advanced review antimicrobial polymers as synthetic mimics
TRANSCRIPT

Advanced Review
Antimicrobial polymers assynthetic mimics of host-defensepeptidesKenichi Kuroda1∗ and Gregory A. Caputo2∗
Antibiotic-resistant bacteria ‘superbugs’ are an emerging threat to public healthdue to the decrease in effective antibiotics as well as the slowed pace ofdevelopment of new antibiotics to replace those that become ineffective. The needfor new antimicrobial agents is a well-documented issue relating to world health.Tremendous efforts have been given to developing compounds that not only showhigh efficacy, but also those that are less susceptible to resistance development inthe bacteria. However, the development of newer, stronger antibiotics which canovercome these acquired resistances is still a scientific challenge because a newmode of antimicrobial action is likely required. To that end, amphiphilic, cationicpolymers have emerged as a promising candidate for further development as anantimicrobial agent with decreased potential for resistance development. Thesepolymers are designed to mimic naturally occurring host-defense antimicrobialpeptides which act on bacterial cell walls or membranes. Antimicrobial-peptidemimetic polymers display antibacterial activity against a broad spectrum of bacteriaincluding drug-resistant strains and are less susceptible to resistance developmentin bacteria. These polymers also showed selective activity to bacteria overmammalian cells. Antimicrobial polymers provide a new molecular frameworkfor chemical modification and adaptation to tune their biological functions. Thepeptide-mimetic design of antimicrobial polymers will be versatile, generating anew generation of antibiotics toward implementation of polymers in biomedicalapplications. © 2012 Wiley Periodicals, Inc.
How to cite this article:WIREs Nanomed Nanobiotechnol 2013, 5:49–66. doi: 10.1002/wnan.1199
INTRODUCTION
The need for new antimicrobial agents is a well-documented issue relating to world health.1–6
The past 20 years have seen a dramatic increase inthe development of antibiotic resistance harbored
Conflict of interest: K. K. is a coinventor on a patent applicationfiled by the University of Pennsylvania covering ‘AntimicrobialCopolymers and Uses Thereof’. The patent application has beenlicensed to PolyMedix Inc. (Radnor, PA). PolyMedix did not playa role in the design and conduct of this study; in the collection,analysis, or interpretation of the data; or in the preparation, review,or approval of the article.∗Correspondence to: [email protected]; [email protected] of Biologic and Materials Sciences, University ofMichigan School of Dentistry, Ann Arbor, MI, USA2Department of Chemistry and Biochemistry, Rowan University,Glassboro, NJ, USA
by relatively common pathogenic bacterial species.These resistances can arise from both naturalevolution/selective pressure as well as from theimproper use and administration of antibiotics at theclinical level. Regardless of the origin of the resistance,these new ‘superbugs’ require the development ofnewer, stronger antibiotics which can overcome theseacquired resistances. Through the development ofthese new compounds, significant effort has been givento developing compounds that not only show highefficacy, but also those that are less susceptible toresistance development in the bacteria they target.Amphiphilic, cationic polymers have emerged asa promising candidate for further development asan antimicrobial agent with decreased potentialfor resistance development.7–10 These polymers aredesigned using naturally occurring host-defense
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 49

Advanced Review wires.wiley.com/nanomed
peptides (HDPs) or antimicrobial peptides (AMPs)as a ‘molecular inspiration’, building on the selectivityof these peptides imparted from their cationic,amphiphilic nature.11 However, polymers exhibitsignificant advantages over the peptide antimicrobialsin that polymers can be cost-effectively produced inmuch greater quantities, have greater compatibilitywith drug-delivery methodology, and provide aflexible framework for chemical modification andadaptation.
The chemical and structural diversity avail-able to polymer chemists allow for great vari-ability in the construction of cationic, amphiphilicmolecules designed to function as antimicrobials.This diversity yields a significant amount of requi-site investigation into mechanism of action, efficacyand selectivity profiles, resistance potential, as wellas structure–activity relationships. In general, thecationic, amphiphilic polymers exhibit selectivity tobacterial targets through favorable electrostatic inter-actions between the polymer and the highly negativelycharged bacterial cell surface.11 This electrostaticinteraction allows for broad-spectrum activity as mostbacterial surfaces maintain anionic properties. Oncebound, many polymers can significantly disrupt thebacterial membrane(s), presumably by inserting thehydrophobic moieties into the bilayer core.12–14 Whilethese are likely mechanisms, mounting evidence showsthat simple membrane disruption may be only onecomponent of a more complex series of effects bring-ing about bacterial cell death.15,16
BACKGROUND
Antimicrobial or antibiotic resistance is an emerg-ing threat due to the inherent loss of efficacy ofclinically prescribed antibiotics but also due to theslowed pace of development of new antibiotics toreplace those that become ineffective.1,2,5,6 The devel-opment of resistant organisms were first detected inhospital settings as nosocomial infections but havesince spread and are now routinely agents of commu-nity acquired infections. Clinically isolated resistantorganisms include bacteria, viruses, fungi, and para-sites. While resistance in any pathogenic organism isproblematic, the problem of bacterial resistance hastaken a primary focus of interest due to widely pub-licized examples [methicillin-resistant Staphylococcusaureus (MRSA), for example] as well as the numer-ous different strains which have shown resistancedevelopment in a clinical setting including Acineto-bacter baumannii, Neisseria gonorrhoeae, Klebsiellapneumoniae, MRSA, multidrug-resistant Mycobac-terium tuberculosis (MDR-TB), Vancomycin-resistant
Enterococci (VRE), and Vancomycin-intermediate/resistant Staphylococcus aureus (VISA/VRSA).2 Therapid emergence of these strains coupled with thewide variety of antimicrobial agents that resistancehas developed against dictates the need for new,highly effective antimicrobials that act on novel targetsor critical bacterial components to which resistancecannot be evolved.
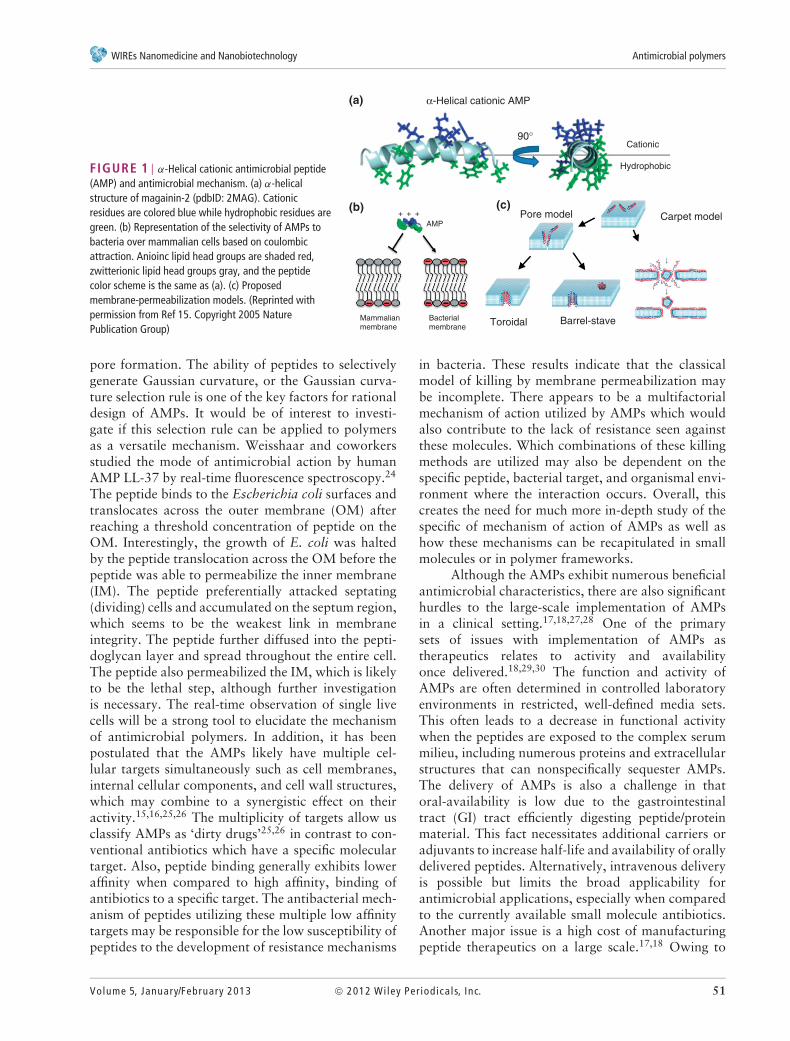
Host-Defense Antimicrobial PeptidesHost-defense AMPs have been extensively studied asnew alternatives for antibiotics because of little or nosusceptibility to the current resistance mechanismsof bacteria.15–22 AMPs are a class of peptides inthe innate immunity, which protect the body frominvasion of microbes by quick killing action. Thestructural feature of AMPs includes a relatively smallmolecular size (10–50 amino acids) and cationicamphiphilicity exerted by cationic and hydrophobicside chains.18 One of the classes is α-helical AMPsincluding magainin and LL-37, and these AMPs formhelix upon binding to bacterial cell membranes, andthe cationic and hydrophobic side chains are localizedon different side of helix15 (Figure 1(a)). AMPs attackbacterial cell membranes, which causes membranepermeabilization, leakage of cellular components,and breakdown of membrane potential, resultingin cell death. This contrasts the mode of action ofconventional antibiotics, which are generally enzymeand DNA replication inhibitors. The activity of AMPsis selective to bacteria over mammalian cells. Thisis because of preferential binding of cationic AMPsto high net negative charge on bacterial surfacesover mammalian cells (Figure 1(b)). Several molecularmechanisms of membrane permeabilization by AMPshave been proposed: the peptides accumulate onthe cell membranes and form discrete pores inthe membrane (toroidal and barrel-stave models)or disrupt the membrane nonspecifically (carpetmodel)15,19 (Figure 1(c)).
Although an in-depth discussion of antimicro-bial mechanism of AMPs is beyond the scope of thisreview, recent studies on natural HDPs provided newinsights into the mechanism of these membrane-activeantimicrobials. Wong and coworkers investigated therole of lipids and topological changes in the membranepermeabilization by peptides and peptidomimeticsusing small-angle X-ray scattering (SAXS).23 Natu-ral peptides selectively permeabilized model bacterialmembranes, while they do not destabilize modeleukaryotic membranes. The HDP defensin generateda negative Gaussian (saddle-splay) curvature in modelbacterial membranes composed of lipids that intrinsi-cally induce negative curvature, which is necessary for
50 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013
WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
FIGURE 1 | α-Helical cationic antimicrobial peptide(AMP) and antimicrobial mechanism. (a) α-helicalstructure of magainin-2 (pdbID: 2MAG). Cationicresidues are colored blue while hydrophobic residues aregreen. (b) Representation of the selectivity of AMPs tobacteria over mammalian cells based on coulombicattraction. Anioinc lipid head groups are shaded red,zwitterionic lipid head groups gray, and the peptidecolor scheme is the same as (a). (c) Proposedmembrane-permeabilization models. (Reprinted withpermission from Ref 15. Copyright 2005 NaturePublication Group)
Carpet model
Cationic90°
AMP+ + +
Hydrophobic
Pore model
α-Helical cationic AMP
ToroidalBacterial membrane
Mammalian membrane
Barrel-stave
(a)
(b) (c)
pore formation. The ability of peptides to selectivelygenerate Gaussian curvature, or the Gaussian curva-ture selection rule is one of the key factors for rationaldesign of AMPs. It would be of interest to investi-gate if this selection rule can be applied to polymersas a versatile mechanism. Weisshaar and coworkersstudied the mode of antimicrobial action by humanAMP LL-37 by real-time fluorescence spectroscopy.24
The peptide binds to the Escherichia coli surfaces andtranslocates across the outer membrane (OM) afterreaching a threshold concentration of peptide on theOM. Interestingly, the growth of E. coli was haltedby the peptide translocation across the OM before thepeptide was able to permeabilize the inner membrane(IM). The peptide preferentially attacked septating(dividing) cells and accumulated on the septum region,which seems to be the weakest link in membraneintegrity. The peptide further diffused into the pepti-doglycan layer and spread throughout the entire cell.The peptide also permeabilized the IM, which is likelyto be the lethal step, although further investigationis necessary. The real-time observation of single livecells will be a strong tool to elucidate the mechanismof antimicrobial polymers. In addition, it has beenpostulated that the AMPs likely have multiple cel-lular targets simultaneously such as cell membranes,internal cellular components, and cell wall structures,which may combine to a synergistic effect on theiractivity.15,16,25,26 The multiplicity of targets allow usclassify AMPs as ‘dirty drugs’25,26 in contrast to con-ventional antibiotics which have a specific moleculartarget. Also, peptide binding generally exhibits loweraffinity when compared to high affinity, binding ofantibiotics to a specific target. The antibacterial mech-anism of peptides utilizing these multiple low affinitytargets may be responsible for the low susceptibility ofpeptides to the development of resistance mechanisms
in bacteria. These results indicate that the classicalmodel of killing by membrane permeabilization maybe incomplete. There appears to be a multifactorialmechanism of action utilized by AMPs which wouldalso contribute to the lack of resistance seen againstthese molecules. Which combinations of these killingmethods are utilized may also be dependent on thespecific peptide, bacterial target, and organismal envi-ronment where the interaction occurs. Overall, thiscreates the need for much more in-depth study of thespecific of mechanism of action of AMPs as well ashow these mechanisms can be recapitulated in smallmolecules or in polymer frameworks.
Although the AMPs exhibit numerous beneficialantimicrobial characteristics, there are also significanthurdles to the large-scale implementation of AMPsin a clinical setting.17,18,27,28 One of the primarysets of issues with implementation of AMPs astherapeutics relates to activity and availabilityonce delivered.18,29,30 The function and activity ofAMPs are often determined in controlled laboratoryenvironments in restricted, well-defined media sets.This often leads to a decrease in functional activitywhen the peptides are exposed to the complex serummilieu, including numerous proteins and extracellularstructures that can nonspecifically sequester AMPs.The delivery of AMPs is also a challenge in thatoral-availability is low due to the gastrointestinaltract (GI) tract efficiently digesting peptide/proteinmaterial. This fact necessitates additional carriers oradjuvants to increase half-life and availability of orallydelivered peptides. Alternatively, intravenous deliveryis possible but limits the broad applicability forantimicrobial applications, especially when comparedto the currently available small molecule antibiotics.Another major issue is a high cost of manufacturingpeptide therapeutics on a large scale.17,18 Owing to
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 51

Advanced Review wires.wiley.com/nanomed
their nature as antibacterial, large scale fermentationsand biologically based production are incompatiblewith these molecules. The length and complexity ofmost AMPs are also incompatible with solution-phasechemical methodology, requiring time- and reagent-intensive solid phase methods for production.
PEPTIDE-MIMETIC DESIGN
To address these obstacles of AMPs, synthetic poly-mers have been designed as mimics of AMPs. In oneof the peptide studies, an all d-enantiomer magaininhomologs showed the same level of activity com-pared to natural magainin.31 This indicated that theantimicrobial action is not receptor-dependent, thatis, based on a specific protein–protein interaction.In addition, many AMP structures share the samephysicochemical properties of a net positive chargeand contain several hydrophobic amino acid residues,but yet no common consensus sequences or motifshave been found.18 While a significant number ofknown AMPs utilize the α-helical conformation tocreate a facially amphiphilic, active structure, recentevidence from the investigation of diastereomeric pep-tides indicate that the stable helix structure may notbe necessary for activity.19,32 In this study, severalamino acids were substituted with d-enantiomers,which disfavors helix formation. However, the pep-tides were still found to adapt amphiphilic conforma-tions with segregation of cationic and hydrophobicresidues, even without stable helix formation, whenbound to lipid membranes.19,32 These results suggestthat the cationic, amphiphilic properties, the three-dimensional conformation of AMPs, or some balanceof these two factors are likely the key determinants forantimicrobial activity rather than the exact sequence,stereochemistry, and/or stable secondary structure.Accordingly, the new design of antimicrobial polymershas been focused to reflect the cationic amphiphilicstructures of AMPs on the synthetic polymer plat-form rather than to mimic the secondary structuralconformations such as the α-helix.
In general, AMPs are relatively small, andthe cationic functionality is imparted by the pri-mary ammonium groups of lysine. Accordingly,polymer mimics of AMPs have been generatedto have low molecular weights (MWs) (a fewthousands) and primary ammonium side chains.11
Hydrophobic comonomers are also incorporated tomimic the amphiphilic property of AMPs, producingamphiphilic random copolymers. Several examplesof antimicrobial polymers are depicted in Figure 2.Polymers with quaternary ammonium side chains(polycations) have been widely utilized as polymeric
O
NR3+ +H3N
+H3N
+H3N
O
R′ OO
O
O
O ONHNH
O
O
4 Ph
O
N+
R
O
OO n m
O
(a) (b) (c)
(d) (e)
FIGURE 2 | Several examples of antimicrobial peptide(AMP)-mimicking polymers based on (a) methacrylate,37
(b) norbornene,38 (c) nylon,39 (d) vinyl ether,40 and (e) alternatingring-opening metathesis polymerization (ROMP) copolymers41.
disinfectants and have high MWs in general.8,33–36
Considering the net positive charge of the amphiphilicpolymers, AMP-mimetic polymers can be also clas-sified as polycations, but their low MW, primaryammonium side chains, and hydrophobic componentsprovide new functionalities and characteristics includ-ing polymer–lipid interactions and altered biologicalactivity. The roles of these factors are discussed below.
In the following sections, we discuss the antimi-crobial activity and toxicity of polymers by focus-ing the recent results from our laboratories.40,42–50
These polymers build on the fundamental amphiphilic,cationic properties found in most AMPs and extendthem to platforms which are compatible for antimi-crobial applications as well as being more scalablefor production. Many research groups are using dif-ferent classes of polymers, different assay conditions,and different strains of target bacteria, which resultin different measures of activity and data analysis. Inother words, antimicrobial effectiveness and toxicityof polymers depend on assay conditions. Therefore,direct comparisons of biological activities of polymersincluding MIC, IC50, and cytotoxicity in literaturemay be ambiguous, and can be qualitative at best.This review discusses results from our work withthe peptide-mimetic design of antibacterial polymersand their potential as new antimicrobials rather thanfocusing on comparisons of efficacy with other classesof polymers. The readers are advised to refer otherexcellent review articles for more detailed discussionson synthesis and mechanistic studies of other antimi-crobial polymers.7–10,33,51
ANTIMICROBIAL ACTIVITY
Antimicrobial AssaysThe activity of antimicrobial polymers against bacte-ria is evaluated as inhibition effect on bacterial growth
52 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013

WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
as well as bactericidal effect. The inhibition effect iscommonly measured as a minimum inhibitory concen-tration (MIC) of polymers, which is the concentrationthat completely inhibits bacterial growth. A micro-dilution method using a microplate has been popularbecause the assay protocol is relatively easy and suit-able for testing large number of compounds on a smallscale. The standard and modified assay protocols areavailable in literature.52,53 In general, known concen-trations of bacteria are incubated with a range ofconcentrations of polymers overnight, and bacterialgrowth is determined by increase in the turbidity ofsolution or optical density. It should be noted thatthis assay determines no growth of bacteria, whichindicate the inhibition of bacterial growth by poly-mers, but not necessarily due to killing of bacteriaby polymers. When the polymers inhibit the bacte-rial growth without killing (bacteriostatic effect), thebacteria are able grow again once the polymers areremoved by dilution or washing. The bacteriostaticand bactericidal effects of polymers should be dis-tinguished for proper data and mechanism analysis.In addition, the micro-dilution assay is generally per-formed by using a plastic multi-well microplate. It hasbeen reported that the activity of AMPs depends on theproperties of microplate plastics.54 AMPs showed lowactivity in polystyrene (PS) microplate compared to apolypropyrene (PP) microplate because amphiphilicpeptides can bind nonspecifically to the walls ofa PS plate, sequestering them from the bacterialsolution.53,54 Considering the amphiphilic propertyof antimicrobial polymers, it is reasonable to assumethat the selection of microplates affects the resultsof antimicrobial assay. On the other hand, bacteri-cidal effect is evaluated as the number of residualviable bacteria at given polymer concentrations orthe polymer concentrations necessary to kill certainnumber of bacteria, typically >99.9% or 3-log reduc-tion. The number of viable cells is usually reported bycolony forming units (cfu) determined through platingassays.
Broad Spectrum of ActivityAntimicrobial polymers generally show broad-spectrum activity against both Gram-positive andnegative bacteria. In contrast, conventional antibioticsare usually more specific because their uniquecellular targets such as enzymes or DNA/RNAreplication inhibitors. The polymers show slightlydifferent activity against bacteria, but there seemsto be no general trend for preference in termsof Gram-positive and negative strains. Methacrylatecopolymers displayed antimicrobial activity against
drug-resistant S. aureus (MRSA) with the samelevel of activity with the drug-susceptible laboratorystrain47 (Table 1). The polymer is effective against A.baumannii, which has developed significant resistanceagainst conventional antibiotic drugs. Recently, A.baumannii infections are found among patientstreated at overseas military medical facilities.55 Thebroad-spectrum activity of antimicrobial polymers canbe attributed to the membrane-disruption mechanism,which is likely to be less sensitive to the specificcell wall structures and lipid compositions in cellularmembranes.
Amphiphilic Polymer Structures and ActivityThe antimicrobial activity of polymers in generalincreases with increasing hydrophobic content in theside chains. This is likely because of the increasedhydrophobicity of polymers enhancing insertion ofpolymers into the hydrophobic region of cell mem-branes. Our recent computational investigation onmethacrylate polymers indicated that the hydropho-bic side chains of methacrylate random copolymersare inserted into lipid bilayers (Figure 3(a) and (b)).47
It could be speculated that the polymer insertion to cellmembranes would be enhanced by the hydrophobicityof side chains. In the same study, methacrylate ran-dom copolymers are more deeply inserted into lipidbilayers when the spacer arms in the cationic sidechain are elongated. This is because of the snorkelingeffect, in which the cationic ammonium groups arebound to the anionic phosphate lipid head groups,
TABLE 1 Activity Spectrum of Representative MethacrylateCopolymer (E429). (Reprinted with permission from Ref 47. Copyright2012 American Chemical Society)
Bacteria or Human Cell MIC or HC50 (μg/mL)
E429 Melittin Magainin-2
Gram-positive
Staphylococcus aureus 63 6 >250
Staphylococcus aureus (CA-MRSA)1 31 n.d. n.d.
Enterococcus faecalis 31 16 >500
Bacillus subtilis 8 6 63
Gram-negative
Escherichia coli 21 12 125
Acinetobacter baumannii 10 n.d. n.d.
Pseudomonas aeruginosa 16 280 500
Salmonella enterica 16 140 500
Human RBC (HC50) 1300 2 >250
n.d., not determined.1Community acquired methicillin-resistant S. aureus (MRSA) strain LAC1236.
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 53

Advanced Review wires.wiley.com/nanomed
OO
OO
OO
OO
OO
OO
OO
OO
OO
CH3
NH3+NH3
+NH3+NH3
+NH3+NH3
+NH3
+m m m m m m m
O
H
Model polymers
Conformation of E4
E2 E4 E6
O
(a)
(b)
(c)
FIGURE 3 | Simulated binding and insertion of methacrylate copolymers into lipid bilayers. (a) Model polymer structures with different cationicspacer arm lengths, m = 2 (E2), 4 (E4), and 6 (E6). (b) Snapshots of polymers inserted into phosphatidylethanolamine/ palmitoyloleoylphosphatidylglycerol (POPE/POPG) lipid bilayers. (c) Conformation of E4 in the lipid bilayer. The polymer structures are presented by green (polymer backbone),red (cationic side chains), and blue (hydrophobic ethyl side chains) colors. (Reprinted with permission from Ref 47. Copyright 2012 AmericanChemical Society)
and the long arm or spacer enables the polymer chainsto separate from the membrane surface and sink intothe lipid bilayer (Figure 3(c)). The concept of snorkel-ing effect has been developed originally in the peptidescience. Transmembrane peptides have been modifiedto have extended spacer arms in ammonium (lysineand guanidine) side chains. These long arms withcationic groups can reach to the water–lipid interface,which stabilizes the hydrophobic peptide helices inthe hydrophobic domains of lipid bilayers. Therefore,the snorkeling effect can modulate the position andorientation of transmembrane peptides.56–63 The sim-ilar effect was found in our methacrylate copolymersdiscussed above (Figure 3). Interestingly, the poly-mer chain is stretched when they are bound to thelipid bilayer, and the cationic groups and hydropho-bic (ethyl) side chains of comonomers are segregatedrelative to the polymer backbone. This amphiphilicstructure is similar to AMPs such as magainin, whichhave also cationic and hydrophobic side chains seg-regated into the different side of helix. Similarly,Yethiraj and coworkers demonstrated that randomcopolymer models of β-peptides bind to lipid bilayersand form the segregated amphiphilic structures.64 Theamphiphilic structure of polymers and peptides when
bound to cell membranes may be the key determinantrather than the identity and heterogeneity of chemicalstructure and conformation.
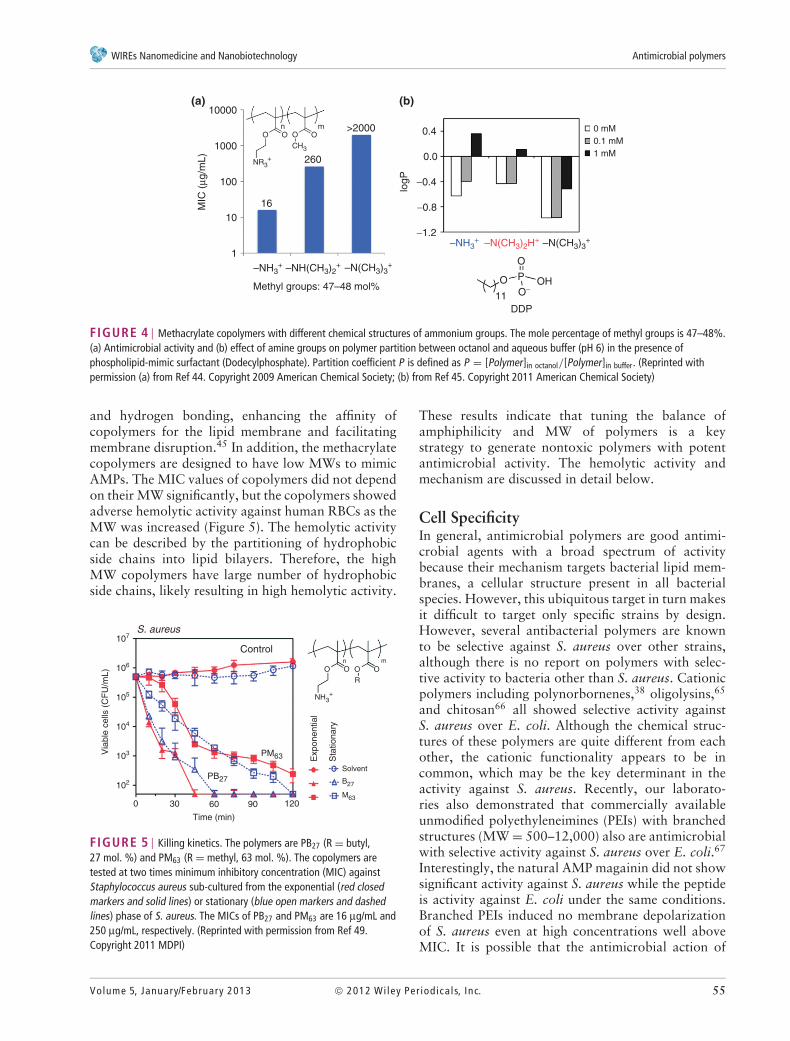
To mimic the functionality of AMPs, antimi-crobial polymers were prepared to have low MWsand primary ammonium side chains, although thisdesign lacked any published experimental data to sup-port the design. To that end, we investigated thestructure–activity relationship of methacrylate ran-dom copolymers. To study the role of cationic sidechains, we hypothesized that the antimicrobial activ-ity of copolymers would not depend on the chemicalstructures of amine groups if the cationic net chargeof polymers were only essential in their underlyingantimicrobial mechanism. To test this hypothesis, weinvestigated the effect of chemical structure of aminegroups (primary, tertiary and quaternary ammo-nium) on the antimicrobial and hemolytic activities ofmethacrylate copolymers (Figure 4(a)). The study indi-cated that the methacrylate copolymers with primaryammonium groups showed the highest activity whilethe copolymers with quaternary ammonium groupsare not active. We speculate that the ammoniumgroups form complexes with phosphate lipid headsthrough a combination of electrostatic attraction
54 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013
WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
0.0
logP
0.4
−0.4
−0.8
−1.2
Methyl groups: 47–48 mol%O
11
O
–N(CH3)3+–N(CH3)2H+–NH3
+
–N(CH3)3+–NH(CH3)2
+–NH3+
P OHO−
DDP
0 mM0.1 mM1 mM
16
1
10
100
1000
MIC
(µg
/mL)
10000
260
O OO
NR3+
CH3
Omn >2000
(a) (b)
FIGURE 4 | Methacrylate copolymers with different chemical structures of ammonium groups. The mole percentage of methyl groups is 47–48%.(a) Antimicrobial activity and (b) effect of amine groups on polymer partition between octanol and aqueous buffer (pH 6) in the presence ofphospholipid-mimic surfactant (Dodecylphosphate). Partition coefficient P is defined as P = [Polymer]in octanol/[Polymer]in buffer. (Reprinted withpermission (a) from Ref 44. Copyright 2009 American Chemical Society; (b) from Ref 45. Copyright 2011 American Chemical Society)
and hydrogen bonding, enhancing the affinity ofcopolymers for the lipid membrane and facilitatingmembrane disruption.45 In addition, the methacrylatecopolymers are designed to have low MWs to mimicAMPs. The MIC values of copolymers did not dependon their MW significantly, but the copolymers showedadverse hemolytic activity against human RBCs as theMW was increased (Figure 5). The hemolytic activitycan be described by the partitioning of hydrophobicside chains into lipid bilayers. Therefore, the highMW copolymers have large number of hydrophobicside chains, likely resulting in high hemolytic activity.
0
102
103
104
Via
ble
cells
(C
FU
/mL)
105
106
107
30 60
PB27
PM63
Control
S. aureus
Time (min)
90 120
Solvent
Exp
onen
tial
Sta
tiona
ry
O O OO
NH3+
n m
R
B27
M63
FIGURE 5 | Killing kinetics. The polymers are PB27 (R = butyl,27 mol. %) and PM63 (R = methyl, 63 mol. %). The copolymers aretested at two times minimum inhibitory concentration (MIC) againstStaphylococcus aureus sub-cultured from the exponential (red closedmarkers and solid lines) or stationary (blue open markers and dashedlines) phase of S. aureus. The MICs of PB27 and PM63 are 16 μg/mL and250 μg/mL, respectively. (Reprinted with permission from Ref 49.Copyright 2011 MDPI)
These results indicate that tuning the balance ofamphiphilicity and MW of polymers is a keystrategy to generate nontoxic polymers with potentantimicrobial activity. The hemolytic activity andmechanism are discussed in detail below.
Cell SpecificityIn general, antimicrobial polymers are good antimi-crobial agents with a broad spectrum of activitybecause their mechanism targets bacterial lipid mem-branes, a cellular structure present in all bacterialspecies. However, this ubiquitous target in turn makesit difficult to target only specific strains by design.However, several antibacterial polymers are knownto be selective against S. aureus over other strains,although there is no report on polymers with selec-tive activity to bacteria other than S. aureus. Cationicpolymers including polynorbornenes,38 oligolysins,65
and chitosan66 all showed selective activity againstS. aureus over E. coli. Although the chemical struc-tures of these polymers are quite different from eachother, the cationic functionality appears to be incommon, which may be the key determinant in theactivity against S. aureus. Recently, our laborato-ries also demonstrated that commercially availableunmodified polyethyleneimines (PEIs) with branchedstructures (MW = 500–12,000) also are antimicrobialwith selective activity against S. aureus over E. coli.67
Interestingly, the natural AMP magainin did not showsignificant activity against S. aureus while the peptideis activity against E. coli under the same conditions.Branched PEIs induced no membrane depolarizationof S. aureus even at high concentrations well aboveMIC. It is possible that the antimicrobial action of
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 55

Advanced Review wires.wiley.com/nanomed
PEIs is not to disturb the cell membranes, but act inthe cell wall.65,66 The cationic polymers may also betrapped by the peptidoglycan layer of E. coli, inhibit-ing polymer diffusion to the cytoplasmic membrane.68
Therefore, the selective activity of cationic polymersmay be linked to the structural difference in the bacte-rial cell walls as well as differences in the antimicrobialmechanism. Of note, since the tested strains are lim-ited, it is not clear at this time that the activity of thesepolymers is specific to Gram-positive strains generalor specifically for S. aureus. Although the mechanismof cationic polymers for anti-S. aureus activity andthe selective activity is not clear yet, it would be ofinterest for further investigation, which will providenew insight into molecular design of next generationantimicrobials.
Biocidal KineticsIn order to assess the timeframe required forantimicrobial polymers to kill bacteria, the numberof viable bacterial cells is determined as a functionof exposure time. In an assay, bacteria are incubatedwith polymers, and an aliquot of this treated cultureis taken and diluted in buffer solution to removethe effect of polymers. Then the diluted solution isspread in an agar plate and incubated overnight toallow formation of bacterial colonies. The number ofcolonies on the agar plate is then counted. The assaysolution needs to be appropriately diluted such thatthe final polymer concentration is much lower thaneffective concentrations of polymers, otherwise, carry-over of polymers may inhibit colony formation. But,the bacteria concentration needs to be high enoughto give colonies in countable numbers for accuracy.In principle, one bacterial cell forms one colony.Therefore, the number of viable cells after incubationwith polymers can be calculated by taking the dilutionfactors into account and reported as a colony-formingunit (cfu). Typically these cfu values are converted intothe units cfu per a given volume (typically cfu/mL)based on the dilution factors used and the volume ofassay solution that was applied to the plate.
In general, antimicrobial polymers display quickbactericidal effect against bacteria: they kill more than99.9% within an hour at the MIC, which is thesame level of effect with antibacterial peptides. Inour study on methacrylate copolymers, the effect ofgrowth phases of bacteria was also investigated.49 Thecopolymers displayed no significant difference in thekilling rate against E. coli and S. aureus in the station-ary and exponential growth phases (Figure 5). Thisindicates that the mechanism of antimicrobial actionof the copolymers does not rely on the metabolicphysiological activity associated with bacterial growth
phases. Bacteria are less metabolically active whenthey are in biofilms or exposed to antibiotics, whichis one of the bacterial defense mechanisms againstantibiotics which can then lead to the developmentof antibiotic resistance. The copolymers we stud-ied would likely be effective against bacteria in thedormant state or in biofilms as well as being lesssusceptible to resistance development through dor-mancy. It should be noted that activity against bacteriawith decreased metabolic activity, as in biofilms, isnot the only challenge in efficacy against biofilms.One of these challenges is that the cationic polymersmay become ‘trapped’ in the biofilm matrix com-posed of anionic biopolymer exopolysaccharide (EPS),which would limit the polymer diffusion and reduc-ing their efficacy. However, compared to conventionalantibiotics, the potential activity of polymers againstdormant bacteria is poised to be a major advantagefor the development of these types of antimicro-bial polymers effective against bacterial biofilms andbiofilm-associated infections.
TOXICITY TO HUMAN CELLS
HemolysisLytic activity to red blood cells or hemolytic activ-ity has been used as a measure of first assessment ofcytotoxicity of antimicrobial polymers. In a hemolysisassay, the polymers are incubated with a suspensionof RBCs for a few hours. After centrifugation of assaysolution, red blood cells sediment to the bottom of ves-sels or wells, and the amount of hemoglobin released inthe supernatant is determined spectroscopically. Thehemolytic activity of polymers is quantified by the per-centage of released hemoglobin relative to 100% lysisof all RBCs by, typically, a surfactant. As the poly-mers are supposed to act on bacterial cell membranes,damages to human cells are the first assessment forcell selectivity. One of the reasons for popularity ofhemolysis is likely due to the easy assay procedurecompared to other cytotoxicity assays using prolifer-able cells. It should be noted that there is no standardprotocol for hemolysis available which yields diffi-cult comparisons between experimental systems andmolecular species in question. Hemolytic activity isknown to be very sensitive to the assay conditions asisolated RBCs so care should be taken to minimizeexperimental variability. One should employ extremecaution when attempting to directly compare specificvalues for hemolytic activity from the literature andinstead should primarily focus on behavioral trends.
In general, when methacrylate copolymers con-tain a large amount of hydrophobic mass (eitherby number of groups, size of groups, or both), the
56 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013

WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
polymers behave as hydrophobic and hemolytic.50 Thepolymers are expected to bind selectively to bacterialcell surfaces over human cells because of electro-static interaction. However, when the hydrophobicityof polymers dominates the cellular binding of poly-mers, the polymers start to bind nonspecifically toboth cell-types and cause hemolytic activity. As thehydrophobic interaction is also the driving forceto enhance their antimicrobial activity, the balancebetween hydrophobicity and cationic functionalities isa key determinant for design of polymers with potentactivity and selective activity against bacteria overhuman cells. Methacrylate copolymers with a rangeof lengths (C1–C6) and contents of alkyl side chainsshowed that the hemolytic activity can be describedby portioning of alkyl groups into the hydrophobicdomain of lipid bilayers (Figure 6).50 In addition, forthe copolymers with high contents of alkyl methacry-lates, we observed polymer aggregates in water, whichdecreases the number of polymer chains active oraccessible to cells, resulting in no enhancement ofactivity by further increases in hydrophobicity.
How can we remove the hemolytic activity frompolymers without losing antimicrobial activity? This isa million dollar question in the field. We have severalinsights into the approach in polymer deign from ourlaboratory and the literature. From the same studyon methacrylate copolymers,50 short alkyl side chainsseem to provide better selective activity compared tocounterparts with longer alkyl side chains. One of thereasons may be because of small incremental changesin hydrophobicity allowing fine-tuning of polymerhydrophobicity for optimal balance between cationicand hydrophobic groups. In addition, the polymers
need more methyl groups to get the same level ofactivity as longer alkyl groups. This may indicatethat the distribution of hydrophobicity along thepolymer chain may be important. Indeed, cationicamphiphilic vinyl ether block copolymers showedlittle or no hemolytic activity compared to randomcopolymers.40 Sen and coworkers demonstrated thatspatial separation of cationic charges and hydrophobicgroups in amphiphilic pyridium-methacrylate randomcopolymers results in increased antimicrobial andhemolytic activities, indicating a higher membrane-disrupting ability.36 Although the chemical structuresof ammonium groups (primary ammonium vs pyrid-ium) of these examples are different, the distribution ofhydrophobicity along the polymer chains and overallamphiphilic sequences are one of the key determinantsto control the antimicrobial and hemolytic activi-ties. The polymeric amphiphilic structure with tunedhydrophobic distribution will be an interesting subjectfor further research. In addition, the amphiphilicity ofpolymers, polymer MW and MW distribution alsoaffect the hemolytic activity of polymers. In general,low MW polymers are less hemolytic than high MWcounter parts.11,50,37 In the study on nylon polymers,copolymers were purified by dialysis and the resultantpolymers are much less hemolytic.39 This indicatesthat the high MW components in polydisperse poly-mer samples are more responsible for hemolysis.
Although hemolysis has been widely used toevaluate polymer toxicity, the hemolysis mechanismhas not been studied in detail. In this regard,methacrylate copolymers have been reported to causeall-or-none hemolysis, in which a fraction of RBCswere lysed and released all hemoglobin into solution,
(a) (b)1000
100
10
HC
50 (
µM)
0.1
0.010 10 20 30
Mole percentage of butyl groups (mol. %)
40 50 60 70
~8.700 (32–46)
~5.000 (19–31)
~1.600 (5–9)
0.1 1
Average number of side chains / polymer chain
10 100
1
1000
100
10MW (DP)
HC
50 /
µM
0.1
0.01
1
R:MethylEthyl
Butyl
HexylBenzyl
O O
R
NH3+
OOn m
O O
NH3+
OOn m
FIGURE 6 | Hemolytic activity of methacrylate copolymers. (a) Molecular weight (MW) dependence of hemolytic activity. High MW copolymersare more hemolytic. (b) Correlation between the number of hydrophobic groups in a polymer chain and HC50. (Reprinted with permission from Ref 50.Copyright 2009 Wiley-VCH Verlag GmbH & Co. KGaA)
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 57

Advanced Review wires.wiley.com/nanomed
120
100
80
60
Dis
appe
ared
RB
Cs
(%)
40
20
0
0 20 40 60Hemolysis (%)
80 100 120
All-or-none release
Graded release
0 25 50
Apparent hemolysis (%)
75
100
80
60
40
20
0
No 400osmolyte
800 1200
PEG MW
1600
Melittin
PB27
Osmotic lysisHypotonic condition
H2O
H2O
H2OH2O
H2OH2O
+ Polymer
(a) (b) (c)
(d)
FIGURE 7 | All-or-none osmotic hemolysis induced by methacrylate polymer. The polymer PB27 contains 27 mol. % of butyl groups. (a) Thecorrelation between percentages of hemolysis (released hemoglobin) and disappeared cells. The good correlation suggests that the hemolysis causedby the polymer is an all-or-none type event. (b) Schematic presentation of all-or-none and graded release of hemoglobin from RBCs. (c) Osmoticprotection by polyethylene glycols (PEGs). High molecular weight (MW) PEGs (> 1000) suppressed hemolysis by the polymer. The threshold MW ofPEG for hemolysis inhibition is 500–1000, corresponding to 1.6–2 nm in diameter. (d) Schematic representation of the osmotic lysis mechanism ofpolymer action. (Reprinted with permission from Ref 48. Copyright 2011 American Chemical Society)
rather than all RBCs released a part of hemoglobin(graded leakage) (Figure 7(a) and (b)).48 A hemoly-sis assay using osmoprotectants such as polyethyleneglycols (PEGs) suggested that the copolymers formnano-sized pores (1–2 nm in diameter) in RBC cellmembranes, causing osmotic lysis of RBCs due toosmotic imbalance between the outer buffer solutionand the cytoplasm of RBCs (Figure 7(c) and (d)).The bee venom toxin melittin also formed nano-pores(∼2 nm), followed by osmolysis of RBCs. Althoughthe peptide and polymers have distinctive difference intheir chemical structures, they seem to share the sameunderlying mechanism.
Cytotoxicity AssaysIn addition to hemolysis, other standard cytotoxicityassays have been used for assessment of toxicity ofantimicrobial polymers to mammalian cells, althoughthe number of reports is limited. One assay methodis to monitor a lease of lactate dehydrogenase (LDH)from the cytoplasm, which reports damage to thecell membrane.67 The released LDH is quantifiedby spectroscopically monitoring product formationcatalyzed by the enzymatic reaction rather than the
direct detection of LDH, which contrasts to thehemolysis assay. An MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay has beenalso used for cell viability.69 This method detects themetabolic activity of cells using a substrate which
00
100
200
300
400
Fol
d in
crea
se M
IC
500
600
5 10 15
Serial passage number
20 25
O
NH3+
O O O
R
n m
Ciprofloxacin
Norfloxacin
PM63
PB27
FIGURE 8 | Susceptibility of methacrylate copolymers andantibiotics to the development of resistance in Escherichia coli. Thepolymers are PB27(R = butyl, 27 mol. %) and PM63 (R = methyl, 63 mol.%). (Reprinted with permission from Ref 49. Copyright 2011 MDPI)
58 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013

WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
produces distinctive color change when reduced byactive components of normal cellular metabolism.Recently, a water-soluble version of substrate, XTT(sodium 2,3,-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)-carbonyl]-2H-tetrazolium), has beenused for the same assay to facilitate the procedure.67
Combining these assay methods, we are able to char-acterize the cytotoxicity of polymers as well as givinginsights into the mechanism of cytotoxicity. Theseassays have been utilized for cationic polymers.69
In our recent report, antimicrobial branched PEIswith low MWs (MW = 500 and 1100) showed norelease of LDH from human epithelial HEp-2 cells,but the cell viability was reduced significantly after24-h incubation.67 This indicates that PEIs do notdamage the cell membranes, but may cause potentiallong-term toxicity to host cells. In the literature, thecytotoxicity of antimicrobial polymers has not beenfully evaluated. It would be important to investigatethe interaction between antimicrobial polymers andhost cells toward clinical applications.
ANTIMICROBIAL RESISTANCEIN BACTERIA
Low susceptibility to the existing or emerging resis-tance mechanisms is one of the hallmarks of AMPs.Although the number of reports is limited, someantimicrobial peptidomimetics and polymers appearto be effective against strains already resistant toconventional antibiotics and do not contribute tothe development of new antimicrobial resistance.49,70
We have investigated the occurrence of resistance inE. coli using methacrylate copolymers (Figure 8).49
Methacrylate copolymers at half the MIC were incu-bated with E. coli, and the solution was culturedovernight. Then the copolymers were tested againstthe bacterial cells, and MIC was determined. The bac-teria were cultured again at half of the new MIC,and MIC was determined after overnight incubation.In this assay, if the bacteria develop resistance, theMIC increases and thus the amount of antibioticsincrease at the subsequent culture. Therefore, thiscondition is likely to stimulate and accelerate theresistance development in bacteria. After 21 passagesof this process, the MIC values of conventional antibi-otics Ciprofloxacin and Norfloxacin increased 200-and 500-fold higher, indicating significant resistancedevelopment. On the other hand, the MIC values ofmethacrylate copolymers did not increase, indicatingno resistance development under this condition. Thebacteria incubated with the copolymers were suscep-tible to the antibiotics Norfloxacin and Ciprofloxacin,and the copolymers were active against bacteria
resistant to these antibiotics. This demonstrates thatthe copolymers do not contribute to cross-resistancedevelopment in bacteria.
FACTORS TO INFLUENCE ACTIVITY
It has been reported that environmental factors inhibitthe activity of AMPs and polymers.17 For the imple-mentation of antimicrobial polymers in biomedicalapplications, it is important to evaluate the polymeractivity in the physiological conditions and to under-stand the factors that affect the activity. These factorsthat affect efficacy can be widely variable, but a goodstarting point are the variables that relate to the mech-anism of action of the polymers in question. In thecase of the antimicrobial polymers described here, thepolymer selectivity relies on electrostatic interactionswhile the membrane disruptive activity relies primar-ily on hydrophobic interactions. As such, the role ofpH, ionic strength, and serum components that havesome hydrophobic character are likely modulators ofin vivo efficacy.
Solution pHThe polymers and peptides described have primaryamine groups, which exhibits a pH dependence onionization behavior. It has been demonstrated thatsolution pH changes polymer’s antimicrobial activity(Figure 9). In the study on methacrylate copolymers,copolymers were tested for their activity against E. colibetween pH 6 and 8.44 The copolymers showed higheractivity at higher pH. These data suggest that thecopolymers are likely more hydrophobic due to theincreased population of nonprotonated amine groupsat higher pH, which are less hydrophilic (and hencemore hydrophobic) than the charged versions of thesemoieties. It should be noted that uncharged primary
10000
1000
100MB
C (
µM)
105.0 6.0 7.0
pH
8.0 9.0
OOOOn m
CH3
NH3+
OOOOn m
CH3
NH+
FIGURE 9 | pH effect on antimicrobial activity (MIC) of methacrylatecopolymers with primary or tertiary ammonium side chains. (Reprintedwith permission from Ref 44. Copyright 2009 American ChemicalSociety)
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 59

Advanced Review wires.wiley.com/nanomed
PM47
On
x1–x
H
OO
R
NH3+
O
Polymer
PM47
PM63
PB27
Magainin-2Melittin
MeMe
Bu
4763
27
2017
16
2.8
MW/ 1000DPR MPalkyl
2.2
2.7
2.5
2.8––
––
––
O
O S
E. coli
S. aureus
PM63
PB27
Melittin
PM47
PM63
PB27
Melittin
0 2 4 6 8
0 2 4
Fold increase MIC
6 8
MgCI2 1 mMCaCI2 2 mMMgCI2+CaCI2NaCI 150 mM
(a)
(b)
FIGURE 10 | Effect of physiological salts on the antibacterial activity of methacrylate copolymers (PB27: 27% of butyl side chains, PM63: 63% ofmethyl side chains) and melittin against Escherichia coli (a) and Staphylococcus aureus (b). (Reprinted with permission from Ref 49. Copyright 2011MDPI)
amines are not truly hydrophobic unlike hydrophobicside chain groups (methyl) in comonomers, which areexpected to actively interact with the hydrophobicdomains of lipid membranes. The deprotonation ofammonium groups reduces the solvation (hydration)of polymers, resulting in a shift of amphiphilic balanceof polymers to a more hydrophobic character andthus increased antimicrobial and hemolytic activitiesas the solution pH increases. This supports the notionthat the hydrophobicity of copolymers increased,which dominate the electrostatic attraction betweenthe copolymers and bacterial cell surfaces, causingnonselective binding to both cells. The number ofcationic protonated ammonium groups decreases aspH increases, and the affinity of copolymers tobacterial cell surfaces could be reduced. However, it isevident that the overall hydrophobicity of copolymersis more effective to increase the antimicrobial activity.
It has been known that the ammonium groupsare cationic sources for selective binding to anionicbacterial cell wall, resulting in selective toxicityto bacteria over human cells. What if the animegroups are always charged? To this end, methacrylatecopolymers with primary, tertiary, and quaternaryammonium groups in the side chains were prepared
and their activity was measured.44 The copolymerswith primary ammonium groups showed highestactivity against E. coli. The activity was further testedin different pH conditions. According to separatedpotentiometirc titrations, most of amine groups areprotonated at pH 6, suggesting all polymers have thesame number of cationic groups in a polymer chain.It is interesting that the copolymers with primaryammonium groups showed the highest activity againstE. coli again. These results suggest that the aminegroups are not only cationic sources, but they areacting in the lipid–polymer interactions. The furtherinvestigation by examining partition of copolymersbetween water and octanol suggested that the primaryammonium groups are likely to form complex withphosphate lipid heads through hydrogen bonding andelectrostatic interaction (Figure 4).45 This complexformation might increase the affinity of copolymerswith primary ammonium groups for bacterial cells aswell as possibly enhances the antimicrobial mecha-nism of membrane disruption or pore formation.
Ionic Strength and Divalent IonsAs polymer binding to bacteria relies on electrostaticinteraction, high salt concentration (ionic strength)
60 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013

WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
1000
100
BC
99.9
(µg
/mL)
10
1
0.120 40 60
MPIBVE (%)80 100 0.10.001
−20
0
20
40
60
Hem
olys
is (
%) 80
100
120
Block copolymer
poly(AEVE)
NH2
CH2 CH
Hydrophilic Hydrophobic
On
CH2 CH
Om
poly(IBVE)
Random copolymer
[Polymer] (µg/mL)
10 1000
BlockRandom
BlockRandom
0
+
+++++ + + + ++
(a)
(b) (c)
FIGURE 11 | Antimicrobial random and block copolymers. (a) Cationic amphiphilic block and random vinyl ether copolymers. (b) Biocidal activityof copolymers. BC99.9 (biocidal concentration of polymers for 99.9% killing) is plotted as a function of MPIBVE (mol. % of IBVE units). (c) Hemolyticactivity of block and random copolymers with ∼25 mol. % of hydrophobic isobutyl vinyl ether (IBVE) units. (Reprinted with permission from Ref 40.Copyright 2012 American Chemical Society)
curtains the attractive interactions between polymersand bacteria, resulting in low affinity and low activity.The methacrylate copolymers showed twofold tofourfold increase in MIC against E. coli in MuellerHinton (MH) broth with additional 150 mM NaCl(Figure 10).49 Divalent cations Mg and Ca also reducethe activity of polymers.49 The activity of methacrylatecopolymers was twofold reduced in the presence ofthese cations (1 mM). These cations are structuralions, which stabilize the lipopolysaccharide (LPS)structures on the cell surfaces, preventing membranedisruption by the polymers. The high concentrationof these cations may also reduce the replacement ofcations in the LPS by the cationic groups of polymers,which prevent disintegration of membrane structure.
Serum ProteinsWhen AMPs are tested in the presence of serum,the peptides’ activity is significantly reduced.17,71,72
This has been attributed to enzymatic degradation ofpeptide chains (proteolysis), high ionic strength, and
nonspecific binding to serum proteins such as bovineserum albumin (BSA). It seems that activity of poly-mers in serum or physiological fluids have not beenevaluated to the same degree, and of those studiesavailable, many focus on the interaction of serumcomponents with surface-attached polymers and notsoluble structures as described herein. Similar to thecase of AMPs, the increased ionic strength of theserum environment may screen the electrostatic attrac-tive interactions between the cationic groups on thepolymers and the intended bacterial surface targets.Alternatively, the nonpolar moieties present in antimi-crobial polymers may drive nonspecific hydrophobicclustering with larger serum proteins. In the case ofantimicrobial polymers, a majority of compatibilitytesting has relied on simple protein adsorption assaysin which a standard serum protein, such as BSA,is used to test for nonspecific interactions withthe polymers.73,74 The complex serum environmentengenders the need for future investigation of thesecomplexities that may modify the efficacy and selec-tivity of antimicrobial polymers.
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 61
Advanced Review wires.wiley.com/nanomed
OOOO
In buffer solution
Antimicrobial polymer
Self-degradation
O
OOOOO
NH3 NH3
NH2
NH2
NH2
H2N
NH2
CI
O
O O O O
O
OOOOCI
NH2NH2
OOOOCI
OHO O
O OOHN
+
CI+ + −−
CI
NH3 NH3CI+ + −−
CI
OO
O Buffer pH 7.0, 37°C
Self-degradation
Time Mp (MALDI)
0 h 5800
0.5 h 4700
3 h 4200
18 h 3500
360 h 1700
m/z700060005000400030002000
(e)
(d)
(c)
(b)
(a)
CIn m
O
NH3+ −
CI
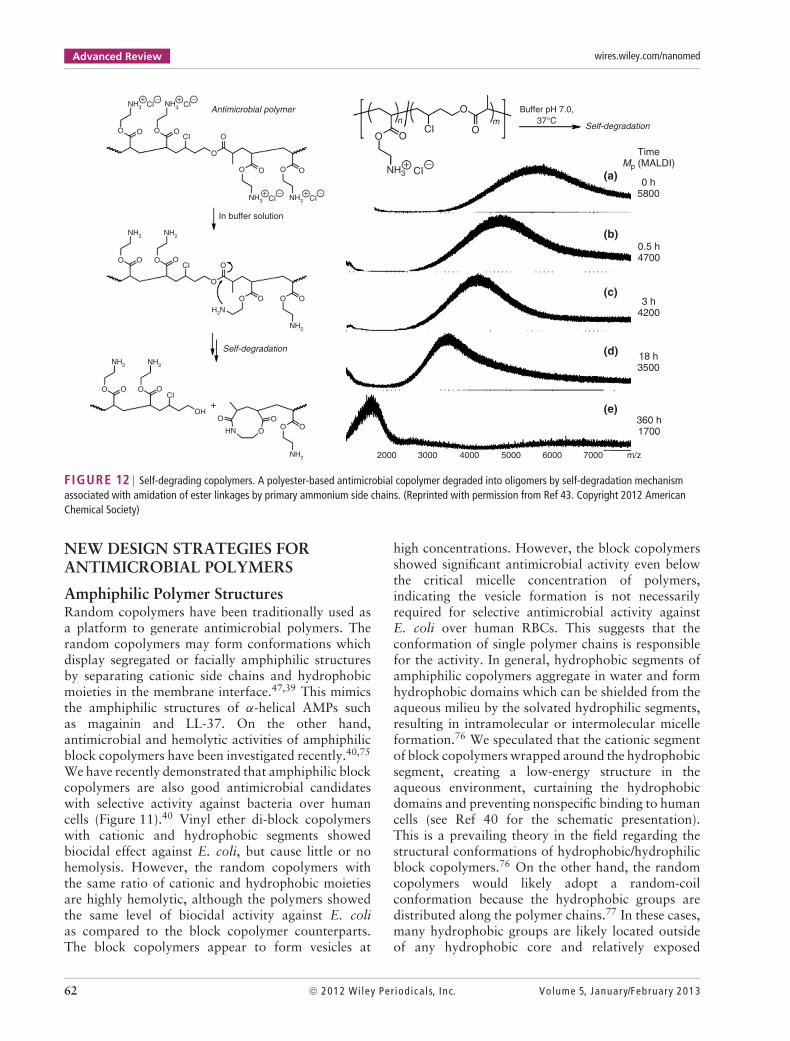
FIGURE 12 | Self-degrading copolymers. A polyester-based antimicrobial copolymer degraded into oligomers by self-degradation mechanismassociated with amidation of ester linkages by primary ammonium side chains. (Reprinted with permission from Ref 43. Copyright 2012 AmericanChemical Society)
NEW DESIGN STRATEGIES FORANTIMICROBIAL POLYMERS
Amphiphilic Polymer StructuresRandom copolymers have been traditionally used asa platform to generate antimicrobial polymers. Therandom copolymers may form conformations whichdisplay segregated or facially amphiphilic structuresby separating cationic side chains and hydrophobicmoieties in the membrane interface.47,39 This mimicsthe amphiphilic structures of α-helical AMPs suchas magainin and LL-37. On the other hand,antimicrobial and hemolytic activities of amphiphilicblock copolymers have been investigated recently.40,75
We have recently demonstrated that amphiphilic blockcopolymers are also good antimicrobial candidateswith selective activity against bacteria over humancells (Figure 11).40 Vinyl ether di-block copolymerswith cationic and hydrophobic segments showedbiocidal effect against E. coli, but cause little or nohemolysis. However, the random copolymers withthe same ratio of cationic and hydrophobic moietiesare highly hemolytic, although the polymers showedthe same level of biocidal activity against E. colias compared to the block copolymer counterparts.The block copolymers appear to form vesicles at
high concentrations. However, the block copolymersshowed significant antimicrobial activity even belowthe critical micelle concentration of polymers,indicating the vesicle formation is not necessarilyrequired for selective antimicrobial activity againstE. coli over human RBCs. This suggests that theconformation of single polymer chains is responsiblefor the activity. In general, hydrophobic segments ofamphiphilic copolymers aggregate in water and formhydrophobic domains which can be shielded from theaqueous milieu by the solvated hydrophilic segments,resulting in intramolecular or intermolecular micelleformation.76 We speculated that the cationic segmentof block copolymers wrapped around the hydrophobicsegment, creating a low-energy structure in theaqueous environment, curtaining the hydrophobicdomains and preventing nonspecific binding to humancells (see Ref 40 for the schematic presentation).This is a prevailing theory in the field regarding thestructural conformations of hydrophobic/hydrophilicblock copolymers.76 On the other hand, the randomcopolymers would likely adopt a random-coilconformation because the hydrophobic groups aredistributed along the polymer chains.77 In these cases,many hydrophobic groups are likely located outsideof any hydrophobic core and relatively exposed
62 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013

WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
to the solution, which can easily facilitate interac-tion with RBC cell membranes and cause hemolysis.This study provided a new insight into the impor-tant role which amphiphilic polymer structures playin their antimicrobial mechanism. In addition, ithas been demonstrated that nanoparticles formed byamphiphilic block copolymers are effective againsta broad spectrum of fungi and bacteria includinga drug-resistant strain.78,79 The molecular designfor the next generation of antimicrobial polymerswill be extended to include macromolecular struc-tures and architectures that build on the increasingmechanistic understanding of both polymeric and pep-tide antimicrobials. These should take advantage ofthe known molecular components that contribute tospecificity and efficacy, while additionally incorporat-ing beneficial properties to aid in bioavailability anddelivery.
Biodegradable Antimicrobial CopolymersAntimicrobial polymers with synthetic biodegradablebackbones including, for example, esters and carbon-ate groups have been reported.78,80–82 Our researchgroup has prepared polyester-based amphiphiliccopolymers with cationic side chains by simulta-neous chain- and step-growth radical polymerization(Figure 12).43 The random copolymers contain pri-mary ammonium groups in the side chains and esterlinkages in the polymer backbone. The copolymersdisplayed antibacterial activity against E. coli, butdegraded into oligomers at neutral pH, and theactivity of copolymer decreased. The detailed inves-tigation using control copolymers with acidic sidechains instead of amine groups, we speculate thatthe primary amine groups in the side chains reactwith the ester groups of the polymer backbone, andthe following amidation cleaves the polymer chains.This self-degradation mechanism will be useful forquick disintegration of polymer chains into inactiveoligomers, avoiding potential toxicity after long-termexposure.
CONCLUSION
Antimicrobial polymers are potential candidatesfor antibiotics, which would be effective againstdrug-resistant bacteria and would not contribute toresistance development. This is likely because the poly-mers act in the cell membranes or cell walls as mimicsof AMPs, although the molecular mechanism is not
fully understood yet at this point. There are distinc-tive differences in their structures and conformationsbetween AMPs and antimicrobial polymers. AMPshave homogeneous structures (defined sequence andmolecular length) and are programmed to form a helixwith regulated amphiphilicity (Figure 1). On the otherhand, synthetic polymers are heterogeneous in termsof chemical structures (random sequences, MW dis-tribution, and tacticity) and structural conformations(random coil). Despite these differences, antimicrobialpolymers are effective against bacteria and selectiveto bacteria over human cells to the same degree asAMPs. This may indicate that the physicochemicalproperties of polymers and peptides are importantfor antimicrobial action rather than the structureswhile it remains still a question whether syntheticcopolymers have the same molecular mechanism(s) ofAMPs.
We envision that these polymers are mostimmediately useful for potential applications as top-ical antimicrobial agents including targets such asperiodontal disease and dermatological applications.However, before this point, there remain many lim-itations and hurdles to overcome before moving toclinical applications. For example, there has beenlittle investigation into potential side effects causedby the polymeric antimicrobials. While the selectivityfor bacterial cells over mammalian cells is a positiveindicator of compatibility, the long-term applicationof these molecules and their possible interactions withnatural flora and/or interaction with complex phys-iological systems must be investigated. This is likelyto be more involved than the peptidic counterparts asthe potential for wildly different polymer backbones,compositions, and sizes will increase the need forindividualized studies.
At this moment, there are only a few polymersidentified as effective agents, and the translationalstudies of these polymers are still limited. However,the scientific reports on antimicrobial polymers haveincreased significantly over the last two decades, andnew advanced chemical methodologies have beenutilized to improve the polymer performance. Manydifferent kinds of polymer structures have alreadybeen developed much like AMPs which also havelarge diversity in their chemical structures. This sug-gests that the AMP-mimetic design of polymers willcontinue to be versatile and applicable to numer-ous polymer types, which will in turn facilitate theimplementation of antimicrobial polymers as a newgeneration of antibiotics.
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 63

Advanced Review wires.wiley.com/nanomed
REFERENCES1. Fernandes P. Antibacterial discovery and development
- the failure of success? Nat Biotechnol 2006, 24:1497–1503.
2. Fischbach MA, Walsh CT. Antibiotics for emergingpathogens. Science 2009, 325:1089–1093.
3. Levy SB. The Antibiotic Paradox. How Miracle DrugsAre Destroying the Miracle. New York: Plenum Press;1992.
4. Levy SB. Antibiotic resistance: an ecological imbalance.In: Chadwick DJ, Goode J, eds. Antibiotic Resistance:Origins, Evolution, Selection and Spread. 207 ed.Chichester: Wiley; 1997,1–14.
5. Levy SB, Marshall B. Antibacterial resistance world-wide: causes, challenges and responses. Nat Med 2004,10:S122–S129.
6. Nathan C. Antibiotics at the crossroads. Nature 2004,431:899–902.
7. Gabriel GJ, Som A, Madkour AE, Eren T, Tew GN.Infectious disease: connecting innate immunity to bio-cidal polymers. Mater Sci Eng R 2007, 57:28–64.
8. Kenawy ER, Worley SD, Broughton R. The chem-istry and applications of antimicrobial polymers:a state-of-the-art review. Biomacromolecules 2007,8:1359–1384.
9. Li P, Li X, Saravanan R, Li CM, Leong SSJ. Antimi-crobial macromolecules: synthesis methods and futureapplications. RSC Adv 2012, 2:4031–4044.
10. Timofeeva L, Kleshcheva N. Antimicrobial polymers:mechanism of action, factors of activity, and applica-tions. Appl Microbiol Biotechnol 2011, 89:475–492.
11. Palermo EF, Kuroda K. Structural determinants ofantimicrobial activity in polymers which mimic hostdefense peptides. Appl Microbiol Biotechnol 2010,87:1605–1615.
12. Epand RF, Mowery BP, Lee SE, Stahl SS, Lehrer RI,Gellman SH, Epand RM. Dual mechanism of bacte-rial lethality for a cationic sequence-random copolymerthat mimics host-defense antimicrobial peptides. J MolBiol 2008, 379:38–50.
13. Gabriel GJ, Pool JG, Som A, Dabkowski JM,Coughlin EB, Muthukurnar M, Tew GN. Interac-tions between antimicrobial polynorbornenes andphospholipid vesicles monitored by light scattering andmicrocalorimetry. Langmuir 2008, 24:12489–12495.
14. Rawlinson L-AB, Ryan SM, Mantovani G, Syrett JA,Haddleton DM, Brayden DJ. Antibacterial effectsof poly(2-(dimethylamino ethyl)methacrylate) againstselected Gram-positive and Gram-negative bacteria.Biomacromolecules 2010, 11:443–453.
15. Brogden KA. Antimicrobial peptides: pore formers ormetabolic inhibitors in bacteria? Nat Rev Microbiol2005, 3:238–250.
16. Zasloff M. Antimicrobial peptides of multicellularorganisms. Nature 2002, 415:389–395.
17. Marr AK, Gooderham WJ, Hancock REW. Antibacte-rial peptides for therapeutic use: obstacles and realisticoutlook. Curr Opin Pharmacol 2006, 6:468–472.
18. Hancock REW, Sahl HG. Antimicrobial and host-defense peptides as new anti-infective therapeutic strate-gies. Nat Biotechnol 2006, 24:1551–1557.
19. Shai Y. Mode of action of membrane active antimicro-bial peptides. Biopolymers 2002, 66:236–248.
20. Rodriguez de Castro F, Naranjo OR, Marco JA,Violan JS. New antimicrobial molecules and new antibi-otic strategies. Semin Respir Crit Care Med 2009,30:161–171.
21. Matsuzaki K. Why and how are peptide-lipid interac-tions utilized for self-defense? Magainins and tachy-plesins as archetypes. Biochim Biophys Acta Biomem-branes 1999, 1462:1–10.
22. Tossi A, Sandri L, Giangaspero A. Amphipathic, α-helical antimicrobial peptides. Biopolymers 2000,55:4–30.
23. Schmidt NW, Mishra A, Lai GH, Davis M, Sanders LK,Dat T, Garcia A, Tai KP, McCray PB, Jr. Ouellette AJ,et al. Criterion for amino acid composition of defensinsand antimicrobial peptides based on geometry ofmembrane destabilization. J Am Chem Soc 2011,133:6720–6727.
24. Sochacki KA, Barns KJ, Bucki R, Weisshaar JC. Real-time attack on single Escherichia coli cells by the humanantimicrobial peptide LL-37. Proc Natl Acad Sci U S A2011, 108:E77–E81.
25. Peschel A, Sahl H-G. The co-evolution of host cationicantimicrobial peptides and microbial resistance. NatRev Microbiol 2006, 4:529–536.
26. Franco OL. Peptide promiscuity: an evolutionary con-cept for plant defense. FEBS Lett 2011, 585:995–1000.
27. Hancock REW, Lehrer R. Cationic peptides: a newsource of antibiotics. Trends Biotechnol 1998,16:82–88.
28. Giuliani A, Pirri G, Nicoletto S. Antimicrobial peptides:an overview of a promising class of therapeutics. CentEur J Biol 2007, 2:33.
29. Papo N, Shai Y. Host defense peptides as newweapons in cancer treatment. Cell Mol Life Sci 2005,62:784–790.
30. Riedl S, Zweytick D, Lohner K. Membrane-active hostdefense peptides - challenges and perspectives for thedevelopment of novel anticancer drugs. Chem PhysLipids 2011, 164:766–781.
31. Wade D, Boman A, Wahlin B, Drain CM, Andreu D,Boman HG, Merrifield RB. All-d amino acid-containingchannel-forming antibiotic peptides. Proc Natl Acad SciU S A 1990, 87:4761–4765.
32. Oren Z, Ramesh J, Avrahami D, Suryaprakash N,Shai Y, Jelinek R. Structures and mode of mem-brane interaction of a short α helical lytic peptide
64 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013

WIREs Nanomedicine and Nanobiotechnology Antimicrobial polymers
and its diastereomer determined by NMR, FTIR,and fluorescence spectroscopy. Eur J Biochem 2002,269:3869–3880.
33. Tashiro T. Antibacterial and bacterium adsorb-ing macromolecules. Macromol Mater Eng 2001,286:63–87.
34. Chakrabarty S, King A, Kurt P, Zhang W, Ohman DE,Wood LF, Lovelace C, Rao R, Wynne KJ. Highlyeffective, water-soluble, hemocompatible 1,3-propylene oxide-based antimicrobials: poly[(3,3-quaternary/PEG)-copolyoxetanes]. Biomacromolecules2011, 12:757–769.
35. Stratton TR, Rickus JL, Youngblood JP. In vitro bio-compatibility studies of antibacterial quaternary poly-mers. Biomacromolecules 2009, 10:2550–2555.
36. Sambhy V, Peterson BR, Sen A. Antibacterial andhemolytic activities of pyridinium polymers as a func-tion of the spatial relationship between the positivecharge and the pendant alkyl tail. Angew Chem Int Ed2008, 47:1250–1254.
37. Kuroda K, DeGrado WF. Amphiphilic polymethacry-late derivatives as antimicrobial agents. J Am Chem Soc2005, 127:4128–4129.
38. Lienkamp K, Madkour AE, Musante A, Nelson CF,Nusslein K, Tew GN. Antimicrobial polymers preparedby ROMP with unprecedented selectivity: a molecu-lar construction kit approach. J Am Chem Soc 2008,130:9836–9843.
39. Mowery BP, Lee SE, Kissounko DA, Epand RF, EpandRM, Weisblum B, Stahl SS, Gellman SH. Mimicry ofantimicrobial host-defense peptides by random copoly-mers. J Am Chem Soc 2007, 129:15474–15476.
40. Oda Y, Kanaoka S, Sato T, Aoshima S, Kuroda K.Block versus random amphiphilic copolymersas antibacterial agents. Biomacromolecules 2011,12:3581–3591.
41. Song A, Walker SG, Parker KA, Sampson NS. Antibac-terial studies of cationic polymers with alternating,random, and uniform backbones. ACS Chem Biol 2011,6:590–599.
42. Avery CW, Palermo EF, McLaughin A, Kuroda K,Chen Z. Investigations of the interactions between syn-thetic antimicrobial polymers and substrate-supportedlipid bilayers using sum frequency generation vibra-tional spectroscopy. Anal Chem 2011, 83:1342–1349.
43. Mizutani M, Palermo EF, Thoma LM, Satoh K, Kami-gaito M, Kuroda K. Design and synthesis of self-degradable antibacterial polymers by simultaneouschain- and step-growth radical copolymerization.Biomacromolecules 2012, 13:1554–1563.
44. Palermo EF, Kuroda K. Chemical structure of cationicgroups in amphiphilic polymethacrylates modulatesthe antimicrobial and hemolytic activities. Biomacro-molecules 2009, 10:1416–1428.
45. Palermo EF, Lee DK, Ramamoorthy A, Kuroda K. Roleof cationic group structure in membrane binding and
disruption by amphiphilic copolymers. J Phys Chem B2011, 115:366–375.
46. Palermo EF, Sovadinova I, Kuroda K. Structural deter-minants of antimicrobial activity and biocompatibil-ity in membrane-disrupting methacrylamide randomcopolymers. Biomacromolecules 2009, 10:3098–3107.
47. Palermo EF, Vemparala S, Kuroda K. Cationic spacerarm design strategy for control of antimicrobial activityand conformation of amphiphilic methacrylate randomcopolymers. Biomacromolecules 2012, 13:1632–1641.
48. Sovadinova I, Palermo EF, Huang R, Thoma LM,Kuroda K. Mechanism of polymer-induced hemolysis:nanosized pore formation and osmotic lysis. Biomacro-molecules 2011, 12:260–268.
49. Sovadinova IP, Urban EF, Mpiga M, Caputo P,Kuroda GA. K Activity and mechanism of antimicrobialpeptide-mimetic amphiphilic polymethacrylate deriva-tives. Polymers 2011, 3:1512–1532.
50. Kuroda K, Caputo GA, DeGrado WF. The role ofhydrophobicity in the antimicrobial and hemolyticactivities of polymethacrylate derivatives. Chem EurJ 2009, 15:1123–1133.
51. Munoz-Bonilla A, Fernandez-Garcia M. Polymericmaterials with antimicrobial activity. Prog Polym Sci2012, 37:281–339.
52. NCCLS. Approved standards M7-A3. Methods forDilution Antimicrobial Susceptibility Tests for Bacte-ria That Grow Aerobically. Philadelphia, PA: NCCLS;2003.
53. Wiegand I, Hilpert K, Hancock REW. Agar and brothdilution methods to determine the minimal inhibitoryconcentration (MIC) of antimicrobial substances. NatProtoc 2008, 3:163–175.
54. Giacometti A, Cirioni O, Barchiesi F, Del Prete MS,Fortuna M, Caselli F, Scalise G. In vitro susceptibilitytests for cationic peptides: comparison of broth microdi-lution methods for bacteria that grow aerobically.Antimicrob Agents Chemother 2000, 44:1694–1696.
55. Centers for Disease Control and Prevention. Acine-tobacter baumannii infections among patients atmilitary medical facilities treating injured U.S. ser-vice members, 2002–2004. Morbid Mortal WeekRep 2004, Available at: http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5345a1.htm.
56. Caputo GA, London E. Cumulative effects of aminoacid substitutions and hydrophobic mismatch uponthe transmembrane stability and conformation ofhydrophobic α-helices. Biochem 2003, 42:3275–3285.
57. Jaud S, Fernandez-Vidal M, Nilsson I, Meindi-BeinkerNM, Hubner NC, Tobias DJ, von Heijne G, White SH.Insertion of short transmembrane helices by theSec61 translocon. Proc Natl Acad Sci U S A 2009,106:11588–11593.
58. Krishnakumar SS, London E. The control of trans-membrane helix transverse position in membranes byhydrophilic residues. J Mol Biol 2007, 374:1251–1269.
Volume 5, January/February 2013 © 2012 Wiley Per iodica ls, Inc. 65

Advanced Review wires.wiley.com/nanomed
59. Mishra VK, Palgunachari MN, Segrest JP, Ananthara-maiah GM. Interactions of synthetic peptide analogsof the class a amphipathic helix with lipids - evi-dence for the snorkel hypothesis. J Biol Chem 1994,269:7185–7191.
60. Monne M, Nilsson I, Johansson M, Elmhed N, vonHeijne G. Positively and negatively charged residueshave different effects on the position in the membraneof a model transmembrane helix. J Mol Biol 1998,284:1177–1183.
61. Schow EV, Freites JA, Cheng P, Bernsel A, von Hei-jne G, White SH, Tobias DJ. Arginine in membranes:the connection between molecular dynamics simula-tions and translocon-mediated insertion experiments.J Membr Biol 2011, 239:35–48.
62. Strandberg E, Killian JA. Snorkeling of lysine sidechains in transmembrane helices: how easy can it get?FEBS Lett 2003, 544:69–73.
63. Strandberg E, Morein S, Rijkers DTS, Liskamp RMJ,van der Wel PCA, Killian JA. Lipid dependence of mem-brane anchoring properties and snorkeling behavior ofaromatic and charged residues in transmembrane pep-tides. Biochem 2002, 41:7190–7198.
64. Mondal J, Zhu X, Cui QA, Yethiraj A. Sequence-dependent interaction of β-peptides with membranes.J Phys Chem B 2010, 114:13585–13592.
65. Epand RF, Sarig H, Mor A, Epand RM. Cell-wallinteractions and the selective bacteriostatic activ-ity of a miniature oligo-acyl-lysyl. Biophys J 2009,97:2250–2257.
66. Raafat D, von Bargen K, Haas A, Sahl H-G. Insightsinto the mode of action of chitosan as an antibac-terial compound. Appl Environ Microbiol 2008, 74:3764–3773.
67. Gibney K, Sovadinova I, Lopez AI, Urban M, RidgwayZ, Caputo GA, Kuroda K. Poly(ethylene imine)s asantimicrobial agents with selective activity. MacromolBiosci 2012, 12:1279–1289.
68. Lienkamp K, Kumar K-N, Som A, Nuesslein K,Tew GN. ‘‘Doubly selective’’ antimicrobial polymers:how do they differentiate between bacteria? Chem EuroJ 2009, 15:11710–11714.
69. Fischer D, Li YX, Ahlemeyer B, Krieglstein J, Kissel T.In vitro cytotoxicity testing of polycations: influenceof polymer structure on cell viability and hemolysis.Biomaterials 2003, 24:1121–1131.
70. Tew GN, Clements D, Tang HZ, Arnt L, Scott RW.Antimicrobial activity of an abiotic host defense pep-tide mimic. Biochim Biophys Acta Biomembranes 2006,1758:1387–1392.
71. Goldman MJ, Anderson GM, Stolzenberg ED, Kari UP,Zasloff M, Wilson JM. Human β-defensin-1 is a salt-sensitive antibiotic in lung that is inactivated in cysticfibrosis. Cell 1997, 88:553–560.
72. Maisetta G, Di Luca M, Esin S, Florio W, Brancati-sano FL, Bottai D, Campa M, Batoni G. Evaluation ofthe inhibitory effects of human serum components onbactericidal activity of human β defensin 3. Peptides2008, 29:1–6.
73. Nie N, Tu Q, Wang J-C, Chao F, Liu R, Zhang Y,Liu W, Wang J. Synthesis of copolymers using den-dronized polyethylene glycol and assay of their bloodcompatibility and antibacterial adhesion activity. Col-loids Surfaces B 2012, 97:226–235.
74. Yang WJ, Cai T, Neoh K-G, Kang E-T, Dickinson GH,Teo SL-M, Rittschof D. Biomimetic anchors forantifouling and antibacterial polymer brushes on stain-less steel. Langmuir 2011, 27:7065–7076.
75. Wang YQ, Xu JJ, Zhang YH, Yan HS, Liu KL. Antimi-crobial and hemolytic activities of copolymers withcationic and hydrophobic groups: a comparison ofblock and random copolymers. Macromol Biosci 2011,11:1499–1504.
76. Chu B. Structure and dynamics of block-copolymer col-loids. Langmuir 1995, 11:414–421.
77. Tominaga Y, Mizuse M, Hashidzume A, Morishima Y,Sato T. Flower micelle of amphiphilic random copoly-mers in aqueous media. J Phys Chem B 2010,114:11403–11408.
78. Nederberg F, Zhang Y, Tan JPK, Xu K, Wang H,Yang C, Gao S, Guo XD, Fukushima K, Li L, et al.Biodegradable nanostructures with selective lysis ofmicrobial membranes. Nat Chem 2011, 3:409–414.
79. Liu L, Xu K, Wang H, Tan JPK, Fan W, Venkatra-man SS, Li L, Yang Y-Y. Self-assembled cationic pep-tide nanoparticles as an efficient antimicrobial agent.Nat Nanotechnol 2009, 4:457–463.
80. Rosenberg LE, Carbone AL, Romling U, Uhrich KE,Chikindas ML. Salicylic acid-based poly(anhydrideesters) for control of biofilm formation in Salmonellaenterica serovar typhimurium. Lett Appl Microbiol2008, 46:593–599.
81. Woo GLY, Mittelman MW, Santerre JP. Synthesis andcharacterization of a novel biodegradable antimicrobialpolymer. Biomaterials 2000, 21:1235–1246.
82. Zawaneh PN, Singh SP, Padera RF, Henderson PW,Spector JA, Putnam D. Design of an injectable syntheticand biodegradable surgical biomaterial. Proc Natl AcadSci U S A 2010, 107:11014–11019.
66 © 2012 Wiley Per iodica ls, Inc. Volume 5, January/February 2013