The presence of zooxanthellae and the reduction in the ventral (digestive) mesentcrial filaments have long been known. Although they possess the typical eight pinnate tentacles, armed with nematocysts, Gohar (194O), working in the Rcd Sea, found no reaction to food of any kind. Later (1048) he found similar conditionR in Cluvularia hamra where also zooxanthellae are abundant and the mesenterial filaments poorly developed with no gland cells. “ The endodermic lining of the coelenteric cavities ”, he states, “ is composed of small cells distended with zooxanthellae, and cofitain nothing else that may be considered as food.”
I n a series of experiments Gohar (1940) kept colonies of various species of the Xeniidae in circulating filtered and unfiltered sea water, similar sets in the light and in the dark. The unfiltered water contained zooplankton. ,411 colonies kept in the dark gradually ceased to pulsate, showed symptoms of starvation and at the end of 2 weeks were begin- ning to disintegrate and were dying. When returned to the light, colonies already showing signs of starvation rapidly regained health and began to pulsate normally. Meanwhile, both Hets of colonies, fed and starved, kept in the light, remained equally healthy.

THE BIOLOGY OF CORAL REEFS 239
This is in striking contrast to parallel experiments carried out over much longer periods on hermatypic corals a t Low Isles (Yonge and Nicholls, 1931b). Colonies kept in the light and fed daily with plankton lived normally for up to 228 days (when the experiment terminated). Those starved in the light immediately began to extrude zooxanthellae. There were clear signs of starvation after only 9 days, the tissues obviously retreating. This was most clearly shown in Fungia danai where after 73 days the greater part of the upper surface of the skeleton was exposed, the “ mouth ” having about half the diameter of the disc. Colonies fed in the dark remained perfectly healthy for up to 228 days but grew paler with the continuous loss of zooxanthellae, starved for lack of light. Other colonies starved in the dark appeared much as those starved in the light except that more zooxanthellae were extruded and these were all obviously dead. More recently Goreau (1959a, 1961a) has made a regular practice of ridding hermatypic corals such aa Manicina areolda of their zooxanthellae by keeping them for appropriate periods in darkness and without harm to the animals.
Thus in the hermatypic corals it is food, usually in the form of zoo- plankton, which ensures survival. Light is less relevant. In the Xeniidae, on the other hand, owing to needs of the contained zooxantheUae, light is essential but the presence or absence of zooplankton is im- material.
In neither Scleractinia nor Alcyonacea, it may be noted, can the a l p survive for long in the dark. They do not have the capacity possessed by the zoochlorellae in Paramecium burwrk and Chlorohydra of living chemotrophically at the expense of the host animal (see Yonge (1944) for references). Even when cultured, Symbiodinium micro- adriuticum shows no capacity for chemotrophy (McLaughlin and Zahl, 1959). Droop (1963) notes that “ poor permeability-one of the factora responsible for obligate phototrophy-could be an advantage for life in a medium as rich in possibly toxic substrates as the interior of an animal cell.”
The Alcyonacea (possibly also the Zoenthidea, according to Goreau and Goreau (1960b) ), do appear to be more omnivorous than other coelenterates. In tropical, although not in temperate, species there is a range of feeding habit correlated in part with the presence of zooxan- thellee. The temperate Alcyonium digitatum appears to be a typical coelenterate in its feeding reactions (Romhdy, 1962) although it possibly also accepts phytoplankton (Roushdy and Hansen, 1960). Nothing definite is known about the feeding habits of the numerom tropical species of Alcyonium but, since the work of Pratt (1905), it haa been known that the ventral (i.e. digestive) mesenterial filaments of

240 C. M. YONOE
genera containing zooxanthellae are reduced. The degree of reduction appears to be directly correlated with the abundance of zooxanthellae in the series Lobophytum-Sarcophytum-Alcyonium (tropical species)- Sclerophyturn leading up to the Xeniidae in which these filaments are so reduced as to appear functionless. All of these genera demand Iight. Other tropical genera, such as Acabaria and other species of Clawularicr (Gohar, i940), apparently live as well in dim as in bright light.
As noted elsewhere (Yonge, 1957), the process of establishing an association between an animal and a unicellular plant must differ according to whether the animals be carnivorous (as in the Sclemctinia) or herbivorous as these Alcyonacea appear to have become. In the former the animal must be initially specialized to take in and tolerate the plants, in the latter it is the plant which must be specialized to resist digestion within the animal. Certainly the end result in the two c a m is very different although some source of energy additional to the zooxan- thellae would appear neoessary in the Xeniidae. The retention of the mouth, although narrow, is possibly significant.
Alcyonacen are often extremely abundant on Indo-Pacific (not West Indian) reefs. Thus on Low Isles they are a dominant part of the fauna, many of the colonies being large and in combined bulk covering wide areas especially in the sheltered anchorage zone as noted at the time of the Expedition and more recently by Stephenson et al. (1958) who noted their presence from M.L.W. to at least 20 f t below L.W.S. Sarwphyton trocheliophorum, Lobophytum cramum and L. pawif i rum valdum were conspicuous, Simularia variabilis, S. jlexibilis, 8. poly- dactyla and Sphaerella echinata were very common and Sinularis conferta gracilis and Sphaerella echinuta were also present. Elsewhere Nephthea mollis is common, while speciea of Xenia, Heteroxenh, Cespitularia, Cluvuluria, Pach ycEavularia and Alcyonium also ocuur. There seems little information about the abundance of these ‘‘soft corals ” on the Pacific atolls.
A thorough and comparative study of feeding and digestion, in Alcyonacea showing all conditions from lack of zooxanthellae to their presence in greatest abundance, as in the Xeniidae, wouId be of the greatest value both for its own sake and for the light the results might show on conditions in the hermatypic corals.
(2) There have been many and extensive estimates of the oxygen produced by the zooxanthellae in the course of photosynthesis notably during the Great Barrier Reef Expedition (Yonge et al., 1932). It wa8 also found that respiration is unaffected by oxygen tension until this drops to between 40 and 50% saturation. mere is certainly no recent evidence to support the view that the oxygen produced by the zooxan-

THE BIOLOGY OF CORAL REEFS 241
thellae is of significant value to the coral, the respimtory needs of which have been often greatly over-estimated by neglecting the effect of oxidation of the secreted mucus during the experimental period (Yonge, 1937).
(3) The presumable increase in metabolic efficiency of hermatypic corals owing to the presence of zooxanthellae which act as automatic agents of excretion (as already noted in their utilization of CO, and of sources of nitrogen and phosphorus) has been particularly stressed by this author (Yonge, 1940, 1944, 1957). Droop (1963) makes the added point that tropical animals frequently contain zooxanthellae whereas their relations in temperate and cold waters do not and associates this with the higher metabolic rate of the former and so their greater need for the complementary services of algae. It can be seen how natural selection would tend to preserve such an aesociation in tropical waters.
It was felt that higher metabolism would involve faster growth which, although possibly of no particular value to the individual coral colony might " be an indispensable factor in the necessarily exceptional powers of skeletal formation possessed by the marine communitiecl known as ooral reefs '' (Yonge, 1940). Proof of this clearly lay in experi- ments involving comparisons of growth rates in corals with and without contained zooxanthellae. It has recently been shown in the im- portant work of Goreau and Goreau (see below) that hermatypic coral8 with zooxanthellae do grow faster than those deprived (by being kept in the dark) of algae. The major reason for this probably resides in the direct effect on skeletal formation, i.e. on calcium metabolism. This leads to consideration of the fourth possible effect of the association, a t any rate in stony corals, namely as an aid to calcium metabolism.
(4) The possible influence of zooxanthellae on skeleton formation was experimentally studied by Kawaguti and Sakumoto (1948). Using four species of corals, they estimated the changes in calcium content in the relatively small volumes of water in which they had been kept. They claimed that uptake was greater in the light than in the dark. However, by using radioactive calcium45 as a tracer, Goreau ( 1969a, 196 1 b) has developed an elegant and precise method enabling him to measure skeletal growth within a few hours of initial exposure. Methods have also been devised for the loading and setting out in natural sites on the reef of weighed and sealed glass jars containing corals with radioactive calcium-45 in the water, the corals later being aampled by means of a hollow steel core punch (Goreau and Goreau, 1959). Leaving for later discussion the growth data ao obtained, our immediate concern is with the process of calcification and the possible role in this of the zooxanthellae.

242 c. ni. YONOE
CALICOBLASTIC J. EPIDERMIS, 1 I{ \
FIG. 12. Diagram (after Goreall, 19B!h) HhoWiiig pCM4Hiblo ptlthWayH of cnlcjwn and carbonate during calcification in a roof-t~uiIdir1g cord. A dirqrarnnlatic cronn- section of tho rn~icob~astic body wall a t tho barn of tho polyp in rhowrr but the parts are not drawn to Rcaln. The coolontoron nnd tho flagollahd gartrodcrmiR containing e zooxaiithella aro &own a t tho top of tho figure, tho calicoblantic epidermin in in the rniddlo and tho organic mornbrano with crystals of culcarooue matter is at the bottom. Ttio direction of growth is upward, i.e. calciuni dopoeition is in a downward direction.

THE BIOLOGY OF CORAL REEFS 243
As shown diagrammatically in Fig. 12, caloium appears to be taken in directly from the water and not to be concentrated in the tissues prior to deposition in the skeleton. Indeed the living tissues effectively isolate the skeleton, preventing interchange or rer~ction with the sea water (Goreau and Goreau, 1960~). The skeleton, which consists exclusively of aragonite-the inability to secrete calcite has been suggested as a possible reason for the paucity of corals in cold waters- is, in the opinion of Goreau, formed outside the ectoderm cells. He postulates a passage, by as yet unknown means, of calcium through the tissues to be '' adsorbed on a mucopolysaccharide-like material that forms part of the organic matrix and acts as the template upon which the initial stages of skeletal mineralization occur " (Goreau, 1961a).
He postulates that the calcium combines with bicarbonate, largely produced in metabolism, to form first calcium bicarbonate and then calcium carbonate but the efficiency of this depends on the effective removal of H,CO, which depends on reactions catalysed by carbonic anhydraae. The various reactions are thus aa follows :
Ca++ + 2 HCO-, 5- --* Ca(HCO,),
Ca( HCO,), +- ~ CaCO, +Fa H,CO, carbonic anhydrase H+ + HCO-,
--* - _- H,CO, carbonic anhydrase CO, + H,O
+ . - - - . . - - . . -.
The prescncc of carl)onio arihytlrnrcc: WILH tlarrrorircCrrt!,r:tl try t,rwd,rwri!, with thc inhibitor, Ihrnox. (hIcjfication WUN rtx~uot!t! by Horri t : GO'%, in light and 76% in darkness. The effect in light was to some cxtent reversed when zooxanthellae were present and photosynthcsising. This is to be expected since both CO, and HCO, are fixed by these algae which would therefore assist calcification in hermatypic corals.
Indeed there can be no question about the advantage conferred by the presence of zooxanthellae. Goreau (1961 b) HtateB that growth in fourteen species is on the average ten times faRter in nunliyht than in darkness, calcification being ctually reduced by 50'g Or1 a clr~udy day. The results of experiments with Manicinn areohla are 8hown in Fig. 13, the corals being deprived of zooxanthellae by a standard procedure of keeping them in darkness for 6 weeks.* By this meens he has
?
* A much longer period wm needed at Low Isles and one wondem why.
17
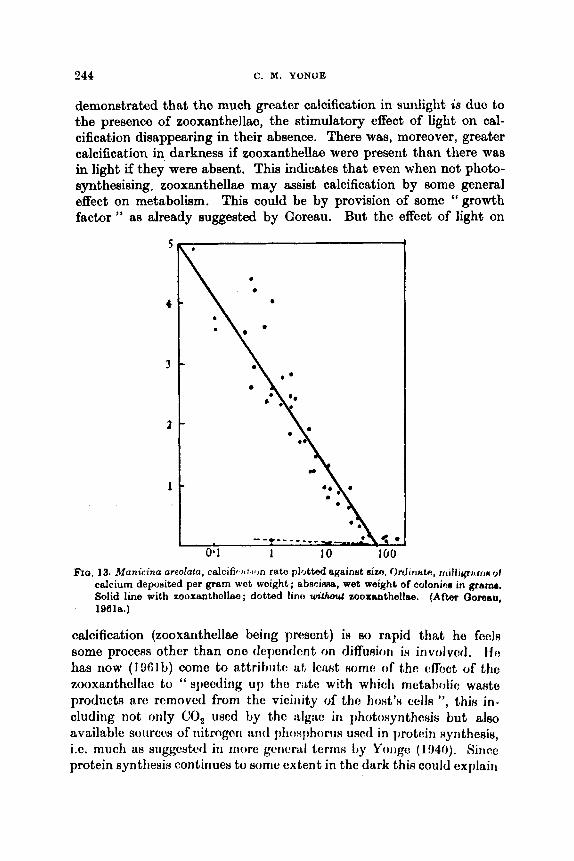
344 C . M. YONUE
demonstrated that the much greater calcification in sunlight is duc to the presence of zooxanthellao, the stimulatory effect of light on cal- cification disappearing in their absence. There was, moreover, greater calcification in darkness if zooxanthellae were present than there was in light if they were absent. This indicates that even when not photo- synthesising, zooxanthellae may assist calcification by some general effect on metabolism. This could be by provision of some " growth factor " as already suggested by Goreau. But the effect of light on
FIG. 13. Manicino areolalo, colcific 1 1 1 ion rato plotted againnt aim. Or~Jiwik, rrrilliyrrrrrtr of calcium deposited per gram wet weight; abscinntr, wet weight of colonias in gram. Solid line with zooxanthollua ; dotted linu without zooxsnthellae. (After Ooreau, 19618.)
calcification (zooxanthellae being present) iR so rapid that he feel8 some process othcr than ono dcpendcnt on diffuaiorr i~ involved. f f f ?
has now (19Olb) come to sttrihutc* at h s t soma of thc cffcct of tho zooxanthellae to " specding u p t h o r:& with which metabolic waHtu prodiict8 arc removcd from the viciiiity of the host'ti celh ", this in- cluding not only CO, used by tho :dgao in Ijhotosynthcsis but also available sourccs of nitrogen arid phosphorus used in protein Hyrithesis, i.e. much as suggcstctl in niorc gcricrd t e r m by Yoligc (1!14O). Sirice protein synthesis continues to some extent in tho dark this could explain

THE BIOLOGY OF CORAL REEFS 245
the greater calcification in darkness when zooxanthellae are present. The ‘ I voracity ” with which zooxanthellae will take in phosphorus (and presumably nitrogen) not merely from within the coral but also from the surrounding medium has already been mentioned (p. 241). It could be illuminating to discover to what extent the content of zoo- xanthellac in hennntypic corals could be actually increased by enriching the external medium with phosphates and ammonia. There is the final point that if, as suggested by Yonge and Nicholls (1931a), corals do obtain much of their energy by deominization of protein, the beneficial effects of the zooxanthellae in removing the end products will be all the greater.
Thcrc is firially the fact, already noted on p. 218, that the presence of zooxanthellac appears to bc essential for normal development in hermatypic corals. This in itself indicates a very intimate metabolic association between animal and plant.
X. GROWTH
A clear distinction must be made between growth of the tissues and that of the skeleton even though this is due to the secretory activities of the tissues. While both obviously increase in young corals there is a later slowing down in tissue increascs which inevitably affects calcifica- tion ; but if the tissues were finally to stop growing completely, calcifica- tion would presumably stop, otherwise the tissues would become increasingly attenuated owing to stretching over the ever-enlarging skeleton.
Early work on growth was cntirely confined to the results of crtl- cification, i.e. to measarcmcnts of lcngth, dinmator or weight of coral8 usually kept uiidcr natiiral conditions. ‘llhcm provided figure3 on which estimates of ttic possible rate of iricreiw of coral reefs hove heon based. Information also covering tissue increase was later obtained by observing increase in the number of polyps, initially in developing colonies in the laboratory, Later Manton (1932) followed tho growth of individual branches of Pocillopora bulbosa kept in the sea while Motoda (1940a) compared the number of polyps (or calyces) in the skeletal mass in varying sized colonies of Qoniastrea aspera. Thc growth of single free individuals of Fun& actinifmis ha8 been followed by Aho ( 1940) who demonstrated some relation between growth and environmont although without analyzing the factors concerned.
It enlerges from these observations that initial rapid growth rate of a colony, or of n singlo individual in the case of Fungia, is followed by ib slowing down leading to almost complete cessation. In Qoniastrea,

246 C . M . YONOE
Motoda thinks this may be due to physiological senescence although admitting i t could also be associated with decrease in the surface/ volume ratio as the colony changed from t,he first flattened plate into the final massive hemispherical form. As shown by Kawaguti (1941a), during growth in these rounded colonies the volume increases in proportion to the cube of the radius whereas the surface area increases in proportion to the square of this. Thus in colonies with fifteen or less polyps he found the weight per polyp to be around 0.15 g, with about 40 polyps around 0.40 g and with between 468 and 486 polyps about 0.80 g. This means, of course, that the needs of calcification increase at a far higher rate than does the surface area of living tissue which
FIQ. 14. Calcification rates in a branch of Acropora conferfu. A, 8.2 i 3.76 p g Ce mg N-* hr-' ; B, 1.9 f 0.92 ; C. 0.5 3~ 0.10. Arrow shows direction of primary growth, scale represents 10 cm. (After Goreau, 1959a.)
represents the feeding surface. This could well explain the eventual cessation in growth.
Goreau's work has completely altered our understanding of both tupects of growth. He has estimated total nitrogen (as a measureof the living tissue) and the rate of calcification by the simultaneous use of '5Ca and W-carbonate (Goreau, 196lb). He had already (see Fig. 12) postulated distinct modes of entry and now finds that the labelled calcium is incorporated up to seventeen times as fast as the carbonate, indicating that greater quantities of carbon than of calcium are available in the tissues.
Apart from the effect of the zooxanthellae, the reasons for which do still remain somewhat uncertain, growth is influ'enced by " inherent species specific factors " (Goreau, 196lb). This is to be expected. All ahermatypic corals have their characteristic growth forms and asRocia-

THE BIOLOQY OF CORAL REEFS 247
tion with zooxanthellae in hermatypic corals would not be expected to modify pre-existing growth forms-except possibly in respect of light as discussed below. Thus in Acroporu confertu (Fig. 14) the terminal polyps, which are large and pale, have a much higher calcification rate
1
5 cm . A B
, Scm 1
C , Scrn
D FIQ. 15. Axial growth gradients measured in p g calcium p r mg of tiHLiue nitrogen per
hour in A, Acropora cewicornia (1, 71.5; 2, 49.0; 3, 30.0; 4. 15.0): B, A . pilmcrlcr (1, 60.0; 2, 30.0; 3, 15.0; 4, 8 .0 ) : C, Por~lea~urcalu ( I , 204; 2, 4.0; 3, 2 . 0 ) : D, Colpophytlio notons (1, Y.0; 2, 8.0; 3, 10.0; 4 , 9.0; 6 , 104). (Aftor Gimtiu, IObJr.)
than the other polyps which although smaller have u, much higher content of zoosanthcllae.
Thus in Imuwliiiig voni in ~ i w i i I ~ H Acroporn m-vicorni,v, A. palmata and Porites furcxzln (Fig. 16, A, U, C) rates at tho apioal arcas are up to ten timm \vliat0 tIlwy nro in Int,cmil or basal arras, whcrctrs in massivo colonios (D)
( ! ~ ~ l ( ~ i f i ( ~ i ~ ~ , i o i ~ nLt,c\x colitrol tho ovcntid forin of the colony.

248 C. M. YONQE
such as Colpophyllia natans (with which Gonimtrea mpera could be compared) there is little difference from one area to another (Goreau, 1959a, 1961a). Nevertheless, the environment may exert a considerable constraining influence, for instance in the formation of flattened colonies of Sidermtrea radians on intertidal beach rock compared with rounded colonies in still, deeper water (Yonge, 1935b). '
These laboratory findings were confirmed by Goreau in the field. Further evidence was also obtained of discontinuous and irregular growth within a single symmetrical colony and also between individual colonies of the same species, conclusions already reached by earlier workers. The probability that calcification is not a continuous process is indicated by the lamellated nature of the skeleton. Field experiments on a wide range of corals revealed that there was no necessary correla- tion between rate of calcification and relative abundance of different species on the reefs. It appears that the race is not necessarily to the most quickly growing species.
Particular interest attaches to studies on Manicinn arwEata (Goreau and Goreau, 1960b). After a period of initial attachment, this interesting coral lives free on the surface of sand and so occupies the same habitat as the Indo-Pacific fungids (Yonge, 1935a). Here also the growth rate falls with increasing size (i.e. age). As shown in Fig. 13, the specific calcification rate in the smallest colonies was seventy-six times greater than in the largest. Senescence (perhaps by reduction in cell division) could account for this, as postulated by Motoda for Gonimtrea. Goreau and Goreau do not think that zooxanthellae are in any way concerned. Unlike related massive corals, the ratio of sur- face area to volume (measured respectively by total nitrogen and weight) remains almost constant in this species. This is due to the increased folding of the surface with age which is rightly regarded aa an adaptation for life in sand. As described and illustrated by Yonge (1935a), M . ( = Maeanclra) areolata can clear itself when covered with sand by a distension of the coenosarc with water. It can also, as observed by Goreau and Goreau, by similar means right itself after being turned over. These activities demand an extensive coenosarc capable of being raised well above the surface of the skeleton. The fungids are capable of similar movements.
XI. EFFECT OF LIGHT
The supreme influence of light on the growth of hermatypic cords has long been apparent. As stated by Umbgrove (1947), " the depth of the living reef is correlated with light penetration ". Earlier evidence

THE BIOLOQY OF CORAL RBEFS 249
has already been reviewed (Yonge, 1940, 1958b) and positive photo- tropism strikingly revealed by the experiments of Kawaguti (1937) on regeneration of branches of Acropora suspended horizontally and upside down (Fig. 16).
This response to light is certainly associated with the presence of zooxanthellae. In both corals and other marine invertebrates con- taining dgsl syinbionts, Knwaguti (1941b, 1944c) considers that the respouse to light varies in intensity with the concentration of algal cells. Gohar (194%) dcscribcs how both the cxpansion of the polyps and the growth of the stolon in the alcyonarian Clavularia hamra, already noted as containing zooxanthellae, are influenced by light. Zahl and
RIcLeughlin (1!)50), who give further references, state in summary " that every species of zooxanthellae-bearing marine animal has its own spccific light needs and therefore tihows characterintic pAitirming on the reef. This may he acc'omplished through m y of I h f g Altjwjrbg means : (a) early selection of position, ( t ~ with plani~~rtc! fjr d h c r frcdy motile larval forms ; (b) gross body movemantH or poduring, a~ with anemones, medusae, and worms ; (c) posseasion of light-filtering pigments, as with coloured corals and molluscs ; (d) possession of light- conccntrating devices, as with tridacnids ; (e) possession of highly contractile and differentially light-absorbing tissues, aa with most coelentcratcs atid some molluscs."
Thcre iu tlia l i iu i l qiicNtion as to whcthc*r the positivo reaction to light is the factor pre-conditioning establishment of the symbiosis or

250 C. M. YONQE
whether it arose aa a result of that association. In the sense that, as Goreau and Goreau (19.59) have demonstrated, light intensity has a profound effect on coral growth owing to the presence of zooxanthellm, then the latter alternative appears the more probable. Droop (1963) makes the interesting suggestion that ‘ I irritability experiments with artificial amociations could decide whether the reaction to an internal oxygen gradient is inherent or not.”
XII. PRODUCTIVITY
Localized centres of high productivity in the open ocean, like oase0 in a desert, atolls are of compelling interest. Each lagoon, in the words of Von Arx (1954), I ‘ is essentially a ‘ lake in mid-ocean ’, cupped in a shallow saucer-like basin supported by a solitary mountain peak ”. It also represents an area of sea bottom raised from aphotic depths into the photic zone. Atolls lie across the trade wind belts, which fact accounts for an asymmetry due certainly to the greater force of seas on the windward side but possibly also to the greater quantity of zooplank- ton on that side. It has been claimed, as discussed later, that an atoll represents a completely closed system effectively neither taking material from nor releasing material into the surrounding ocean. But atolls have grown to their present size in the past and probably con- tinue to grow and this must have involved, and still involves, some interception and utilization of organic and inorganic matter from the sea. The problem also arises with fringing and barrier reefs but there, particularly with the former, the factors are more difficult to analyse owing to the presence of complications such aa the effect of land drainage and of greater sedimentation.
The extent to which lagoon waters are isolated from those of the surrounding ocean has been demonstrated by the work of Von Arx (1954) at Bikini and Rongelap. He notes they are infiuenced by four sources of energy, namely wind, waves, tides and the North Equatorial Current, with the trade winds much the most important. Apart from their effect on the seaward reefs, they cause a steady inflow of water through channels on the reef surface into the lagoon. There is no such major inflow on the lee where water movements are largely influenced by the tides. The wind also drives the surface waters in the lagoon, to &
depth dependent on the force of the wind, in steady movement from seaward to leeward side. The greater part of this water cannot escape so is driven below to return upwind as a bottom current (Fig. 17). There is also a secondary circulation consisting of “ two counter-rotating compartments, which move clockwise in the southern portions and

THE BIOLOGY OF CORAL REEFS 25 1
counter-clockwise in the northern portions." Measurements of water flow into and out of Bikini lagoon indicated that the total volume of water (approximately 28 x los m3) is completely replaced every 39 days during the trade wind season and in about double this time during the summer. Thus there is retention with continuous replenishment.
The same picture emerges from the study of the plankton by Johnson (1954). Certain species are found in both open ocean and lagoon but four times more abundantly in the latter owing to retention of water within it. There are also endemic lagoon species able to pass the entire life history there. They live almost entirely in the deeper slowly moving levels whic'h represent up to 90% of the water in the lagoons. Even upward migration at night may not take them out of
FIG. 17. Diagram (based on Von Arx, 1954) showing circulation in an atoll lagoon under the influence of trade wind (T). Surface current caused by this but the greater past of the water returns a8 a deep current which upwells on the sheltered side of the lagoon. While energy thus largely conserved within lagoon, maintenance and growth of the atoll depends on a constant supply of energy (zooplankton) from the open ocean (A).
this water which also contains the jdanktonic H t a p (if thc: liot~tom- living invertebrates and.fishes found in the lagoori. Little of t th c!ntlorn i f ! and temporary plankton is loat into the ooeari.
Various workers have attempted cstimutos of organic productivity on atolls and other recfs basing thwe on measurements of oxygen production, i.e. of carbon utilization, across the upper surfaces of windward reefs (Sargent and Austin (1!)4!j, 1954) on Rongelap Atoll; Odum and Odum (1956) on Eniwetok Atoll; Kohn and Helfrich (1957) on the fringing reef at Kapaa, Kauai, Hawaii). Their eRtimatcs indicate a high primary productivity of 1500-3500 g carlton fixed per ~qiiare metre annually. This is up to 100 times highcr than in the nurroundirig tropical ocean and four to eight times groatw than i n tho r n o d pro- ductive temperate seas.
But these upward-facing surfaces are inhabited hy spwirtlixc:d plant communities largely of encrusting nullipores. I n addition zooxarlthclleo

262 0. M. YONQE
occur in the corals and almost all other coelenterates as well as in compound ascidians and bivalve Tridacnidae. There are also the blue- green and filamentous green algae on or in the coral skeletons and boulders. On the other hand the bulk of the fauna consists of the sessile animals which harbour these zooxanthellae. Corals are con- spicuous but largely because of the great surface exposed. The biomass is small, there is no succession of overlapping surfaces and colonies there is off the windward reef margin. All of these animals feed on zoo- plankton or phytoplankton and almost the only animals capable of feeding on the calcareous algae are parrot fishes (Smrw) which “ move about the reef, grazing like a herd of sheep . . . Their tooth marks remaining on the rocks, are easily observed” (Schultz, 1948). These predominantly plant communities are not surprisingly autotrophic.
Unable to collect adequate supplies of zooplankton, Sargent and Austin and also Odum and Odum postulate that corals obtain nutriment from associated plants. The former indeed state that “ under conditions of growth on the eastern reef of Rongelap Atoll, the association between corals and zooxanthellm seems to be essential to the individual colonies precisely because the corals must derive organic matter from the algae or die.” They do not explain how this may be achieved. To meet the same postulated need, Odum and Odum, who estimated that the zooxanthellae comprised no more than 6% of the total plant mass, bring in the filamentous boring algae universally present in coral skeletons. They do not state how the animal codd utilize products from plants separated from them by an appreciable depth of skelet.al material. Moreover, Goreau and Goreau (1960b) found that in Municina areolata the content of filamentous boring algae is very much less than that estimated by Odum and Odum.
This fundamental problem of coral nutrition has already been discussed in connexion with Goreau’s work and his conclusion that the contribution of the zooxanthellae can be no more than some d i h b l e organic matter which may serve as a vitamin or hormone. Stephens (1960) has recently shown that Fmgia can remove labelled glucose and also amino acids from solution. There remains the undoubted, well demonstrated, fact that corals are most highly adapted for the capture and extremely rapid digestion of exclusively animal prey and that, in relation to the bulk of the tissues, they have a literally enormous feeding surface in most species only exposed at night when zooplankton is abundant. Nevertheless in view of these contmry views the question posed by Hand (1956) as to whether corals are herbivorous must, following further investigation, be conclusively answered.
On the basis of their 6 weeks’ survey of a reef on Eniwetok, Odum and

THE BIOLOQY OF CORAL REEFS 253
Odum attempted to draw up a balance sheet of gain and loss. While in the words of Hedgpeth (1957) “This tour de force will probably excite comment for some years ”, it was surely somewhat premature. So little is yet known about food chains within a coral reef community ; indeed the authors themselves are not too accurate about tho trophic gtatutl of certain of the animals. Thus in their pyramids of biomass, apart from their assumption that coral are primary producers, they list the gastropods Cypraea and Thais and the Ophiuroidea all as herbivores whereas they are specialized carnivores, on the other hand crabs, indeed decapod Crustacea generally, are omnivores rather than carni- vores. However i t could be that the final estimate in dry biomass in grams per square metro of carnivores 11, herbivores 132, primary producers 703, may be a reasonable approximation.
But the prime criticism of this admittedly bold and very interesting attempt lies in tho basic failure to distingukh between the outer sea- ward growing (or certainly maintaining) margin of the reef and the upper reef flat with the encircled lagoon. With little doubt the latter regions are in a more or less steady state with gain from and loss to the ocean roughly balanced. In the words of Odum and Odum, under present ocean levels the reef community may represent ‘ I a true ecologi- cal climax or open steady-state system ”. Certainly the lagoon may be regarded as a localized area within which, over a long period of time (since late Eocene it would appear in the case of Eniwetok), productivity has been built up by a gradual accumulation of nutrients needed for protein synthesis. This has been achieved by intercepting nutrients in surface waters and, whcrc zooxunthcllao arc present, retaining them in closed cycle withiri t ho ‘ I orgnnim ”. I n (:onRequ(!nco tho lagoor1 arid mcircling H I I ~ ~ I L W r c v f i N riow i L n iircii of high productivity in ocoanic watcm o f t\.utrtwwly low productivity. Sorno measure of the difference niiiy bc- iinplicd in the cstirnato of Emcry et al. (1954) that sedimentation on the summit of an atoll is about 1000 times faster than in the mrrounding dcptlis. In the warm, well illuminated and well mixed lagoon wntcrx thrrc is a rapid turnover of tho cndcmic planktonic and benthic population, although whother this amounts to an annual replacement by as much as 12.5 times the average standing crop, which is what Odum and Odum estimate, remains to be confirmed.
All this takes 110 account of the outer seaward surface. There, the hermatypic corals and associated animals can only be maintained from oceanic sources-and if they are not maintained the whole reef formation will be disrupted. There is no nutritional connexion with the reef flat and the lagoon. This exposed, seaward community i R outside the closed system where essential nutrients aro retained and

264 C. M. YONOE
circulated. There is much evidence that the most actively growing corals are those some metres below the surf- on the windward side. They have the major responsibility for maintaining the stability of the entire atoll against the constant battering of trade wind sem. This involves constant replacement of eroded or damaged surf-. With the zooxanthellae of essential assistance in maintaining a high rate of calcification, there must also be a constant supply of food in the form of zooplankton if only to replace the organic matter continually being lost, carried around the atoll or dropped into profound depths. There. can be no closed or autotrophic system in this environment. Although this is the most difficult of all coral reef environments to penetrate, every endeavour should be made to sample its waters and obtain some estimate of the amount of zooplankton, largely no doubt some distance below the surface, which is carried against these seaward reef surfaces. The position of the partly downward directed ourrents containing this essential zooplankton is indicated in Fig. 17, A.
We are thus brought logically to the supreme question which ariws in any consideration of the biology of coral reefs, namely to what extent are the powers of growth capable of withstanding the full force of the sea; in other words, are modern reefs static, advancing or retreating? It has long been the opinion of the writer (Yonge, 1940) that seaward reefs do grow outwards against constant forces of wind and sea which mould their form. This is the opinion of Kuenen (1960), and also of Crossland (1962) who makes the point that " Permanent beds of living coral can exist only immediately along an outgrowing reef where their foundations are being continually extended." Emery et al. (1954) consider that the windward reeh at Bikini a p p r to be growing seaward. Newell (1956), on the other hand, considera that the seaward reefs at Raroia are getting narrower and that the surge o h e l s are. formed by erosion, i.e. in opposition to the more general view that it is the intervening algal ridges which are growing outwards.
By placing permanent marks on reef surfaces it should not be difficult to discover precisely what is happening. But certainly in the past the associated powers of growth and high calcification rate have permitted the formation of reefs exposed to the full force of oceanic seas. Munk and Sargent (1954) estimate that the wave% beating against the windward reefs a t Bikini dissipate a force of approximately half a million horsepower which the ridges and surge channels sucoe~ully withstand. It is indeed difficult to see how living reefs could successfuUy retreat before such forces. Once the powers of erosion gained the upper hand, the living reef surface would be breached and damage would be progressive and irrevocable. As suggested earlier, obeervation of the

THE BIOLOGY OF CORAL REEFS 255
results of a localized area of experimental destruction on an exposed reef surface, i.e. of breaching the surface of living tissue, could be most illuminating.
ACKNOWLEDGMENT
The author wishes to acknowledge the great assistance received from his research assistant, Miss J. I. Campbell, who copied and modified the figures and checked the final manuscript.
XIII. REFERENCES
Abe, N. (1940). Growth of Fungia actinifornab var. palawe& Daderlein and its environmental conditions. Palm trop. biol. Stud. 2, 106-146.
Abel. E. F. (1959). Zur Kenntnis der marinen Hohlenfauna unter besonderer Beriicksichtigung der Anthoaoa. Pubbl. Slaz. 2001. Napoli, 36, Suppl. 1-84.
Allen, M. B. (1956). Excretion of organic compounds by Chlrmaydomonua. Arch. Mikrobiol. 24, 163-168.
Atoda, K. (1947a). The larva and postlarval development of some reef-building corals. I. Pocillopora demicomia cespitoaa (Dana). Sci. Rep Tdhoku Univ. (4), 18, 24-47.
Atoda, K. (1947b). Tho larva and postlarval devolopment of Borne reef-building corals. 11. Stylophora pistillata (Espor). Sci. Rep. TGhoku Univ. (4), 18,
Atoda, K. (19618). The larva and postlarval dovolopment of some reef-building corals. 111. Acropora briiggemunni (Brook). J . Morph. 89, 1-16.
Atoda, K. (1951b). The larva and postlarval development of 8ome reef-building corals. IV. Cfahxea wpera Quelch. J . Morph. 89, 17-36.
Atoda, K. (1961~). The larva and postlarval development of some reef-building corals. V. Seriatoporq hyst+x Dana. Sci. Rep. T6hoku Uniu. (4), 19, 33-39.
Atoda, K. (1951d). Asexual reproduction in somo young reef corals of Seriato- poridae. Sci. Rep. T6hoku Univ. (4). 19, 178-186.
Atoda, K. (1953). The larva and poRtlarva1 devoloprncjrit, of tho rmf'hilding coral. S c i . Rep. Z'bhoku Univ. (4), 20, 10.5 121.
Bardach, J. E. (1961). Transport of calcaroou~ frrrgrnentH by rvef fhhw. Science,
Boschma, H. (1948s). Specific charactem in Millepora. Proc. Kon. Ned. Akad.
Boschma, H. (1948b). Tho species problem in Millepora. Zool. Verh. Mua.
Boschma, H. (1949). The ampullae of Millepora. Proc. Kon. Ned. Akad. Wetensch.,
Boschma, H. (1950). Further notes on the ampullae of Millepora. 2001. Med. M w .
Boschma, H. (1951). Notes on Hydrocorallia. Eool. Verh. Mur. l,e&n, no. 13,
Cmssland, C. (1952). Marlreporaria. Hydrocorallinnc, Hdiopra and l'ubiporrr.
Doty, M. S., and Morrison, J. P. E. (1964). Int,errelationship~ of the organisms
48-64.
133, 98-99.
Weknsch., Ameterdum, 51, pp. 818-823.
Leiden. no. 1, 115 pp.
Amsterdam, 52, pp. 3-14.
Leiden, 31, pp. 49-61.
49 PP-
Sci. Rep. Qr. Barrier Reef Exped. 1928-29, 6, pp. 86-267.
on Raroia mido from man. Atoll. Rea. Bull. no. 36.

286 C. M. YONOE
Droop, M. R. (1963). Algae and invertebrates in symbiosis. “ Symbiotic Associa- tions ”, Cambridge University Press, 171-199.
Durham, J. Wyatt (1947). Corals from the Gulf of California and the North Pacific coast of America. Mem. geol. SOC. Amer. 20, 46 pp.
Durham, J. Wyatt (1962). Corals from the Galhpagos and Cocos Islands. Proc. Calif. A c d . Sci. 4th ser., 32, pp. 41-56.
Durham, J. Wyat,t,, and Allison, 14:. C. (1900). The gcologic history of Baja California and its m~ririo failtias. In Syrripo~iiim : The Biogcography of Baja California arid adjacorit H O ~ R . Syatem. Zool. 9, 47-91.
Durham, J. Wyatt, and nctrnard, J. Imirons (1952). Stony corals of the eaetern Pacific collected by tho Velero I11 and Velero IV . Allan Hancock Pai f . Exped. 16, 1-110.
Edmondson, C. H. (1946). Bohaviour of coral planulae under altered saline and thermal conditions. Occ. Pap. Bishop M w . 18, pp. 283-304.
Emery, K. P. (1948). Submarine geology of Bikini Atoll. Bull. geol. SOC. Amer. no. 59, 855-860.
Emery, K. O., Tracey, J. I., and Ladd, H. S. (1954). Geology of Bikini and nearby atolls: Part 1, Geology. Prof. Pap. 7J.S. geol. Sum. 26&& 1-265.
Fairbridge, R. W. (1950). Recent and ploistocene coral reefs of Australia. J . Ueol. 58, 330-401.
Fairbridgo, R. W., and Teichert, C. (1947). The rampart system at Low Ides, 1928-45. Rep. Qt. Barrier Reef Comm. 6, 1-16.
Fairbridge, R. W., and Teichert, C. (1948). The Low Isles of the Great Barrier Reef: a new analysis. Ueog. J . 111, 67-88.
Freudenthal, H. D. (1962). Symbiodinium gen. nov. and Symbiodinium micro- adriaticum sp. nov., a zooxanthella : taxonomy, life cycle and morphology. J . Protozool. 9, 45-52.
Gerlach, S. A. (1961). The tropical coral reef as a biotope. Atoll Red. Bull. no. 80.
Gohar, H. A. F. (1940). Studies on the Xeniidae of t,he Red Sea. Publ. Mar. biol. Sla. Qhurduqa, no. 2, 26-1 18.
Gohar, H. A. E’. (1948). A chic:rijJl.ion n r i d norno khhqkd nliifliws r t f R new alcyonarian HpocieH “ Chvuhrirl tumra (hfinr ’ I . 1’dd. Mnr. 6 h l . ,4&4.
Qhardaqn, no. 6, pi). 3-33. The physiology of skoleton formation in corals. I. A
inot~liotl for niwi,siiriiig the r8t.o of ralciiim doposit,ion by corals under different contlit,iontl. I W . l h l l . 116, 6U--75.
Goroau, ‘1’. P. ( 1 9591)). Tho ocology of Jamaican coral reefs. T. Spccies com- posit ion and zonat.ion. Ecology, 40, 0!1-90.
Goreail, T. F. (19611~). l’roblcms of growth and calcium deposit,ion in reef corals. Rticlcnvoztr, 20, 32--39.
Goreau. 1‘. F. (1901b). On tho rolntiori of calcification to primary production in roof-building organisms. In “ Tho Biology of Hydra otc.” (H.M. Lenhoff and W. F. Loomis). Univorsity of Miami Press, 269-285.
Goreau, T. F., and Gorcau, N. I. (1969). The physiology of Bkeleton formation in corals. 11. Calcium deposition by hermatypic corals under various con- ditions in the reef. Biol. Bull. 117, 239-260.
Goreau, T. F., and Goreau, N. I. (198Oa). Distribution of labellod carbon in reef- building corals with and without Zooxanthellae. Science, 131, 668-609.
(:orom, ‘l’. F. (I!)RQa).

THE BIOLOGY OF CORAL REEFS 257
Goreau, T. F., and Goreau, N. I. (1960b). The physiology of skeleton formation in corals. 111. Calcification rate aa 8 function of colony weight and totd nitrogen content in the reef coral Manicinu areokrta (Linnww). Biol. Bull. 118, 419-429.
Goreau, T. F., and Goresu, N. I. (1960~). The physiology of ekeleton formation in corals. IV. On isotopic equilibrium exchanges of calcium between corallum and environment in living and dead reef-building corals. Biol BUU. 119, 416427.
Hamilton. E. L. (1956). Sunken lslanda of the Mid-Peoitic Mountains. Ma. geol. SOC. Anter. 64.
Hand, C. (1956). Are corals really herbivores? Ecology, 37, 384-385. Harry, R. R. (1963). Ichthyological field data of Raroia Atoll, Tuamotu Archi-
pelago. Atoll Res. Bull. no. 16. Hedgpeth, J. W. (1957). Concepts of marine ecology. Mem. geol. SOC. Am?. 07,
Vol. 1, pp. 29-52. Hertlein, L. G., and Emerson, W. I(. (1967). Additional notes on the invertebrate
fauna of Clipperton Island. Amer. MUB. Novit. no. 1869, 1-9. Hodgkin, N. M. (1962). Limestone boring by the Mytilid Lithophga. Tb
Veliger, 4, 123-129. Horridge, G. A. (1956). The msponsea of Heferozenio (Alcyonarie) to etimuhtion
and to some inorganic ions. J. exp. Biol. 33, 604-614. Horridge, G. A. (1957). The co-ordination of the protective retraction of cord
polyps. Phil. Tram. B. 240, 495-529. Hovasee, R. (1937). Lea zooxanthellee sont des dinoflagell&. C.R. A d . SCi.,
Pariu, 205, 1015-1018. Johnson, M. W. (1954). Prof. Pap.
U.S. geol. Sum. 260-E, 301-314. Kawaguti, S. (1937). Regeneration and phototropism in reef corals. P a h trop.
Kawaguti, S. (19418). On the physiology of reef cords. IV. -The growth Of Gonhatrea @era measured from numerical and areal increase of calyces. Pa2ao trop. biol. Stud, 2, 309-317.
Kewaguti, S. (1941b). On the physiology of reef corals. V. Tropism8 of C O ~
planulae, considered aa a factor of distribution of the wh. Palao Crop. biof. Stud. 2, 319-328.
Kawaguti, S. (1944a). On the physiology of reef corals. VII. Zooxanthelh of the reef corals in Gymnodinium sp., Dhoflagellata; its culture in viho. p u k ~ trop. biol. Stud. 2, 676-679.
Kawaguti. 8. (1944b). VI. Study on the pigments. Palao trop. biol. Stud. 2, 617-073.
Kawaguti, 5. (1944~) . Zooxanthellae as a factor of poeitive phototropism in those animals containing them. Palao trop. bwl. Stud. 2. 681-682.
Kawaguti, S. (1953). Ammonium metabolism of the coral reefs. BWZ. J. Okuyama
Gwaguti, S., and Sakumoto, D. (1948). The effect of light on the calcium
K o h , A. J., and Helfrich, P. (1957). Primary organic productivity O f 8 Hawaiian
Kuenen, P. H. (1950). " Marine Geology ", 668 pp. John Wiley, New York. m d , H. S. (1950). Recent reefs. Bull. Amer. Aeeoc. Petrol. G W . 34, 203-214.
Plankton of Northern Msrshall Ishda.
b i d . Stud. 1, pp. 209-216.
On the physiology of reef corals.
unit^. 1,171-176.
deposition of corals. Bull. oceanogr. Inat. Taizmn, 4, 66-70.
coral reef. fimnol. & Oceanogr. 2, 241-261.

258 C. M. YONOE
Ladd, H. S. (1961). Reef building. Science, 134, 703-716. Ladd, H. S., Ing&rson, E., Townsend, R. C., Russell, Martin, and Stephenson,
H. K. (1953). Drilling on Eniwetok Atoll, Marshall Islands. BuU. Amer. Assoc. Petrol. Cfeot. 37, 2267-2280.
Ladd, H. S., and Tracey, J. I. (1949). The problem of coral reefs. Sci. Monthly, 69, no. 6, 8 pp.
McLaughlin, J. J . &, and Zahl, P. A. (1967). Studies in marine biology. I T . In vitro cultlure of zooxanthellao. Proc. SOC. exp. Biol. 95, 116-120.
McLaughlin, J. J . A., and Zahl, P. A. (1959). Axenic zooxanthellae from various invortobrate hosts. Ann. N.Y. A d . Sci. 17, 65-72.
Manton, S. M. (1932). On the growth of the adult colony of PociUopora bulbow. Sci. Rep. Gr. Barrier Reef Exped. 1928-29, 3, 167-166.
Marsden, J. R. (1962). A coral-eating Polychaete. Nature, Lond. 193, 698. Moorhouso, F. W. (1936). The cyclone of 1934 and its effect on LOW Isles, with
special observations on Porites. Rep. at . Barrier Reef Comm. 4, 31-44. Morrison, J. P. E. (1964). Ecological notes on the mollusks and other anhale of
Raroia. Atoll Res. BuU. no. 34. Motoda, S. (1940a). A study of growth rate in the massive reef coral, aonklretz
aspera Verrill. Palm trop. biol. Stud. 2, 1-6. Motoda, S. (1940b). The environment and the life of the mwive reef coral,
Goniastrea aspera Verrill, inhabiting the reef flat in Palao. Palcro trop. biol.
Munk, W. H., and Sargent, M. C. (1964). Adjustment of Bikini Atoll to ocean waves. Prof. Pap. U.S. geol. Sum. 2 6 0 4 276-280.
Muscatine, L. (1961). Symbiosis in marine and freshwater coelenterates. In " The Biology of Hydra etc." (H.M. Lenhoff and W. F. Loomis). University of Miami Press, 266-268.
Muscatine, L., and Hand, C. (1958). Direct evidence for the transfer of meterials from symbiotic algae to the tissues of a coelenterate. Proc. nut. A d . Sci., Wash. 44, 1269-1263.
Newell, N. L). (1 956). Bahamian $latforrns. Bull. geol. Soc. Amcr. Spec. Pap. 62,
Nowcd, N. I). ( I U M I ) . (~oi~l~igii~ol t ~ i ~ i ~ i ~ i i t i i ~ i ~ ~ ~ i i i ~ i ~ r i f Itiiriiir~ (Kori 'l'rkt) At oil.
Newoll, N. 1). (lDG8). American coral Neae. i'roc. X V l h In$. C'onyr. Zool. Boat,
Newoll, N. I). (1969). Coral reefs. Nut. Hiat., N.Y. 1969, 119-131; 227-236. Newoll, N. D.. and Imbrie, J. (1966). Biogeological reconnaissance in the
Rimini area, Groat Bahamas Bank. Trana. N.Y. A d . Sci. Ser. 11, 18,3-14, Newell, N. D., Rigby, J. K., Whiteman, A. J., and Bradly, J. S. (1961). Shoal
water geology and environments, eaatern Andros Island, Bahamae. BuU.
Nicol, J. A, C. (1969). Digeation in sea anemones. J. Mar. biol. Aaa. U.K. 38,
Odum, H. T., and Odum, E. P. (1956). Trophic structure and productivity of 8
Ecol. Monogr. 25,
Pratt, E. M. (1905). The digestive organs of the Alcyonaria and their relation to
Stud. 2, 61-104.
Jjp. 30:s -316.
' I ' i i t~ i r io l i i Arc*tripoli~go. Ilull. Anivr. M U R . nut. Ifant. 109, 317-372.
111, Paper 43.
A m r . MU^. m t . Hiet., Vol. 91, pp. 1-30.
469-47 6.
windward coral reef community on Eniwetok Atoll. 291-320.
the mesogloeal cell plexus. Quart. J. micr. Sci. 49, 327-362.

THE BIOLOGY OF CORAL REEFS 259
Pringsheim, E. G. (1955). Die “ gelben Zellen ” der Koralle Cladocosa. Pubbl. Staz. zool. Napoli, 27, b-9.
Ranson, G. (1955). Observations sur lea principaux agents de la dissolution du calcaire sous-marine dans la zone c6tiere des ilea coralliennes de I’Archipel des Tuamotu. C.R. Aead. Sci., Par&, 240, 808-808.
Roushdy, H. M. (1962). Expansion of Akyonium digitalurn L. (Octocorallia) and its significance for the uptake of food. Vidensk. Medd. dansk mturh. Foren.
Roimhdy, H. M., and Hansen, V. Kr. (1960). Filtration of phytoplankton by the
Sargent, M. C., and Austin, T. S. (1949). Organic productivity of an atoll.
Sargent, M. C., and Austin, T. S. (1954). Biologiceconomy of coral reefs. Prof. Pap.
Schultz, L. P. (1948). The biology of Bikini atoll, with special reference to the
Searle, A. G. (1956). An illustrated key to Malayan hard corale. Malay. Nat. J.
Smith, F. W. W. (1948). “ Atlantic Roef Corals ”. University of Miami Press. Squires, D. F. (1959).
Kbh. 124, 409-420.
octocoral Abyonium digitutum L. Nature, Lond. 190, 649-660.
Trans. Amer. geophy8. Un. 30, 245-249.
U.S. gwl. SUW. 260-E, 263-300.
iishea. Rep. Smiulson Inatn. 1947, 301-316.
VOl. 11, pp. 1-28.
Results of the Puritan-American Museum of Natural History Expedition to Western Mexico. 7. Corals and coral reef8 in the Gulf of California. BuU. A w r . Mus. nut. Hiet. Vol. 118, pp. 367-432.
Stephens, G. C. (1960). Uptake of glucose from solution by the solitary coral, Fungia. Science, 131, 1532.
Stephenson, T. A. (1958). Coral reefs regarded as seashores. Proc. XVth Int. Congr. Zool. Soct. TTI, pp. 244-246.
Stephenson, T. A.. and Stephenson, A. (1960). Life between tidemarh in North America. I. The Florida Keys. J. ECOI!. 38, 364-402.
Stephenson, W., and Wells, J. W. (1966). The corals of Low Islea, Queeneland. August, 1964. Pap. Dept. 2001. Univ. Queenstand, 1, no. 4, 60 pp.
Stephenson, W., Endean, R., and Bennett, I. (1958). An ecological survey of the marine fauna of Low Isles, Queensland. Auet. J. Mar. Fruhw. Res. 9,
Stoddart, D. R. (19628). Three Caribbean atoll8 : Turneffo Islands, Lighthome
Stoddart, D. R. (I962b). Catastrophic storm effects on the Britiah Hondurm
Tracey, J. I., Ladd, H. S., and Hoffmeieter, J. E. (1 948). Reefs of Bikini, Merehal
Tracey, J. I., Cloud, P. E., and Emery, K. 0. (1965). Conspicuous featurea of
n m e y , J. I., Abbott, D. P., and Arnow, W. (1961). Natural hietory of Ifaluk
261-318.
Reef, and Gover’s Reef. British Honduras. Atoll Rcs. Bull. no. 87.
reefs and cays. Nature, Lond. 196, 612-616.
Islands. Bull. geol. SOC. Amer. 59, 861-878.
organic reefs. Atoll Rea. Bull. no. 40.
Atoll : physical environment. BuU. Biahop Mus. 222. Umbgrove, J. H. F. (1947). Coral reefs of the East Indies. BuU. gwl. Soc. Amur.
58. 729-778. Vaughan, T. W. (1919). Corals and the formation of coral reefs. Rep. Smithson.
Vaughan, T. W., and Wella, d. W. (1943). Ikviniorr r p f tho nllhorrjnra;, fumilh, UWJ I m h . 1917, 189-276.
genera of the Scloractinia. Spec. Pap. gml. h c . Amrr. no. 44.
I H

260 C. M. YONGE
Von Arx, W. S. (1954). Circulation systems of Bikini and Rondelap bgoona.
Wells, J. W. (1950). Reef corals from Cocos-Keeling atoll. BuU. RUBS Mus.
Wells, J. W. (1954). Recent corals of the Marshall Islands, Bikini and nearby
Wells, J. W. (1055). A survey of the distribution of reef coral genera in the Great
Wells, J. W. (1956). Scleract,inia. “ Treatise on Invertebrate Paleontology,”
Wells, J. W. (1957). Coral reefs. Mem. geol. SOC. Amer. 67, Vol. 1, pp. 009-631. Wiens, H. J. (1962). “ Atoll Environment and Ecology ”. Yale University P m ,
Yongo, C. M. (1930-31). Studies on the physiology of cords. 1-111. Sci. Rep.
Yonge, C . M. (1935a). Studies on the biology of Tortugee corals. I. Observations
Yonge, C. M. (1935b). Studies on the biology of Tortugaa corals. 11. Variation in
Yonge, C. M. (1937). Studies on the biology of Tortugm corals. 111. The effect
Yonge, C. M. (1940). The biology of reef-building corals. Sci. Rep. Gr. Barrier
Yonge, C. M. (1944). Experimental analysis of the association between inverte-
Yonge, C. M. (1951). The form of coral reefs. Endeavour, 10, 136-144. Yonge, C. M. (1966).
Prof. Pap. U.S. geol. Surv. Vol. 260-E, pp. 205-273.
22, 29-52.
atolls. U.S..geoE. Surv. Pap. Vol. 260-1, pp. 385-486.
Barrier Reef region.‘ Rep. Ur. Barrier Reef Comm. 6, Pt. 2, pp. 1-9.
Part, F, pp. 328-444.
New Haven and London, 532 pp.
Gr. Barrier Reef Exped. 1928-29, 1, 13-91.
on Maeandra areolata Linn. Pap. Tortugaa Lab. 29, 186-198.
the Genus Sideraatrea. Pap. Tortugaa Lab. 29, 199-208.
of mucus on oxygen consumption. Pap, Tort- Lab. 31, 207-214.
Reef Exped. 1928-29, 1, 363-391.
brates and unicellular algae. Bid . Rev. 19, 68-80.
Adaptation to rock boring in BotuZu and Lithophagu (Lamellibranchia, Mytilidae) with a discussion on the evolution of this habit. Quart. 2. micr. Sci. 96, 383-410.
Yonge, C. M. (1957). Symbiosis. Mem. gcot. SOC. Amer. 67, 1, 429-442. Yonge, C. M. (1958a).
Yonge, C. M. (1958b).
Darwin and coral reefs. I n “ A Century of Darwin ” (S. A. Barnott). Hcinemann, I,ondon, pp. 246-206.
I n “ Perspectives in Marine I3iology ” (Buzzati-Travomo). Univoraity of California, pp. 117-135.
Yonge, C. M., and Nicholls, A. G. (19314. Studiea on the phyaiolol~y of corah. IV. The structure, distribution and physiology of the eooxsnthellae. Sci . Rep. Gr. Barrier Reef Exped. 1928-29, 1, 135-170.
Yonge, C. M., and Nicholls, A. G . (1931b). 8tudim on tho phyfiiology I J f coruln. V. The effect of Htarvat,ivn in light and in iltlrknc?~n on L h * y reliitiw~hip between corals and zooxanthallwt. Sci. Jtep. Ur. Iimriw ltmf KrpwL. lO2Y 29,
Yongo. C. M., Yongo, M . .I., ant1 Nicholin, A. (1. (1032). Htirdictrc on t 3 h phyRiology of c o r h V I. ‘I’ho r ~ ~ l i i t ~ i ~ i r i ~ t i i ~ ~ lmtwoon ronpiration in (:orals and the pro- cirwt.im of oxygoii by t~tloir zoc,siLiit.tiollir. Sci. Rep. Ur. Ilarrier Reef Ezpcd. 1112N-21), 1, 213 261.
Zahl, 1’. A.. arid Mc~Aa~~gtil i~i . . I . ,J . A. (1067). Isolstion and cultivation of zooxtirit.Ji~~1Ii~1~. Ntr trm, Loud. 180, 199 -200.
z~hl. 1’. A.. tiritl MoIAaughli~i. J . J. A. (1959). Studies in marine biology. IV. On t,he roln of algal colls in t hc t insucs of marine invertebrates. J . Protozoal.
1hli)gy and phynioloyy of rw:f-bi~ilrhy coral&
1, 177-211.
6, 344-352.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS
Depart men
J. H. S . BUXTER Marine Laborato y, Aberdeen, Scotland
AND
F. G . T. HOLTJDAY of Natural History, Aberdeen University, Scotland
I. Introduction . . .. . . . . .. .. .. .. . . A. General . . .. .. .. .. .. .. .. .. B. Characteristics of Clupeida . . .. .. .. .. ..
11. The Gametes .. . . .. .. .. .. .. .. .. A. Basic Structure and Composition . . .. .. .. .. B. Tolerance to External Conditions . . .. .. .. .. C. Viability of the Gametea and Artificial Storage . . .. .. D. Fertilization . . .. .. . * * . .. .. ' . E. Parthenogenesis. . .. .. .. . . .. .. ..
111. The Developing Egg . . .. .. .. .. .. .. .. A. Embryology .. .. .. .. .. .. .. .. B. Effect of Temperature on Rate of Development . . .. ..
D. Effect of Temperature, Pressure and CO, 011 Eggs. . .. .. E. Egg Mortality , . .. .. .. . . . . .. ..
IV. The Larva . . .. .. .. . . . . .. .. .. .. A. Development of Organ SyHtenls . . .. .. a . .. B. Feeding of Larvae . . .. . . . . . . .. . .
D. Rearingof Larvae . . .. . . .. .. .. .. E. Farming . . .. .. .. .. .. .. .. .. F. Mortality of Larvae . . .. . . .. .. .. . .
H. Salinity Tolerance and Osmo-regulatioii of Larvae. . .. . . I. Oxygen Uptake.. .. .. . . .. . . .. . . J. Dermal Receptors . . .. . . .. .. . . . . L. hcomotory Behaviour and Ltheotropic ~wponne . . .. . . M. Vertical Migration of Larvae . . .. .. .. . . . . N. Response of Larvae to Light . . . . . . . . . . , I
v. Metamorphosis . . . . .. .. . . . . . . .. . I
vI. Poet-metamorphic Stages . . .. . . . . .. . . . . A. Establlrrhment . . . . . . .. .. . . * . . . B. FwdinK . . .. .. C. The Alimentary System . . . . . . . . . . . . D. Fat Storage and Metabolism , . .. .. .. .. . .
C. Salinity Tolerance and Oamo-regulation of the Developing Egg . .
C. Growth of Larvae .. .. .. .. .. .. ..
G. Predationon Larvae .. .. . . . . .. .. . .
K. Temperature, pH, Oxygen, Presauro and Light S(I Limiting FZW~KJW
. . .. . . .. . . . .
261
262 262 264 264 264 266 261 201 268 268 268 269 21 1 212 213 214 814 215 211 218 280 280 283 283
281 287 2He
28 1 504 284 284 ZYG 207 2YO
2 n ~
m

262
E. F. G. H. I. J. K. L. M.
N. 0. P. Q. R. S. T. U. V.
W. x. Y. 2.
J. H. 5. BLAXTER AXD F. 0. T. HOLLIDAY
.. .. .. .. .. .. Growth .. .. .. Natural Mortality . . .. .. .. .. . . .. Composition of the Tisaues . . .. .. .. .. .. Salinity Tolerance and Osmo-regulation . . .. .. . . Temperature, Oxygen, CO, and H,S aa Limiting Factors. . . . The Brain . . .. .. .. .. . . . . .. Vision . . .. .. .. .. .. .. .. .. Olfaction. . .. .. The Labyrinth, Hearing, theEffeot of Sound, and Sound Pro-
duction . . .. .. .. .. .. .. .. Buoyancy and Equilibrium . . .. .. .. . . .. swimming . . .. .. .. .. .. .. .. Activity . . .. .. .. .. .. .. .. .. Shoaling . . .. .. .. .. .. .. .. .. Migrations . . .. .. .. * . . . .. .. Vertical Migration . . .. .. .. .. .. .. Effect of the Moon . . . . . . . . . . * . .. Attraction to Artificial Light8 .. .. . . . . .. Reaction to Nets and Other Obstacles . . .. .. . . Learning. . .. .. .. .. .. ,. .. . . Maturation of the Gonads . . .. .. .. .. ..
. . .. .. .. . . ..
302 305 307 309 312 313 314 316
316 320 322 325 326 332 342 347
36 1 356 366
348
Spawning .. .. .. . . .. .. . . . . 364 Racial Characters, the Genotype and the Environment . . .. 361
VII. Conclusions . . .. . . .. . . .. .. . . .. 370 VIII. References . . .. .. .. .. .. .. .. .. 372
I. INTRODUCTION A. General
Of all the clupeids the herring, Clupea harengw, has been most extensively studied. Perhaps i t has been more the subject of research than any other fish species, partly due to its commercial importance and partly to the need for so much work before even tho ~ ~ s c r i t i a l ~ of i t H complex biology could be exposed ; tliw the herring i H bound to take u p tho main H ~ C O in this review. Although most of the retlearch has been conconic~d with the racial question and population studies, these aspects are only mentioncd where experimental work has assisted such research. Therefore while natural mortality due to disease and predation, and fecundity in relation to reproductive biology, will be considered, fishing mortality and its estimation, and fecundity as a means of distinguishing races and for estimating stock size, will not.
For a general review of clupeid biology reference may be made to a recent bibliography (Scattergood, 1957), to a series of translations from Russian (Scattergood, 1959), to the monograph on tho Clupeidac by Svetovidov (1952) and to general books Ruch aa those of Hodgson (1067) and Nikolsky (1957). Two recent Hpecial scientific meetirign of t h International Council for t h o 12xploration of tho Soa in (hpnhngun (J.C.N.S.) in 1 f W I and 1901 c:ovcmrtl much huhaviour and physiological

THE BEHAVIODR AND PHYSIOLOGY OF RERRINQ AND OTHER OLUPEIDS 263
work, some of which is described here. For a general review on fish physiology and behaviour, reference should be made to Bull (1962), who described techniques used for behaviour studies aa well as giving estimates of the sensitivity of teleosts to environmental factors such as temperature, salinity and pH. He also (Bull, 1961) described the role of ethology in oceanographic research. Brown (1967) edited a valuable publication on the comparative behaviour and physiology of fish in general, much of which is applicable to clupeids.
The main species covered in this review are given below : Clwpea harengw L.-Atlantic herring (the Baltic herring is referred
to by the Russians aa Clupea harengus membraa or salaka) Clupea p a l h i i Val.-Pacific herring (considered by Russian workers
to be a sub-species of harengw) Sardim pilchardus ( Wa1baum)-European sardine or pilchard Spattus sprattus (L.)-Sprat Surdinella aurita Val.-Gilt sardine Surdimps cuerulea (Girard)-Californian sardine (the taxonomic
status of the Pacific sardines is discussed by Man, 1957) Sardinops metmwsticta (SchIegel)-Japanese sardine Brevoortia tyrannwr (Latrobe)-Menhaden Alosa spp.-Shads Pomolobus pseudoharengw (Wilson)-Alewife Caspialosa spp.-Caspian and Black Sea herrings Clupeonella spp.4aspian sprat or kilka Some other species of clupeids and engraulids are also mentioned.
In this review Clupea harengw will usually be referred to as ‘* herring ”, all other species being given their Latin names.
The behaviour and physiology of these fish have been studied, not only from experiments in aquaria, but also from more indirect observa- tions at sea. Inferences have also been drawn from the extensive anatomical studies made on clupeids. Because of the commercial value of clupeids some importance has been given to their behaviour and physiology in relation to fishing gear.
The review is not arranged solely on the basis of organ systems; where possible i t follows the life history of the fish starting with fertilization, the egg and larval* stages and ending with maturation and spawning.
Due to the dificulty of covering the literature, particularly Ruesias and Japanese, i t is regretted if any relevant papers have been omitted. However, the authors would like to acknowledge the help of the
The word ‘‘ larva ” will be wed to include Stcrges from hatohing to metamorphoais.

264 J. H. S. BLAXTER AND F. 0. T. HOLLIDAY
following in tracing literature: Dr. Vivien Brawn (Vancouver), Mr. J. Jakobsson (Reykjavik), Dr. R. Lasker (La Jolla), Dr. Radosna Muiini6 (Split), Prof. G. Nikolsky (Moscow), Dr. P. A. Orkin (Aberdeen) and Dr. C. J. Sindermann (Boothbay Harbor). Dr. Z. Kabata (Aberdeen) has assisted greatly in translating Russian papers.
Thanks are due to Mr. B. B. I’arrish (Abcrdeen) and Dr. G. Hempel (Hamburg) who read and criticized this paper in manuscript, and to Dr. G. Krefft (Hamburg) for his advice on nomenclature.
B. Characteristics of Clupeids The Clupeidae are to be found in all habitats from fresh water,
txg. some Pomolobus and Harengula species, to sea water. Many of them are anadronious such as Alosa, Caspiolosa and Pornolobus. The family contains species which lay demersal eggs offshore (herring) , demersal eggs in shallow water (Clupea p a l h i i ) and pelagic eggs (Surdinope).
Hatching takes place in a few days (the eggs being small) and there is little yolk available before the larva must find an external source of food. These larvae are usually very small and transparent, meta- morphosing after some months to the adult form. The larvae and juveniles of marine species are often found on shallow inshore nursery grounds.
Thc fccundity is low compared with many teleosts, but the numbers of individuals arc high. The adults of most species adopt a pelagic and shoaling habit and often make extensive migrations for feeding and spawning. They arc mostly plankton and therefore seasonal feeders, laying down fat reserves which allow them to survive when food is scarce. Lacking much in the way of defence mechaniams, except pra- tective colouration, speed and shoaling, and being present in such large numbers, most clupeids are heavily preyed upon by other fish, mrtm- mals and birds.
11. THE GAMETES A. Basic structure and composition
1. The sperm The morphology of herring sperm has been studied by Retzius (1906)
and Ballowitz (1915) who described the successive stages in the trana- formation from spermatid to spermatozoon by the development of the tail piece. It is unlikely that the sperm of other clupeid8 differs sig- nificantly from that of the herring.
The head is compoRed almoHt cntircly of nuclear material and In approximately 1p in diamotor; R short middlo picco i~ followod by a
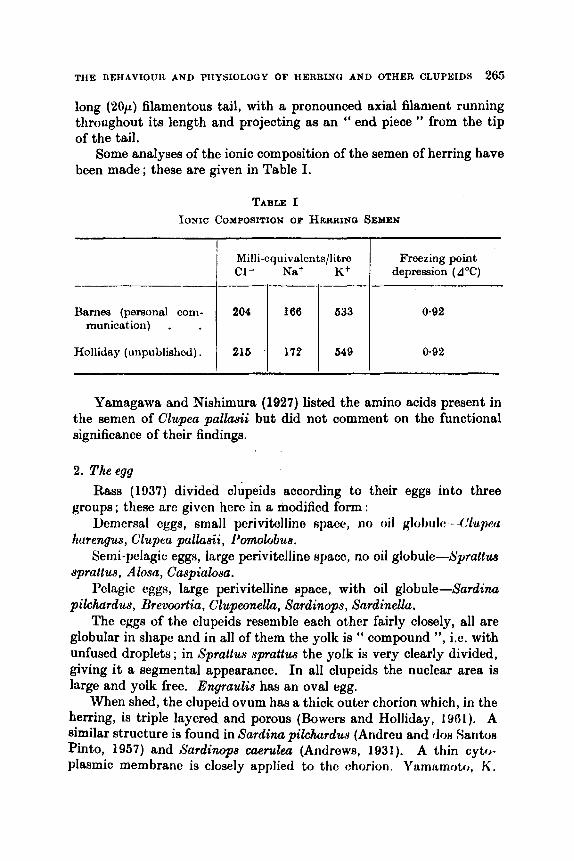
THE IIEHAVIOUlt AND PlIYYIOLOOY OF HERRING AND OTHER CLUPEIDS 265
Milli-cquivalents/litre C1- Na+ K+
Barnes (personal com- 204 166 633 munication) .
Holliday (unpublishod) . 215 112 549
long (20p) filamentous tail, with a pronounced axial filament running throughout its length and projecting as an " end piece " from the tip of the tail.
Some analyses of the ionic composition of the semen of herring have been made ; these are given in Table I.
Freezing point depression ( d "C)
0.92
0.92
Yamagawa and Nishimura (1927) listed the amino acids present in the semen of Clupea pallasii but did not comment on the functional significance of their findings.
2. The egg Rass (1937) divided clupeids according to their eggs into three
groups ; these are givcn here in a modified form : Dcmersal eggs, small perivitcllino upacc, no oil globule - 4,'lupea
hurengus, Clupeu pullusii, Pomolobucv. Serni-pelagic eggs, large perivitelline Npace, no oil globule-Sprullus
spratt us, Alosa, Caspiaha. Pelagic eggs, large perivitelline space, with oil globule-Sardinu
pilchardus, Brevoortia, Clupeonellu, Sardinops, Sardinellu. The eggs of the clupcids resemble each other fairly closely, all are
globular in shape and in all of them the yolk is " compound ", i.e. with unfused droplets ; in Sprattus sprattus the yolk is very clearly divided, giving i t a segmental appearance. In all clupeids the nuclear area is large and yolk free. Engraulis has an oval egg.
When shed, the clupeid ovum has a thick outer chorion which, in the herring, is triple layered and porous (Bowers and Holliday, 1 Y f l 1 ) . A similar structure is found in Sardina pilchardua (Andreu and d o u , 9 antos Pinto, 1957) and Sardinops caerulea (Andrews, 1931). A thin cyto- plasmic membrane is closely applied to thc chorion. Yamnmoto, K.

200 J. H. S . BLAXTER AND F. 0. T. HOLLIDAY
(lS56) described the position and structure of the micropyle, penetrating all three layers of the chorion in Clupea p a l h i i .
Few analyses have been made on the composition of the clupeid egg. The peripheral cytoplasm in the egg of Clupea p a l h i i contains a number of cortical alveoli with neutral muco-polysaccharide contents (KanoX, 1953 ; Yamamoto, 1956); the same is true of the herring egg (Holliday, unpublished). The alveoli contain no lipids ; the main yolk particles are separate from these, and give only a weak response to the P.A.S. test. Hempel (personal communication) found that the mean fat content of herring eggs was 1.32% of the wet weight and probably about 8.3% of the dry weight, in the southern North Sea in 1961. Lasker (1962) analysed the egg of Surdimp caerulea and found that over 70% of the dry weight was protein ; lipids constituted 13% of the dry weight and the water content of the yolk averaged 91.2%.
B. Tolerance to external conditions
In the body of the parent herring the gametes are maintained in a relatively favourable environment. Herring eggs are slightly hypo- tonic to the body fluids of the female parent; Holliday (unpublished) obtained freezing point values of -0.75"C for ripe eggs in the gonads of female herring with blood freezing points of -0-92"C. Barnes (personal communication) and Kolliday (unpublished) both obtained values of -0.92"C for the sperm of herring, i.e. the malc gametes were isotonic with the blood of the parents.
On Being rcloased into the water the gametes often sxperience a vast change in concentration of the external medium. Htmmg ofitnotic forcen are often exerted on thcm, and, with their Itirgf: H I J ~ ~ ~ W : t i r w i / v o l t ~ r ~ ~ ~ ~ ratios, one might imagine them to be enpcaially HllSC~!JJt i tJh t,o oomotio death. On the contrary, they appear very tolerant (see Section 111, C).
It is difficult to determine just what mechanisms are employed by the qmmatozoa to survive these osmotic forces. They could survive by being impermeable or by being freely permeable, thereby changing drastically in composition but avoiding any osmotic gradient ; alter- natively there may be some non-electrolyte present which is capable of setting up a force opposing the osmotic forces of the environment.
Much more has recently been found out about regulation in the egg, and this is discussed in detail in a later section. When released into the water, whether fertilized or not, the herring egg rapidly approacheu isotonicity with the medium. The osmotic forces are t h w cqualizcd passively. True regulation depends on the suhequcrit emhryonic development (see Section 111, C).

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 267
C. ViabiZity of the gam&es and artifxiab storage
Herring sperm remains fertile for at least 3 hr in sea water at 8°C (Blaxter, unpublished). Yanagimachi (1953) found that the sperm of Clupea p a l h i i remained fertile to a limited extent after 12 hr in sea water at 8-10°C. In Ringer solution or in Iower salinities some fertility was retained after 24 hr. However, Stroganov (1938) showed that the sperm of Caspialosa volgensis lived for 1 min or less in fresh water and that sperm from mixed males survived less well than sperm from one male ; of the eggs 60% could still be fertilized after about 30 hr.
Gamete storage has been developed mainly in salmonids as a pisci- cultural technique. In herring, short-term storage haa been used (Blaxter, 1955) for transporting gametes from where the parents are caught to the laboratory for controlled rearing studies, and long-term storage for making cross fertilizations between herring races spawning at different seasons. For short-term storage, different diluents such aa phosphate and borate buffers with and without chicken egg yolk were used, but the most satisfactory results were obtained by storing whole gonads dry at about 4°C. While fertilization of eggs may be obtained even after 6 to 7 days of dry storage, a high hatching rate is only to be expected after periods not exceeding 36 hr storage. Long-term storage has been achieved successfully with sperm, but not with eggs, by freezing in glycerol and diluted sea water to a temperature of -79°C. At least 6 months later the sperm were viable and crosses were made between spring and autumn spawning herring (Blaxter, 1957) and between different races of spring spawners (Blaxter and Hempel, 1961).
Storage of gametes within the dead parent’s body does not seem to be successful and the fertilization rate was very low after I8 hr in herring. Stroganov (1938) found in Cuqklosa voZgemia that the sperm started to die 10-60 min after the death of the parent, if the testis wm left in situ.
D. Fertilization
Fertilization in Clupea pallaaii was described by Yamamoto (1958) as being normally monospermic. He showed that immediately following sperm entry the fertilization membrane, which was a protein structure, withdrew from the chorion thus forming a perivitelline space. Into this space are extruded the droplets of the cortical alveoli (Volodin, 1956). Yanagimachi and Kanoh (1953) found that the calcium ion concentra- tion was critical for successful sperm entry. Yanagimachi (1957) described a factor bound to the egg membranes of C l u p p a W i that caused quiescent sperm to become active if brought near it. This

268 J. H. 9. BLAXTER AND F. 0. T. HOLLJDAY
factor may be contained in the secretion of the oviduct (described in Section VI, X). Kryzhanovsky’s observation (see Galkina, 1957) that the eggs of Clupea pallasii seemed to be fertilized by immobile sperm may have been due to the sperm in this instance not yet having been activated.
The present authors and Hempel showed that when released from the body of the female the egg of the herring had an osmotic con- centration ( A 0.75) equivalent to about 12ym sea water. The process of fertilization tends to accelerate the change to isotonicity (discussed previously). Permeability is increased and i t is probable that the mucopolysaccharide contents of the cortical alveoli are responsible for drawing the outside medium through the freely permeable chorion by some process of imbibition.
E. Parthenogenesis Volodin (1956) described an instance of parthenogenesis in the
herring. Nearly 60% of a number of unfertilized eggs, when placed in sea water, extruded droplets into the perivitelline space. A small proportion of these “ activated ” eggs stopped developing after bipolar differentiation had occurred, the remainder beginning cleavage. In 5% of the eggs this waa abnormal, the cleavage furrows being in no definite order and blastomeres of different sizes being produced ; these eggs died, usually before reaching gastrulation. The remaining eggs followed a pattern of apparently normal development, producing active embryos. Unfortunately observations were not made aa far aa hatching.
Just what the factor is that ‘‘ activates ’’ an unfertilized egg is not clear. It has been suggested that certain differences in osmotic pressure between the egg and the medium may stimulate diviHion (Morgan, 1927). Unfortunately Volodin did not give the salinity of the water in which this development took place ; apparently i t could have been as low as 4-9%,.
Galkina (1957) described parthenogenesis in C’Zupea palkz&i and found that it occurred to a greater extent in the lower Halinitir:fi. Development proceeded as far as hatching but thr: larvltr: wcrc ~ I P normal.
111. THE DEVELOPING Eao A. Embryotogy
Thc embryonic development of the herring has been studied and described in some detail (Kupffer, 1878; Brook, 1885, 1886; Prince, 1907 ; Volodin, 1956 ; Toom, 1958) and also of Clupea pallnsii (Hamana,

THE BEHAVIOUR AND PHYSIOLOQY OF HERRING AND OTHER CLUPEIDS 269
1936; McMynn and Hoar, 1953; Outram, 1955). The rate of develop- ment is closely linked with temperature bee Section 111, B); for example at 14% gastrulation begins 22 hr after fertilization, whereas at 11-S"C the eggs after 22 hr are still in the blastula stage. In the following account no references are made to either time or temperature scales.
Immediately after fertilization there is a rearrangement of the cytoplasm and yolk. The blastula resulting from cleavage forms a cap of small cells at the animal pole. Gastrulation then occurs. The cells of the blastula lift slightly from the surface of the yolk, retaining contact with it by a cellular " root " in the centre. Intucking occurs and first a dorsal and then a ventral lip of the blastopore is formed. The inwardly migrating cells from the lips meet to form a two-layered embryo, and growth of the peripheral cells over the surface of the yolk takes place. The blastopore closes when the migrating cells meet.
A head region is differentiated at the dorsal lip of the blastopore. Mesoderm cells migrate to form a layer between the ectoderm and endoderm, and segmentation then occurs. The embryo is about 2 mm long at this stage. As segmentation continues the nervous system is established and the rudiments of the sense organs (optic cups, olfactory capsules and otic vesicles) are developed. The head and trunk segments remain attached to the yolk, but the tail soon becomes free. The embryo shows occasional movements. The eyes are formed but lack pigment ; two otoliths are present in the otic vesicles. The gut tube is formed and the heart commences beating (about 40 beats per min at 15°C). A dorsal aorta is present and a sub-intestinal vein; thc only corpuscular elcmcntfi viAhlc in tho Mood arc a form of leucocytc!. 'I'hn ( y s then hctaornc! pigrncwtcd arid haLct1ing glunthi q y w a r on tho ht:wl. l'hc gut is open posteriorly, but although tho Rtornodaeurn is viaiblct, the gut is still closed antcriorly.
According to McMynn and Hoar (1 053) hatching of Clupcu p d l r ~ i i was nearly always head first through a softened part of the chorion.
The development of the herring is not necessarily typical for clupeids. Many of the species ( A h a , Clupeonella, Cmpiabsa) have a much shorter incubation time and Yerceva (1939) and Lasker (1962) have found that Cmpialosa caspia and Surdinope cuerulea larvae respectively hatch with unpigmented eyes.
B. Effect of temperature on rate of development Some of the published work is given in Table 11. From the equation of Hela and Laevastu for herring the biological
zero (the theoretical temperature when incubation takes infinite tirnc)

TABLE I1
EBBECT OF TEMPERATURE ON RATE OF DEVELOPMENT AND TIME TO HATCHING
Author
Hela and
(1962)using J e a n ' s (1966) and other data
Blaxter
LReVRStU
(1966)
Toom (1968)
Ryder (1882)
Perceva (1939)
Ahlstrom (1943)
Miller (1962)
Ahlstrom (1954)
Murphy (1961)
It0 (1958)
MnMynn and Hoar (1958)
Ou trim i ( 1965)
Mot octn and Hirano (1961)
Nikolsky (1957)
Species ~
Herring
Herring
Homing
A loaa
Caspialoaa
Sardinopa caerulea
cagpk
sardinop8 caeruku
Sardinopa caerulea
sardinop8 caerulea
Srzrdinnpa wr hiti onlict4
Clupea pl lapi i
Clupea plllkzsii Clupea pallaeii
Caapiuloaa 7lupeonella
12.8-19'0
11-23 normal range)
18-4-21.4
13.6-1 6.0
16.8
12.6-18.0
8.6
4.4-10.7
3.1-7.8
19-22.7 17.9-19.8
Time 1) Days
7 7-4- 2
16-3
24-14
3
2.3
4-2
10-11
40-1 1
22.9-16
c. 1.75 e . 1.25
Equation Remarks
H a t c h e d in the sea
Estimated f r o m samples at sea
Used Ahl- s t r o m ' s r e s u l t s and grttp. tlWJ I h W l
I1 J L t J 11
d I1 u t 0
IIctltt and I.aovantu (1 902)
Summary of J a p a - nese work
Summary
Aian work of RUS-

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 271
is -m°C, but these authors give a time of 49 days at 0°C. Soleim (1942) found - 1-2°C for Atlanto-Scandian herring, Blaxter (1 956) -1.34"C for northern North Sea herring, and Blaxter and Hempel (1961) about 0°C for German coastal herring.
The product of temperature (using the biological zero) and time (" day degrees ") has been used as a means of comparing stages of development in larvae reared at different temperatures. This concept is unsatisfactory in some ways because of the difficulty in identifying equal stages of development from which to calculate the initial biological zero. Also as the temperature changes, so may the relative growth of different parts of the body. Even the time to hatching itself may vary aa a result of environmental conditions other than temperature. Barlow (1961) has recently suggested that the time to reach a given developmental stage should be plotted against absolute temperature.
Ahlstrom (1943) found the Arrhenius relationship for hatching of Sardinops caerulea to bc about 22,500. Blaxter (1966) found values ranging from 20,800 to 9,500 in herring, with Qlo values ranging from 6.6 to 2.0.
C. Salinity tolerance and osmo-regulation of the developing egg
1. Salinity tolerance
Ford (1929) found that herring eggs could be successfully fertilized and incubated in salinities down to 4-8%,. Kryzhanovsky (1956) reported normal fertilization of Baltic herring eggs in salinities as high as 25%, (normally i t is 4-5%,,), although later development was ab- normal. Brandhorst (1 959) reported RucceHsful spawning in th: Kiel Canal at a salinity of 5%,. Holliday and Blaxter (1961) meawrc:d the changes taking place in the embryonic tissues of herring rear& in salinities from 5-55%,; they found a 100% fertilization in salinitiw from 25-55%,, becoming 70% in 12%, and 5%,. Over 50% of the eggs hatched in salinities from 10-45%,. McMynn and Hoar (1953) showed that the eggs of Clupea pallasii would tolerate salinities at lea& aR low as 6%,, and in a larger number of experiments found that t h o optimurn salinity for development and hatching lay betwecn 11.53 and I&Y4%,,. Galkina (1957) found in Clupea pallavii that percentage fertilizutiori Ody fell off below salinities of 6%,.
2. Osmotic regulation
After fertilization the herring egg is almout isotonic with tho water into which it has been shed, although it never reaches isotonicity with

272 J. H. 8. BLAXTER AND F. Q. T. HOLLIDAY
very low (<6%,) salinities. The increased water content of eggs developing in these low salinities is reflected by the increase in size of the egg (Holliday and Blaxter, 1961). This is also found in Clupea p a l h i i (Galkina, 1957). Probably the mechanical pressure exerted by the chorion limits water inflow ; this is certainly the case in the eggs of Surdimps caerulea (Laaker and Theilacker, 1962a). There is inflow of water into both the perivitelline space and the embryo. The chorion appears to be freely permeable, although there may be a binding of some ions by the colloid in the perivitelline space. Galkina (1957) found that the chorion in Clupea pallasii waa strongest in salinities 1O-28%, .
Unpublished work by the authors and Hempel showed that, during the early stages of development, the herring egg had little capacity to regulate the osmotic concentration of the yolk, although the individual cells of the blastula were apparently regulating. It was only after gastrulation was completed that the whole egg was regulated to an osmotic concentration equivalent to about one-third of the external mcdiuni in salinities above 12%,, and, in lower salinities to a value equivalent to 12%, (4=0.72). At this stage the whole of the exposed surface of the yolk was enclosed within the endo- and ectodermal layers. It seems probable that the ability to regulate is a property of these cells. Lasker and Theilacker (1962s) showed that the degree of regulation maintained by embryos of Sardimps caerulea was similar to that in the herring (to about one-third of the external medium).
D. Effect of temperature, pressure and CO, on eggs* The effect of temperature on the incubation time for clupeid eggu is
discussed in Section 111, B. Meyer (187%) fwind that normal tfcwlop. ment took place from 22Oto l”C, but death occurred at - O.R‘”f!. ‘I’cmrn (1958) found that eggs of Baltic herring would hatch at temperatures as high as 18.5OC and Blaxter (1956) that the optimum temperatare varied in different races.
Ewart (1886) found that eggs lowered to a depth of ahout 200 rn (21 atm) and left for some days hatched normally in the s q u ~ r i u m after being brought to the surface.
Kelley (1946) studied the effcct of carbon dioxide tension on the eggs of Clupea p a l h i i and found that they could tolcratc pH valucs ranging from 5.12-9.00 (the average ranging from 6.68-8.68), low pR values retarding development. The eggs would tolerate CO, values ranging from 0.00-67.76 ppm, the average values ranging from 0.088- 27.06 ppm.
Polikarpov and Ivanov (Voproay Ikhliol. 1, 683-9, 1961) found that exposure to radiation (2 x 10-0 curies/litre) from @OSr and *OY led to abnormalities In developing Ewaulis .

THE BEHAVIOUR AND FHYSIOLOQY OF IIERRMQ AND OTHER CLUPEIDS 273
E. Egg mortality 1. Egg density
Lea (1030) and Runnstr~m (1941b) showed that overcrowding of herring eggs on the sea bed off Norway resulted in a high mortality. Eggs in layers 0.6-2.5 cm thick showed mortalities of 4340%. Cleaver and Franett (no date) found a similar occurrence in eggs of Clupea p a h i deposited in large clumps inshore. However, Parrish et al. (1959), who succeeded in taking egg samples by grab on the Scottish west coast, found no evidence of a differential mortality in eggs four to eight layers thick (i.e. 0-5-1.0 cm). McMynn and Hoar (1053) and Blaxter (1950, 1962) showed that tho succcssful hatching of eggs of Clupca pallasii and Scottish herring respectively in aquaria depended to some extent on the density, crowding resulting in a low percentage hatch. In particular, the former authors found what appeared to be an intermediate optimum density for the best hatching ; above and below this percentage hatch was reduced.
2. Fungal and bacterial attack Toom (1958) found that eggs of Baltic herring were liable to attack
by fungi when kept in the sea away from the spawning ground, although on the spawning ground they were not affected. In rearing experiments on herring fungal attack may reduce percentage hatching considerably, especially when the incubation period is long. Anon. (1952) reported on the high rate of infection by bacteria of some Sardinope caerulea eggs, and showed that this could be checked in aquaria by antibioticn.
3. Predation Reports of fish wi th their &omactis gorgc:d with twriiig 1:gp arc
to be foirntl iri tho litcrtitiiro ; iridecd concc:ritriitiotiH of hetlilook anti coal fish were one of thc fir& means of locnting hcrrirlg np~wnirrg grounds (e.g. Bowman, 1923). Even the stomach8 of qmwriing hcrririg sometimes contain eggs. Parrish et al. (1959) found on a herring spawning ground with a heavy population of ophiuroids that the ophi- uroids were inversely distributed in relation to the eggs. The eggs of Clupeu pullasii are deposited inshore (e.g. Rounsefell, 1930) and are liable to be eaten by birds a t lo& tide (Cleaver and Franett, no date). Presumably the hazards facing demersal and pelagic eggs are some- what different, the former being concentrated and available in large quantities, the latter being dispersed, at least some time after spawning, and yet liable to stranding.

274 J. H. 9. BLAXTER AND F. Q. T. HOLLIDAY
IV. THE LARVA
A. Development of organ system
The anatomy and development of the skeleton and organ systems after hatching is best known in herring. At hatching the larvae vary in length from 5-9 mm, the yolk being absorbed at 8-12 mm. The size of the larva and its yolk supply and the rate of absorption of yolk depend on the original egg size and race of the parents (Toom, 1958 ; Blaxter and Hempel, 1963). The larvae of Sprattus aprattus, Sardina pilchardus (Lebour, 1921) and h'urdinopa caerulea (Miller, 1952 ; Arthur, 1966), however, hatch a t 3 4 mm and have used up their yolk at 6-6 mm.
In herring larvae only a primordial fin is present at hatching in addition to the pectorals ; the eyes are large, the mouth is non-functional and the gut is straight. However, the jaw soon begins to grow forward. At first the lower jaw does not articulate, being suspended by an elastic ligament (Fluchter, 1963). Harder (1952) described the development of the alimentary system, showing that the pyloric oaecae and caecum gradually started to develop at about 25 mm, the liver already being present. At this stage, too, the lateral musculature began to invest the gut. Harder (1 960) also gave a comparative description of the develop- ment of the gut of many clupeids and related the gut proportions to the cross-sectional area of the body and body mass.
Bamford (1941) studied the lateral line in herring which was com- pletely developed a t 50 mm length. Sensory papillae are found in the head region (de Kock, unpublished) of newly hatched herring larvae, and over the body of Sprattus sprattus and Lsardina pilchrdzcs (Lebour, 1921). Holstvoogd (1957) described a well developed pronephros and archinephric duct in herring larvae 10 mm long ; this is not present at hatching (Holliday and Blaxter, 1961). Maier and Scheuring (1923) quoted work on the development of the swim bladder and its mn- nection with the labyrinth. At 7 mm the larvae had a much shortened swim bladder rudiment and at 14 mm the connection with the laby- rinth had already been established. The gill arches are present at hatching but the gills are not functional until about 20 mm (Hazder, 1954). The blood contains a form of leucocyte but no erythrocytes.
The general development of the fins and skeletal system of herring has been described by Lebour (1921), Schnakenbeck (1929), Buckmann (1950), Blaxter and Hempel (1961) and Blaxter (1962). Wells (1922) gave a description of the development of the chrondrocranium ; in the newly hatched larva the ethmoid plate formed the roof of the buccal cavity and the upper jaw (palato-pterygo-quadrate) began to develop only at 13 mm. The detailed development of the vertebral column

THE BEEUVIOWR AND PHYSIOLOOY OB HERRING AND OTHER CLUPEIDS 275
wm worked on by Ramanujam (1929) in herring and by G w p (1940) in Clupea p&ii. In herring the pectoral fins were present at hatching, but the dorsal, caudal and anal fins developed between 12 and 20 m, the vertebrae starting to develop at about 26 mm. Harder (1953) hae shown the relative changes in the proportions of the gut, skeleton and muscle at lengths from 6 2 4 6 mm. Lebour (1921) made a comparative study of the relationship between myotomes, vertebrae and fins in herring, 8prattu.a qrattus and 8ardina pilchardus, as an aid to the identification of the larvae.
B. Feeding of larae 1. Yolk supply
The early growth and survival of 1mae depends partly on the yolk supply and, therefore, on the size of the egg. In herring this varies from ram to race and from parent to parent* ; spring and winter spawning herring produce larger eggs (up to four times the dry weight) than summer and autumn spawners (Rannak, 1968; Blaxter and Hempel, 1963). Larger eggs produce mainly larger larvae, but also larvae living longer on their yolk reserves. Both these factors could be of im- portance to brood survival where the food supply was minimal. 2. Feeding beisaviour
On hatching, herring l m a e show much vertical movement, but after some days more horizontal movement is observed. At the same time searching movements for food occur. Many authors have reported food taken by herring l m m still with yolk sacs. Food is taken by a darting movement following an S-shaped flexing of the body (Schach, 1939; Soleim, 1942 ; Biickmann et al., 1953). The prominent eyes would appear to indicate the importance of vision to the larvae in seeking food. Similar feeding behaviour has been described by Breder and Krumholz (1943) in Harengh pemmlae and Maxfield (1953) found that the larvae of Absa sapidisdm took only zooplankton, suggesting a predatory type of feeding. No feeding wae observed in darkness in rearing experiments on Clupea prdhsii (Kurata, 1969) and Hentschel (1960) and Bhattacharyya (1967) showed a cycle of feeding activity in herring larvae caught at sea, suggesting no feeding activity by night. while this evidence supporta the importance of vision and active selection of food to clupeid larvae, Laaker’s (1962) observations, that the young lmae of Surdinups mrub fed passively with gaping jaws, the eyes at this stage being non-functional, indicate a different mech- anism. However, Arthur (1966) reported that older larvae of this
Yolk sac size for larvae of e given length varied from year to yesr in Clupea mi ( h u k s & al., BuU. Hokkoido Reg. Fish. Rw. Lub. 25, 1-10. 1982).
19

276 J. H. 9. BLAXTER AND F. 0. T. H O U I D A Y
speoies seemed to feed only by day, indicating the importance of vision. 3. Type and quantity of food taken
Bowers and Williamson (1951) and Bhattacharyya (1957) have reviewed the large amount of work done on feeding of herring larvae caught at sea from the analysis of gut contents. While it is not in- tended to discuss the results in detail, it may be said that the size of food organisms taken seems to increase with the size of larvae and a limited selection (or rejection) takes place. For example, organisms such as the copepod Acartia, the diatom Biddulphia and dinoflagel- late Ceratium, though present in the plankton, are not found in the gut. As larvae grow older they tend to take larger food organisms.* The apparently low percentage of larvae feeding reported by some authors, though not for the larger larvae examined by Bowers and Williamson, may be due to rapid digestion or defaecation on capture or fixation, or to night sampling. Arthur (1986) suggested that mainly sickly or starving Sardinqps caerulea larvae might be caught, giving a biased estimate of percentage feeding. He also gave details of the feeding of these larvae and showed how the size of food increased from up to Sop when 4mm long to 200p when 10 mm long. Cushing (1960) also described the increase in size of food taken by larvae of this species as it grew.
The importance of phytoplankton for clupeid larvae has been under discussion for many years. For example, Hardy (1924) found diatoms taken by herring larvae, but Bowers and Williamson (1951) did not. Arthur (1956) considered Sardinops caerulea larvae to be carnivores, less than 1% containing plant food, but Brevoortia larvae are supposed to utilize diatoms (Harder, 1960). Morris (1955) suggested that the ingestion of dissolved organic matter could be of importance to fish larvae (Putter’s Theory), but to date little evidence is availabIe for or against this.
The quantity of food taken waa found to vary with temperature and size in Baltic herring larvae (Waldmann, 1961). Meyer (1878a) reported herring larvae in his rearing experimcnts with as many a8 twenty organisms in the gut. In rearing cxperimentn Blaxtcr (i!N2) f i ) i i r i c l up to ten organisms at any one time in larvae 10-12 mrri long f i r i d Kiirntts (1959) found up to nineteen organimu takcri per day by f : l u p ~ ~ pulhu6 larvae 15-40 days old. 4. Digestion
JII hcrring larva0 tho food i s pnrmd almost immcdiatcly to the and Iizukn el ul. (Bull. tlolskuido Rrg. FiNh. Res. Ln6. 25, 1-10. 1962) reportod an
incream in mouth size with growth but no great change in Him of food takon by larvne of Clupea pallaeii.

THE BEHAVIObR AND PIIYSIOLOt3Y OF HERXXNO AND OTHER CLUPEIDS 277
end of the gut, where digestion takes place (criteria being transparency of gut contents) in 2-5 hr at 5-15°C in larvae 10-12 mm long (Blaxter, 1962). Blaxter and Hempel (1961) found digestion times of 4-6 hr at 10-12°C in slightly larger larvae, but Kurata (1959) gave times of 7-8 hr at about 9°C for Clupea pallasii larvae, clearance of the gut taking 12-20 hr, depending on the original quantity of food taken. Arthur (1956) calculated from feeding frequencies of Sardinqs caerulea larvae caught at sea, that digestion took from 11 i u in the smallest larvae ( 1 P 5 mm) to 3 hr in those 25 mm long. This type of analysis from sea- caught larvae may not be reliable due to the difficulty of recognizhg partly digested food, and the possibility of food being lost due to unusual stimulation.
5. Starvation and the need for food (also gee Section I I I , F ) Very little is known about the time clupeid larvae can live without
food, though clearly this is of great importance in relation to survival in areas of poor plankton. Blaxter (1962) found that herring larvaa in rearing experiments could survive for 2 to 9 days without feeding. Lishev et a,!. (1961) reported a relationship between the abundance of herring (salaka) " fry " in the Bay of Kiga and the number of food organisms from 1955-1961, and Nikitinskaya (1968a) that the larvae of Clupea pallasii required a food concentration in the sea of 20-60 mg/m8. Dementeva (1958) considered that the mortality of Sea of Azov anchovy waa not connected with food supply, due to the plaaticity of feeding. Murphy (1961) estimated that the food organisms of Sardinope caerulea were spaced so that the larvae were on average not more than 3.5-5 cm away from a food organism.
6. Adaptations in relation to food Nikolsky et al. (1961) compared the number of pyloric caecae and
length of the alimentary tract in larval and young herring of different length from the Atlantic and parts of the Baltic. They showed in the oceanic form that the number of caecae was greater, but the variability less, while the alimentary tract was longer. They related this to the better feeding conditions in the Atlantic, in particular high variability being an adaptation to poor conditions (this haa a1ao been reported for egg size).
C. Qrowth of larvae It is not proposed to discuss the growth of clupcid larvae wtimated
from material caught on larval aurveys, though mention should bo mule of tho work by MarRhrill el al. (1037) on tho fat content and incream in

278 J. H. 8 . BLAXTER AND F. a. T. HOLLIDAY
weight of Scottish (Clyde) herring larvae with length. Growth will vary as a result of biotic and abiotic factors as well as inherent ones such aa yolk supply (Toom, 1958; Blaxter and Hempel, 1963). Few attempts have been made to correlate growth rates from year to year and place to place with such factors.
Growth rates have been studied in aquaria to a small extent. Meyer (1878a) and Schach (1939), using wild plankton as food, reared herring larvae whose growth rate was similar to that in the sea. They found the growth rate dropped when plankton was scarce. The best growth waa found by Schach to be in the largest tanks. Blaxter and Hempel (1961) found an increased growth rate in reared herring larvae when coarser plankton was used as food. Blaxter (1962) found some evidence for better growth of herring larvae at higher temperatures and in larger tanks. Nikitinskaya (1958b), hatching larvae of CZwpea pauaSii, obtained less active and less well-developed larvae in tem- peratures outside the average of 3-6"C, and from the later hatchings a t 3-6°C. Farris (1960) suggested that the overall growth of young Say- dinops cueruba larvae was best approximated by two curves, an initial logarithmic and a subsequent exponential one. A common finding of many authors rearing fish larvm is the great size range of larvae of the same origin kept in the same tank, in conditions of apparently abundant food (discussed in Section VI, E). Another finding has been some abnormality of growth, mainly a foreshortening of the head region, as that described by Buckmann et al. (1953) for herring larvae. Nikolsky et d. (1961) analysed the variation in length of young herring from Norway and the Baltic (" salaka 'I). The Norwegian herring showed less variability, whiah they related to the greater stability of feeding condition8 in the ocoan.
D. Rearing of larvae The successful rearing of herring and CZqea pdlaeii beyond the
yolk sac stage has been achieved by a number of workers aa e m - marked in Table 111.
Morris (1956) gave a valuable review of marine fish rearing, par- ticularly from the feeding point of view. From a study of this and published work in the table, the following facilities appear to be desirable for successful rearing : tanks with opaque walls, running sea water, even overhead illumination and a source of wild plankton (Bahnw nauplii, obtained from the adults before spawning, also being eatisfactory ).
Little success seems to have been met with in rearing clupeib othor than herring, though A h u species have been reared in pondn in
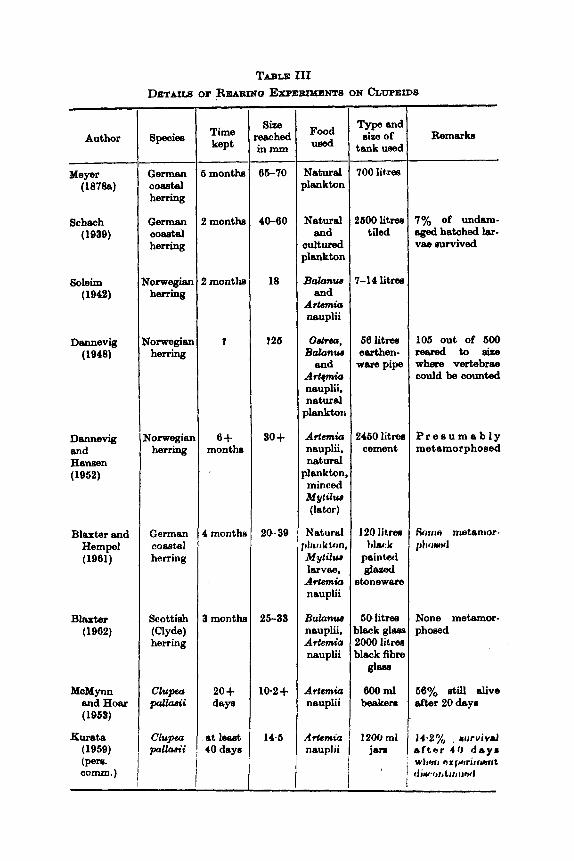
TABLE 111 DETA~CS OF REAEING Ex~ExuMErns ON &WPEIDS
-~
Author
- Meyer
(18788)
Schech (1939)
soleim (1942)
Dannevig (1948)
Dannevig and H e w n (1952)
Blaxter and Hempel (1961)
Bliuter (1962)
McMynn and Hoar ( 1963)
Kumta (1959) (p=. c a m . )
Speciea
German coa8td herring
German coastal herring
iorwegian herrhq
?orwegiau herring
qorwegiar herring
German coaetal herring
Scottish
herring (Clyde)
Clupfa pdkwii
Clupur. pallaa’i
- Time kept
month
I months
; montha
f
6+ monthe
I monthn
I month
20 + day8
at 1- 4 0 day8
- Size
w h e d inmm
66-70 -
40-60
18
f26
30 +
20.39
25-33
10.2 +
14.6
-
Food used
Natural plankton
Natural and
cultured plankton
Balanzcs end
ArtGmio nauplii
08btS0, Balunzcs
and A d Q h nauplii. natural
plankton
Artmio. nauplii, natural
plankton, minced
(later)
Natural
Mytilum larvae. Artamio. nauplii
Balanw nauplii, Ar&m&z nauplii
MytilUiI
7 h k tctn,
Artemio. neuplii
A*& nauplii
~~
Type and size of
tank used
700 l i tm
t600 l i tm tiled
1-14 lit=
66 lit= earthen- ware pipe
2460 litm cement
120 litrea t h k
painted
itonewars g l a d
50 litren )lack glesl ZOO0 litm deck fibm
gl=
600 ml beakern
12WJ rnl jrrn
Remarks
106 out of 600 r e a r e d t o e i z e where vertebrae could be counted
P r e s u m a b l y metamorphosed
None metamor- phosed

280 J. H. S. BLAXTER AND F. 0. T. IIOLLIDAY
uncontrolled conditions (e.g. Pearson, 1952). The very small larvae of Sardinops, for example, make the successful rearing of these species a great challenge.
E. Farming In 30me clupeids a form of fish farming has been attempted. For
instance, Williamson (1909), using low temperatures, retarded the development of herring eggs so that they might be transported to New Zealand ; this attempt failed. Japanese workers (see under Isasaki in Scattergood, 1957) tried to improve the fishery for Clupea pallasii at Akkeshi-machi in the early 1930's. Motoda and Hirano (1961) des- cribed experiments from 1939-1949 in which 2 x 10" eggs of Clupea pallasii were transported on the west coast of Hokkaido in order to see whether the fishery could be improved, but without any apparently beneficial result. Bibov (1960), howcver, reported a successful project where 19 million herring eggs were transported from the Baltic to the h a 1 Sea between 1954-1966, resulting in thc formation of a new stock there, the fish having established themselves to the extent of growing and spawning. The growth in length was up to one and a half times that in the Baltic and the growth in weight four times, and the fecundity five times, as great as the original parent stock. The importance of this type of project, which gives valuable ideas as to the plasticity of the characteristics of a species, cannot be overestimated.
In the shad A h a sapidissima, pond culture has been successfully carried out on the eastern coast of the United States since the end of the last century. Recent publications on the subject are those of Pearson (1952) and Maxfield (1953).
Murphy (1960) and Hida and Thomson (1902) rcportad the estah- lishment of Harengula vittata and Doroarno petlemcnar, renpccti vcly in Hawaiian waters as bait fish for tuna.
F. Mortality o j lurvue
During the period from hatching to metamorjhnirr, it, hrtn tjmn questioned whether clupeid larvae go through critical p c r i o h whwi high mortalities occur, for instancc at the stagt: wtwrt: thc lervac: change from their yolk to external food sourcc8. High mortalitics of hcrring larvac have been reported at sea by Soleim (1942). Bowers and Williamson (1951) commented on published work Hhowing a low percentage of guts of herring larvw containing food. Idow jwrcentagw may be due to extrusion of gut contents on capture and need not necessarily bc a sign of poor feeding conditions or low survival rate of larvae. Hempel and Blaxter (1961b) measured the condition factor of

THE BEHAVIOUR AND PHYSIOLOOY OF HERRING AND OTHER OLUPEIDS 281
herring larvae caught at sea in different a r e a and found in some instances very low values indicating the larvae were near starvation. b k e r (1962) quoted reports of Sardinops caerulea starving in the sea.
The general subject of critical periods has been reviewed by Marr (1956), with new data on Sardinops caerulea,, and he concluded that this concept shoilld be treated cautiously. Farris (1960) took this study further and calculated the effect of different types of growth curves on estimates of larval survival. Taking data on the abundance of larvae of Sardinops caerulea, Sardinops naelanostida and Engraulis japonica caught at sea, he concluded that high mortality took place before the final absorption of the yolk, assuming that net selection, which might allow the escape of small larvae, was unimportant. This observation was supported by laboratory cxperiments on rearing Sccrdinops caerulea when a high mortality took place before absorption of the yolk. While such a mortality in the sea could be explained by predation, this was not so in tanks and Farris suggested that a high proportion of eggs might be imperfect, both in the sea and in tanks. This would be rather surprising on general biological grounds. Murphy (1961) disccssed the critical period concept for Sardiwps caerulea, suggesting how both poor food supply and low temperatures might retard development and prolong this period.
Bridger (1960,1961) considered the fluctuations in the abundance of herring larvae in the southern North Sea and English Channel and concluded that the abundance dcpendcd on the number of eggs spawned by the older fish rather than on the total stock. This might be due to the older fish spawning later, a t a timc of better food supply for the larvae, or i t might be due to intrinsic diffcrc?nce8 in the yolk supply, the older fish producing larger c g y ~ with rrtorc yolk. A nirr i i lru vir!w- point has also been put forward ~ J Y Marty ( 1 959). Again o r ) h t h q i c d grounds i t seems somewhat unlikely that large Rcctionn of a #pawning population should bo producing inviable larvae.
Thc irnportancs of cgg H i m on eurvival of larvao has been studied by Blaxter and Hempel (1!163). Thoy tihowcd that within a race larger eggs tended to produce larger larvae on hatching an wcll a8 larvae that lived longer on their yolk, Whether larger larvae arc' morc! viable as a result of a larger mouth or greater activity remains to IJC tested. Certainly larger larvae take larger food organisms according to samples caught at sea (see Section IV, B). Between races the yolk supply had a very strong effect on survival, larvae living for from 12-25 days at 8°C for instance, depending on their race. Nikitinskaya ( 1958b) found tha t the larvae of Clupea pallasii reared at temperatures above normal were smaller, less well-developed and less active, as

282 J. H. 8. BLAXTER AND F. 0. T. HOLLIDAY
were the later hatching 1ama.e at normal temperatures. She suggested that a wide variability in larval viability permitted the stronger larvae to survive when food was poor. Anokhina (1961), working on Baltic herring, found in a restricted sample that the variability of egg size was greater from parents with low fat content, suggesting that where fat was low, food was poor, and at least the larger eggs would have a chance of producing larvae which would survive. However, this result would bear further examination, particularly in relation to fat utiliza- tion and the different stages of maturation when yolk may be laid down. Ivlev (1966) measured the loss of weight in starving larvae of Caspiatosa volgemis. After 20 days 64% survived, the average weight loss being 18.1% ; after 30 days when only 7% were left alive, the weight loss was 22.1%.
Laboratory studies of larval fish survival in general were referred to by Morris (1966) and Farris (1960). The occurrence of high mortalities in rearing experiments may not necessarily be paralleled in the sea. In rearing experiments on herring, the following causes of mortality have been reported-gas disease (bubbles of gas in the intestine) by Kotthaus (1939), Schach (1939), Soleim (1 942), Dannevig (1 948), Dannevig and Dannevig (1960), Dannevig and Hansen (1962) and Blaxter and Hempel (1961); attack by hypotrichous ciliates by Kotthaus (1939); becoming caught or out by artificial glass-wool substrata by Schach (1939); attack by Lentospora cerebralis (?), and the use of unsuitable food by Dannevig and Hansen (1962); being sucked against the water outflow by Schach (1939) ; and capture by Coryne and blockage of the gut by Blaxter (1962).
Blaxter and Hempel (1961) found two phases of mortality in herring larvae during rearing experiments, the firut after yolk HUC ahwrption and the second at 18-20 mm. Both phases occurred denpitr! an ehundant, though not necessarily suitable, food upp ply. Many fttiluyw f ~ , rwr clupeid larvae after absorption of the yolk munt haw: hx:n dnr: t r , unsatisfactory food, but from cams where 8omc larvw: havc: AIJrviv(!d, i t would appear that the ability to Burvivo varicH widely from larva to larva, even from the samc parente. The second phaso of mortality mentioned above might have bcen due to the imperfect development of a particular organ system.
At present, proof of the existence of critical periodn for Burviva1 of larvae in the sea must await the development of a sufficiently con- tinuous and comprehensive sampling programme on a larval population. Clearly, however, the survival of a brood must depend on a number of factors, two of which are feeding, which has already been discussed, and predation.

THE BEHAVIOUR AND PEIYSIOLOQY OB HERRING AND OTHER CLUPEIDS 283
c. Predation on larvae While this may well be an important source of mortality (Marty,
1966), nothing is known of its extent in the sea, although some pre- datory species have been noted. Lebour (1923) described plankton investigations where herring larvae were observed in the guts of various species of medusae, in the ctenophore Pleurobrachia and in T q - teris. Larvae were also taken with Sqitta attached to them. This was also observed in Sardinops caerulea by Arthur (1956). Marshall et al. (1937) reported an absence of herring larvae in the Firth of Clyde, possibly due to predation by medusae and ctenophores, while Steven- son (1947, 1962) cited ctenophores as preying heavily on the larvae of Clupea p a l h i i . Lebour (1924) and Arthur (1966) report cannibalism in herring and Bardinups respectively, adults eating the young. The latter also reviewed the work done on the possible effects of phytoplankton and metabolites on clupeid lazvae.
H. Salinity tolerance and om-regulation of larvae 1. Salinity tolerance This has been studied in newly hatched herring larvae by Holliday
and Blaxter (1 960) and Bishai (1 961a), and in Clupeapalhii by Kurata (1969). Figure 1 shows the salinity tolerance of herring larvae. The most striking feature of the results is the very wide tolerance of the larvae, from 14-60%, over 24 hr, and 2-5-52.5%, over 168 hr. Bishai (196la) found that herring larvae lived longer in salinities from 10-15%,, than in higher or lower salinities. The 24-hr values quoted for Clupeu pallasii by Kurata were from 1.06-69.5%,. Kurata also gave figures for 10-day-old Iarvae (1.06-66-5%,,) and 20-day-old larvae (2.45-40*25%,). No abnormalities of development were reported by these workers. However, Kryzhanovsky (1956) reported that if Baltic herring eggs, which normally develop in water of salinity 4-5%,, were transferred to salinities of 25%, then abnormal development and death of the larvae occurred. Stevenson (1962) suggested that larvae of Clupeu palhi i might die in the open sea due to its high salinity. It would appear that other clupeids have a wide salinity tolerance, for instance Perceva (1939) found that larval Cmpiakwa caspia would survive in fresh and " salt " water.
2. Osmotic regulation Holliday and Blaxter (1960) showed that the tolerance of herring
larvae was based on a process of active regulation. If the larvae were transferred from sea wafer to any salinity within their tolerance range

284 J. H. 9. BLAXTER AND F. a. T. HOLLIDAY
Rearing salinity p/oo) A
I
n -
i
24 h 48 h 168 h
FIG. 1. The salinity tolerance of herring larvae reared in differont salinities after varying (Reproduced with perrniwion from Holliday and
r r i , , tc - tlrnrl %i'){, itfive, ),lit Inr viui iriiu f ive ,
periods in transfer salinities. Blaxter, 19GO.)
black-more than 50'%, &MI ; h i s t i )d clear-more tharr so'%, alive vr1t1 IU I.IVIL

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 285
the body fluids first showed a rapid change in concentration. Regulation started within 6-12 hr and the body fluids were returned to close to their original value. In salinities above 12%,, (the isotonic point) the body fluids stabilized at a value roughly one-third of the external salinity. Below la%, the values were only slightly lower than the sea water values. The larvae showed changes in body weight corresponding
Control in sea water
0.30 5
__-_ L I . I 1 - 1 20 40 60
0 I Sl l ln l ry ( /,,)
Fro. 2. O~niotir c-oticentrations of larval body flIlidH. larvcu, trnurrforrd f rom no8 wator (32.3%,). (Reproduced with permission from Holliday and BIaxter, 1960.)
to a loss of water in the high salinities and a gain of water in the low salinities, the weight returning to near normal after successful regulation.
Within the salinity range 2.5-52.5%,, the larval tissues were regulsbd to equivalent salinities 9.8-15.0%, (A = 0-59-0-90). Before regulation was complete, however, the larval tissues experienced and survived internal concentrations equivalent to 8.7-27-5% (A = 0.52-1.06) ; in normal sea water the value waq equivalent to 12%,, ld --- 0.72), see Fig. 2. Lasker and Theilackcr ( I ! j O h ) oht,sinrxl rr:niJltn o r 1 ,Yw. dinops caerulen larva(. showing that irr sw, wati.r of :iGX,, mlqj ti'%,., t,hr:

286 J. H. 8. BLAXTER AND F. 0. T. HOLLIDAY
body fluids and yolk were regulated to a concentration equivalent to la%,. They showed that whereas newly hatched larvae died in distilled water, larvae two days old survived for at least 24 hr, although the body fluids were reduced in concentration to about 9.7%,.
3. Sites of regulation These have not been determined experimentally in fish larvae.
Shelbourne (1957) suggested the epidermis, and this seems to be most likely.
4. Effects on metabolism It might be expected that as the degree of regulation required
depended on the difference between the environmental salinity and the tissue concentration, more metabolic effort would be required to survive in the extreme salinities. Lasker and Theilacker (19624 found no difference in oxygen uptake in Sardinops caerulea larvae living in sea water or in sea water diluted until it w a isotonic with the tissues. This work was extended by Laaker and the present authors, who found that there was no difference in oxygen consumption of herring larvae incubated and hatched in salinities ranging from 5-50L. However, some variations occurred in the few hours immediately following abrupt transfer from one salinity to another, but as regulation was achieved the value returned to its previous level.
5. Effects of starvation Ivlev (1955) showed that there was little difference in salinity
tolerance between starved and normal C m p i a h a volgemis larvae, although the range of salinitieH te~ted WRR #mall.
1. Oxygen uptake ReRpirntory axehnngu in youngcor tarvat, afrnctst csrtainly takes
placa ovcr tho tmiy Hirrfncch. Ilartlor (1054) roportcal that thc gills of hcrring were iiot functional until a lcngth of about 20 mm. Volodin (1956) suggosted that carotenoids in tho yolk ected aIs respiratory pigments. A scnsitive technique is required to meamro ths oxygon consumption of singlo larvae. AH the activity of tho larvao is dominant in determining the oxygcn requirement, comparisons of metabolism in larvae of different age or in different environmental conditions are best made on individuals anaesthetized to the same level (determined by heart rate). Table I V gives values of oxygen uptake in some clupeid larvae.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 287
Speoiee
TABLE IV OXYGEN CONSUDWCION OF CLUPEID LARVAE
0, uptske p litree/larva/hr Author
Mershall sl al. (1937)
Laaker and present authora (unpub.)
Volodin (1966)
Laaker and "heilecker (19628)
Herring (Clyde, Scottish)
c o d ) Herring (German
Herring (Beltic)
Sardinop cacrulur
0-048 0.03 7
0.07 0.70
0.19 0.32 0.43
0.08 0.03
Remarks
In the dark (a12"C) In the light (8-12°C)
Aneeethetized (8°C) Unenesethetimd (S'C)
1 day poet hatching (14OC) 2 days post hatching (14%) 3 days poet hatching (14'C)
Active (14OC) Ineotive (14°C)
The effects of salinity on oxygen uptake were considered in Section IV, H.
J. D e m l receptors Cunningham (1889, 1894) and Lebour (1921) described in larval
Sardina pilchardus and Sprattw aprattw a number of small papillae placed alternately down the length of the body, about six on each side. de Kock (personal communication) found them also in the head region of herring larvae where they resembled small taste buds in structure, having a number of projecting sensory hairs (see Fig. 3). It is possible that they are chemoreceptors, or they may have some hydrodynamic function (they also resemble the lateral line organs of the adult , and the cupulae described by Verheijen (1956) on the adult Sardinu pilchardue).
K. Temperature, pH, oxygen, pressure and light a8 limiting factore Marshall et al. (1937) kept newly hatched herring larvae for a short
time at 26°C without harm. Kurata (1959) found that Clupea p a k i i larvae died after transfer from fM"C to 17-18°C. Blaxter (1960) showed that recently 'hatched herring larvae would tolerate 22-24°C and -0.75 to -1.8"C when acclimatized to temperatures htweon 7.6 and 154°C. The range of lethal temperatures tcndwl h tm n m o w w l by unfavourable salinities.
Bishai (1960b) found that l1 the activity " of homing Iarvm becarno rustriatad a t pH v ~ ~ I u ~ R of 0.4 to 6.6, tho luthal pH boing lower. Raising the pH of the water from tho rearing value of 8.1 , to 8.6, cautmd death.
High oxygen tension wm less important to newly hatohod herring

288 J. H. 9. BLAXTER AND F. a . T. HOLLIDAY
(Bishai, 1960a), whose activity was " restricted " between 26-9-32*2% oxygen at 14°C ; this occurred at 55-64y0 oxygen in larvae 3-4 days old. Larvae died in 6 hr at 11.6% satura,tion. The larvae would live at oxygen values up to 41 1.4%.
Bishai (1961b) showed that herring larvae could withstand pressures up to 4 atmospheres. No gas bubbles were found in the intestine when the pressure was released from 2 atm to 1 atm in 10 min.
+--+
3ge jaw
FIQ. 3. "raneveme section of the head of a herring larva ehowing a dermal receptor (de Kock-unpubliahed).
Ljubickaja (1957) reported that there were more Sardinella '' spawn " and larvae dead near the surface in the Sea of Azov than near the bottom. She suggested this might be a mortality due to infra red light which waa absorbed rapidly by sea water and would only affect organisms near the surface. Dementeva (1958) condercd that sunlight wr~uld only hc deleterious because it warmed up thc nurfrtce layfm.
Bishai (1960a) gave a good review of work on K ~ A H rliwrurr, and ih possible causeR in rearing experimcrrtti on Iiorring larvac. Six poesi ble

THE BEHAVIOUR AND PHYSIOLOOY OF HERRING AND OTHER CLUPEIDS 289
causes were discussed-formation of gas by monads, swallowing of air bubbles, break-down of yolk material, lack of oxygen and therefore intake of air from above the surface, supersaturatioq of the wster and swallowing of air for the swim bladder. Bishai concluded that super- saturation of the water, followed by involuntary uwallowing by the larvae, was the most likely cause. Blaxter and Hempel (1961) also concluded in older larvae that active swallowing of air trapped on the surface, particularly in supersaturated water, was also a, cause of gas disease and this might be eliminated by careful cleaning of the surface.
L. Locomotory behaviour and rheotropic response On hatching, herring larvae tend to swim vertically to the surface
and then sink head first to the bottom. At the end of the yolk sac stage this behaviour bcbcomeu replaced by horizontal searching movements for food. Breder and lirumholz (1943) rcported that Harengula pensa- colae, about 3 mm long, were unstable, the centre of buoyancy being lower than the centre of gravity and equilibrium being maintained by the pectoral fins. They moved by alternate sinking and swimming periods. Clupeid larvae with an oil globule swim upside down until this is absorbed.
Biickmann et al. (1953) found that herring larvae swam both with and against a current induced in a circular aquarium tank, while in an open air tank, and in harbour, they were always seen to swim anti- clockwise. These authors mentioned the need for optical marks to get a rheotropic response. Blaxter and Dickson (1959) and Bishai (1960~) found that herring larvae about 8 mm long would orientate against a current passing through a tube. Bishai also found that thc larvae did not show a pseudo-rheotropic reRponRc when ~trip:rl paper wan p a d along the tube. Morris ( 1956) ~uggest(:d that a cum:tit WA# of irnport,,rtncct in rearing tanks as larvae would thcn tcrrd to H t c m n u c h a c:urrmt instead of making frequent, probably di~arlvaritagcou~, coritactn with the walls of tlic tank.
M. V'erlicul migration of lurvtie 1. Qenerul
Reports of the distribution of clupeid larvae in the sca are difficult to compare and cinalyse due to sampling problems. In relation to diurnal vertical migration these are perhaps most serious. Johansen (1925) gave some evidence for a downward movement by day of herring larvae 10-18 mm long, while Russell (1926, 1928) had similar evidence for the larvae of Sardina p i l chrdw and Sprattus sprattus under 24 mm long. Silliman (1950) found that the larvae of Sardinops caerulea,

290 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
greater than 8 mm long, tended to move down in daytime, but not the smaller larvae, while Bridger (1958) found that older herring larvae (16-26 mm) around the English‘coast tended to go nearer the bottom than the younger ones by day, and Colton et at. (1961), in the Gulf of Maine, reported a diurnd migration of herring larvae, the extent of the migration increasing with size.
However, one of the most striking observations of Johansen, Russell and Silliman, and also of Stevenson (1947) on Clupea pk i i , waa that greater numbers of larvae were caught by night, particularly large ones. For example, Johansen caught about four times as many 8-19 mm herring by night as by day, and no larvae longer than 20 mm at all by day. Russell caught ten to thirty times more 8ardirm pikhrdwr and Sprattus wprattus up to 24 mm long by night, Silliman about ten times more Sardinops caemlea over 8mm long by night, while Stevenson (1947, 1962) caught five times the number of CZupea paZhii l m a e up to 15 mm long by night, and none over 15 mm at all by day. Ahlstrom (1954) reported the undersampling of Bardimp caermlea in daylight, and Murphy (1961) showed in the same speoies how the ratio night hauls : dayhauleincreasedwithlength of the 1ma.e from 4.75-21-26 mm.
The first explanation given for this by Johansen (1925) and Russell (1926, 1928) was that vertical migration waa taking place in daytime and the larvae were not being sampled, but both these authors thought that net avoidance by day might be signifiwt, and Russell (1926) considered that swarm formation by day might amount for lower catches, due to unevenness of the larval dietribution. The vertical migration explanation was not supported by hauls made at greater depths (Russell, 1926, 1930), where no abundance of larvae was found by day. Stevenson (1947) and Sillimsn (1950) ah0 found no conmn- trations of clupeid larvae at great depths. It seems that while restrioted vertical migration and swarm formation might account for some of the difference found, avoidance of nets by day waa the mein factor, especially aa the nets were moving slowly.
2. Avoidance of nets and swimming qeeds Net avoidance by sight was particularly emphwized by Silliman
(1960), while Stevenson (1947) thought that phosphoresenoe at night in hi8 area would make the net as visible by night aa by day. New methods were clearly required to show the distribution of larvae. Bridger (1958), using a high-speed sampler, showed that herring larvse up to 25 mm long were caught in equal quantities by day and night, which clearly supported the view that net avoidance. oaused e serious sampling bias with tow nets used at low speeds. Tibbo et d. (1968) and

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLWEIDS 291
Colton et al. (1961) found that larger herring larvae in the Gulf of Maine were caught mainly a t night by a metre net ; the latter authors using a Hardy high-speed plankton recorder also found the ratio of larva0 caught by this gear, compared with the metre net, increased with increasing size of the larvae. Silliman estimated that Sardinops caerulea larvae would have to swim a t about 5 cm/sec to escape a net towed a t 50 cmlsec.
Blaxter (1962), using herring larvae from 8-20 mm long in tanks, tested their reaction to visual stimuli such aa moving obstacles, and vibrations up to 10 c/sec, and measured their swimming speeds. He found that they were remarkably unreactive to such stimuli under tank conditions. The maximum swimming speed varied from about 3 cm/sec for larvae 8 mm long to 30 cm/sec for larvae 20 mm long. There was a tendency for an increase in maximum swimming speed a t the stage when the caudal fin developed a t about 15 mm. Bishai (1960~) worked on sustained swimming speed of herring larvae 6-8 mm long and found that they could maintain 0.58-1.03 cm/sec for a t leaat 1 hr.
From these data it seems likely that clupeid larvae have the swim- ming ability to escape nets towed at low speeds of, say, up to 2 knots (100 cmlsec) ; it is unlikely that they would escape from a high speed sampler towed a t 6-6 knots (250-300 cmlsec). If the evidence put forward in this review of the importance of sight in clupeids is accepted, and sight is the most important means of detecting such nets, then even nets hauled slowly will catch larger larvae a t night. .
N. Rmponse of larvae to light Spooner (1933) studied the reaction of herring larvae to light using
convergent and parallel beams to separate intensity and direction effects. Their photo-positive behaviour was in general more a reaction to direotion, activity falling off at low light intensities. Woodhead and Woodhead (1966) also found a drop in activity of herring larvae kept in a dish with diffuse light from below-from 35% of the time active at 4,000 lux to 20% at 20 lux. This photo-orthokinesis waa accom- panied by phototaxis, the larvae swimming towards directional light. In a, tall vessel the larvae were active in complete darkness. Below 100 lux most movement WES vertical ; at higher light intensities more and more horizontal movement was observed aa if there were a dorsal light reaction. It was suggested that this movement provided a mech- anism for vertical migration. It is necessary to note also, however, that herring larvae gradually develop more horizontal movement during the yolk-m stage as part of' feeding behaviour. Soleim (1942) found herring larvae more often in lighted parts of an aquarium. Buckmann 20
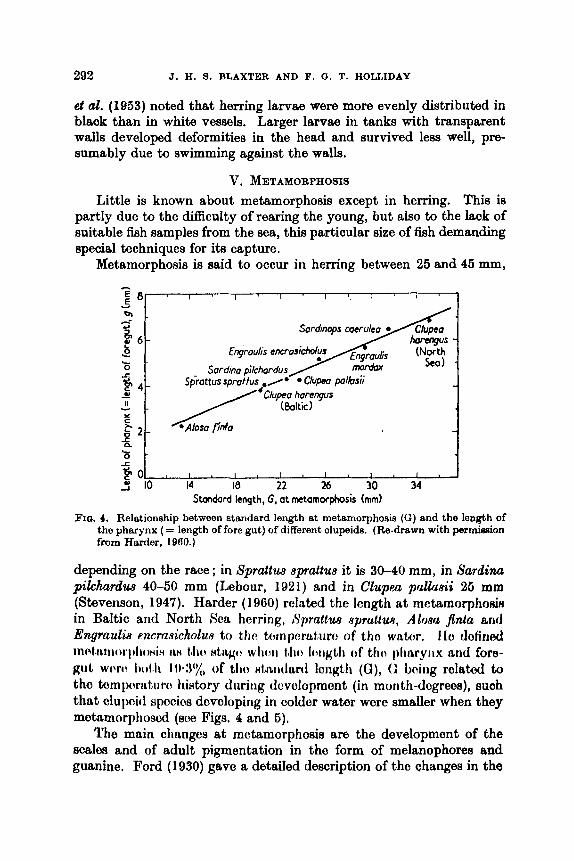
292 J. H. 9. BLAXTER AND F. (3. T. HOLLIDAY
el al. (1953) noted that herring larvae were more evenly distributed in black than in white vessels. Larger larvae in tanks with transparent walls developed deformities in the head and survived less well, pre- sumably due to swimming against the walls.
V. METAMORPHOSIS Little is known about metamorphosis except in herring. This is
partly due to the difficuhy ofrearing the young, but also to the lack of suitable fish samples from the sea, this particular size of fish demanding special techniques for ita capture.
Metamorphosis is said to occur in herring between 25 and 45 mm,
, , , , ,
14 18 22 26 30 34 PO f 10
Standard length, G, at metamorphosis (mmf
FIG. 4. Relationship between standard length at metamorphosis ( G ) and the length of the pharynx (= length of fore gut) of different clupeids. (Re-drawn with permhion from Harder, 1960.)
depending on the race ; in Sprattus eprattus it is 30-40 mm, in sardina pilchardus 40-50 mm (Lebour, 1921) and in Clupea pallnsii 26 m m (Stevenson, 1947). Harder (1960) related the lcngth at metamorphowiR in Baltic and North Sea herring, Sprultue npruitw, Alonu jinta ant1 Engradis encmsicholue to the tc:inpcr&t,iirrt of tho wntcw. I lo dofined inctt,rwiroi.l,lioHI" iLN (Jro HtiLgo wlrcw tho I ~ g b h of tf i t r plrnryirx arid fore- girt w t w I)olth I w : ~ ' ~ , of tho Htmitlarcl loiigth (a), (; baiiig rolatod to tho tornprat,ut.cr history during tlovolopmont (in month-dcgrees), suoh that clupoid spocics dnvoloping in colder water were smaller when they motamorpliosod (so0 Figs. 4 and 5).
Tho main cliangcs at metamorphosis are the development of the scalee and of adult pigmentation in the form of melanophores and guanine. Ford (1930) gave a detailed deecription of the changes in the

THE BEHAMOUR AND PRYSIOLOQY OF HERRING AND OTHER CLUPEIDS 293
vertebral column and position of the fins in herring at this stage. There was a forward movement of the dorsal fin and anus and a backward movement of the pelvic fins.
The speed of metamorphosis at sea, is unknown but Blaxtor and Hempel (1961) found it lasted about 10 days at 15°C in German coastal herring reared in tanks.
The heart, which prior to metamorphosis is composed of an outer myocardium with an endothelial lining, seems to be the source of erythrocyte formation (John, 1932). The endothelium thickens and megaloblasts bud off into the lumen of the atrium and ventricle, where they pass through an erythroblast stage to become erythrocytes. The spleen, which is the site of formation of erythrocytes in the adult, is not present at this stage.
2 0 1 . I , I , I , I , l , l l
10 14 I8 22 26 30 34 Standard length, G, a t metamorphosis (mm)
FIQ. 6. Relationehip between standard length at metamorphose (G) and temperature (Re-drawn with permiasion from hietory of larval clupeids in month-degrees.
Harder, 1960.)
Buchmann (1940) observed a large increase of thyroid activity a t metamorphosis, the cell heights in the follicles increasing and colloid being formed and released. There was also a change in the pro- adenohypophysis which he believed was due to the production of thyrotropic hormone. According to Holliday ( 1960a), however, the application of a histochemical (P.A.S.) technique indicated that the site of thyrotropin production was the meso-adenohypophysis, and Buchmann may have been reporting the production of a growth hormone.
The formation of the gonads seems to take place at about this stage, at least in Sardina pilchardus where dos Santos Pinto and Andreu (1956) established the formation of the gonad at a length of 37 mm and sexual differentiation at 45 mm.

294 J. 11. S. BLAXTER AND F. 0. T. HOLLIDAY
Little can be said about changes in the habits of clupeids after metamorphosis. Certainly herring at this stage start to shoal and are found inshore in large numbers. Whether there is a need to move into water of low salinity is not clear, though such a need has been correlated in other fish species with increase in thyroid activity. It may be that feeding conditions are more suitable, but there can be no doubt that young herring are particularly heavily preyed upon at this time.
VI. POST-METAMORPHIC STAQES A. Establishment
The general delicacy of clupeids was the reason given by such workers as Parr (1927), Spooner (1931), Loukashkin and Groody (1965) and Parrish et a2. (1958) for the lack of experimental work on this important group of fish. Clupeids have not often been established in the aquarium. However, a number of attempts have been made not merely to keep the fish, but also to determine the reasons for the success or failure of the attempts and to perform expcriments on them. These are summarized in Table V. Loukashkin ancl Groody (1955), Ver- heijen (1950), Farrin et al. (1957) and I’arrish et al. (1958) reviewed tho problems of establishing clupeids in aquaria, listing the factors that contributed to mortality at each of the stages of capture, transport, initial establishment and long-term maintenance and feeding. In general these authors concluded that epidermal damage was the prin- cipal cause of initial mortality. Parrish et al. suggested that osmotic dehydration followed the damage and these mortalities could be overcome by careful handling and keeping the fish in sea water of salinity roughly isotonic with the blood-about IS%,. It is interesting that, much earlier, Elmhirst (1908) accidentally di~covcrcd that herring survived best in an equal mixture of sea water and frcsh water.
The size and structure of the aquarium, the rapidity with which the herring start to feed and the prevention of disease were the main factors controlling the long-term establishment of the herring (Parrhh et al. 1958). SUCCeHSfUl experimental handling waH h i t carrkd out under anaesthesia with tricainr: mcthane Hulphonata. ‘I’ho Ircrring always appeared very senRitive to ditjturbances, ancl thowcd no &gnn of becoming conditioned (or “ tame ”) in any way. ‘1’hr:y werct in- fluenced by thc size of the tunk, an fcc:ding could not trr: i i l i t i i i k d if‘ tho fish length: tank spaco ratio was too nmall.
Both Verheijen (1956) working on Sardina p i l c h r d w and I’arrinh et at. (1958) on herring, found that feeding fish (in the cam of herring also feeding Gadus virens) influenced non-feeding fish and caused them to begin feeding.

TABLIE V SOME PUBLISHED WORK ON KEEPIXC CLUPEIDS IN AQUARIA
~
I Fish Time
kept Means of transport
Experiments carried out capture
Survival in low salinities c. 3 months i - - C l u p harcnqua
- - I Effect of environment on atructure. Feeding behaviour
At lea& 1 yerr
0.75-14.5 Seine from I weim 3alvanired
iron tub Food preference, digeation, opti-
mum light conditions Some months
- lTz:rm Effect of temperature and light on
Effect of light, currents, salinity
EBect of echo-sounders
Effects of sounb, light and
feeding
barriers. Feeding behaviour
ObEt&?lHl
Johnson (19398) 112-1.600
1 5-2.800 ~
Biickmann Ltol. (1953)
Skoylund(l953) I 2630 I 1,100 - 80-litre
containers Verheijen (1963) I 2-12 5oo-2~ooo i
- 1 Sink net Shoaling behaviour
Farrin el 01. (1967) I 620 I SSO-7.M)o 1-18 I Seine net M-litre containers
Study of feeding and mortality 2 years
Parrish c: 01. (19S8) 6-23 I I 460-7'600
06-16 General maintenance and feeding u p t o 4 y s e r s Seine, trawl and drift net
-
la-litre containers
- 100 daja General maintenance in tanks and livecar
Temperature and salinity tolerance
Mortality, oxygen and salinity
Growth and otolith studies
barriers
8.1-11.2 Dipped from weir
At l e u t 9 month.
Over 1 ysu I, .* 7.12-14 Dip net inshore
Plsstic ear- boys, bucket
communication)
9.0-14.3 Dipped fro1 weira
Bucket Feeding and growth studied 3 month ~
Clupm pruadii
Branding 7 mon th
shelfordand Powen (1916)). rr Rerction to tempemture, sdinity and oxy en gradients. Rcsis- tance to 80. and H,S
Growth and spawning
Population studies, marking mor- tality. feeding and shoaling
Some month
- -
"Boat pound"
- Seine net Hourston(1968) 1 1 hrgepo;dsand
Anon (l9S2)
cim. tanka 0.9 m dia., 0 4 m deep
10-12 Schooling pattern, stimuli involved in feeding, effects of electric fields
Feeding, mortality, general maintenance " 11-20.5 --litre con-
b i n e n with lights
Neuly 3
Diet and growth studies n
Me& of coloured light. 4+ month "
Diet and fat storsge "
Circular racsptaeles
Schooling and feeding By lighb
-
11.1-2&4
- swimming atudies 2 month Cylindrical 3.2 m I 7-10 1 dim. Kiluaurs (1934)
450-lib con- t a ine r s . b v e n B h per 6 lit=
General handling and maintenance Some month. Seine net

This Page Intentionally Left Blank

TXE EIELAVXOUR AND PHYSIOLOGY OF HERRING AND OTRER CLUPEIDB 295
B. Feeding 1. Behatiour
In most clupeids this seems to be a direct, predatory snapping action. This was deduced by Hardy (1924) from the analysis of stomach contents of herring, and has been observed quits frequently in aquaria in ti number of species, in herring for instance, by Battle et al. (1936), Buckmann et al. (1953) and by Verheijen (1953), in Sardina pilchardus by M u h i 6 (1960), in Harengulu penaacolae by Breder and Krumholz (1943), in Sardinop8 caerulea by Loukashkin and Grant (1959), in Porno- pseudoharengus by Graham (1957) and in Sardinellu w r - opthdmia and Brevoortia tyrannw by Breder (1959). Kirchhoff (1958), in a detailed study of the functional morphology of the herring, des- cribed a sucking action which would occur as food wm being taken.
Kirchhoff (1958) also stated that herring had a suitable arrange- ment of gill rakers for filter feeding, and a study of gut contents in clupeids does suggest that some species are at least partly filter feeders. Radovich (1 962) showed this for Sardinops urermlea and Komarovsky (1959) for Surdinella aurita. Gunter and Christmm (1960) reported findings of detritus and bacteria in the gut of Brevoortia patronus, and Harder (1960) cited Brevoortia and the engraulid Cetengraulis as diatom feeders. Lagler et al. (1962) stated that Brevoortia tyrannw swam with mouth agape through rich plankton, straining as much as 6.8 gallons of water a minute.
During feeding shoal formation tends to be disrupted. Hourston (1958) found this in Clupea pa lh i i , Graham (1957) in Pmlobw pseudoharengw and Breder (1 959) for Sardinellu mropthalmia and Brevoortia tyrannw. In fact, feeding could cause a considerable delay or change in any type of migration pattern (as discussed under Section VI, R). Jakobsson (1961) noticed how herring chasing and feeding on fish larvae changed their behaviour so markedly that they became very spread out, and difficult to catch. Glover (1957) suggested that herring might be able to detect concentration gradients of their favourite food organisms. Thus feeding might cause an aggregation of dispersed fish, or a break up and possible dispersion of shoals which were already established.
2. Selection of food Williamson (1917) observed herring in captivity. He found they
could detect food from about 30-90 cm away depending on its size and that they rejected some food after taking it into the mouth. Blaxter and Holliday (1968) found that detection of food by herring kept in

296 J. 11. 5. BLAXTER AND F. 0. T. HOLLIDAY
aquaria was by eye; the fish attempted to take food regardless of shape, size or colour, once they were established. However, first texture, and then taste, were very important in selection within the mouth. Taste seemed to be dependent on water soluble, rather than fat soluble, substances. Anon. (1052) found in Surdimps caerulea, however, that the olfactory sense was of great importance in detection; this increased feeding activity after appropriate stimulation. The import- ance of smell was also mentioned by Cushing (1960).
Savage (1931) made a comprehensive study of the feeding habits of North Sea herring and reviewed earlier work by Hardy and others. He found, by also studying the plankton, that different food organisms were in the same proportion in the herring stomachs as in the plankton.
3. Effect of light and temperature on feeding Clupeids not only appear usually to break shoal a t night, they also
atop feeding. This was reported in general terms by Zusser (1958a) for pelagic plankton-catching fish. Girsa (1961) reported also that visual feeders tend to disperse and stop feeding at about 0.1 lux. Girsa (see Section VI, U) developed the theme that light might be a signal for feeding and cause both attraction of fish and greater activity.
In aquaria Battle eft al. (1936) found that captive herring did not feed in the dark but they, Johnson (1 939a) and Blaxter and Holliday (1!158), found feeding took place in moonlight (incident light intensity probably about 0.25 lux). Blaxter and Holliday found a threshold light intensity for feeding (using tungsten filament light) of about 3 lux; optimum feeding conditions appearing to be from 100 to 1,000 lux, feeding alwaya heing bettar when t h o daylight W[LR not too bright,. There appeared to be nomc cvidciico for II tliiirrml f'i~wlitig r h y i h , feeding activity being lowest from 0400-0800 hours. Williamson (1917) also reported that captive herring fed better in dim light. Loukaahkin and Grant (1959) found that h'ardinops caerulea would feed as well at 5 lux as a t 200 lux, but did not try feeding the sardines in Jarknees. In view of their sensitivity to olfactory stimulation (Anon., 1952), thia would make an interesting experiment.
At temperatures below 4"C, captive herring ceamd to feed (,JohnHon, 1939a; Blaxter and Holliday, 1958), and Williamson (1017) alm reported inactivity at low temperature. Howevcr, Battle p t al. (1936) and Farrin et al. (1957) found feeding in captive herring at, H. tA:mporatiirct of 1°C and Kamshylov and Gerasirnov (11160) w ~ n at . fb4"C. 'I'hc:~: findings may have been due to a longer acclimatization to the low temperatures, or to a difference in the physiological (or psychological) state of the fish.

THE BEHAVIOUT AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 297
4. Effect of inter& factors on feeding Williamson (1917) found that “ hungry ” herring were less easy to
frighten and would feed better. It has often been said that spawning herring c e m to fecd, but sometimos they have becn found with food in the gut-even herring eggs, which suggests feeding at some time during the mass spawning act, for herring do not appear to take food off the bottom.
The importance of psychological stimuli in feeding or initiating feeding is mentioned in Section VI, A, where i t is seen that the presence of feeding f%h, not necessarily of the =me species, is a great incentive to feeding.
6. Competition for food Variations in growth rate of clupeids kept in aquaria suggest the
importance of dominant feeders, or of fish with more efficient utiliza- tion of food (see Section VI, E). But apart from intra-specific compe- tition of this sort, about which practically nothing is known, there are indications of negative correlations between the abundance of related clupeid species in the same area. There have been changes in the dominant species recently, but whether these are reversible or not remains to be seen. Such correlations were discussed by MacGregor (1959) and Marr (1961) for Sardinops eaerulea and Engraulis mordaz, for Clupea pallusii and Sardinops melunosticta by Motoda and Hirano (1961), and for Clupea harengus and Sardina pilchardus by Cushing (1961). One possible explanation for these fluctuations could be competition for food, but the extent to which long-term changes in hydrographical conditions are responsihlo in not clear. It, rnrty t i c of significance that these changes occur near the extrcmw of t h r : rnrigc of these species, where changes in hydrographical c o n d i t i o r i n m i g h t I)(: expected to exert the maximum effect. hcr:ntly SOlJth WJLrfl I I ! N l ) has suggested that the increase of Sardinu pilchrlrdw arid dw:rww of herring described by Cushing (1061) might be due to long-torm tem- perature charlgcs which put the herring at a disadvantage frorrl the point of view of metabolism.
C. The alimentary system I . Morphalqpy ( i d (inntom?/
Jmolwhqpi ( I !)13) dcscribcd tho hcrring intestine as being short (one-half to thrcc-quarters the body length) and associated this fact with the carnivorous diet of the fish. Herder (1960) compared gut morphology with body form, food and environment in forty-six
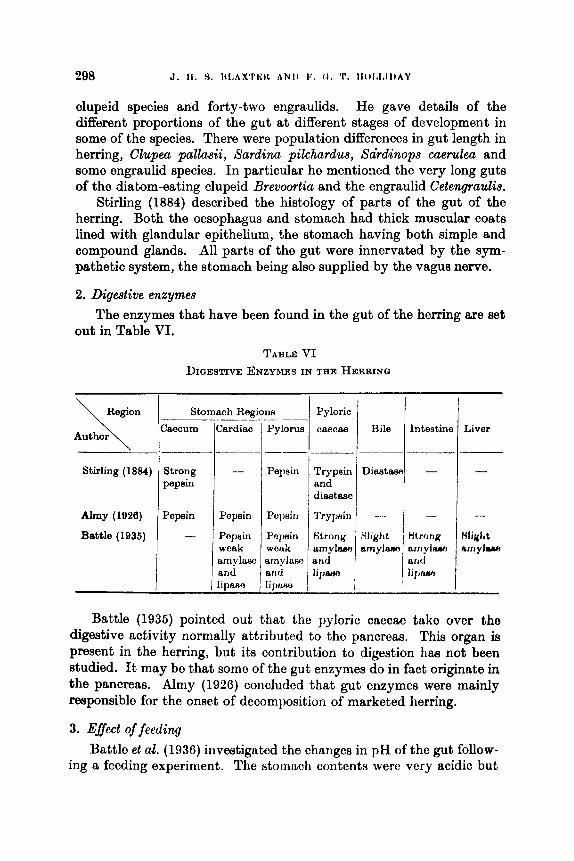
clupeid species a d forty-two engraulids. He gave details of the different proportions of the gut a t different stages of development in some of the species. There were population differences in gut length in herring, Clupea pallasii, Sardina pilchardus, Sdrdinops cuerulea and some engraulid species. In particular he mentioned the very long guts of ths diatom-eating clupeid Brevoortia and the engraulid Cetengraulia.
Stirling (1884) described the histology of parts of the gut of the herring. Both the oesophagus and stomach had thick muscular coats lined with glandular epithelium, the stomach having both simple and compound glands. All parts of the gut were innervated by the sym- pathetic system, the stomach being also supplied by the vague nerve.
2. Digestive enzymes
out in Table VI. The enzymes that have been found in the gut of the herring are set
TABLE VI DIGESTIVE ENZYMES IN THE HERRING
Pyloric
caecae Bile
Stomach Reg
Caecum Cardiac - Region
Author \ Intestine Liver
' I I-- Stirling (1884)
Almy (1926)
Battle (1935)
Strong - pepsin
Pepsin Pepein
- Pepsin weak
Pepsin
PUr)HiYl
Popnin wonk arnylaru, anti l i 1 J f M
1 i
amylase and lipam
I -
Battle (1935) pointed out that the pyloric caecae take ovcr the digestive activity normally attributed to the pancreas. This organ is present in the herring, but its contribution to digestion has not been studied. It may be that some of the gut enzymes do in fact originate in the pancreas. Almy (1926) concluded that gut enzymes were mainly responsible for the onset of dacomposition of marketed herring.
3. Eflect of feeding
Battle et a,?. (1936) investigated the changes in pH of tho gut follow- ing a feeding experiment. The stomach contents werc very acidic but

THE BEHAVIOER" AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 299
this acidity was neutralized by the bile in the intestine. More peptic activity was found in the full stomach than in the empty, or partly full one. The pH optimum for pepsin was between 2.5 and 2.85. Its activity increased with rise in temperature (Ql0 = 1.5). The level of trypsin in the pyloric caecae was highest in herring with full stomachs. Calcium salts present in the food increased the activity of trypsin. The pH optimum lay between 8.5 and 9.5, the Qlo being 1.5. The lipase of the pyloric caecae was rendered more active by the bile.
4. EfJect of enzymes on food Battle ( 1935) reported that live euphausiids, Meganyctiphanes, were
killed in 3-4 min, and the copepods, Calunus and Temora, in 30 sec, by the action of the stomach secretions. Breakdown of the food started in the caecal and pyloric regions of the stomach, while in the pyloric caecae the food was in the form of an oily chyme, mixed with mucus. The bacteria in the gut might aid in digestion. Chitin from the food, and a large proportion of the oil, passed out with the faeces, often causing an oily film on the water in which herring were feeding.
5. Time for digestion The length of time that the food stays in the gut depends on the
temperature (Fig. 6) and on the feeding history of the fish. Blaxter and Holliday (1958) showed that food took longest to pass through the gut when fish were starved before andfor after the test feed. Battle et al. (1936) suggested that small herring retained food in the gut longer than large herring.
D. Fat storage and metabolism 1. Cyclical changes
The clupeids are seasonal feeders, building up large stores of fat to sustain them when food is in poor supply. The overall changes in body fat of the herring in relation to feeding, age, size, race and maturity stage have been extensively studied (for bibliography see Wood, 1958). Similar studies have been made on Sardinu pilchardus (Hickling, 1945) and Sprattw sprattw (Liihmann, 1952). A good deal of individual variation exists, but it is the intensity of feeding that is the overriding factor in determining the fat reserves. The spring and autumn plankton peaks provide potentially rich feeding and whether the fish make use of this food depends to a large extent on the condition of the gonads, fish with well developed gonads feeding only to a very small extent. Thus the autumn-spawning herring feed and lay down fat reserves in
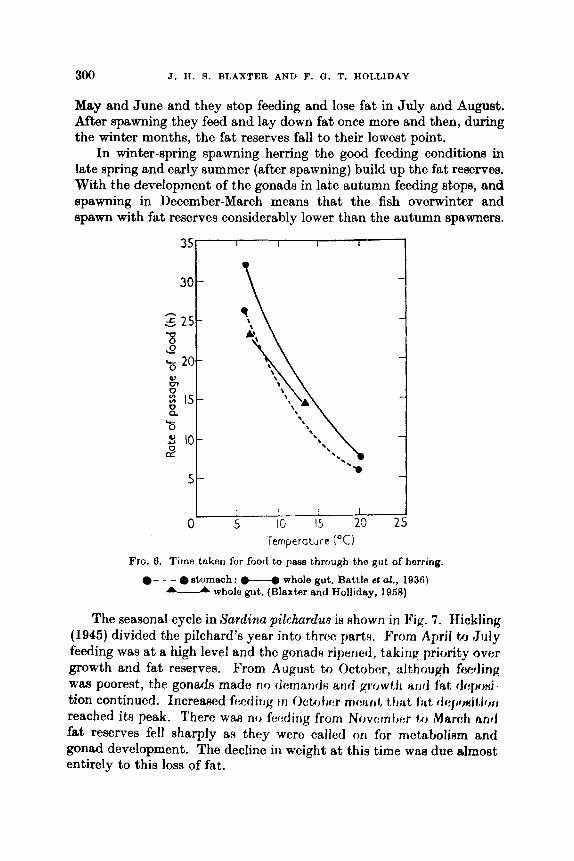
300 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
May and June and they stop feeding and lose fat in July and August. After spawning they feed and lay down fat once more and then, during the winter months, the fat reserves fall to their lowest point.
In winter-spring spawning herring the good feeding conditions in late spring and early summer (after spawning) build up the fat reserves. With the development of the gonads in late autumn feeding stops, and spawning in December-March means that the fish overwinter and spawn with fat reserves considerably lower than the autumn spawners.
Temperature ( "C)
FIQ. 6. Time taken for food to pwa through the gut of horring.
- - - stomach: whole gut. Battle ct al., 1936) whole gut. (Blaxter 4nd Holliday, 1958)
The seasonal cycle in 8ardina pilchardus is shown in Fig. 7. Hickling (1945) divided the pilchard's year into three parts. From April tr, J ~ d y feeding was at a high level and the gonads ripened, taking priority over growth and fat reserves. From August to October, although fwding waa poorest, the gonwh made no rlernsridH and growth arid fitt, c f q m i tiOn continued. Increamd ffzdirrg in 0chhr:r morirrt that fit, hjmifhjf1
reached its peak. There W;MI no fecding from N r i v a m t m to March aid fat reserves fell sharply as they were called on for metaboliem and gonad development. The decline in weight a t this time was due almost entirely to this loss of fat.
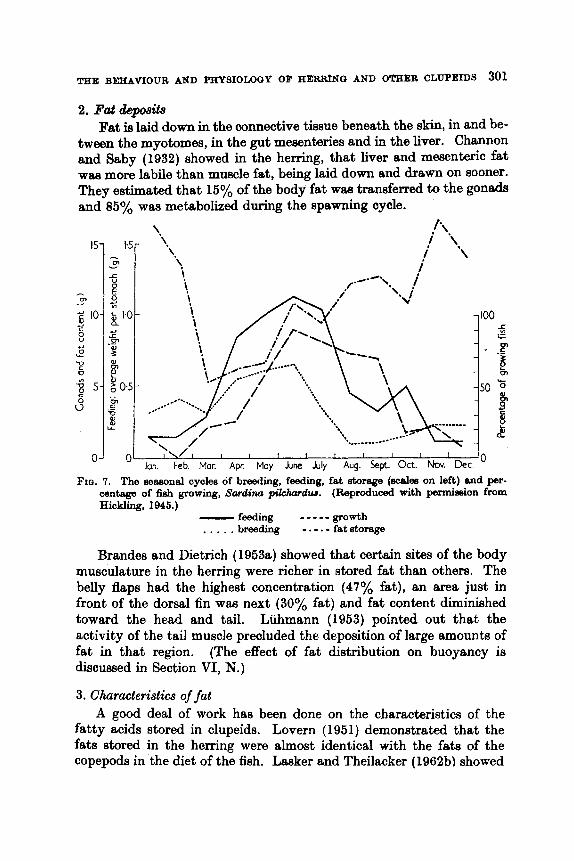
TEE B E R A ~ O U R AND PHYSXOLOOY OF HERRING AND OTHER CLUPEIDS 301
2. Fat dep8&¶ Fat is laid down in the connective tissue beneath the skin, in and be-
tween the myotomes, in the gut mesenteries and in the liver. Channon and Saby (1932) showed in the herring, that liver and mesenteric fat waa more labile than muscle fat, being laid down and drawn on sooner. They estimated that 16% of the body fat waa transferred to the gonads and 85% was metabolized during the spawning cycle.
15
- vl u
s 10 Y 0, 2 a &J
$ 5
s
0
FIQ. 7. The seasonal cycles of breeding, feeding, fat storage ( d 0 a on left) and per- (Reproduced with permiasion from centage of fish growing, Sardinu pilehardus.
Hickling, 1946.) - feeding - - - - - growth - . - . - fat storage . . . . . breeding
Brandes and Dietrich (1 953a) showed that certain sites of the body musculature in the herring were richer in stored fat than others. The belly flaps had the highest concentration (47% fat), an area just in front of the dorsal fin waa next (30% fat) and fat content diminished toward the head and tail. Luhmann (1953) pointed out that the activity of the tail muscle precluded the deposition of large amounts of fat in that region. (The effect of fat distribution on buoyancy is discussed in Section VI, N.)
3. Characteristics of fat A good deal of work has been done on the characteristics of the
fatty acids stored in clupeids. Lovern (1951) demonstrated that the fats stored in the herring were almost identical with the fats of the copepods in the diet of the fish. Lasker and Theilacker (1962bl showed

302 J. H. S. BLAXTER AND F. 0. T. HOLLIDAY
that the dietary fat of Sardinqs caerulea (derived to a large extent from Calanus) was stored largely unaltered. They also showed that both the blood lipids and the lipids deposited in the gonad were prac- tically identical, indicating that mobilization of fats from the intestine to the gonads was relatively direct. The lipids of the developing eggs caught st sea were almost identical with those from the eggs at different stages of maturation in the gonad. Changes were shown to occur in the fatty acid pattern of the lipids of the blood, gonad and mesenteries when the sardines were fed on an artificial diet. The herring does have the capacity to alter the fatty acid composition of the depot lipids, especially in regard to the degree of unsaturation of the fat, but the rapid ingestion of the highly unsaturated fats during the intense feeding periods causes a temporary swamping of the fishes' hydro- genation mechanisms (Lovern, 1938).
Cholesterol does not show the same kind of changes as the fatty acids. The level remains constant in the muscles, there being a slow transfer of cholesterol from the liver and mesenteries to the gonads, where cholesterol levels rise with the degree of maturation. The testis contains considerably more cholesterol than the ovary. Channon and $aby (1932) demonstrated that during the final stages of maturation more cholesterol was present in the gonad then could be accounted for in the depots, and they concluded that a t this time the herring waa synthesizing cholesterol.
4. General The food available for metabolism, either from an external source
or from the fat depots, obviouxly profoundly influenccu the timing and rate of many of the other physifhJgicd ar:tivitif:n. J 1, i8 riot nurprinirrg then that a t least loose correlation^ can t x made tJCtWf:<:rl fatnfm arid fecundity, maturation rate, age, growth r:tc. MoHt estimaks of thc " condition factor " of clupcids reflect to a largc: di?grcA: thc Htate of the fat stores. A good deal of work fitill remain8 to tjc donc on tho coofro- lations between fatness and gonad state, growth rtitc, ctc., but at a more fundamental level than that of Rtatistical correlation. A lot also remains to be done on the mechanisms involved in the metabolism of tho oily diet of thc clupeids, especially on the fate of the metabolic by-products.
1. Metiwds of calculation
Growth rates for thc herring in the sea have been calculated by plotting tho mean length or increment of length, against age, or by back
E. Growth

THE BEEAVIOIJR AND PEYSIOLOGY OF HERRING AND OTHER C L U P ~ S 303
Author
dculating the growth rate.of older fish from the s d e s (11, 1J. The growth constants Loo (the asymptotic length of the herring) and k (the growth constant) can then be obtained by fitting the growth equations of B e r t a l a q (1938, 1949) to the back calculabd lengths. Calculations of maintenance rations, conversioE rates, etc., are best made en herring kept in the laboratory, where feeding rate, tem- perature and interval changes in size can be meaeured. Savage (1931), however, correlated the increase in length of North Sea herring with the amount of food taken.
Remlukll Maintenance ration wet weight Species
2. bdaintemnce, growth and feeding rater,
am e u m m ~ e d in Table VII. Experimental studies on the maintenanoe requirementa of clupeida
Blaxter and Holliday
Holliday (unpub.)
Takabeehi and
(1958)
Hataneka (1900)
TABLE VII MAINTENANCE REQK!lBEMENTS OF SOXB SPBOIE8
Herring 200 mg/g fhh/week For 40 g fish at 12OC
Herring 120 mg/g fbh/week For 10-20 g at 11.4°C
Engrau2ia 14&210mg/gfiah/week For 14-64gfiahat ll-22°C
140 mg/g f%h/week
64 mg/g @/week
For 100 g fish at 12°C
For 61 g fbh at ll,4'C
j w w n h
The figures obtained by Blaxter and Holliday (1958) are estimates and are almost certainly too high. The later experiments (Holliday, unpublished, and Takahashi and Hatanaka, 1960) are more acclurate as they anaesthetized and accurately weighed the fish to obtain their information.
Ivlev (1960) calculated the energy expenditure of herring living in the Gulf of Finland and concluded that these fish needed to feed continuously for 15 hr a day to obtain sufficient food to meet their energy and growth demands, but presumably only at certain seasons of the year.
3. Salinity and growth Canagaratnam (1959) quoted values obtained by Hodgson (1934)
on adult herring from the Atlantic and Baltic to support his own ex- perimental evidence (on salmonids) that there was a positive correlation

304 J. H. 9. BLAXTER AND F. a. T. HOLLIDAY
between the growth of a fish and the salinity of the water in which i t lived (all other things being equal). Graham (1956) showed that marine Pornolobus had a more rapid rate of growth than the freshwater form of this fish, and Hoar (1952) thought that this was related to the high energy requirements of osmo-regulation in the freshwater environment (see Section VI, H).
4. Growth of herring in the North Sea During and after 1950 a large increase in the growth rate of the
herring of the North Sea and English Channel occurred (e.g. Ancellin, 1953; Gilis, 1954; Andersson, 1954; Cushing and Burd, 1957; Parrish and Craig, 1957). The increase in growth rate has been correlated with an increase in the abundance of Calanua in the North Sea (Cushing and Burd, 1956, 1957 ; Glover, 1958 ; Cushing, 1962). If this change in the growth rate is in fact a real one (and not an error in aging the fish, due to a change in the pattern of scale growth), it poses many physiological problems. For instance, was the herring " underfed " prior to 1950, retarded and living below its physiological limits? Has i t now reached these limits or could i t show an even higher growth rate? Waa there any increaaed pituitary activity from 19501
5. Growth and maturation Burd (1956, 1958, 1962) studied the relationship betweengrowth and
the maturation cycle. He concluded that there was a minimal length, irrespective of age, that a herring must reach before maturation of the gonad commenced. Burd termed this the " Critical Length " (21.6 cm for herring of the southern North %a). OhviouHly t-incc? the increased growth rate of 1950, herring have roached t h i H lurigtti Hoortcr tharr before 1950, and hence the age at first maturity hae been advanced (by 1 year).
6 . Effect of age and sex
Hickling (1940) showed that the male herring of the southern North Sea grew faster than the female. He found a disharmony of growth between the fish as a whole and the growth of the gonadH, uc) that an increasing strain was put on older fish as they matured. Because of their better growth rate the Atrain wae leeu on the malee, h u t Hickling found no significant difference in mortality betwc:c:ll malr: and fi:male fish at any age. However, Wynne-FXwardH (1029) nhowed that in tht, Manx herring growth of the body and the gonads was in harmony, and that spawning did not draw on the reserves of the fish more than could be replaced each year.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 305
Westman and Nigrelli (1 966) showed that in Brevowtia tyrannw the females grew faster than the males.
7 . Competition and growth The size hierarchy effect described by Brown (1957) in 8 d m o trulta
was noticed by Holliday (unpublished) in herring. There was no sign of fierce competition between the fish and no ‘‘ pock order ”, but despite an abundant food supply the variability of size soon became obvious. An increase in size variability was also reported by Kamshylov and Geraaimov (1960) who kept young herring in aquaria, and MacGregor (1959) found an inverse correlation between the population density and the length and condition factor of Sardinopa GaeruEeCl (although this may have been due to competition for a limited supply of food). An increase in size range would, of course, inevitably arise even with no difference in growth rate, assuming growth to be exponential in some young stages, if there was an initial size difference at the start of an exponential period. The size hierarchy phenomenon is another example of “ social effects ” existing between fish (another example is given in relation to the initiation of feeding in Section VI, A).
8 . Abnormalities of growth in the aquarium Farris ( 1956) obtained differences in the weight/length relationship
in Sardirwpa caerulea fed on different diets. On a high protein diet their length increased at a higher rate than the weight, but an opposite effect was obtained on a high carbohydrate diet. Farris suggested that the differences were due to new cell formation on a protein diet, but in- creased weight of existing cells on the carbohydrate diet.
Other abnormalities of growth in the aquarium can uuually be associated with the lack of space-tails become abraided, and the bones of the head distorted, by collisions with the walls of the tanks.
F. Natural mortality 1. Qeneral
Beverton and Holt (1969) reviewed the relationship between long- evity, body size, natural mortality rate and growth rate in different clupeids and in many other fish species. They showed a tendency for the natural mortality rate (M), in so far as i t was possible to measure this, and rate of growth (K) to be connectcd, faRtor-growing c1upr:ids tending therefore to die off more quickly. They concluded that in a wide range of fish species the natural span of life was adjuuted to tho time needed to complete, or nearly cornpletc, tho growth pattenl. In

306 J. H. 8. BLAXTER AND F. 0. T. HOLLIDAY
clupeids, too, the maximum length attained seemed to be positively correlated with the life span and larger alupeids eeemed to live to a greater age than fish of other species of comparable size.
Few attempts have been made to estimate the natural mortality rates of clupeids resulting from old age, internal " stress ", predation, disease and parasites. Presumably old age is of little importance in h e a d y fished stocks, though it is known, for instance, that the Atlanto- Scandian herring lives a t least to an age of 20 years, from following the progress of the 1904 year class in the Norwegian fishery. The Iifespan of other herring races and of other clupeid species seems to be con- siderably less, nearer 10-12 years, while in lsprattw 8prattua it may be 5-6 years (see review by Beverton and Holt, 1959).
Estimates of natural mortality rate may be made by plotting the relationship between fishing intensity and total mortality and extra- polating back to zero fishing intensity. Independent estimates of natural mortality would be of value in confirming such extrapolations, but unfortunately this approach has been completely negleoted in the past due to the extreme difEculty of making a census of predators and their food intake. However, in view of the fact that clupeids are so heavily preyed on, one might expect that relatively high estimates of natural mortality would be obtained.
2. Predation In particular, Russian workers have stressed the need for more
importance to be attached to studies on predation. Awerinzew (1935) described the mammals, birds and fish preying on the Barents Sea herring, and their effect on the migration pattern and capture by drift nets. Mitchell (1864, p. 37) reported the possible effect of the gannet, Sula alba, a diving bird, on herring stocks round the Scottish Coast. It was assumed that 200,000 of these birds near St. Kilda were feeding for 7 months of the year, taking five fish each per day on average. The total taken by this one colony of sea birds would thus have amounted to 214 million fish. Gunter and Christmas (1960) gave details of predation by sea birds, cetaceans and fish such at3 tuna on Brevm&&z ptrmw. The accumulation of guano on the Galapagos Islands from ma birds feeding on Sardinqs sagax off the Peruvian coast is a sign of the high level of predation there. This source of fertilizer haa now been replaced by fish meal obtained by direct capture of the fish, but predation by the birds continues.
The behaviour of captive herring kept with cod (Qdw eallarias) in a tank was described by Schiifer (1965). "he fish usually reacted 80-90 cm away from the cod, the avoiding movement of the shod

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 307
depending on its position in relation to the cod, and the angle of its approach. Loukashkin and Grant (1959) observed the effect of the kelp bass Paralabrax clathratos on Sardinops caeru2ea in aquaria and found little agitation among the sardines, none being taken by the predator. Apart from these aquarium studies, observations have been reported at aea on the effect of predators on shoals (see Section VI, Q).
3. Disease and parasites It is not proposed to deal in any detail with this problem due to the
difficulty of covering the literature. Disems of herring due to fungi, bacteria and parasites have been described by Sindermann and other workers (see Scattergood, 1957). Reports of mass mortality of clupeids are uncommon, though Sindermann (1956) described a heavy mortality of herring in the Gulf of St. Lawrence due to Ichthyosporidium infection.
The distribution of parasites may vary geographically in herring populations, for instance, Ichthyosporidium hofari, Kudoa clupeidae, larval cestodes and nematodes, and the trematode Brmhyphallw crenatwr, in herring of the St. Lawrence and Gulf of Maine (Sindermann, 1967).
G. Cmposition of the t~ssuw 1 . General
The clupeids, and especially the herring, have been extensively analysed chemically, mainly to provide information on the quality of the fish as a marketable product. As Love’s (1957) review points out, analytical figures vary enormously with age, sex, season and stock of fish. Unfortunately there are relatively few analyses aimed at eluci- dating the biological significance of the subfltanccs mea~urctd.
2. Fat and water The most variable constituent i H the hody fat,, doalt, with i t , IL
previous section. The inverfle linear relationship which cxiHtH hctwccn fat and water content (Brandes and Dietrich, 1953b) in the herring, and reported in other clupeids, is in fact an artificial relationship, apparcnt because most results have been expressed on a percentage (or other proportional) basis. An absolute fall in fat undoubtedly occurs, the percentage of other substances thus rises, although their absolute values do not. As Hickling (1946) pointed out for Sardina pilchardw the water and “ a s h ” provided the stable framework of the fish body. The herring, Saydim pilchardua and Sp.rattw sprattus draw to a small extent on protein reserves during times of starvation and gonad maturation, but protein loss makes, only a small contribution to any total weight lass. Changes in intracellular lipids would lead to biologically significant
21

308 J. H. S. BLAXTER A N D F. 0. T. HOLLIDAS
changes in the proportions of intracellular water and proteins, but 110
measurements exist.
3. Skin thickness and composition McBride et al. (1960) showed that sexually mature Clupea p a l h i i
caught in January had 40% more collagen in the body than immature fish caught in June. The changes were found to be due almost entirely to seasonal changes in collagen content of the skin and scales. Hughes (personal communication) reported a similar finding in the herring, and it would appear that collagen content of the skin is highest when the subdermal fat content is lowest. Histological studies of the skin of these herring showed a much greater thickness of skin in the early spring than in the late summer.
4 . Changes in the blood Naumov (1966) found that the haemoglobin content of the herring's
blood was highest (61-64%) during early maturation, but fell sharply (to 43%) at spawning and in the spent stages. Percentage haemoglobin increased with age and fatness. These changes may not represent actual changes in either circulating erythrocytes or their individual haemo- globin content, for any increase in the plasma volume would lower the percentage of haemoglobin per unit of blood. Estimations of blood volume and haematocrit values are clearly needed. Naumov also describes cyclical changes in the numbers and types of leucocytes with gonad maturation.
Sindermann and Mairs (1959a) studied pre- and post-spawning Pornolobus pseudoharengw (which ie anadromou) and found no change in sedimentation rake, erythrocytr: fragility, tw:rnoglotjiri c:orrb:iit, serum electrophoretic patterns hetween tbe two phawn of thf: lifc of t h o fish. Sindermann (1958), Sindermann and Maim (1958, l!KiOL, 1940) and itfairs and Sindermann (1960) studied the serology of Atlantic clupeids and compared them with Oncorhyncus nerkn (the blue back). They found that Pornolobus and Oncurhy~zce had a high degree of serological correspondence. A h~a wttw clwit$r to thi: On.corhyrt.cu~ - F'cmdobw complex than tr, ei thcr Iirevoortia or th: !wrrhg. '1'h herring was furthest serologically from any of the other spccics studied. These authors were able to use the serological properties of the blood to differentiate between different stocks of the herring. Age appeared to alter the serological patterns in the herring, as did disease. Sprague and Vrooman (1962) studied the erythrocyte antigens in Sardinops caerulea and found two systems of blood groups (indicating two reproductively isolated groups).

THE BEKAVXOUR AND PHYSIOLOGY OF HERRING AND OTEER CLUPEIDS 309
5. Nitrogenous extractives of tissues Shewan (1951) gave the proportion which nitrogenous extractives
formed of the total muscle nitrogen. The clupeid fishes, with a value of 14 to 18%, lay between the flat fish-gadoid group (9 to 14%) and the elasmobranchs (34 to 38%). (Hunter and Dauphinke (1925) found that Clupea pallaaii also occupied this position with respect to the arginase concentration of its tissues.)
Shewan (1955) found a close similarity in the composition of the basic amino acids between the herring and Ab8afi7&zJ and also that the herring (and other pelagic species) contained higher concentrations of histidine, and lower concentrations of carnosine and anserine than demersal species. It is paradoxical that the muscles of a pelagic, active fish should have low levels of carnosine and anserine; Yudaev (1949, quoted by Shewan, 1951) found that this condition was usually associated with lack of mobiliiiy and endurance. Yudaev (1950) also found that high histidine and low carnosine and anserine was character- istic of freshwater fishes. Hughes (1959a) suggested that there may be seasonal variations in the amounts of free amino a ids , especially histidine, in the herring.
Data from various authors were compiled by Shewan (1951), who compared the trimethylamine oxide content of fish from fresh and salt water. Pornolobus had less (185 mg%) than the herring (250 mgyo), but the difference between these clupeids was a good deal less than between other comparable fresh and sea water species. Seasonal variation of trimethylamine oxide content of Clupea harengus and Sprattus spattus has been reported (Ronold and Jakobsen, 1947; Hughes, 1959b). However, the presentation of the data as a proportion (mg per 100 g fish) does not allow one to decide if this is a change in absolute values or another reflexion of the loss of fat.
6. Effect of diet F m i s (1958), working with Sardinups caeruku, found that if these
fish were fed a high protein diet then the concentration of free aspartic acid in the tissues fell. It did not fall in fish fed a carbohydrate diet. Farris found that the proportion of fish having free aapctrtic acid and alanine in the tissues varied inversely with the condition of the iish, suggesting that during starvation amino acids were mobilized.
H. Salinity tolerance and osmo-regulation 1. Tolerance
The clupeids have freshwater, marine and anadromous represen- tatives. Many of the marine forms come close inshore either when

310 J. H. S. BLAXTER AND F. a. T. HOLLIDAY
Tramfor SinliriiOy
3P/, .
Oo/oo 3 hr . 3 hr .
24 hr . . 16°/00 3 hr .
16°/m 24 hr . . 50"/m 3 hr .
juvenile, or to spawn (see Sections 111, C ; IV, H). The herring of the Baltic sea spend almost their entire life in brackish water. Massman (1 954) recorded young Brevoortia tyrannue and Anchova mitchilli in completely fresh water.
The salinity tolerance of the herring was determined by Brawn (19GOd) and Holliday and Blaxtcr (1901). The lowor level lay between 5 and 6%,, the uppcr level between 40 and 45%,. Tagatz (1961) deter- mined the tolerance of Alosa to abrupt changes of salinity. The juvenile form tolerated changes from sea water to fresh water, but not the reverse. The adult survived a transfer from fresh water to sea water, but not the reverse. This difference between juvenile and adult prob- ably indicates differences in the physiological capabilities of regulation.
2. Regulation Holliday and Blaxter (1961) gave the mean freezing point of
herring blood as -0-95°C (equivalent to a salinity of 15.8%,). The value was slightly lower (- 1.Ol"C) in the winter months. When the herring were transferred to salinities within the tolerance range the blood first showed changes in freezing point consistent with the osmotic loss of water in high salinities and gain of water in the lower salinities. These changes were reversed and regulation of the blood to a value close to its
Blood Ntr'
129
66
102
116
128
128
288
--__-
TABLE VIII
IONIC COMPOSITION OF THE BLOOD OF HERRING TRANSFERRED TO
DIFFERENT SALINITIES BROM SEA WATER (34°/,0)
M i l l i .ocliii viLltirit.H/Iil.iw
I3lood c1-
123
48
92
116
I10
112
21 1
Musclo NaC
37
16
19
26
26
44
146
_- ~
Musclo C1-
24
12
14
19
19
21
9U

THE BEHAVIOUR AND PHYSIOLOQY OF HERRINQ AND OTHER CLUPEIDS 311
original took place within 24 hr. Total body weight changed in a similar way.
Unpublished data by Holliday on the ionic composition of the blood and muscle of the transferred fish (summarized in Table VIII) showed corresponding changes.
Outside the tolerance range the changes were more extreme and not reversible, the fish dying within 24 hr. It can be seen from Table VII I that muscle concentrations closely paralleled blood changes ; it would appear that muscle concentration changes are largely the result of changes in intracellular water.
3. Sites of regulation The problem facing the herring in low salinities is the excretion of
excess water through the kidney. Holliday and Blaxter (1961) found that the kidney of the herring had a very high glomerular count, being obviously well adapted to dealing with this water (see Fig. 8). Nash (1931) also found that Sadinella macrophthalmus and Sardinellu anchovia had high glomerular counts. The clupeids strongly resemble freshwater fishes in this respect. Unlike that in a freshwater fish the nephron of the herring contpins no distal tubule. This segment is normally responsible for resorption of salts from the urine in fresh- water fishes. It seems likely that the absence of this precludes salt conservation, and hence most herring cannot live in water below 5-6%, although some do, e.g., in the eastern Baltic.
The regulation of ionic composition of the tissues is almost certainly carried out through the general gill epithelium. It now seems unlikely that the acidophil cells, described by Keys and Willmer (1932) as being chloride secreting, are concerned with this function (see Parry et al., 1959). This would not affeot the general conclusion of Graham (1956), on Pornolobus, that those fish having the greater relative head size (and hence gill area) would be more efficient at ionic regulation and hence survive better in certain environments.
4 . Effects of endocrines Hoar (1952) suggested that the high degree of osmotic regulation
taking place in Pornolobus in fresh water made such a heavy demand on the hormones of the thyroid gland that growth was retarded; this situation did not exist in the marine forms of this fish, which had a higher growth rate (discussed in Section V, E).

312 J. H. S. BLAXTER AND F. 0. T. HOLLIDAY
20
h YI
0 C
> a 0 IS 5
8
v Y
51
L rn - Q, 2 - E, M 10- - rn Y
s
5 -
O O
0
08
o 8 0 0 0
- - 200
0 00
0 0
00
80 88 . * 08 -
a e 2 a
c
8 0 2 - loog
5
n
. - - 0 , t t q 8 1 3 L
0 u
l!J -
08 * 08
8Oo
080
I I I
10 20 30 40
I. Temperature, oxygen, CO, and H,S as limiting fuetorcl
(Reactions to gradients of these factors are considered in other HectionN) 1. Temperature
Kamshylov and Gerasimov (1 960) found that Mumian herring ( 2 10 cm long) died at .- 1.6"(:. llrawn ( I OROc) rnc:ritio,~c:<l t t ~ r : impclrt,anr:r: of determining Icthnl tempcraCurce on nowly caught C l i r p c i d H , a8 tho type of fat present in such fatty fish, which might be modified by an

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 313
aquarium diet, could affect the temperature tolerance. Using young Passamaquoddy herring she found that unacclimatized fish would withstand -1°C for a short time. There was a 50% mortality of unacclimatized herring 11.1-21-9 cm long at temperatures of 21.2- 19.5"C in 48 hr, the smaller fish being more tolerant. As mentioned in the section on osmo-regulation, the freezing point of herring blood is about 4-95"C ; temperatures below this would quickly freeze the blood unless super-cooling or depression of the freezing point took place.
Suehiro (1951), observing the factors causing mortality in bait &shes, found that unacclimatized Sardinops melanosticta 114-15-1 cm long had an approximate temperature range of 7-29°C ; Engraulis japonicw 5.6-13.7 cm long had a range of ll-31°C. Phillips and Radovich (1952) thought that Sardinops caerulea had an upper level of tolerance of about 26°C. Graham (1956) acclimatized Pornolobus psezcdoharengus to 5, 10, 15 and 20"C, and found the adults had an upper lethal limit (until 50% died on continual exposure) of 20-23°C (acclimatized at 10-20°C) and the yearlings of 15-23°C (acclimatized 5-9°C).
As reported in Section VI, B, low temperatures cause a reduction in both general and feeding activity, the temperature at which feeding ceases depending on the time of acclimatization.
2. Oxygen Kamshylov and Gerasimov (1960) found that young Murman
herring (? 10 cm) died at oxygen values below 2 ml/litre (30-33"/, saturation). Suehiro (1951) measured the oxygen requirements of clupeids. He reported that Sardinops melanosticta and Engraulis japonicw showed sigria of difficulty a t about 3 rnl/litro, ttic mtr! of respiration increased and t h y put thcir W)H(:H Ht)ovt: t,tm wt i tm. ' I ' h average lethal oxygen concentration for ,Yurdimp wau 2-02 ml/lit,rc and for Engraulis 1.82 ml/litre.
3. CO, and H,S Shelford and Powers (1915) used young Clupea p a k i i 6 cm long,
and found that they died in 6 min whcn t h c H,S conrx-mtration W ~ L H
7.6 ml/litre and in 159 min when the CO, concc:ntration wa8 20 ml/IJt,rc,.
J. Thebrain The physiology of the nervous system is virtually unknown in
clupeids. Lissner (1925) and Blaxter and Holliday (1968) described the brain in relation to feeding (see Fig. 9), following the similar study of Evans (1952) on cyprinid fishes. The optic lobes in the herring dominate the fore-brain, indicating the importance of sight ; the large

314 J. R. 9. BLAXTER AND F. B. T. HOLLXDAY
cerebellum, controlling movement and manoeuvrability, dominates the hind brain. The olfactory, facial and vagal lobes are small, in- dicating that chemoreception is not a dominant sense.
Acoustic tubercle
Fro. 9. Dorm1 view of the brain of the herring. (Reproduced with p e r h i o n from Blaxter and Holliday, 1958).
[ ~ i t h i H roview, ovitlcrico hm 1 ~ 1 i cotnpilcd t,o atiow tho importance of vision to clupoids. Shoaling, feeding and spawning, as well as avoidance of obstacles, seem to be behaviour patterns mainly controlled by sight.
1. Structure of the eye
The retina of Sardina pilchardus has been studied by Vilter (1950) who found a ventral fovcal ragion where the cone : rod ratio was 8 : 1. He related this to the visual acuity necessary for selective feeding on plankton. Baburina (1955), working on the ‘I common pilchard ” (2 Sardinapilchurdus), found that the cone : rod ratio varied from 22.4 : 1 to 32.7 : 1, the highest visual acuity being in the ventral posterior part of the retina, where the cones were most abundant of all. The eyes of the sardine ’) and I ‘ sprat ” were similar. Verheijen (19594 found a

THE BEEAVIODR AND PHYSIOLOGY OF HERRING AND OTHER CLITPEIDS 315
rotating action of the eyeball (nystagmus) in herring and an associated ventral foveal region where the cones were ten times as dense as elsewhere. No doubt the apparent high acuity of clupeids is an adap- tation to feeding on small plankton organisms. As the light decreases, rod vision presumably becomes dominant and acuity drops ; later, behavioix patterns associated with vision cease. Presumably the threshold light intensity at which such behaviour patterns cease will vary considerably ; the continuation of feeding, for example, will depend to a great extent on the size of the food, but this has not been worked on.
2. Vision at low light intensities The threshold light intensity at which various behaviour patterns
cease are discussed under the particular behaviour pattern concerned, e.g. shoaling, feeding. Protasov et al. (1960), however, using a flicker fusion frequency technique, found that the transition from day to “ night ” vision occurred at 1.0-0.1 lux in Engraulis encrasicholus.
3. Spectral sensitivity Some data are available on the spectral sensitivity of clupeids,
though colour vision has not yet been demonstrated. Borisov and Protasov ( 1960) showed by electrophysiological techniques that the ‘ I anchovy ” had a spectral sensitivity curve with a maximum at 500 m p . There was a Purkinje shift., the maximum when light adapted being at 560 mp. Blaxter (unpublished data), by finding the feeding threshold of young herring in different coloured light, and by other behaviour techniques, showed there was a maximum sensitivity in the blue-green, but the curve was plateau-like with a sharp fall off only in the red. A fairly high sensitivity in the violet suggests that experiments t o test the reactions of clupeids to ultra-violet light would be worth- while, though the existence of a positive reaction would depend on whether the lens would transmit short wavelength light. Fisher (personal communication) found that retinal pigment extracted from the eyes of dark-adapted herring had a maximum absorption at 500 mp.
4. Distance vision
A little is known of the distance herring can see. For instance, Williamson (1917) found in aquaria that food could be seen 30-90 cm away depending on its size, and Schkfer (1955) that herring could see 1.0-1.5 m. Blaxter et al. (1960) found a reaction distance to nets of 1.5-2.0 m.

316 J. H. 3. BLAXTER AND F. a. T. HOLLIDAY
L. Olfaction Burne (1909) described the olfactory pit of clupeids, showing it to
be a simple pit similar to that of the salmon. Olfaction is of little importance in the detection or primary selection of food by herring (Blaxter and Holliday, 1958), this being done by sight. However, Anon. (1952) reported that olfaction was 'important in the initial approach to food made by Sardinops caerulea. Lagler et al. (1962) described a hydraulic system of lymphatic sinuses which filled and emptied the nasal sacs in response to pressure changes during respira- tory movements.
Verheijen (1959b) could find no evidence of an alarm substance (" Schreckstoff ", reviewed by Schutz, 1956) in four clupeids*. It would be interesting to know whether species-specific odours exist which might help in any way to keep shoals together. Jones (1962) suggested that olfaction might be important for keeping herring together by night.
M . The labyrinth, hearing, the effect of sound, and sound production 1. Labyrinth and hearing
the labyrinth in teleosts as : The recent review of Lowenstein (1957) gave the main function of
Regulation of muscle tone Reception of angular accelerations Reception of gravitational stimuli Reception of sound
In clupeids work ha8 bccn rcHtriutcrl alrnoHt aonipletr:ly to anatomical ( l twri I i t .i ( ii I H . '1 '1 I ( 9 ( v 1 I I i I i I) r i I i i t I I'ii I I c.l,ic )I I I I I' f.1 I ( 1 I d )y ti I it I I t i w ct l i'ciiu J y bccri mentioned. It HOC'IIIH tliat c : x p ~ i ~ n c : r i t ~ ~ l work, H U C ~ a~ removal of the otoliths at the point where thc wall of the sacculus is separated from the buccal cavity by only a thin membrane, or the injection of anaes- thetics, would be possible.
In the herring the labyrinth consists of a sacculus containing the sagitta, the largest otolith. The lagena, posteriorly, contains a smaller otolith, the asteriscus, while the dorsally placed utriculus contains the third otolith, the lapillus. Arising from the utriculus are the three semicircular canals. The otoliths are in contact with mnculae receiving branches of the V I I I nerve.
In relation to sound, the connection between the labyrinth and swim bladder seems to be of great importance. O'Connell (1956) described this in Sardinops caerulea and Engraulis mordax, at the same
* Pfefler (2. veryl. Phyaiol. 43, 574 64, IR6fj) i d n o fvuiid 110 i i l ~ r r r i rutmt,nrir:a in four ClLlpeid H F I f X l e H .
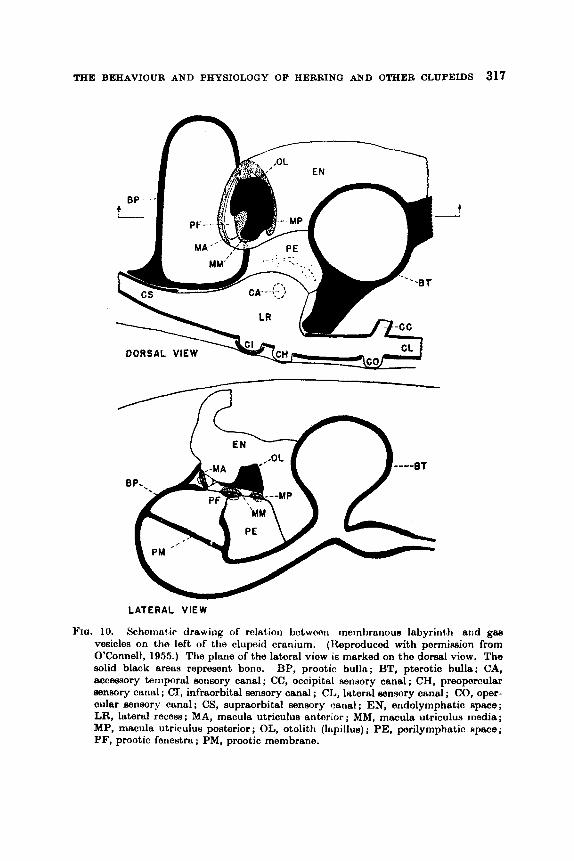
THE BEHAVIOUR AND PHYSIOLOGY OF HERBINO AND OTHER CLUPEIDS 317
n
t
LATERAL VIEW
YIO. 10. Schorrra! ir drawing of rolation botwoon mombranow labyrinth and gaa vesicles on the left of the clupnid cranium. (Reproduced with permimion from O’Connell, 1955.) The plane of the lateral view is marked on the dorsal view. The solid black areas represent bone. BP, prootic bulla; BT. pterotic bulla; CA, accessory temporal seiisory canal ; CC, occipital sensory canal ; CH, preopercular Bensory canal ; CI, infraorbital sensory canal ; CL, lateral sensory canal ; CO, oper- cular sensory canal ; CS, supraorbital sensory canal ; EN, endolymphatic space; LR, lateral recess ; MA, macula utriculus anterior ; MM, macula utriculus media ; MP, macula utriculus posterior; OL, otolith (Icqillus) ; PE, perilymphatic space; PF, prootic fenestra; PM. prootic membrane.

318 J. H. S. BLAXTER AND F. 0. T. HOLLIDAY
time reviewing the earlier work of Maier and Scheuring (1923) on herring and Wohlfahrt (1936) on Sardina pilchardus. There seem to be few essential differences between the species (see Fig. 10). The anterior end of the swim bladder bifurcates into two tubes which widen out into vesicles within the pterotic and pro-otic bullae. The pro-otio vesicles are applied to perilymph cavities which have contwt with the utricular part of the labyrinth through a pro-otic fenestra. The suggestion here is that the utriculus plays a part in hearing (Wohlfahrt, 1936 ; Verheijen, 1953), which is probably not true, for instance, of the Ostariophysi, where the swim bladder is associated with the sacculus-lagena part of the labyrinth (Lowenstein, 1957). Usually the utriculus has been considered to have an equilibrium function due to its connection with the semicircular canals as the pars superior of the labyrinth. According to Ridewood (1891) the utriculi in the herring are joined to one another by a canal which passes through the membranes of the brain just behind the pituitary fossa.
Wohlfahrt (1936) discussed in detail the possible auditory function of the utricular maculae. He considered that the lapillus also retained a residual equilibrium function. The pterotio and pro-0th bullae acted as resonators. O'Connell (1955), extending this hypothesis, saw the utriculus-swim bladder complex as widening the frequency range of the species, and perhaps their overall sensitivity, with at least a basic auditory response resting in the sacculus-lagena part of the labyrinth.
2. Response to auditory and vibrating stimuli Little is known of this, though Verheijen (1953) considered that
herring kept in aquaria could perceive up to 400 clsec at medium intensity. Blaxter and Holliday (1958), using juvenile herring in aquaria, observed the effect of sounds of different frequency played through an underwater loudspeaker on feeding behaviour. At frequen- cies from 10-100 clsec feeding temporarily stopped, the. fish reacting strongly to the starting or stopping of the stimuli. From 600-5000 clsec a " startle '' reaction waa observed but feeding was not inter- rupted.
It appears that commercial echo-sounders have no observable. effects on herring. Dow (quoted by Moulton and Baokhus, 1956) found no effect with a 50 kc/sec sounder, and Skoglund (1953) and Blaxfer and Holliday (1958) none with a 30 kclsec sounder. Bull (1961), however, has reported a response by some other teleosts to echo-sounders aa shown by training techniques. If this is confirmed it points to the need to distinguish between sensitivity and response.
Rumyantsev (1960) reported serious lesions of the body wall and

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 310
ovaries of Clwpea pallasii, which he thought were due to over exposure to high frequency transmissions from echo-sounders. It should be noted, however, that usual echo-sounding frequencies are well below the ultra- high frequencies used in sonic disintegrators, and that these observations have not been supported by experimental evidence.
3. Sounds d e by herring Recently Freytag (1961) recorded " piping '' noises made apparently
by herring and i t was intended that these noises should be played back to the fish. Such reports are not new, and according to Freytag, the Russians are already using recorded noises to lure herring into places where they would be more easily caught. The connection between " piping " noises and the release of gas bubbles from the swim bladder is not clear.
4. Lateral line Bamford (1941) demonstrated the complex lateral line system
present on the head of herring (see Fig. 11) . It was complcte a t 60 mm length and was the best developed of any teleost described to date. The canals opened to the exterior by pores and the latcral line organs were peculiar in being at the sides of the canals. Wohlfahrt (1937) found mainly a head system in Sardina pilchardus, the anterior part being innervated by the trigeminus-facialis nerve and the posterior part by the vagus. Although no canals were present on the surface of the body, except in a limited way in the scapular region, the body surface was innervated by the ramus lateralis and ramus dorsalis of the vagus. This is probably true of other clupeids. Thcre were alm frco lateral line organa on the head but thme wcre not men on t h o trunk. Verheijen (1956), howevor, by very careful handling and anaesthesia of Sardina pilchardus kept in aquaria, photographed trunk cupulae.
Both herring and Sardina pikhurdu.9 have enormous supra-orbi tal dilatations of the lateral line system which Bamford thought might be pressure receptors, because they overlay the temporal foasa and were in contact with the brain. This seems unlikely unless thc brain were enclosed in a non-compressible bony caw, the only contact with thH exterior being through tho temporal fossa.
The lateral line is considered to be reflponsiblo for localization Of objects or disturbances a t a distance-or " distant touch " ImrceIJtion (Lowenatein, 1057). It would rospond both 1.0 disturbances oroatad by other orgariiems as woll as porhaps rcflectione of dieturbances made by
own swimming movoments. Clupeids kept in the dark in aquaria do not seem to swim into the walls, which may be due to the lateral
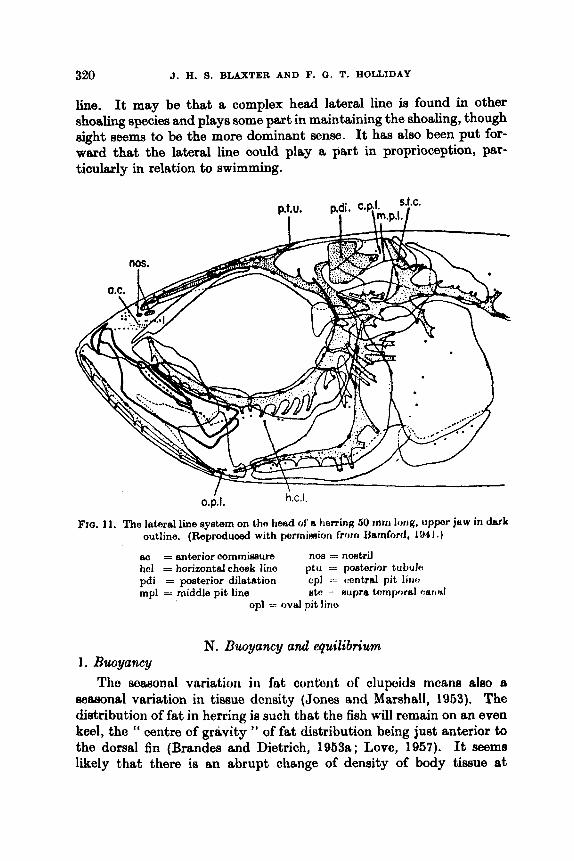
320 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
line. It may be that a complex head lateral line is found in other shoaling species and plays some part in maintaining the shoaling, though dght seems to be the more dominant sense. It has also been put for- ward that the lateral line could play a part in proprioception, par- ticularly in relation to swimming.
I 0.p.l. h.c.1.
Fro. 11. T h e lateral line ayatem on the head of a herring 50 mm long, u p p r jaw in dark outline. (Reproduced with permimian from Iknforrl, 1 V4 1 . )
BD = anterior commiasure nos = noatril hcl = horizontal cheek line ptu = posterior tubule pdi = posterior dilatation cpl := central pit l irro mpl = middle pit line ~ t c - nupra ternrmrrrl wmnl
opl r tmcil pit linu
N. Buoyancy a d equilibrium 1. Buoyancy
Tho suaclonal variation in fat aonthnt of clupoids means also 8
seasonal variation in tissue density (Jones and Marshall, 1053). The distribution of fat in herring is such that the fish will remain on an even keel, the centre of gravity ” of fat distribution being just anterior to the dorsal fin (Brandes and Dietrich, 1963a; Love, 1957). It seems likely that there is an abrupt change of density of body tissue at

THE BEHAVIOUR AND PHYSIOLOGY OF HEBRINO AND OTHER CLUPEIDS 321
spawning, for the ovary, at any rate, is mainly composed of protein- aceous yolk.
Immediate changes in density such as those required during vertical migration, or during movement into water of a different tem- perature or salinity, may be controlled by the swim bladder or perhaps by fin movements. The function of the swim bladder may be deduced from its structure. This has been described by many earlier workers, but especially in herring and Sprattm sprattus by Maier and Schcuring (1923) and in Sardinops caerulea arid Enyraulis m d a x by O’Connell (1966). The clupeids are pliysostomes with an open pneumatic duct between the swim bladder and the gut. In clupeids there is also an anal duct leading to the exterior, but this is not present in Engraulie.
The presence of smooth muscle in the wall of the swim bladder suggests that the volume may be controlled, but the anal duct does not seem to be equipped with a sphincter muscle while the pneumatic duct seems to be designed for peristalsis. There is no well-developed gas gland. An increase of pressure in the swim bladder, required to com- pensate for a reduction in volume at greater depths, might be brought about by secretion of gas from the epithelial lining, or perhaps by swallowing air at the surface and keeping i t under pressure by means of the muscular wall until a relaxation of the muscles would be neutral- ized by the increase of hydrostatic pressure at greater depths. O’Connell suggefited that the pneumatic duct might pump in air (from outside or perhaps from decomposition products in the gut), while excluding food particles and parasites by peristaltic action. The anal duct might collapse if internal pressure was too low and act as a safety valve if too high. The release of gas from fifih rifling to the Rurface ifl a wc?ll known phenomenon observed hy finhcrmcrr . Vdwi jw1 ( 1 Ijh:$) f i J t 1 r d t h i t l h
specimen of Sprattw spraltus relrmccl gas hi1 hblofi from thc, arisl oparting after 70 sec when the preRsuro was rodaocd st n rate of 40 mm Hglmin. Relcafie of gas ~ I H O occurs tlirough tho yrieiimntic ducts where an anal clrrot is present (Brawn, 1962), but whcther gm might diffuse through the swim bladder wall is not known.
Jones and Marshall (1963) discussed the high sinking factor (Density of fish x 100
Density of water ---) of herring (reported by Magnan to be 1034
compared with the range 980-1013 for most other fish) and considered that the value should be redetermined. This high sinking factor will, presumably, vary greatly with the fat content m d an average value would be hard to arrive at. If i t is usually as high as this, the fish would require to make considerable compensation to maintain a level in the water. Brawn (1962) found a mean sinking factor of 1003 for herring

322 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
and thought that the value obtained by Magnan was high due to loss of gas from the swim bladder. Brawn (1960s) watched by television the behaviour of herring lowered and raised in an underwater oage, and found that descent seemed to be mainly by sinking. Presumably to d o this they would have to be dense to start with, this density and the speed' of descent increasing as the swim bladder became compreaeed, unless there was some compensation by the swim bladder.
2. Equilibrium* Fat deposits are such (Foster, quoted by Love, 1967) that there is
n o difference in buoyancy of the head and tail regions which might give equilibrium difficulties. Maintenance of equilibrium is presumably by mema of the labyrinth, but a dorsal light reaction may also be present. Clupeida kept in aquaria often start to swim vertically when the light is reduced to a low level, this being later compensated for ; Loukashkin a n d Grant (1959) reported this in Surdimps caerulea, and Radakov and Solovyev (1959) and Zaitsev and Radakov (1960) observed herring by night from a submarine to be sometimes vertical.
0. Swimming 1. Rheotaxia and swimming ability
The ability of clupeids to stem currents is well known, but it Seems that this depends on the presence of visual or tactile reference points (Brawn, 1960a). She showed that herring 15-30 cm long, kept in an underwater cage and watched by television, ceased to swim against the current at low light intensities. Jones (1962) a h questioned the ability of herring shoals in mid-water to stem currents, although it might be possible in a deep shoal for the fish near the bottom to act &B
markers for other fish above them. His observations on the movements of herring in relation to tide were not conclusive enough to show whether the stemming was maintained in darkness after the fish lefi the bottom.
The maximum and cruising speeds of some clupeida have been measured. Blaxter and Dickson (1959) found that captive herring ranging from 1-26 cm swam at speeds (for distances at leaat ten times their body length) of 2-200 cm/sec (temperature 6-18OC). Herring 20-25 cm long could swim at near their maximum speed for 1000 lengths. Brawn (1960a) (as part of the recent Paasamaquoddy herring investigations) used freshly caught fish in the underwater cage, which W a a towed by a research vessel, and found that herring 15-27 cm long at ll-12°C had maximum speeds of 90-143 cm/sec (maintained by 60%
Sea sleo Section VI, Q.

"HE BEHAVIOUR AND PHYSlOLOOY OF HERRING AND OTHER CLUPEIDS 323
of fish for 1 min). Boyar (1961) (also as part of these investigations) found that captive herring, 6.0-21.9 cm long, in an annular rotating trough, maintained a maximum speed for 30 sec of 69-132 cm/sec at temperatures ranging from 1.4-643"C and 9.5-11-2"C. Boyar dso measured the endurance of fish of different lengths in different water velocities. These results are shown in Fig. 12.
Schiirfe (1 960), from echo-sounding records, found that shoals of herring seemed to be keeping ahead of a trawl moving at 3 knots (170 cm/sec), and that the best catches were obtained at trawling speeds of 3-6-44 knots (200-250 cmlsec).
Length (cm)
velocities (re-drawn with permiwnion from Royar, 1 gel ) . FIQ. 12. Calculated endurance of herring of differont length in diffwmt, wntnt
In contrast Stringham (1 924) observed Pomolobw pseudoharengu.4 of unknown length making headway against a current 9f 900 crnlwc, but not against one of 405 cmlsec. Dow (1962) meamred the spwd of thn same species, 27.1-31.2 cm long, swimming against & very rapid fhJW of' water in an artificial sluice. In current vc:locitir?n of 36fJ 4f)(J cm/nr:c: the maximum speeds ranged from 415-486 cm/Hr:c, for periodrr of 5 - 4 880. These and the results for captive herring are plottttd in Fig. 1.3. The results are a little difficult to compare as I h W h fish wore riot captive and wem in faot thcr beet porformers of many others involved in the experiments, and thc oxtont to which the high current spced may have influenced them is not known. Cloady tho quostion is raised of how far the swimming performance of captive fish may be underestimated, and
22

324 J. H. S. BLAXTER A N D F. G . T. HOLLIDAY
also the possibility of fish used to strong currents having a very well- developed means of finding routes where the velocity of the current is minimal. This might perhaps be a function of the lateral line system.
The mechanism of clupeid locomotion has not been worked on. It is presumably little different from other groups. Recent work by Rosen (1961) has shown that fish may progress by means of their reaction to vortices created by the head. The whole body would then be
500
h
U u 2 400 U -
*++
Length (cm)
FIQ. 13. Maximum swimming apeeds of dupeids - herring (Bkxter and Dickwn, I M I J )
Mxt for further detaila).
0 herring (Hrawn, l Y 6 k ) x herring (Boyer, 1961) + Pomolobuu (Dow, 1902) + Pomolobm of urtspecified h g t h (SYtringhem, 1924)
involved in locomotion rather than the tail alone, t h r : rcmtiori bring similar to the movement of fiRh on H, peg board af! fh:ficrihl by ( h y (see in Rosen).
2. The skebton and swimming
Hardcr (1953) rcportod on tho chnngo in proportionR of the cross- scctiotinl area of hcrring occupicd by gut, skcleton and muscle. Tho proportions of the gut remained conRtant, tho Akcloton Yhowing a negative allometry and the muscle a positive allometry, as the fish bccamc adult. Blaxter (1962) showed how the swimming speed of

TIIE BEHAVIOUR AND PHYSIOLOGY OF HERRTNQ AND OTHER CIIJPE1T)S 3%
herring larvae suddenly increased when thc caudal fin developed. Symmons (personal communication) has drawn attention to the presence of a strong elastic longitudinal ligament ventral to, and in close aasoc- iation with, the vertebral column of almost all clupeiform species. Its skeletal importance has never been suggested before, but Symmons believes i t might be associated with fish having good powers of swim- ming and leaping. The ligament is probably capable of producing a spring effect, making rapid movements of the tail more effective by a side-to-side snapping action. She suggests that this is a source of poten- tial energy which may account, in part, for the endurance of these fish.
P. Activity The study of the levels of activity of clupeid fishes is becoming
It influences both increasingly important to the fishery biologist. fish location and fish capture.
1. Diurnal variations in activity The intriguing problem of whether the herring ever ‘ I sleeps ” was
apparently solved by Radakov and Solovyev (1969) and Zaitsev and Radakov (1960), who observed hcrring from a submarine. As night fell the herring became motionless in the surface waters, remaining at various angles to the horizontal ‘ I aa if drowsy ”. The fish became increasingly active just before dawn, and in the early morning all migrated down several hundred metres. If a searchlight was played on the motionless herring, they became active and moved away from the beam.
Blaxter et al. (1960) found that at night hwririg w w : riot t i f ~ r M lry towed ropes, or even underwater lights (though young herring may he attracted by artificial lights). It would appear that the physiological state of the herring is different by day and night.
2 . Control of activity Parry and Holliday (1960) and Holliday and Parry (1962) suggested
that the pseudobranch in fishes controlled their activity by governing in some way the availability of oxygen to the fish and it8 utilization at the tissues. Removal of the pseudobranch8 rewltcd in a reduction in metabolism coupled with an ability to survivo in conditiona of oxygm lack, fatal to normal fish. The pseudobranch waH linked hy itfi I h r d system to the choroid gland of the eye, wherc it, could t j c oontroll~:~l hy the amount of light falling on the retina. ‘I’hiH Hyntr:rn might oxplairl diurnal variations in thc activity of f h h .

320 J. H. 9. BLAXTER AND F. a. T. HOLLIDAY
Buchmann (1940) described an increase in thyroid activity in the herring a t spawning, when there was a high level of activity.
Studies on Brevoortia tyrannwr (Gray, 1954) showed that it had a very high gill surface area per unit of body weight (ten times greater than in Opeanus). Also Gray and Hall (1930) found the blood sugar level was higher-76 mg/100 ml blood, compared with 16 mg/lOO ml in Opsanue, and so was the iron content (41 mg/lOO ml, compared with 13.8 mg/lOO ml in Opaanue). Dawson (1933) reported a high level of erythrocyte precursors in the blood of Brevoortia, presumably main- taining a high proportion of functional red blood cells. Vernberg and Gray (1953) and Vernberg (1954) showed that the oxygen uptake of the intact Brevoortia was very high, as it was dso in its excised brain tissue (twice that of similar tissue from Opsanwr). The oxygen uptake of isolated muscle tissue was no higher in Brevoorth than in any other fish. It waa concluded that the nervous system imposed a higher level of activity on the muscle by direct stimulation.
&. 8hling 1. General
A shod or school is here defined as a group of fish which are polar- ized or orientated in the same direction as a reaction to one another, rather than a common reaction to an external stimulus. The fish tend to have a regular spacing and move a t about the same speed.
The general concept of shoaling waa discussed by Breder (1959) who referred to clupeids and quoted work on rather unusual social groupings in Pornolobus and Alosa. He pointed out that the shoaling bbi ta of clupeids might be to some extent correlated with the immobility of their pectoral fins; fish with mobile and flexible pectorals, being able to " back up ", had a form of locomotion less suited to a shoaling habit. The importance of the silvery skin of clupeids waa also discussed. In some conditions this might cause flashing and help to keep the ehod together in poor light; on other occasions the mirror effect might reflect the colour of the surroundings and so act as a camoufiage. Manteufel and Radakov (1960) discussed the adaptive importance of shoaling in relation to defence, feeding, migration and spawning, and mentioned clupeids. The high incidence of shoaling in these fkh may be connected with their susceptibility to predation. It is well known that they either form shod or shoal more tightly aa a result of stimuli which may be harmful, but the survival value of shoaling is by no means certain.

THE BEEAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 327
2. Survival value of shoaling Ulrich (1951) made underwater observations on herring in the
Baltic and found that they formed up into shoals in response to stimu- lation from himself or the presence of fish. Schafer (1955) also observed the disturbigg effect of predators on herring in aquaria. The pattern of movement was related to the position of cod on approach of the shoal, an indentation being caused in that part of the shoal nearest the cod. Manteufel and Radakov (1960) found that only straggling herring were taken by predators in an aquarium, tight shoals causing I' con- fusion ,' of the predators. Blaxter and Parrish (1958) reported an extreme tightening of shoaling in Sprattus sprattus preyed on by Caranx, but few of the sprats survived as a result. Allen (1920) observed a " ball " of Sardinops meruka, 2 m in diameter, under attack from diving birds. The birds caused indentation of the ball but apparently were unable to extract fish from it. Phillips and Radovich (1952) found that shoals of Sardi~wp8 and Engraulis were not overexcited by attacks from carangids and sphyraenids and only stragglers were taken. If the shoals became disorganized, they were then more prone to attack. Breder (1959) reported seeing shoals of Sardinella attacked by Caranx, without affecting the behaviour of the shoals. While most evidence has been interpreted as shoaling preventing predators picking out their prey, there is the possibility that large shoals might cause a concentration of predators in a particular area, so that the overall survival value of shoaling must remain somewhat uncertain. Shoals might, of course, act as a frightening stimulus to predators, large, tightly-packed shoals appearing as a single organism.
Shoaling may aid the fish in efwapo frorn movirig netfl, hot rwtain t y p e s of fiRhing practice, for irrntrtncf: ptirnr: miflifig, r j f t f r r m r j 0 1 1 f h presence of shoals cJf remonabh size. Mantt:ufi:l and f l a h k o v ( I W ) ) reported that it WRH more difficult to catch tightly packed shoals of herring in tho Baltic, though Galkov (1068) was of the opinion that the mobility of the herring determined their susceptibility to capture, shoaling perhaps playing a subsidiary rola. (See Section VI , V for further discussion.)
Breder (1959) and Manteufel and Radakov (1960) also pointed out the importance of transmission of stimuli by shoals. The passage of a stimulus seemed to depend on the transmission of visual movements, rather than of noise through a shoal, noises probably being absorbed in many cases. While solitary fish might well react to noise, visual stimuli would be more quickly passed through a shoal and would also be more quickly received than a sound ono, as long as the water was clear. It seeme rather unlikely that any clupeids produce an " alarm

328 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
substance ” (see Section VI, L) but this would be more effective in a shoaling species. Just how well chemical stimulation would pass through a shoal is not clear, but it would almost oertainly be slower than a visual one.
Another possible advantage of shoaling might be in the deteotion of gradients of hydrographical conditions, or of food, suoh that the part of a shoal in favourable conditions might linger, the other side moving faster and so wheeling round towards the more favourable conditions (see Section VI, R).
Shaw (1962) has recently suggested that shoaling may be the most efficient way to move through the water, turbulence being used in a reciprocal way to reduce drag in neighbouring fish.
One is the possibility of attracting predators to large concentrations of their prey, the other is the limiting effect on food supply. MacGregor (1969), for instance, found an inverse correlation between adult population size and average length and condition factor of Sardirwps cuerzkz, suggesting that there was less food available and poorer growth where the population denaity was high. 3. Structure of s b a b
This is not well known in clupeids though Schlifer (1966) obeerved that small herring were near the top in herring shoals of mixed size. This type of size delimitation is discussed by Breder (1959). Phillips and Radovich (1952) found mixed shoals of Surdinups ccleruleu and Engraulis mordas, and in European waters it is common to find mixed shoals of small herring and Spattus eprattus. It seems that clupeid shoals may be mixed both in age and species.
An interesting observation was that of Wood et al. (1955) who found no change in the shoaling behaviour of tagged herring, or of their untagged neighbours, in aquaria. 4. Size and shape of shack
Off the Norwegian coast they might be miles wide, though the more usual size in the North Sea was probably “ that of a house ”. Jakobsson (1961) found that some fishable shoals of herring in Icelandic waters contaiaed 60-200 tons of fish. Tibbo and Brawn (1960) estimated the density of a shoal off the East American coast as about 2 kg/ms, and that one shoal contained 22,000 metric tons. From a submarine, Radakov and Solovyev (1959) estimated the length of some herring shoale in the northern Atlantic to be 1500 m long. While shoals are probably in many cases amorphous, Bolster (1968) found from echo ameye in the
The disadvantages of shoaling seem to be twofold.
The size of herring shoals is discussed by Balls (1951).

THE BEIIAVIOUR AND YHYSIOLOOY OF HERRING AND OTHER CLUPEIDS 329
southern North Sea that herring shoals in strong tidal streams were cigar-shaped with their long axes parallel with tho t idd strcnms. Whether this shape is brought about by thct pattorn of tidal currciits or whether it in somo way reflects tho switiitning abilit,y of tho fish, a few being faster, a few Rlowcr, tho majority bciiig average, is not ktwwti. Runnstr~m (1941b) also found herring shouls off the Norwegian coast in the form of a long narrow streak, and Khoals of Sardina pilchardus may also be cigar-shaped (Cushing, 1960).
Glover (1957) called for a study of the factors causing dispersion or aggregation in herring. Jakobsson (1961) has given an account of the factors which might determine shoal size off Iceland. The formation of large shoals seemed to be positively correlated with the numbers of some zooplankton organisms such as Calanus, with the mixed water in the border areas between warm and cold water masses, and with shallow and irregular bottom configurations. There seemed to be a negative correlation between large shoals and fast-swimming food organisms such as fish larvae, and with deep and uniformly cold water. These gross factors which may control shoal size must be accompanied by other more intimate stimuli between individual fish.
6. Maintenance of shoals It has been extensively reported that shoals tend to break up to
feed (see Section VI, B) and that shoaling behaviour ceases in dark- ness. Loukwhkin and Grant (1959) cited many authors who had reported this.
The effect of low light intcntiticfi on clupoidfl ifl shown in Tal)lc: IX. Apart from some sea obRervationR and in Anchnviellu, an c : t i ~ r t ~ t ~ l i d ,
there seem8 to be diRpcrHion of shoalH at low light iritcmnition, niiggwtirq the extreme importance of viHiori in kceping shoals togothcr. Tho observation of Zusser (1968a), Radakov and Solovyev (1959) and Radakov 11960) of some " schools " existing in darkness needs to be further examined, in particular as to whether the fish were in true schools or unpolarized groups, and what effect the means of observation had on behaviour. It is always possible that phosphorescence in the water may help fish to keep together at night (Cushing, 1960).
Jones (1962) has also suggested that the breaking up of clupeid shoals, as observed especially in aquaria, might not necessarily be invariable. He watched the movement8 of herring 8hfJdH in tho southern North Sea by echo-sounder. Both duna: trmwi and " h k " traces were probably attributablo to hurring; tho " ( I d " traces (small groups of fish) dkappeard :It night, whilr; thc: f h M 9
traces (large groups) became more diffuw. I!(: ~ i ~ g g 0 ~ t . 4 t h n L while f,ttcl

TABLE IX EBKEOT OF L O W LIGli’l! INTENSITY ON SHOarJrJG
Author
Verheijen (1953)
SohLifer (1968) Newman, Puchkov
quoted by Loukashkin and Grant (1959)
Blaxter and Parrieh (1969)
Dragesund (1968)
Radakov and Solovyev (1959)
Blaxter et d. (1960)
Radakov (1960)
Jones (1962)
Parrish (personal communication)
Owatiri et al. (1953)
Zuaser (1958a)
Loukaahkin and Grant (1959)
Moulton (1960)
Girsa (1961)
Species
Herring
Herring Herring
Herring
Herring Sp&U-9 Bpraltlcs
Herring
Herring
Herring
Herring
Herring
Sardine ? S a r d i m p Tnelanoetieta
Caspian sprat ?Spattua sprattud phalerhl.9 Sardinops caerufea
AnchovieUa
Anchovy (?species)
Observation
Stopped shoaling in aquarium BB
light intensity dropped Ditto Stopped 3hOfding in darkncm
Stopped shoaling in 8 mD qua- rim at 0.01 to 0.001 photopic lUX
Echo traces in Norwegian waters more diffuae at night, but could be concentrated by artifickd lights
Observed from research sub- marine “Severyanka” to re- main “widely spread out, in small or even large schoole at night ”
Stopped shoaling in 226 m* aqua- rium at 1.0 to 0.1 photopic lux, depending on number of f%h and their maturity
Observed from research eub- marine “Severyanka” to be paesive and usually scattered at night but also in “ small group and shoals ”. Activity in- cr& at dawn
Echo t r m in southern North Sea more diffuse at night, but herring still appear to keep together
Echo traces in northern North Sea sometimes dense at night
Echo tram in Japanese watem more diffuse at night
Activity decreased at night, but remained in “ motionless shoals ”
Stopped shoaling in darkness
Shoals remain intact at night,
Dieperso at low light intensities
(0.1 lux) in aquarium
blinded fish still shoal
below 0.1 lux

THE BEHAWOUR AND ~HYSIOLOQY OF HERRINO AND OTHER CLUPEIDS 331
small groups disappeaqd at night, the larger ones were kept together by a chemical sense reacting to a substance produoed by the herring themselves, only large groups producing enough to keep the individuals together. This might explain why clupeids at sea could keep together in the dark, whereas they could not in aquarium tanks, where the smell would be pervasive. It w m s , however, basically unlikely that the fish could polarize by smell, and unorientated aggregations at night would be perhaps more likely. The importance of the lateral line in shoaling is completely unknown, but i t might play some part in keeping fish together in darkness.
Galkov (1958) reported that small dense schools of herring, some- times aa deep as 600 m, have been seen to migrate actively across the Norwegian current by day and night. Craig and Priestley (1961) also photographed herring by flash on a spawning ground off the west coast of Scotland and found that the fish were orientated sometimes by day and night, the light on the bottom being below threshold. These fish were near the bottom and could all have been reacting to an external stimulus, such as tide, in the same way and not to each other. Indeed Valdez and Cushing (1969) reported that layer t r w s in the southern North Sea, which were probably herring, formed up in general at the first slaok water after dusk and remained aa a layer until the first slack water after dawn. It seemed aa if the herring shoals could not break up until slack water and once dispersed in the tidal stream, could not re- form into shoals until the next slack water with good light conditions.
No evidence is available that sound might be important for clupeids keeping together, but recently Freytag (1961) has reported that herring may make sounds, and Moulton (1 960) hm suggested that Amhaviella shoals may keep together by mund.
At the present time it may be said that viRion appoam tAJ h: of grt:at importance to the maintenance of the shoal but the exhnt to which clupeid shoals may keep together, or remain polarized, as a result of intervtion between the individuals due to other stimuli must remain an open question.
The more important question for the fisheries biologist is the control of shoal size, for while herring may be present in fairly high density, i t is the aim of shoal which may decide whether certain fisheries are to be successful or not. Some factors such as spawning, predation and feeding on slow-moving organisms, as well as gradients in hydrograph- ical conditions or certain bottom configurations may keep H ~ O R I R to- gether, or even catalym the formation (Jf I q p r whoaln by thr: fnniotl of smaller ones. Other factors such &H fcedirig on ftuit-movirjg orgariirrrrrrr, darknew and uniform hydrographic conditiona may oithor tc!trd hsrtrly

33:: J . H . 8. BIAXTRIL AND Y. (1. T. HOLLIDAY
to maintain the status quo, or even cause rapid dispersion. The relative importance of shoal size and shoal numbers depends on the type of fishery and the searching power available to detect the presence of fish. Large shoals are usually more valuable to fisheries using cncircling gear, while more numerous but smaller shoals, but not too small, are more valuable to trawl and drift net fisheries.
R. Migrations 1. General
The migrations of clupeids have been studied by plotting distribution of the fish, and by internal and external tagging. Such work started on Clupea pallasii about 1930, on Sardinops cuerulea a little later and on &?lapea harengus just after the Second World War. How the migrations are controlled is not well known. Cycles of hormone activity (see Sections VI, P ; VI, X) suggest that there is a cycle of internal endocrine secretion, which may well give the fish an urge to migrate ; the question of whether this cycle is an endogenous one, or one brought about by environmental conditions, such as tcmperature or day length, is also discussed in these sections. The problem to be discussed here is how the urge to migrate is directed along certain fairly well-defined routes. Is this essentially an undirected process, m a result of kineses (random movements where the speed of movement or turning frequency depends on environmental conditions), or is it an active, orientated movement (taxis) on the part of the fish, or is it a mixture of the two?
An undirected process could come about m follows. The fixh might overwinter and remain relativoly inactive in conditicmn of low b m - perature and poor food. The end of tJih pcriofl wotlltj qq)c:irr itlg t h : fidi became more activc in roHponsc: to riHing txmpcrciturc: or innnitirm. Their movements would then be limited by unfavourable water m w e s (of low temperature or sihinity and with poor food) as well aa by the depth of water or by land masses. The fish would tend to aggregate where food was good, but as the season went by it would be necessary to postulate some factor which would direct them to the spawning ground. Assuming that a feeding phase were followed by a further spawning migration, this might be triggered off by the €at content reaching a certain value. The direction of the spawning migration might be partly controlled by non-optimal water masmH and b y aggre- gation, in the case of herring, in arcas of ~ u i t a t h rit:pt,h arid nuhhtratc: for depositing eggs. After ripawning, f h: urgc f,o migrih: woi1J4 bill,&Jt:
and the fish .~.vould tmd to drift, off with tho provtiilirig c:urrcvif.#. The active Pr0CCH.S would poatuhtsc nuch fnctorrt art ntift o r ;r:rrt,st,ic,n

THE BEHAVIOUR AND PIIYSIOLGOY OF HERRING AND OTHER CLUPEIDS 333
and contranatant migrations, as those described by Russell (1937), and the reaction in a directed way to gradients of biotic and abiotic factors.
2. Aggregations What evidence is available for clupeids aggregating in certain areas?
Most work to date has been on Atlanto-Scandian and North Sea herring. The effect of hydrographic conditions, particularly temper- ature, on the North Sea herring has been partly reviewed by Hela and Laevastu (1962). The work of Dietrich et al. (1959) showed that the herring of the North Sea collected in the core of cold bottom water ( 6 - 7 O C ) on the Fladen ground in summer and autumn. An example of their distribution in August 1955 is given in Fig. 14. The time during which they remained in this area depended on tho temperature, and tho extent of their vertical migration dopended on the position and steap- ness of tho thermocline. Rogalla and &hrliage (1960) found a some- what different picture in early summer 1959 where the herring seemed to avoid the colder bottom temperatures as they did the cold, low- salinity outflow from the Skagerak. Schubert (personal communication) found later that herring particularly concentrated where there were hydrographical discontinuities in the North Sea. Biryukov (1958) found a partial tendency for herring to collect in cool water in the North Sea (for instance in 7-5-8.5"C in June) but also for them to collect where there were the sharpest temperature gradients (see Fig. 15). Craig (1960) showed an association between rapid warming of the surface water and northerly winds with reduced catches of older herring in the Scottish summer fishery in the north-western North Sea. Steele (1961) observed that herring leaving their overwintering area in the Skagerak tended to collect along the outside of the Baltic outflow in t h late spring, and that they would follow the rnovcmontn arid hakirrg down of this barrier to some extent. I)wald (]!Nil) and I 'o lomki j (1961), for example, showed the now weU-kriowri r f h l , ~ f J r i d ~ ~ p h 4 , w w l the Norwegian herring and hydrogralhic corr4t iorrn, Lhc: finh c ~ d l f d , i t t ~ along the " Polar Prorit ".
Cjlortrly tho try(1rogrrtphio nncl pinnkton cotditioiin t m c : o r i t i w t ~ ~ ~ ~ I f ~ l d p l i L i i k t o i i , trc*itiK HO varicwl i i r q)ooicH, i H o f h i ih t)wt iti(lic*af,or of tho origitl o f wntor tiiwnoN. Tlik i H discuwd by Clovcr (1!)67) uncl Pavshtiks (1956).
I'avshtiks roviowed much of the cvidence supporting tho corre- lation between hcrring and plankton as shown by Russian workers and by British workers such as Praaer, Hardy, Henderson, Lucas, Savage and Wimpenny; to this evidence should be added Cushing's (1955) report and the work of Jakobsson (1961). There appear to be well
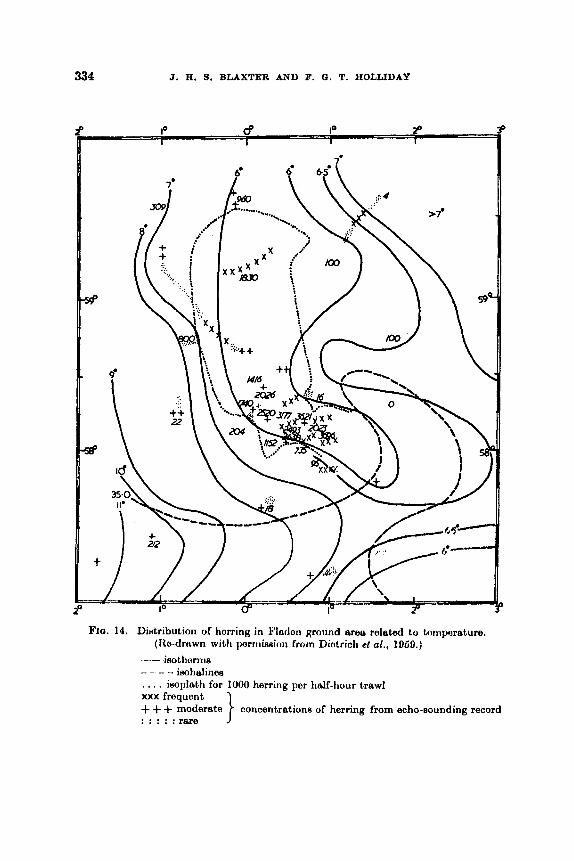
334 J. R. 9. BLAXTER AND F. 0 . T. HOLLIDAY
7'
F I ~ . 14. Dietribution of horring in Flndon ground area related to tornperaturn. (Itu-drtrwn with porrriis&n from Diotrich el al., 1069.)
- - ieotlitrriiis -- - - - isohalines . . . . isoplath for I000 herring per half-hour trawl MX frequent
. . . . :rare concentrations of herring from echo-sounding record . . . .
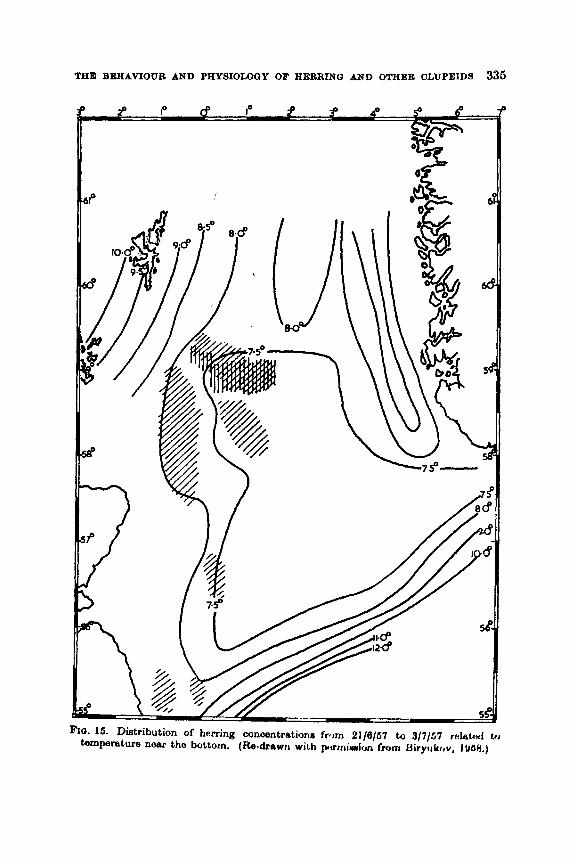
THE BEHAMOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 335
' 9 P B I" 2 9 4 0
61' m
D
FIO. 16. Distribution of herring concentratioria frwn 21/6/67 tu 3/7/67 r&hd to temperature noar the bottom. (Re-drswrr with pomiw*i~,n frclnr Uiryllkr,v, 1 U6H.j

336 J. H. 9. BLAXTICR AND F. 0. T. HOLLIDAY
established connections betwecn zooplankton, particularly Calanw, and herring, both in the North Sea and north-east Atlantic. Sometimes, however, the correlation fails and it is clear that the physiological state of the fish as well as the size and type of plankton need to be taken into consideration. Blooms of Phaeocystis seem often to be avoided, perhaps partly or wholly due to the absence of large zooplanktons in such blooms. However, the possibility of the phytoplankton being un- palatable or of gills becoming clogged by such blooms cannot be disregarded and could be tested by experiment. Pavshtiks also dis- cussed how annual fluctuations in plankton abundance could shift the feeding grounds of herring. It is difficult to do justice to the enormous amount of work done in correlating zooplankton and herring. Glover (1957) described the possible effect of oceanic influence and its con- comitant plankton changes on the Scottish East Coast and Shetland fisheries, suggesting broadly that an increased oceanic (Atlantic) influence in the northern North Sea might cause a shift of fish south- wards. Williamson (1 961), by means of statistical analysis, suggested some relationship between “ northerliness ” of the stock, oceanic influence and the abundance of certain zooplankton groups. The suggestion here is that oceanic conditions are avoided by North Sea, herring. Fraser (1961) has put forward the hypothesis that pure oceanic water is not conducive to good growth of plankton and this, and gossibly an element of ‘ I distastefulness ”, might cause the avoid- ance. On the other hand, mixed water appeared to give good conditions for zooplankton production, which might partly account for herring being found along hydrographical fronts. These fronts might also delimit areas of unfavourable water, thus acting as migratory blocks. Steele (1961) showed that euphausiids were to be found along the Baltic water front off the Norwegian coast in the spring, possibly partly caw- ing the collection of herring there. Devoid (1961), too, reported on the feeding of Norwegian herring along the “ Polar Front ”. Jakobsson (1961) showed very interesting correlations between the abundance of zooplankton and the concentration of herring off the north coast of Iceland (see Fig. 16) ; he suggested that some zooplankton would tend to cause the formation of larger shoals where it was abundant.
FIG. 16. a. 4-11/7/1957. 6 . 7-19/7/1958. The relationship between herring shoal8 off Iceland and the concentration of zooplankton. (Re-drawn with permiseion from Jakobsson, 1961.)
continuous line-path of research vessel transverse lin-oncentration of herring trecea
0 --zooplankton concentrations X -phytoplankton concentrations

Fro. 1kL. 4-11/7/1957. (See legend foot of facing page.)
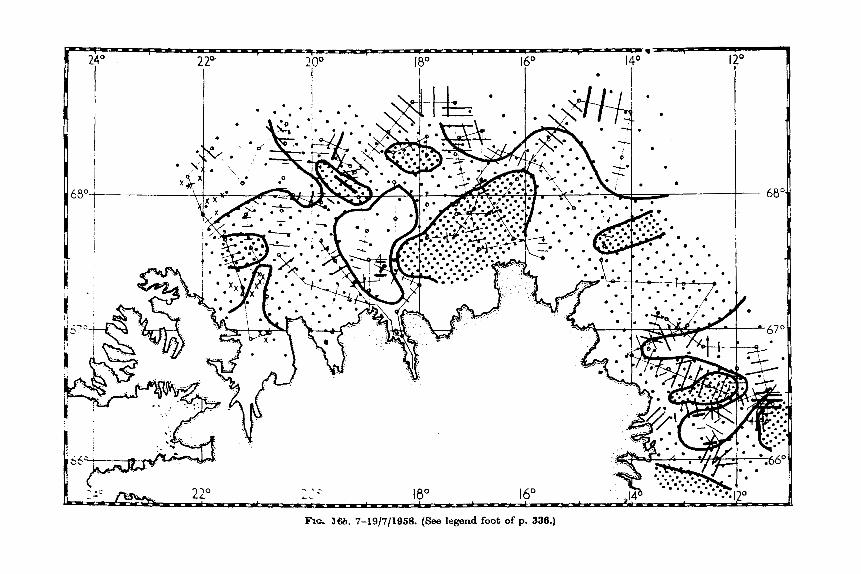
Ru, 16%. 7-19/7/1958. (See legend foot of p. 336.)

THE BEHAVIOUR AND PRYSI0u)OY OF HERRIXO AND OTHER CLUPEIDS 339
In species other than herring correlations have been found, per- ticdimly between temperature and fish concentrations. Hela and Lwvaatu (1962) reported Russian work in the Bdtio showing how Spattm eprattus was mainly caught in summer in water of 10-15"C, and Japanese workers showed that the best catches of S a r d i q s rnelanoe- tii% on southern grounds were made where the surface temperature ranged from 12-16°C. Pavshtika (1966) quoted Russian work where this species was found to delay its spawning migration if food was abundant. Phillips and Radovich (1962) elso reported that the dis- tribution of Sardinups caerulea was correlated with temperature. Hourston (1963) collected data showing that Clupea p l h i i was found in less saline water off the Britieh Columbia coast.
It seems possible, too, that long-term hydrographical changes may affect the distribution of fish on a more permanent basis. Devold (1961), for example, has shown that the annual migration to the Norwegian c m t of the Norwegian herring is becoming progreseively later eaoh year. He suggested that this might ultimately bring the fish into suoh different environmental conditions for a given stage in the life cyole that the spawning time miFht be radically altered (thie is also d i s c d under Section VI, X). Motoda and Hirano (1961) reported on long-term temperature changes (and reviewed similar reports &om the eaetern Pacific) affecting the distribution of the spawning of Clupea pallaaii. Such hydrographical changes also seem to have affected the distribution and spawning of Sardinup8 caerulea (Murphy, 1961). It is clew that temprature changes on a long-term basis may not only affect distri- bution of fish by putting certain, formerly colonized areas " out of bounds ", but may also affect growth, maturation and time of fir& maturity (see Section VI, X).
The mechanism of aggregation in areas of favourable conditiom might be explained on the theory of the kinesk, tho AAh tanding to move lees in favoured arcaR, ctnd moro rapidly in arms which are unfavourablo. In the case of feeding the shoals would, in any caso, tend to break and slow down to allow the fish to feed. It is not clear whether such mechanisms could account for the total migration pattern observed in the life cycle. Could, for instance, tho movement of northern North See herring from the edge of the Norwegian deeps to the feeding grounds, the westward and then southward movement to the spawning grounds, and the eastward drift after spawning, be only the result of a series of kineses? It is possible that the herring might be prevented from leaving the North Sea by unfavourable Atlantic water end from further westward movement by the land mass of Scotland. With an Urge to move, they would then turn muth or east to a favourable substratum 23

340 J . H. S. BLAXTER AND F. Q. T. HOLLIDAY
for spawning and then drift with the current back to the Skagerak to overwinter. But this approach may be too simple, particularly in relation to the further southward movement of the southern North Sea spawners, some of which pass through Scottish waters, and in relation to the migration of Norwegian herring from Icelandic waters.
Another factor in migration is the sudden movement of fish from productive fishing grounds often without any apparent cause. Some- times these movements may be correlated with unfavourable weather and they may also be due to lack of food or the arrival of predators.
There is a wide field open for the experimental study of phenomena relating to kineses in fish and how their speed of movement may be controlled by the environment. For instance, Loukashkin and Grant (1959) found that Sardinops caerulea swam more rapidly in unfavourable red light.
3. Directed orientation Much more difficult is the explanation of directed orientation which
may well be of great importance. Can clupeids detect, and react to, gradients of hydrographical conditions or of food or of some unspecified chemical and thus actively seek an optimal environment? Glover (1957) suggested that herring might be able to detect concentration gradients of food organisms, though it is not clear what the mechanism would be. Presumably the fish .would move slower and slower as food became more abundant, which would be more of a kinetic response. It seems unlikely that gradients of temperature or salinity would ever be abrupt enough, except perhaps vertically, for single fish to detect them along the length of their bodies. Shelford and Powers (1915) reported that Clupea pallasii in tanks could detect a temperature change of 0*6"C, but their temperature preference experiments were inconclusive. Anon. (1956) reported that Sardinops caerulea acclimated to 17-19"C usually occupied a region of temperature 18-22°C in a tall tank with a gradient from 4°C at the bottom to 34°C at the top. Engraulie w d u x acclimated to 12°C remained in 12-14"C, acclimated to 17°C in 15-18°C and acclimated to 20°C in 18-20°C. It is conceivable that a very large shoal could occupy an area where a gradient existed such that one side of the shoal experienced more favourable conditions than the other. In this event, the fish in the favourable conditions might start to move more slowly, introducing a " wheeling " effect and a turn of the shoal towards the more favourable area (Steele, 1961).
Contranatant migration seems unlikely in a pelagic fish out of contact with stationary reference points (see Section VI, 0) but a denatant drift, which would essentially be passive, seems quite probable.

THE BEHAVIOUR AND PHYSIOLOOY OF HERRINO AND OTHER CLUPEIDS 341
For instance, a t certain times of year spent herring are to be found on the Fladen ground and off the Belgian coast where the prevailing currents might have taken them after spawning.
Nothing is known of the ability of marine fish to orientate to the sun, though this ability is present in Lepmis (Haaler et al., 1968). It is also known that sunlight may have a directional component to considerable depths (e.g. to 90 my Sasaki et d., 1958). This would provide a mechanism for the southerly migration of maturing herring in the North Sea, but probably not for the migration of Norwegian herring during the winter darkness, where the herring are a t the surface a t night and 300-600 m deep by day and moving 0-7 miles in 24 hr (Devold, personal communication). Devold has shown that the initial migration of the Norwegian herring towardr, the spawning grounds might be merely the following of a hydrographioal front a t speeds of 6-7 miles per day but, in the later stages near the coast, they broke through the barrier into the Atlantic water towarda the coast, aa if by some directional urge, attaining sometimes speeds of 20-40 miles per day. Galkov (1968) reported that small schools of herring migrated across the Norwegian current by day and night a t depths down to 500 m.
One inevitably turns to the question of lemming in this respect. Wynne-Edwards (1962) suggested that tradition played an important part in the movements of fish migrating in a regular manner. If this were so, a highly deleterious effect would be produced if stocks were reduced to too low a limit, for a breakdown in the migration pattern would result. It is well known that in herring the older fish usually mature and enter the spawning fishery first, aa if they might have been leading the recruit spawners ; but how is on0 to explain even a learned ability to orientate?
4. Summary It may be said, by way of summary, that there is good evidence for
the aggregation of clupeids into defined areas, partly as controlled by land maases and partly as the result of kineees. These favourable areas may themselves be moving, giving yearly variations of the migration pattern. This, and a possible drift with the residual current aftm spawning to overwintering or feeding areas (depending on the spawning time), may partly explain migration. A more directed movement, particularly to the spawning grounds, needs also to be postulated, but it haa yet to be explained, either aa a result of a contranatant migration, or a8 a reaction to gradients of temperature or chemical factore by means of a taxis, or aa a result of an orientation to the sun or other

342 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
celestial bodies. No evidence either is forthcoming, so far, of a learning factor in migration.
S. Vertical migration 1. Occurrence and extent of vertical migration
Since the advent of the echo-sounder i t has been possible to follow the variations and extent of vertical migration in clupeids, a study which would have little meaning in the aquarium. Runnstrram (194lb) was one of the pioneers of research work with the echo-sounder. H e traced some of the characteristic movements of Atlanto-Scandian herring, showing that the shoals rose at dusk and sank in daylight, being found then at depths of 150 m or more. This has also been well demonstrated and photographed by Solovyev and Degtyarev (1957). Devold (1963) and Ryzhenko (1961) found such shoals down to 400 m in the daytime, rising to 50 m or less by night. Dragesund (1958) watched the diurnal movement of herring shoals when a ship lay over them for long periods. Later Devold (1961) reported that Atlanto- Scandian herring were being found during their migrations down to 500 m by day, tending to keep below 100 m even at night, particularly in later years.
In the North Sea the diurnal movements, though similar, are greatly restricted by the depth of water. Balls (1961) has described the move- ments in relation to depth, particularly in the southern North Sea. Biryukov (1958) reported a migration from 140-160 m up to 10-40 m in the northern North Sea. Okonski and Konkol (1957) showed how herring rose towards the aurface at night in the Skagerak. Brawn (1960b) studied the seasonal and vertical distribution of immature herring in Pmamaquoddy Bay. Tho cxtcnt of tho upward migration by night i i i thiR shallow watm variod from 5-20 m doponding on the season, tho fish boing deeper in tho month8 January to April. Tester (1943) showed diurnal vortical movements of Clupea pallmii in British Columbia waters and Nomura (1958) in Sardinopa melanoeticla in Japanese waters.
The general pattern of vcrtioal migration seems to be an upward movement at night followed by a spreading out of the fish. At dawn the fish come together and descend. Balls (1951) called the upward movement in the shallow southern North Sea (20-60 m) a I' wine glass " effect, the herring being spread over the bottom by day, rising together at dusk and spreading out again in darkneas. Cases have been reportad (see Section VI, Q) of herring shoals or aggregationn retaining their entity at night, but this seems less common and i t is not certain what the nature is of these group of fish. It is aim clear (lmxw, 1836;
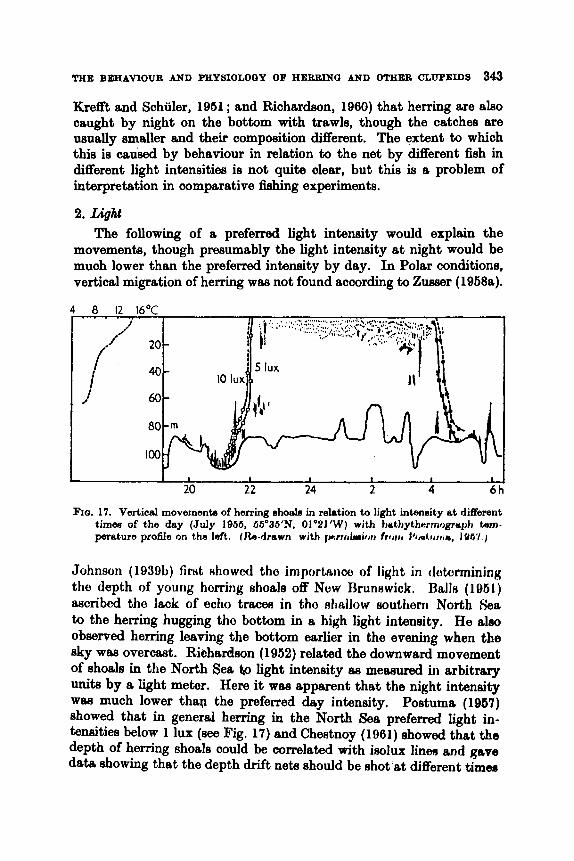
THE BEHAVIOUB AND PHYSIOLOQY OF HEBRINQ AND OTHER CLTJPEIDS 343
Krefft and Schuler, 1961 ; and Richardson, 1960) that herring are also caught by night on the bottom with trawls, though the catches are usually smaller and their composition different. The extent to which this is caused by behaviour in relation to the net by different fish in different light intensities is not quite dear, but this is a problem of interpretation in comparative fishing experiments.
2. Light The following of a preferred light intensity would explain the
movements, though presumably the light intensity at night would be much lower than the preferred intensity by day. In Polar conditions, vertical migration of herring was not found according to Zueeer (19584.
FIG. 17. Vertical movements of herring shoals in relation to light intensity at different times of the day (July 1956, 65”35‘N, OlO2J’W) with hathytherrnogrnph tern- perature profile on the left. Ilb.drawrr with perrirlYLrr) f m t r J’~dimtu, lW/ /
Johnson (1930b) firHt Hhowd tho importatico of light in dotormining tho depth of young herring shoals off Now Brunewick. Balla (1951) -scribed the lack of echo tracee in tho H ~ ~ I J O W southern North Nee to the herring hugging the bottom in a high light intensity. He elm observed herring leaving the bottom earlier in the evening when the sky was overcrtst. Richardson (1962) related the downward movement of shoals in the North $ea 40 light intensity as measured in arbitrary units by a light meter. Here it was apparent that the night intensity was much lower thap the preferred day intensity. Poetuma (1957) showed that in generd herring in the North Sea preferred light in- tensities below 1 lux (see Fig. 17) and Chestnoy (1961) showed that the depth of herring shoals could be correlated with ieolux linen and gave date showing that the depth drift neta should be shot at different timen

344 J. H. 5. BLAXTER AND F. 0. T. HOLLIDAY
of day and night. Brawn (1960b), in Passamaquoddy Bay, found no relationship between mean solar radiation and the depth of shoals of immature he~ ing . Nomura (1 958) reported Sardinops melanosticta sinking to a depth by day where the illumination was 10-1000 lux.
3. Ternperdure Schubert (1950) reported that northern North Sea herring would not
migrate upwards through thermoclines of 6 deg. C in 3 m, but would through 2 deg. C in 3 m. I n shallower depths herring were found to
Depth of upper IweI of herring shoals (lothorns)
FIO. 14. The dttpth of fiorririy W ~ O H I H rrt rliffiwmt. virrt,icd oxt,imt,H flf t h mtrfnen lrynr of horriogonnoiin wator tornperatiiro. ( I t0 drtrwn wiLh ~ i n r r i i i ~ ~ i i m from l'orrtuma, 1067.)
0 1954 . 1 I ) X x 1057
remain below a thermocline of 2 cleg. C in 6 m. Richardson (1952) found that herring passed upwards through a thermocline of 2.45 deg. C in 6 m. Postuma (1957) showed that North Sea herring would not migrate upwards beyond the point where the gradually rifling tempera- ture ceased to rise (see Fig. 18). ThuA where there waH a deep layer of water a t the surface where the temperature wau homogeneous, the herring remained deep. Solovyev and Uegtyarev (1057) found that herring in tLe Viking Bank area would not move up into cold, lem d i n e water of Baltic origin, and Devold (pmonal communication) found that spawning Atlanto-Scandian herring would not move up into low mlirlity

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 346
water of 5°C or less at the surface. Temperature may elao affect move- ment downwards. Biryukov (1968), also in the North Sea, found that herring would not move downwards by day through thermoclines which were too steep (for instance, 3 deg. C in about 10 m). They then remained by day at the depth of the thermocline, moving upwards to the surface at night. Solovyev (1959) found in the East Iceland Current that herring would not move down into colder water, and Devold observed that these herring would not move into cold water when feed- ing in the summer. Brawn (1960b) found a complex relationship be- tween the daytime depth of immature herring shoals in Passamaquoddy Bay and the water temperature (measured at 30 m). At temperatures below about 7°C there was an inverse relation between the median depth of the shoals and temperature, the herring tending to go deeper when the water was colder. Above about 7°C the herring did not stay higher by day. This could not be explained by a simple temperature- activity hypothesis, the herring sinking lower in cold water due to inactivity, because at night they rose to the surface layers, regardless of the temperature. ZusBer (1958a) reported that Atlantic and Baltic herring and the Caspian sprat Clupeonella did not swim below water of a certain temperature, and Lovetskaya (1956) that Clupeonella showed greater vertical movements at higher temperatures.
Aquarium observations (Kamshylov and Gerasimov, 1960) showed that young Murman herring would not swim from water a t 4-2°C to water of 1.8"C in a tank, while Shelford and Powers (1915), using Clupeu pallasii, found they reacted to temperature differences of 04°C.
4. Feeding Girsa (1961) related the vertical migration of marine organinmR in
general to the change in availability of their food. Richardmn (1 952) and Ryzhenko (1961) reported vertical migration in herring which were not feeding. However, Ryzhenko also described herring which may have been following luminescent patches of plankton. It wemH un- likely from both sea and aquarium observations (see Section VI, B) that clupeids feed by night and it is probable that feeding is at it8 peak at dusk and dawn. Why less foeding seems to be found by day is not certain.
ti. Other factors Aquarium observations on herring one year old (Biickmann et al.,
1953) suggested that they might bo prwented from vertical migration by sharp salinity gradients, and Kamshylov and Gerasimov (1980) reported that young Murman herring in aquaria would not swim

346 J. E. 8. BLAXTER AND F. Q. T. HOLLlDAY
through a sharp salinity barrier, except temporarily, to get food. It is also clear that vertical migration might also be modified by such factors as tide ; Valdez and Cushing (1959) showcd that tho hroakirig and re-forming of what was thought to be herring shoals depended on the state of tide and the amount of light. Catches of herring vary with moon phase and the depth of shoals may be lower where there is a moon (see Section VI, T).
It seems likely that the control of vertical migration is extremely complex, depending on a number of external factors, the effect of which will be modified by internal factors such aa the age and physiological state of the fish and the size of shoals. Any prediction of the depth to which shoals will rise at night or sink by day needs to bc based on data from a given area at a given time.
6. Abptive value of vertical migration Zusser (1968a) described the advantage to the fish of changing their
environment twice per day, giving them a greater choice of food and the possibility of better distribution by currents. Wynne-Edwards (1962) developed the idea that the numerical density of animals was regulated at certain specified times, in the case of plankton, when they migrated to the surface at night. This hypothesis could also be extended in a modified way to vertical migration of clupeids.
7 . #peed of vertical migration and pressure changes (see also Section VI, N) Runnstrcam (1941b) reported an upward movement of 100 m in
1 hr in Atlanto-Scandian herring and Dragesund (1958) an upward movement of sprats ( 2 ) of 16 m in 46 min. Krr:fff; and 3 c h ~ i h r (1051 j gave speeds of 75 m/hr up and 200 m/hr down in North Sea herring. On a more experimental basis, Blaxter and Parrish (1958) memured tha speed of clupeids following the vertical movements of artificial lightrc. Herring 15-25 cm long moved up at speeds up to 13.5 m/min and down at speeds up to 36 m/min ; sprats, 5-15 cm long, swam up at npeed8 up to 30 m/min. The pressure changes involved ranged from increarccn of' 16-180% and decrease of pressure from 18 to 73% for horring, and increases of 22 to 225% and decreaseR of 11 to 07%, for RpratR. Drawn (1962) put immature herring from Passamaquoddy Bay into c a p s which she raised and lowered. They withstood being lowercd from the surface to about 50 m (change of' prossure 0 to 4.7 kg/cm2) at a rate of 0.06 kg/cm2/sec, and boing rnisod to the surface again at a spoed of 0.07-0.08 kglcm21sec. The herring wcre also subjected to greatcr stressw i i i a Rtorilizer. Aftor a prosRuro incronm of 1.4 kg/cma they

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER OLUPEIDS 347
were deoompressed at’ a rate of 0.9 kg/oma/8eo without harm. Rates greater than 1.8 kg/cma/8eo were probably injurious.
There is no reason to suppose that clupeids, which are physosto- matous, should suffer from inflated swim bladders on decompression, but it is possible that decompression might cause embolisms, which could account partly for mortality of herring caught by trawl at considerable depths.
It appeared from Brawn’s (19604 work that herring in cages eank mimming horizontally and rose swimming vertically. The data on swimming speeds show that upward movement tends to be slower. This might be partly a gravitational effect and partly a need to com- peneate for decreaaing pressure, which might be a more serious problem than increasing pressure.
8. The “ &rn ”
This h w been discussed in Section VI, V, but i t is a phenomenon pa,rtioularly aaeociated with vertical migration in the shallow waters of the North Sea (Graham, 1931 ; Balls, 1961).
‘r. ~ffect of the Probably the first scientific enquiry into the lunar influence on
clupeid catches was that of Fulton (1906), who found the total herring catch in the Inveraray district of Scotland from 1860-1900 to be 293,035 crms over the new-moon period, and over the full-moon period 215,152 crans. He thought the difference might be due to fishermen fishing less at full moon as they thought the fish would me the nets. The analysis of the pre-war southern North 8ca hcrriihg catchaa hy Havago anrl Hodgson (1934) showed the striking dependence of the catch CHI thr: moon phase. Around full moon the catch might be double that a t other times, but the course of the seaaon’s fishing waa influenced by the date of the full moon. If i t was in the 2nd week of October good fiRhing could be expected for about 5 weeks. If the full moon waa near tho end of October, there tended to be only one peak of good catchee, which made the fishing more dependent on bad weather. Hickling (1947), however, found little evidence of the moon phase on the trawl fishery for herring from Milford Havcn (South Wales) from 1933-1940. It momod that if the moon waa full in the last part of Scptornher tho trawl firrhcry suffercd, but them was no effect comparablo with southern North Soa.
Since the war only Jcns (1964) has publiHhed further arialyses of herring catch statistics baaed on moon phase. He used tho Gorman lugger drift net catches in the North Sea in 1963, and found a peak

348 J. H. S. BLAXTER AND F. a . T. HOLLIDAY
about 4 days after full moon and a second lower peak one day after new moon.
Tester (1938b) analysed the catches of Clupea pallasii off Vancouver Island from 1933-1937 and found the lowest catches at new and full moon, which he thought was due to a reduction in the influx of herring into the channel during the spring tides. Clark (1956) reported that Californian sardine fishermen tied up during the full-moon period because they caught so few fish. Normally the fish were found by the phosphorescence they created and this was best observed before moonrise or after it set.
It is clear that a lunar influence on catch may reflect the behaviour of fish, or that of the fishermen. The following might be the causes of a lunar influetice on fish behaviour, with a resultant effect on the catch :
(a) ‘I’lic light of thc mooti mighl. cuuso a change in tho activity lovul of fish, particularly in shr~llow water, for instance in the southern North Sea. It might change the extent of vertical migration (particularly in deep water) or horizontal migration. These effects might vary with the physiological state of the fish, e.g. whether they were near spawning or not.
(b) The light of the moon (probably 0.5 lux or less) might raise the level of illumination near the surface sufficiently to allow the fish to see the nets (see Section VI, V), or to change the pattern of shoaling.
( c ) The tidal effect associated with the moon’s phases might also have a variable influence on the excitability of the fish, or on their horizontal or vertical migration, or o n their shoaling bchaviour.
(d) Tho light or tidal sffcct of 1.110 rnooti might affcot predator-prey rolationships.
‘l’htw cliffwent factors, comhirit:il with tho pofisihlo (!ffcct o f woutthcr and of moon 1)haso on fiHhitig pracl,icci, mako R caii~al unnlysir, of thiR typo of data vory tiifEcult. I’omihly lunar cffccta may have been smootlied out since tho war hy chatigcn in tho Icngth o f buoy ropes u~ed on the ncts, by the us0 of ccIio-Houti(lcw, or by change^ in fishing grounds.
U . Attraction to artificial lights
Published accounts of the attraction or repulsion of clUpC!idR to
1. Occurrence
artificial lights are shown in Table X.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 349
TABLE X
ATTRACTION 01 CLIJPEIDS BY ARTIFICIAL LIGHTS
Author
Krofft aid dchiibort
Day (personal
Borisov (1956)
(1960)
communication)
Blaster and Parriah (1968)
Schiiler and Krefft (1961)
Richardson (1962)
Drsgeaund (1968)
Tikhinov (1960) Ryzhenko (1961)
Richardson (1962)
Radovich and Gibbc (1964)
Borisov (1966)
Borisov (1966)
Verheijen (1956)
Blaxter and Parrish (1958)
Dregesund (1968)
Nikonorov (1968)
Specios
Young hcrring
Young herring
Young herring
Young herring
c. 10cm
6-26 cm
Adult herring
Adult herring
Adult herring
Adult herring ?Adult herring
Sardina
Young pilchardua
Sardimpa cae&
h8a. Clupeo- TW3ll.U
Clupm pauaaii
Sardim pikhurdus
Sprattua epnram
sprattw
~lupconella
Ah-, CoSpia-
aprattw
Area
North Sca
E-t Coaat of America Murmansk (Atlantic)
Scottieh inshore waters
North Sea
North Sea
Norway
Atlantic Atlantic
Southern North Sea
Eastern Pacific
Caspian
Paciflc
Mediter- ranean
Inshore Scottieh waters
Norway
Caspian
Observation
Attractd to Rrirfacc by 100 W
Attracted to surface by lights
Attracted by lights
seerchligh t
of 61000 w
Attracted by underwater lights from 1 6 1 600 W
Repelled by 40 W under- water light and 600W tiearchlight
Shoals deecend 7-10 m due to 60 W searchlight
Sometimm attracted, some- times repelled by l600W searchlight
Repelled by lights “Attacked ” lights of re-
search submarine “ Severy- enke ”
Attracted by lights
Attracted by 760 W liplhtn shove tho Hirrfwo
Attracted by light8
Attracted by light#
Attracted by lightw
Attracted by lightrc
Attracted by lights
Detailed description of re- action to liglite

360 J. R. 5. BLAXTER AND F. 0. T. HOLLIDAY
These observations were all made at sea, mainly in an attempt to develop light fisheries. Loukashkin and Grant (1959) made a detailed aquarium study of the effect of coloured and white light on Sardinops caerulea; this species preferred blue and green light and disliked red light, exhibiting a fright reaction both to this and flashing white light.
2. Varaability of the reaction While attraction to artificial lights has been frequently observed,
especially in young clupeids, such reactions will vary widely depending on the environment and the physiological state and age of the fish, BB
well as on the type of stimulus itself. Lovetskaya (1956) found 8etwoml. differences in the Caspian sprat ( 2 Clupeonella) and Shubnikov (1960) found that more ripe Clupeonella males were attracted than females. This could have been associated with the higher feeding activity of the males. Belyaeva and Nikonorov (1960) found that the behaviour to light& changed in relation to the condition of the fish, ecological factors, amount of moonlight, temperature, turbidity, current and the nature of the light source. Schuler and Krefft (1951) found that herring (1) would not pass through a thermocline of 4.63 deg. C in 6 m under the attraction of a lamp.
The intensity of the light source was found to be important by Borisov (1 955) ; higher intensities attracted Clupeonella more effectively. Of different colours used, yellow was most effective, but probably because it was the brightest light. Nikonorov and Patejev (1959) gave details of the best light condition for Clupeonella. Blaxter and Parrish (1958) found, too, that higher light intensities collected more organisms, presumably due to their greater penetration.
3. Explanation for light attraction This has been roviewed by Belyaova and Nikonorov (1960), who
suggested that ( ( curiosity ”, feeding, positive phototaxis, hypnosis, following of optimum light intensity, disorientation or conditioned reflex might be responsible. They favoured the idea of a (( testing reflex ”, referring particularly to Clupeonella. Zusser (1958b) and Borisov and Protasov (1960) considered that phototaxis was no explanation, but that attraction waa due to a conditioned response ; the fish normally associated feeding with light, to the extent that attraction to light had become a conditioned response associated with feeding, the unconditioned response. This requires to be proved experimentally. Verheijen (1958) developed the idea that attraction was due to a masa disorientation, such as that caused by lighthouses to migrating birds and insects. The mechanism of this disorientation would be aa follows :

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 351
The lamp would appear to the fish as a point source, stimulating only one eye at a time. This would give the effect of unilateral blinding, causing the fish to swim in a deviating path to the light. While this might occur in clear water and might be the explanation for those fish seen swimming in a disorienhted and disturbed manner near lights (e.g. Ryzhenko, 1901), it does not so eatlily explain the orientated swimming in the vicinity of lamps reported by some authors, nor attraction in very turbid water. It seems not unlikely that different explanations may apply to different species.
4. Lightfihen'es Scharfe (1963) has given a comprehensive review of the use of
lights by fishermen. Dragesund (1958) gave some details of this in the Norwegian fishery for Atlanto-Scandian herring, while Radovich and Gibbs (1954) described the blanket net used in Japanese fisheries for sardines. The use of lights has been developed most by Russian fishermen (Borisov, 1956) and by the Japanese. The most recent development is the use of light to attract clupeida initially into a region where they become influenced by an electric current. This leads them to the inlet of a pump, from whence they are pumped on deck. This was described by Smith (1955) on an experimental basis for young Clupeu p a k i i , and engraulids in Puget Sound. This method of fishing is now in extensive use, for instance in the Caspian, as a means of catching Clupeonella (e.g. Nikonorov, 1968). A theoretical discussion of the various factors involved in determining the optimum conditions for establishing a light fishery is given by Hels and Lwvastu (1962).
V. Reaction to nets and other ob8tack.a A review of the reaction of fish, in many inatancea clupeidn, to
nets was given by Mohr (1960), who discussed the I.U?IIWR involvcd and the importance of ecological and physiological factors and of learrning in captive fish.
1. Reaction to stationary nets Experiments at sea all point to the importance of sight in avoiding
such nets. Tokarew and Aslanova, quoted by Mohr (1960), reported the desirability of nets being as invisible as possible in the North Sea herring and in the Black Sea sardine fishorios. Aslanova gave reaction distances to nets of 4 m to a few cm, depending on turbidity and light intensity. v. Brandt (1954) showed the better catching power of Perlon nets in the Baltic herring fishery. Andreev (1066) discussed also the advantage of having nets of low visibility.

352 J. H . S. BLAXTER AND F. 0. T. HOLLIDAY
Mohr (1960, 1961) developed the idea of a relationship between quantity of fish present (and therefore shoal size) and net avoidance, small shoals avoiding nets better than large shoals, where the “ shoaling pressure ” was greater and the fish less able to perceive danger in their surroundings. The concept of the “ swim ”, a sudden panic reaction of herring, usually associated with slack tide in shallow depths of water, was discussed by Graham (1931) and Balls (1951). At the time of the swim the fish show a panic reaction due, perhaps, to the state of tide, crowding and sexual or migrating impulses, and they swim into the nets.
Aquarium observations on captive herring were made by Verheijen (1953), who mentioned the importance of sight for perceiving obstacles. Blaxter and Parrish (1059) also found that sight seemed to be the dominant sense in avoiding nets placed across a tank. The more contrast the net had (by reason of mesh size, twine thickness, or colour) in relation to the background, the more it was avoided. The herring failed to perceive monofilament nylon nets or shects of transparent plastic, suggesting an inability to react to any form of reflected sound or pressure waves from obstacles. Fish “ blinded ” by means of opaque plastic over the eyes, failed to respond to the nets. The reaction distance of normal fish was of the order of 1 m in a small tank in high light intensities. Nets which were avoided in daylight were swum through at night, and i t was possible to measure a threshold light intensity when the herring started to pass through the nets. This was between 0.001 and 0.0001 photopic lux in small tanks (4 m x 2 m) and 0.01 to 0.001 lux in much larger tanks (16 m x 13 m). Mohr (1961) did similar experiment8 with nets, but did not meaRim? n i ~ ( : h thrediold values. He found that thc monofilsrni:rrt riylott n d , ~ wf:r<: r i r A p w w i v d when the twine was 1e.m than 0.2 m m in diameter. ‘l’h H i m ( : author (1962) showed that transparent l’crlon netting of 0.3 mm arid mcnh size 4 cm was avoided by 50% of individual fish, the reaction distance being 1-10 cm. Small shoals of 40-100 fish almost never swam into the net while larger shoals frequently did, showing the effect of shoal
pressure ”. i t
2. Reaction to moving nets
This has been studied at sea by echo-soundcr and tJy indirrxd, methods such as analysis of catch from nets towed at, tlifferent H J W A ~ .
Scharfe (1955) followed the reaction of herring Hhonln to trcrwln hy echo-sounder in a small boat and observed no general waring cffwt. Springer (1957) followed the reactions of Sardinella anchovia to a trawl by underwater tclevision. The fish showed no strong fright reaction but

TJIE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 353
they increased their swimming speed slightly to avoid the different parts of the gear. Schiirfe (1960), in a review paper, reported the tendency of herring to swim downwards when frightened by a pelagic trawl, reacting 2-4 m from the headline or ground rope. At 3 knots (170 cmlsec) the herring appeared to swim ahead of the net, but at 3.5+ knots (200+ cmlsec) they were overtaken. Okonski and Konkol (1957), however, reported an upward movement by herring in the Skagerak when scared by a pelagic trawl. Clearly the direction of movement depends on the position of the shoal in relation to the net as well as on other possible factors. Galkov (1958) stressed that the capture of Atlanto-Scandian herring by pelagic trawl depended very much on their mobility, catches being best in November and December when mobility was low. Mohr (1962) compared the echo traces obtained from a transmitter on board ship with those from a transmitter on the headline of a herring trawl. The reaction of the herring differed, depending on their concentration. Small groups of fish kept about 5 m away from the headline and ground rope ; large groups appeared not to react at all, due to greater shoaling " pressure ". Mohr pointed out the variability of behaviour of herring to nets with such factors aa maturity stage, shoal sizes and time of day. Manteufel and Radakov (1960) reported that tightly packed shoals of herring were more diffi- cult to catch by trawl in the Baltic, suggesting that shoaling assists the fish in reacting to the net and avoiding it. Manteufel et al. (1958) showed that herring shoals in the Baltic wcro frightened by the boards, warps and net of a trawl, and also by the fishing vessel itself 30 m above. The shoals soon returned to their previous position after the net had passed. In contrast, Itadakov ( 1 962) obnervcd tihoal~ of ClupeoneZh duZicatu2a ; i t i t 1 Bn,!yravli.Y encm.uicho1u.u H wi i t1 r r r i trg, clpparci t t I y w i tt io tit agitation, within a trawl towed at 1.5 knots.
Lucas (1936), Krefft and Schuler (1951) and Richardson (1960) showed that trawl catches of North Sea herring were better by day, there being size and maturity differences in the catches. While vertical migration may play the principal part here, i t is also posdlh: that herring catches are poorer a t night becaufic: th: finh arc more qmw1 orif,
(if they are not shoaling), or because they arc not hc;rdcd by t h extremities of the net, the net having, therefore, a l oww cffi:ctive width.
In relation to this, the aquarium observations of Blaxter at ul. (1960) showed the importance of sight to hcrring in avoiding moving obstacles. 'L'hcy t m t d thr hrrding ability of ground ropes, floats, panels of Ilctting, underwater lights and air bubbles. The more conspicuous the ob8tacle and the greater its effective height, the better tho herding, the

364 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
fish reacting at distances usually of about 2 m and swimming on average a t distances of about 2 m in front of all types of obstacle in good light conditions. At speeds of 160 cmlsec (2.7 knots) or above, they were soon overtaken by the gear. At night, shoaling and all herding ceased, herring not responding even to strings of underwater lights. The threshold light intensity for loss of the herding response was between 0.5 and 0.05 photopic lux.
The importance of sound and vibrating and pressure stimuli to clupeids in avoiding nets is difficult to assess due to the difficulties of avoiding echoes in tanks and the difficulty of producing suitable sound sources of low frequency to simulate the sounds made by warps and boards. However, Schiirfe (1960) suggested that light netting was desirable for the maiiufacture of the pelagic Engels trawl in order to prevent too great a pressure wave in front of it. Although the main object of this would be to reduce drag, it might also reduce the stow in front of the net to which fish could respond.
3. Reaction to air bubbles The work on the Maine coast of the U.S.A. on the reaction of young
herrings to “ curtains ” of air bubbles has been summarized by Smith (1961). The shoals of fish might be led into anchored nets or kept within the confines of an estuary or bay ; now this seems to be developing into a commercial technique. According to Smith (personal com- munication), the curtain was sometimes effective at night. Due to moonlight or phosphorescence, however, the air bubbles were usually visible a t night, and it was possible that the method was ineffective on very dark nights. This would agree with the observations of Rlaxter and Parrish (1959) and Blaxter el al. ( i900) , that captivo hwring do not react to air bubble barriers in durkiieHs. Nikonorov ( I 058) found that Clupeonella in aquaria would pass through air bubble curtains in daylight.
4. Learning in net avoidance Our lack of knowledge of this has been discussed by Mohr (1960),
who quoted Aslanova’s observation of sardine avoiding an area where they had been frightened. Mohr (1961) reported some signs of learning in captive herring, which were being subjected to the presonco of notn in their aquarium. Devold (personal communication) pointed to the gradual increase of the depth of migration of the N;orwr:yian hrmirtg u it approached the coast in late winter. The main ccmpotionf, of thh stock is the 1950 year class and various explanations may hc offered to

THE BEHAVIOUR AND PHYSIOLOQY OF HERRINQ AND OTHER CLUPEIDS 365
account for this behaviour. Devold has put forward learning as a poa- sibility-the fish learning to keep lower in order to avoid the nets after frequent migrations to the coast during their lifetime. Another possibility is that the fish with a tendency to swim higher in the water might have been fished out, or there may be a hydrographical ex- planation. Polonskij (1961) found an increased warming of surface layers during this time.
5. Effect of electricity Groody et al. (1952) and Loukashkin and Grant (1964) studied the
effect of electrical fields on Sardinop caerdea. The fish orientated to face the anode and were forced to it, when subjected to pulsating direct current and rectified 60-cycle alternating current of different wave forms. The current density required to produce galvanotropism waa critical and varied inversely with the size of fish. Densities below the optimum produced little response; those above the optimum might cause temporary paralysis or death.
W . Learning Clupeids in aquaria do not seem to become tame as do many
other marine species. This may be a characteristic of shoaling species which are the prey of many other types of animals. There are apparently no reports of conditioned responses being set up in clupeids, though this would clearly be of great value in fish behaviour studies, particularly in relation to the sensitivity of clupeids to stimuli of differeiit intensity and quality (e.g. Bull, 1952). Unfortunately the locomotmy behaviour and general manoeuvrability of clupeids, arid thdr lack of marked behaviour patterns except shoaling, would mrtko the work more diffi- cult. However some Russian worker8 (mc Section VI, U) conniderorl that attraction to artificial light W&R the result of a crmditioned rewymrirwt set up by the association between light and feeding.
Clearly learning and memory could be of the greatest importance irt capture and avoidance of fishing gear. Both Dcvold and Mohr (see Section VI, V) have stressed this point, but i t is doubtful whether its importance can ever he assessed, except indirectly by fishing trials on a population of known size in an enclosed arect, where fish that had been caught could bo marked and rctleaqed.
Tho role of learning and mcmory in migration has been discussed by Wynne-Edwards (sce Section VI, R). Possibly the only way of assessing this would be in plotting the migration pttterns of a stock being reduced to a very low level and consisting of rocruit fish.
24

356 J. H. 5. BLAXTER AND F. 0. T. HOLLIDAY
X. Haturation of the gonads 1. Maturity scales
The assessmeiit of sexual maturity has been for some time part of the routine sampling of herring populations by fisheries workers. Gonads are classified into arbitrary maturity stages (I, 11, 111, etc.), by estimation of such criteria as the shape and colour of the gonad, its size in relation to that of the body cavity, size of eggs, etc. The classi- fication of Hjort (1910) was (with slight modifications) recommended for general adoption by the International Council for the Exploration of the Sea (Ehrenbaum, 1930). Jensen (1950) discussed the relative merits of the international scale, and that proposed by Johansen (1919) which was used for a while by Danish fishery biologists. Naumov (1956) discussed another scale in relation to fecundity and ovarian development. More recently Heincke’s (1898) scale (modified by Aasen, 1952) has been suggested as being preferable to the Hjort scale, in that it allows closer distinctions to be made between stages.
2. Morphology The general gonad morphology and gross micro-anatomy was
described in the herring by Cunningham (1896), Hjort (1910), Bullough (1940) and Williamson (1945).
3. Histological changes during maturation Naumov (1956), Polder (1961) and Bowers and Holliday (1961)
independently published detailed accounts of the herring gonad. Andrews (unpublished) gave a similar account for Rardinop cuerulea and Andreu and dos Santos Knto (IU57) arid (JOH ~ I L l l f A J H I’irif,o (IV57) for Sardina pilchardus. ‘I’ho herring tias n total qmwrririg, i.c. all the developirig eggs mature us or10 l ~ ~ t e I i uiid aro spawnetl together. In A9urtlinops ctiwuletc and Sarrlinn pilrhardus the eggs arc produced and rc1casc.d in siicccssivc crops throughout the scasoii. The general rlevolopinent is, however, aplmrcxitly the same for all clupcids. (a) The testis. The germ cells dcvelop in a system of coiinectivo tiseue tubules (see Fig. 19), these tubules being lined with connective tissue cells which may function as Sertoli cells. Figure 20 summarizes the changes in the relative occurrence of the germ cells at each stage of development in the herring. The increase in size of the testes is brought about by the rapid,increa,se in total numhcr of germ crdh in the gonad. Spermatozoa, first appear at Stage I I (Fig, 21). .Jwt prior 1.11 npiwrhrg (Stage VI) the tuhulks are packed with Hprrn ; aftfir Hpiiwitirtg htw tubules shrink as the conncctivc tiwue contractn. I’ltrlict, nporrn arr.

THE BEHAVIOUR AND PHYSIOWOY OF HERRINO AND OTHER CLUPEIDS 357
FIQ. 19. Diagram of the herring tatie (from Cunningham, 1896).
ingested by phagocytes. Recovery of the testis is preceded by con- solidation of the connective tissue tubules and their relining with connective tissue cells.
None of the authors working on the herring found cells that could be undisputedly called Lcydig cells (or lobule boundary cell^) aa described by Marshall and Lofts (1956) in Rprdtue 8p~dtu8.
Flu. 20. Changee in the relative occurrenm of the g e m cellr in tierring. ( J h j w h t t d with permiasion from Bowere and tioltiday, I084 .)
358 J. I€. S . BLAXTElt AND F. G . T. HOLLIDAY
x2- XI-
s-
f-
c I I
w I FIG. 21. Section of herring terrtin. (Reproduced with perminxion from Bowors and
Hollid~y, 1961.)
c-corinectivo tisHuo c o l l ~
nc 2---xecoiidary Hperrnatocyto sc 1-primary Hpermatonyto
ng---aperrnatogonia
8-8permatozoa f-fibrous connective tiwuo
(b ) The ovary. The basic structure of the herring ovary i8 d r o w r i i rr Fig. 22. The oocytes develop withiri the wptn, arrd lhow 1.fiibt rrrtAtirl+
in any one year become ~urroundetl by K foilicic w m l j o w f j of' i ~ i i oiit,w connective tiveue theca exterria and an i r t~tcr grari ~~IOHII, Irtyw of'
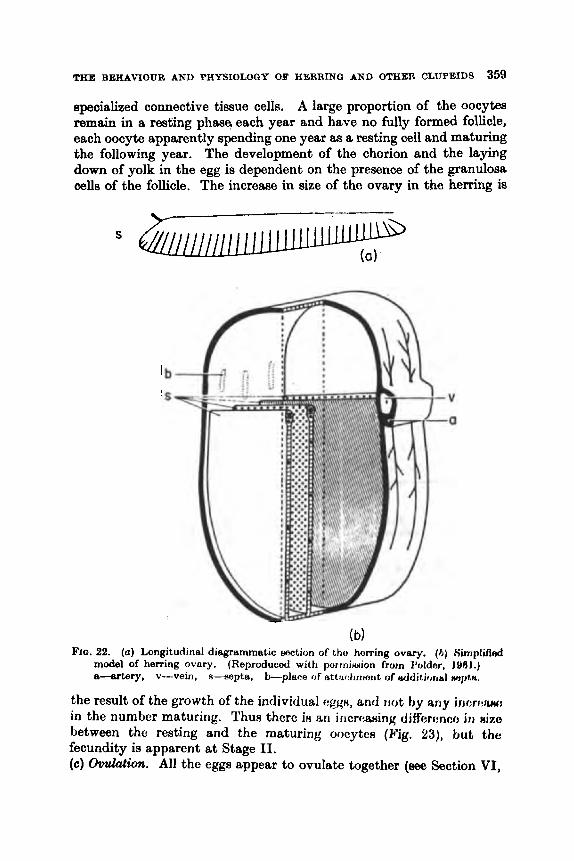
THE BEHAVIOUR AND PHYSIOLOQY OF HERRmO AND OTHER CLUPEIDS 359
specialized connective tissue cells. A large proportion of the oocytes remain in a resting pliaseq each year and have no fully formed follicle, each oocyte apparently spending one year as a resting cell and maturing the following year. The development of the chorion and the laying down of yolk in the egg is dependent on the presence of the granulosa cells of the follicle. The increase in size of the ovary in the herring is
S
(b) FIG. 22. (a) Longitudinal diagrammatic section of tho horring ovary. (a) RimpliBed
model of herring ovary. (Reproduced with porrriiwrrion from l’older, 1961.) a-artery, v-vein, s--septa, b-place of attdinrerlt of dditirmd )(rt lh~
the result of the growth of the individual egg^' and riot hy any iricrmw in the number maturing. Thus there ie an iricrcaeing diffcranw3 iri nizs between the resting and the maturing oocytes (Fig. 23), but the fecundity is apparent a t Stage 11. (c) ovulation. All the eggs appear to ovulate together (see Section VI,

360 J. H. 8. BLAXTER AND F. 0. T. HOUIDAY
X, 8). The glandular lining of the oviduct produces a secretion which bathes the ovulated eggs. Following ovulation the granulosa cells are cast off the follicles and pass to the exterior. (d) Recovery. As in the male there is a contraction and thickening of the connective tissue. A further crop of oooytes is formed to augment the resting oocytes of the previous year. ( e ) Degeneration of eggs. Pre- and post-ovulatory degeneration of eggs occurs in all clupeids that have been studied, but nothing resembling a glandular " corpus luteum " is formed as a result of this degeneration. There is so far no evidence of any endocrine activity in the herring
HJORT'S STAG€
Fro. 23. Change in diameter of oocyteH (rnm) with rnaturatictn wtagtt. (ItAprdul.ml with permiaeion from Bowurn und Hollidsy, lirf3l.)
ovary. Weismari et al. (1936) assayed extracts on immature mice but obtained no response.
4. Fecundity This has been studied in considerable detail recently a8 R character
with which to distinguish one stock of herring from anothw, r q . naxhr (1959). Marty (1956) gave an excellent Hummary of fcciirrrlity in rdatiorl to race, age and length (see Fig. 24). NagaHaki ( 1 958) Htlldi4:d fr:ciindity in Clupea pallasii and MacOregor (1950) corrcltlted fecundity with length, ago and weight in h'ardinmps caerulea.
Thorct is probably a niaximilm fecundity for a given fish that is germtically dotrrmincd, and attaiiied under perfectly suitable en- vironmetital conditions. It is likely that available food supply, age,

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 361
water temperature, etc., can all in some degree modify fecundity. Bibov (1960) found that the offspring of the Baltic herring did not have the =me fecundity as their parents when they were reared in a different environment. Anokhina (1959, 1960, 1961) reviewed fecundity as an adaptive character that might vary as environmental factors varied. She studied the relationships between fecundity and size of ova, tho
38 I I 1
36 -
34 t 30 t
Summer Islandian
22f
Atlantic -Scandinavian -c cr - A / d
Y C h e c h a n Bay
2ot I . @Kandalaskii Bay
I I I I I I I I I I I I I I
Age
2 3 4 5 6 7 8 9 1 0 1 1 I2 13 14 I
FIG. 24. Feoundity of ma homing by age and size. (Reproduaed with permission from Mnrty, 1966.)
fatness of the fish and the feeding conditions. There were some in- dividual variations, but in general there was a positive correlation between fecundity, fatness and feeding. In poor oonditions there was a larger range in the size of the egg8 produced; presumably i t is more advantageous to produce a small number of larvae from large eggs than a large number of larvae from smaller eggs.
Naumov (1956) found that the total fecundity of C a p k h ~ ~ upecim

362 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
in which the eggs were matured and spawned in batches, was higher than in the Murmansk herring which had a total spawning. Naumov attributed this to the need to spawn enough eggs at any one time to ensure a successful spawning. Andreu and dos Santos Pinto (1957) found in Sardina pilchardus that the general stock of oocytes from which the annual fecundity takes a proportien increased by the fourth power of the body length.
5. The pituitary This was studied histologically by Buchmann (1940) and Holliday
(1960a). Both described changes in vascularity and cellular activity which appeared at certain stages in the maturation cycle. Gonado- trophic activity is apparently localized in the meso-adenohypophysis. From about gonad Stage I11 onwards this region becomes increasingly vascular and the cells more abundant and larger in size. After spawning this area appears relatively inactive. Assay of the whole pituitary confirmed that the gonadotrophic activity was highest in Stages IV-VI. Holliday (1960a) studied the maturation of herring in the aquarium. The ovaries developed to about Stage 111-IV only after a high level of feeding had been maintained ; they then regressed. The testis developed normally but in miniature. Histological examination of the pituitaries showed that they were still in the immature, inactive state, suggesting that development beyond Stages 111-IV depended on a higher level of pituitary activity.
6. Timing of maturation in the sea Naumov (1956) attempted to correlate the ntntc of m:itmity in tlvr
herring with temperaturo, fkding, .dinity of th : wnfrvr t t t r t i m i r j i i t i t , r j f
solar radiation. Although wwh stage WFLR found unt1r.r fairiy well defined m d differing degrees of thcm environmental variables, it was difficult to ascribe to any of them the actual control of maturation.
Sahrhage (1954) found that southern North Sea herring were not ripe in early Deconilm 1952 when sea temperatures were 2 (leg. C below nornial, whcrcns in early I)ocembc*r 1953, when tcrnpcr:itures werc 2-3 deg. C abovo normal, nioro than 60% of thc herring were ripe. Devold (1 ‘301) suggested that t h o Norwegian herring, whose spawning was gradually becoming later each year, might eventually change their spawning time and grounds completely ; from being spring spawners off the Norwegian coast, they would spawn in early winter in the Skagerak.
Different “ races ” of herring mature and spawn at different times of the year, even though they sometimes share the same environment, for example, Iles (1962) suggested that autumn and winter upawning

THE BEHAVIOUR AND PHYSIOLOQY OF IIERRI??Q AND OTHER CLUPEIDY 363
herring were both to be found at Stage V in September on the East Anglian grounds, the winter spawners spending a further 2 to 3 months in this stage before spawning in the English Channel. One can only conclude that these fish are giving different responsea to the same environmental stimuli, implying a genetical difference (Blaxter, 1958).
The control of the process of maturation may well be due to a num- ber of essential factors interacting. An adequate food supply and a certain temperature history prior to spawning seem to be important, while the influence of day length is possible from studies on other fish, though uncertain in herring.
The age of first spawning may well depend on a “ critical length ” being reached before maturation commences, certain phases of matura- tion requiring a given level of fat before they proceed further. This critical length may be a product of the genotype and vary between Merent races (for instance, Atlanto-Scandian herring first spawn at a much greater length than many other races, and also have a greater maximum length). Superimposed on this, a certain temperature history might advance or retard maturation a little. Stimulation of the gonads by the pituitary, at least in the later stages, would be timed by these two factors as well as being possibly more basically timed by day length.
Two apparently contradictory observations are relevant here. One was the observation of herring workers at Lowestoft in the 1950’s (e.g. Cushing and Burd, 1957) that the southern North Sea herring seemed to have changed its age at first spawning from predominantly 4 years old to predominantly 3 years old, the significant points bcing increased growth rate and a decrease in age of fimt 8j)awners of e&ly one ?/ear. The implication here is firntly of it gfmtdhil tmfi!,roJ of ~ ~ ~ L W I I
ing time and of an inflexible reaction to the wivirorirnwit, rriich ~ l p rriigtit, be expected from day length as a critical controlling fac:tr,r, othcrwinc increased growth rate and maturation might have led to spawning weeks or months earlier rather than one year. Set against this must be the Norwegian observations by Devold (1961) t ha t the spawning time of the Atlanto-Scandian herring is becoming progressively later each year off tho Norwegian coast, the hypothesis of Piirrish (personal communica- tion) that southcrn North Sea herring may Romotimes tend to Rpawn progrcssivcly carlier each year aa thcy get older, and the general observation that recruit spawners spawn l i b t e r than older fish.
7. Duration of maturity stages
Naumov (1956) recorded the number of months in the year when the Murmansk herring could be found at any particular maturity stage.

364 J. H. 9. BLAXTER AND F. 0. T. HOLLIDAY
His figures do not allow the calculation of the mean duration of each stage for individual fish. Iles (personal communication) concluded that there was no simple relationship between the time of maturation and the time of spawning. The herring could remain for a variable length of time in a maturity stage, depending on the suitability of the en- vironment for further maturation.
8. Ovulation The timing of ovulation in the clupeids is closely linked to the time
of spawning, Holliday (1 960b) described ovulation in anaesthetized herring. A split appeared in the follicle and the egg was extruded through this (becoming distorted as it did so)-the process taking about 15 min. The ovulated eggs could be retained for a t least 7 days before spawning, probably maintained at this time by the secretions of the oviduct. This fluid was apparently taken up by the eggs after ovulation. Holliday (1 960b) showed that hypophysectomy prevented ovulation (and sperm release) in the herring. Injection of pituitary extracts restored these processes.
9. Control of shedding The elasticity and muscles of the gonad wall, together with the
muscles of the cloaca, exert a close control on the release of the gametes ; shedding stops if the conditions become unfavourable.
10. Hermaphroditism This has been described in Sardina pilchardus by dos Santos Pinto
(1953, 1954) and reviewed in the herring by Holliday (1962). It wm clear that in the herring the male portion8 of the gonad wwcf rnaturhg at 8 faster rate than the femalc pcmti~ri~ --fiuggwkirIy 14 h i g h r h p ! of responsiveness to the pituitary horrnonf:a by the 8pwrri:~trqrmia BnrJ
spermatocytes than by the oocyfm. However, al~awning ayparcntly did not take place until both portions of tho gonad wore fully mature. It is interesting to note that Iles (personal communication) has found that in the sea the male herring normally mature, stage for stage, faster than the females and the male herring could be in spawning condition before the female. Bowers and Holliday (1961) described the presence of ripe sperm in the testes of male herring as early as Stage 11.
Y. Spawning 1. Spawning grounds
The generJ properties of herring spawning grounds have been described, for example, by Bolster and Bridger (1057) and by Parrish

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 365
et a1. (1959). Hydrographic surveys of these grounds before and after spawning have led to many correlations between such factors as tem- perature, salinity and coastal influence and the onset of spawning.
2. Factors controiling spawning Runnstr~rm ( 194 1 b) suggested that Norwegian herring responded
to a particular type of oceanic water. Hela and Laavaatu (1962) reviewed the effect of temperature on spawning. They found that in the spring, herring would spawn in water from &12"C, and in the autumn from 8-15°C. Apparently different stocks responded to differ- ent temperature conditions. Although in general the herring spawns in relatively shallow, coastal water, Devold (personal communication) and Solovyev and Degtyarev (1957) reported herring spawning to 200-240 m. Ci'upea pallasii spawns on weeds in the littoral zone (Rounsefell, 1930), the eggs being often exposed at low tide. Ahlstrom (1954) described the spawning places, times and temperatures of Surdinope caerulea. Spawning was found from March to September but mainly in the spring in temperatures from about 13-20°C. Cushing (1960) gave values of 8-17"C for the spawning limits of Sardina pikhurdw. Most authors (e.g. Delsman, 1930 ; Devold, personal communication ; Schaeffer, 1937) agree that clupeids do not spawn in total darkness. Vision presumably plays a part in the selection of the spawning bed, and the exclusion of light stopped the spawning of herring in aquaria (Holliday, 1960b).
Dragesund (1960) kept a number of herring in a netted cove and found that spawning was delayed for 3 months and was then induced by a rise in tcmpcraturc. Outram (1051) found that CZupea pai 'hi i wore stimulatcd to Rpawn by a Ruddun iriflirx of frwh watw i n b tho tank. Holliday (1 9601) could not find any direct stimulation of the apawning of herring by either temperature or salinity fluctuations. Herring selected a spawning bed On the basis of its general pattern-which was most effective when it presented the fine broken appearance of the gravel on which they normally spawned.
3. Spawning behuviour Ewart (1884) and Holliday (1958) described the spawning behaviour
of the herring. The female pressed close to the bottom, depositing a ribbon of eggs on to the substrate, while the male fish swimming above the females released ribbons of milt into the water. There was no pairing or courtship behaviour ; i t was an indiscriminate, mam spawning. At the end of the spawning the fish appeared exhausted and showed signs of distress. Rounsefell (1930) and Schaeffer (1937) dewribcd the

366 J. H. S. BLAXTER AND F. 0. T. HOLLIDAY
spawning of Chpea p&i. The eggs were deposited in much the same way as in Clwpea hare-, but the males deposited the sperm directly on top of the eggs. No pairing of the fish waa observed and again it appeared a mass spawning. The madromous shads (Alosu sp.) spawn in rivers, on fine gravel. There appears to be a form of nuptial behaviour in these freshwater spawners (Medcof, 1957).
Less is known of the spawning of the clupeids that lay pelagic eggs. Ahlstrom (1943) deduced that Sardinops cuerulea spawned in mid- water between 2000 and 2400hr. This was confirmed by Cushing (1960) in Sardina pdc?turdue.
4. Homing Although the spawning grounds of herring, at least, remain much
the same from year to year, the extent to which herring spawn year after year in the same place is not well known, and even less is known about whether individual herring return to the grounds on which they were spawned.
A study of meristic and other characters on different spawning grounds in the southern North Sea led Zijlstra (1961) to the view that homing might take place in herring, but only tagging of spawning fish, to date largely unsuccessful, would c o n h this. However, Krefft (personal communication) who tagged spawning herring in the Elbe Estuary in April 1957, had a fish returned almost exactly 2 years later from the Skagerak. There are also cases of sudden outbursts of spawning in new places, e.g. of herring in the Texel area of the southern North Sea in 1961.
The ability of recruit spawners to return to the grounds where they themselves were spawned would dcmnnd a rc:rnarkrthlo f i d , of larval or racial I ‘ memory ”, greatly exceeding that of salmon id^. A t thin stage one might aasume that if no change in spawning time took place be- tween parents and offspring, the recruits would return, by reason of the spatial and temporal dimension of their migration pattern, and possibly by following older spawners, roughly to the area where they were spawned. Once having achieved this, memory might play aomo part 80 that the recruits themselves the following year helped to l e d youngor fish to the spawning grounds. At present it b not clear how different spawning grounds might appear to fish due h thcir t~pograptry, typo of bottom, fauna, chemical or hydrodynamic rmti~rc, IV, that w j t ;
spawning ground could be distinguished from another. W i t h ~ t r r u m evidence it would seem likely that any return would bo to tho approx- imate area of previous spawning.
Instances have been recorded, however, of spawning times and

THE B E H A ~ O U R AND PHYSIOLOOY OF RERSINCI AND OTHER CLUPEIDS 367
habits changing. Devold (1961) pointed to the gradual northward movements and latening of the spawning herring off Norway in the spring, and he suggested that this Iatening might eventually result in the spawning grounds changing from the Norwegian coast to the Skagerak, giving a “ Swedish herring period ”. Parrish (personal communication) has suggested that tu~ the southern North Sea (Downs) herring become older they may sometimes spawn farther and farther to the north,
Z. Racial characters, the genotype and the environment The nature of the racial complex in herring, Clupea pa22asii and
Sardiwps caerutea has not been considered to any great extent by trained geneticists or systematists. Genetical work hardly exists, due to the difficulty of rearing clupeids. The so-called racial characters are used in the separation of stocks for the purpose of assessing migration and the effects of fishing. A great expenditure has been made on research into scale and otolith types, body proportions and meristic characters. Reviews of racial investigations are given by Ahlstrom (1967) in Clupea palhii and by Man- (1967) in Sardinops cuerulea.
Only the meristic characters and their plasticity will be discussed further, the influence of the environment on them being shown in Table XI.
TABLE XI
EFFEIJT OP THE ENVIRONMENT ON MERISTIC CHARACTERS
Author
Runnstram (1933)
Le Gall (1936)
Runnetram (1941a)
Rpccies
Herring
Herring
Herring
PL:0
X.E. Atlantic
N.E. Atlantic
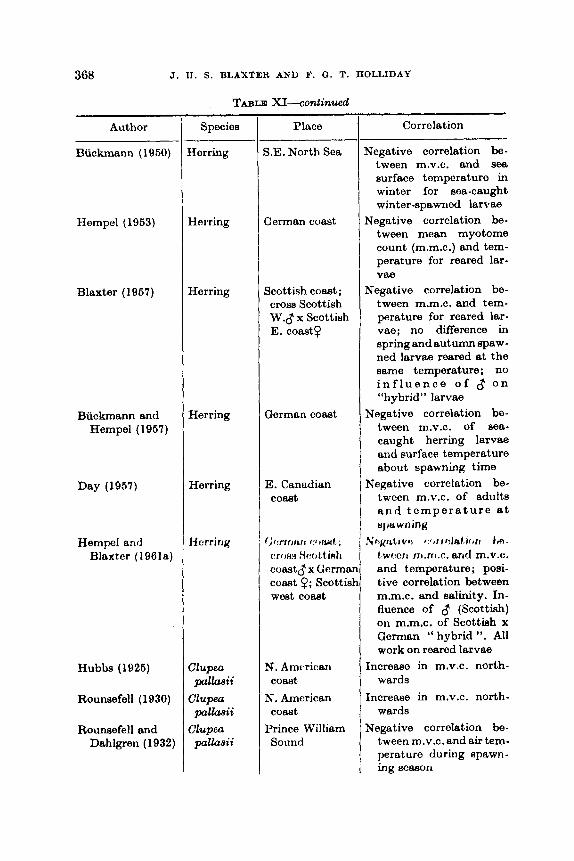
368 J. 11. S. BLAXTER AND F. Q. T. ROLLIDAY
Author
Buckmann (1960)
Hempel (1953)
Blaxter (1967)
Buckmann and Hempel (1967)
Day (1967)
Hempel and Blaxter (196la)
Hubbs (192G)
Rounsefell (1930)
Rounsefell and Dahlgren (1932)
TABLE XI-conlinued
Species
Herring
Herring
Herring
Herring
Herring
Iicrring
Clupea pllaaii
Clupea pallaaii
pallaaii Clupeu
Place
S.E. North Sea
German coast
Scottish coast; cross Scottish W.d x Scottish E. coast?
German coast
E. Canadian coast
N. Amc&an
N. American
Prince William
comt
coast
Sound
Correlation
Negative correlation be- tween m.v.c. and sea surface tomperature in winter for Boa-caught winter-spawned larvae
Negative correlation be- tween moan myotome count (m.m.c.) and tem- perature for reared lar-
Negative correlation be- tween m.m.c. and tem- perature for reared lar- vae; no difference in spring and autumn spaw- ned larvae reared at the same temperature; no i n f l u e n c e o f o n “hybrid” larvae
Negative correlation be- tween m.v.c. of 688-
caught herring larvae and surface temperature about spawning time
Negative correlation be- tween m.v.c. of adults a n d t e m p e r a t u r e a t spwning
tWPC7l rn.ni.c. and m.v.c. and temperature; posi- tive correlation between m.m.c. and salinity. In- fluence of 8 (Scottish) on m.m.c. of Scottiah x German “ hybrid ”. All work on reared larvae
Increaso in m.v.c. north- wards
Increase in m.v.c. north- wards
Negative correlation be- tween m.v.c. and air tem- perature during spawn- ing seaeon
vae
?P’hJ/JLf,IVO I t , f f h h f r ) f r l l

THE BEHAVIOUR AND w w s i o L o a Y OF HERR IN^ AND OTHER CLUPEIDS 369
Author
Tester (1937)
Teater (19384
McHugh (1942)
McHugh (quoted by Marr, 1957)
TABLE XI-mntinued
Species
Clupea pallasii
Clupea pauasii
Clupea pllasii
Sardinops caerulea
Place ,
Jap Inlet British Colum- bia
Barkley Sound British Coium- bia
British Columbia
American west coat
Correlation
Negative correlation be- tween m.v.c. and air temperature at spawn- ing seaon; gradation in vertebral count with latitude
Negative correlation be - tween m.v.c. and water temperature during spawning m w n
Negative correlation be- tween m.v.c. of EBB-
caught larvae and temp- erature at Bpawning
Latitudinal differences in m.v.c.
From this table it may be seen that both temperature and salinity may modify meristic characters. Gabriel (1944) has developed the idea that temperature affects the relationship between the processes con- trolling growth and differentiation, so that low temperatures, giving high meristic counts, would retard differentiation relative to growth. The use of meristic characters in racial investigations need not be invalidated if this plasticity is appreciated, and if the environment does not fluctuate too greatly. It seems likely that the modifications take place only during early development, cepecially h ~ f ~ r e j hatchirig ; Hempel and Blaxter (1961s) found a sensitive period 6 to 7 daya after fertilization at 10°C.
The importance of the genotype is difficult to judge. It seems likely that i t provides a general basis for the number of meristic units, the actual number being controlled by environmental factors. Larvae of widely different origin reared under similar conditions retained the difference in mean vertebral count of the parents (Hempel and Blaxter, 1961a). Egg size might provide an inherited effect, herring races with larger eggs tending to have higher mean vertebral counts, but thin does not always hold (e.g. within the North Sea). It is certainly true that larger individuals have higher vertebral countfi (Tester, 1037 ; McHuqh, 1942). Mean vertebral counts may also changc with age and time ; e.g. Buckmann and Hempel (1957) found them gradually increaniag in larvae off the German coast sampled in succewivc monthn. 7'ht:ne

370 J. H. 9. BLAXTER AND F. Q. T. HOLLIDAY
authors suggested that larvae with low counts were selected out, while Barlow (1961) suggested that selection may take place for factors connected with high or low vertebral counts, such as sensitivity to temperature changes giving different metabolic rates or in different larvae.
Apart from meristic characters, which appear to have at least some genotypic basis, Blaxter (1958) has put forward the following as geno- typically based characters-egg size, length at first spawning, spawning season, maximum length attained-to which might be added fecundity differences. These are discussed in different sections in this review, but little is known about their control, and so their value in racial and fisheries investigations remains questionable-do herring change their spawning season, can egg size and fecundity change out of all relation- ship to the normal increases which may occur with increasing age, why do Atlanto-Scandian herring spawn when older than other races? Some answers may be obtained from Russian experiments on transplanting herring from the Baltic to Aral Seas (Bibov, 19601, but the question remains whether they could occur in the normal environment. Graham (1962) suggested from studies on otolith characters that herring do not necessarily spawn at the same season as that in which they were hatched.
Interfertility of spring- and autumn-spawning herring (Scottish West and East Coasts) and of different spring-spawning races (Scottish West Coast and Baltic) has been shown by storage of sperm (Blaxter, 1957 ; Hempel and Blaxter, 1961a).
On the basis of a study of tht: characteristics of herring races, Blaxter (1958) concluded that spring- and autumn-npawning herring should in general he referred to diffr:rt:n t ~ j J e c h , A I, f , h : (Jtfivr cxf,rwrit), Russian workers consider &pea p d b x i i t,o ho a NIJ tJnp:cic:fl of' h-wenqu4. Differences in opinion as to the syetcmatio HtatuH of diffwont olupcidfi, which are also found in Surdinop~, may Heem academic, h i i t t h y ltavr: more far-reaching connotations for the k~chaviourint, rtnd phyHiOhJ~ifit. For instance, what significance ehould bc attachod to t h e wry di@w:nt, types of spawning grounds frequcnttd by pallacrii and harengus.
VII. CONULUSIONS
From the point of view of distribution and numbers the clupeids are obviously a very successful group of fishes. One reason for this is their unspecialized nature so that they aro able to deal with a wide variety of environmental conditions. For example, the primitive kidney with its high glomerular count enables them to exploit a wide range of

THE BEK~VIOUR AXD PHYSIOLOOY OF HERRWQ AND OTHER CLUPEIDS 371
salinities, and the tolerant eggs and larvae can survive in marine, littoral, estuarine and in some caws freshwater conditions. They me, however, well adapted to a pelagic, shoaling mode of life-mechanically, hydrodynamically, as well as in the biochemical composition and potential activity level of their tissues. Although they depend on a fluctuating food supply, the ability to store fat enables them to survive at all seasons. Presumably this adaptability has led to the formation of the complex racial system.
Clupeids are found in large numbers despite the level of predation to which they are subjected. Presumably the advantages of shoaling far outweigh the disadvantages. Relatively low fecundity does not seem to have been a handicap and this is true where the egg is dememl, as well as where it is pelagic. Nor does a small egg, with little yolk and a small larva dependent on external food soon after hatching, appear to be a disadvantage. The size of a spawning stock seems to have little effect on the subsequent year class strength, so that egg potential does not appear to be in any way limiting. What determines the year class strength, in herring at least, remains a mystery, and yet the fluctuations can be large.
Unfortunately, many other aspects of the herring basic biology remain a mystery-in particular its means of migration, the control of its reproductive processes and the interrelationship between different races and the extent to which flow may take place between them. Other problems which have scarcely been touched on are the role of olfactory and auditory stimuli and the function of the complex lateral line in the life of clupeids, and the advantage of the soft body and deciduous scales to their survival.
One aspect of specialization iu thc apparent d o m i n ~ n m ~ of v i k i among the other senses. It seems to be the sense uaed beforo all othcrs in feeding, shoaling, spawning and escape.
It is interesting to speculate upon the evolutionary position of the clupeids. Even the accepted marine species, Ruch as the herring, have some associations with a freshwater way of life. This is shown in serological, biochemical and anatomical correspondences with fresh- water fishes, as well as in certain aspects of behaviour, such as demersal spawning. It seems most likely that the teleosts originated in fresh water and that the clupeids have retained stronger links with their origins than have other teleosts ; perhaps this is due to a much more recent colonization of the sea,
It has become clearer in the past few years tha t fisheries problem8 cannot be solved without a better knowledge of behaviour and phywi- Ology. This has been emphasized by Hardy (1949) and Bull (1061).
25

372 J. H. 9. BLAXTER AND F. Q. T. HOLLIDAY
and that this .is now generally appreciated is shown by the increase in such work in fisheries laboratories. Much of the work described in this review has been done with equipment only available a t such lab- oratories, is. research boats and marine aquaria, and it remains a cause for regret that marine fish of commercial importance are still to a large extent neglected by many universities due to lack of facilities.
VIII. REPERENCES
Amen, 0. (1952). The Lusterfjord herring and its environment. Fiskeridir. Skr. Havunderaek. 10 (2). 63 pp.
Ahlstrom, E. H. (1943). Studies on the Pacific pilchard or sardine (Sardinqps caeruh) . 4. Influence of temperature on the rate of development of pilchard eggs in Nature. Spec. aci, Rep. U.S. Fish Wildl. Seru. no. 23, 26 pp.
Ahlstrom, E. H. (1954). Distribution and abundance of eggs and larval popula- tions of the Pacific sardine. Fiah. Bull., U.S. Fish Wildl. Sew. no. 93,
,&lstrom, E. H. (1967). A review of recent studies of subpopulations of Pacific
Allen, W. E. (1920). Behaviour of loon and sardines. Ecology, 1, 309-10. Almy, L. H. (1926). The role of the proteolytic enzymes in the decomposition of
the herring. J. Amer. chem. SOC. 48, 2138-46. Ancellin, J. (1953). Hareng du sud de la Mer du Nord et de la Manche orientale,
Campagne de 1952-53. Ann. Biol. Copenhague, 9, 202-5. Andemson. K. A. (1954). Studies on the fluctuations in the rate of growth of the
herring in the Kattegat and the Skagerak. Rep. Inst. mar. Rea. Lyaekil, Ser. Biol. no. 3, 13 pp.
Andreev, N. N. (1966). Some problems in the theory of the capture of fieh by gill nets. (In Russian.) Trans. Inat. Mar. Fiah. U.S.S.R. 30, 109-27.
Andreu, B., and Pinto, J. do8 Ranto8 (1957). CaracteriRticas histologicas y biornotricrw tlol ovario (lo sarclinu (Surdirur pikhurdur Walt).) MI In mimliire- cion, pirosta y rocuporacion. Origon tlo 10s ovocitos. (Eng. summary.) N o h Eat&. Inat. Biol. marit., Liaboa, 6 , 3-38.
Andrews, C. B. (1931). The development of the ova of the California sardine (Sardina caerutea). Unpublished thesis, Stanford Medical School, U.S.A.
Anokhina, L. E. (1959). On the relationship between fecundity and the fat content of Clupea haren.gw membraa L. (In Russian, Marine Laboratory, Aberdeen trans. no. 735.) C.R. Acad. Sci., U.R.S.S. 129 (6), 1417-20.
Anokhina, L. E. (1960). On the relationship between fecundity, difference6 in the size of ova and fat content of Chpea harengue p a l h i i maris-albi Berg. (In Russian, Marine Laboratory, Abordeen trans. no. 734.) C.R. Acad. Sci., U.R.S.S. 133 (4), 960-3.
Anokhina, L. E. (1961). Relationship between the fertility, fat content and the variations in the size of ova in the clupeid fishes. (In Russian, Marine Labora- tory, Aberdeen trans. no. 733.) Trud. Soveahch. ikhtiot. Kom. Akad. Sci. S.S.S.R. 13, 290-5.
Anon. (1952). Californian Cooperative Oceanic Fisheries Investigation. Prog. Rep., 1st Jan. 1951-30th June 1952. 51 pp.
83-140.
fishes. Spec. aci. Rep. U.S. Fish Wildl. Sew. no. 208,44-73.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 373
Anon. (1966). Behaviour studies. Californian Cooperative Oceanic Fisheries Investigations. Prog. Rep., let April 196630th June 1966. 44 pp.
Arthur, D. K. (1966). "he particulate food and food resources of the larvae of three pelagic Bshea, especially the Pacific sardine, Sardimp8 cueruka (Girard). Scrippe. Inet. Oceanogr. Ph.D. thesis.
Awerinzew, S. (1936). Einige Ergebniase von Reobaohtungen an C l u p b r e w des Earentsmeoree. J . Cons. int. E z p h . Met, 10, 68-74.
Baburina, E. A. (1966). Structure of the eye and its retina in the common nor- thern pilchard. (In Ruseian. Biol. Abetr. no. 23677, 1968.) C.R. A d . S& U.R.S.S. 102, 626-8.
Bdowitz, E. (1916). Zur Kenntnia der Spermien des Heringa. Arch. Zeusford .
Balls. R. (1961). Environmental qhanges in herring behaviour. A theory of light avoidance aa suggested by echosounding observations in the North Sea. J . Cons. int. E x p h . Mer, 17, 274-94.
Bamford, T. W. (1941). The lateral line and related bones of the herring (CZUZW, harmpa L.). Ann. Mag. nat. HiBt., Ser. I I , 8, 414-38.
Barlow, G. W. (1961). C a w s and signi5cance of morphological variation in fishes. S p t . Zool. 10, 106-17.
Battle, H. I. (1936). Digestion and digestive enzymes in the herring ( C h p harmgu.9 L.). J . biol. Bd Can. 1, 146-67.
Bettle, H. I., Huntsman, A. G., Jeffere, Anne M., Jeffers, G. W., Johneon, W. H., and MacNairn, N. A. (1936). Fatnew, digestion and food of Psesamequoddy young herring. J. biol. Bd Can. 2, 401-29.
Batter, I. G. (1969). Fecunditim of winter-spring and summer-autumn herring spawners. J. Cons. int. Bxph. Mer, 25, 73-80.
de Beaufort, L. F. (1909). Die Schwimmbleee der Malacopterygii. Morph. Jb. 39, 626-644.
Belyaeva, V. N., and Nikonorov, I. V. (1980). Reeeons of attraction of 5sh by underwater light. I.C.E.S. Comparative Fishing Committee, Paper no. 160
BertalanfFy, L. von (1938). A quantitative theory of organic growth. (Inquiries
Ber t a ldy , L. von (1949). Problems of organic growth. Nafurc, Lmui. 163, 160. Beverton, R. J. H.. and Holt, 9. J. (1969). A review of the lifeupens and mortality
ratee of fish in Nature, and their relation to growth and other physiologicel characteristics. Ciba Symp. on The Lifeapan of Animals, 142-77.
Bhattacharyya. R. N. (1967). The food and feeding habits of larval arid post- larval herring in the northern North Sea. Mar. Rw. Scot. no. 3, 14 pp.
Bibov, N. E. (1960). Acclimatization of aalaka (Baltic herring) Clupea hasenpa mmbrau L. in the h a l Sea. I.C.E.S. Herring Committee, Paper no. 136 (mimeo . ) .
B*kov, N. P. (1968). On the distribution of herring in the North Sea in 1967. I.C.E.S. Herring Committee, Paper no. 77 (mimeo.).
Biehai. H. M. (1960a). The effect of gee content of water on larvas arid young fish. 2. Wb8. Zool. 163, 37-64.
Biehai. H. M. (1960b). The effect of hydrogen ion concentration on the survival and distribution of larvae and young fbh. 2. un&. 2001. 164, 107-18,
Bishai. H. M. (1960~) . The effect of water currenb on tho nurvival and din- tribution of fish larvas. J. Cons. int, Ezpkr. Met. 25, 134-46.
14, 177-84.
(mimeo.).
on growth laws 11.) Hum. Biol. 10, 181-213.

374 J . H. 9. BLAXTER AND F. 0. T. IIOLLIDAY
Bishai, H. M. (196la). The effect of salinity on the survival and distribution of larvae and young fish. J. Cons. int. Explor. Mer, 26, 16679.
Bishai. H. M. (1961b). The effect of pressure on the survival and distribution of larvaa and young fish. J. Cons. int. Explor. Mer, 26, 292-311.
Blaxter, J. H. S. (1955). Herring Rearing-I. The storage of herring gametes. Mar. Re.*, Scot. no. 3, 12 pp.
Blaxter, J. H. S. (1956). Herring Rearing--11. The effect of temperature and other factors on development. Mar. Res. Scot. no. 5, 19 pp.
Blaxter, J. H. S. (1957). Herring Rearing-111. The effect of temperature and other factors on myotome counts. Mar. Res. Scot. no. 1, 16 pp.
Blaxter, J. H. S. (1958). The racial problem in herring from the viewpoint of recent physiological, evolutionary and genetical theory. Rapp. C m . Explor. Mer, 143, 10-19.
Blaxter, J. H. 8. (1960). Tho uffoct of extromcs of temperature on herring Iarvac.
Blaxter, J. H. S. (1962). Herring Rearing-IV. Rearing beyond the yolk sac stage. Mar. Res. Scot. no. 1, 18 pp.
Blaxter, J. H. S., and Dickson. W. (1959). Observations on the swimming speeds of fish. J. Corn. int. Explor. Mer, 24, 472-9.
Blaxter, J. H. S., and Hempel, G. (1961). Biologische Beobachtungen bei der Aufzucht von Heringsbrut. (Eng. summary.) Helgoliind Wise. Meereeunter8. 7. 26&83.
Blaxter, J. H. S., and Hempel, G. (1963). The influence of egg size on herring larvae. Submitted for publication to J. Cons. int. Exp:plor. Mer.
Blaxter, J. H. S., and Holliday, F. G. T. (1958). Herring (Clupea harengw, L.) in aquaria.-11. Feeding. Mar. Res. Scot. no. 6, 22 pp.
Blaxter, J. H. S., and Parrish, B. B. (1968). The effect of artificial lights on fish and other marine organisms a t sea. Mar. Res. Scot. no. 2, 24 pp.
Blsxter, J. H. S., and Parrish, B. B. (1969). Observations on the avoidance of netting by herring in aquaria, I.C.E.S. Comparative Fbhinp; Committee, Paper no. 18 (mimoo.).
Blaxter, J. H. S., Parrish, B. B., and MO~U~JWH, 1’. H. (1060). ‘ J h r4wthm of herring to moving obstactclos. i .C.JS.H. Cc,mpwstlvu IEivhiriK ~ h r r i r t i i t f m ,
Paper no. I6 (mimeo.). Bolster, G. C. (1958). On the shapo of herring f i c h f J f > k . J. f h i n . z n t . K x p h r . Mcr.
Bolster, G. C., and Bridger, J. P. (1967). Nature of tho spawning art:% of hnrringn. Nature, Lond. 179, 638.
Borisov, P. G. (1955). Tho behaviour of futhcn iirldor tho i r l f t i m n w (Jf nrt,iflciaI light. (In Rumian, Marine hhOrtLtQry, AtJc:rdet?n trctriw. no. 41 1 .) Ymo. Conf. on the behaviour of fish and on the locating of its commercial con- centrations. Ed. by E. N. Pavlovskii (MOSCOW 1965).
Borisov, P. G., and Protaeov, V. (1960). Some aspects of light perception in fish and selective light sources. I.C.E.S. Comparative Fishing Committee, Papr no. 139 (mimeo.).
Bowers, A. B., and Holliday, F. G. T. (1961). Histological changes in the gonad associated with the reproductive cycle of the herring (Clupea harengua L.). Mar. Res. Scot. no. 5, 16 pp.
J . mar. biOl. A88. U.K. 39, 605-8.
23, 228-34.

THE BEHAVIOUR AND PHYSIOLOQY OF EIERRINQ AND OTHER CLUPEIDS 375
Bowera, A. B., and Williamson, D. I. (1961). Food of lervd and early post- larval stagea of autumn spawned herring in Manx waters. Rep. mar. bid. Stat. Pt Erin, 63, 17-26.
Bowman, A. (1923). Spawny haddocks; the occurrence of spawny haddocks and the locus and extent of herring spawning grounds. Fi8h, Inueat. Scot. 1922, no. 4, 16 ?p.
Boyar, H. C. (1961). Swimming speed of immature Atlantic herring with reference to the Pamxnaquoddy Tidal Project. Trans. A m r . Fish. SOC. 90,21-6.
Boyar, H. C., and Sindermann, C. J. (1969). Additional notes on the maintenance of immature sea herring in captivity. Prog. Fiah. Cult. 21, 186-7.
Brendes, C. H., and Dietrich, R. (1963a). nber die Fettverteilung kn K6rper des Herings. Ver6ff. Imt. Heeresf. Bremrhven, 2, 109-21.
Brandes, C. H., and Diotrich, R. (1963b). A review of the problem of fat and water content in the edible part of the herring. Fettc u. Beif. 55, 633-41.
Brandhorst, W. (1969). Spawning activity of the herring8 and growth of their larvae. Int. oceanogr. Congreee, New York, Preprints, 218-21.
v. Brandt, A. (1964). Perlonnetze in der Ostseetreibnetzfiaoherei fiir Heringe. Bet Fischwirt, 4, 296-300.
Brawn, V. M. (1960a). Underwater televieion observation8 of the sWimming speed and behaviour of captive herring. J. Fhh. Res. Bd Can. 17, 689-98.
Brawn, V. M. (1960b). Seasonal and d i d vertical distribution of herring (Clupea hrengus L.) in Peeeamequoddy Bay, N.B. J. Fiuh. Res. Bd Can.
Brawn, V. M. (1960~). Temperature tolerance of unacclimatized herring (CZupeo krengm L.). J . Fish. Rea. Bd Can. 17,721-3.
Brawn, V. M. (198Od). Swive l of herring (CZum krengus L.) in water of low salinity. J. Fish. Rea. Bd Can. 17, 726-6.
Brawn, V. M. (1962). Physical properties and hydrostatic function of the swim bladder of herring (CZupea hrengus L.). J . Fish. Res. Bd Can. 19, 636-68.
Breder, C. M. Jr. (1969). Studies on social groupinp in hhea. BuU. Amer. Mus.
Breder, C. M. Jr., and Krumholz, L. A. (1943). On the locomohr and f ~ r l i n g behaviour of certain postlarval Clupeoidea. ZoologicCr, N. Y. 28, 61 - 7.
Bretschneider, L. H., and Duyven6 de Wit, J. J. (1!147). Sexual endocrindoKy of non-mammalian vertebrates. M m g r . Prog. Rea. HoUand during Wur (2). 146 pp. Elsevier Pub. Co. Inc., Amsterdam.
Bridger, J. P. (1958). On efficiency tests made with a modified Gulf 111 high speed tow-net. J. Cone. int. h'xpbr. Mer, 23, 357-66.
Bridger, J. 1'. (1960). On the rwltrtionshiy betwoon Htock, larvae arid recruih in the " D o m " herring. I.C.E.S. Herring Committee, Yapor no. 169 (mimoo.).
Bridger, J. P. (1961). On fecundity and larval ahundanco of Dowm herring. Fish. Invest. Lond., Ser. I I , 23 (3). 30 pp.
Brook, G. (1885). On the development of tho herring. Part I. Rep. Piah. Bd Scot. no. 3, pp. 32-61.
Brook, G. (1886). On the development of the herring. Part 11. Rep. Fbh. Bd Scot. no. 4, pp. 31-43.
Brown, M. E. (Ed.) (1967). " The Physiology of Fishes " 2 Volumes, 447 and 526 pp. Acadomic Preess, New York.
17, 699-711.
nd. Hiat. 117, 397-481.

376 J. €I. S. BLAXTER AND F. Q. T. HOLLIDAY
Buchmann, H. (1940). Hypophyse und Thyreoidea im Individualzyklus des Herings. (Marine Laboratory, Aberdeen trane. no. 367.) Zool. Jb. (Anat.)
Biickmann, A. (1960). Die Untersuchungen der Biologischen Anstalt uber die okologie der Heringsbrut in der siidlichen Nordsee. 11. Hekokind. Wise. Meereauntera. 3, 171-206.
Biickmann, A., and Nempel, G. (1957). Untersuchungen an der Heringslarven- bevolkerung der Innenjade. (Eng. summary.) Helgoldnd. Wiss. Meermn- ters. 6 (l), 62-70.
Biickmann, A., Harder, W., and Hempel, G. (1963). Unsere Beobachtungen am Hering, Clupea harengua L. (Eng. summary.) Kurz. Mitt. In&. Fiachereibiol. Hamburg, 3, 22-42.
Bull, H. 0. (1962). An evaluation of our knowledge of fish behaviour in relation to hydrography. Rapp. Cons. Explor. Aler, 131, 8-23.
Bull, H. 0. (1961). Oceanography, American Association for the Advancement of Science. pp. 239-66..
Bullough, W. S. (1940). A caae of hermaphroditism in the herring (Clupeu hatenqua. Linn.) Proc. Leeds Phil. Zit. SOC. 3, 638-41.
Burd, A. C. (1966). The North Shields fishery 1949-66. I.C.E.S. Herring Com- mittee, Paper no. 66 (mheo.).
Burd, A. C. (1968). Recruitment and the North Shields fhhery. I.C.E.S. Herring Committee, Paper no. 67 (mheo.).
Burd, A. C. (1962). Growth and recruitment in the herring of the southern North Sea. Fish. Invest., Lo&. Ser. I I , 23 (6), 42 pp.
Burne, R. H. (1909). The anatomy of the olfactory organ of Teleostean fkhes. Proc. zool. Soe. Lo&. 2, 610-62.
Canagaratnam, P. (1959). Growth of fishes in different salinities. J. Fiah. Res. Bd Can. 16,121-30.
Chamon, H . J., and El Saby, M. K. (1932). Fat metabolism of the herring. 1. A preliminary survey. Biochem. J. 26, 2021-34.
Chestnoy, V. N. (1961). Vertical migrations of herring and the methods of their utilization in the fishing practice. (In Ruwian.) Ryb. Khoz. 37 (6). 63-7,
Clark, F. N. (1966). Average lunar month catch of sardine fwhermen in nouthcrri California 1932-33 through 1964-65. CaZ$ Fish &me, 42, 309.23.
Cleaver, F. C., and Franett, D. M. (no date). Tho predation by wa birds upon tho eggs of the Pacific herring. Clupea t ~ a l b i i at Holmos Hnrhor during 1946. Wash. Slate Dept. Fish. Div. Sci. Rea. Biol. Rep. no. 46 B, 18 pp.
Colton, J. B., (Jr.), Honey, K. A., and Temple, R. F. (1961). The effectivencm of sampling methods used to study the distribution of larval herring in tho Gulf of Maine. J . Cons. int. Ezplor. Mer, 26, 180 90.
Cowan, D. (1938). Tank life of fish in the aqiiariiim of tho 1)ove Miirino I,&m. tory, Cullercoats. Rep. Dove Mar. Lab. 3rd Aeries ( b ) , 38-9.
Craig, R. E, (1960). A note on the dependence of cetchw (Jn thrnparrrturo and wind in the Buchan prespawning fkhery. J. Cone. int. Ezplor. Mer, 25,
Craig, R. E., and Priesthy, El. (1961). I'trotographic studion of finh ptJphtifJriN. Nature, Lvnd. 188,333-4.
Cunningham, J. T. (1889). Studies of tho reproduction and development of tebostean hh0S occurring in the neighbourhood of Plymouth, J. mar.
66, 191-262.
The role of ethology in oceanography.
188-90.
b i d . ABS. l7.h'. 1, 10-64.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 377
Cunningham, J. T. (1894). The lifs history of the pilchard. J . mar. 6iot. Ass. U.K. 3, 148-63.
Cunningham, J. T. (1896). “ The Natural History of the Marketable Marine Fishes of the British Islands.” 376 pp. Macmillan, London.
Cushing, D. H. (1966). Production and a pelagic fishery. Fwh. Invest. Lond., Ser. I I , 18 (7). 104 pp.
Cushing, D. H. (1960). Fishing gear and fieh behaviour. Proc. world sci. meeting on the biology of sardines and related species. Rome 1969. (Methodologicel
Cushing, D. H. (1961). On the failure of the Plymouth herring fiehery. J. mar.
Cushing, D. H. (1962). Recruitment to the North See herring stocka. F a . Invest. Lond., Ser. 11, 23 (6), Part 2, pp. 43-71.
Cushing, D. H., and Burd, A. C. (1966). Recruitment and maturity of the East Anglian herring. Nature, Lond. 178, 80-7.
Cushing, D. H., and Burd, A. C. (1967). On the herring of the southern North See. Fish. Invest. Lond., Ser. I I , 20, (11). 31 pp.
Dannevig, A. (1948). Rearing experiments at the Flradevigen sea fish hatchery 1943-1946. J. Cons. int. Explor. Mer, 15, 277-83.
Dannevig, A., and Dannevig, 0. (1960). Factors affecting the sunrival of fiah larvae. J. Cone. int. E x p h . Mer, 16, 211-16.
Dannevig, A., and Hansen, S. (1952). Factors involved in hatching and rearing fiah eggs and larvae. (In Norwegian, Marine Laboratory, Aberdeen trans. no. 416.) Fiskeridir. Skr. Huvunderaek. 10 (l), 8-16.
Dee, N., and Tibbo, S. N. (1962). On the feeding and growth of young herring (Clupeu hurengw, L.) in captivity. J. Fish. Res. Bd Can. 19, 981-3.
Dawson, A. B. (1933). The relative numbers of immature erythrocytee in the circulating blood of several species of marine teleosts. Bwl. Bull., Woods Hole, 64, 36.
Day, L. (1967). Vertebral numbera and 5rst year growth of Canadian Atlantic herring (Clupea hurengus L.) in relation to water temperature. BuU. Fiah. Res. Bd Can. 3, 166-76.
Delsman, H. C. (1930). The study of pelagic f%h eggs. Proc. 4th Puc;f. eci. Congr. 3 (Biol.), 61-7.
Dementeva, T. F. (1968). Methods of studying the effect of environmental factors on the fluctuations in abundance of the Azov anchovy. (In Russian. Fish. Res. Bd Can. trans. no. 239.) Trudy V.N.I.R.O., 34, 30-52.
Devold, F. (1963). Tokter med “ G.O. Sam ” i Norskehavet vinteren 1962163. Fiskele Gang, 39, 236-9.
*Devold, F. (1961). The life history of the Atlanto-Scandian herring. I.C.E.S. Herring Symposium, Paper no. 40 (mimeo.).
Dietrich, a,, Sahrhage, D., and Schubert, K. (1969). Locating fbh by thermo- metric methods. I n ‘‘ Fishing Gear of the World ”. 607 pp. Fishing News Books Ltd., London.
DOW, R. L. (1962). Swimming speed of river herring Pomoloinu paeudoharengw (Wilson). J . Cons. int. @xplar. Mer, 27, 77-80.
Dragwund, 0. (1958). Reactions of fish to artificial light with s p b l reforonce tr, Lerge Herring and Spring Herring in Norway. J. Cone. int. Ezplor. Mer, 23, 213-27.
Paper, 3.) 3, 1307-26.
b&d. A S S . U.K. 41,799-816.

378 J. I€. S. BLAXTER AND F, 0. T. HOLLIDAY
Dragesund, 0. (1960). Observations on the spawning of herring in captivity. Fauna, 4, 137-44.
Ehrenbaum, E. (1930). (Chairman). Meeting of Herring Experts, held at Fisheries Laboratory, Lowestoft, England, Nov. 4th-7th 1930. Rapp. Cons. Explor. Mer, 74, 116-20.
Elmhirst, R. (1908). The keeping of young herring alive in captivity. Nature,
Evans, H. E. (1962). The correlation of brain pattern and fooding habits in four
Ewart, J. C. (1884). Natural history of the herring. Rep. Fiah. Bd Scot. no. 2,
Ewart, J. C. (1886). Are herring ova likely to develop normally on the deep fishing banks? Rep. Fish. Bd Scot. no. 4, 43-6.
Farrin, A. E., Scattergood, L. W., and Sindermann, C. J. (1967). Maintenance of immature sea herring in captivity. Prog. Fish. Cult. 19, 188-9.
Farris, D. A. (1966). Diet induoed differences in the weight-length relatiomhip of aquarium fod sardines (Sardinopa caerulea). J. Fish. Res. Bd Can. 13,507-13.
Farris, D. A. (1968). Diet induced variation in the free amino acid complex of Sardinops caerulea. J . Cona. int. Explor. Mer, 23, 236-44.
Ferris, D. A. (1960). The effect of three different types of growth curves on estimates of larval fish survival. J . Cons. int. Explor. Mer, 25, 294-306.
Fliichter, J. (1 963). Funktioneanatomkche Untersuchungen am Kieferapparat der Heringslarven. (Eng. summary.) Kun. Mitt. Inst. Rischereibiol. Hamburg, 12 (in press).
Ford, E. (1929). Herring Investigations a t Plymouth. VII. On the artificial fertilization and hatching of herring eggs under known conditions of salinity with some observations on the specific gravity of the larvae. J . mar. biol. Ass. U.K. 39, 591-603.
Ford, E. (1930). Herring investigations at Plymouth. VIII. The transition from larva to adolescent. J. mar. b i d . ,488. 1J.K. 16, 723-62.
*Fraser, J. H. (1961). Plankton arid th: Shc.lInricl Herririg F’dfiry. I,C.J?.ij. Herring Rympobiium, J’qm no. 14 ftnirrw,.) .
Freytag. G . (1961). fIerirrK~H~:hw~t~r~lK~:ri ioHC~i~: . Tnfi,rrni~ti~mwl filr f l i r t
Fuchwirtschaft (from tho U u r i d ~ : ~ f ~ ~ r ~ c t i i ~ r ~ ~ ~ ~ ~ r ~ ~ t s l t f Iir F~iwhww, I l i u r i . burg), nr. 616, 139-40.
Fulton, T. W. (1906). Report on an onqliiry into tho horring fisheries of the Firth of Clyde. Rep. E’iah. Bd Scot. no. 26, t)8-9.
Gabriel, M. L. (1944). Factors affecting the number and form of vertebrae in Fundulua heteroclitus. J. exp. Zool. 95, 106-47.
Galkina, L, A. (1967). Effect of salinity on the sperm, eggs and larvae of the Okhotsk herring. (In Russian, trtms. available from Office of Technical Services, U.S. Dept. of Commerce, Washington 26. D.C.). Izveatiya T.N.I.R.O. 45, 37-50.
Galkov, A. A. (1968). Some data on the influence of the behaviour of the Atlanto- Scandian herring on its catch by the pelagic trawl. (In Ruwian.) Ryb. Khot.
Gilis, C . (1954). Spent herring concentrations exploited by the Belgian herring
Lond. 79, 38.
species of cyprinid fishes. J. comp. Neurol. 97, 133-42.
pp. 61-73.
34 ( l l) , 8-12.
trawlers in 1953-64. Ann. Biol. Copenhugue, 10, 180-3.

TEE B3XTAVXOUR AND PEYSXOLOOY OF HERRING AND OTHER OLUF'EXDS 379
Girse, I. I. (1961). Availability of food animals to some fish- at different levels of illumination. (In Ruseian, Marine Laboratory, Aberdeen trans. no. 717.) Trudy sovesch. Ikhtwl. Kom. Akad. Nuuk. S.S.S.R. 13, 365-9.
Glover, R. S. (1967). An ecological survey of the drift-net herring fishery off the north-east coast of Scotland. Part 11. Tho planktonic environment of the herring. Bull. mar. Ecol. 5 (no. 39), 43 pp.
Glover, R. 8. (1968). Fish hohaviour and tho phktoriic environment, : t h c t rob of field observations. Proc. Indo-t'ucijc E'ish. Coun. 3, 72-8.
Graham, M. (1931). Some problems in herring behaviour. J. C m . int. Ezplor. Me*, 6, 252-66.
Graham, J. J. (1966). Observations on the alewife Pornolobur, psesezcdoharengur, (Wilson) in fresh water. Publ. Ontario Fish. Res. Lab. no. 74. Biol. Ser. no. 62, 43 pp.
Graham, J. J. (1967). Some observations on the schooling movements of the aledfe in Lake Ontario. C u d . Fieid. Nut. 71, 116-6.
Graham, T. (1962). A relationship between growth, hatching, and spawning seaaon in Canadian Atlantic herring (Clupeu harengua L.). J. Fish Rea. Bd Can. 19, 986-7.
Gray, I. E. (1964). Comparative study of the gill area of marine fish. Biol. BuU., Woods Hole. 107, 219-26.
Gray, I. E., and Hall, F. G. (1930). Blood sugar and activity in fishes with notes on the action of insulin. Biol. Bull., Woods Hole, 58, 217-23.
Groody, T., Loukashkin, A., and Grant, N. (1962). A preliminary report on the behaviour of the Pacific sardine (Surdinopa caerdeu) in an electrical field. Proc. Calif. A d . Sci. 27, 311-23.
Gunter, G., and Christmas, J. Y. (1960). A review of literature on menhaden with special roference to the Gulf of Mexico menhaden Brevoortia putronur, Goode. spec. mi. Rep. U.8 . Fish. Wildl. serv. no. 363, 31 pp.
Gwyn, A. M. (1940). The developmont of the vertebral column of the Pacific herring (Clupeu pu lh i i ) . J. Fish. Rea. Bd Can. 5,ll-22.
Hamana, H. (1936). On the h t stages of devolopment of the herring. (In Japaneso.) Suisankai, no. 628, 13-14.
Harder, W. (1962). Ziir piiRt,om~iryorieli~ri Ent,wii:khmg den Durrnon beirn HiwinK. (:liqwu luirisnyun 1,. %. AM^. h'nttidhrh. 116, 370 U8.
Iitrrtlor, W. ( IM3). Zu1r1 Rwmwuoluhurn t l w IIoringR (Clupea harengm L.). %. Morph. i)kol. l'iere, 42, 200-24.
Hardor, W. (1964). Die Entwieklung der Rospirationsorgane beim Hering (Clupea harengus L.), 2. Anut. Entwacach. 118, 102-23.
Harder, W. (1060). Vergleichende Unterauchungen zur Morphologie des Dames h i Clupooidoa. 2. wias. 2001. 163, 66-167.
Hurdy, A. C. (1924). The herring in relation to its animate environment. Part I. The food and feeding habits of the herring with special reference to the east coast of England. Fiah. Invest. Lond., Ser. 11, 7 (3), 63 pp.
Hardy, A. C. (1949). Towards a programme of herring research. Rupp. C m . Explor. Mer, 128, 9-18.
Hasler, A. D., Horrall, R. M., Wisby, W. J., and Breemer, W. (1968). Bun- orientation and homing in fishes. Limml. Oceanogr. 3, 363-61.
Heincke. F. (1898). Naturgeschichte des Heringq. Teil I. Die Lokalforrnen und die Wanderungon des Herings in den europlimhen Meeren. Abh. dkch. sqfiach Ver. 2, 128-223.

380 J. H . S. BLAXTER AND F. 0. T. HOLLIDAY
Hela, I., and Laevastu, T. (1962). " Fisheries Hydrography " 137 pp. Fishing News Books Ltd., London.
Hempel, G. (1953). Die Temperaturabhiingigkeit der Myomerenzahl beim Hering (Clupea hrengus L.). (Marine Laboratory, Aberdeen trans. no. 326.) Naturwiasenschaften, 17, 466-8.
Hempel, G., and Blaxter, J. H. S. (1961a). The experinlentel modification of meristic characters in herring (Clupea hnrengua L.). J . Cone. int. E x p h . Met, 26, 336-46.
*Hempel, G., and Blaxter, J. H. S. (1961b). On the condition of herring larvae. I.C.E.S. Herring Symposium, Paper no. 34 (mimeo.).
Hentschel, E. (1960). Die Nahrung der Heringslarven. Helgoliind. WisS. Meeresunters. 3, 59-81.
Hickling, C. F. (1940). The fecundity of the herring of the southern North Sea. J. mar. biol. Ass. U.K. 24, 019-32.
Hickling, C. F. (1945). The seasonal cycle in the Cornish pilchard. J. mar. biol.
Hickling, C. F. (1947). The herring fisheries a t Milford Haven. J. m r . biol. AM.
Hide, T. S., and Thomson, D. A. (1962). Introduction of the threadfin shad to Hawaii. Prog. Fish. Cult. 24, 159-63.
Hjort, J. (1910). Report on herring invnstigations until January 1910. Publ. Circ. Cons. Explor. Mer, no. 53, 175 pp.
Hoar, W. S. (1962). Thyroid function in some anadromous and landlocked teleosts. Trans. roy. SOC. Can. 46, 39-53.
Hoar, W. S. (1955). Mem. SOC. Endocrinol. 4, 45-22.
Hodgson, W. C. (1934). " The Natural History of the Herring of the Southern North Sea. The Buckland Lectures for 1933." 120 pp. Arnold & Co., London.
Hodgson, W. C. (1967). " The Herring and Its Fishery ". 197 pp. Routledge and Kegan Paul, London.
Holliday, P. 0. T. (IihS8). The npuwnirig of tho tiorring. Scot. h' i~h . / h l l . no. 10.
Holliday, F. G. T. (1960a). The control of maturation in the herring. I.C.E.S. Herring Committee, Paper no. 38 (mimeo.).
Holliday, F. G. T. (1960b). The control of spawning in the herring. I.C.E.S. Herring Committee, Paper no. 39 (mimeo.).
Holliday, F. G. T. (1962). Hermaphroditism in the herring (Clupeu harengus L.). Proc. r q . SOC. Edinb., Ser. B. 68, 208-14.
Holliday, F. G. T., and Blaxter, J. H. S. (1960). The effects of salinity on the developing eggs and larvae of the herring. J. mar. biol. Ass. U.K. 39, 691- 603.
Holliday, F. G. T.. and Blaxter, J. H. H. (l!Ml). The effects of Halinity on herring after metamorphosis. J. mar. biol. Aes. U.K. 41, 37-48.
Holliday, F. G. T., and Parry, Gwyneth (1962). Electron-microrrcoi,o ntudiee of the acidophil cells in the gills and pReurlobranchs of fbh. Nature, L i d . 193, 192.
Holstvoogd, C. (1957). The poatornbryonio devd~tlirner~t of the l~rcirifil~bro4 ir, Clupea h a r e n p 1,. Arch. nhr1. dad. 12, 4db 66.
ASS.. U.K. 26, 115-38.
U.K. 26, 408-20.
Reproduction in teleost fishes.
pp. 11-13.

THE BEHAVIOUR AND PKYSIOLOOY OF HERRING AND OTHER CLUPEIDS 381
Hourston, A. S. (1963). The relationship of herring in inshore waters to the temperature and salinity of their environment. Progr. Rep. bid. Stas Nanaimo and Prince Rupert, no. 96, p. 16.
Hourston, A. S. (1968). Population studies on juvenile herring in Barkley Sound, British Columbia. J . Fish. Ree. 3 d Can. 15, 909-60.
Hubbs, C. L. (1926). Racial and seaeonal variation in the Pacific herring, Cali- fornia sardine and California anchovy. Cali,f. Fiah Q a m , 8.23 pp.
Hughea, R. B. (1969e). Chemical studies on the herring (Clupea h r w ) . 2. The free amino acids of herring flesh and their behaviour during post- mortem spoilage. J . Sci. Fd Agric. no. 10, 558-64.
Chemical studies on the herring (Clupea hut--). 1. Trimethylamine oxide and volatile amines in fresh, spoiling and cooked herring flesh. J . Sci. Fd Agric. no. 8, 431-6.
Hunter, A. and Dauphin&, J. A. (1926). Quantitative studies concernhg the distribution of arginase in fishes and other animals. Proc. roy. Soc.,Ser. B . 97,
Hughes, R. B. (1969b).
227-42. Iles, T. D. (1962). I.C.E.S. Herring Committee, Paper no. 86 (mimeo). Ito, S. (1968). On the relationship between water temperature and the incubtion
time of sardino (Sardinops melanosticta) Ann. Rep. Jap. Sea Reg. Fiah. Rw. Lab. 1 (4), 26-31.
Ivlev, V. S. (1966). “ Experimental Ecology of the Feeding of Fiehee.” Tram- lmted by D. Scott, 302 pp. Yale University Fress, New Haven.
Ivlev, V. S. (1960). On the utilization of food byplanktophagefishes. Bull. math.
Jeoobshagen, E . (1913). Untersuchungen iiber daa Darmsystem der Fieche und Dipnoer. J e m . 2. Naturw. 49, 373-810.
+Jakobeson, J. (1961). Some remarks on the distribution and availability of the North Coaet herring of Iceland. I.C.E.S. Herring Symposium, Paper no. 44 (mimeo.).
Jean, Y. (1966). A study of spring and fall spawned herring (CZupea harengus L.) at Grand RiviBre, Bay of Chaleur, Quebec. Crmtr. Dep. Fbh. Quebec, no. 49,
Jens. G. (1954). Untersuchungen iiber rhythmische Eracheinungen hirn Herinp- fang in der Nordsee. Arch. Fiach Wias. 5 , 11 .%9.
Jensen, A. G. C. (1950). Changes in the quality of the herring in the course of the year and from year to year. Rep. Danieh bid. Sta. 51, 3-18.
Johansen, A. C. (1919). On the large spring spawnlng sea herring (Clupea haretagua L.) in the North West European Water. Medd. Komm. Havunderseg. Kbh., Ser. Fiak. 5 (8), 66 pp.
Johaneen, A. C. (1926). On the diurnal vertical movements of young of aome fishes in Danish waters. Medd. Komm. Havundereog. Kbh., Ser. F&3k. 8 (2).
John, C. C. (1932). The origin of erythrocytes in herring (Clupea harengw). Proc.
Johneon, W. H. (193911). Feeding of the herring. . I . Fieh. Rea. Bd Cun. 4, W 2 . 6. Johnson, W. H. (1939b). Effect, of light on t h e rriovnrrronts of herring. , I . bud.
Bd Can. 4, 349-64. Jones, F- R. Harden (1962). Further ohsorvatiorui on the ??lovfmfJr:t## of‘ horrinK
(czum harengua L.) shoals in relation to tide1 currorit. , I . I:ms. int. kkphr . Mer, 27, 62-76.
Bwphya. 22, 371-89.
76 PP.
28 PP.
roy. SOC., Ser. 3. 110, 112-19.

382 J. H. 9. BLAXTEIt AND P. Q. T. IIOLLTDAY
Jones, F. R. Harden, and Marahall, N. B. (1963). The structure and functions of the teleostean swimbladder. Biol. Rev. 28, 16-83.
Kamshylov, M. M., and Gerasimov, V. V. (1960). An attempt at keeping young Murman herring in aquarium. (In Russian.) Proc. Symp. on the biological bases of the oceanic fisheries. pp. 84-7.
-oh, Y. (1953). tfber den japankchen Hering (Clupea p a l h i i Cuv. et Val.).
Kelley, A. M. (1946). Effect of abnormal carbon dioxide tension on development of herring eggs. J. FGh. Rea. Bd Can. 6, 435-40.
Keys, A. B., and Willmer, E. N. (1932). ‘‘ Chloride secreting cells ” in the gills of fishes, with special reference to the common eel. J. Phyeiol. 76.368-78.
Kirchhoff, H. (1968). Funktionell-anat,omkche Untersuchung des Visceral- apparates von Clupeu brengua L. 2001. Jb. 76, 461-640.
Kiumura, K. (1934). On the manntr of swimming of sardines in a confined space. Bull. Jap. Soc. aci. Fish, 3, 87-92.
Komarovsky, B. (1969). The feeding habits of sardines. New Scientist, 6, 1366-7.
Kotthaus, A. (1939). Zuchtversuche mit Heringslawen (Clupeu hurengua L.). (Marine Laboratory, ‘Aberdeen trans. no. 306.) Helgokind. Wiaa. Meeresun- tera. 1, 349-611.
Krefft, G., and Schubert, K. (1960). Beobachtungen uber den Einflum kiinst- licher Beleuchtung dor Meeresoberfliiche auf Fische. Fiachereiwelt, 2nd year, 86-8.
Krefft, G., and Schiiler, F. (1961). Beobachtungen uber die Tiefenverteilung von Heringsschwarmen in der nordlichen und mittleren Nordsee im August 1960. Fiachereiwelt, 3rd year, 93-6.
Kryzhanovsky, S. G. (1966). Development of Clupea hurengua nzenzbras in the water of high salinity. (In Russian, Marine Laboratory, Aberdeen trans. no. 792.) Voprosy. Ikhtiologii, no. 6, 100-4.
Kupffer, C. (1878). Die Entwicklung des Herings im Ei. Wiaa. Meereantera. Abt. Kiel, 3, 176-226.
Kurata, H. (1969). Preliminary report on the rearing of the herring larvae. Bull. Hokkuido Reg. Fish. Ree. Lab. 20, 117-38.
Lagler, K. E., Bardach, J. E., and Miller, It. 1%. (1962). “ Ichthyology.” 646 pp. John Wiley and Sons Inc., New York.
Lesker, R. (1962). Efficiency and rato of yolk utilization by dcvaloping omhryon and larvaa of the Pacific sardine, ,Yardinope caeruku (Cirtrrd). . I . Fi8h. Ilea. Bd Can. 19, 867-76.
Oxygon cormrirnl)ticm arul r~urrio- regulation by single Pmific mrdirio VKKH and larvae ( i%ird ir~~pa rM*5+&i Girsrd). J. &ms. int. Ezpbr. Mar, 27, 26-93.
Lasker, R., and “heilacker, 0. II. (IW&), ‘J’tio fatty r x i d corrtr,onikiori i d t . h l i p i r k of some Pacific sardine tissues in r&hn to ovarian nratiwcltiim arid t h t .
J. Lipid nee. 3. 60-4.
Cytologia, 18, 67-9.
Laaker, R., and Thcilacker, 0. H. (196214.
Lea, 1:. (1U30). Mortality in the tribo of”orwoKian herring. Rupp. C m . E x p h . Mer, 65, 100-23.
Lobour, M. V. (1921). Tho larval and post-liirval stage8 of tho pilchard. sprat and herring from the Plymouth district. J. mar. biol. Aaa. U.K. 12, 427-68.
Lebour, M. V. (1923). The food of plankton organisms 11. J. mar. biol. Aaa. U.K. 13, 70-92.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 383
bbour , M. V. (1924). The food of young herring. J. mar. biol. Ass. U.K. 13,325-30. Le Gall, J. (1935). 1. Les populations de
I‘Atlantique Nord-Eat. Ann. Imt. Ochnogr. 15 (I) , 216 pp. Lishev, M. N., Rannak, L. A., and Lisivnenko, L. N. (1961). Condition of the
Baltic herring stock in north-eastern Baltic and tho Gulf of Riga. I.C.E.S. Baltic-Belt Committee, Paper no. 124 (mimeo.).
Lissner, H. (1.925). Die Nahrungaaufnahmo boim Hering. Ber. dtech. Kmm. Meeresforach. 1, 199-208.
Ljubickaja, A. I. (1957). Beobachtungen imd Versuche iiber den Einfiuss dee Lichtes auf daa Verhalten der Fische. (In Ruseian, Rostock In& f* Hochueefischerei trans. no. 1436.) Izveatija. A M . pedugog. Nauk. R.S.F.S.R. no. 85.
Loukashkin, A., and Grant, N. (1964). Further studies of the behaviour of the Pacific sardine (Sardimps caerub) in an electrical field. Proc. CaZij. A d .
Loukaahkin, A. S., and Grant, N. (1969). Behaviour and reactions of the P8cifio eardine, Sardinops c a e d e u (Girard) under the influence of white and coloured lights and darkness. Proc. Calif. A d . Sci. 29, 509-48.
Loukeshkin, A. S., and Groody, T. C. (1955). On the Pacific sardine (Sardimp cuermlea Girard) in aquaria : transportation, handling, maintenance and survival. Proc. Calif. A d . Sci. 28, 339-53.
Love, R. M. (1957). The biochemical composition of fish. I n ‘ I The Physiology of Fishes.” Vol. 1, Ed. M. E. Brown. 447 pp. Academic Press, New York.
Lovern, J. A. (1935). Fat metabolism in fishes, VI. The fats of some plankton crustacea. Biochem. J. 29, 847-9.
Lovern, J. A. (1938). Fat metabolism in fishes, XII. Seasonal changes in the cornpiition of horriug fat. Riochent. J . 32, 678-80.
Lovern, J. A. (1951). Tho chemistry and Inutabolism of fate in fish. Biochcm. SOC. symposia Cumbridge Engl. no. 6, 49-62.
Lovern, J. A., and Wood, H. (1937). Variations in the chomical composition of the herring. J. mar. biol. A88. U .K . 22, 281-93.
Lovetskaya, A. A. (1956). On the reaction to light in the sprat. (In Russian, & h e Laboratory, Aberdeen tram. no. 637.) Trudy ,%veech. Ikhl id . Kom. A M . Nauk. S.S.S.R., 8, 121-3.
Lowenstein, 0. (1957). The acouatico.IatmaIis rywttm. In ‘ I ‘1710 I’hyHifJlqy (if Fishes.” Vol. 11, Ed. M. E. Brown. 526 pp. Academic Proa, Now York.
LUC~S, C. E. (1936). On the diurnal variation of size groups of trawl caught herring. J. Cons. int. E x p h . Mer, 2, 53-9.
Liihmann, M. (1952). Fettgehalt und Preisgestaltung bei Ostseesprotten. Pis- chereiwelt, 4th year, 81-3.
Liihmann, M. (1953). a e r die Fettspeicherung bei Ostseeheringen und ihre Beziehung zum Fortpflanzungszyklus. Kieler Meereeforcrch. 9, 213-27.
WGregor, J. S. (1959). Relation between fish condition and population size in the sardine (Sardimpa caerula). Fish. Bull. U.S. Fish Wildl. Serv. no. 166, 215-30.
Maier, H. N., and Scheuring, L. (1923). Entwickolung der Schwirrimbluwt urid Beziehungen zum s ta tkhen Organ und dor Khake boi ~ ~ I I J ~ H ~ ~ ~ J ~ ~ npez.
beim Hering. (Marine Laboratory, Abrmlmrr trann. no. 369.) Wwu. iWmunter8. Abt. Helgoland, 15 ( l ) , no. 6, 22 pp.
Maire, D. F., and Sindcmann, C. J. (1960). J~) trw?p: iw variahlit,y in ctJnc:tro- phoretic pat tern of 6sh serum. Anal. iiw. 137, 377-8.
Le hareng Clupea harengua Linnb.
S C ~ . 28, 323-37.

384 J . H. S. BLAXTER AND F. 0 . T. HOLLIDAY
Manteufel, B. P., Lishev, M. N., Radakov, D. V., and Yudanov, K. I. (1968). Observations on behaviour of Clupea hurengua membraa in the Gulf of Riga, when fished by trawl. (In Russian, Marine Laboratory, Aberdeen trans. no. 807.) Trudy V.N.I.R.O. 36, 19-24.
Manteufel, B. P., and Radakov, D. V. (1960). On the study of adaptive im- portance of schooling behaviour in fishes. (In Russian.) Adv. mod. Biol. Moecow, 50, 362-70.
Marr, J. C. (1956). Tho “ critical period ” in tho early life history of marine fbhea. J . Cone. int. Explor. Mer, 21, 160-70.
Marr, J. C. (1967). The subpopulation problem in the Pacific sardine, h’ardinopa caeruleu. Spec. eci. Rep. US. Fiah Wildl. Serv. no. 208, 108-25.
+Marr, J. C. (1961). A model of the population biology of the Pacific mrdine, Sardinops cuerulea. I.C.E.S. Herring Symposium, Paper no. 2 (mimeo.).
Marshall, A. J., and Lofts, B. (1966). The 1,eydig cell homologue in certain teleost fish. Nature, Lond. 177, 704-5.
Marshall. S. M., Nicholls, A. G., and Orr, A. P. (1937). On the growth and feeding of the larval and post-larval stages of the Clyde herring. J. tnur. bwl . Ass. U.K. 22, 246-68.
The fundamental stages of the life cycle of Atlantic Scandinavian herring. (In Russian, tram. in Spec. 8& Rep. US. Fiah Wildl. Serv., no. 327, pp. 6-68a, 1969). Trudy P.I.N.R.O. 9.
M y , Ju. Ju. (1969). Reproduction of marine herring in the light of the in- creasing intensity of fisheries. I.C.E.S. Herring Committee, Paper no. 66 (mimeo.).
Messman, W. H. (1954). Marine fishes in fresh and brackish waters of Virginia rivers. Ecology, 35, ?S-8.
Maxfield, G. H. (1963). The food habits of hatchery-produced pond-cultured shad (Aloea aapid+Gm) reared to a length of two inches. Publ. Cheeupeake biol. Lab. no. 98, 38 pp.
McBride, J. R., Macleod, R. A., and Idler, D. R. (1960). Seasonal variation in the collagen content of Pacific herring tismes. J. Fieh. Re8. Bd Can. 17, 913-8.
McHugh, J. L. (1942). Vertobral number of young herring in southern British Columbia. J. Fiah. Rea. fid C‘un. 5, 414 H4.
McMynn, R. C., and Hoar, W. 3. (1053). EffwtR of rmliriity on tho clovtrlynnent of the I’aciflc herring. Cunacl. J . ZooE. 31, 417-32.
Modrof, J . C . (1967). Nuptial and prnnuptial beheviour of the shad Aloes sap.- dieaim. Copeia, no. 3, 2611-3.
Meyor, H. A. (1878a). “ Biologische Buobachtungen bei kiinstlicher Aufzucht dee Herings dor westlichen Ostsee.” Wiegandt, Hempel and Parey, Berlin.
Meyer, H. A. (187815). Beobachtungen uber daa Wachsthum des Herings in weatlichen Theile der Ostsee. Jber. C m m . wiae. Untersuch. dtech. Meere Kiel, 4, 5, 6, 229-60.
Miller, D. J. (1952). Development through the prolarval stage of artificially fertilized eggs of the Pacific sardine (Surdinope cuerulea). CaZq. Fiah Qame,
Mitchell, J. M. (1864). “The Herring. Its Natural History and Importance.” 372 pp. Edmonston and Douglas, Edinburgh.
Mohr, H. (1960). Zum Verhalten der Fische gogenuber Fanggoraten. (Marine Laboratory, Aberdeon trans. no. 889.) T’rvt. Piachereibchnik, pt. 6, 296-328.
Marty, Ju. Ju. (1956).
38, 687-96.

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 388
Mohr, H. (1961). Note on the behaviour of herring in a round tank. I.C.E.S. Comparative Fishing Committee, Paper no. 86 (mheo.).
Mohr, H. (1962). Beobachtungen an Heringsschwknen. Allgenteine Fisch- urirtechaftteitung, no. 24 (16/6/62-n0 page nos.).
Morgan, T. H. (1927). “ Experimental embryology ”. 766 pp. Columbia University Prese, New York.
Morris, R. W. (1966). Some considerations regarding the nutrition of marine fleh larvae. J. Cone. int. Ezplor. Mer, 20, 266-66.
Morris, R. W. (1966). Some aepects of the problem of rearing marine fishes. BuU. Inut. ochnogr. Motzaco, no. 1082, 61 pp.
*Motode, S., and Hirano, Y. (1961). Review of Japanese herring investigatiom. I.C.E.S. Herring Symposium, Paper no. 13 (mimeo.).
Moulton, J. M. (1960). Swimming aounda and the schooling of fishes. Bwl. Bull. Woo& Hole, 119, 210-23.
Moulton, J. M., and Backhus, R. H. (1966). Annotated references concerning the effects of man-made sounds on the movements of fishes. Coll. R e p . Woo& Hole oceanogr. Inatn. no. 802, 8 pp.
Murphy, C. I. (1960). Introduction of the Marquesen sardine, Harenph vi#ata (Cuv. et Val.) to Hawaiian waters. Pa. Sci., 14, 186-7.
Murphy, C. I. (1961). Oceanography and variations in the Psciflc eardine populations. Californian Cooperative Oceanic Fisheries Investigatione, 8, 66-64.
M u h i 6 , R. (1960). On the schooling and feeding habits of sardine (Sardina p i k h r d w Walb.) in the aquarium (Preliminary ob~ervatiom). Proc. world sci. meeting on the biology of sardines and related species, Rome 1969. (Experience Paper, 17.) 3, 1119-23,
N-ki, F. (1968). The fecundity of the Pacific herring (Clupea palkzuii) in British Columbia coastal waters. J. Fbh. Ree. Bd Can. 15, 313-30.
Nseh, J. (1931). The number and size of glomeruli in the kidney of Mes, with observations on the morphology of the renal tubules of Wes. Amcr. J . And. 47,426-46.
Naumov, V. M. (1966). “ The ovogenesia and ecology of the sexual cycle of the Murmansk herring (Clupea harengzcs L.).” (In Russian, trans. in Spec. uci. Rep., U.S. Fiah Wildl. Sew., no. 327,203-62, 1969.) Trudy P.I.N.R.O. 9.
Nikitinskaya, I. V. (19688). On the onset of active feeding of the larvae of Clupea b r e w pcrllaeii Val. (In Russian, Eng. summary.) 2001. Zh. 37,
Nikithkaya, I. V. (1968b). On the variability of the larvae of the 13akhalin herring. (In Russian, trans. U.S. Fish and Wildlife Bervico, Jloothbay Harbor, Maine.) Nauch. Doklady Vyehey Shkoly, Uiol. Nmk. , no. 4.
Nikolsky, G. W. (1967). “ Spezielle Fiechkundo ” 632 pp. V.E.13. Jlsutrchnr Verlag der Winsernhaften, Berlin.
*N&oleky, 0. W., Chepunov, A. V., and Rhatunovnky, M. I. (1861). Wuleribiw in variability of features in certain forma of North Atlmtio herring. I.C.E.S. Herring Symposium, Paper no. 38 (mimeo.).
Fishery for Caspian Clupeonclla by meBne of mction pilmp and underwater illumination.” (In Ruseian, Mberine Leboretory, Aberdeen trans. no. 647.) 96 pp. Pishcheproniizdat, Moscow.
1668-71.
Nikonomv, I. V. (1068).

386 J. H. 9. BLAXTICR AND F. 13. T. HOLLIDAY
Nikonorov. I. V., and Patejev, A. Kh. (1969). The Gshery for Kilka (sprat) with fish pump, combined with underwater light and impulse-current. (In Russian, Marine Laboratory, Aberdeen trans. no. 686.) Ryb. Khoz. 7, 53-8.
Nomura, M. (1968). Some knowledge on beheviour of fish schools. Proc. Indo-
O'Connell, C. P. (1955). The gas bladder and its relation to the inner ear in sardinops caermh and Engraulis mordux. Fish. Bull. U.S. Fish Wildl. Sew. no. 104, 603-33.
Okoneki, S., and Konkol, H. (1967). Daily migrations of herring shoals and their reaction to fishing gear. (In Polish, Eng. summary.) Prace Morsk. Inat. Rybackiego Qdyni. 9, 649-03.
Outram, D. N. (1951). Observations oii the rotention and spawning of the Pacific herring. Progr. Rep. biol. Stm Nanaimo and Prince Rupert, 87, 32.
Outram, D. N. (1956). The development of the Pacific herring ogg and its use in estimating age of spawn. Fish. Res. Ud Can. Pacific Biot. Stat., circular no.
Owatari, A., Furuya, K., and Furuya, K. (1953). The behaviour of the sardine schools by fish detector. Bull. Jap. SOC. sci. Fish. 18, 669-74.
Pam, A. E. (1927). A contribution to the theoretical analysis of the schooling behaviour of fishes. Occasional Papera of the Bingham Oceanographic Foundation, 1, 1-32.
Parrish, B. B., and Craig, R. E. (1957). Recent changes in the North Sea herring fisheries. Rapp. Cow. Explor. Mer, 143, 12-21.
Parrish, B. B., Blaxter, J. H. S., and Holliday, F. G. T. (1958). Herring (Clupea hurengus L.) in aquaria. I. Establishment. Mar. Res. Scot. no. 6, 11 pp.
Parrish, B. B., Saville, A., Craig, R. E., Baxter, I. G., and Priestley, R. (1959). Observations on herring spawning and larval distribution in the Firth of Clyde in 1958. J. mar. biol. Ass. U.K. 38, 446-53.
Parry, Gwyneth, and Holliday, F. G. T. (1 960). An experimental analysis of the function of the pseudobranch in teleosts. J . exp. Biol. 37, 344-54.
Parry, G., Holliday, F. G. T., and Blaxter, J. 13. 5. (1969). Chloride secretory cells in tho gills of toleoets. Nature, 1,ow.l. 183, 1248-9.
Pavshtika, E. A. (1966). fjeturonfil c h q w 111 p h ~ I k t A m atid fiwhiig t i i i ~ M i i * i m C b f herring. (In Russian, trans. in 8 p w . *ti. &TI. I1.N. ff%h WiUL. timi., 1 j l J . 321, 104-39.) Trudy P.I.N.R.O. 9.
Pearson, J. C. (1952). Rearing young Hhrtd iri rmnrlH. Prog. d a n h . f h l t . 14, 3.3 6. Perceva, T. A. (1939). The devolopmont of tho Orutpiari did, f , ' ~ w / , U i b ~ ~ i n y h i
(Eichw.). (In Russian. Eng. summary.) Truly V.N.I . II .0 . 8, 27 05. Phillips, J. B., and Radovich, J. (1962). Survoy~ through 1961 of tho distribution
and abundance of young sardines (Surdinope eaerulea). Fiah. BuU. Sacra- mento, no. 87, 63 pp.
Pinto, J. dos Santos (1963). Um cmo de Hermafroditismo em Sardino pikhurdw, (Walb.). (Eng. summary.) N o h Estud. Inat. Biol. mri t . , Lisboa. no. 6,
Pinto, J. dos Santos (1964). Note on a new cam of hermaphroditism in Sardina pi&rdwr (Walb.). Rev. Fac. Cienc. Univ. Lbboa, 4, 120-2.
pinto, J. do8 Santos (1957). Essai d'une Bchelle de maturation sewelle pour m&s de le sardine (Satdim pilchsdzrs Walb.) baa& sur l'histologie des testiculee. (Eng. summary.) N o h Edud. Inat. Biol. w r i t . , Lisboa. no. 16,
Fish. Coun. 3, 95-6.
40.9 pp.
5 PP.
37 PP-

Pinto, J. dos Santos, and Andreu, B. (1966). Algunos espectos des deaarrollo initial del ovario de sardina (Sardina pi,!&zrdua Walb.). Inu. Pmq. 5, 3-12.
Polder, J. J. W. (1961). Cyclical changes in testis and ovary related to maturity Stages in the North Sea herring, Clupea hare- L. Arch. nbr l . .%~l. 14, 46-60.
Polonskij, A. 3. (1961). Verhalten des Hering8 im Winter. (In Russian. Restock Inst. fur Hochseefischerei trans. no. 1422.) Ryb. Khoz. 37 (2), 22-7.
Postma, K. H. (1967). The vertical migration of feeding hemhg in relation to light and vertical temperature gradient. I.C.E.S. Herring Committee (mimeo.).
Prince, E. (1907). The egga and early life history of the herring, gespereau, shad and other clupeoids. Cmtrib. Can. Bwl., Suppl. 39th Ann. Rept. Dept. Mar. Fish., Fish. Branch, pp. 96-110.
Protasov, V. P., Altukhov, Y. P., and Kovaleva, N. D. (1960). MorphololJical and functional features of the transition from day vieion to twilight vision in some Black Sea fish. (In Russian, trans. in Dokl. biol. Sci. Sect. 134, 694-6.) Dokl. A M . Nauk. S.S.S.R.. 134, 196.
Radakov, D. V. (1900). Observations on herring during a voyage of research submarine " Severyanka ". (In Rumian, Marine Laboratory, Aberdaen trans. no. 718.)
Radakov, D. V. (1962). On tho behaviour of some f l h in trawls. I.C.E.S. Com- parative Fishing Committee Paper no. 120 (mimeo.).
Radakov, D. V., and Solovyev, B. S. (1969). A first attempt at using a submarine for observations on the behaviour of herring. (In Russian, Fish. Ree. Bd Can. trans. no. 338.) Ryb. Khoz. 35 (7). 16-21.
Redovich, J. (1952). Food of the Pacific sardine Sardinops eaefillecr from central Baja, California, and from southern California. Cat$. FGh Game, 38,676-86.
Radovich, J., and Gibbs, E. D. (1964). The use of a blanket net in sempling 5 h populations. Calif. Fish (Jam, 40, 363-66.
Ramanujam, S. G. M. (1929). The study of the development of the vertebral column in teleosts as shown in the life history of the herring. Proc. M O I . Soc. W. pp. 365-414.
Rannak, L. A. (1968). Quantitative study of the Baltic herring eggs and larvae in the northern part of tho Gulf of Riga and principle factore determining their survival. Trudy
Race, T. S. (1937). Pigmentation of embryos and larvae in the heminp (faniily Clupeidae) aa an adaptation to a pelagic mode of life. (In Humian, JSI~K. summary.) BuU. SOC. Nat. Moacozu, 46, 165-64.
Retzius, G. (1906). Die spermien der Loptoknrdiw, Toleo~tior untl ~ h t i o d ~ m . Biol. Unkreach. 12, 103-16.
Richardson, I. D. (1962). Borne reactions of pnlagic: fiHh to light am mcorrfiwl hy echo-noundirig. Fbh. Inverrt. rand.. h'w. 11. 18 ( I ) , 19 pp.
Itic~litrrclHoit. 1, I ) . (IfW)). O l i ~ t ~ r v i ~ t iona o i i tho HIZO irnd numbers of herring taken Iby t Iw Iicw'riiig (rttwl. . I . ( h ~ . int. Erpbr. Mar. 25, 204-9.
Iticlowcwtl. 11'. (i. ( I H U l ) . 'I'lio trir bladtior and Oar of Brit,ish clupeoid fiehea. J.
Itogtilln, 15. 11 .. t i r i d Stihrhngo, D. (1900). lieringworkommen und Wasaertem-
Byull. okeanogr. Kom. A M . Nauk. S.S.S.R., 6, 39-40.
(In Ruesian, Fish. Res. Bd Can. trans. no. 238.) V.N.I.R.O. 34, 7-18.
Ai&tl. LoPtd . 26, 20-42.
jwrtit iir. InJolm. E'k!ac:hru. 7, 136-8.
26

388 J. R. 9. BLAXTER AND F. 0. T. HOLLIDAY
Ronold, 0. A., and Jakobsen, F. (1947). Trimethylamine oxide in marine products. J. SOC. chem. I d . Lond. 66, 180-8.
Rosen, M. W. (1981). Experiments with swimming fish and dolphins. Amer. SOC. mech. Engineers, Paper no. 81-WA-203.
Rounsefell, G. A. (1930). Contribution to the biology of the Pacific herring Clupeu pallasii, and the condition of the fishery in Alaska. BuU. U.S. Bur.
Rounsefell, G . A., and Dahlgren, E. H. (1932). Fluctuations in the supply of herring Clupea pallaeii in Prince William Sound, Alaska. BuU. U.S. Bur.
Rumyantsev, A. I. (1980). Damage to herring smolt in their natural environment. C.R. A&. SCi.,
tfber die Rasmnverhaltnisse bei dem norwegiachen Friihjahrshering mit besondorer Beriicksichtigung der Konstanz der Ramen- merkmale. J. C m . int. Explor. MeV. 8, 236-49.
Runnstrem, S. (1941a). Racial analysis of the herring in Norwegian wetere.. Fiakeridir. Skr. Hawnderaok. 6 (7). 110 pp.
Runnstrem, S. (1941b). Quantitativo investigations on herring spawning and its yearly fluctuations at the west cowt of Norway. FGkeridir. Skr. Hawn- deruek. 6 (S), 71 pp.
FGh. 45, 227-320.
Fish. 41, 283-91.
(In Russian. tram. in Dokl. biol. Sci. Sect. 134, 683-6.) U.R.S.S. 134, 966-8.
Runnetr0m, S. (1933).
Russell. E. S. (1937). Fish migrations. BWZ. Rev. 12, 320-37. Russell, F. S. (1928). The vertical distribution of marine macroplankton. III.
Diurnal observations on the pelagic young of teleostean fisheg in the Ply- mouth area. J. mar. biol. Ass. U.K. 14, 387-414.
The vertical distribution of marine macroplankton. VIII. Further observations on the diurnal behaviour of the pelagic young of teleostean fishes in the Plymouth area. J. mar. biol. Ass. U.K. 15, 829-60.
Russell, F. S. (1930). The vertical distribution of marine macroplankton. IX. The distribution of the pelagic young of teleostean fishes in the daytime in the Plymouth area. J. mar. biol. Aea. U.K. 16, 639-70.
Ryder, J. A. (1882). On the retardation of the development of the ova of the #had (Alosa aapidiaaima) with ObservationH on the egg fungun and bwteria. ~ U U .
Ryzhenko, M. (1981). " Severyanka " in the schools of herring and cod. (In Rybovodstvoi i
Sahrhage, D. (1954). Beobachtungen zur Yrage der Iiering~l~ichj)l t l t ,~. Mitt. Inat. Seejiach. 6 , 24-32.
Sasaki, T., Watanabe, S., Oshiba, G., ant1 Okaini, N. (1968). Mwuriircminntn of angular dktribution of submarino daylight by mnaris of II new innt,ninimt,. J . oceanogr. Soc. Japan, 14 (2). 1 -6.
Savage, R. E. (1931). The relation botwwn tho feeding of tho t ictrrin~ off tho a ~ , comt of EnKlantl and the plankton ol' tho nurroirnding watom. FiRh. fm&. Lond., Set. I I , 12 (3), 88 pp.
Savage, R. E., and llodgtion, W. C. (11)34). Lunnr influence on tho East Anglian herring fishery. J. Cone. in& Explor. hfer. 9, 223-40.
SCBttOrRood. L. W. (1967). A bibliography of the herring (Clupeu hnrcngua and C ~ u p m p a l h i i ) . ItoHnarc!h Bulletiri no. 28, 108 pp. Department of See and Shorp Fisheries, Augusta, Maine.
Russell, F. S. (1928).
US. FGh. COWWTI. 1, 177-90, 422-4.
Russian, Marine Laboratory, Aberdeen trans. no. 718.) ~ b o l o ~ t v o , 4, 29-30.

THE BEHAVIOW AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 389
Scattergood, L. W. (1969). (Translator.) Herring of the North European Baain and adjacent seas. (Translation of nine Ruseien papers.) S p . 8ci. Rep. US. Fish Wildl. Sew. no. 327. 284 pp.
Schach, H. (1939). Die kiinstliche Aufzucht von CZupeu harengus. (Marine Laboratory, Aberdeen trans. no. 307). Helgoldnd. W k . Meereaunters. 1, 359-72.
Schaefller, M. B. (1937). Notes on the spawning of the PacSc herring (CcUpea pallwii). CopacC, no. 1, 67.
Schiifer, W. (1966). ober das Verhelten von Jungheringaechwamen hn Aqua- rium. Arch. FischJf”is8. 6, 276-81.
Schiirfe, J. (1953). tfber die Verwendung kiinetlichen Lichtes in der Fiecherei. Prot. Fischereiteohnik, 8, 1-29.
Schiirfe, J. (1955). Observations with echo-sounding on the behaviour of a herring shoal towards a bottom trawl. 1.C.E.S. Comparative Fishing committee, Paper no. 42 (mimeo.).
Scharfe, J. (1960). A new method for aimed one-boat trawling in midwater and on the bottom. S t d . Rev. gen. Fiah. Coun. Medit. no. 13, 38 pp.
Schnakenbeck, W. (1929). Entwicklurigsgaschichtliche und morphologieche Untereuchungen am .Hering.
Schubert, K. (1960). FiidbereiurcZt, 2nd year, 149-61.
Schiiler. F.. and Krefft, 0. (1961). Versuche zur Beeinflussung von M e e m - fischen durch Schalldruckwellen und kunxtlichea Licht. Fiachereiwelt, 3rd year, 8-10.
Schutz, F. (1966). Vergleichende Untersuchungen iiber die Schreckreaktion bei Fhchen und deren Verbmitung. 2. vergl. Phyaiol. 38, 84-136.
Shew, E. (1862). The schooling of fishes. Sci. A m r . 206, 128-38. Shelbourne, J. E. (1957). Sitea of chloride regulation in marine fish larvee.
Nature, Lund. 180, 920-2. Shelford, V. E., and Powers, E. B. (1916). An experimentel study of the move-
ments of herring and other marine fishee. BWE. BuU., W d Hole, 28,316-34. Shewan, J. M. (1951). The chemistry and metabolism of the nitrogenous ex-
tractives in fish. Bwchem. SOC. Syrnpoak C a d r i d g e &&. no. 0, 28-48. Shewan, J. M. (1955). The nitrogenous extractives from fresh h h mrinc:lc~ 3.
Comparison of several flat fishes and member8 of the herring-mackerol fffJUp. J. mi. Fd Agric. no. 2, 99-104.
Shubnikov, D. A. (1960). Different responses in mde and female kilka ClupeMIsua engradifomis (Borodin) to elect,ric light. 2001. Zh. 38, 781-2.
Silliman, R. P. (1960). Thermal and diurnal chenqea in the vertical chtribiition of eggs and larvae. Spec. sci. Rep. U .S . Pirh H’ildi. Sew., nlJ. 16, 181-.200.
Sindemann, c. J. (1956). I k x w f m of finhw ol 1 . h j wc:xtiirri Nor1.h Atli~iitic. IV. Fungi18 diswiu nnd rtwltarit rriorhIit,if~t of‘ tJ<.rriiig irr ttiu (;iilf‘ o r Hf.. Lawrence in 1055. I h p l . SPLJ Shore IGtth. Mrrinc / h a . I h U . 25, I . 23.
Sindemann, C. J. (1987). Uiso,cteRcw of fixhos of t h r r wiutt.orii North A t h l t i f : . V. Paraaitex an iriilicatort~ of tiorriiig r i i o v m t ~ ~ ~ i i t , ~ . IJspt. Serr Shore Pwh. Maine Hea. RuU. 27, 1-30.
SiniIwwuw~, (1. , I . ( I !)AH). A i i l , i ~ t i ~ ~ ~ i i i t ~ i i ~ l i i L I i cvyt.lirrwyto I)roprbitw of Roa hnrring NIWIII~. / I J I ~ I / . 1ti.c:. 131, 60U.
Siiidoririalm ( 1 . J . . i ~ i i t l Mibiiw, I). 1‘’. (IUBH). Serum ~irot~uiii c!hanga in dkcleed 808
litwring. Attat. Itrc. 131, 509.--600.
Ber. dtuch. Ki~mm. Meere8fwsch. 5, 23-78. I)or Heringsfang iin Juli in der Nodsee.

390 J. R. 8 . BLAXTER AND F. 0. T. ROLLIDAY
Sindennann, C.,J., and Mairs, D. F. (1959a). Blood properties of pro- and poet-
Sindermann, C. J., and Mairs, D. F. (1969b). A major blood group systum in
Sindermann, C. J., and Maim, D. F. (1960). Comparative serology of five species
Skoglund, G. (1953). Skrammer ekold fisken? Ostkwten, 25 (6). 14. Smith, K. A. (1955). Use of an electrical attracting and guiding device in ex-
Smith, K. A. (1961). Air curtain fishing for Maine sardines. Comm. Fiah. Rev.
Soleim, P. A. (1942). Arsaker ti1 rike og fattige &ganger av Sild. Fiskeridir. Skr. Havundersek. 7 (2), 39 pp.
Solovyev, B. S. (1969). On vertical distribution of herring at the end of its feeding season. (In Russian.) Ryb. Khoz. 35 (3). 9-12.
Solovyev, B. S., and Degtyarev, A. A. (1957). Diurnal vertical migrations of the Atlanto-Scandian herring during the pre-spawning and spawning sea~om. (In Russian.) Ryb. Khoz. 33 (ll), 81-4.
The distribution of some plankton anhala in the English Channel and approaches. 111. Theories about long-term biological changes, including fish. J. mar. biol. Ass. U.K. 43, 1-29.
Spooner, G. M. (1931). Some observations on schooling in fish. J. mar. biol. dee.
Spoonur, 0. M. (1933). Obsorvations on tlio reactions of marine plankton to light. J. mar. biol. Ass. U.K. 19, 386-438.
Spraguo, I,. M., and Vronrnan, A. M. (1962). A racial analysis of the' Pacific sardine (Sardinops caerulea) baaod on studies of erythrocyte antigens. Ann.
Springer, S. (1957). Some observations on the behaviour of schools of fishes in
Steele, J. H. (1961). The environment of a herring fishery. Mu?. Rea. Scot. no. 6,
Stevenson, J. C. (1947). Preliminary study of larval herring an tho went cnwt of Vancouver Island, 1947. Progr. Rep. bwl. Slae N n n a h and pram^ t lupr t , no. 73, 65-7;
Distribution and survival of herring larvae (CZupm palhsii, Valenciennes) in British Columbia waters. J. Fish. Res. Bd Cun. 19, 735-810.
Stirling, W. (1884). On the chemistry and histology of the digestive organs of fishes. Rep. Fish. Bd i3cot. no. 2, 31-46.
Stringham, E. (1924). The maximum q m d of freshwater fishes. Amer. Nai.
Stroganov, N. S. (1938). The Burviva1 capacity of sperm in tho Volga herrirrg
spawning ariadromous alewives. Anat. Rec. 134, 639-40.
Atlantic sea herring. Copeiu, 3, 228-31.
of Atlantic clupeoid fishes. Anat. Rec. 137, 393.
perimenting with a " fish pump ". Comm. Fieh. Rev. 17 (2), 1-7.
23 (3). 14 PP.
Southward, A. J. (1963).
U . K . 17, 421-48.
N . Y . A d . Sci. 97, 131-8.
the Gulf of Mexico and adjacent waters. Ecology, 38, 166-71.
19 PP.
Stevenson, J. C. (1962).
58, 156-61.
(CuspioZosa volyensis) under external corrdit i o r i ~ . Zool. %h. 17, 31 6.
Rep. 1J.S. Zhh W i l d . , S e r ~ , w l . P,2, 57 pp. Suehiro, Y. (1951). A HtUdY of t , t l l> f:iLIJHC'i ( J f t)io l j W $ L h f>f tmit. fj#hf%. 6Spw:. jy&.
Svetovidrjv, A. N. (19.52). " F~iiiria S.H.S.R. K i h m ". 2, IIO. 1. ( ; I i i r m i ~ I t w . ( I r l
I JigtLtz. hl. 14:. ( l i j 6 1 ) . ' ~ ' i ! i ~ + r i h r i i x ~ of Itl.rij!otl ~JRNH iLnc1 Airwric?en shlL(1 L o C:~&IIKOR of I ~ ~ I I I I ~ ~ ~ ~ . ~ L I I I I ' ( ' ruul H d i i i i t . y . SIif'c. 8c.i. &p. lJ.A'. IG9h Wi,!&. Setz). 110. 388, 8 pI1.
H I I S S ~ ~ L ~ I . ) %/d. I I IN~. . 4 k ~ t L . ,S.,Y.,';.jt., N ~ w . ,SW. 110. 4H. 3:11 rip. < ,

THE BEHAVIOUR AND PHYSIOLOGY OF HERRING AND OTHER CLUPEIDS 391
Tekeheahi, M., and Hatanaks, M. (1960). Experimental study on utilization of food by young anchovy, Enqradk japonicua Temminck et Schlegel. Tohoku
Teeter, A. L. (1937). Population of herring (Clupea pmUaSii) in the coastal watsre of British Columbia. J . FWh. Rea. Bd Can. 3, 108-44.
Teeter, A. L. (19388). Variation in the mean vertebral count of horring (Clupeu paltclpii) with wnter tomperaturo. J. C m . inl. Expbr. Met. 13.71-5.
Teeter. A. L. (1938b). Herring, the tide and tho moon. Prop. Rep. 6wl. S ’ b S
Nanaim and Prince Rwpert, no. 38. 10-4. Teeter, A. L. (1943). Uae of the echosounder to locate herring in British Columbia
waters. BuU. Fish. Rea. Bd Can. no. 63, 21 pp. Tibbo, 8. N., and Brawn, V. M. (1960). Exploration for herring in the Bay of
Fundy and Gulf of Maine. J. Fish. Res. Bd Can. 17,736-7. Tibbo, 8. N., Legad, J. E. H., Scattergood, L. W., and Temple, R. F. (1968).
On the occurrence and distribution of larval herring (Clupeu harengua L.) in the Bay of Fundy and Gulf of Maine. J. Fhh. Re8. Bd Can. 15, 1461-69.
Supplementary data on the reaction of herring to electric light. (In Russian, Marine Laboratory, Aberdeen trans. no. 694.)
Toom, M. M. (1958). Experiments in the incubation of Baltic herring eggs. (In Russian, trans. available from Office of Technical Services, US. Dept. of Commerce, Washington 25, D.C.) Trudy V.N.I.R.O. 34, 19-29.
Ulrich, B. (1951). Unterwasserbeobachtungen von Heringsschwamen. Fi8cherei- welt, 3rd year, 164-5.
Valdez, V., and Cushing, D. H. (195Q). The effect of tidal stream on the prosence of an extensive layer of midwater fish traces. I.C.E.S. Herring Cornniiltce, Paper no. 45 (mimoo.).
Verheijen, F. J. (1953). Laboratory oxporirnonts H ith ttro horring CZU~J~*~L hrrrengua L. Experierdci, 9, 193-7.
Verheijen, F. J. (19G6). On a method for collorting rtnd koeping cliiprmid~ for uxperimnntnl purpoeos . . . . Pubbl. Skrz. z o d . Napdi, 28, 22G- 40.
Verheijen. F. J. (1068). Tho mochaniRmR of l h o 1 rnpping otToc1 of artili(*iul light
Vortioijori, 14’. . I . (111601~). A Imvulisr I I ~ H ~ . I L K I I I I I H rrr i t l ~b i ~ i ~ r ~ ~ ~ ~ ~ i i ~ r i t l i ~ i ~ f1)vvtt.l
Atriirtim) iri tho (‘yo of tho tiorririy ((!lupetr lulrmyun I,.). h ’ q ~ ~ r i s n t ~ t i , 15, 443 -4.
Vorlioijori, k’. .I. (1f)Af)b). AbHoncW of alann Hirbstanco in some clupoids. I’ubll. Nbrz. zool. Napoli, 31, 146-52.
Vernborg. F. J. (1954). Tho respiratory metabolism of tiwucs of marine telcosts in relation to activity and body size. Biol. Bull., Wood8 Hole, 106, 360-7.
Vernberg, F. G., and Gray, I. E. (1963). A comptirative study of the respiratory metabolism of excised brain tiseue of marine teleosts. Biol. Bdl . , JvOod8 Hole. 104, 446.
Vilter, V. (1950). Adaptation biologique de l’appamil visuel et les structures dtiniennea de la sardine. C.R. SOC. BWL Pa&, 144, 200-3.
Vincent, R. E. (1960). Experimental introduction of fresh water alowives. Prog.
Volodin, V. M. (1966). Embryonic development of‘the autumn Baltic herring and their oxygen requirement during the coiirm of c1ovelol)mimt. ( IJI ~ ~ I J H H I L I I ,
Fish- RM. Bd Can. trarw. no. 262.) V o p r o q /k/ttai&/yi, no. 7, 123 33.
J . q r i C . R-. 11, 161-70.
Tikhinov, D. P. (1960).
Ryb. Khoz. 4, 13-14.
HOIIPCOH ilp(WI I L I ~ i t t t ~ h . Arch. 7 ikCd. %JO1. 13, 1 107.
FWh. Cult. 22, 38-41.

392 J. H. S. BLAXTER AND F. 0 . T. HOLLIDAY
Waldmann, J. (1961). Untersuchungen an Heringslarven und Zooplankton dee (Eng. summary.)
Watson, J. E. (1961). Branding as a short term mark. Prog. Fish. Cult. 23, 106. Weisman, A. L., Coates, C. W., and Moses, R. L. (1936). An investigation of the
presence of oestrogenic hormones in the ova and ovsrios of fish. Endo- crinology, 20, 661-2.
Wells, E’. It. (1022). On tho morphology of tlie chondrocranium of tho larval herring (Clupea hurengua). l’roc. 2001. SOC. Lond. 1213-29.
Westman, J. R., and Nigrelli, R. F. (1965). Preliminary studies of menhaden und their mas8 mortalities in Long Island and New Jersey waters. N.Y. Fish &me J . 2, 142-53.
Williamson, H. C. (1909). Exporimonts t,o show the influence of cold in retarding tho dovelopment of the eggs of the herring (Clupea harengw, L.), the plaice (Pleuronectes platessa L.) and the haddock (Gadw aeglejinus L.) Rep. Fish. Bd Scot. no. 27, 100-28.
Williamson, H. C. (1917). On the herring in captivity and an effect of the en- vironment on the structure of this fish. J. 2001. Res. 2, 101-10.
Williamson, H. C. (1945). Devolopmcnt of the roo and milt of the herring. Independent lives. Fish. News, Oct, 6, 1945, p. 10.
*Williamson, M. H. (1961). The relation of plankton to some parameters of the herring population of the north-western North Sea. I.C.E.S. Herring Symposium, Paper no. 11 (mimeo.).
Das Ohrlabyrinth der Sardine (Clupea pikhardw Walb.) und seine Beziehungen zur Schwimmblase und Seitenlinie. 2. Morph. Okol. Tiere, 31, 371-410.
Wohlfahrt, T. A. (1937). Anatomische lintersuchungen iiber die Seitenkanale der Sardine (Clupea pikhardU8 Walb.). 2. Morph. Okol. Tiere, 33, 381-Pll.
Wood, H., Parrish, B. B., and MacPhorsoil, G. (1958). Review of Scottifih herring tagging exporinients 1948-1983. IZapp. Cons. Explor. Mer, 140, 35-84.
Wood, Ti. J. (1988). Fat cycleR of North Sea herring. J. Cons. int. Explor. Mer, 23,
Greifswalder Boddens in den Jahren 1958 und 1959. Zeit. Fisch. Hi{f&s. 10, 523-36.
Wohlfahrt, T. A. (1936).
390 -8. kV~JfJdhf!lld, 1’. M. . j . , UlJ(1 WoO~lh~*llfl, A . f j. fl!jfifiJ, ‘ j ’ h rwu;fl~tfi* Of l i r r Y l i l l K InrVffn
M’~I I IW I ~ ~ ~ I w I ~ I ~ I I ~ . V . ( I , ( IIjW). ‘I’lii~ rlrylri~iIiic:t,ivct I I ~ K I L I I H of I I I O tIi*rrIiIg in rolulion
Wynrio-I‘,clwnr(l~, V. C. (IO(12). “ Aniriiul 1hq)orsion in ltolatioll ‘to Social Hehaviour.” 663 pi). Olivur and Ihyd, Ndinburgh and London.
YiuiiiLpLwit, M., nrul Nintiiniurrc, S. (1!)27). Chemical study on tho Hpenn of the mririnu itiiirxiul. V. 0 1 1 the hydrolysis of testis of the herring, Clupea pallaeii. (In Japanese.) J . Fish. Inat. Tokyo, 22, 85-8.
Yememoto, K. (1955). Studies on the formation of fish eggs. 5. The chemicei nature and the origin of the yolk vesicles in the oocytes of the herring, Clupeu p a l b i i . Annot. Zool. Jap. 28, 158-62.
Yamamoto, K. (1956). Studies on the formation of fish eggm. 7. Tho fate of the yolk vesicles in the oocytes of the herring, Clupea pallaeii during vitello- genesis. Annot. zool. Jap. 29, 91-6.
(In Japanese.) Jap. J . Ichth. 4, 182-92.
t o light : 1% r r i w ~ t i i i r i i H r r i of‘ vorl i c d rr~iyrrrt ifm. A‘rti?Jrt, /,/ut</. 178, J I H 60.
p ~ ) w l l i . J . ?mr. hid, .4Na. (J.k’. I(;, 4 0 06.
Yamamoto, T. S. (1955). Ovulation in the salmon, herring and lamprey.

THE BEHAVIOUR AND PRYSIOLOOY OF HERRINQ AND OTHER CLUPEIDS 393
Yamamoto, T. S. (1958). Biochemical property of the membranes of the herring egg with special reference to the role of the micropyle in fertiliza- tion. J. Fac. Sci. Hokkaido Univ., Ser. 6, Zool. 14 (I), 9-16.
Effect of environmental salt concentration on fer- tilizability of herring gametes. J. Fac. Sci. Hokkaido Univ., Set 0 , Zool. 11,
Yanagimachi, R. (1957). Studies of fertilization in Clupea 13&uaaii. IV. Some properties of the sperm-stimulating factor in the micropyle area of the mature egg. Bull. Jap. SOC. aci. Fieh. 23, 81-5.
Yanagimarhi, R., and Kanoh, Y. (1953). Manner of sperm entry in herring egg, with special reference to the role of tho calcium ion in fertilization. J. Fa. Sci. Hokkaido Univ., Ser. 6 , Zool. 11, 487-94.
Yudaev, H. A. (1950). The content of histidine, carnosine and anaerine in the muscle of several fish. Trans. A M . Sci. U.S.S.R. 10, 219-82.
Zaitsev, V. P., and Radakov, D. V. (1960). Submarines in fiahery research. (In Russian, Marine Laboratory, Aberdeen trans. no. 069.) “VNIRO and PNIRO. Soviet fisheries investigations in north European eaas.” 408 pp. Moscow.
*Zijlstra, J. J. (1961). On the recruitment mechanism in the North Sea herring. I.C.E.S. Herring Symposium, Papor no. 29 (mimeo.).
Zusser, S. G. (1958a). A study of the causes of diurnal vertical migration in fiehes. (In Russian, trans. by U.S. Fish and Wildlife Service, Boothbay Harbor, Maine, U.S.A.). Ikht. Kom. A M . Nauk.S.S.S.R. no. 8, 116-20.
Zuaser, S. G. (1958b). A contribution to the study of 5sh behaviour. PYOC. Indo- Pmi& Fish. Coun. 3, 34-1.
Yanagimachi, R. (1953).
481-6.
*Papers in 1901 I.C.E.S. Herring Symposium eventually to be published as a special volume of Rupp. Cons. Exploy. iMer.

This Page Intentionally Left Blank

Author Index x-0 in W ~ W mjw to poqsr on d h h qfmnca oppsar d h~ end of ths ah-.
A
Aaaen, 0.. 360, 372 Abbott. D. P., 224, 259 Abe. N., 246, 255 Abel, E. F., 217, 255
300. 307, 372
197. 201, 202.204 Wen, M. B., 237,255 Wen, R. D., 121,130 Allen, W. E., 327, 372 Allieon, E. C., 216. 256 Almy, L. H., 298, 372 Altukhov. Y. P., 316,387 Anwllin, J., 304,372 Andemon, K. A., 304,372 Andreev, N. N., 361.372
Andrew, C. B., 286, 372 Andrew, J. D., 81.132 Anokhina. L. E., 282, 301, 372
Barlow, Q. W., 271, 370, 373 Barnerd, J. L~wM, 216.266 Barnett, C. H., 194, 201 Bateeon, W., 176, 178, 196, 202 Battle, H., 124, 130 Battle, H. I., f.294, 296, 286, 298, 299,
Bauchot, M.-L., 106,166
Bayliea, L. E., 178, 202 Beylor, E. R.. 183, 202 Besufort, L. F. de, 373 Belding, D. L., 2, 22, 98, 99, 123. 124,
Belyaeva, V. N., 360, 373 Bennett, I., 231. 240, 259 Bergmaam, A. M., 163, I66 Berland, B., 170, 202 Berndt, Oswald, 162, 166
Bertin. L., 142, 166 Beverton, R. J. H., 306. 300, 373 Bhattecharyya, R. N., 276, 270, 373 ~ i b ~ , N. E., 280, 361, 370, 373 Bigelow, H. B., 143, 147, 166.166 T3iryirkov. N. P., 333, 336,342, 346,373 I h t r o i , I f . M., 283, 207, 288, 289, 291,
373, 374 Blexter, J. H. S., 207, 270, 271, 272.
273, 274, 276. 270, 277, 278, 279, 280, 281, 282, 283, 284, 286, 287, 289, 291, 293, 294, f.294, 296, 290, 299, 300, 303, 310, 311, 312, 313, 314, 316, 310, 318, 322, 324, 326, 321, 330, 340, 349. 360, 362, 363. 364, 303, 308, 309, 370, 374, 380, 386
Ahletrom, E. H., 270, 271, 290, 306, 300. 373
Mi, A*, 179, 180, 181, 18% 184s 193, M ~ , 1. GI., 273, 360, 365, 373, 386
130
Andreu, B e , 206,2Q3,366 30% 372,387 %wffy, L. van, 303, 373
Anon., 273, f.294, 286, 316, 340, 372,
Anmll, A. n., 53,132 Anvy. I,. 11.. 178, IHO. 201
Arnow, W., 224, 259 Arthur, D. K., 274, 276, 270, 277, 283,
Atode. K. 216, 218, 219, 232, 255 Austin, T. 8.. 251, 259 Awerinzew, 8.. 300. 373
3 73
Armstrong. I!. A. J., 191. 801
373
Boslch. G. T., 191, 201 Boden, B. P., 189, 202 Boleter. G. C., 328, 304, 374 Boriejak, A., 90, 130 Borieov, P. Q., 316, 349, 360, 361. 374 Boaohma, H., 216, 217, 266 Boury, M., 102, 130
B Baburim, E. A., 314, 373 Baokhus, R. H., 318. 385 Ballowitz. E., 264, 373 Belle, R.. 328, 342. 343. 347, 362, 373 Bamford, T. W.. 274, 319, 320, 373 Berdech J. E., 230. 255, 286, 310, 382 21 186

396 AUTHOR INDEX
Bowers, A. B., 266, 276, 280, 356, 358, 360, 364, 374, 375
Bowman, A., 273, 375 Boyar, H. C., f.294, 323, 324, 375 Bradly, J. S., 225, 258 Brmmer, W.. 199, 202, 204, 206, 341,
Brandes, C. H., 301, 307, 320, 375 Brandhorst, W., 271, 375 Brandt, A. von, 197, 198,202, 351, 375 Brauer, A.. 175, 202 Brawn, V. M., f.294, 310, 312, 321, 322,
324, 328, 342, 344, 346, 346, 347, 375, 391
Uretlor, C. M., Jr., 173, 174, 175, 106, 196, 202. 275. 289, 296, 320, 327, 328, 375
379
Bretschneider, L. H., 375 Brett, J. R., 171, 182, 197, 202 Bridger, J. P., 281, 290, 364, 374, 375 Brook, G., 268, 375 Brooks, W. K., 20, 22, 26, 35, 130 Brown, M. E. (Ed.), 263, 305, 375 Bruce, J. R., 35, 61, 130 Briicke, E., 176, 202 Bruun. Anton Fr., 141, 163, 166, 160,
Buchmann, H., 293, 326, 362, 376 Biickmann. A., 274, 276, 278, 289,292,
f.294, 295, 345, 368; 369, 376 Bull, H. O., 263, 318, 355, 371. 376 Bullough, W. S., 356, 376 Burd, A. C., 304, 363, 378, 377 Burkenroad, M. D., 22, 130 Burne, R. H.. 316, 376
163,166
C ( ‘ ~ l i n , A. It.. 28, 108, 110, 1 1 1 , 130 ~‘n~rbllt~rl l t~c~i~~, S.. 163, 167 (hlif. Coi)p. Ococmic Fish. Invost., 107,
Culluinatrcl. Oilotto. 161, 162, 166, 167 C!rrnnKurcbt I I R I I I . P., 303, ,376 (‘iwlido, I). I s . , 188, 209 Currikor, M. it., 71, 130 Chanloy, 1’. E.. 3, 22, 30, 34, 41, 46, 47,
62, 64, 09, 77, 79, 81, 130, 131, 134 Chunnon, H. J., 301, 302, 376 Chepurnov, A. V., 277, 278, 385
202
Cheatnoy, V. N.. 343, 376 Cheatnut, A. F., 81, 131 Chiarini, P., 179, 202 Christ-, J. Y., 296, 306, 379 Churchill, E. P.. Jr., 22, 132 Clark, F. N., 348, 376 Clarke, C . L., 191. 192, 193, 194, 202,
Clarke, R., 141, 166 Cleaver, F. C.. 273, 376 Cloud, P. E., 220, 259 Coates, C. W.. 360. 392 Coe, W. R., 104, 131 Colo, I€. A., 30, 36, 61, 02, 72, 103, 131 Colton, J. B., Jr., 290, 291, 376 Conover, R. J., 194, 203
203
COEtellO, D. P., 26, 82, 90, 99, 113, 117, 119, 121, 127, 128, 132
Cowan, D.. f.294, 376 Craig. R. E., 273, 304, 331, 333, 305,
Crescitelli, F., 187, 188, 203 Crossland, C., 216, 217, 231, 264, 255 Cunningham, J. T., 287. 357, 376, 377 Cushing, D. H., 197, 203, 276,298,297,
304, 329, 331, 333, 346, 363, 365, 366, 377, 391
376, 386
D
Dahlgren, E. H., 368, 388 D’Anconu, U., 138, 150, 166 Dannavig, A., 270, 282, 977 I)mnovig, ( l , , 282, 677 l>art~idl, I f . .i. A., 187, 188, l!jO, 803
Ihuptiiii(,c~, J , A., 309, .?XI Ihvitl, (!. N., 194. 203 Ihvitlaon, M. 15., 26, 82, 90, 99, 113,
117, 119, 121. 127, 128, 131 UaviH, 11. C., 3, 14, 15, 17, 22, 26, 30,
32, 34, 36, 39. 46, 46, 47, 48, 62, 63, 67, 68, 59, 01, 62, 64, 69, 71, 72, 73, 76, 77, 79, 80, 82, 85, 105, 107,131,152, 134
I h , N., f.204, 677
Daweon, A. B., 3L6, 377 Day, L., 368, 377 Deelder, C. L., 138, 144, 166, 196, 203 Degtyerev, A. A., 342, 344, 305, 390 Delaman, H. C., 365, 377

AUTHOR INDEX 397
Dementeva, T. F., 277, 288, 327 Denton, E. J., 186, 187, 188, 190, 191,
192, 193, 202, 203 Devold, F., 333, 336, 339, 342, 362,
363, 366, 367, 377 Dickson, W., 289, 322. 324, 374 Dietrich, G., 333, 334, 377 Dietrich, It., 301, 307, 320, 375 Dodt, E., 174, 203 Doty, M. S.. 224, 230, 256 Dow, R. L., 318, 323, 324;377 Drew, G. A., 98, 132 Drageaund, 0.. 197, 203, 330, 342, 346,
Droop, M. R., 236, 239, 241, 260, 256 Dunkan, R. E., 197, 203 Durham, J. Wyatt, 216, 256 Duyven6 de Wit, J. J.. 375
349, 361, 366, 377, 378
E Edmondaon, C. H., 218, 256 Ege. V., 139, 140, 166, 167, 168, 166,
Eggers, A., 26, 82, 90, 99, 113, 117, 119,
Ehrenbaum, E., 146,167, 366, 378 Elnnan, S., 143, 167 Elmhirst, R., 294, f.294, 378 Ellis, G. H., 196, 203 El Saby, M. K., 301, 302, 376 Exner, S., 182, 203 Emerson, W. K., 216, 257 Emery, K. 0.. 220, 263, 264, 256, 259 Endean, R., 231, 240, 259 Engle, J. B.. 17, 39, 47, 66, 64, 96, 134 EngatrBm, K., 184, 203 Evans, H. E., 313, 378 Ewart, J. C., 272, 366, 378
167
121, 127, 128, 131
F Fahy, W. E., 81, 131 Fairbridge, R. W., 229, 231, 256 Farrin, A. E., 294, f.294, 296. 378 Farris, D. A., 278, 281, 282, f.294, 306,
Field, G. A., 90, 132 Fish. M. P., 166, 167 Fleming, R. L., 142, 169
309, 378
Fluchter, J., 274, 378 Fontaine, M., 161, 162, 166, 167 Ford, E., 271, 292, 378 Fox, M. H.. 26, 82.90.99, 113, 117, 119,
121, 127, 128, 131 Frmett, D. M., 273, 376 Franz, V., 176, 176, 203 Fraeer, J. H., 336, 378 Freudenthal, H. D., 234, 236, 256 Freytag, G., 319, 331, 378 Frisch, K. von, 173, 203 Frost, W., 161, 167 Fullerton. J. H., 98, 99, 132 Fulton, T. W., 347, 378 Furuya, K., 330, 386
G Gabriel, M. L., 369, 378 Galkina. L. A., 268, 271, 272, 378 Galkov, A. A., 327, 331, 341, 363, 378 Galtsoff, P. S., 18, 23, 24, 132 Ganaroe, A. E., 66,134 Gandolfi-Hornyold. A., 148, 149, 167 Garten, s., 178, 180, 204 George, C. J., 86, 136 Gerasimov, V. V., f.294, 296, 306, 312,
313, 346. 382 Gerlach, S. A., 224, 256 Gibbs, E. D., 349, 361, 387 Gilbert, P. W., 176, 177, 178, 204 Gilis, C., 304, 378 Gha, I. I., 296, 330, 346, 379 Glover, R. S.. 296, 304, 329, 333, 336,
Gohar, H. A. F., 238, 240, 249, 256 Goodall, R. G., 196, 204 Goreau, N. I., 237, 239, 241, 243, 248,
260, 262, 256, 257 Goreau,T. F.. 226, 227, 228, 229, 232,
237, 238, 239, 241, 242, 243, 244,
340, 379
246, 247, 248, 250, 262,256, 257 Gosline, W. A., 146,167 Graham, J. J., 296, 304, 311, 313, 379 Graham, T., 370, 379 Graham, M., 347, 362, 379 Grant, N.. 196, 197, 198, 205, f.294,
296, 296, 307, 322, 329, 330, 340, 360, 366, 379, 383

398 AUTHOR INDEX
Graasi, B., 153, 167 Grave, B. H., 127, 128, 132 Gray, I. E., 326, 379, 391 Groody, T., 366, 379 Groody, T. C., 294, f.294, 383 Guillard, R. R., 3, 34, 39,57, 69, 61, 64,
Gunter, G.. 295, 306, 379 Gustafson, G., 194, 204 Gutsell, J. S., 99, 132 Gwyn, A. M., 276, 379
77, 131, 132
H Hall, F. G., 326, 379 Hamana, H., 268, 379 Hamikon, E. L., 219, 257 Hand, C., 237, 262, 257, 258 Hanks, J. E., 56, 234 Hansen, S., 279, 282, 377 Hansen, V. Kr., 239, 259 Hanyu, I., 184, 204 Harder, W., 274, 275, 276, 278, 286,
289, 292, 293, f.294, 296, 297, 324, 345, 376, 379
Hardy, A. C., 276, 296, 371, 379 Harry, R. R., 224, 257 Harvey, E. N., 193, 204 Haaler, A. D., 144, 167, 199, 200, 204,
341, 379 Hatanaka, M., 26,66.80, 103,106,107,
121, 123, 127, 128, 129, 132, 303, 391
Haven, D., 81,132 Hedgpeth, J. W., 263, 257 Heerd, E., 174, 203 Heiberg, Barge., 153, 166 Heincke, F., 366, 379 Hela, I., 270, 333, 339, 361, 365, 380 Heldt, H., 148, 149, 160, 167 Heldt, H., (Mme.), 149, 160, 167 Helfrich, P., 261, 257 Hempel, G., 267, 271, 274, 275, 277,
278, 279, 280, 281, 282, 289, 292, 293, f.294, 296, 346, 368, 369, 370,
Henley, C., 26,82,90,99, 113, 117, 119,
Hennacy, R. E., 129, 133 Hentschel, E., 275, 380
374,376, 380
121, 127, 128, 131
Hertlein, L. G., 215, 257 Hickling, C. F., 300, 301, 304, 307, 347,
Hida, T. S., 280, 380 Hildebrand, S. F., 147, 167, 196, 205 Hirano, Y., 26, 133, 270, 280, 297, 339,
Hjort, J.. 366, 380 Hoar, W. S., 174, 182, 196, 197, 201,
204, 269, 270, 271, 273, 279, 304, 311, 380, 384
380
385
Hodgkin, N. M., 231,257 Hodgson, W. C., 262,303,347,380,388 Hoffmeister, J. E., 220, 221, 222, 223,
259 Holliday, F. ct. T., 194, 206, 265, 271,
272, 274, 283, 284, 286, 293, 294, f.294, 295, 296, 299, 300, 303, 310, 311, 312, 313, 314, 316, 318, 326, 356, 368, 360, 362, 364, 365, 374, 380, 386
Holstvoogd, C., 274, 380 Holt, S. J., 306, 306, 373 Honey, K. A., 290, 291, 376 Hopkina, A. E., 104, 105, 132 Hori, J., 104, 106, 132 Horrall, R. M., 199, 204, 341, 379 Horridge, G. A., 232, 238, 257 Houraton, A. S., f.294, 295,,339, 381 Hovaeee, R., 233, 257 Hubbard, C. J., 192, 203 Hubbs, C. L., 368, 381 Hughes, R. B., 309, 381 Hunter, A., 309, 381 Huntmmn, A. G., f.294, 296, 296, 298,
299. 300, 373
I Idler, D. R., 308, 384 Iles, T. D., 362, 381 Imai, T., 26, 41, 66, 80, 81, 103, 106,
106, 107, 121, 123, 127, 128, 129, 132
Imamura, G., 196, 198, 204 Imbrie, J., 226, 229, 230, 258 Ingeraon, E., 214, 219, 220, 258 Ies, ct., 198, 204 Ito, S., 270, 381 Ivlev, V. S., 282, 286, 303, 381

AUTHOR INDEX 399
J
Jscobsen, J. P., 160,167 Jscobshegen, E., 297, 381 Jekobeen. F., 309, 388 Jekobeson. J., 296, 328, 329, 333, 336,
Jenuechke, H., 182, 203 Jeen, Y., 270, 381 Jeffem, Anne M., f.294, 296, 296. 298,
299, 300. 373 Jeffem, G. W., f.294,296,296,298,299,
300,373 Jem, G., 347, 381 Jensen, A'. G. C., 366, 381 Jerlov, N. G., 189, 204 Jargemem, C. B., 26, 63, 84, 88, 89. 90,
382
93,94,96,96,97,99,108,109,119. 123, 124, 126, 127,132
366, 382 Johensen, A. C.. 163, 167. 289, 290,
John, C. C., 293, 382 Johnson, M. W., 142,169, 261,257 Johneon, W. H., f.294, 296, 296, 298,
Jonee,F. R. Harden, 316,320,381,322,
Jones, J. W., 138, 141, 144, 146, 160,
299,300, 343, 373, 381
329. 330, 381, 382
267
K Kamshylov, M. M., f.294,296,306,312,
313, 346, 382 K&ndler, R., 123, 233 -pa, E. M., 189, 202 Kenda, K., 196, 204 Kenoh, Y., 206, 267,382, 393 Kawaguti, S., 218, 232, 233, 236. 241,
Kawamoto, N. Y., 198, 205 Keenleyeide, M. H. A., 196, 204 Kelley. A. M., 272, 382 Kennedy, D., 191, 205 Keys, A., 150, 267. 311, 382 Kirrodiits, T., 26, 233 Kirchhoff, H., 295, 382 Kiumurs. K., f.294. 382 Kniiht, 116.. 36, 61. 230 Kmh, H., 161. 152, 167
246, 249, 257
Koch, H. J., 161,161 Koh, A. J., 261, 267 Koike, A., 196, 204 Komarovskg. B., 296,382 Konkol, H., 342. 363, 386 Korringe. P., 47, 101, 233 Kotthsue, A., 282, 382 Kovaleve, N. D., 316, 387 Kreftenmecber, W., 191, 206 W t , G., 343, 346, 349, 360, 363, 382,
Rrietjoneeon, H., 198,205 Krogh, A., 160, 162,167 Krumhob, L. A., 276, 289, 296, 375 I(ryzhenovsky, 8.0.. 271, 283, 382 Kiihne, W., 177, 205 Kuenen, P. H., 264,257 Kupffer, C., 268, 382 Kurete, H., 276, 276, 277, 279, 283,
KueaLels, D., 106, 232
389
287, 382
L Ledd, H. S., 210, 213, 214, 219, 220,
221, 222, 223. 263, 264, 256. 257, 258. 259
Laevwtu, T., 270, 333, 339, 361, 366, 380
Wler, K. E., 296, 316, 382 h e , C. E., 129, 133 Leeker, R., 266, 269, 272, 276, 281, 286,
Leufer, M., 191, 206 Lea, E., 273, 382 Lebour, M. V., 96, 97, 108, 119, 233,
Le Gall, J., 367, 383 Legad, J. E. H., 290, 391 Lindsey, C. E., 36,133 Liahev, M. N., 277, 363, 383, 384 Lhivnenko, L. N., 277, 386 Ljubickaja, A. I., 288. 383 IdFt~i, B., 367, 384 Longloy, W. H., 196, 205 Im~enolT, V. L., 3, 7, 14, 16, 17, 18, 21,
26, 30, 34, 36, 37, 39, 40,46,47,48, 52.63. 64,66, 66.62, 63.64, 69, 71, 72.73. 74.16, 77.81, 82,85,96,98, 101. 121, 231, 133, 234
286, 287, f.294, 301, 382
274, 276, 283, 287, 292, 382, 383

400 AUTHOR IMDEX
Loukaahkin, A. S., 196, 197, 198, 205, 294, f.294, 296, 296. 307, 322, 329, 330, 340, 350, 366, 379. 383
Love, R. M., 307, 320, 322, 383 LovBn, S., 108, 134 Lovern, J. 9.. 301, 302, 383 Lovetskaya, A. A., 346, 360, 383 Lowenstein, O., 316, 318, 319, 383 Luces, C. E., 47, 64, 234, 342, 363, 383 Luhmann, M., 300, 301. 383 Lythgoe, R. J.. 178, 202
M
MacBride, E. W., 26, 134 McBrido; J. R., 308, 384 McDonald. J., 193, 196, 205 MacGregor, J. S., 297, 306, 328, 360,
McHugh, J. L., 369, 384 MacKinnon, D., 197, 202 McLaughlin, J. J. A., 233, 236, 237,
Macleod, R. A., 308, 384 McMynn, R. G., 269,270,271,273,279,
MacNairn, N. A., f.294, 296, 296, 298,
MwNichol, E. F., Jr.. 190, 191, 20.5:
MacPherson, G., 328, 392 Miiiir, A., 144, 146, 268 Maim, 11. N., 274, 3 l H , 321, 383 Mfiirn, I ) . It ' . , 3 ( ) H , J N J , J H f / , ,YO/) Mntitviift4, I!. I+.. 320, 5'17, 3fi3, M d Mnrr, . I . ( !., 2W. 281, 21)7, 387, 309, 3RJ Mtuiton. M. M., 246, 2bR M a m h i . $1. It., 230, 259 Mtrruhall, A. J . , 367, 386 Marshall. N. B., 320, 321, 382 Marshall, S. M., 277, 283, 287, 384 Martin, C., 76, 132 Marty, Ju. Ju., 281. 283, 360, 361, 384 Meeeman, W. H., 310, 384 Matsui, I., 142, 167 M8xfield, G. H., 276, 280, 384 Mazzarelli, G., 152, 168 Meadows, P. S., 316,325,330,353,354,
Medcof, J. C., 366, 384
383
239, 249, 258, 260
384
299, 300, 373
206, 207
374
Meyer, H. A., 272, 276, 278. 279, 384 Milkman, R. D., 191, 205 Miller, D. J., 270, 274, 384 Miller, R. R., 295, 316. 382 . Miller, W. S., 3, 39, 46, 48, 62. 69, 71,
72,86, 134 Mitarai, G., 191, 205 Mitchell, J. M., 306, 384 Miyazaki, I., 96, 134 Mohr, H., 361, 362, 363, 364, 384, 385 Moorhouse, F. W., 231, 268 Morgan, T. H., 268, 386 Morris, R. W., 276, 278, 282, 288, 385 Morrison. J. P. E., 224, 230, 256, 268 Mortonsen, T., 86, 136 Moses, R. L., 360, 392 Motaie, R., 191, 205 Motoda, S., 224,230,246,258,270,280,
Moulton, J. M., 318, 330, 331, 385 Munk, W. H., 220, 221, 222, 223, 264,
Mmz, F. W., 186, 187. 189, 190, 205 Murphy, G. I., 270, 277, 280, 281, 290,
Muscattine, L., 237, 238, 258 MuhiO, R., f.294, 295, 385
291, 339, 385
258
339, 385
N Nagmaki, F., 360, 385 Naah, J., 311, 386 NI~IUIIOV, V. M., 3OH, :Ilgfi, MI, 362, $63,
2Xh NOIMOII, '1'. t !., 4 I , 44, 1*1,5 Ntrwoll, N. l)., 214, 224, 226, 220, 229,
Nowth, D. R., 172, 194, 205 Nicholls, A. G., 236. 239, 240, 246, 260,
277, 283, 287, 384 Nicol, J. A. C., 177, 178, 179, 193, 194,
203, 205, 237, 258 Nicoll, J., 162, 168 Nigrelli, R. F., 306, 392 Nikitin, V. N., 2 ,135 Nikitinskaya, I. V., 277, 278, 281. 385 Nikolsky, G. W., 262,270,277,278,385 Nikonorov, I. V., 349, 350, 361, 364,
373, 385, 386 Nishimura, S., 265. 392
230, 264, 258

AUTHOR INDEX 40 1
Nomejko, C. A., 17, 40, 45, 48, 62, 69,
Nomura, M., 342, 344, 386 Northcote, T. G., 193, 196. 205 Nybelin, O., 163, 168
71, 72, 86, 134
0
O’Connell, C.P., 316,317,318,321,386 Odhner, N. H., 99, 135 mum, E. P., 213, 261, 258 Odum, H. T., 213, 261, 258 Ogura, M., 196, 204 Okada, H., 41, 106, 132 O b i , N.. 341, 388 Okonski, S., 342, 363, 386 Om. A. P., 277, 283, 287, 384 Orton, J. H., 101, 108, 135 Oshiba, C., 341, 388 Otterstram, C. V., 142, 147, 168 Outram, D. N., 269, 270, f.294, 366,386 Owatari, A.. 330. 386
P
Parke, M. W., 35, 61, 130 Parker, G. H., 173, 206
P h h , B. B., 273, 294, f.294, 304, 316, 326, 327, 328. 330, 346, 349, 360, 362, 363, 364, 366, 374, 386, 392
Pany, Gwyneth, 194, 206, 311, 326, 380, 386
Patejev, A. Kh., 360, 386
Peeraon, J. C.. 280, 386 Pehneer, P., 48, 135 Perceva, T. A., 269, 270, 283, 386 Pergens, C., 179, 206 Phillips, J. B., 313, 327, 328, 339, 386 Pinto, J. dos Santos, 266,203,366,362,
I’iiwiiir, M. I t . . 11)1, 102, I U J , I’irio, A.. 177, 2UfJ l’()ldor, J. J. W., 367, 360, 387 Polonskij, A. LL, 333, 368, 387 Porter. H. J.. 81. 231 Postume. K. H., 343, 344, 387 Powers. E. B., f.294, 313. 340, 346,389
PaR, A. E., 294, 386
P8vshtiks, E. A., 333, 339, 386
364, 372. 380, 587
Pratt, E. M., 239, 258 Press, J. S.. 179, 202 Priestley, R., 273. 331, 366, 376, 386 Prince, E., 268, 387 Pringsheim, E. G., 233, 259 Protaaov, V. P., 316, 387 Protaaov, V., 316, 360, 374 Prytherch, H. F., 2, 36, 72, 235 Pumphrey, R. J., 191,206
0 Quayle, D. B., 72, 135
R Redakov, D. V., 322, 326, 326, 327,
Itadovich. J., 296, 313, 327, 328, 339,
Remanujam, 8. a. M., 276, 387 Rennak, L. A., 276, 277, 383, 387 Ranson, G., 230, 269 Raamuesen, C. J., 148, 168 Rasquin, P., 173, 174, 176, 202, 206 €hiss, T. S., 266, 387 Rees, C. B., 89, 90. 96, 120, 123, 135 Retzius, G., 264, 387 Richardson, I. D., 343, 344, 346, 349,
Ridowocid. W. G., 318, 387 Rigby, J. K., 226, 258 Rochon-Duvigneaud, A., 171, 176, 206 Rogalla, E. H., 333, 387 Ronold, 0. A., 309, 388 Rosen, M. W., 324, 388 ROBR, D. M., 172, 194. 205 Rounuefdl, C. A., 273, 366, 368, 388 RoiiHhdy, H. M., 239, 259 Rurrlyantwov, A. I., 318, 388 I t i r r r n w l m t c i , N.. 273. 320. 342, 346, 366.
Itusliton. W. A. H., 174, 187, 206 Rus~oll. E. S., 333, 388 Ituecrell, b’. S., 289, 290, 388 Rueeell. Martin, 214, 219, 220, 258 Rydor, J. A., 36, 136, 270, 388 Ryzhenko, M., 342, 346, 349, 361. 388
328, 329, 330, 363, 384, 387, 393
349. 361, 386, 387
363, 387
307, JCVY

402 AUTHOR INDEX
S
Sahrhage, D., 333, 334, 362. 377, 387,
Sakei, S., 26, 41, 66, 80, 81, 103, 106,
Sakumoto, D., 241, 257 Sara, R., 197, 198, 206 Sargent, M. C., 220, 221, 222, 223, 261,
264, 258, 259 Ssertki, T., 341, 388 Sato, R., 26, 66, 80, 103, 106, 107, 121,
Sato, T., 7, 135 Saunders, J. W., 196, 206 Savage, R. E., 296, 303, 347, 388 Saville, A., 273, 366, 386 Scettergvod. L. W., 262,280, 290,294,
f.294, 296, 307, 378, 388, 389, 391 Scheoh, H., 276, 278, 279, 282, 389 Schiifer, W., f.294, 306, 316, 327, 328,
Scheeffer, M. B., 366, 389 Schiiperclaus, W., 163, 169 Schiirfe, J., 323,361,362,363,364,389 Scharrer, E.. 173, 206 Scheming, L., 274, 318, 321, 383 Schlaifer, A., 193, 196, 206 Schmidt, J., 139, 141, 147, 164. 166,
166, 167, 168, 169, 160, 162. 163, 164, 105,168
Schnakenbeck, W., 162, 163, 269, 274, 389
Schott, G., 146,168 Schroeder, W. C., 143, 147, 166, 167 Schubert, K., 333, 334, 344, 349, 377,
Schiiler, F., 343, 346, 349, 360, 363,
Schultz, L. P., 230, 262, 259 Schutz, F., 316, 389 Schwaesmm, H. 0.. 199, 200, 201,
Hnrrlr, A . (1.. 91 (I, $69 Hdicw. 11. J I . , 176, PO6
388
106, 107, 121, 123, 128, 129, 232
123, 127, 128. 129, 232
330, 389
382, 389
382, 389
goe
Howlbll. ti., 177, %U6 SIItbt,U1koVHky, M. I., 277, 278, 385 Shw, E.. 183, 106, 202, 206, 328, 389 Shew, T. I., 187, 203 Shelbourne, J. E., 286, 389
Shelford, V. E., f.294,313, 340,344 389 Shewan, J. M., 309, 389 Shubnikov, D. A., 360,389 Sigerfoos, C. P.. 128, 235 Silliman, R. P., 289, 290, 389 Simpon, D. M., 177, 206 Sindermenn, C. J., 294, f.294, 296.307,
Skoglund, 0.. f.294, 318, 390 Smith, F. W. W.. 224, 259 Smith, K. A., 361, 364, 390 Smith, P. B., 3, 39, 46, 48, 62, 69, 71,
Smith, M. W., 196, 206 Snodgraes, J. M.. 189, 202 Soleim, P. A., 271, 276, 279, 280, 282,
Soiovyev, B. S., 322,326,328,329,330,
Southward, A. J., 297, 390 Spooner, Q. M., 291, 294, 390 Sprague, L. M., 308, 390 Springer, S.. 362, 390 Squires, D. F., 216, 224, 259 Stafford, J., 83,86,96,96,104,126,135 Stauber, L. A., 17, 135 S b l e , J. H., 333, 336, 340, 390 Stephens, G. C., 262, 259 Stephenson, A., 229, 259 Stephenson. T. A.. 224, 229, 259 Stephenson, H. K., 214, 219, 220, 258 Stephenson, W., 216, 231, 240, 259 Steven, D. M., 190, 206 Stevenson, J. C., 283, 290, 292, 390 Stevenson, W. R., 179, 202 Stewart, K. W., 184, 206 Stirling, W., 298, 380 Stoddart, D. R., 224, 229, 231, 259 Stringham, E., 323, 324, 390 Stroganov, N. 0., 267, 390 Buehiro, Y., 313. 3Y0 Hullivm, C. M., 83, 86, 88, 89, 90, 05,
DA. 98, 108, 113, 115, 116, 117, 120, 123, 124, 196, 127, 129,135
308, 375, 378, 383, 389, 390
72, 86,134
291, 390
342, 344, 345, 366, 387, 390
Xvwtichin, U., 190, 191, 205, 206 Svaotiohin, J., 190, 200 SvlLrdeon, a., 148, 169 Svedrup, H. V.. 142,169 Svetovidov, A. N., 262, 390 Swallow, J. C., 142, 143, 269

AUTHOR INDEX 403
T Tagatz, M. E.. 310, 390 Takahaehi, M., 303, 391 Temura, T., 181, 206 Thing, A. V., 164, 166, 168, 169, 161,
Teneley, K., 178, 202 Teichert, C., 231, 256 Temple, R. F., 290, 291, 376, 391 Terry, R. J., 173, 207 Teeter, A. L., 197, 207, 342, 348, 369,
Theilacker, GI. H., 272, 285, 286, 287.
Thomeon, D. A., 280, 380 Thoreon, G., 72,135 Tibbo, S. N., 290, f.294, 328, 391 Tierney, J . Q., 129. 133 Tikhinov, D. P., 349, 391 Tommere, F. D., 63, 64,134 Toom, M. M., 268, 270. 272, 273, 274,
Townaend, R. C., 214, 219, 220, 258 Tracey, J. I., 213, 220, 221, 222, 223,
224, 263, 264, 256, 258, 259 Tucker, D. W., 137, 138, 141, 142, 143,
145, 146, 147, 148, 149, 160, 161, 162, 163, 164, 166, 166, 158, 169, 160. 162, 163,169
Turner, H. J., 86, 136 Turpae~va, E. P., 2, 135
169
391
f.294, 301, 382
278, 391
U
Uchida, K., 166,168 Utelee, R., 10, 58, 64, 131, 136 UIrich, B., 327,391 Umbgrove, J. H. F., 214,220,221,230,
248. 259
V VBihYlb, L., 141,169 Veldez, V.. 331, 346. 391 Vaughan, T. W., 211, 214, 219, 269 Verheijen. F. J., 198, 207, 287, 294,
f.294. 296, 314, 316, 31% 319. 321, 3.70, 34V. 360, 3U2. 391
Vernberg, F. G., 326, 391 Vernberg, F. J.. 326, 391 Verrier, M.-L., 176, 207 Vilter, V., 314, 391 Vincent, R. E., f.294, 391 Vishniac, H. 9.. 75,136 Vladykov, V. D., 147, 149, 169 Voisin, P., 102,136 Volodin, V. M., 267, 268, 286, 287, 391 Von Ant, W. S., 250, 261.260 Vmman, A. M., 308, 390
W Wagner, H. G., 191, 205, 207 Weld, G., 146, 169, 187, 188, 207 Waldmann, J., 276, 392 Walls, G. L., 176, 184, 207 Walne. P. R., 30, 67, 66, 66, 77, 101,
Wering, H,, 196,207 Warren, F. J., 185, 187, 190, 192, 203 WBtrtnebe, 8.. 341, 388 Waterman, T. H., 144, 169 Weteon, J. E., f.294, 392 Waugh, 0. D., 7, 77, 136 Weisman, A. L., 360, 392 Wells, F. R., 274. 392 Wells, J. W., 210, 211, 214, 216, 216,
217, 219. 220, 221, 224, 231, 259, 260
103,136
Wells, W. F., 2, 35, 99, 138 Welsh. J. H., 181, 207 Werner, B., 93, 120, 136 Weatman, J. R., 306, 392 Weeton, W. H., 76,132 Whiteman, A. J., 226, 258 Wiene, H. J., 212. 219, 224, 260 Wilcocka, J. C., 196, 207 Willbmon, D. I., 276, 280, 375 Williamson, H. C., 280, f.294, 296, 290,
297, 316, 356, 392 Willismaon, M. H., 336, 392 Willmor, E. N., 311, 382 Wileon, I). P., 45, 72, 86, 136 WiMklW, F.. 36, 138 Wieby. W. J., 199, 801, 341, 379 Witterttwrg, 1%. A., 195, 207

404 AUTHOR INDEX
Wittenberg, J. S., 196, 207 Woelke, C. E., 36, 133, 136 Wohlfahrt, T. A., 318, 319, 392 Wolbereht, M. L., 191, 205, 207 Wood, H., 328, 383, 392 Wood, P. C., 7, 77, 136 Wood, R. J., 300, 392 Woodhead, A. D., 291, 392 Woodhead, P. M. J., 193,207, 291, 392 Worthington, L. V., 143, 169 Wynne-EdwardS, V. C., 304, 341, 346,
392
Y YaXIUbgBWe, M., 286, 392 Yememoto, K., 286, 268, 392 Yememoto, T. S., 286, 267, 392, 393 Yanegimschi, R., 287, 393
Yonge, C. M., 101, 136, 211, 212, 217, 218, 220, 231. 234, 236, 237, 239, 240, 241, 244, 246. 248, 249, 264, 260
Yonge, M. J., 240, 260 Yoehide, H., 2, 136 Yoshide, T., 41, 105, 112, 126, 132 Young, J. Z., 172, 173, 176, 207, 208 Yudaev, H. A., 309, 393 Yudanov, K. I., 363,384 Yuki. R., 80, 106, 107. 128. 129, 132
Z Zehl, P. A., 233,236,237,239,249,268,
Zeitsev, V. P., 322, 326, 393 Zijlstra, J. J., 388, 393 Z m r , 5. G., 296, 329, 330, 343, 346,
346, 360, 393
260

Subject Index
A
Accommodation in fish eye, 182-186 Activity of clupeids, 326, 328 Alimentary system of clupeids, 297-300 AnguiUa anguiUa
deacription of, 138, 139-141, 147,
leptocephali of, 16k-166, 169, 100 migration of, 141-146
diettibution of, 142 leptocephali of, 164-166, 180, 182,
183 number of vertebrae in, 139, 140,
141,156-161,183 epawrhg areas of, 180-182
Anguillu austdiu, 139,167 AnguiUcr j a p k a , 139,140,1b8 Anguik roetmta
description of, 139-141, 148 effect of tampereture on, 143-144,
leptooephali of, 164-160,169-160 migration of, 141, 142
Arm tranevcssa, 3, 86,87
148,168
AnguiUcr genus, aee a h Eels
156-161
~nomia *lez, 3,95-9a
Atoh, a b o cod re43f8, 21tk214, 219-224,250-255
B Bacteria, effect on lawee of bivalvea,
Bikini atoll, 214, 21S224, 260, 261 Bivalve molluaka
77-78
out of &won pwning, 1C28 reering, 1-136 sse abo named 8peGie8, Egge, Lama4
Brain of olupeidn, 3 13-31 4 Breeding
of freeh-water eels, 137-189 eelective, of bivalves, 80, 81
Buoyancy of clupeida, 320-322 406
c Calcification of corals, 238, 237, 241-
Catching migrating eele, 146,146 Catf%h, pineal region of, 173 Chemical conditions, aee also 0sm0-
245,246-248
regulotim. Salinity effect on bivalvea, 44-47 effect on clupeids, 271, 272, 312, 313
effect on zooxanthellae, 238 l a ~ a e , 287-289
Chemicals, effect in controlling dieeagee
Chondrichthyea, tapetum lucidum of,
Chorioidal gland of fishes, 194, 196 Chrysopins in Aeh eye, 188 Clams; dcc Arm tranaverea, Eneir
directus, Mactra eolidiesima, Merccnaria campchien&, Mer- cenaria me*cena*io, M y a arcnarirr, Pitar m m h w n a , Taw semi- ckcuu-
of bivalvea, 76-80
176-182
Clupeids activity, 326, 326 alimentary ayatem, 297-300 behaviour and phyaiology, 201-393 brain, 313, 314 buoyancy, 320-322 characteristics, 264, 367-370 eggs, uee Eggs, clu- equilibrium, 320-322 establishment. 294 fat Atorage, 3 W 3 0 2 feeding, 294-297, 300-307, 346 gametee. 284-208 gonclde, maturation of, 366-304
hearing, 315320 labyrinth, 316-318, 322 larvae, we hm, clupirlr light, mponse to, 195, 348-351 ‘‘ light Aching ”, 198
mwth , 302-306

406 SUBJECT INDEX
Clupeids-wntind metabolism, 300-302 metamorphkm, 292-294 migration. 332-342 migration. vertical, 320-322, 342-
moon, effect of, 347,348 mortality, 306-307 nete, reaction to, 361-366 olfaction, 316 osmo-regulation, 309-312 oxygen uptake, 313 reviews, 262-264 rearing, 278-280 salinity tolerance, 309-312 shoaling, 328-332 spawning. 322-326 tissues, 307-309 vieion, 314, 315, 329, 362, 363
Colour vision in fishes, 186191 Conger eel, 162-164, 166, 178, 188, 192 Coral reefs and corals
Atlantic and Pacific, comparison of,
atolls, 210, 211, 212, 213, 214,
biology, 209-200 calcification, 236, 237, 241-245, 240,
247, 248 distribution, 2 14-2 17
erosion, 229-232, 260-266 food, 238-245, 252-2114 fringing barrier formation, 223-229 growth, 241, 245-255 literature on, general, 210-214 movement of, 260-266 physiology, 232 planulation, 217-219 productivity, 260-266 and zooxanthek, 232245,246-248
Crassostrea genus, 61,80,81 ~radsosh.ea: ha .6 , 3,80-82,106-107 Ctlreeosh.eo O;rghiCrr, 2, 3, 16, 16, 20,
22, 28, 30, 37, 39, 40-46, 60-64, 60-62, 65, 08, 71, 72, 73, 76, 78, SO, 81, 82, 83, 101, 103, 104, 106, 107
Crowding of larvae of bivalves, 88-71 Cycbnea, effect on reefs, 231-232
347
212,224429,231,232
219424,250-255
ecology, 219-224
D
Demineralization of female eele, 161,
Diseeeea, see also Mortality 162
bivalves, 76-80 clupeida, 307 eeh, 163
Distribution of corals, 214-217
E Eele, see also AnpiUa
breeding of fresh-water, 137-169 characterietica, 139-141, 147, 148 demineralization of female, 161, 162 leptocephali, 154-156,169-161,162-
migration 166
capture during, 146-146 direction of, 142-146 seaward, 147-153,198
mortality, 162, 163 retinomotor changes, 178 salinity, effect of, 144,149-160 etagee, 148,149 temperature, effect of, 142-144, 149 vertebrae, 139, 140, 141, 156-161,
visual pigmenta, 188
bivalvcss
163
Egf?fJ
characterietica, 82128 cultivation methods, 36-88 development, 28-29, 30-36, 8 2
fertilization, 36
numbera, 22-25, 118 eelinity, effect of, 6 2 6 3
128
herdin-, 41-47
etripping, 20, 21 temperature, effect Of, 47-62 turbidity, effect of, 63-55
characteristior, 266,266 development, 268-274 fertilization, 267, 268 mortality, 273
clupeida

DUIWEWL 407
Egge-conts 'nued clupeida-mnts 'nued
osmo-regulation, 260, 271, 272 salinity tolerance, 271 temperature, effect of, 269-271
Eleemobranche hypermetropy, 182,183,184 pupillmy movement, 176 t8petum lucidum, 177, 178
Eniwetok atoll, 214, 219-221 En& directus, 3, 31, 117-120 Equilibrium of clupeida. 320-332 Equipment for rearing of bivalvee,
C14,36-38,66-66, 67-68 Eroaion of mefa, 229-232 Eetablishment of clupeids, 294 Extre-ocular reception in fishee, 172-
Eye of fish 174
chorioidal gland, 194, 196 n o d functioning, 183-185, 314,
photoeeneitivity. 19 1-194 regulation of light, 176-182 review on, 171 spectral eenaitivity, 186191 visual pigments, 186-191
316
Visual threeholde. 191-194
F
Fat efor8ge in clupeide, 300-302 Fecundity
bivalvee, 22-26 clupeida, 264, 360-362
bivalve larvae, 39-41, 44, 61, 62,
clupeida, 294, 29S297, 300-307, 346
C o d , 236-245,252254
bivalvee, 36 clupeida, 267, 268
Feeding
55-69
larvae, 276-277
Fertilization
Pinh UM Iliualveo, CAondri&hgw,
Fungun infeotiotm in bivdvee, 76-71
~ h ~ i d s . h b , E h b n m a l b . Tmhta
G
Gsmetea bivalvee, 14 clupeide, 264-268
Gibraltar, Strait of, eel8 in. 145. 146 Gonads
bivalvee, 14 clupeids, 366-304
Crest Barrier Reef, 216,216,221,243 Expedition, 212, 217, 218, 220, 240
H
Herring, uee Clu# Hybridizetion of bivalvm. 80.81
J
Jingle Shell, 3, 95-98 Juvenile molluske. diseeeee of, 76-80
L
Labyrinth of clupeida, 316-318, 322 Laevkwdiniun, mortuni. 3, 107-109 la go or^, .260-254 k a e , wb alao Ewe, M&nnor&&,
spawning. Surfming bivalvee
chemical conditions, effect of 44-
colour. 68,83421 crowding, d e c t of, 69-71 cultivation methods, 3 6 8 8 development, 26-29,82-129 development, a b n o d . 80-85,
91.102-103,120 diseeses, 75-80,100,112,120, 129 food. 39-41, 44, S!$-68,117 hardiness, 41-47 eelinity, d e c t of, 52, 63
47
fempenrture, effect of, 39-44, 47-62.122
turbidity, &eot of, 63-66
ohtmbrietioe, 264 development of organ eyatems,
fad, 276-377
clupoido
274, 275

408 SUBJEOT INDEX
LeNae--cmtinued c l u p e i a i n u e d
growth, 277,278 light, response to, 291,292 migration, vertical, 289-291 mortality, 280-283 osmo-regulation, 283-286 oxygen uptake, 286, 287
salinity tolerance, 283-286 temperature, effect of, 287
eels, am Leptocephdi lamprey, response to light, 172
m h g , 278-280
Larval period of bivalves, 38-41, 44, 72 Lens of fish eye, 183,191 Leptocepheli of eela, f54-156,189,160,
Lcptocephalus species, 162164,166 Light
162-166
effect on clupeids, 288, 291, 292,
effect on corals, 237-239, 243-246.
effect on planulation, 218, 219 reception by fish, 171-208, 314, 316
296,343,344, 348-357
248-260
‘‘ Light 5hing ”, 198,361 LumineeCent mixnab, 193-194
M Mactra eolidieeima, 3, 26-29, 120-123 Maturation of g o d of clupidn,
Mercenaria cumpechicnsis, 3, 74, 76,
Mercenark genus, cross-breeding, 81 Mercenaria mrcenaricr, 2,16-17,21-22,
26, 26, 30, 32, 34, 39-40, 46,
69-78,81, 83-85, 109-110
367-370
356-364
81, 109-110
48-50. 63, 64, 61-66, 66-68,
Meristic characteristics of olupeids,
Metaboiiam in clupoids, 300-302 Metainorplroain
hivalvw, 2, 28. 29, 36, 39, 71-75,
112, 114, 117, 119, 122, 123, 124,129
clupeida, 292,294
83--86, 87, 94-98, 103, 108, 100,
Migration clupeida, 289-291, 332-342. 342-347
factors influencing, 199-200 eels, 141-163, 200
ModWlu~ &miem, 3,87-89 Modwlua diolw, 88.89 Mollusks, bivalve, em BivotW mdcudks Moon, effect on clupeicb, 347,348 Mortality
bivdvea, larvae of, 75-80,116 clupeida, 306-307
eggs, 273 larvae, 280-283
eels, 162, 163 Morton’s cockle, 3, 107-109 Muaeela, .me M o d ~ w ~ , MyitW
Mya arenark, 3,123-127 Mytilics eddk, 3, 20, 61, 89, 9 b B S
eddn
N
Navigation by eele. 142-146
0
OIfaction clupeide, 316 eels, 144 elaemobranchs, 177
clupeida, 300-31 2 eggm, 266, 271, 272 larvae. 283-286
Osmo-regulation
O~lrcCr ddW, 3, 7, 30, 41, 77, 101-104 effect of food on growth of lam- of,
61, 66, 66 Oetrea genue, 61 6shur luda, 3, 104-106 Oxygen uptake
clupeicb, 313 larvae, 286, 287
Optere, dcd Craaaostreu &M, C m - SOStVea Virginia, Ostrca ddia, oatreu lurida
P Pectcn iwadiataa, 3,76,@8-100 Petricolo pholad~ormw. 3.116117

SUBJEOT INDEX 409
Photoreception in fishes, 171-208,
Photawmitivity of fish, 191-194 Pineal complex, 173, 174, 176, 196 P h r morrhuarrcr. 3,21.112-115 Plankton, effect on migration of
clupeide, 333, 336-338 Planulee, eettlement of, 217, 219 Porphympeine in fieh eye, 186-188 Poet-metamorphic ategee of clupeide,
Pupillsry movement of 58h. 176
314-316
294-370
R Rearing
bivdvee, 81-129 clupeid larvae, 278-280
Receptor layer in fish eye, 182-186
Refreotion in fish eye, 182-186 Reguletion of light reeching seneory
Retinornotor chengm in teleoete 178-
Rhodopine in fish eye, 186-188.190 Rook borer, 3.115-117
A R e e f i , mml, wc coral reef8
nvfecee of fish, 174-182
182
S Salinity, effect on
bivalves, larvae of, 62, 63.112 clupeide, 303-304, 309-312
egge of, 271 larvae of, 283-286 meristic characteristics of, 367-370
eeb, 142,143, 144.149, 160 planulae, 218
migration, 200 mpom to light, 196, 197 retinomotor changes, 179, 180-181,
vim81 pigments, 187, 188 viaunl thresholds of larvae. 193
Salmon
182
Smiina, bee Clupeid.8 SC~IIOP, bey, 3,m, 98-100
Settlement of planulae, 217-219 Sharks
tapetum lucidum of, 176-178 visual pigments, 186
Shipworm, 3, 76, 127-129 Shoaling of clupeids, 196,294, 32C332,
Spwn quality, 24, 26 342
spawning, dce crbo Sh*+g&lg, swarming area8
clupeids, 332-342 eele, 160, 161, 162
bivalves, out-of9eeeon. 7, 14-26,
clupeide, 339, 367. 362-370 Spectral sensitivity of fish eye, 185-
82-127
- 191,198-199.il6 Sprete, 85f3 clupcids Stripping, 113, 119, 121 Sun, effect of, on migrating fishes,
199-200,341 Swarming, 101. 102, 103, 104-106, 127 Swimming by clupeids, 322-326 SymbMinium mhvad-m, 23C
Spqhbmnchuu speoies, 146,152-164, 235,236,239
160,166
clupeid, 367 c o d , 214-217
S ~ e t i c e
T Tapee semidcclrswta, 3.21,76.110-112 Tapeturn lucidum of chondrichthyea.
Teleosts 176-178
chorioidel gbnd, 194, 106 myopy, 182, 183, 184 photoreception and vision, 171-208 pupillery movement, 176 retinomotor chenges, 178, 182 visual pigments, 186
Temperature, effect on bivalvee
181~88. 39-44, 47-52,112,117 metamorphosis. 72.73 spawning, 14-26.82-127

410 SWJEClT INDEX
T e m p e r a t v i t a d olupeids, 312,313
e m , 269-271, 272 feeding, 296 Isrvae, 287 meristio cheracteristice, 367, 370 migration, 333, 334, 336, 339. 344
corals, 218 eels, 142, 143, 163, 166, 161
Teredo wvaliu, 3,76, 127-129 T h e s of clupeids, 307-309 TrenwPiesion by lene of 5 h eye, 191 Turbidity, effeot on lervee of bivalves,
6 3 6 5
Vertebrw, number in-cmtind eels, 139, 140-141, 156161, 163-104
Vision in fiches, 171-208, 314-316, 329 Vieuel pigment8 of fMwe’ eyee, 186-191 Visual thwholds of Aehes, 191-194,
196-198
W
Winds, effeat on -fa, 219, 220. 221, 223, 260,261
V Z
Vertebrae, number in clupeids, 367-370
Zooxanthellee, 232. 245, 246-248, 249-264