a sphingolipid having a novel type of ceramide and lacto-n ... · the journal of biological...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. U6. No. 5, Issue of March 10, pp. 1192-1200, 19il
Printed in U.S.A.
A Sphingolipid Having a Novel Type of Ceramide and
Lacto-N-Fucopentaose III*
(Received for publication. August 3, 1970)
HUNG-JU YANG~ AND SEN-ITIROH HAKOMORI~
From the Biochemical Laboratory, Department of Pathobiology, School of Public Health and Community Medicine, University of Washington, Seattle, Washington 98105
SUMMARY
A new sphingoglycolipid was isolated from human adeno- carcinomas. The glycolipid had a novel ceramide which contained predominantly 4-hydroxysphinganine (phyto- sphingosine) and a high content of o-hydroxy fatty acids. The structure of the carbohydrate moiety was identified as 0-fi-galactosyl-(1 -+ 4)-[0-cr-L-fucosyl-(1 --f 3)1-O-P- (N-acetyl)glucosaminosyl-(1 --f 3)-0-/3-galactosyl-(1 + 4)- glucose, which was identical with that of lacto-N-fucopen- taose III (KOBATA, A., AND GINSBURG, V., J. Biol. Chem., 244, 5496 (1969)).
The precipitating antibody against this glycolipid was produced in rabbit, the antiserum was allowed to react specifically with this glycolipid but not with Lewis a or Lewis b glycolipid, and the precipitin reaction was inhibited by lacto-N-fucopentaose III.
In epithelial glandular tissue, or in adenocarcinoma derived from the epithelial tissue, the presence of a number of fucose-con- taining glycosphingolipids has been reported (l-4). The fucose-glycolipid is completely absent in neural tissue and fibroblastic cells, but can be found in a limited quantity in various organs (4), erythrocytes (5, 6), and serum (7) as the blood group ABH or Lewis antigens.
The fucose-glycolipids of adenocarcinoma have been char- acterized by the absence of blood group A or B activities and by the copresence of Lewis a- and Lewis b-active glycolipids, in contrast to the blood group status of normal tissue (4, 8). Iso- lation and characterization of Lea-active* glycolipid and two Leb- active glycolipids have been reported2 (4, 9, 10). During the
* This work was supported by United States Public Health Service Grant CA-10909 and by the American Cancer Society Grant T-475.
$ Present address, Department of Biochemistry, National Re- search Council of Canada, Ottawa, Canada.
0 To whom communications and reprint requests should be ad- dressed.
1 The abbreviations used are Lea and Leb, blood group Lewis a and Lewis b.
2 S. Hakomori and R. W. Jeanloz, unpublished details of the structure of Lea glycolipids.
course of these studies, the presence of a glycolipid having no blood group ABH and Lewis specificities, but still having a chemical composition similar to Lea-active glycolipid, has been noticed (Al) (4). Studies on chemical structure and immuno- logical properties of this glycolipid are reported in this paper. The glycolipid has been characterized by having a novel cer- amide and an unique carbohydrate moiety.
EXPERIMENTAL PROCEDURES
Isolation of Glycolipid A1
The water-soluble glycolipid fraction of adenocarcinoma tissues was prepared from the upper layer of the Folch chloro- form-methanol extract by 10 times partitioning with aqueous phases according to the method of Svennerholm (11). The water-soluble glycolipid fraction was fractionated on a DEAE- cellulose column to separate water-soluble neutral glycolipid from gangliosides according to Rouser et al. (12). The fucose- containing glycolipids were eluted by chloroform-methanol- water (7 :3 :O.l) and by chloroform-methanol-water (5 :5 :O.l). These fractions were combined. The gangliosides were re- tained on the column. For retention of gangliosides, it is es- sential to prepare a homogeneous densely packed column by mechanically pressing the column. The yield of the combined fucose-containing glycolipid fractions was about 30 to 35 mg from 500 g of the original tissue. The fucose-containing glyco- lipid fraction was further fractionated on a Biosil A column (30 mg of substance was loaded on a column, 1.5 x 20 cm, and eluted with 100 ml each of the following chloroform-methanol mixtures: 8:2, 775~225, 75:25, 725~275, 7~3, 675:325, 65:45, 6:4, and 55 :45. Ten-milliliter aliquots were collected on a frac- tion collector, and each fraction was analyzed by thin layer chromatography on a Silica Gel H thin layer plate, developed with chloroform-methanol-water (60 : 35 : 8). Ceramide trihexos- ide, globoside, hematoside, gangliosides, and a ceramide penta- saccharide of rabbit red blood cells (13) were run as references.
Fractions appearing as elongated spots which had a similar migration rate to that of a ceramide pentasaccharide (13) were eluted with chloroform-methanol, 725 :275 to 675 :325 (Frac- tion A). After Fraction A was eluted, another fraction (B) was subsequently eluted with chloroform-methanol, 60:40 to 55:45, which showed a migration rate similar to disialoganglioside on thin layer chromatography.
Fraction A consisted of several components and was most effectively separated into components after acetylation by
1192
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of March 10. 1971 H. Yang and S. Hakomori 1193
thin layer chromatography on Silica Gel H using less polar organic solvents. Fraction A was acetylated in pyridine and acetic anhydride (3 :2, v/v), and, after the solvents were evap- orated with a large quantity of toluene in a rotary evaporator, the residue was applied to a thin layer plate (Silica Gel H) and developed with 1,2-dichloroethane-methanol-water (93 :7 :0.2) five times with intermediate drying for the best resolution. Five distinct bands were revealed by exposure of the plates to a 3-mm channel of iodine vapor contained in a plastic container. The bands were eluted with chloroform-methanol (2 :l) and de- acetylated in chloroform-methanol-O.5 y0 sodium methoxide (2 : 1:0.6) at room temperature for 30 min, followed by neutral- ization with acetic acid. The samples were dialyzed against distilled water and evaporated in a vacuum. According to the order of mobilities on thin layer chromatography, these sub- fractions of A were called Ar, AZ to As. The A1 fraction mi- grated about 12 cm from the start line and Ag migrated about 3 cm from the start line under the above-mentioned condition. In most adenocarcinoma cases, Al, Aa, and A4 were found to be predominant, but the relative quantity of these fractions varied depending on the case. One case of cecal adenocarcinoma (Case 6089, King County Hospital, Seattle) yielded 21 mg of a pure A1 glycolipid from 500 g of the original wet tissue. Sub- fraction Ar, the major component from this case, was freed from Lea-active glycolipid and has been used throughout the present investigation.
Glycolipids, Oligosaccharides, Fatty Acids, and Long Chain Bases Used as Reference Compounds
The following glycolipids, used as references in analyses, were prepared in this laboratory: glucosylceramide from human spleen, lactosylceramide, galactosyl-(1 -+ 4)galactosyl(1 + 4)glucosyl- ceramide(ceramide trihexoside), N-acetylgalactosaminosyl-(1 -+ 3)galactosyll(1 -+ 4)galactosyl-(1 + 4)glucosylceramide (globo- side of human erythrocytes) (5, 14), N-glycolylneuraminosyl- lactosylceramide (hematoside from equine erythrocytes) (14, 15), N-acetylneuraminosyllactosylceramide (hematoside from human spleen) (16), cr-galactosyl-(1 -+ 3)galactosyl(1 ---) 3)N-acetyl- glucosaminosyl-(1 -+ 3)galactosyl-(1 -+ 4)glucosylceramide (13), mono-, di-, and trisialoganglioside of human brain (17).
O+Galactopyranosyl-(1 + 3)-O-~-N-acetylglucosaminosyl- (1 ---f 3) -O-P-galactopyranosyl- (1 + 4)glucose(lacto-N-tetraose) (18)) 0-P-galactopyranosyl- (1 --) 4).O-~-N-acetylglucosaminosyl- (1 + 3).0-/3-galactopyranosyl-(1 --f 4)glucose(lacto-N-neotet- raose) (19), 0-a-L-fucopyranosyl-(1 + 2)0-P-n-galactopyran- osyl-(1 -+ 3)0-P-N-acetylglucosaminosyl-(1 + 3).0-P-galacto- pyranosyl-(1 -+ 4)glucose(lacto-N-fucopentaose I) (20), O-B- galactopyranosyl-(1 --) 3)-[0-or-L-fucopyranosyl-(1 -+ 4)]-0- /3-N-acetylglucosaminosyl- (1 + 3)-0-fi-galactopyranosyl-(1 -+ 4)glucose(lacto-N-fucopentaose II) (21), O-fi-galactopyranosyl-
(1 + 4)-[0-cr-L-fucopyranosyl-(1 --) 3)]-O-B-N-acetylglucos- aminosyl- (1 + 3) -O+galactopyranosyl- (1 + 4) glucose(lacto- N-fucopentaose III) (22), and 0-ar-L-fucopyranosyl-(1 + 2)-0- fl-n-galactopyranosyl-(1 --f 3))[0-a-L-fucopyranosyl-(1 + 4)]- 0-~-N-acetylglucosaminosyl- (1 + 3)0-@-galactopyranosyl-(1 --f 4)glucose(lacto-N-difucohexose II) (23) were the products iso- lated from milk and were donated by Dr. Adalin Gauhe, Max Planck Institute, Heidelberg, Germany, and by Drs. Akira Kobata and Victor Ginsburg, National Institute of Arthritis and Metabolic Diseases, National Institutes of Health, Bethesda, Maryland. 0-/3-N-Scetyl-galactosaminosyl-(1 ---) 3)galactosyl-
(1 + 4)galactosyl-(1 -+ 4)glucose(globo&tetraose) was pre- pared in this laboratory from globoside (24). Normal and (Y- hydroxy fatty acids were purchased from the Applied Science Laboratory, State College, Pennsylvania. Sphingosine (octa- decasphingenine), 3-0-methylsphingosine, and dihydrosphingo- sine (octadecasphinganine) were prepared from methanolysate of brain cerebroside by Dr. Bader Siddiqui in this laboratory. A long chain fatty base fraction of brain monosialoganglioside was used for reference of C&sphingosine (eicosasphingenine). Phytosphingosine(4-hydroxyoctadecasphinganine) was a gift from Dr. C. C. Sweeley, Michigan State University, and was prepared from yeast (25).
Methods of Analyses
Optical Rotation-Optical rotation was determined in the electrometric polarimeter (Perkin-Elmer) with a IO-cm cell; chloroform-methanol (2 : 1) and pyridine (freshly distilled) were used as solvents.
Homogeneity of Glycolipid-The homogeneity of the glyco- lipid was determined by thin layer chromatography on Silica Gel H or borate-impregnated Silica Gel H (26) developed with chloroform-methanol-water (65 : 30 : 8, lower layer), chloroform- methanol-water (60:35:8), propanol-water (7:3), and tetra- hydrofuran-2-butanone-water (8 :2 : 1). Acetylated glycolipid was separated on activated Silica Gel H with 1,2-dichloroethane- methanol-water (90 : 10 : 0.1). Iodine vapor and 0.2 y. orcinol in 2 M sulfuric acid were used as spot detectors.
sugar Composition-The ratios of glucosamine, galactosamine, glucose, galactose, and fucose were determined by gas chromatog- raphy on an ECNSS-M3 column as the reduced and acetylated compounds (27-29). In order to increase the recovery of hexosamines without destruction of neutral sugars in glyco- lipids, acetyolysis was followed by hydrolysis. Acetolysis dis- solved lipids and preferentially hydrolyzed the hexosaminidic linkage rather than the N-acetyl group; thus a higher yield of hexosamines and neutral sugars was obtained (30). Glycolipid (50 to 100 pg) was dissolved in 0.3 ml of 0.5 N HzS04 in 90% acetic acid and heated in a Teflon-lined, screw-capped tube for 16 hours at 80”. The mixture was heated for another 5 hours after the addition of 0.3 ml of water. The hydrolysate was filtered through 200 mg of dried Dowex l-X8 (acetate form) in a small column and washed with 2 ml of methanol. The filtrate and washings were combined and evaporated under nitrogen. The residue was dissolved in 0.3 ml of water and reduced by the addition of 10 mg of sodium borohydride for 3 hours at room temperature. The excess borohydride was destroyed by 2 drops of glacial acetic acid and 0.3 ml of chloroform was added, mixed, and centrifuged to remove the fatty acid released. The water layer was evaporated to dryness under nitrogen four times with 3 ml of methanol containing one drop of acetic acid, and finally dried completely over Pz05 in a vacuum. The residue was dis- solved in 0.3 ml of acetic anhydride and heated at 100” for 2 hours. The acetic anhydride was evaporated under nitrogen with the addition of an excess of toluene and the residue was partitioned between equal volumes of chloroform and water. The chloroform layer was removed and evaporated to dryness in a small conical test tube. For gas-liquid chromatography, the
8 An organosilicon polyester resulting from the combination of ethylene glycol succinate and a silicone of a cyanoethyl type, purchased from Applied Science Laboratories, State College, Pennsylvania.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

1194 A New Xphinyolipid Vol. 246, No. 5
residue was dissolved in 20 to 30 ~1 of acetone. The solution (2 ~1) was injected and developed on a 4-foot column of 3% ECNSS-M coated on Gas Chrom Q at 180” for 16 mm, and then programmed up to 210”. By this method 80 to 100% of the glycolipid hexosamines were recovered and well separated. The method of Sweeley and Walker (31) was used occasionally for analysis of neutral sugars.
Fatty Acids and Long Chain Bases-These were analyzed by gas chromatography. A sample (l- to 2-mg) was hydrolyzed in 1 N hydrochloric acid in 82% aqueous methanol for 18 hours (32) and the hydrolysate was evaporated to dryness under nitrogen. The residue was dissolved in chloroform and put onto a small Biosil A column (1 x 7 cm) prepared from a chloro- form suspension. Fatty acids and methyl esters were eluted by chloroform, and sphingosines (and a part of the methyl glycosides) were eluted by 2 column volumes of methanol (32). The fatty acids and their methyl ester fraction were treated with dry methanolic HCl or diazomethane to convert free carboxyl groups into ester and analyzed by gas chromatography with or without trimethylsilylation on an SE-52 column (33). In a separate run, normal and ar-hydroxy fatty acid methyl esters were separated on a Silica Gel G thin layer plate with diethyl ether-hexane (15 :85) (34), followed by analysis by gas chroma- tography. The sphingosine fraction was dissolved in chloroform, shaken repeatedly with 0.1 N NaOH followed by water, evap- orated to dryness, and converted to an N-acetyl derivative with acetic anhydride methanol (35). The N-acetyl sphingosines were analyzed by gas chromatography on a SE-30 column (6 feet) according to Gaver and Sweeley (32), with an improved tri- methylsilylating reagent of Carter and Gaver (35). A peak was separated in a capillary tube and analyzed by mass spec- trometry.
Methods of Degradation and Methylation Study
Liberation of Oligosaccharide-Oligosaccharides were released from the glycolipid by osmium-catalyzed periodate oxidation followed by alkaline treatment (24), the modified degrada- tion procedure of Wiegandt and Baschang (36). The oligosac- charides were separated on Whatman No. 3MM paper with the following solvents: ethylacetate-pyridine-water (12:5:4), ethyl- acetate-pyridine-acetic acid-water (5 :5 :3 : 1) in a tank saturated with ethyl acetate-pyridine-water (40 : 11: 8)) methyl ethyl ketone-acetic acid-water (7:3:1), and on thin layer chromatog- raphy on Silica Gel G, developed three times with butanol- pyridine-water (5:4:3). The indirect Ehrlich reaction of the oligosaccharides was carried out by the microscale (total volume, 0.5 ml) method of Morgan and Elson (37).
Hydrolysis of Fucosyl Residue-Glycolipid was hydrolyzed in aqueous 0.1 N hydrochloric acid for 1 hour at 80”, and that of the liberated oligosaccharide was hydrolyzed in aqueous 0.05 N HCI for 1 hour at 80”. The hydrolysate was neutralized by pyridine and was either analyzed directly or after dialysis.
Elimination of Nonreducing Terminal-Glycolipids were de- graded according to the method of Smith and Unrau (38). The oxidation was carried out in 0.02 M sodium periodate in 0.5% aqueous solution for 24 hours, pH 5, at 4”. Under these condi- tions, oxidation of the internal part of the carbohydrate chain was prevented by a strong aggregation of aqueous micelles of glycolipids.
Methylation Study-The glycolipid was methylated in di- methyl formamide and methyl iodide (39), and also in dimethyl
sulfoxide with a methyl sulfinyl-carbanion base and methyl iodide (40, 41). The methylated glycolipid was isolated by counter- current partition between chloroform and water. Completion of methylation was checked by the disappearance in infrared spectra of the absorption bands due to the presence of an hy- droxyl group. The permethylated product was subjected to formolysis in 90% formic acid at 100” for 3 hours, followed by hydrolysis with 0.3 N hydrochloric acid for 18 hours, diluted with 3 volumes of water, and lyophilized (Residue I). Residue I was passed through a small column of Dowex 50 (H+ form, 0.5 x 3 cm) and washed with 5 bed volumes of water. The eluate
and washings were combined and evaporated in a vacuum to a small volume (0.5 ml), and 10 mg of sodium borohydride were added. After 5 hours, 1 drop of glacial acetic acid was added to the solution; it was evaporated to dryness and repeatedly evap- orated with methanol. The residue was acetylated with acetic anhydride. The acetylated, partially methylated sugar alcohols thus prepared were analyzed by gas chromatography (27, 28, 29) and by mass spectrometry according to the method of BjGrn- dal, Lindberg, and Svensson (42). Mass spectra of the partially methylated alditol acetates were kindly furnished by Professor B. Lindberg, Stockholm University, Sweden. The standard compounds of partially methylated alditol acetate were either synthetic compounds or those prepared from oligosaccharide or glycolipids of well established structure and separated on gas chromatography. In some cases, methyl-cY-glucoside or methyl- cr-galactoside was methylated with methyl iodide and silver oxide (Purdie methylation) for 3 hours (Aliquot A) and for 10 hours (Aliquot B). Aliquot A contained mono- and dimethyl compounds and Aliquot B contained tri- and tetramethyl compounds as major components, which were converted to alditol acetate by the procedure described above and analyzed by gas chromatography and mass spectrometry. The patterns of these compounds were used as references. The hexosamine or its methylated product was eluted from the Dowex 50 column by 3 bed volumes of 1 N hydrochloric acid, and evaporated to dryness. The residue was directly trimethylsilylated by the improved reagent of Carter and Gaver (35), and analyzed in gas chromatography with a Gas Chrom Q column (6 feet) coated with SE-30 at 180” and a Gas Chrom Q column (6 feet) coated with 10% butanediol succinate polyester at 220”. The standard reference was 6-0-methylglucosamine hydrochloride prepared according to Jeanloz (43).
Determination of Antigenic Property
Blood Group A, B, Lea, and Leb Activzties-These activities have been determined by inhibition of respective hemagglutina- tion caused by respective specific normal antisera, with micro- titer plates with or without addition of auxiliary lipids (9,44,45). Activities for Lea and Leb specificities were further tested by precipitin reaction with immune anti-Lea or -Leb goat sera of Marcus and Grollman (46) on agar diffusion plates. The normal anti-A and -B sera were purchased from Ortho Diagnostics, Inc., Raritan, New Jersey, the normal anti-Lea and anti-Leb sera were donated by Dr. Eloise Gibblet, King County Central Blood Bank, and the precipitating anti-Lea and -Leb sera were donated by Dr. Donald M. Marcus, Albert Einstein College of Medicine, Bronx, New York. Blood group H activity was determined by hemagglutination inhibition caused by anti-H Ulex europeus phytoagglutinin.
Reaction with Anti-Type XIV Pneumococcal Polysaccharide
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from
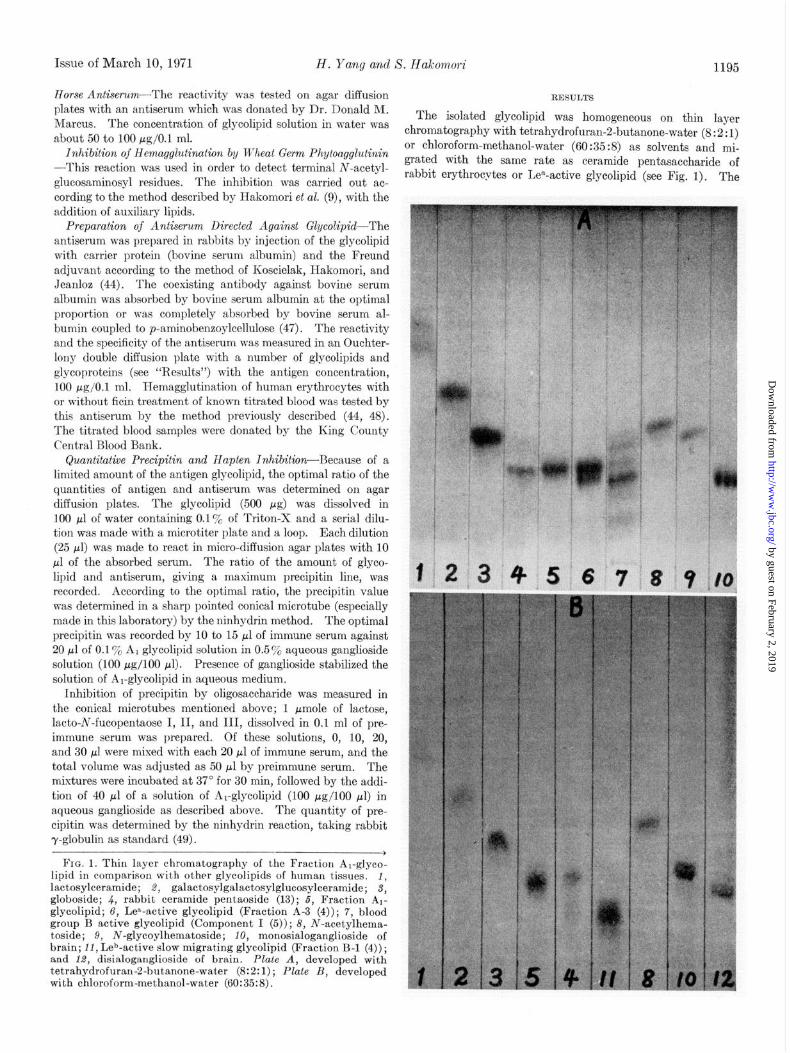
Issue of March 10, 1971 H. Yang and S. Hakomori 1195
Horse Antiserum-The reactivity was tested on agar diffusion RESULTS
plates with an antiserum which was donated by Dr. Donald M. Marcus. The concentration of glycolipid solution in water was
The isolated glycolipid was homogeneous on thin layer
about 50 to 100 clg/O.l ml. chromatography with tetrahydrofuran-2-butanone-water (8:2:I)
Inhibition of Hemagglutination by Wheat Germ Phytoagglutinin or chloroform-methanol-water (60:35:8) as solvents and mi-
--This reaction was used in order to detect terminal N-acetyl- grated with the same rate as ceramide pentasaccharide of
glucosaminosvl residues. The inhibition was carried out ac- rabbit erythrocytes or Lea-active glycolipid (see Fig, 1). The
cording to the method described by Hakomori et al. (9), with the addition of auxiliary lipids.
Preparation of Antiserum Directed Against Glycolip&-The antiserum was prepared in rabbits by injection of the glycolipid with carrier protein (bovine serum albumin) and the Freund
h, ., ;id 4
adjuvant according to the method of Koscielak, Ffakomori, and ,, r5: 1. >*
Jeanloz (44). The coexisting antibody against bovine serum ?A albumin was absorbed by bovine serum albumin at the optimal proportion or was completely absorbed by bovine serum al- bumin coupled to p-aminobenzoylcellulose (47). The reactivity and the specificity of the antiserum was measured in an Cuchter- lony double diffusion plate with a number of glycolipids and ” glycoproteins (see “Results”) with the antigen concentration, 100 pgiO.1 ml. Hemagglutination of human erythrocytes with or without ficin treatment of known titrated blood was tested by this antiserum by the method previously described (44, 48). The titrated blood samples were donated by the King County 1 Central Blood Bank,
Quantitative Precipitin and Hapten Inhibition-Because of a limited amount of the antigen glycolipid, the optimal ratio of the quantities of antigen and antiserum was determined on agar diffusion plates. The glycolipid (500 pg) was dissolved in ’ 100 ~1 of water containing 0.1 o/0 of Triton-X and a serial dilu- tion was made with a microtiter plate and a loop. Each dilution (25 ~1) was made to react in micro-diffusion agar plates with 10 ~1 of the absorbed serum. The ratio of the amount of glyco- lipid and antiserum, giving a maximum precipitin line, was recorded. According to the optimal ratio, the precipitin value was determined in a sharp pointed conical microtube (especially made in this laboratory) by the ninhydrin method. The optimal precipitin was recorded by 10 to 15 ~1 of immune serum against 20 ~1 of 0.1% Ai glycolipid solution in 0.5 y0 aqueous ganglioside solution (100 pg/lOO ~1). Presence of ganglioside stabilized the solution of Ai-glycolipid in aqueous medium.
Inhibition of precipitin by oligosaccharide was measured in the conical microtubes mentioned above; 1 pmole of lactose, lacto-N-fucopentaose I, II, and III, dissolved in 0.1 ml of pre- immune serum was prepared. Of these solutions, 0, 10, 20, and 30 ~1 were mixed with each 20 ~1 of immune serum, and the total volume was adjusted as 50 ~1 by preimmune serum. The mixtures were incubated at 37” for 30 min, followed by the addi- tion of 40 ~1 of a solution of Al-glycolipid (100 pg/lOO ~1) in aqueous ganglioside as described above. The quantity of pre- cipitin was determined by the ninhydrin reaction, taking rabbit -y-globulin as standard (49).
FIG. 1. Thin layer chromatography of the Fraction A,-gfyco- lipid in comparison with other glycolipids of human tissues, 1, lactosylceramide; 8, galactosylgalactosylglucosylceramide; d, globoside; 4, rabbit ceramide pentaoside (13); 6, Fraction A,- glycolipid; 6, Lea-active glycolipid (Fraction A-3 (4)); 7, blood group B active glycolipid (Component I (5)); 8, N-acetylhema- toside; 9, N-glycoylhematoside; 10, monosialoganglioside of brain; 11, Leb-active slow migrating glycolipid (Fraction B-l (4)) ; and 1.8, disialoganglioside of brain. Plate A, developed with tetrahydrofuran-2-butanone-water (8:2:1) ; Plate B, developed with chloroform-methanol-water (60:35:8).
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

1196 A New Xphingolipid Vol. 246, No. 5
a
C
X
\_.:.:
8 16 24 32 Time (minutes1
FIG. 2. Gas-liquid chromatogram of sphingosines. A, stand- ard run: 1, three-Cl*-sphingosine; 2, erythro-Ci8sphingosine; and 3, phytosphingosine from yeast. B, sphingosine fraction of brain gangliosides: 1, threo-C18sphingosine; 2, erythro-CIs-sphingosine; 3, threo-C&-sphingosine; and 4, erythro-Czo-sphingosine. C, sphingosine fraction of Fraction Al-glycolipid. Separation was performed on an OV-17 column (6 feet) at 210” as N-acetyl-o-tri- methylsilyl derivatives according to the method of Gaver and Sweeley (32) and Carter and Gaver (35).
73
7%
50-
a
218 . I 187
: 160
B
1,I ,I 240 320 400
m/e
FIG. 3. Mass spectra of N-acetyl-tri-O-trimethylsilylphyto- sphingosine (A) and of the same derivative of a substance in Peak X/C of Fig. 2. Massspectra were recorded in a Hitachi-Perkin El- mer apparatus. Spectra were recorded at 60 e.v. (chamber volt- age), with an accelerating voltage of 3.6 kv, an ion source tempera- ture of 200”, filament current 3 amps, emission current 80 pa, target current 50 pa, and target voltage of 90 volts.
TABLE I Fatty acid compositions of Fraction AI-qlycolipid
Fatty acids are designated by chain length and the number of double bonds.
C1e:o CL:0 CL:0 Unknown czz 0 Unknown czz:o C24:0 C24:1
- 1N0rIXil
14.7
11.0
1.7
3.3 4.0
I a-Hydroxy
70 total
7.8 21.6
1.6
2.6 4.2
12.8 14.8
acetylated glycolipid was also homogeneous on Silica Gel H plate by multiple development with less polar solvents (1,2- dichloroethane-methanol-water, 90: 1O:O.l). Under the latter condition, the acetylated glycolipid was separated from other acetylated ceramide pentasaccharides.
The ceramide moiety of this glycolipid was characterized by having phytosphingosine (4-hydroxysphinganine) as the major (75%) and other sphingenines as only minor components. The gas chromatograms and mass spectra of the long chain bases are shown in Figs. 2 and 3. The presence of a large proportion of ol-hydroxy fatty acids was observed (Table I).
Because of the high content of 4-hydroxysphinganine, the major part of this glycolipid was not degraded by the osmium periodate-alkaline degradation (24), and the nondegraded glyco- lipid was detected in the reaction mixture by thin layer chroma- tography on Silica Gel G, developed with butyl alcohol-pyridine- water (5:4:3). However, a small but significant quantity of a homogeneous oligosaccharide (10% of the theoretical), which should be derived from glycolipid containing unsaturated long chain bases, was released by this degradation procedure. The structural characterization of the carbohydrate moiety of the glycolipid was carried out on the intact glycolipid as well as on the oligosaccharide released from the sphingenine base.
The carbohydrate moiety of this glycolipid was identified as la&o-N-fucopentaose III (22) on the following basis. (a) The sugar analysis indicated that glycolipids contained a pentasac- charide which consisted of 2 moles of galactose and 1 mole each of glucosamine, fucose, and glucose (Fig. 4). The Smith deg- radation (38) eliminates 1 of 2 galactose moles present and the fucose mole. It is known that under the controlled condition only the nonreducing terminal of glycolipid is eliminated in aqueous micelles of glycolipids. The ceramide trisaccharide found in the Smith degradation product was capable of inhibiting hemagglutination caused by wheat germ agglutinin; i.e. this trihexosylceramide has N-acetylglucosamine as the terminal. This agglutinin has been known to react with any glycolipid or oligosaccharide having a /3&acetylglucosaminosyl residue at the nonreducing terminal (50, 51).4 Removal of a fucosyl residue gave a ceramide tetrahexoside which was able to react with anti-type XIV antisera reactive to a ,&galactosyl-(1 --f 4)- N-acetylglucosaminosyl residue at the nonreducing terminal. (b) Permethylated glycolipid gave, after formolysis, hydrolysis, reduction, and aeetylation, four peaks of partially methylated alditol acetate according to the procedure of Bjorndal, Lindberg,
4 H. Yang and S. Hakomori, unpublished observations.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of March 10, 1971 H. Yang and S. Hakomori 1197
FIG. 4. Gas chromatography of acetylated alditols and hexos- aminitols. A, globoside: 1, galactitol; 2, glucitol; and 3, galactos- aminitol. B, A1-glycolipid: 1, fucositol; b, galactitol; 3, glucitol; and 4, glucosaminitol. C, standard sugars: 1, fucositol; b, galacti- tol; 3, glucitol; 4, glucosaminitol; and 5, galactosaminitol. All components were acetates.
and Svensson (29). The peaks (Fig. 5E) were identified as 2,3,4-trimethylfucositol, 2,3,4,6-tetramethylgalactitol, 2,4, 6-trimethylgalactitol, and 2,3,6-trimethylglucitol by relative retention value and by mass spectrometry of the peak according to Bjijrndal et al. (42). The absence of 2,3,6-trimethylgalactitol was significant, since this sugar is characteristically present in the hydrolysate of permethylated globoside and asialoganglio- side. The gas chromatogram of these sugar acetates is shown in Fig. 5, and the mass spectra are given in Table II. The partially methylated glucosamine, separated from the hydrolysate by a Dowex 50-H+ column, gave a component appearing with the same retention time as the trimethylsilyl derivative of 6-O- methylglucosamine by gas chromatography on a SE-30 column (Fig. 6) and on butanediol succinate polyester; under these conditions 3-0-methylglucosamine was shown to be eluted with much higher retention time (22). (c) The oligosaccharide liberated by the modified Wiegandt’s degradation (24, 36) was clearly separated from lacto-N-fucopentaose III (22). On prolonged chromatography with ethyl acetate-pyridine-water (12 : 5 :4) or with methylethylketone-acetic acid-water (7 : 3 : 1)) the oligosaccharide of the glycolipid was somewhat slower than lacto-N-fucopentaose II, but identical with lacto-N-fucopentaose III. After acetylation, lacto-N-fucopentaose III migrated faster than lacto-N-fucopentaose II on a Silica Gel H plate
FIG. 5. Gas chromatography of partially methylated alditol acetate on a 3% ECNSS-M column (6 ft). A-l, 2,3,4,6-tetra- methylgal; A-2, 2,3,6-trimethylglu; B-l, 2,4,6-trimethylgal; B-2, 2,3,6-trimethylgal; B-3, 2,3,6-trimethylglu; C-l, 2,3,4,6-tetra- methylgal; C-2, 2,3,6-trimethylgal; C-3, 2,3,6-trimethylglu; D-l, 2,4,6-trimethylgal; D-S, 2,3,6-trimethylglu; E, sugars in the hy- drolysate of permethylated AI-glycolipid; 1, 2,3,4-trimethylfuc; 9, 2,3,4,6-tetramethylgal; 3, 2,4,6-trimethylgal; and 4, 2,3,6-tri- methylglu.
developed with 1,2 dichloroethane-methanol-water (97 :3 :O.l). Two spots, possibly corresponding to cx and p anomers, were given by the acetylated lacto-N-fucopentaose II and III, re- spectively. The oligosaccharide was hydrolyzed with 0.05 N
hydrochloric acid to remove the fucosyl residue. The chromato- graphic behavior and the indirect Ehrlich reaction of the re- sulting tetrasaccharide were compared with those of lacto-N- tetraose (18), lacto-N-neotetraose (19), and globe-N-tetraose (24). These tetraoses were well separated in most of the solvent systems and the tetraose yielded by hydrolyzing the fucosyl residue of the pentaose of this glycolipid was shown to have a migration rate identical with that of lacto-N-neot.etraose on paper and thin layer chromatography (Fig. 7). The indirect Ehrlich reaction (37) of this tetraose was completely negative as in the lacto-N-neotetraose. Lacto-N-tetraose and globo-N-tetraose gave a positive reaction. (d) The glycolipid showed a weak levorotation, [a]? - 1 .O” (in chloroform-methanol, 2 : 1) ; -5.2” (in pyridine). Dextrorotatory shifting of the optical rotation
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from
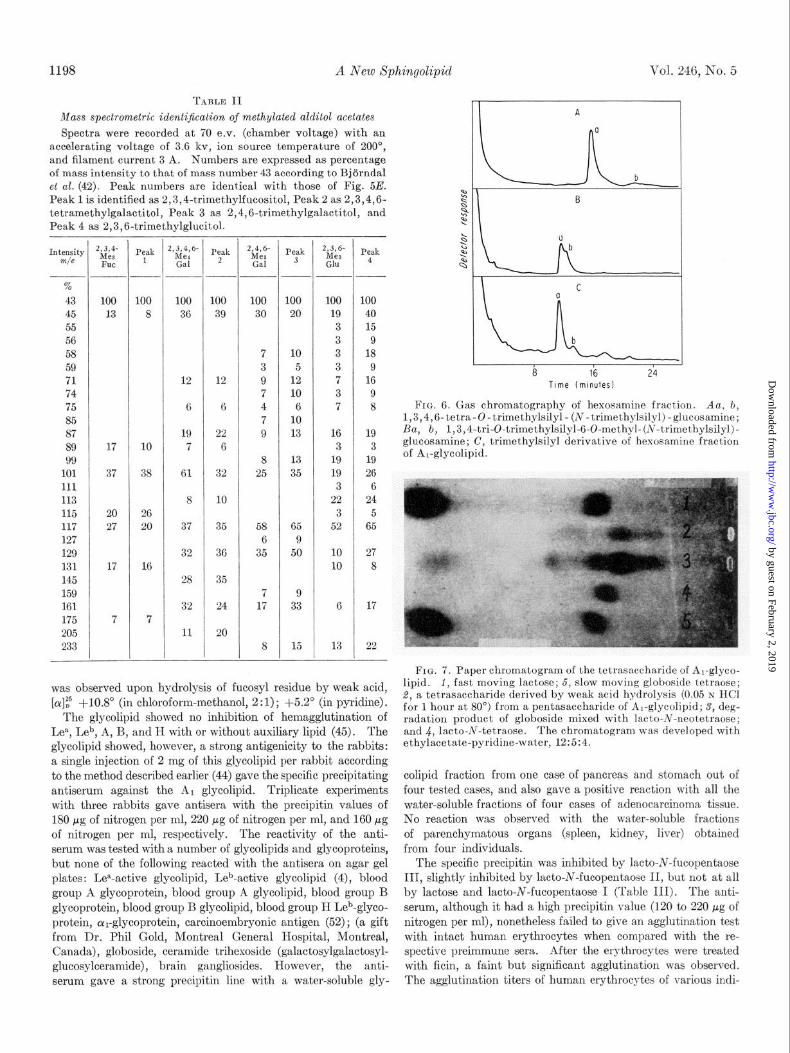
1198 A New Xphingolipid Vol. 246, No. 5
TARLE II
Mass spectrometric identification of methylated alditol acetates Spectra were recorded at 70 e.v. (chamber voltage) with an
accelerating voltage of 3.6 kv, ion source temperature of 200”, and filament current 3 A. Numbers are expressed as percentage of mass intensity to that of mass number 43 according to Bjiirndal et al. (42). Peak numbers are identical with those of Fig. 5E. Peak 1 is identified as 2,3,4-trimethylfucositol, Peak 2 as 2,3,4,6- tetramethylgalactitol, Peak 3 as 2,4,6trimethylgalactitol, and Peak 4 as 2,3,6-trimethylglucitol.
IntZty % 43 45 55 56 58 59 71 74 75 85 87 89 99
101 111 113 115 117 127 129 131 145 159 161 175 205 233
- Peak
1 1,3,4- MC8 FUC
100 100 100 LOO 100 100 13 8 36 39 30 20
12 12
6 6
19 22 7 6
61 32
8 10
37 35
32 36
28 35
32 24
11 20
7 10 3 5 9 12 7 10 4 6 7 10 9 13
17 10
37 38 8 13
25 35
20 26 27 20 58 65
6 9 35 50
17
7
16
7 17
7
-!-
8
9 33
15
-t-- v&4,6 Me4 Gal
-
-.
Peak 2
- 2&-
Gal Peak
3 “&-
Glu
Peak 4
100 100
19 40 3 15 3 9 3 18 3 9 7 16 3 9 7 8
16 3
19 19 3
22 3
52
19 3
19 26 *;,;>. . . ./ I 6
24 ‘” 5
65
10 10
6 17
13 I - 22
1 A
1 8 16 24
Time (minutes)
Fro. 6. Gas chromatography of hexosamine fraction. Aa, b, 1,3,4,6-tetra-O-trimethylsilyl- (N-trimethylsilyl)-glucosamine; Ba, b, 1,3,4-tri-0-trimethylsilyl-6-O-methyl-(N-trimethylsilyl)- glucosamine; C, trimethylsilyl derivative of hexosamine fract.ion of Al-glycolipid.
was observed upon hydrolysis of fucosyl residue by weak acid, [cr]E5 f10.8” (in chloroform-methanol, 2:l); f5.2” (in pyridine).
The glycolipid showed no inhibition of hemagglutination of Lea, Leb, A, B, and II with or without auxiliary lipid (45). The glycolipid showed, however, a strong antigenicity to the rabbits: a single injection of 2 mg of this glycolipid per rabbit according to the method described earlier (44) gave the specific precipitating antiserum against the A r glycolipid. Triplicate experiments with three rabbits gave antisera with the precipitin values of 180 pg of nitrogen per ml, 220 pg of nitrogen per ml, and 160 pg of nitrogen per ml, respectively. The reactivity of the anti- serum was tested with a number of glycolipids and glycoproteins, but none of the following reacted with the antisera on agar gel plates : Lea-active glycolipid, Leb-active glycolipid (4)) blood group A glycoprotein, blood group A glycolipid, blood group B glycoprotein, blood group B glycolipid, blood group H Leb-glyco- protein, cur-glycoprotein, carcinoembryonic antigen (52) ; (a gift from Dr. Phil Gold, Montreal General Hospital, Montreal, Canada), globoside, ceramide trihexoside (galactosylgalactosyl- glucosylceramide), brain gangliosides. However, the anti- serum gave a strong precipitin line with a water-soluble gly-
FIG. 7. Paper chromatogram of the tetrassccharide of Ar-glyco- lipid. I, fast moving lactose; 5, slow moving globoside tetraose; 2, a tetrasaccharide derived by weak acid hydrolysis (0.05 N HCl for 1 hour at 80”) from a pentasaccharide of Al-glycolipid; S, deg- radation product of globoside mixed with lacto-N-neotetraose; and .J, lacto-AT-tetraose. The chromatogram was developed with ethylacetate-pyridine-water, 12:5:4.
colipid fraction from one case of pancreas and stomach out of four tested cases, and also gave a positive reaction with all the water-soluble fractions of four cases of adenocarcinoma tissue, No reaction was observed with the water-soluble fractions of parenchymatous organs (spleen, kidney, liver) obtained from four individuals.
The specific precipitin was inhibited by lacto-N-fucopentaose III, slightly inhibited by lacto-N-fucopentaose II, but not at all by lactose and lacto-N-fucopentaose I (Table III). The anti- serum, although it had a high precipitin value (120 to 220 pg of nitrogen per ml), nonetheless failed to give an agglutination test with intact human erythrocytes when compared with the re- spective preimmune sera. After the erythrocytes were treated with ficin, a faint but significant agglutination was observed. The agglutination titers of human erythrocytes of various indi-
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of March 10, 1971 H. Yang and S. Hakomori 1199
TABLE III
Inhibition of precipitin between Fraciion Al-glycolipid and its antiserum (No. 1)” by oligosaccharides
Quantity of oligosaccharides and percentage of inhibition of precipitin
caused by those quantitiesb
0.1 /ml& 0.2 @de 0.3 jmole
Lactose........................ 0 0 0 Lacto-N-fucopentaose I.. 5 Lacto-N-fucopentaose II.. 5 15 20 Lact,o-N-fucopentaose III.. 12 35 65
o Serum No. 1 had a precipitin value of 180 pg of nitrogen per ml.
b The quantity of sugar added against 10 ~1 of immune serum and 0.013 pmole (20 r;l of 0.1% glycolipid solution) and total volume 50 ~1.
viduals was determined with one serum (Serum 1) and the titers were found to be varied between I:8 and 1:256. No correlation was found between the intensity of the agglutina- tion and the known blood type as tested (A, B, H, Lea, Leb, Rh, I, P, M, and N). The samples of typed blood were furnished by Dr. E. Giblett of King County Central Blood Bank, and by the Hematology Department of the University of Washington.
DISCUSSION
The ceramide moiety of this glycolipid was characterized by the presence of a large quantity of phytosphingosine (4 hy- droxyoctadecasphinganine) and cY-hydroxy fatty acids. The presence of phytosphingosine in mammalian sphingolipid has been shown in ceramide, cerebroside, and sphingomyelin (53-56) but is very limited in the higher glycolipids. The long chain bases of brain gangliosides, hematosides, globoside, ceramide di- and trihexosides, of spleen and erythrocytes, and fucose- containing glycolipids of human erythrocytes carrying blood group A, B, H, and Leb specificities (5) have been investigated and no phytosphingosine, or only a trace, was detected in any of these glycolipids (5)?
The glycolipid, without being acetylated, migrated the same distance as Lea-active ceramide pentasaccharide and rabbit ceramide pentasaccharide (13). They were only partially sep- arated on the usual thin layer chromatography, but were com- pletely separated as acetylated compounds on thin layer chroma- tography. It is possible that the Lea-active glycolipid previously reported2 (9, 10) could be contaminated with the glycolipid described in this paper. The acetylated glycolipid migrated slightly faster than the Lea-active glycolipid, probably because lacto-N-fucopentaose III migrates faster than lacto-N-fuco- pentaose II after acetylation and chromatography in a 1,2 dichloroethane-methanol system.
The glycolipid and its acetylated derivative have been shown to be homogeneous on thin layer chromatography under various conditions (see “Results”), and the released oligosaccharide obtained by osmium periodate degradation was also homoge- neous. It is reasonable to assume, on an empirical basis, that the carbohydrate structure of a nondegraded glycolipid is identical with that of the released oligosaccharide. It is empir-
ically known that, so far as the carbohydrate of glycolipid is
743
(7H2)n
FHOH c=o
OH OH iH I I I
CH3-(CH2),3-CH-CH-CH-CH2
o~oAMgAIH~
OH 0
/r--Q f
NHCOCH3 OH OH
FIG. 8. A proposed structure for Fraction Al-glycolipid. The ceramide moiety was shown as N+-hydroxy fatty acyl)-4-hy- droxysphinganine since this structure represents the major com- ponent. Variation of the ceramide and fatty acid; see Fig. 2 and Table I.
homogeneous, an identical carbohydrate should be found ir- respective of the kind of long chain base present. The homoge- neity of the carbohydrate structure of this glycolipid was further evidenced by the stoichiometric yield of the partially methylated sugars in methylation analysis and by the results of a quanti- tative hapten inhibition.
The results of the methylation study, the Smith degradation, the creation of a cross-reactivity with type XIV pneumococcal polysaccharide by weak acid hydrolysis, the creation of a reactiv- ity with wheat germ agglutinin by Smith degradation, and the indirect Ehrlich reaction of a released pentasaccharide, or a tetrasaccharide derived therefrom, all supported the structure as shown in Fig. 8. Further evidence for this structure was furnished by the inhibition of antigen-antibody reaction by the lacto-hi-fucopentaose III.
This fucose-glycolipid has been found as the common blood group ABH Lewis-inactive component of five cases of adeno- carcinoma tissue (4). The quantity of lipid-bound fucose in parenchymatous organs (kidney, spleen, and liver) was extremely low and often below the detectable limit, and the distribution is probably limited to glandular epithelial tissue because the quan- tity of lipid-bound fucose is significantly high in glandular tissue (24). The glycolipid was isolated in relatively high yield from human adenocarcinoma tissue, and it is predicted that the occurrence of this glycolipid in tumor tissue is in higher incidence and in higher quantity than in normal glandular tissue, based on some immunological studies. It is conceivable that this glycolipid may play a role in determining the immuno- pathological specificity of human tumors. The glycolipid antigen is, however, independent from the carcinoembryonic antigen of Gold and Freedman (49), since the antiserum against this glycolipid did not react with the carcinoembryonic antigen and the carcinoembryonic antiserum did not react with this glycolipid.
The occurrence of this glycolipid in normal and adenocar- cinema tissue seemed independent of the known blood group
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

1200 A New Xphingolipid Vol. 246, No. 5
systems. The glycolipid failed to inhibit either Lea- or Leb- hemagglutination, and the specific antisera directed against this glycolipid showed only a faint hemagglutination (or practically none) in the known types of human erythrocytes. A faint and irregular hemagglutination was observed after ficin treatment, and this agglutination was independent of any known blood group. This agreed with the finding of Kobata and Ginsburg (22) that lacto-N-fucopentaose III failed to inhibit various blood group systems. It is highly probable that this glycolipid could be a new isologous or allogeneic tissue antigen which is primarily present in the glandular tissue of a small fraction of the population, but has appeared or greatly increased in adeno- carcinoma as a consequence of incomplete synthesis of blood group ABH antigens, since the structure of the carbohydrate moiety of this glycolipid was similar to the proximal part of the Type II chain of blood group A and B haptens (51, 57) and incomplete synthesis of carbohydrate chains in malignant cells has been suggested by the model experiment with cultured cells and their viral and chemical transformants (58-60). Fur- ther extensive investigation on immunogenetical and immuno- pathological significance of this glycolipid hapten is in progress.
1.
2.
3. 4.
6.
6.
HAKOMORI. S., AND JEANLOZ, R. W.. J. Biol. Chem.. 239. PC3606 (1964).
I I
SUZUKI, C., MAICITA, A., AND YOSIZAWA, Z., Arch. Biochem. BioDhus.. 127, 140 (1968).
McKI&&; J. %I., Biocherkstry, 8, 679 (1969). HAKOMORI, S., AND ANDREWS, H. D., Biochim. Biophys. Acta,
202, 225 (1970). HAKOMORI, S., AND STRYCHARZ, G. D., Biochemistry, ‘7, 1279
(1968),
7. 8.
9.
10.
11. 12.
KOSCIELAK, J., PIASEIC, A., AND GORNIAK, H., in D. AMINOFF, (Editor), Blood and tissue antigens, Academic Press, New York,1970,p.163.
MARCUS, D. M., AND CASS, L. E., Science, 164, 553 (1969). MASAMUNE, H., in F. EG~AMI AND Y. OSHIMA (Editors),
Biochemistry and medicine of mucopolysaccharides, Maruzen Publishers, Tokyo, 1962, p. 85.
HAKOMORI, S., KOSCIELAIC, J.,B~ocx, K., AND JEANLOZ, R. W., J. Immunol., 98, 31 (1967).
HAKOMORI, S., Abstracts, Seventh International Congress of Biochemistry, Vol. Q, Tokyo, 1967, p. 713.
SVENNERHOLM, L., Acta Chem. &and., 17, 239 (1963). ROUSER, G., KRITCHEVSKY, G., HELLER, D., AND LIEBER, E.,
J. Amer. Oil Chem. Sot.. 40, 425 (1963). 13. ETO, T., ICHIKAWA, Y., N&HI-MURA; K.,‘ANDo, S., AND YAMA-
14.
15.
K~WA; T., J. Biochem. (Tokyo), 64, 265 (1968). YAMAKAWA. T.. IRIE. R.. IWANAGA. M.. J. Biochem. (Tokuol.
48, 919 (196Oj. ’ , , . “,,
YAMAKA’IVA, T., AND SUZUKI, S., J. Biochem. (Tokyo), 38, 199 (1951).
16. 17.
SVENNERHOLM, L.. Acta Chem. &and., 17, 860 (1963). PENICK, R. J.; M&SLER, M. H., AND-MCCLUER, R. H., Bio-
chim. Biovhus. Acta. 116. 279 (1966). 18. 19. 20.
21.
KUHN, R., A&~BAER, k. H.., Chek Bkr., 89, 504 (1956). KUHN, R., AND GAUHE, A., Chem. Ber., 96, 518 (1962). KUHN; R., BAER, H. H., AND GAUHE, A., Chem. Ber., 89, 2514
(1956). KUHN, R., BAER, H. H., AND GAUHE, A., Chem. Ber., 91, 364
(1958).
REFERENCES
22. KOBAT~, A., AND GINSBURG, V., J. Biol. Chem., 244, 5496 (1969).
23. KUHN, R., AND GAUHE, A., Chem. Ber., 93, 647 (1960). 24. HAKOMORI, S., J. Lipid Res., 7, 789 (1966). 25. THORPE, S. R., AND SWEELEY, C. C., Biochemistry, 6, 887
(1967). 26. 27.
28.
29.
30.
31. 32.
33. 34.
35. 36.
37.
38. 39.
40. 41.
42.
43. 46.
44.
45. 47.
48.
49.
50.
51. 52.
53. 54.
55.
56.
57.
58.
59.
60.
KEAN, E. L., J. Lipid Res., 7, 449 (1966). SAWARDEKER, J. S., SLONEKER, J. H., JEANES, A., Anal. Chem.,
37, 1602 (1965). KIM, J. H.,‘SHOME, B., LIAO, T. H., AND PIERCE, J. G., Anal.
Biochem., 20, 258 (1967). BJ~RNDAL, H., LINDBERG, B., AND SVENSSON, S., Acta Chem.
Stand., 21, 1801 (1967a). BJ~RNDAL, H., HELLERQVIST, C. G., AND LINDBERG, B. Angeur.
Chem. Int., Ed. Engl., 9, 610 (1970). SWEELEY, C. C., AND WALKER, B., Anal. Chem., 36,1946 (1965). GAVER, R. C., AND SWEELEY, C. C., J. Amer. Oil Chem. Sot.,
42, 294 (1965). SIDDIQUI, B., AND MCCLUER, R. H., J. Lipid Res., 9,366 (1968). VIOQUE, E., AND HOLMAN, R. T., J. Amer. Oil Chem. Sot.,
39, 63 (1962). CARTER, H. E., AND GAVER, R. C., J. Lipid Res., 8,391 (1967). WIEGANDT, H., AND BASCHANG, G., 2. Naturforsch., 206, 164
(1965). MORGAN, W. T. J., AND ELSON, L. A., Biochem. J., 28, 988
(1934). SMITH, F., AND UNRAU, A. M., Chem. Znd. (London), 881 (1959). KUHN, R., TRISCHMANN, I., AND LSw, I., Angew. Chem., 67,
32 (1955). HAKOMORI, S., J. Biochem. (Tokyo), 66, 205 (1964). SANFORD, P. A.. AND CONRAD. H. E.. Biochemistry. 6, 1508
(1966): “I
BJ~RNDAL, H., LINDBERG, B., AND SVENSSON, S., Carbohyd. Res., 6, 433 (196713).
JEANL~Z,. R. W.., J. Amer. Ch,em. Sot., 76, 558 (1956). MARCUS, D. M., AND GROLLMAN, P. A., J. Immunol., 97, 867
(1966). KOSCIELAK, J., HAKOMORI, S., AND JEANLOZ, R. W., Zmmunc-
chemistry, 6, 441 (1968). RAPPORT, M. M., AND GRAF, L., Progr. Allergy, 13, 273 (1969). DAVIS, R. V., BLANKEN, It. M., AND BEAGLE, R. J., Biochemis-
try, 8, 2706 (1969). BOORMAN, K. E., AND DODD, B. E., An introduction to blood
group serology, J. and A. Churchill, London, p. 801, 1961. KABAT, E. A., Experimental immunochemistry, Ed. 2, Charles
C Thomas, Publisher, Springfield, Illinois, 1961, p. 560. BURGER, M. M., AND GOLDBERG, A. R., Proc. Nat. Acad. Sci.
U. 8. A., 67, 359 (1967). WATKINS, W. M., Science, 162, 172 (1966). GOLD, P., AND FREEDMAN, S. O., J. Exp. Med., 121, 439; 122,
467 (1965). KARLSSON, K.-A., Acta Chem. &and., 18, 2397 (1964). KARLSSON. K.-A.. AND MARTENSSON. E.. Biochim. Bionhus.
Acta, 16i, 230 (1968). I I I I
KARLSSON, K.-A., AND STEEN, G. E., Biochim. Biophys. Acta, 162, 798 (1968).
MICHALEC, C., AND KOLMAN, Z., Clin. Chim. Acta, 13, 529 (1966).
LLOYD, K. W., AND KABAT, A., Proc. Nat. Acad. Sci. U. S. A., 61, 1470 (1968).
HAKOMORI, S., AND MURAKaMI, W. T., Proc. Nat. Acad. &i. U. S. A., 69, 254 (1968).
HAKOMORI, S., TEATHER, C., AND ANDREWS, H. D., Biochem. Biophys. Res. Commun. 33, 563 (1968).
SIDDIQUI, B., AND HAKOMORI, S., Cancer Res., 30, 2930 (1970).
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Hung-ju Yang and Sen-itiroh Hakomori-Fucopentaose IIINA Sphingolipid Having a Novel Type of Ceramide and Lacto-
1971, 246:1192-1200.J. Biol. Chem.
http://www.jbc.org/content/246/5/1192Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/246/5/1192.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from